Differential expression of U2AF 35 in the arthritic joint of avian reovirus-infected chicks Yi-Hsin Fan a , Chun-Da Lin a , Shiow-Her Chiou a, * , Kuan-Chih Chow b, * , Yung-Sheng Chi a , Long-Huw Lee c , Jui-Hung Shien c , Happy K. Shieh c a Graduate Institute of Veterinary Microbiology, National Chung Hsing University, 250 Kuo Kuang Road, Taichung 40227, Taiwan, ROC b Graduate Institute of Biomedical Sciences, National Chung Hsing University, 250 Kuo Kuang Road, Taichung 40227, Taiwan, ROC c Department of Veterinary Medicine, National Chung Hsing University, 250 Kuo Kuang Road, Taichung 40227, Taiwan, ROC Received 21 April 2006; accepted 5 July 2006 Abstract To identify cell types and genes that are differentially expressed during immunopathogenesis of avian reovirus (ARV)-induced viral arthritis (VA), we inoculated arthrotropic strain S1133 of ARVinto 1-day-old broilers, and examined tissue histology as well as RNA expression at different days post-inoculation (PI). Using immunohistochemical staining, we detected many CD68 expressing macrophages in and around the blood vessels of the arthritic joints. By RT-PCR, we found that expression of matrix metalloproteinase-2 (MMP-2) and bone morphogenetic protein-2 (BMP-2) was induced earlier in footpads and hock joints of ARV-infected chickens. By employing suppression subtractive hybridization (SSH) technique and RT-PCR, we further identified that small subunit of U2 snRNP auxiliary factor (U2AF 35 or U2AF1) mRNAwas differentially induced in the joint of ARV-infected chickens. By in situ hybridization (ISH), mRNA signals of U2AF 35 and BMP-2 were located in chondrocytes within/near the epiphyseal plate and secondary center of ossification, and in epidermal cells and dermal fibroblast-like cells of arthritic joints. In addition, U2AF 35 mRNAwas expressed in the inflammatory infiltrates of the bone marrow of ARV-infected arthritic joints, while MMP-2 was mainly detected in chondrocytes. Interestingly, among U2AF 35 , MMP-2, and BMP-2 that were differentially expressed in the joint of ARV-infected chickens, only U2AF 35 induction correlated well with arthritic manifestation. Because U2AF 35 may assist in mRNA splicing of proinflammatory chemokines and cytokines, our results indicated that U2AF 35 induction might play an immunopathological role in ARV-induced arthritis. This study has first associated U2AF 35 to viral arthritis. # 2006 Elsevier B.V. All rights reserved. Keywords: Avian reovirus (ARV); Bone morphogenetic protein-2 (BMP-2); Matrix metalloproteinase-2 (MMP-2); Small subunit of U2 snRNP auxiliary factor (U2AF 35 or U2AF1); Viral arthritis (VA) 1. Introduction Avian reovirus (ARV) infections have been asso- ciated with viral arthritis (VA) (also known as tenosynovitis), enteric diseases and other pathological conditions such as myocarditis, and pericarditis in chickens and turkeys (Clark et al., 1990; Robertson and Wilcox, 1986; van der Heide et al., 1981; Walker et al., 1972). Among these diseases, the causative role of ARV has only been established in VA (Robertson and Wilcox, 1986). Young chicks are especially susceptible to ARV infection, and resistance to infection increases with age (Rosenberger and Olson, 1991). The apparent lesions of ARV-induced VA include gross swelling and inflammatory infiltration of the www.elsevier.com/locate/vetimm Veterinary Immunology and Immunopathology 114 (2006) 49–60 * Corresponding authors. Tel.: +886 4 2285 1343; fax: +886 4 2285 9270. E-mail addresses: [email protected] (S.-H. Chiou), [email protected] (K.-C. Chow). 0165-2427/$ – see front matter # 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.vetimm.2006.07.003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/vetimm
Veterinary Immunology and Immunopathology 114 (2006) 49–60
Differential expression of U2AF35 in the arthritic joint of avian
reovirus-infected chicks
Yi-Hsin Fan a, Chun-Da Lin a, Shiow-Her Chiou a,*, Kuan-Chih Chow b,*,Yung-Sheng Chi a, Long-Huw Lee c, Jui-Hung Shien c, Happy K. Shieh c
a Graduate Institute of Veterinary Microbiology, National Chung Hsing University, 250 Kuo Kuang Road, Taichung 40227, Taiwan, ROCb Graduate Institute of Biomedical Sciences, National Chung Hsing University, 250 Kuo Kuang Road, Taichung 40227, Taiwan, ROC
c Department of Veterinary Medicine, National Chung Hsing University, 250 Kuo Kuang Road, Taichung 40227, Taiwan, ROC
Received 21 April 2006; accepted 5 July 2006
Abstract
To identify cell types and genes that are differentially expressed during immunopathogenesis of avian reovirus (ARV)-induced
viral arthritis (VA), we inoculated arthrotropic strain S1133 of ARV into 1-day-old broilers, and examined tissue histology as well as
RNA expression at different days post-inoculation (PI). Using immunohistochemical staining, we detected many CD68 expressing
macrophages in and around the blood vessels of the arthritic joints. By RT-PCR, we found that expression of matrix
metalloproteinase-2 (MMP-2) and bone morphogenetic protein-2 (BMP-2) was induced earlier in footpads and hock joints of
ARV-infected chickens. By employing suppression subtractive hybridization (SSH) technique and RT-PCR, we further identified
that small subunit of U2 snRNP auxiliary factor (U2AF35 or U2AF1) mRNAwas differentially induced in the joint of ARV-infected
chickens. By in situ hybridization (ISH), mRNA signals of U2AF35 and BMP-2 were located in chondrocytes within/near the
epiphyseal plate and secondary center of ossification, and in epidermal cells and dermal fibroblast-like cells of arthritic joints. In
addition, U2AF35 mRNA was expressed in the inflammatory infiltrates of the bone marrow of ARV-infected arthritic joints, while
MMP-2 was mainly detected in chondrocytes. Interestingly, among U2AF35, MMP-2, and BMP-2 that were differentially expressed
in the joint of ARV-infected chickens, only U2AF35 induction correlated well with arthritic manifestation. Because U2AF35 may
assist in mRNA splicing of proinflammatory chemokines and cytokines, our results indicated that U2AF35 induction might play an
immunopathological role in ARV-induced arthritis. This study has first associated U2AF35 to viral arthritis.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Avian reovirus (ARV); Bone morphogenetic protein-2 (BMP-2); Matrix metalloproteinase-2 (MMP-2); Small subunit of U2 snRNP
auxiliary factor (U2AF35 or U2AF1); Viral arthritis (VA)
1. Introduction
Avian reovirus (ARV) infections have been asso-
ciated with viral arthritis (VA) (also known as
tenosynovitis), enteric diseases and other pathological
* Corresponding authors. Tel.: +886 4 2285 1343;
fax: +886 4 2285 9270.
E-mail addresses: [email protected] (S.-H. Chiou),
[email protected] (K.-C. Chow).
0165-2427/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.vetimm.2006.07.003
conditions such as myocarditis, and pericarditis in
chickens and turkeys (Clark et al., 1990; Robertson and
Wilcox, 1986; van der Heide et al., 1981; Walker et al.,
1972). Among these diseases, the causative role of ARV
has only been established in VA (Robertson and Wilcox,
1986). Young chicks are especially susceptible to ARV
infection, and resistance to infection increases with age
(Rosenberger and Olson, 1991).
The apparent lesions of ARV-induced VA include
gross swelling and inflammatory infiltration of the

Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–6050
tibiotarsal and hock joints of infected chickens (Glass
et al., 1973; Kerr and Olson, 1969; Pertile et al., 1996).
