1998, 18(2):694. Mol. Cell. Biol. Aberdam, Jean-Paul Ortonne and Robert Ballotti Corine Bertolotto, Roser Buscà, Patricia Abbe, Karine Bille, Edith Boxes (GTCATGTGCT) and of Microphthalmia Activities by Cyclic AMP: Pivotal Role of M the Regulation of TRP1 and TRP2 Promoter -Acting Elements Are Involved in cis Different http://mcb.asm.org/content/18/2/694 Updated information and services can be found at: These include: REFERENCES http://mcb.asm.org/content/18/2/694#ref-list-1 This article cites 37 articles, 19 of which can be accessed free at: CONTENT ALERTS more» cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new articles http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on November 1, 2014 by guest http://mcb.asm.org/ Downloaded from on November 1, 2014 by guest http://mcb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1998, 18(2):694. Mol. Cell. Biol.
Aberdam, Jean-Paul Ortonne and Robert BallottiCorine Bertolotto, Roser Buscà, Patricia Abbe, Karine Bille, Edith Boxes (GTCATGTGCT) and of MicrophthalmiaActivities by Cyclic AMP: Pivotal Role of Mthe Regulation of TRP1 and TRP2 Promoter
-Acting Elements Are Involved incisDifferent
http://mcb.asm.org/content/18/2/694Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/18/2/694#ref-list-1This article cites 37 articles, 19 of which can be accessed free at:
CONTENT ALERTS more»cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new articles
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY,0270-7306/98/$04.0010
Feb. 1998, p. 694–702 Vol. 18, No. 2
Copyright © 1998, American Society for Microbiology
Different cis-Acting Elements Are Involved in the Regulation ofTRP1 and TRP2 Promoter Activities by Cyclic AMP: PivotalRole of M Boxes (GTCATGTGCT) and of MicrophthalmiaCORINE BERTOLOTTO, ROSER BUSCA, PATRICIA ABBE, KARINE BILLE, EDITH ABERDAM,
JEAN-PAUL ORTONNE, AND ROBERT BALLOTTI*
INSERM U385, Biologie et Physiopathologie de la Peau, Faculte de Medecine, 06107 Nice Cedex 2, France
Received 27 May 1997/Returned for modification 31 July 1997/Accepted 10 November 1997
In melanocytes and in melanoma cells, cyclic AMP (cAMP)-elevating agents stimulate melanogenesis andincrease the transcription of tyrosinase, the rate-limiting enzyme in melanin synthesis. However, two otherenzymes, tyrosinase-related protein 1 (TRP1) and TRP2, are required for a normal melanization processleading to eumelanin synthesis. In B16 melanoma cells, we demonstrated that stimulation of melanogenesis bycAMP-elevating agents results in an increase in tyrosinase, TRP1, and TRP2 expression. cAMP, through acAMP-dependent protein kinase pathway, stimulates TRP1 and TRP2 promoter activities in both B16 mousemelanoma cells and normal human melanocytes. Regulation of the TRP1 and TRP2 promoters by cAMPinvolves a M box and an E box. Further, a classical cAMP response element-like motif participates in the cAMPresponsiveness of the TRP2 promoter, demonstrating that the TRP2 gene is subjected to different regulatoryprocesses, which could account for its different expression patterns during embryonic development or underspecific physiological and pathological conditions. We also found that microphthalmia, a basic helix-loop-helixtranscription factor, strongly stimulates the transcriptional activities of the TRP1 and TRP2 promoters,mainly through binding to the M boxes. Additionally, we demonstrated that cAMP increases microphthalmiaexpression and thereby its binding to TRP1 and TRP2 M boxes. These convergent and compelling resultsdisclose at least a part of the molecular mechanism involved in the regulation of melanogenic gene expressionby cAMP and emphasize the pivotal role of microphthalmia in this process.
In mammals, pigmentation results from the synthesis anddistribution of melanin in the skin, hair bulbs, and eyes. Mel-anin synthesis (melanogenesis) takes place in the melanocyteafter differentiation of the nonpigmented precursor, the mel-anoblast (27). Three melanocyte-specific enzymes, tyrosinase,tyrosinase-related protein 1 (TRP1), and TRP2, are involvedin this enzymatic process that converts tyrosine to melaninpigments. Although these proteins have similar structures andfeatures, they are expressed by different genes and possessdistinct enzymatic activities. Tyrosinase, encoded by the albinolocus of the mouse, catalyzes the conversion of tyrosine to3,4-dihydroxyphenylalanine (DOPA) and of DOPA to DOPAquinone (14, 25, 31). TRP2, encoded by the mouse slaty locus,possesses a Dopachrome tautomerase activity, converting theDopachrome to 5,6-dihydroxyindole-2-carboxylic acid (DHICA)(3, 19, 42). TRP1, which has been mapped in mouse to thebrown locus, catalyzes the oxidation of DHICA to indole-5,6-quinone-2-carboxylic acid (21, 24).
In vivo, melanogenesis is regulated by UVB radiation thatcan act either directly on melanocytes or indirectly through therelease of keratinocyte-derived factors such as interleukins,prostaglandins, and alpha melanocyte-stimulating hormone(a-MSH) (1, 12, 22, 35). Interestingly, a-MSH, one of the mostpotent activators of melanogenesis, binds to an as-coupledreceptor and increases the intracellular level of cyclic AMP(cAMP) (9, 17, 20, 37). Further, the melanogenic effects ofa-MSH can be mimicked by pharmacological cAMP-elevatingagents such as forskolin, cholera toxin, and isobutylmethylxan-
thine (8, 13, 15, 18, 38), indicating that the cAMP pathwayplays a pivotal role in the regulation of melanogenesis.
It has been thought for many years that the regulation ofmelanin synthesis occurs at the level of tyrosinase, which is therate-limiting enzyme in melanogenesis. However, TRP1 andTRP2 have been recently shown to play an important role inthe control of melanin type (23). Indeed, two types of melaninare produced by melanocytes: pheomelanins, which are red oryellow, and eumelanins, which are brown or black (32). Thelatter pigments can absorb UV photons and scavenge damag-ing free radicals generated by UV within the cells, thus pre-venting DNA damage and sheltering the skin from the harmfuleffects of UV radiation. TRP1 and TRP2 control distal steps ofeumelanin synthesis, thereby fulfilling a key photoprotectivefunction. Noteworthy, the stimulation of melanogenesis bycAMP-elevating agents leads to an increased eumelanin syn-thesis, suggesting that cAMP regulates TRP1 and TRP2 activ-ity and/or expression. However, the regulation of TRP1 andTRP2 expression by cAMP has not been clearly demonstratedand remains controversial. Interestingly, TRP2 is expressedbefore tyrosinase and TRP1 during embryogenesis (34). Fur-ther, in agouti mice that synthesize only pheomelanins, extinc-tion of TRP1 and TRP2 expression has been reported, whiletyrosinase is still expressed (23). These observations suggestthat different mechanisms are involved in the regulation oftyrosinase, TRP1, and TRP2 gene expression. Tyrosinase andTRP1 promoters share an 11-bp motif (AGTCATGTGCT)termed the M box located upstream of the TATA box. Thismotif binds microphthalmia, a basic helix-loop-helix transcrip-tion factor that increases tyrosinase and TRP1 promoter ac-tivities, thereby playing a key role in the tissue-specific expres-sion of these genes (11, 29, 40). In the TRP2 promoter, ahomologous sequence (GTCATGTGCT) is also found up-
* Corresponding author. Mailing address: INSERM U385, Biologieet Physiopathologie de la Peau, Faculte de Medecine, Ave. de Valom-brose, 06107 Nice Cedex 2, France. Phone: 33-49337-7790. Fax: 33-49381-1404. E-mail: [email protected].
694
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

stream of the TATA box (41). However, it has not been clearlyestablished whether microphthalmia binds to and stimulatesthe TRP2 promoter.
Since a precise control of melanogenic gene expression iscrucial for normal melanization, it is essential to study themolecular mechanisms involved in the control of TRP1 andTRP2 gene expression by cAMP. In this study, we demon-strated that cAMP increases the level of TRP1 and TRP2mRNA. Then, using reporter constructs containing either the1.1-kb fragment 59 of the transcriptional start site of the TRP1gene or the 0.6-kb fragment 59 of the transcriptional start siteof the TRP2 gene, we demonstrated that cAMP-elevatingagents, through the cAMP-dependent protein kinase (PKA)pathway, stimulate the transcriptional activity of the TRP1 andTRP2 promoters. Deletion and mutation analysis allowed us tolocalize the cis-acting elements involved in the cAMP respon-siveness of TRP1 and TRP2 promoters. Our data indicate thatthe M box (GTCATGTGCT) plays a key role in the regulationof TRP1 and TRP2 expression by cAMP. However, we iden-tified in the TRP1 and TRP2 promoters other cis-acting ele-ments involved in the cAMP response, indicating that specificregulatory mechanisms participate in the regulation of TRP1and TRP2 expression by cAMP. Finally, we showed that mi-crophthalmia binds to the M boxes and transactivates theTRP1 and TRP2 promoters. Further, we observed that cAMPincreases microphthalmia expression, thus demonstrating thepivotal role of microphthalmia in the regulation of melano-genic gene expression by cAMP.