In addition to VA lesions developed in the joints, ARV
can lead to a transient reduction of immune responses in
chickens (Montgomery et al., 1985; Neelima et al.,
2003; Pertile et al., 1995; Springer et al., 1983), which
may predispose chickens to secondary infections
(Kibenge et al., 1982). Interestingly, arthritic lesions
and temporal immunosuppression developed in ARV-
infected chickens resemble symptoms that are exhibited
in patients with rheumatoid arthritis (RA) (Harris, 1990;
Verwilghen et al., 1990). RA is a chronic inflammatory
joint disease mediated by activated macrophages, T
lymphocytes, and B lymphocytes. Macrophages, in
particular, play a pivotal role during RA pathogenesis
by expressing major histocompatibility complex
(MHC) class II molecules, proinflammatory cytokines,
and matrix degrading enzymes (Kinne et al., 2000). In
fact, Pertile et al. (1996) have studied in detail the
pattern of lymphocyte (CD4- and CD8-positive T
lymphocytes, B lymphocytes/plasma cells) infiltration
in ARV-induced VA and found it to be similar to that of
RA. Apart from the well-characterized lymphocytes,
they have also described the infiltration of Ia (class II
MHC antigen)-positive non-lymphocyte cell popula-
tions during acute phase arthritis (Pertile et al., 1996).
However, it remains to be determined whether these
non-lymphocyte cells are macrophages.
Studies in RA and other human arthritis have shown
that matrix metalloproteinases (MMPs) are major
catabolic enzymes participating in the destruction of
cartilage and bone matrix during arthritis pathogenesis
(Giannelli et al., 2004; Vincenti and Brinckerhoff,
2002). As a counterbalance, anabolic factors such as
bone morphogenetic proteins (BMPs) are induced to
repair damage in arthritis (Nakase et al., 2003).
However, it is still not clear whether MMPs and BMPs
are involved in ARV-induced VA as well.
In this study we employed immunohistochemistry,
suppression subtractive hybridization (SSH) technique
(Diatchenko et al., 1996), and reverse transcription-
polymerase chain reaction (RT-PCR) to recognize the
identity of the CD68-positive macrophage infiltrates
and to identify the gene(s) that may participate in
immunopathogenesis of ARV-induced viral arthritis.
2. Materials and methods
2.1. Virus and virus titration
The arthrotropic virulent strain S1133 of ARV (van der
Heide and Kalbac, 1975), originally obtained from
Vineland Laboratories (Vineland, NJ, USA), is a
challenge strain used for the vaccine evaluation. The
virus was propagated briefly in chicken embryo fibrob-
lasts (CEF). Virus titration was performed as described
(Robertson and Wilcox, 1984), and virus titer was
expressed as 50% tissue culture infectious dose (TCID50).
2.2. Experimental chickens
Sixty specific pathogen free (SPF) broiler chickens (1-
day-old) were used in this study. Each chicken was
inoculated either with 103 TCID50/50 ml of S1133 strain
ARV (ARV-infected group) or 50 ml of phosphate
buffered saline (PBS) (control group) into each footpad.
ARV-infected and PBS-inoculated chickens were housed
in separate isolation units with biologically filtered air. At
12 h, 1–4, 6, 8, 10, 12, 14, 16, 18, 21, 28, and 35 days after
inoculation, two chickens, respectively from the ARV-
infected group and the control group were sacrificed. The
experiment was carried out following the guideline of the
Institutional Animal Care and Usage Committee,
National Chung Hsing University, Taiwan.
2.3. Enzyme-linked immunosorbent assay (ELISA)
Serum was collected from each chicken before
sacrifice. A commercial ELISA kit (avian reovirus
antibody test kit, Idexx Laboratories, Westbrook, ME,
USA) was used to determine the titer of anti-ARV
antibody.
2.4. Histology and immunohistochemistry
Lung, heart, liver, spleen, kidney, bursa of Fabricius,
proventriculus, duodenum, jejunum, ileum, footpads,
and hock joints were dissected from each chicken and
cut into half. One half of the tissue was frozen in liquid
nitrogen and stored at �70 8C before RNA extraction.
The other half of the tissue was fixed with 10% PBS
buffered formalin, embedded in paraffin wax, sectioned
at 5 mm, and then mounted on glass slides. Two slides
for each tissue were stained with hematoxylin and eosin.
For immunohistochemistry, slides were incubated
with anti-CD68 monoclonal antibody (KP1) (Dako,
Carpenteria, CA, USA) by using the labeled strepta-
vidin–biotin (LSAB) method as described previously
(Chow et al., 2004; Peng et al., 1999).
2.5. RNA extraction
Frozen tissues of hock joints and footpads were
removed from�70 8C and ground under liquid nitrogen

Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–60 51
in a mortar. Total RNA was extracted by using Trizol
reagent (Invitrogen, Rockville, MD, USA) and RNA
concentration was determined by spectrophotometry.
2.6. Reverse transcription-polymerase chain
reaction (RT-PCR)
RT-PCR was performed in a 50 ml reaction volume
containing RT-PCR buffer, dNTPs (0.2 mM each), RNA
template (30 ng of total RNA), sense primer (20 pmol),
antisense primer (20 pmol), Taq DNA polymerase
(1 U), and AMV reverse transcriptase (1.5 U). RT
reaction was carried out at 42 8C for 45 min. After a
denaturation step at 94 8C for 3 min, 35 PCR cycles
were performed (denaturation at 94 8C for 1 min,
primer annealing at 52 8C for 1 min, extension at 72 8Cfor 1 min), followed by a final extension at 72 8C for
10 min. Primer sequences are listed in Table 1. RT-PCR
products were resolved by agarose gel electrophoresis.
Representative RT-PCR product was sequenced to
confirm the identity of each gene.
2.7. Suppression subtractive hybridizaition (SSH)
Equal amounts of total RNA extracted respectively
from hock joints and footpads of 2, 3, 4, 6, 8, 10 days
post-inoculated chickens were pooled together. Mes-
senger RNA (mRNA) was isolated from the pooled
RNA using a Chemagic mRNA T kit (Chemagen,
Baesweiler, Germany). About 5 mg of mRNA isolated
from each group (ARV-infected or control group) of
chickens were annealed with oligo dT primer and
reverse transcribed into cDNA by using reagents from a
commercial kit (SuperScriptTM
plasmid system with
GatewayTM
technology for cDNA synthesis and cloning,
Table 1
Primer sequences used in RT-PCR
Gene Amplicon (bp) Sense or antisense Primer seque
S1 (ARV) 941 Sense 50-GATACTG
Antisense 50-CACCTTA
S4 (ARV) 290 Sense 50-GCAGCCA
Antisense 50-ACATGGT
MMP-2 381 Sense 50-GCACCGT
Antisense 50-TGGCGTG
BMP-2 512 Sense 50-GTTGGTG
Antisense 50-GTTCTGT
U2AF35 416 Sense 50-AGACGTG
Antisense 50-ACTGGCG
b-actin 295 Sense 50-GTACCAC
Antisense 50-CTCGTCT
Invitrogen). After Rsa I digestion, cDNAs were
subjected to SSH using a PCR-select cDNA subtraction
kit (BD Clontech, Palo Alto, CA, USA). Briefly, Rsa I
digested-cDNA of ARV-infected group was divided into
two populations, ligated to specific adaptors, and
hybridized separately with excessive Rsa I digested-
cDNA of the control group. The two populations were
finally hybridized with each other and underwent two
rounds of PCR amplification. The resultant PCR
products were subcloned into pCRII vector, using a
TA cloning kit (Invitrogen).
2.8. In situ hybridization (ISH)
Based on a random primed DNA labeling method,
RT-PCR amplified ARV S1, MMP-2, BMP-2, and
U2AF35 DNA fragments were incorporated respectively
with fluorescein isothiocyanate (FITC) using a Fluor-
escein-High Prime kit (Roche, Mannheim, Germany).
ISH was performed as described previously (Chiou
et al., 2005). Briefly, formalin-fixed sections were
dewaxed and incubated with 20 mg/ml of proteinase K
for 15 min at room temperature, and rinsed with
distilled deionized water. DNA probe in hybridization
buffer (200 ng/ml of FITC-labeled DNA fragment, 50%
formamide, 6� SSC, 0.25% non-fat dry milk) was
added onto sections, denatured at 70 8C for 10 min, and
hybridized at 37 8C in a moist chamber overnight.