MATERIALS AND METHODS
Materials. MCDB 153, forskolin, bovine serum albumin, hydrocortisone, in-sulin, phorbol 12-myristate 13-acetate, p-nitrophenyl phosphate (PNPP), 4-(2-aminoethyl)benzenesulfonyl fluoride (AEBSF), aprotinin, and leupeptin werepurchased from Sigma Chemical Co. Dispase was from Boehringer, and basicfibroblast growth factor and the basic vector PGL2 were from Promega. Dulbec-co’s modified Eagle’s medium, fetal calf serum (FCS), trypsin, and Lipo-fectamine reagent were from GIBCO. Peroxidase-conjugated anti-rabbit andanti-mouse antibodies were from Dakopatts. Synthetic oligonucleotides werefrom Oligo Express. Expression vectors encoding the catalytic subunit of PKAand the PKA peptide inhibitor (PKI) were previously described (6, 30).
Cell cultures. B16-F10 murine melanoma cells and NIH 3T3 fibroblast cellswere grown at 37°C under 5% CO2 in Dulbecco’s modified Eagle’s mediumsupplemented with 10% FCS, penicillin (100 U/ml), and streptomycin (50 mg/ml). Epidermal cell suspensions were obtained from foreskins of caucasoidchildren by overnight digestion in phosphate-buffered saline containing 0.5%dispase grade II at 4°C, followed by a 1-h digestion with 0.05% trypsin–0.02%EDTA in phosphate-buffered saline at 37°C. Cells were grown in MCDB 153medium supplemented with 2% FCS, hydrocortisone (0.4 mg/ml), insulin (5mg/ml), 16 nM phorbol 12-myristate 13-acetate, basic fibroblast growth factor (1ng/ml), penicillin (100 U/ml), and streptomycin (50 mg/ml) in a humidifiedatmosphere containing 5% CO2 in air at 37°C.
Western blot assays. B16 mouse melanoma cells were grown in six-well disheswith or without 20 mM forskolin. After 24 or 48 h of cAMP treatment, cells werelysed in phosphate buffer (pH 6.8) containing 1% (wt/vol) Triton X-100, leupep-tin (5 mg/ml), 1 mM AEBSF, and aprotinin (100 IU/ml). Proteins (30 mg) wereseparated on sodium dodecyl sulfate (SDS)–7.5% polyacrylamide gels and trans-ferred to a nitrocellulose membrane. Tyrosinase, TRP2, and ERK1 proteins weredetected with polyclonal antibodies PEP7, PEP8 (23), and C-16, respectively, ata 1/3,000 dilution in saturation buffer and with a secondary peroxidase-conju-gated anti-rabbit antibody at a 1/3,000 dilution. TRP1 protein was detected withmonoclonal antibody B8G3 (36) at a 1/1,000 dilution in saturation buffer andwith a secondary peroxidase-conjugated anti-mouse antibody at a 1/3,000 dilu-tion. Proteins were visualized with the Amersham ECL system. Anti-ERK1antibody C-16 was from Santa Cruz Biotechnology (Santa Cruz, Calif.).
RNA isolation and reverse transcription-PCR assays. Total cellular RNA wasextracted from control and cAMP-treated cells by a modification of the methodof Chomczynski. Ten micrograms of RNA was reverse transcribed by using thePromega reverse transcription system. Twenty-eight cycles of PCR amplificationof the resulting cDNA allowed us to quantify tyrosinase, TRP1, and TRP2mRNAs (53°C, 45 s; 72°C, 1 min; 94°C, 30 s). The 1,191-bp tyrosinase fragmentwas amplified with primers 59-CATTTTTGATTTGAGTGTCT-39 and 59-TGTGGTAGTCGTCTTTGTCC-39, the 784-bp TRP1 product was amplified withprimers 59-CTTTCTCCCTTCCTTACTGG-39 and 59-TGGCTTCATTCTTGGTGCTT-39, and the 518-bp TRP2 fragment was amplified with primers 59-TGA
GAAGAAACAAAGTAGGCAGAA-39 and 59-CAACCCCAAGAGCAAGACGAAAGC-39. Specific primers for glyceraldehyde-3-phosphate dehydrogenase(GAPDH) were from Clontech and gave an amplified PCR product of 983 bp.Preliminary trials showed that after 28 cycles of PCR, the reaction remainedexponential. The PCR products were electrophoresed on 1% agarose gels andstained with ethidium bromide before visualization under UV light.
Construction of the reporter plasmids. We have previously described thereporter plasmid containing the 2.2-kb fragment of the mouse tyrosinase pro-moter (pTyro; 22236 to 159) (5). A 1.1-kb fragment 59 of the transcriptionalstart site of the mouse TRP1 gene and a 0.6-kb fragment 59 of the transcriptionalstart site of the human TRP2 gene were isolated from genomic DNA by PCRusing primers 59-TGAAGCCACAGAGAATAAGG-39 (21247 to 21228) and59-CCAGACAGTAAATCCCAAGC-39 (182 to 163) for TRP1 and primers59-GGTTCCAGTGCCTTCCATAC-39 (2585 to 2566) and 59-TTTCAGTCTTTTCTTTTCAG-39 (1359 to 1340) for TRP2. Promoter fragments were clonedinto the unique SmaI-BglII sites of the PGL2-basic vector (PGL2B) upstream ofthe luciferase coding sequence (pTRP1 [21247 to 182] and pTRP2 [2585 to1359]). All deletions and mutations of TRP1 and TRP2 were constructed witha Transformer site-directed mutagenesis kit (Clontech) and were verified byplasmid sequencing. Briefly, the deletions of the TRP1 and TRP2 promoterswere obtained with primers selected to hybridize to the multiple cloning sites ofPGL2B in the 59 half and to the 59 end of the selected deletion (indicated inboldface) in the 39 half, resulting in the following constructions: D1pTRP1(59-GGTACTGTAACTGAGCCTTAAGACTTTAACC-39; 283 to 182),D2pTRP1 (59-GGTACTGTAACTGAGCGTTGGGGCAGGGGGG-39; 2864 to182), D3pTRP1 (59-CTGTAACTGAGCTAACATAACAGGCATCTTATATCAAGCA-39; 2608 to 182), D4pTRP1 (59-GTACTGTAACTGAGCTAACGTAGAGTAATCATGTATTC-39; 2363 to 182), and D5pTRP1 (59-GGTACTGTAACTGAGCTAACGAGTTTTCAACTTCCAGGAG-39; 2304 to 182) for the TRP1promoter and D1pTRP2 (59-ACTGAGCTAACATAAACTTTGGGTCATGTG-39; 2144 to 1349), D2pTRP2 (59-GGTACTGTAACTGAGCTAACGAGTAAGTTATTATTTGGAG-39; 2475 to 1349), D3pTRP2 (59-CTGTAACTGAGCTAACGAGCTCACTGCATC-39; 2273 to 1349), and D4pTRP2 (59-CTGTAACTGAGCTAACATAAGGAGCACATGAGCCCAGATA-39; 2220 to 1349) for theTRP2 promoter. Mutations were introduced with primers carrying point muta-tions in the core motif of target sequences. In the TRP1 promoter, the M box(GTCATGTGCT) located between bp 244 and 233 upstream from the initia-tion start site was mutated in GTCGGATCCT (59-GGAGGGAGTCGGATCCTGCCTAGTAG-39) and the E box (CAAGTG) located between bp 2238 and2233 was mutated in CGGATC (59-CAGAAAATACGGATCTGACATTGGCC-39), giving mMBOXpTRP1, mEBOXpTRP1, or the double mutant mM/EBOXpTRP1. In the TRP2 promoter, the M box (GTCATGTGCT) locatedbetween bp 2135 and 2129 upstream from the initiation start site was mutatedin GTCGGATCCT (59-CACTTTGGGTCGGATCCTAATGATGA-39), the Ebox (CACATG) between bp 2346 and 2340 was mutated in CGGATC (59-GGTCTTTTTTGCACGGATCTCAGAAAGC-39), and the cAMP response ele-ment (CRE; TGAGGTCA) located between bp 2239 and 2232 was mutated inTGTGTTCG (59-CCAGGATGTCTGTGTTCGCAAGTTTGGC-39), givingmMBOXpTRP2, mEBOXpTRP2, and mCREpTRP2, respectively.
Transfections and luciferase assays. B16 melanoma cells were seeded in24-well dishes, and transient transfections were performed the following day,using 2 ml of Lipofectamine and 0.55 mg of total plasmid DNA in a 200-ml finalvolume as indicated in figure legends. pCMVbGAL was transfected with the testplasmids to control the variability in transfection efficiency. At 48 h after trans-fection, soluble extracts were harvested in 50 ml of lysis buffer and assayed forluciferase and b-galactosidase activities. All transfections were repeated at leastfive times with different plasmid preparations. NIH 3T3 fibroblast cells wereseeded in six-well dishes and transiently transfected with 8 ml of Lipofectamineand 2 mg of an expression vector encoding microphthalmia in a 800-ml finalvolume. At 24 h after transfection, cells were labeled with [35S]methionine-cysteine mix. Human melanocytes were transiently transfected by electropora-tion. Two million melanocytes were resuspended in 400 ml of MCDB 153. Cellsuspensions were placed in a 0.4-cm-gap cuvette with 30 mg of total plasmidDNA. Electroporation was performed with simple electric shock (280 V; 1,050mF) by using an Easyject electroporator system (Eurogentec). Cells were incu-bated for 48 h, and then luciferase and b-galactosidase activities were assayed asdescribed for B16 cells.
Metabolic labeling and immunoprecipitation. B16 mouse melanoma cells andnontransfected or microphthalmia-transfected NIH 3T3 cells were grown insix-well dishes and labeled for 30 h with [35S]methionine-cysteine (0.1 mCi/ml;Amersham) in methionine-cysteine-free medium. Cells were then solubilized at4°C in radioimmunoprecipitation assay buffer (pH 7.5) containing 10 mM Tris-HCl, 1% sodium deoxycholate, 1% Nonidet P-40, 150 mM NaCl, 0.1% SDS, 5 mgof leupeptin per ml, 1 mM AEBSF, 100 IU of aprotinin per ml, 1 mM NaVO4,5 mM NaF, 20 mM b-glycerophosphate, and 10 mM PNPP. Total extracts wereprecleared by incubation with 50 ml of protein A-Sepharose (Pharmacia) andthen incubated with 20 ml of antibodies to the C terminus of the microphthalmia(5) complexed to 50 ml of protein A-Sepharose for 1 h at 4°C. The immunecomplexes were washed five times with radioimmunoprecipitation assay buffer,eluted in SDS-sample buffer at 95°C for 5 min, and analyzed on a 7.5% SDS gel.The specifically bound immune complexes were visualized by autoradiography.
VOL. 18, 1998 REGULATION OF TRP1 AND TRP2 PROMOTER ACTIVITIES 695
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

Nuclear extracts and gel mobility shift assay. B16 cells were stimulated with20 mM forskolin, and the nuclear extracts were prepared essentially as describedpreviously (7) except that phosphatase inhibitors (1 mM NaVO4, 5 mM NaF, 20mM b-glycerophosphate, and 10 mM PNPP) were added to the nuclear extrac-tion buffer. Double-stranded synthetic M-BOXTRP1 (59-GGAGGGAGTCATGTGCTGCCTAG-39) or E-BOXTRP1 (59-GAAAATACAAGTGTGACATTG-39) and M-BOXTRP2 (59-CTTTGGGTCATGTGCTAATGATG-39) orE-BOXTRP2 (59-GTCTTTTTTGCACACATGTCAGAAAGC-39) were end la-beled with T4 polynucleotide kinase and [g-32P]ATP. Five micrograms of nuclearproteins or 4 ml of the in vitro-translated microphthalmia was preincubated inbinding buffer containing 10 mM Tris (pH 7.5), 100 mM NaCl, 1 mM dithio-threitol, 1 mM EDTA, 4% glycerol, 80 mg of salmon sperm DNA per ml, 0.1 mgof poly(dI-dC), 10% FCS, 2 mM MgCl2, and 2 mM spermidine for 15 min on ice.Then 30,000 to 50,000 cpm of 32P-labeled probe was added to the bindingreaction for 10 min at room temperature. DNA-protein complexes were resolvedby electrophoresis on a 4% polyacrylamide (37.5:1 acrylamide-bisacrylamide) gelin TBE buffer (22.5 mM Tris-borate, 0.5 mM EDTA [pH 8]) for 2 h at 150 V.When indicated, a 50-fold excess of unlabeled competitor oligonucleotides wasadded during preincubation. For supershift assays, 0.3 ml of preimmune serum or0.3 ml of specific antibodies against microphthalmia (5) was preincubated withnuclear extracts or with in vitro-translated microphthalmia in binding reactionbuffer before addition of the labeled probe.
RESULTS
Stimulation of tyrosinase, TRP1, and TRP2 expression bycAMP-elevating agents in B16-F10 mouse melanoma cells.Since conflicting data concerning the effects of cAMP-elevat-ing agents on TRP1 and TRP2 gene expression have beenreported, we investigated the effects of forskolin on theamount of tyrosinase, TRP1, and TRP2 proteins and mRNAsin B16-F10 mouse melanoma cells. Western blot experimentsshowed that 24 or 48 h of treatment with forskolin markedlyincreased the amount of tyrosinase protein. Forskolin alsoincreased the amount of TRP1 and TRP2 proteins (Fig. 1A,upper panel). The detection of the ERK1 protein at 44 kDaensured even loading of lanes (Fig. 1A, lower panel).
In addition, tyrosinase, TRP1, and TRP2 mRNAs were an-alyzed by reverse transcription-PCR experiments. Using spe-cific primers, we amplified PCR fragments of 1,191, 784 and518 bp, corresponding to fragments of the tyrosinase, TRP1,and TRP2 cDNAs, respectively (Fig. 1B). The amounts of thePCR fragments were increased when we used cDNA from B16cells treated with forskolin for 48 h, reflecting an augmentationof tyrosinase, TRP1, and TRP2 messengers. A control of PCRamplification showed that the GAPDH transcripts were notmodified following forskolin treatment.
These results indicate that cAMP-elevating agents increasethe amount of tyrosinase, TRP1, and TRP2 proteins andmRNAs, suggesting a coordinated regulation of the expressionof these melanogenic enzymes by cAMP in B16-F10 mousemelanoma cells.
Regulation of tyrosinase, TRP1, and TRP2 transcriptionalactivities by cAMP-elevating agents and PKA. To study theeffects of cAMP on TRP1 and TRP2 gene transcription, weconstructed two luciferase reporter plasmids, containing eitherTRP1 (pTRP1; 21247 to 182) or TRP2 (pTRP2; 2585 to1349) promoter fragments that contain the elements account-able for melanocyte-specific expression (29, 41). Then B16cells were transiently transfected with pTRP1 and pTRP2 andexposed to forskolin or a-MSH for 48 h. As a control, wetransfected a reporter plasmid containing the tyrosinase pro-moter (pTyro; 22236 to 159) that we had previously found tobe responsive to cAMP-elevating agents (5). As expected, for-skolin and a-MSH, respectively, induced 30- and 20-fold in-creases in the luciferase activity in cells transfected with pTyro.Further, we observed a 20-fold stimulation of the luciferaseactivity by forskolin treatment in cells transfected with pTRP1and pTRP2, demonstrating that TRP1 and TRP2 promotersare responsive to cAMP elevation (Fig. 2). The effects evoked
by a-MSH on TRP1 and TRP2 promoters were slightly lesssignificant (15- and 10-fold stimulation, respectively). Addi-tionally, in cells transfected with pTyro, pTRP1, and pTRP2,the luciferase activities were dramatically (80-, 42-, and 37-fold,respectively) increased by cotransfection with an expressionvector encoding the catalytic subunit of PKA. The effects offorskolin, a-MSH, and PKA were severely impaired by trans-fection with an expression plasmid encoding PKI, emphasizingthe role of PKA in the effects of cAMP-elevating agents onthese promoter activities. The effect of PKI on the cAMPpathway was specific, since transfection with PKI-encodingvector did not affect the response to phorbol ester of a TRE-LUC reporter plasmid in B16 cells (not shown). The resultspresented above demonstrate that tyrosinase, TRP1, andTRP2 promoters contain cis-acting elements involved in theregulation of their transcriptional activities by cAMP throughPKA activation.