Sections were washed twice at room temperature for
5 min in wash buffer containing 2� SSC and 0.25%
Brij35, and once at 42 8C for 10 min in wash buffer
containing 0.1� SSC and 0.25% Brij35. Hybridized
products were incubated with alkaline phosphatase-
conjugated anti-FITC polyclonal antibodies (Dako) and
visualized by chromogen NBT/BCIP (Sigma, St. Louis,
nce Reference
TCATTGACTTCGA-30 Shapouri et al. (1995)
GGTGTCGATGCC-30
TGGACAACACCG-30 Chiu and Lee (1997)
GTGCGTAGTTGATTAGATCCGAG-30
CACCAATCATTAA-30 Aimes et al. (1994)
ACATCACTCCAG-30
CATCATAATGCAAG-30 Francis et al. (1994)
CGGCACACAGCAA-30
AGTTGTACGGGCG-30 Pacheco et al. (2004)
CTCCTCCAATACA-30
AATGTACCCTGGCA-30 Kost et al. (1983)
TGTTTTATGCGCA-30

Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–6052
Fig. 1. The titers of anti-ARV antibody at different days post-inocu-
lation (PI). Each chicken was inoculated with either S1133 strain ARV
(ARV-infected group, &) or phosphate buffered saline (control group,
*) into both footpads. Anti-ARV antibody, as determined by ELISA,
was first detected at day 6 PI. The titer of antibody increased steadily
after day 8 PI and finally reached 10854 (13.4 log2) at day 35 PI in
ARV-infected chickens. Anti-ARV antibody remained undetectable in
the control chickens throughout the experimental period. Each data
point represents an average result obtained from two chickens.
MO, USA). Positive staining (purple blue precipitate)
was identified by a light microscope.
3. Results
3.1. Virus inoculation and clinical manifestation
A total of 60 SPF broiler chickens (1-day-old) were
inoculated either with S1133 strain ARV (30 for the
ARV-infected group) or PBS (30 for the control group)
into each footpad. At 12 h, 1–4, 6, 8, 10, 12, 14, 16, 18,
21, 28, and 35 days after inoculation, two chickens
respectively from the ARV-infected group and the
control group were sacrificed and analyzed. ARV-
infected chickens manifested primarily with gross
swelling in the footpad (inoculation site) from day 3
to 6 post-inoculation (PI), and in the hock joint from day
4 to 8 PI. Between days 4 and 6 PI, most ARV-infected
chickens showed severe lameness. The infected chick-
ens gradually recovered macroscopically from day 8 to
12 PI. In contrast, chickens inoculated with PBS (the
control group) remained healthy throughout the
experimental period (to day 35 PI).
3.2. Humoral immune response in ARV-infected
chickens
To measure host humoral immune response, serum
was collected from each chicken, and titer of antibody
to ARV was determined by ELISA. Each data point
represents an average antibody titer of two chickens.
Anti-ARV antibody was detected at about 6 days after
ARV infection. The titer of antibody increased steadily
from day 8 to 35 PI. At 35th day (5 weeks) PI, anti-ARV
antibody reached an average titer of 10,854 (13.4 log2)
in infected chickens (Fig. 1). In contrast, anti-ARV
antibody was undetectable in the control chickens
throughout the experimental period (Fig. 1).
3.3. Histology, and immunohistochemistry
Histological staining revealed inflammatory cells in
synovium, dermis, bone marrow, and blood vessels of
footpads and hock joints of ARV-infected chickens
between day 1 and 8 PI. At day 1 PI, endothelial lining
of blood vessel in the secondary center of ossification
was seriously destructed, and many infiltrates leaked
out from blood vessel (Fig. 2B and D). At day 6 PI,
macrophage-like infiltrates were observed near the
secondary center of ossification in ARV-infected joints
(Fig. 2F). Using monoclonal antibody KP1 that is
specific for CD68, we performed immunohistochem-
istry to characterize these inflammatory infiltrates.
Based on the strong immunostaining of CD68 as well as
the macrophage-like morphology, we confirmed that
many of the inflammatory infiltrates were CD68-
positive macrophages. Fig. 3 showed the CD68 staining
on the footpad section at day 4 PI. CD68-expresssing
macrophages were abundant in blood vessel of the
ARV-infected chicken during arthritic manifestation
(Fig. 3B).
3.4. Persistent presence of ARV in the joints of
ARV-infected chickens
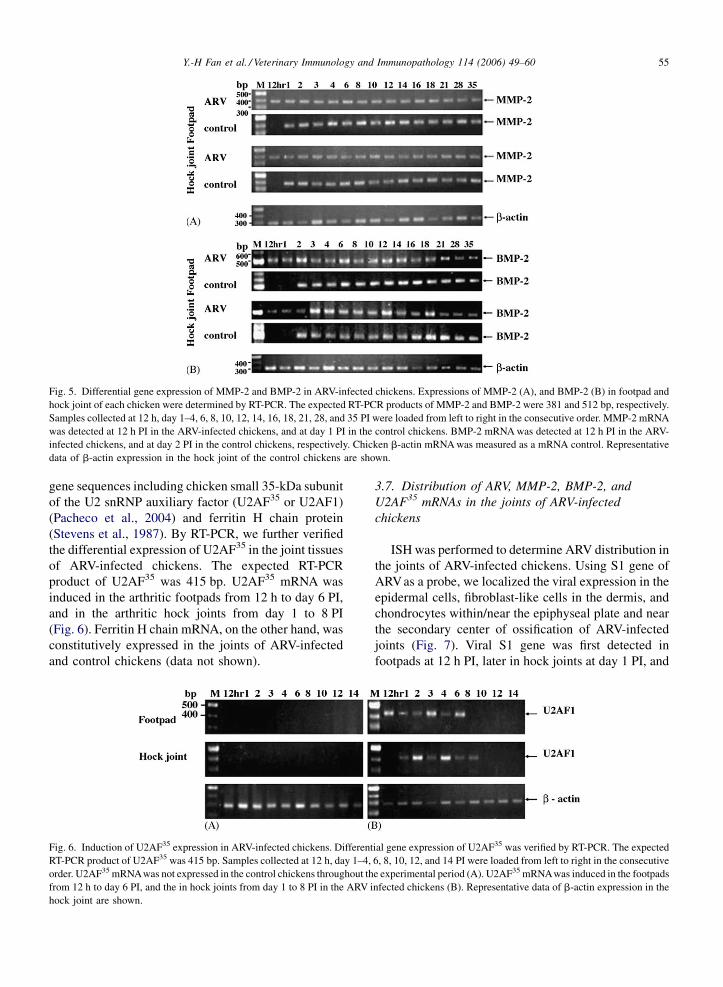
Using primers specific for viral S4 gene, we
performed RT-PCR to analyze the tissue distribution
of ARV S1133 strain. The expected RT-PCR product of
S4 gene region was 290 bp. At 12 h PI, S4 gene of the
virus was first detected in hock joints, footpads, liver,
and spleen. At day 1 PI, the virus continued to spread in
lung, heart, kidney, and proventriculus. It became
widespread in all tissues tested, including hock joints,
footpads, liver, spleen, lung, heart, kidney, proventri-
culus, bursa of fabricius, duodenum, jejunum, and
ileum, from day 2 to 6 PI (Fig. 4). However, the virus
gradually diminished in most tissues except for in
footpads and hock joints from day 8 to 16 PI, and was
exclusively confined to footpads and hock joints after
day 18 PI (Fig. 4). The RT-PCR product was sequenced
and the identity of S4 gene was confirmed. ARV signal
was undetectable in the control chickens (data not
shown).