Localization of cis-acting elements in TRP1 promoter re-sponsible for cAMP response. In an attempt to localize thecis-acting elements responsible for cAMP response of theTRP1 promoter, we constructed reporter plasmids containingdeletions and mutations in the 59-flanking region of the TRP1promoter. Our previous results (5) have demonstrated that theM box upstream of the TATA box confers on the tyrosinasepromoter its responsiveness to cAMP. Comparison of the ty-rosinase, TRP1, and TRP2 promoter sequences shows thatthe M box (GTCATGTGCT) is highly conserved in thesepromoters. Hence, we constructed two reporter plasmids; inone, the M box was mutated in the pTRP1 construct(mMBOXpTRP1); the second construction had a deletion just
FIG. 1. Forskolin treatment increases tyrosinase, TRP1, and TRP2 geneexpression in B16 mouse melanoma cells. (A) Thirty micrograms of proteinsfrom control cells and cells treated for 24 or 48 h with 20 mM forskolin (FSK) wassubjected to Western blot analysis using antibodies PEP7 for tyrosinase detec-tion, B8G3 for TRP1, and PEP8 for TRP2 (upper panel). The detection of the44-kDa ERK1 protein ensured even loading of lanes (lower panel). (B) Tenmicrograms of total RNA from control B16 cells or cells treated for 48 h with 20mM forskolin was reverse transcribed. The resulting cDNAs were subjected to28-cycle PCRs using specific primers that gave amplified products of 1,191 bp fortyrosinase (TYRO), 784 bp for TRP1, and 518 bp for TRP2. A control of PCRamplification with specific primers of GAPDH transcript showed a 983-bp frag-ment. PCR products were electrophoresed on a 1% agarose gel and stained withethidium bromide before UV light visualization. DNA molecular weight markers(lane M), from bottom to top: 510, 1,018, 1,636, 2,036, 3,054, and 4,072 bp.
696 BERTOLOTTO ET AL. MOL. CELL. BIOL.
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
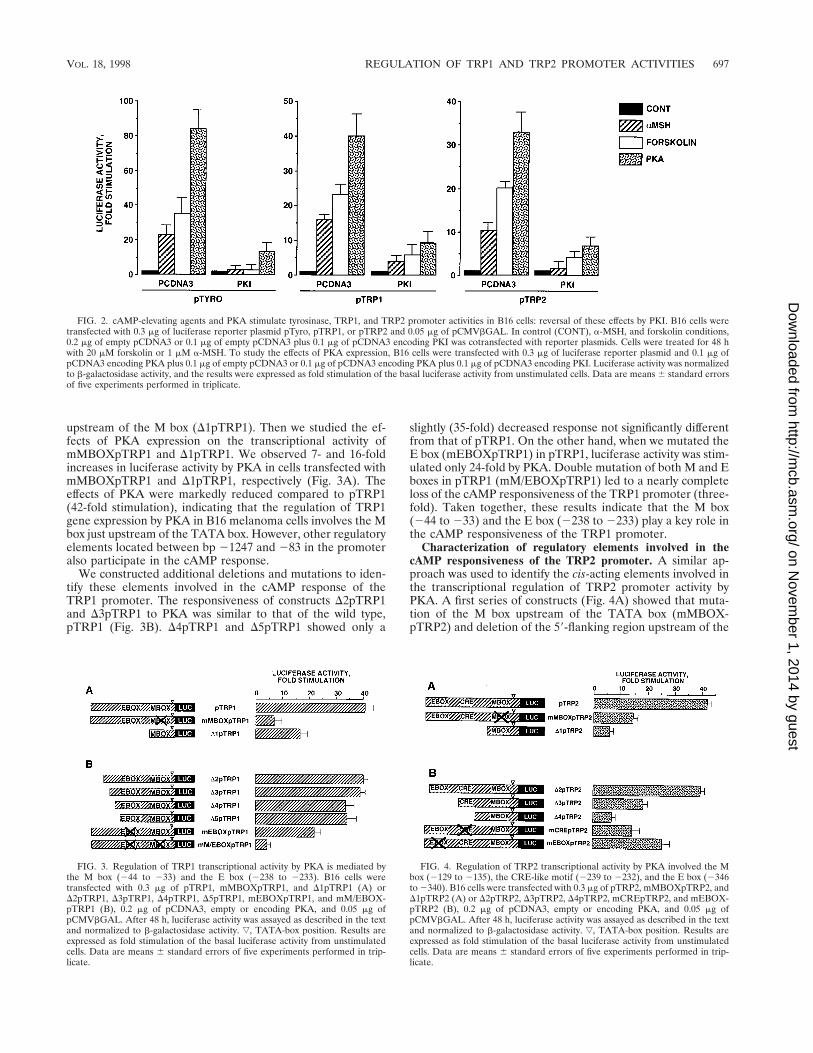
upstream of the M box (D1pTRP1). Then we studied the ef-fects of PKA expression on the transcriptional activity ofmMBOXpTRP1 and D1pTRP1. We observed 7- and 16-foldincreases in luciferase activity by PKA in cells transfected withmMBOXpTRP1 and D1pTRP1, respectively (Fig. 3A). Theeffects of PKA were markedly reduced compared to pTRP1(42-fold stimulation), indicating that the regulation of TRP1gene expression by PKA in B16 melanoma cells involves the Mbox just upstream of the TATA box. However, other regulatoryelements located between bp 21247 and 283 in the promoteralso participate in the cAMP response.
We constructed additional deletions and mutations to iden-tify these elements involved in the cAMP response of theTRP1 promoter. The responsiveness of constructs D2pTRP1and D3pTRP1 to PKA was similar to that of the wild type,pTRP1 (Fig. 3B). D4pTRP1 and D5pTRP1 showed only a
slightly (35-fold) decreased response not significantly differentfrom that of pTRP1. On the other hand, when we mutated theE box (mEBOXpTRP1) in pTRP1, luciferase activity was stim-ulated only 24-fold by PKA. Double mutation of both M and Eboxes in pTRP1 (mM/EBOXpTRP1) led to a nearly completeloss of the cAMP responsiveness of the TRP1 promoter (three-fold). Taken together, these results indicate that the M box(244 to 233) and the E box (2238 to 2233) play a key role inthe cAMP responsiveness of the TRP1 promoter.
Characterization of regulatory elements involved in thecAMP responsiveness of the TRP2 promoter. A similar ap-proach was used to identify the cis-acting elements involved inthe transcriptional regulation of TRP2 promoter activity byPKA. A first series of constructs (Fig. 4A) showed that muta-tion of the M box upstream of the TATA box (mMBOX-pTRP2) and deletion of the 59-flanking region upstream of the
FIG. 2. cAMP-elevating agents and PKA stimulate tyrosinase, TRP1, and TRP2 promoter activities in B16 cells: reversal of these effects by PKI. B16 cells weretransfected with 0.3 mg of luciferase reporter plasmid pTyro, pTRP1, or pTRP2 and 0.05 mg of pCMVbGAL. In control (CONT), a-MSH, and forskolin conditions,0.2 mg of empty pCDNA3 or 0.1 mg of empty pCDNA3 plus 0.1 mg of pCDNA3 encoding PKI was cotransfected with reporter plasmids. Cells were treated for 48 hwith 20 mM forskolin or 1 mM a-MSH. To study the effects of PKA expression, B16 cells were transfected with 0.3 mg of luciferase reporter plasmid and 0.1 mg ofpCDNA3 encoding PKA plus 0.1 mg of empty pCDNA3 or 0.1 mg of pCDNA3 encoding PKA plus 0.1 mg of pCDNA3 encoding PKI. Luciferase activity was normalizedto b-galactosidase activity, and the results were expressed as fold stimulation of the basal luciferase activity from unstimulated cells. Data are means 6 standard errorsof five experiments performed in triplicate.
FIG. 3. Regulation of TRP1 transcriptional activity by PKA is mediated bythe M box (244 to 233) and the E box (2238 to 2233). B16 cells weretransfected with 0.3 mg of pTRP1, mMBOXpTRP1, and D1pTRP1 (A) orD2pTRP1, D3pTRP1, D4pTRP1, D5pTRP1, mEBOXpTRP1, and mM/EBOX-pTRP1 (B), 0.2 mg of pCDNA3, empty or encoding PKA, and 0.05 mg ofpCMVbGAL. After 48 h, luciferase activity was assayed as described in the textand normalized to b-galactosidase activity. ƒ, TATA-box position. Results areexpressed as fold stimulation of the basal luciferase activity from unstimulatedcells. Data are means 6 standard errors of five experiments performed in trip-licate.
FIG. 4. Regulation of TRP2 transcriptional activity by PKA involved the Mbox (2129 to 2135), the CRE-like motif (2239 to 2232), and the E box (2346to 2340). B16 cells were transfected with 0.3 mg of pTRP2, mMBOXpTRP2, andD1pTRP2 (A) or D2pTRP2, D3pTRP2, D4pTRP2, mCREpTRP2, and mEBOX-pTRP2 (B), 0.2 mg of pCDNA3, empty or encoding PKA, and 0.05 mg ofpCMVbGAL. After 48 h, luciferase activity was assayed as described in the textand normalized to b-galactosidase activity. ƒ, TATA-box position. Results areexpressed as fold stimulation of the basal luciferase activity from unstimulatedcells. Data are means 6 standard errors of five experiments performed in trip-licate.
VOL. 18, 1998 REGULATION OF TRP1 AND TRP2 PROMOTER ACTIVITIES 697
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

M box (D1pTRP2) greatly impaired the responsiveness of theTRP2 promoter to PKA (14- and 6-fold stimulation, respec-tively). These results indicate that the M box is involved in thetranscriptional regulation of the TRP2 promoter activity byPKA and that important cis-regulatory elements conferringcAMP responsiveness on the TRP2 promoter are located be-tween bp 2585 and 2144.
To further characterize these elements, we constructed asecond series of deletions and mutations (Fig. 4B). The re-sponsiveness of D2pTRP2 to PKA was similar to that observedwith the initial reporter plasmid pTRP2 (37-fold), while aftertransfection with D3pTRP2, we observed only a 17-fold stim-ulation of the luciferase activity by PKA. Further deletion(D4pTRP2) led to a dramatic (sixfold) decrease in the cAMPsensitivity of the promoter. Hence, our data suggest that im-portant regulatory elements are located between bp 2475 and2273 and between bp 2273 and 2220. The mutation of the Ebox (mEBOXpTRP2) clearly impaired the effects of PKA onthe TRP2 promoter (25-fold stimulation). Interestingly, wefound a sequence (TGAGGTCA) very close to a classical CRE(TGACGTCA) located between bp 2239 and 2232 in thepromoter. When mCREpTRP2, containing mutations in theCRE-like motif (2239 to 2232), was transfected, stimulationof the luciferase activity was markedly (14-fold) reduced inresponse to PKA. Taken together, the results show that the Mbox (2129 to 2135), the E box (2346 to 2340), and theCRE-like motif (2239 to 2232) play a pivotal role in theregulation of the transcriptional activity of the TRP2 promoterby cAMP.
Stimulation of tyrosinase, TRP1, and TRP2 promoter activ-ities by PKA in normal human melanocytes. All of the resultspresented above pertain to the transformed B16 melanomacell line. Thus, we wished to verify that tyrosinase, TRP1, andTRP2 promoters respond to cAMP in normal, i.e., nontrans-formed, cells. To accomplish this, normal human melanocytesfrom skin phototype II or III were transfected with pTyro,pTRP1, and pTRP2 with or without the expression plasmidencoding PKA. The results from three different experimentsperformed on three different melanocyte cultures are shown inTable 1. Expression of PKA increased the luciferase activity innormal human melanocytes transfected with pTyro, pTRP1,and pTRP2. Similar effects were observed with cAMP-elevat-ing agents such as forskolin or a-MSH (not shown). Theseresults indicate that the transcriptional activities of tyrosinase,TRP1, and TRP2 promoters are stimulated by the cAMP path-way in normal human melanocytes.
Microphthalmia transactivates TRP1 and TRP2 promoters.To thoroughly study the role of microphthalmia in the cAMPresponsiveness of TRP1 and TRP2 promoters, we investigatedthe effects of microphthalmia on the different TRP1 and TRP2constructs and compared these effects with their cAMP re-sponses. TRP1 and TRP2 reporter constructs were cotrans-fected with an expression plasmid encoding microphthalmia.Microphthalmia induced a 40-fold stimulation of the TRP1promoter (Fig. 5A). In three other constructs, mMBOX-pTRP1, D1pTRP1, and mEBOXpTRP1, the effects of mi-crophthalmia and those of cAMP were markedly decreased.The TRP2 promoter was also transactivated (20-fold) by mi-crophthalmia (Fig. 5B). For the mMBOXpTRP2, D1pTRP2,and mEBOXpTRP2 constructs, which showed decreasedcAMP responsiveness, the activation by microphthalmia wasclearly reduced. It appears that the effects of cAMP on TRP1and TRP2 promoter activities correlate with the ability of mi-crophthalmia to transactivate the promoters.
cAMP increases the binding of microphthalmia to TRP1and TRP2 M boxes. M and E boxes involved in the cAMPresponsiveness of TRP1 and TRP2 promoters contain a coresequence, CANNTG, that binds helix-loop-helix transcriptionfactors. Thus, we analyzed the binding activity of microphthal-mia produced by in vitro transcription-translation to M and Eboxes from TRP1 and TRP2 promoters (Fig. 6A). We ob-served strongly labeled complexes with TRP1 and TRP2 Mboxes that were shifted by antimicrophthalmia antibody butnot by preimmune serum. Further, these complexes were dis-placed by homologous unlabeled probe but not by oligonucle-otides containing mutations corresponding to those introducedin mMBOXpTRP1 and mMBOXpTRP2. Complexes were alsoobserved with the TRP1 E box (2238 to 2233) and with theTRP2 E box (2346 to 2340), but these complexes were mark-edly less labeled than the M-box complexes. E-box complexeswere specifically shifted by antimicrophthalmia antibody anddisplaced by the corresponding unlabeled oligonucleotides butnot by oligonucleotides bearing mutations in the E-box coremotif. No DNA binding activity was observed when the tran-
FIG. 5. The effects of microphthalmia on TRP1 and TRP2 promoter con-structs correlate with their cAMP responsiveness. Histograms on the left repre-sent the cAMP responsiveness of the different TRP1 (A) and TRP2 (B) con-structs as determined in Fig. 3 and 4. To study the effects of microphthalmia onthese different constructs, B16 cells were cotransfected with 0.3 mg of the re-porter plasmid and 0.05 mg of pCMVbGAL plus 0.04 mg of empty pCDNA3 orpCDNA3 encoding microphthalmia. Reporter plasmids were pTRP1, mMBOX-pTRP1, D1pTRP1, and mEBOXpTRP1 (A) and pTRP2, mMBOXpTRP2,D1pTRP2, and mEBOXpTRP2 (B). After 48 h, luciferase activity was assayed aspreviously described and normalized to b-galactosidase activity. Results areexpressed as fold stimulation of the luciferase activity from cells transfected withempty pCDNA3. Data are means 6 standard error of five experiments per-formed in triplicate.
TABLE 1. Expression of PKA increases luciferase activity innormal human melanocytes transfected with pTyro, pTRP1,
and pTRP2a
VectorFold stimulation
Expt 1 Expt 2 Expt 3
pTyro 3.8 5 6.6pTRP1 9.2 6.9 10pTRP2 4.6 7.2 8.2
a Three different cultures of normal human melanocytes were transfected byelectroporation with 20 mg of reporter plasmid, 10 mg of pCDNA3, or 10 mg ofpCDNA3 encoding the catalytic subunit of PKA and 3 mg of pCMVbGAL; 48 hlater, luciferase activity was measured and corrected by b-galactosidase activity.The results represent the fold stimulation of luciferase activity in cells transfectedwith PKA expression vector compared to cells transfected with empty pCDNA3.The effects of PKA on the promoterless construct PGL2B represent means of1.3 6 0.5 for three independent experiments.