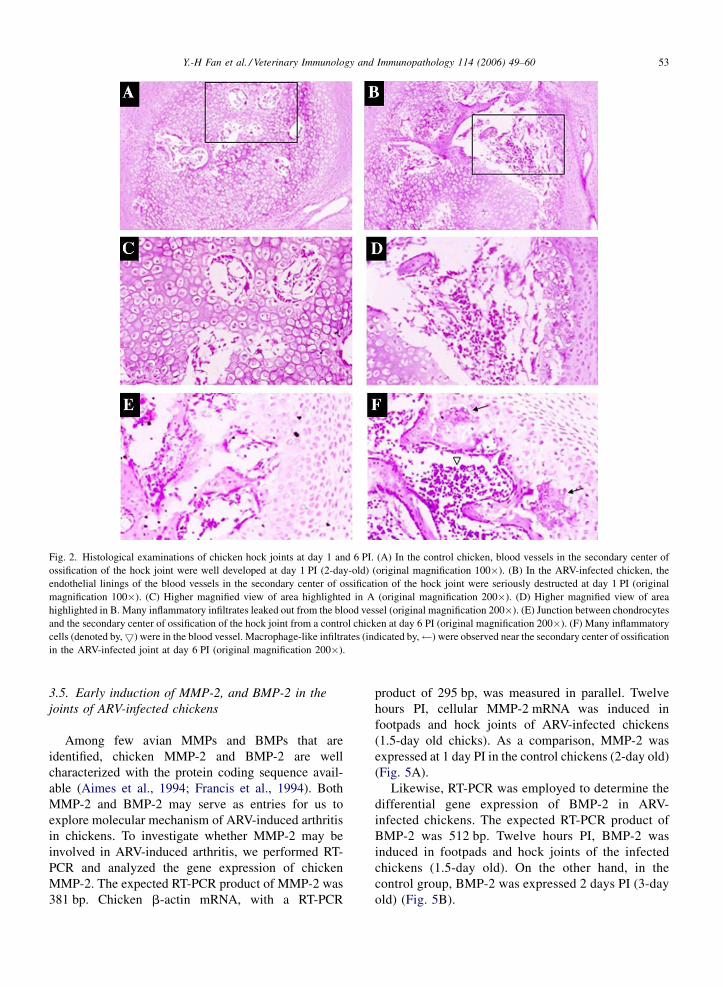
Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–60 53
Fig. 2. Histological examinations of chicken hock joints at day 1 and 6 PI. (A) In the control chicken, blood vessels in the secondary center of
ossification of the hock joint were well developed at day 1 PI (2-day-old) (original magnification 100�). (B) In the ARV-infected chicken, the
endothelial linings of the blood vessels in the secondary center of ossification of the hock joint were seriously destructed at day 1 PI (original
magnification 100�). (C) Higher magnified view of area highlighted in A (original magnification 200�). (D) Higher magnified view of area
highlighted in B. Many inflammatory infiltrates leaked out from the blood vessel (original magnification 200�). (E) Junction between chondrocytes
and the secondary center of ossification of the hock joint from a control chicken at day 6 PI (original magnification 200�). (F) Many inflammatory
cells (denoted by,5) were in the blood vessel. Macrophage-like infiltrates (indicated by, ) were observed near the secondary center of ossification
in the ARV-infected joint at day 6 PI (original magnification 200�).
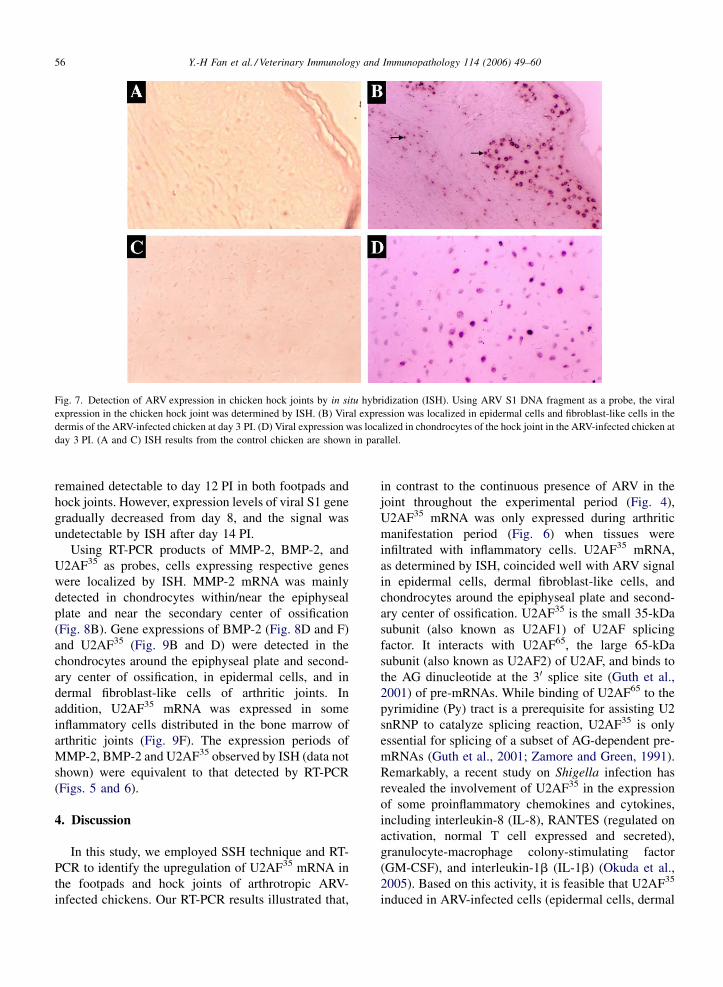
3.5. Early induction of MMP-2, and BMP-2 in the
joints of ARV-infected chickens
Among few avian MMPs and BMPs that are
identified, chicken MMP-2 and BMP-2 are well
characterized with the protein coding sequence avail-
able (Aimes et al., 1994; Francis et al., 1994). Both
MMP-2 and BMP-2 may serve as entries for us to
explore molecular mechanism of ARV-induced arthritis
in chickens. To investigate whether MMP-2 may be
involved in ARV-induced arthritis, we performed RT-
PCR and analyzed the gene expression of chicken
MMP-2. The expected RT-PCR product of MMP-2 was
381 bp. Chicken b-actin mRNA, with a RT-PCR
product of 295 bp, was measured in parallel. Twelve
hours PI, cellular MMP-2 mRNA was induced in
footpads and hock joints of ARV-infected chickens
(1.5-day old chicks). As a comparison, MMP-2 was
expressed at 1 day PI in the control chickens (2-day old)
(Fig. 5A).
Likewise, RT-PCR was employed to determine the
differential gene expression of BMP-2 in ARV-
infected chickens. The expected RT-PCR product of
BMP-2 was 512 bp. Twelve hours PI, BMP-2 was
induced in footpads and hock joints of the infected
chickens (1.5-day old). On the other hand, in the
control group, BMP-2 was expressed 2 days PI (3-day
old) (Fig. 5B).
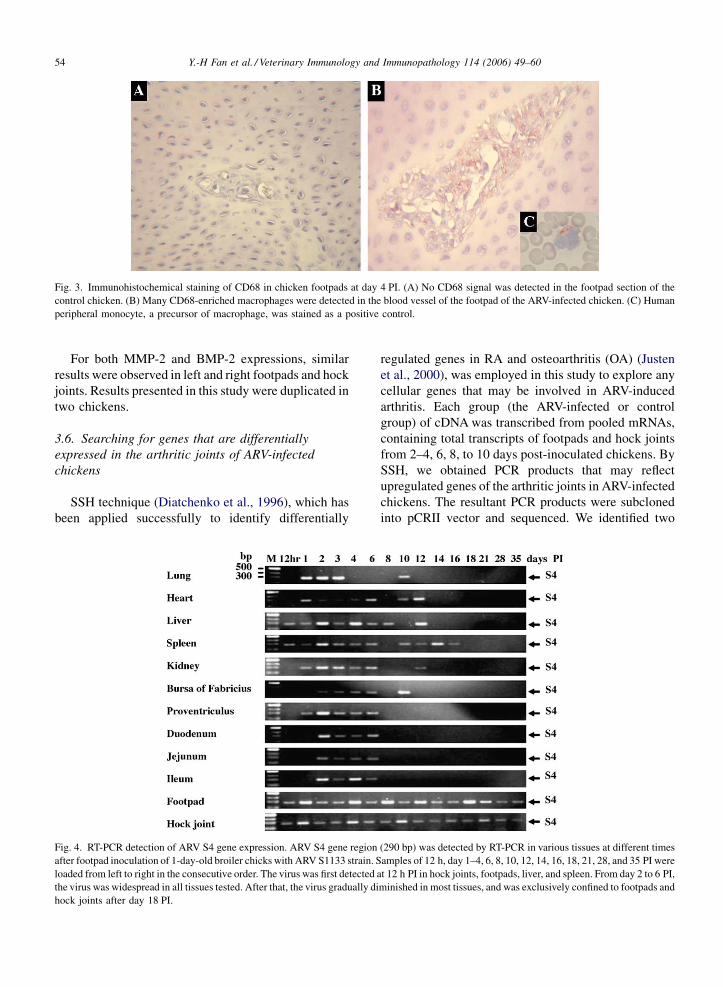
Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–6054
Fig. 3. Immunohistochemical staining of CD68 in chicken footpads at day 4 PI. (A) No CD68 signal was detected in the footpad section of the
control chicken. (B) Many CD68-enriched macrophages were detected in the blood vessel of the footpad of the ARV-infected chicken. (C) Human
peripheral monocyte, a precursor of macrophage, was stained as a positive control.
For both MMP-2 and BMP-2 expressions, similar
results were observed in left and right footpads and hock
joints. Results presented in this study were duplicated in
two chickens.
3.6. Searching for genes that are differentially
expressed in the arthritic joints of ARV-infected
chickens
SSH technique (Diatchenko et al., 1996), which has
been applied successfully to identify differentially
Fig. 4. RT-PCR detection of ARV S4 gene expression. ARV S4 gene region
after footpad inoculation of 1-day-old broiler chicks with ARV S1133 strain. S
loaded from left to right in the consecutive order. The virus was first detected
the virus was widespread in all tissues tested. After that, the virus gradually di
hock joints after day 18 PI.
regulated genes in RA and osteoarthritis (OA) (Justen
et al., 2000), was employed in this study to explore any
cellular genes that may be involved in ARV-induced
arthritis. Each group (the ARV-infected or control
group) of cDNA was transcribed from pooled mRNAs,
containing total transcripts of footpads and hock joints
from 2–4, 6, 8, to 10 days post-inoculated chickens. By
SSH, we obtained PCR products that may reflect
upregulated genes of the arthritic joints in ARV-infected
chickens. The resultant PCR products were subcloned
into pCRII vector and sequenced. We identified two
(290 bp) was detected by RT-PCR in various tissues at different times
amples of 12 h, day 1–4, 6, 8, 10, 12, 14, 16, 18, 21, 28, and 35 PI were
at 12 h PI in hock joints, footpads, liver, and spleen. From day 2 to 6 PI,
minished in most tissues, and was exclusively confined to footpads and
Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–60 55
Fig. 5. Differential gene expression of MMP-2 and BMP-2 in ARV-infected chickens. Expressions of MMP-2 (A), and BMP-2 (B) in footpad and
hock joint of each chicken were determined by RT-PCR. The expected RT-PCR products of MMP-2 and BMP-2 were 381 and 512 bp, respectively.
Samples collected at 12 h, day 1–4, 6, 8, 10, 12, 14, 16, 18, 21, 28, and 35 PI were loaded from left to right in the consecutive order. MMP-2 mRNA
was detected at 12 h PI in the ARV-infected chickens, and at day 1 PI in the control chickens. BMP-2 mRNA was detected at 12 h PI in the ARV-
infected chickens, and at day 2 PI in the control chickens, respectively. Chicken b-actin mRNA was measured as a mRNA control. Representative
data of b-actin expression in the hock joint of the control chickens are shown.
gene sequences including chicken small 35-kDa subunit
of the U2 snRNP auxiliary factor (U2AF35 or U2AF1)
(Pacheco et al., 2004) and ferritin H chain protein
(Stevens et al., 1987). By RT-PCR, we further verified
the differential expression of U2AF35 in the joint tissues
of ARV-infected chickens. The expected RT-PCR
product of U2AF35 was 415 bp. U2AF35 mRNA was
induced in the arthritic footpads from 12 h to day 6 PI,
and in the arthritic hock joints from day 1 to 8 PI
(Fig. 6). Ferritin H chain mRNA, on the other hand, was
constitutively expressed in the joints of ARV-infected
and control chickens (data not shown).
Fig. 6. Induction of U2AF35 expression in ARV-infected chickens. Different
RT-PCR product of U2AF35 was 415 bp. Samples collected at 12 h, day 1–4,
order. U2AF35 mRNAwas not expressed in the control chickens throughout th
from 12 h to day 6 PI, and the in hock joints from day 1 to 8 PI in the ARV i
hock joint are shown.
3.7. Distribution of ARV, MMP-2, BMP-2, and
U2AF35 mRNAs in the joints of ARV-infected
chickens
ISH was performed to determine ARV distribution in
the joints of ARV-infected chickens. Using S1 gene of
ARVas a probe, we localized the viral expression in the
epidermal cells, fibroblast-like cells in the dermis, and
chondrocytes within/near the epiphyseal plate and near
the secondary center of ossification of ARV-infected
joints (Fig. 7). Viral S1 gene was first detected in
footpads at 12 h PI, later in hock joints at day 1 PI, and
ial gene expression of U2AF35 was verified by RT-PCR. The expected
6, 8, 10, 12, and 14 PI were loaded from left to right in the consecutive
e experimental period (A). U2AF35 mRNAwas induced in the footpads
nfected chickens (B). Representative data of b-actin expression in the
Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–6056
Fig. 7. Detection of ARV expression in chicken hock joints by in situ hybridization (ISH). Using ARV S1 DNA fragment as a probe, the viral
expression in the chicken hock joint was determined by ISH. (B) Viral expression was localized in epidermal cells and fibroblast-like cells in the
dermis of the ARV-infected chicken at day 3 PI. (D) Viral expression was localized in chondrocytes of the hock joint in the ARV-infected chicken at
day 3 PI. (A and C) ISH results from the control chicken are shown in parallel.
remained detectable to day 12 PI in both footpads and
hock joints. However, expression levels of viral S1 gene
gradually decreased from day 8, and the signal was
undetectable by ISH after day 14 PI.
Using RT-PCR products of MMP-2, BMP-2, and
U2AF35 as probes, cells expressing respective genes
were localized by ISH. MMP-2 mRNA was mainly
detected in chondrocytes within/near the epiphyseal
plate and near the secondary center of ossification
(Fig. 8B). Gene expressions of BMP-2 (Fig. 8D and F)
and U2AF35 (Fig. 9B and D) were detected in the
chondrocytes around the epiphyseal plate and second-
ary center of ossification, in epidermal cells, and in
dermal fibroblast-like cells of arthritic joints. In
addition, U2AF35 mRNA was expressed in some
inflammatory cells distributed in the bone marrow of
arthritic joints (Fig. 9F). The expression periods of
MMP-2, BMP-2 and U2AF35 observed by ISH (data not
shown) were equivalent to that detected by RT-PCR
(Figs. 5 and 6).
4. Discussion
In this study, we employed SSH technique and RT-
PCR to identify the upregulation of U2AF35 mRNA in
the footpads and hock joints of arthrotropic ARV-
infected chickens. Our RT-PCR results illustrated that,
in contrast to the continuous presence of ARV in the
joint throughout the experimental period (Fig. 4),
U2AF35 mRNA was only expressed during arthritic
manifestation period (Fig. 6) when tissues were
infiltrated with inflammatory cells. U2AF35 mRNA,
as determined by ISH, coincided well with ARV signal
in epidermal cells, dermal fibroblast-like cells, and
chondrocytes around the epiphyseal plate and second-
ary center of ossification. U2AF35 is the small 35-kDa
subunit (also known as U2AF1) of U2AF splicing
factor. It interacts with U2AF65, the large 65-kDa
subunit (also known as U2AF2) of U2AF, and binds to
the AG dinucleotide at the 30 splice site (Guth et al.,
2001) of pre-mRNAs. While binding of U2AF65 to the
pyrimidine (Py) tract is a prerequisite for assisting U2
snRNP to catalyze splicing reaction, U2AF35 is only
essential for splicing of a subset of AG-dependent pre-
mRNAs (Guth et al., 2001; Zamore and Green, 1991).