698 BERTOLOTTO ET AL. MOL. CELL. BIOL.
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
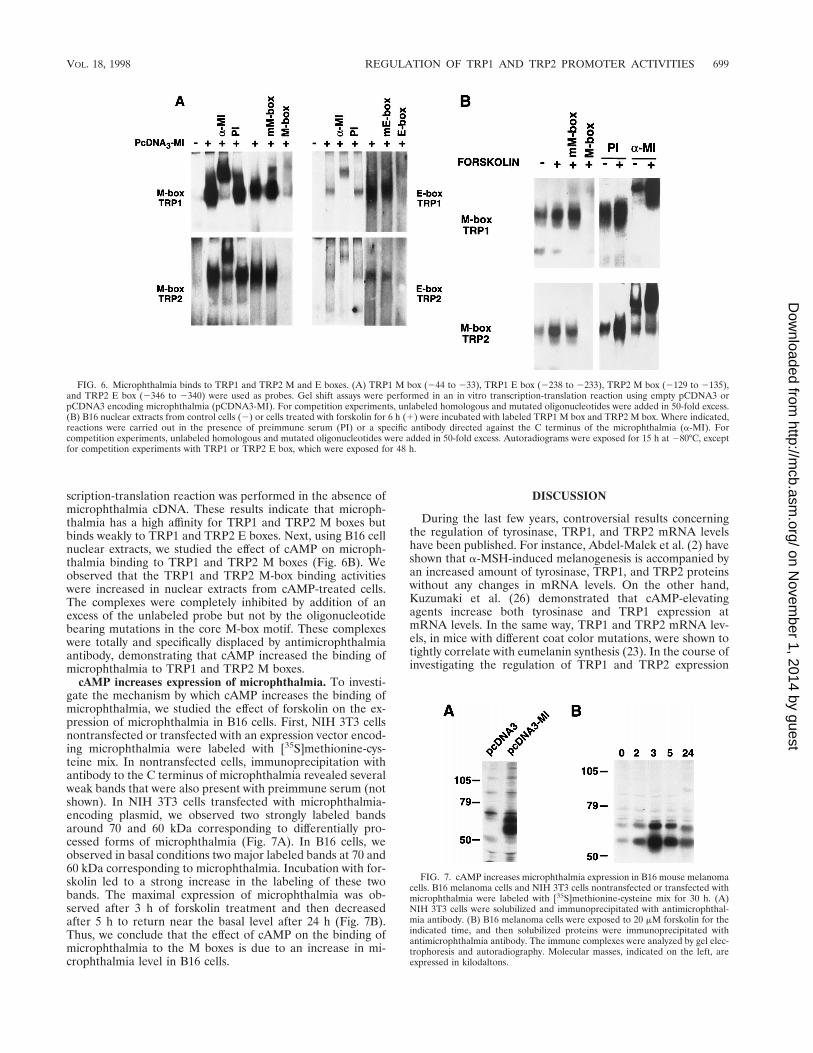
scription-translation reaction was performed in the absence ofmicrophthalmia cDNA. These results indicate that microph-thalmia has a high affinity for TRP1 and TRP2 M boxes butbinds weakly to TRP1 and TRP2 E boxes. Next, using B16 cellnuclear extracts, we studied the effect of cAMP on microph-thalmia binding to TRP1 and TRP2 M boxes (Fig. 6B). Weobserved that the TRP1 and TRP2 M-box binding activitieswere increased in nuclear extracts from cAMP-treated cells.The complexes were completely inhibited by addition of anexcess of the unlabeled probe but not by the oligonucleotidebearing mutations in the core M-box motif. These complexeswere totally and specifically displaced by antimicrophthalmiaantibody, demonstrating that cAMP increased the binding ofmicrophthalmia to TRP1 and TRP2 M boxes.
cAMP increases expression of microphthalmia. To investi-gate the mechanism by which cAMP increases the binding ofmicrophthalmia, we studied the effect of forskolin on the ex-pression of microphthalmia in B16 cells. First, NIH 3T3 cellsnontransfected or transfected with an expression vector encod-ing microphthalmia were labeled with [35S]methionine-cys-teine mix. In nontransfected cells, immunoprecipitation withantibody to the C terminus of microphthalmia revealed severalweak bands that were also present with preimmune serum (notshown). In NIH 3T3 cells transfected with microphthalmia-encoding plasmid, we observed two strongly labeled bandsaround 70 and 60 kDa corresponding to differentially pro-cessed forms of microphthalmia (Fig. 7A). In B16 cells, weobserved in basal conditions two major labeled bands at 70 and60 kDa corresponding to microphthalmia. Incubation with for-skolin led to a strong increase in the labeling of these twobands. The maximal expression of microphthalmia was ob-served after 3 h of forskolin treatment and then decreasedafter 5 h to return near the basal level after 24 h (Fig. 7B).Thus, we conclude that the effect of cAMP on the binding ofmicrophthalmia to the M boxes is due to an increase in mi-crophthalmia level in B16 cells.
DISCUSSION
During the last few years, controversial results concerningthe regulation of tyrosinase, TRP1, and TRP2 mRNA levelshave been published. For instance, Abdel-Malek et al. (2) haveshown that a-MSH-induced melanogenesis is accompanied byan increased amount of tyrosinase, TRP1, and TRP2 proteinswithout any changes in mRNA levels. On the other hand,Kuzumaki et al. (26) demonstrated that cAMP-elevatingagents increase both tyrosinase and TRP1 expression atmRNA levels. In the same way, TRP1 and TRP2 mRNA lev-els, in mice with different coat color mutations, were shown totightly correlate with eumelanin synthesis (23). In the course ofinvestigating the regulation of TRP1 and TRP2 expression
FIG. 6. Microphthalmia binds to TRP1 and TRP2 M and E boxes. (A) TRP1 M box (244 to 233), TRP1 E box (2238 to 2233), TRP2 M box (2129 to 2135),and TRP2 E box (2346 to 2340) were used as probes. Gel shift assays were performed in an in vitro transcription-translation reaction using empty pCDNA3 orpCDNA3 encoding microphthalmia (pCDNA3-MI). For competition experiments, unlabeled homologous and mutated oligonucleotides were added in 50-fold excess.(B) B16 nuclear extracts from control cells (2) or cells treated with forskolin for 6 h (1) were incubated with labeled TRP1 M box and TRP2 M box. Where indicated,reactions were carried out in the presence of preimmune serum (PI) or a specific antibody directed against the C terminus of the microphthalmia (a-MI). Forcompetition experiments, unlabeled homologous and mutated oligonucleotides were added in 50-fold excess. Autoradiograms were exposed for 15 h at 280°C, exceptfor competition experiments with TRP1 or TRP2 E box, which were exposed for 48 h.
FIG. 7. cAMP increases microphthalmia expression in B16 mouse melanomacells. B16 melanoma cells and NIH 3T3 cells nontransfected or transfected withmicrophthalmia were labeled with [35S]methionine-cysteine mix for 30 h. (A)NIH 3T3 cells were solubilized and immunoprecipitated with antimicrophthal-mia antibody. (B) B16 melanoma cells were exposed to 20 mM forskolin for theindicated time, and then solubilized proteins were immunoprecipitated withantimicrophthalmia antibody. The immune complexes were analyzed by gel elec-trophoresis and autoradiography. Molecular masses, indicated on the left, areexpressed in kilodaltons.
VOL. 18, 1998 REGULATION OF TRP1 AND TRP2 PROMOTER ACTIVITIES 699
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

during cAMP-induced melanogenesis, we clearly showed inthis report that cAMP increases tyrosinase, TRP1, and TRP2at both protein and mRNA levels. Our results demonstrate acoordinated regulation of tyrosinase, TRP1, and TRP2 expres-sion at mRNA levels during cAMP-induced melanogenesis.Next, we showed that cAMP-elevating agents such as forskolinand a-MSH or expression of the catalytic subunit of PKAstimulate the transcriptional activity of the TRP1 and TRP2promoters. The role of PKA in the melanogenic pathway wasconfirmed by transfection with an expression plasmid encodingPKI, which dramatically reduces the effect of forskolin,a-MSH, and PKA. These results clearly demonstrate thata-MSH effects on melanogenesis are mediated through theactivation of the cAMP pathway and PKA.
The regulation of tyrosinase, TRP1, and TRP2 promoters byPKA was observed in other human or mouse melanoma celllines such as G361 or S91 (data not shown). Furthermore, wealso observed a stimulation of the melanogenic promoter ac-tivities by PKA in normal human melanocytes, emphasizing thephysiological relevance of the cAMP effects. It should be notedthat human melanocytes express high basal levels of melano-genic proteins (13), probably because culture conditions ofnormal melanocytes are unable to prevent a spontaneous dif-ferentiation of the cells. Thus, consequent to the high basalmelanogenesis, the effect of PKA on tyrosinase, TRP1, andTRP2 promoters appears markedly less pronounced in humanmelanocytes than in mouse melanoma cells. In other cell types,such as NIH 3T3 mouse fibroblasts, cAMP does not change theactivity of the melanogenic promoters (not shown). These re-sults indicate that a cell-specific mechanism is involved in thecAMP response of the melanogenic promoters. We then at-tempted to localize and to identify the cis-regulatory elementsconferring on TRP1 and TRP2 promoters their cAMP respon-siveness. We demonstrated that the M box and the E boxupstream of the TATA box play a key role in the cAMPresponse of TRP1 and TRP2 promoters. Recently, we alsoreported that the cAMP response of the tyrosinase promoterinvolved a M box and an E box surrounding the TATA box (5).Thus, the mechanisms of regulation of tyrosinase, TRP1, andTRP2 promoters by cAMP appear to rely on similar regulatoryelements. However, it should be noted that in the tyrosinasepromoter, the cAMP-sensitive E box located at the initiatorsite binds tightly to microphthalmia and is absolutely necessaryfor the cAMP response. In TRP1 and TRP2 promoters, the Mboxes bind avidly to microphthalmia, but the E boxes, involvedin the cAMP response, interact weakly with microphthalmia.Thus, microphthalmia appears to require the core motif CATGTG to provide a strong interaction with DNA, since thismotif is found in all of the M boxes and in the tyrosinase E box,while TRP1 and TRP2 E boxes have CAAGTG and CAATTG, respectively, as core sequences. We cannot exclude thepossibility that in intact cells and in the context of the intactpromoter, microphthalmia tightly binds to TRP1 and TRP2 Eboxes. However, mutations of TRP1 and TRP2 E boxes mod-erately affect cAMP sensitivity and the microphthalmia effecton the promoters, suggesting that in intact cells, microphthal-mia does not interact strongly with these motifs. In the TRP2promoter, a CRE-like sequence also participates in the cAMPresponse, indicating that transcription factors of the CREBfamily are involved in the stimulation of the TRP2 promoteractivity by cAMP. However, this CRE motif has to cooperatewith the M box to give full cAMP sensitivity to the TRP2promoter. Noteworthy, the same cis-acting elements (M and Eboxes) were thought to mediate the tissue-specific expression(4) and the cAMP responses of the tyrosinase, TRP1, andTRP2 genes. Thus, it should be considered that the cAMP
pathway participates in the regulation of the tissue-specificexpression of the melanocyte-specific genes.