Remarkably, a recent study on Shigella infection has
revealed the involvement of U2AF35 in the expression
of some proinflammatory chemokines and cytokines,
including interleukin-8 (IL-8), RANTES (regulated on
activation, normal T cell expressed and secreted),
granulocyte-macrophage colony-stimulating factor
(GM-CSF), and interleukin-1b (IL-1b) (Okuda et al.,
2005). Based on this activity, it is feasible that U2AF35
induced in ARV-infected cells (epidermal cells, dermal
Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–60 57
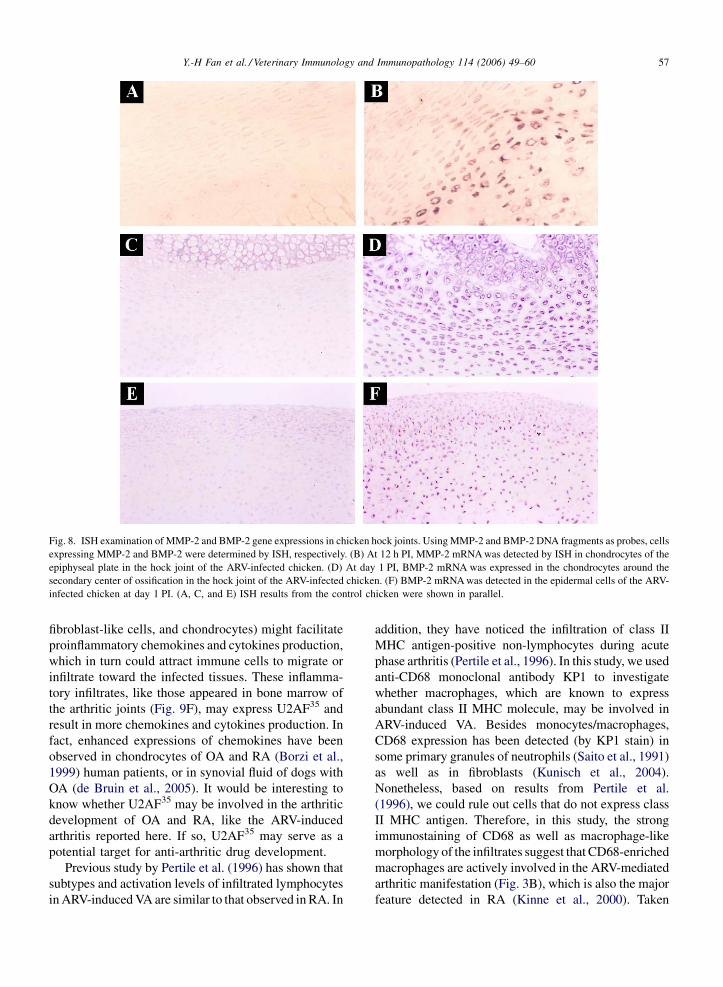
Fig. 8. ISH examination of MMP-2 and BMP-2 gene expressions in chicken hock joints. Using MMP-2 and BMP-2 DNA fragments as probes, cells
expressing MMP-2 and BMP-2 were determined by ISH, respectively. (B) At 12 h PI, MMP-2 mRNA was detected by ISH in chondrocytes of the
epiphyseal plate in the hock joint of the ARV-infected chicken. (D) At day 1 PI, BMP-2 mRNA was expressed in the chondrocytes around the
secondary center of ossification in the hock joint of the ARV-infected chicken. (F) BMP-2 mRNA was detected in the epidermal cells of the ARV-
infected chicken at day 1 PI. (A, C, and E) ISH results from the control chicken were shown in parallel.
fibroblast-like cells, and chondrocytes) might facilitate
proinflammatory chemokines and cytokines production,
which in turn could attract immune cells to migrate or
infiltrate toward the infected tissues. These inflamma-
tory infiltrates, like those appeared in bone marrow of
the arthritic joints (Fig. 9F), may express U2AF35 and
result in more chemokines and cytokines production. In
fact, enhanced expressions of chemokines have been
observed in chondrocytes of OA and RA (Borzi et al.,
1999) human patients, or in synovial fluid of dogs with
OA (de Bruin et al., 2005). It would be interesting to
know whether U2AF35 may be involved in the arthritic
development of OA and RA, like the ARV-induced
arthritis reported here. If so, U2AF35 may serve as a
potential target for anti-arthritic drug development.
Previous study by Pertile et al. (1996) has shown that
subtypes and activation levels of infiltrated lymphocytes
in ARV-induced VA are similar to that observed in RA. In
addition, they have noticed the infiltration of class II
MHC antigen-positive non-lymphocytes during acute
phase arthritis (Pertile et al., 1996). In this study, we used
anti-CD68 monoclonal antibody KP1 to investigate
whether macrophages, which are known to express
abundant class II MHC molecule, may be involved in
ARV-induced VA. Besides monocytes/macrophages,
CD68 expression has been detected (by KP1 stain) in
some primary granules of neutrophils (Saito et al., 1991)
as well as in fibroblasts (Kunisch et al., 2004).
Nonetheless, based on results from Pertile et al.
(1996), we could rule out cells that do not express class
II MHC antigen. Therefore, in this study, the strong
immunostaining of CD68 as well as macrophage-like
morphology of the infiltrates suggest that CD68-enriched
macrophages are actively involved in the ARV-mediated
arthritic manifestation (Fig. 3B), which is also the major
feature detected in RA (Kinne et al., 2000). Taken
Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–6058
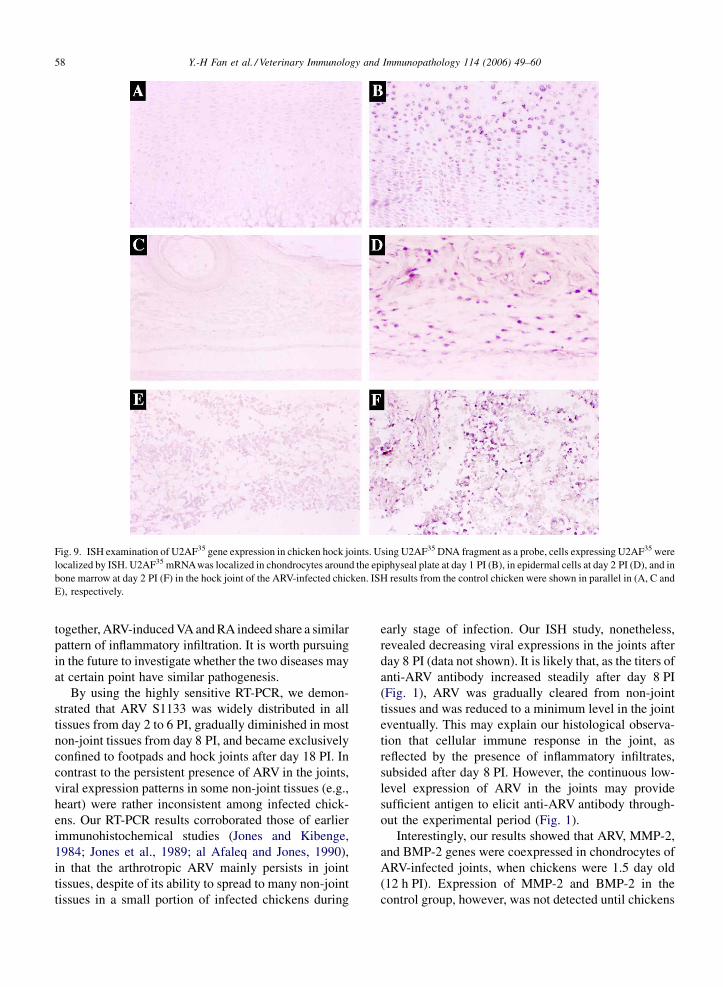
Fig. 9. ISH examination of U2AF35 gene expression in chicken hock joints. Using U2AF35 DNA fragment as a probe, cells expressing U2AF35 were
localized by ISH. U2AF35 mRNAwas localized in chondrocytes around the epiphyseal plate at day 1 PI (B), in epidermal cells at day 2 PI (D), and in
bone marrow at day 2 PI (F) in the hock joint of the ARV-infected chicken. ISH results from the control chicken were shown in parallel in (A, C and
E), respectively.
together, ARV-induced VA and RA indeed share a similar
pattern of inflammatory infiltration. It is worth pursuing
in the future to investigate whether the two diseases may
at certain point have similar pathogenesis.
By using the highly sensitive RT-PCR, we demon-
strated that ARV S1133 was widely distributed in all
tissues from day 2 to 6 PI, gradually diminished in most
non-joint tissues from day 8 PI, and became exclusively
confined to footpads and hock joints after day 18 PI. In
contrast to the persistent presence of ARV in the joints,
viral expression patterns in some non-joint tissues (e.g.,
heart) were rather inconsistent among infected chick-
ens. Our RT-PCR results corroborated those of earlier
immunohistochemical studies (Jones and Kibenge,
1984; Jones et al., 1989; al Afaleq and Jones, 1990),
in that the arthrotropic ARV mainly persists in joint
tissues, despite of its ability to spread to many non-joint
tissues in a small portion of infected chickens during
early stage of infection. Our ISH study, nonetheless,
revealed decreasing viral expressions in the joints after
day 8 PI (data not shown). It is likely that, as the titers of
anti-ARV antibody increased steadily after day 8 PI
(Fig. 1), ARV was gradually cleared from non-joint
tissues and was reduced to a minimum level in the joint
eventually. This may explain our histological observa-
tion that cellular immune response in the joint, as
reflected by the presence of inflammatory infiltrates,
subsided after day 8 PI. However, the continuous low-
level expression of ARV in the joints may provide
sufficient antigen to elicit anti-ARV antibody through-
out the experimental period (Fig. 1).
Interestingly, our results showed that ARV, MMP-2,
and BMP-2 genes were coexpressed in chondrocytes of
ARV-infected joints, when chickens were 1.5 day old
(12 h PI). Expression of MMP-2 and BMP-2 in the
control group, however, was not detected until chickens

Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–60 59
were 2-day old and 3-day old, respectively. Based on the
collagenase activity of chicken MMP-2 (Aimes and
Quigley, 1995), the earlier induction of MMP-2 is likely
to cause matrix damage around the viral-infected
chondrocytes of 1-day-old chicks. Conversely, chicken
BMP-2 is capable of promoting cartilage extracellular
matrix production (Roark and Greer, 1994). Similar to
human arthritis (Nakase et al., 2003), BMP-2 may be
induced here to counterbalance MMP activity and to
repair damage. MMP family members are enzymes that
decompose extracellular matrix (Nagase and Woessner,
1999). Among many MMP members that have been
detected in the RA synovium, MMP-1, MMP-3, and
MMP-13 are particularly important for joint destruction
(Hegemann et al., 2003; Vincenti and Brinckerhoff,
2002; Westhoff et al., 1999). On the other hand, BMPs
are members of the TGF-beta (TGF-b) superfamily.
Apart from playing important roles in almost all
processes of skeletal morphogenesis (Wan and Cao,
2005), some BMPs, such as BMP-2, BMP-6, and BMP-
7, have been identified in arthritic tissues as well (Gortz
et al., 2004; Nakase et al., 2003). These BMPs are
thought to be involved in cartilage and bone repair in
human arthritis (Nakase et al., 2003). Therefore, it is
worth pursuing in the future to investigate whether
chicken MMP-1, MMP-3, MMP-13, BMP-6, BMP-7, or
any chicken homologs of other MMP and BMP
members may participate in ARV-induced arthritis.
In conclusions, among U2AF35, MMP-2, and BMP-2
that were differentially expressed in the joint of ARV-
infected chickens, only U2AF35 expression correlated
well with the arthritic development. Because U2AF35
may assist in the mRNA splicing of some proinflam-
matory chemokines and cytokines, we propose that
U2AF35 induction may play an immunopathological
role in ARV-induced arthritis. This study has first
associated U2AF35 to viral arthritis.
Acknowledgments
We would like to thank Mr. Chun-Hao Lin for his
technical assistance. This study was supported in part by
grants from the National Science Council (NSC87-
2313-B-005-070) and Council of Agriculture, Execu-
tive Yuan (88-AS-2.2-BQ-01-11 and 89-AS-2.2-BQ-
62), Taiwan, ROC.
References
Aimes, R.T., French, D.L., Quigley, J.P., 1994. Cloning of a 72 kDa
matrix metalloproteinase (gelatinase) from chicken embryo fibro-
blasts using gene family PCR: expression of the gelatinase
increases upon malignant transformation. Biochem. J. 300,
729–736.
Aimes, R.T., Quigley, J.P., 1995. Matrix metalloproteinase-2 is an
interstitial collagenase. J. Biol. Chem. 270, 5872–5876.
al Afaleq, A.I., Jones, R.C., 1990. Localisation of avian reovirus in the
hock joints of chicks after entry through broken skin. Res. Vet. Sci.
48, 381–382.
Borzi, R.M., Mazzetti, I., Macor, S., Silvestri, T., Bassi, A., Cattini, L.,
Facchini, A., 1999. Flow cytometric analysis of intracellular
chemokines in chondrocytes in vivo: constitutive expression
and enhancement in osteoarthritis and rheumatoid arthritis. FEBS
Lett. 455, 238–242.
Chiou, S.-H., Chow, K.-C., Yang, C.-H., Chiang, S.-F., Lin, C.-H.,
2005. Discovery of Epstein-Barr virus (EBV)-encoded RNA
signal and EBV nuclear antigen leader protein DNA sequence
in pet dogs. J. Gen. Virol. 86, 899–905.
Chiu, C.J., Lee, L.H., 1997. Cloning and nucleotide sequencing of the
S4 genome of avian reovirus S1133. Arch. Virol. 142, 2515–2520.
Chow, K.-C., Hsiao, C.-H., Lin, T.-Y., Chen, C.-L., Chiou, S.-H., 2004.
Detection of severe acute respiratory syndrome-associated corona-
virus in pneumocytes of the lung. Am. J. Clin. Pathol. 121, 574–580.
Clark, F.D., Ni, Y., Collisson, E.W., 1990. Characterization of avian
reovirus strain-specific polymorphisms. Avian Dis. 34, 304–314.
de Bruin, T., de Rooster, H., van Bree, H., Cox, E., 2005. Interleukin-8
mRNA expression in synovial fluid of canine stifle joints with
osteoarthritis. Vet. Immunol. Immunopathol. 108, 387–397.
Diatchenko, L., Lau, Y.F., Campbell, A.P., Chenchik, A., Moqadam,
F., Huang, B., Lukyanov, S., Lukyanov, K., Gurskaya, N., Sver-
dlov, E.D., Siebert, P.D., 1996. Suppression subtractive hybridiza-
tion: a method for generating differentially regulated or tissue-
specific cDNA probes and libraries. Proc. Natl. Acad. Sci. U.S.A.
93, 6025–6030.
Francis, P.H., Richardson, M.K., Brickell, P.M., Tickle, C., 1994.
Bone morphogenetic proteins and a signalling pathway that con-
trols patterning in the developing chick limb. Development 120,
209–218.
Giannelli, G., Erriquez, R., Iannone, F., Marinosci, F., Lapadula, G.,
Antonaci, S., 2004. MMP-2, MMP-9, TIMP-1 and TIMP-2 levels
in patients with rheumatoid arthritis and psoriatic arthritis. Clin.
Exp. Rheumatol. 22, 335–338.
Glass, S.E., Naqi, S.A., Hall, C.F., Kerr, K.M., 1973. Isolation and
characterization of a virus associated with arthritis of chickens.
Avian Dis. 17, 415–424.
Gortz, B., Hayer, S., Redlich, K., Zwerina, J., Tohidast-Akrad, M.,
Tuerk, B., Hartmann, C., Kollias, G., Steiner, G., Smolen, J.S.,
Schett, G., 2004. Arthritis induces lymphocytic bone marrow
inflammation and endosteal bone formation. J. Bone Miner.
Res. 19, 990–998.
Guth, S., Tange, T.O., Kellenberger, E., Valcarcel, J., 2001. Dual
function for U2AF(35) in AG-dependent pre-mRNA splicing.
Mol. Cell. Biol. 21, 7673–7681.
Harris, E.D., 1990. Rheumatoid arthritis: pathophysiology and impli-
cations for therapy. N. Engl. J. Med. 322, 1277–1289.
Hegemann, N., Wondimu, A., Ullrich, K., Schmidt, M.F., 2003.
Synovial MMP-3 and TIMP-1 levels and their correlation with
cytokine expression in canine rheumatoid arthritis. Vet. Immunol.
Immunopathol. 91, 199–204.