Although microphthalmia and M boxes play a pivotal role inthe regulation of tyrosinase, TRP1, and TRP2 promoter activ-ities by cAMP, it appears that each promoter responds tocAMP through specific mechanisms because of the relativeposition of the regulatory elements or the intrinsic nature ofthe elements cooperating with the M box. This is particularlytrue for the TRP2 promoter, which contains a CRE-like ele-ment involved in cAMP sensitivity; in tyrosinase and TRP1promoters, no CRE participates in the cAMP response. Theseobservations reveal, besides a common regulatory mechanism,the existence of specific processes involved in the control oftyrosinase, TRP1, and TRP2 gene expression that could allowa differential expression pattern of the melanogenic enzymesunder particular physiological and pathological conditions.
We have previously demonstrated that cAMP increases mi-crophthalmia binding to the M box and to the E box. Sincemicrophthalmia has a strong stimulating effect on the tyrosi-nase promoter, we have proposed that microphthalmia,through the binding to M and E boxes, mediates the effects ofcAMP on tyrosinase gene expression (5). In the present report,we clearly showed a strong stimulation of TRP1 and TRP2promoter activities by microphthalmia. Consistently, microph-thalmia or its human homolog MITF (microphthalmia-associ-ated transcription factor) was shown to transactivate the TRP1promoter (39, 40). However, the TRP2 promoter has beenshown to be unresponsive to MITF (39), while we clearlyshowed in this report that microphthalmia stimulates the TRP2promoter activity. This discrepancy could be explained eitherby specific behaviors of human versus mouse microphthalmiaor by the different cellular contexts.
Interestingly, the effect of microphthalmia on differentTRP1 and TRP2 promoter constructs tightly correlates withtheir cAMP responsiveness. Further, we showed that cAMPincreases the binding of microphthalmia to TRP1 and TRP2 Mboxes. These data strongly suggest that the effect of cAMP onTRP1 and TRP2 gene expression is mediated through an in-creased interaction of microphthalmia with M boxes, therebyleading to a transactivation of TRP1 and TRP2 promoters. Atleast two hypothesis could explain the stimulation of microph-thalmia binding on the M box after cAMP treatment. cAMPcould increase either microphthalmia expression or affinity ofmicrophthalmia for its target sequences. Previous immunoflu-orescence studies with antimicrophthalmia antibody did notshow a stimulation of microphthalmia expression after 24 hwith forskolin. Although this result does not support the firsthypothesis, we have reassessed the effect of cAMP on microph-thalmia expression. Metabolic labeling followed by immuno-precipitation with specific antibody to microphthalmia demon-strates that cAMP increases microphthalmia expression.Microphthalmia appears as two bands around 60 and 70 kDa.The lower mobility of the higher band results from its phos-phorylation on serine 73 by mitogen-activated protein kinases(16). The maximal expression of microphthalmia was obtainedafter 3 h of forskolin treatment, and consistent with our im-munofluorescence studies, the level of microphthalmia re-turned near the basal level after 24 h with forskolin. Thepresence of a CRE in the microphthalmia promoter (10) sug-gests that PKA regulates microphthalmia expression throughthe classical pathway involving transcription factors of theCREB family. However, this hypothesis remains to be demon-strated. Interestingly, Rungta et al. (33) showed that a-MSHtreatment significantly increases the tyrosinase mRNA levelswithin 16 h, and we previously observed an effect of forskolinon tyrosinase promoter after 6 h. Further, careful examination
700 BERTOLOTTO ET AL. MOL. CELL. BIOL.
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

of the report of Abdel-Malek et al. (2) shows that the TRP1mRNA amount was increased after 6 h with a-MSH. Thus, ifwe compare the kinetics of microphthalmia induction withthose of tyrosinase and TRP1, the upregulation of the mela-nogenic enzymes is observed clearly after the maximal expres-sion of microphthalmia. These observations are consistent withour former hypothesis suggesting that microphthalmia plays akey role in the stimulation of melanogenic gene expression.
Considering the physiological aspect of our findings, itshould be mentioned that in humans, melanogenesis is stimu-lated by UVB, which upregulates the production of a-MSH byepidermal keratinocytes. Further, subcutaneous injection ofa-MSH has been shown to stimulate local pigmentation (28).Thus, we can hypothesize that a-MSH, through the binding toits receptor coupled to the G protein as and adenylate cyclase,increases the cAMP content in melanocytes. Then cAMP,through the activation of PKA, leads to an augmentation ofmicrophthalmia expression. Consequently, the amount of mi-crophthalmia bound to M or E boxes increases resulting in astimulation of the melanogenic promoter activities. Taken to-gether, our results disclosed the cascade of molecular eventsinvolved in the regulation of the melanogenic genes that couldbe of paramount importance in the control of skin pigmentation.
ACKNOWLEDGMENTS
We thank V. Hearing (Bethesda, Md.) for providing antityrosinase(PEP7) and anti-TRP2 (PEP8) antibodies and P. G. Parsons (Bris-bane, Australia) for anti-TRP1 antibody B8G3. We also thank P.Sassone-Corsi and E. Lalli (Illkirch, France) for providing the expres-sion vector encoding the catalytic subunit of PKA and R. Maurer(Portland, Oreg.) for the expression vector encoding PKI. We aregrateful to A. Grima and C. Minghelli for illustration work and to J. C.Scimeca and C. Sable for critical reading of the manuscript.
This work was supported by Association pour la Recherche sur leCancer (grant 6760) and Ligue Nationale Contre le Cancer.
REFERENCES
1. Abdel-Malek, Z., V. B. Swope, N. Amornsiripanitch, and J. J. Nordlund.1987. In vitro modulation of proliferation and melanization of S91 mela-noma cells by prostaglandins. Cancer Res. 47:3141–3146.
2. Abdel-Malek, Z., V. B. Swope, I. Suzuki, C. Akcali, M. D. Harriger, S. T.Boyce, K. Urabe, and V. Hearing. 1995. Mitogenic and melanogenic stimu-lation of normal human melanocytes by melanotropic peptides. Proc. Natl.Acad. Sci. USA 92:1789–1793.
3. Barber, J. I., D. Townsend, D. P. A. Olds, and R. A. King. 1984. Dopachromeoxydoreductase: a new enzyme in the pigment pathway. J. Invest. Dermatol.83:145–149.
4. Bentley, N. J., T. Eisen, and C. R. Goding. 1994. Melanocyte-specific expres-sion of the human tyrosinase promoter: activation by the microphthalmiagene product and role of the initiator. Mol. Cell. Biol. 14:7996–8006.
5. Bertolotto, C., K. Bille, J. P. Ortonne, and R. Ballotti. 1996. Regulation oftyrosinase gene expression by cAMP in B16 melanoma cells involves twoCATGTG motifs surrounding the TATA box: implication of the microph-thalmia gene product. J. Cell Biol. 134:747–755.
6. Day, R. N., J. A. Walder, and R. A. Maurer. 1989. A protein kinase inhibitorgene reduces both basal and multihormone-stimulated prolactin gene tran-scription. J. Biol. Chem. 264:431–436.
7. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcrip-tion initiation by RNA polymerase II in a soluble extract from isolatedmammalian nuclei. Nucleic Acids Res. 11:1475–1489.
8. Englaro, W., R. Rezzonico, M. Durand-Clement, D. Lallemand, J. P. Or-tonne, and R. Ballotti. 1995. Mitogen-activated protein kinase pathway andAP-1 are activated during cAMP-induced melanogenesis in B-16 melanomacells. J. Biol. Chem. 270:24315–24320.
9. Fuller, B. B., and D. H. Viskochil. 1979. The role of RNA and proteinsynthesis in mediating action of MSH on melanoma cell cultures. Life Sci.24:2405–2416.
10. Fuse, N., K. I. Yasumoto, H. Suzuki, K. Takahashi, and S. Shibahara. 1996.Identification of a melanocyte-type promoter of the microphthalmia-associ-ated transcription factor gene. Biochem. Biophys. Res. Commun. 219:702–707.
11. Ganss, R., G. Schutz, and F. Beermann. 1994. The mouse tyrosinase gene.J. Biol. Chem. 269:29808–29816.
12. Gordon, P. R., C. P. Mansur, and B. A. Gilchrest. 1989. Regulation of human
melanocyte growth, dendricity, and melanization by keratinocyte derivedfactors. J. Invest. Dermatol. 92:565–572.
13. Halaban, R., S. H. Pomerantz, S. Marshall, D. T. Lambert, and A. B. Lerner.1983. Regulation of tyrosinase in human melanocytes grown in culture. J.Cell Biol. 97:480–488.
14. Hearing, V. J. 1987. Mammalian monophenol monooxygenase (tyrosinase):purification, properties and reactions catalyzed. Methods Enzymol. 142:154–165.
15. Hearing, V. J., and K. Tsukamoto. 1991. Enzymatic control of pigmentationin mammals. FASEB J. 5:2902–2909.
16. Hemesath, T. J., E. R. Price, C. Takemoto, T. Badalian, and D. E. Fisher.MAPK links microphthalmia to c-kit signaling in melanocytes. Nature, inpress.
17. Hirobe, T., and T. Takeuchi. 1977. Induction of melanogenesis in vitro in theepidermal melanoblasts of newborn mouse skin MSH. J. Embryol. Exp.Morphol. 37:79–80.
18. Hunt, G., C. Todd, J. E. Cresswell, and A. J. Thody. 1994. a-MSH and itsanalog Nle4DPhe7 a-MSH affect morphology, tyrosinase activity and mela-nogenesis in cultured human melanocytes. J. Cell Sci. 107:205–211.
19. Jackson, J. I., D. M. Cambers, K. Tsukamoto, N. Copeland, D. J. Gilbert,N. A. Jenkins, and V. J. Hearing. 1992. A second tyrosinase-related protein,TRP-2, maps to and mutated at the mouse slaty locus. EMBO J. 11:527–535.
20. Jimenez, M., K. Kameyama, W. L. Maloy, Y. Tomita, and V. J. Hearing.1988. Mammalian tyrosinase: biosynthesis, processing, and modulation bymelanocyte-stimulating hormone. Proc. Natl. Acad. Sci. USA 85:3830–3834.
21. Jimenez-Cervantes, C., F. Solano, T. Kobayashi, K. Urabe, V. J. Hearing,J. A. Lozano, and J. C. Garcıa-Borron. 1994. A new enzymatic function inthe melanogenic pathway: the 5,6-dihydroxyindole-2-carboxylic acid oxidaseactivity of tyrosinase-related protein-1 (TRP1). J. Biol. Chem. 269:29198–29205.
22. Kameyama, K., M. Jimenez, J. Muller, Y. Ishida, and V. J. Hearing. 1989.Regulation of mammalian melanogenesis by tyrosinase inhibition. Differen-tiation 42:28–36.
23. Kobayashi, T., W. D. Viera, B. Potterf, C. Sakai, and G. Imokawa. 1995.Modulation of melanogenic protein expression during the switch from eu- topheomelanogenesis. J. Cell Sci. 108:2301–2309.
24. Kobayashi, T., K. Urabe, A. Winder, C. Jimenez-Cervantes, G. Imokawa, T.Brewington, F. Solano, J. C. Garcıa-Borron, and V. J. Hearing. 1994. Ty-rosinase related protein 1 (TRP1) functions as a DHICA oxidase in melaninbiosynthesis. EMBO J. 13:5818–5825.
25. Korner, A., and J. Pawelek. 1982. Mammalian tyrosinase catalyzes threereactions in the biosynthesis of melanin. Science 217:1163–1165.
26. Kuzumaki, T., A. Matsuda, K. Wakamatsu, S. Ito, and K. Ishikawa. 1993.Eumelanin biosynthesis is regulated by coordinate expression of tyrosinaseand tyrosinase-related protein-1 genes. Exp. Cell Res. 207:33–40.
27. Le Douarin, N. 1982. The neural crest. Cambridge University Press, Cam-bridge, England.
28. Levine, N., S. N. Sheftel, T. Eytan, R. Dorr, M. E. Hadley, J. C. Weinrach,G. A. Ertl, K. Toth, D. L. McGee, and V. J. Hurby. 1991. Induction of skintanning by subcutaneous administration of a potent synthetic melanotropin.JAMA 226:2730–2736.
29. Lowings, P., U. Yavuzer, and R. Goding. 1992. Positive and negative ele-ments regulate a melanocyte-specific promoter. Mol. Cell. Biol. 12:3653–3662.
30. Molina, C. A., N. S. Foulkes, E. Lalli, and P. Sassone-Corsi. 1993. Induc-ibility and negative autoregulation of CREM: an alternative promoter di-rects expression of ICER, an early response repressor. Cell 75:875–886.
31. Prota, G. 1988. Some new aspects of eumelanin chemistry. Prog. Clin. Biol.Res. 256:101–124.
32. Prota, G. 1992. Melanins and melanogenesis. Academic Press, New York,N.Y.
33. Rungta, D., T. D. Corn, and B. B. Fuller. 1996. Regulation of tyrosinasemRNA in mouse melanoma cells by a-melanocyte-stimulating hormone.J. Invest. Dermatol. 107:689–693.
34. Steel, K. P., D. R. Davidson, and I. J. Jackson. 1992. TRP-2/DT, a new earlymelanoblast marker, shows that steel growth factor (c-kit ligand) is a survivalfactor. Development 115:1111–1119.
35. Swope, V. B., Z. Abdel-Malek, L. M. Kassem, and J. J. Nordlund. 1991.Interleukins 1 alpha and 6 and tumor necrosis factor-alpha are paracrineinhibitors of human melanocyte proliferation and melanogenesis. J. Invest.Dermatol. 96:180–185.
36. Takahashi, H., and P. G. Parsons. 1990. In vitro phenotypic alteration ofhuman melanoma cells induced by differentiating agents. Pigment Cell Res.3:223–232.
37. Wong, G., and J. Pawelek. 1973. Control of phenotypic expression of cul-tured melanoma cells by melanocyte stimulating hormone. Nature 241:213–215.
38. Wong, G., and J. Pawelek. 1975. Melanocyte-stimulating hormone promotesactivation of pre-existing tyrosinase molecules in Cloudman S91 melanomacells. Nature 255:644–645.
39. Yasumoto, K. I., K. Yokoyama, K. Takahashi, Y. Tomita, and S. Shibahara.1997. Functional analysis of microphthalmia-associated transcription factor
VOL. 18, 1998 REGULATION OF TRP1 AND TRP2 PROMOTER ACTIVITIES 701
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

in pigment cell-specific transcription of the human tyrosinase family genes.J. Biol. Chem. 272:503–509.
40. Yavuzer, U., E. Keenan, P. Lowings, J. Vachtenheim, G. Currie, and C. R.Goding. 1995. The microphthalmia gene product interacts with the retino-blastoma protein in vitro and is a target for deregulation of melanocyte-specific transcription. Oncogene 10:123–134.
41. Yokoyama, K., K. I. Yasumoto, H. Suzuki, and S. Shibahara. 1994. Cloning
of the human DOPAchrome tautomerase/tyrosinase-related protein 2 geneand identification of two regulatory regions required for its pigment cell-specific expression. J. Biol. Chem. 269:27080–27087.
42. Yokoyama, K., H. Suzuki, Y. Tomita, and S. Shibahara. 1994. Molecularcloning of and functional analysis of a cDNA coding for human DOPA-chrome tautomerase/tyrosinase-related protein-2. Biochim. Biophys. Acta1217:317–321.
702 BERTOLOTTO ET AL. MOL. CELL. BIOL.
on Novem
ber 1, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
Related Documents