Jones, R.C., Islam, M.R., Kelly, D.F., 1989. Early pathogenesis of
experimental reovirus infection in chickens. Avian Pathol. 18,
239–253.
Jones, R.C., Kibenge, F.S.B., 1984. Reovirus-induced tenosynovitis in
chickens: the effect of breed. Avian Pathol. 13, 511–528.

Y.-H Fan et al. / Veterinary Immunology and Immunopathology 114 (2006) 49–6060
Justen, H.P., Grunewald, E., Totzke, G., Gouni-Berthold, I., Sachinidis,
A., Wessinghage, D., Vetter, H., Schulze-Osthoff, K., Ko, Y., 2000.
Differential gene expression in synovium of rheumatoid arthritis and
osteoarthritis. Mol. Cell Biol. Res. Commun. 3, 165–172.
Kerr, K.M., Olson, N.O., 1969. Pathology of chickens experimentally
inoculated or contact-infected with an arthritis-producing virus.
Avian Dis. 13, 729–745.
Kibenge, F.S.B., Robertson, M.D., Wilcox, G.E., 1982. Staphylococ-
cus aureus isolated from poultry in Australia. II. Epidemiology of
strains associated with tenosynovitis. Vet. Microbiol. 7, 485–491.
Kinne, R.W., Brauer, R., Stuhlmuller, B., Palombo-Kinne, E.,
Burmester, G.R., 2000. Macrophages in rheumatoid arthritis.
Arthritis Res. 2, 189–202.
Kost, T.A., Theodorakis, N., Hughes, S.H., 1983. The nucleotide
sequence of the chick cytoplasmic beta-actin gene. Nucleic Acids
Res. 11, 8287–8301.
Kunisch, K., Fuhrmann, R., Roth, A., Winter, R., Lungershausen, W.,
Kinne, R.W., 2004. Macrophage specificity of three anti-CD68
monoclonal antibodies (KP1, EBM11, and PGM1) widely used for
immunohistochemistry and flow cytometry. Ann. Rheum. Dis. 63,
774–784.
Montgomery, R.D., Villegas, P., Dowe, D.L., Brown, J., 1985. Effect of
avian reovirus on lymphoid organ weights. Avian Dis. 29, 552–560.
Nagase, H., Woessner Jr., J.F., 1999. Matrix metalloproteinases. J.
Biol. Chem. 274, 21491–21494.
Nakase, T., Miyaji, T., Tomita, T., Kaneko, M., Kuriyama, K., Myoui,
A., Sugamoto, K., Ochi, T., Yoshikawa, H., 2003. Localization of
bone morphogenetic protein-2 in human osteoarthritic cartilage
and osteophyte. Osteoarthritis Cartilage 11, 278–284.
Neelima, S., Ram, G.C., Kataria, J.M., Goswami, T.K., 2003. Avian
reovirus induces an inhibitory effect on lymphoproliferation in
chickens. Vet. Res. Commun. 27, 73–85.
Okuda, J., Toyotome, T., Kataoka, N., Ohno, M., Abe, H., Shimura, Y.,
Seyedarabi, A., Pickersgill, R., Sasakawa, C., 2005. Shigella
effector IpaH9.8 binds to a splicing factor U2AF(35) to modulate
host immune responses. Biochem. Biophys. Res. Commun. 333,
531–539.
Pacheco, T.R., Gomes, A.Q., Barbosa-Morais, N.L., Benes, V.,
Ansorge, W., Wollerton, M., Smith, C.W., Valcarcel, J., Carmo-
Fonseca, M., 2004. Diversity of vertebrate splicing factor U2AF35:
identification of alternatively spliced U2AF1 mRNAs. J. Biol.
Chem. 279, 27039–27049.
Peng, C.T., Chow, K.C., Chang, W.C., Tsai, C.H., Lin, T.Y., Lin, S.S.,
Chiu, C.F., 1999. Expression of Fas ligand in Langerhans’ cell
histiocytosis: A case report of a boy with multisystem involve-
ment. Am. J. Hematol. 61, 256–261.
Pertile, T.L., Sharma, J.M., Walser, M.M., 1995. Reovirus infection in
chickens primes splenic adherent macrophages to produce nitric
oxide in response to T cell-produced factors. Cell. Immunol. 164,
207–216.
Pertile, T.L., Walser, M.M., Sharma, J.M., Shivers, J.L., 1996. Immu-
nohistochemical detection of lymphocyte subpopulations in the
tarsal joints of chickens with experimental viral arthritis. Vet.
Pathol. 33, 303–310.
Roark, E.F., Greer, K., 1994. Transforming growth factor-beta and
bone morphogenetic protein-2 act by distinct mechanisms to
promote chick limb cartilage differentiation in vitro. Dev. Dyn.
200, 103–116.
Robertson, M.D., Wilcox, G.E., 1984. Serological characteristics of
avian reoviruses of Australian origin. Avian Pathol. 13, 585–594.
Robertson, M.D., Wilcox, G.E., 1986. Avian reovirus. Vet. Bull. 56,
726–733.
Rosenberger, J.K., Olson, N.O., 1991. Reovirus infections. Dis. Poult.
9, 639–674.
Saito, N., Pulford, K.A., Breton-Gorius, J., Masse, J.M., Mason, D.Y.,
Cramer, E.M., 1991. Ultrastructural localization of the CD68
macrophage-associated antigen in human blood neutrophils and
monocytes. Am. J. Pathol. 139, 1053–1059.
Shapouri, M.R.S., Kane, M., Letarte, M., Bergeron, J., Arella, M.,
Silim, A., 1995. Cloning, sequencing, and expression of the S1
gene of avian reovirus. J. Gen. Virol. 76, 1515–1520.
Springer, W.T., Olson, N.O., Kerr, K.M., Fabacher, C.J., 1983.
Response of specific pathogen free chicks to concomitant infec-
tion of reovirus (WVU 2937) and infectious bursal disease virus.
Avian Dis. 27, 911–917.
Stevens, P.W., Dodgson, J.B., Engel, J.D., 1987. Structure and expres-
sion of the chicken ferritin H-subunit gene. Mol. Cell. Biol. 7,
1751–1758.
van der Heide, L., Kalbac, M., 1975. Infectious tenosynovitis (viral
arthritis): characterization of a Connecticut viral isolant as a
reovirus and evidence of viral egg transmission by reovirus-
infected broiler breeders. Avian Dis. 19, 683–688.
van der Heide, L., Lutticken, D., Horzinek, M., 1981. Isolation of
avian reovirus as a possible etiologic agent of osteoporosis
(‘‘brittle bone disease’’; ‘‘femoral head necrosis’’) in broiler
chickens. Avian Dis. 25, 847–856.
Verwilghen, J., Vertessen, S., Stevens, E.A., Dequeker, J., Ceuppens,
J.L., 1990. Depressed T-cell reactivity to recall antigens in rheu-
matoid arthritis. J. Clin. Immunol. 10, 90–98.
Vincenti, M.P., Brinckerhoff, C.E., 2002. Transcriptional regulation of
collagenase (MMP-1, MMP-13) genes in arthritis: integration of
complex signalling pathways for the recruitment of gene-specific
transcription factors. Arthritis Res. 4, 157–164.
Walker, E.R., Friedman, M.H., Olson, N.O., 1972. Electron micro-
scopic study of an avian reovirus that causes arthritis. J. Ultra-
struct. Res. 41, 67–79.
Wan, M., Cao, X., 2005. BMP signaling in skeletal development.
Biochem. Biophys. Res. Commun. 328, 651–657.
Westhoff, C.S., Freudiger, D., Petrow, P., Seyfert, C., Zacher, J.,
Kriegsmann, J., Pap, T., Gay, S., Stiehl, P., Gromnica-Ihle, E.,
Wernicke, D., 1999. Characterization of collagenase 3 (matrix
metalloproteinase 13) messenger RNA expression in the synovial
membrane and synovial fibroblasts of patients with rheumatoid
arthritis. Arthritis Rheum. 42, 1517–1527.
Zamore, P.D., Green, M.R., 1991. Biochemical characterization
of U2 snRNP auxiliary factor: an essential pre-mRNA splicing
factor with a novel intranuclear distribution. EMBO J. 10, 207–
214.
Related Documents











