AMER. ZOOL., 25:291-301 (1985) Differences and Similarities: Comparative Methods in Mastication 1 CARL GANS Division of Biological Sciences, The University of Michigan, Ann Arbor, Michigan 48109 SYNOPSIS. Comparison is the key to biological study and both similarities and differences aid in sharpening our understanding of structures and processes. The initial comparison represents an observational level; followed by decisions about possible homology, analogy and homoplasy which represent levels that have less certainty. Analogous homoplasies are presumably the most useful conditions for characterizing functional constraints. Opera- tional criteria are discussed, as are minimal technical standards for procedures permitting characterization of functions and roles in the masticatory system. INTRODUCTION Most of what we know is based upon comparison. When asked to describe a food not previously tasted or a new kind of music, one often responds that the taste is "like" some other food, or that the sensation "dif- fers" in a particular way from something that is familiar. Indeed, comparison and the similarities and differences it discloses is ingrained in our approach to description of objects, events and processes. Hence the questions "what can we compare?" and its ancillary "how shall we compare?" prove to be the key to any study of natural phe- nomena. Biologists apply special standards to establish the similarities and differences of organisms. We recognize that these organ- isms and all of their structures evolved rather than having been created. The idea of evolution incorporates the implicit assumption that organisms started out with an ancestral phenotype and diverged from this. This makes it appropriate to ask why some aspects of organisms differ from those in related species, whereas other aspects appear to be similar. Assuming that all organisms share some level of common ancestry, we can argue that the apparent similarities of structures or behaviors reflect the retention of ances- tral characteristics (perhaps by genetical and developmental mechanisms), or that the similarity is secondary, due either to 1 From the Symposium on Mammalian Mastication: An Overview presented at the Annual Meeting of the American Society of Zoologists, 27-30 December 1983, at Philadelphia, Pennsylvania. common environmental effects or to ran- dom accident. In contrast, the apparent dif- ferences may reflect either the effects of random genetic changes or the adaptive response of organisms to different envi- ronments. In short, the process of evolu- tion appears to endow the observed phe- notypes with evidence both for conservative aspects and for changed ones. The traces of conservative aspects in the phenotype reflect the fact that all aspects of the phe- notype were formed by modification and adjustment, rather than by total replace- ment of structures and processes; traces of common historical features always remain. The evidence for phenotypic change reflects the diversifying forces; these result either from "random" alterations, for instance those occurring in the genetic material, or from adaptation to the influ- ences of the environment. Comparison of morphology and func- tion is based upon observation and may be tested by experiment. Uncertainties often reflect methodology, so that refinements generally remain possible and reliability may be improved. However, search for the meaning or possible causes of observed dif- ferences and similarities introduces a much greater level of uncertainty. Any answer represents a statement, not only about the relative likelihood of historical processes and events, but about the actual sequence in which changes occurred. Rarely can putative causes be confirmed directly; cer- tainly, the fossil record shows intrinsic uncertainty. These generalizations are hardly new. Except for nomenclatorial issues (cf. Pat- terson, 1982) they tend to be generally 291 Downloaded from https://academic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AMER. ZOOL., 25:291-301 (1985)
Differences and Similarities: Comparative Methods in Mastication1
CARL GANS
Division of Biological Sciences, The University of Michigan,Ann Arbor, Michigan 48109
SYNOPSIS. Comparison is the key to biological study and both similarities and differencesaid in sharpening our understanding of structures and processes. The initial comparisonrepresents an observational level; followed by decisions about possible homology, analogyand homoplasy which represent levels that have less certainty. Analogous homoplasies arepresumably the most useful conditions for characterizing functional constraints. Opera-tional criteria are discussed, as are minimal technical standards for procedures permittingcharacterization of functions and roles in the masticatory system.
INTRODUCTION
Most of what we know is based uponcomparison. When asked to describe a foodnot previously tasted or a new kind of music,one often responds that the taste is "like"some other food, or that the sensation "dif-fers" in a particular way from somethingthat is familiar. Indeed, comparison andthe similarities and differences it disclosesis ingrained in our approach to descriptionof objects, events and processes. Hence thequestions "what can we compare?" and itsancillary "how shall we compare?" proveto be the key to any study of natural phe-nomena.
Biologists apply special standards toestablish the similarities and differences oforganisms. We recognize that these organ-isms and all of their structures evolvedrather than having been created. The ideaof evolution incorporates the implicitassumption that organisms started out withan ancestral phenotype and diverged fromthis. This makes it appropriate to ask whysome aspects of organisms differ from thosein related species, whereas other aspectsappear to be similar.
Assuming that all organisms share somelevel of common ancestry, we can arguethat the apparent similarities of structuresor behaviors reflect the retention of ances-tral characteristics (perhaps by geneticaland developmental mechanisms), or thatthe similarity is secondary, due either to
1 From the Symposium on Mammalian Mastication:An Overview presented at the Annual Meeting of theAmerican Society of Zoologists, 27-30 December1983, at Philadelphia, Pennsylvania.
common environmental effects or to ran-dom accident. In contrast, the apparent dif-ferences may reflect either the effects ofrandom genetic changes or the adaptiveresponse of organisms to different envi-ronments. In short, the process of evolu-tion appears to endow the observed phe-notypes with evidence both for conservativeaspects and for changed ones. The tracesof conservative aspects in the phenotypereflect the fact that all aspects of the phe-notype were formed by modification andadjustment, rather than by total replace-ment of structures and processes; traces ofcommon historical features always remain.The evidence for phenotypic changereflects the diversifying forces; these resulteither from "random" alterations, forinstance those occurring in the geneticmaterial, or from adaptation to the influ-ences of the environment.
Comparison of morphology and func-tion is based upon observation and may betested by experiment. Uncertainties oftenreflect methodology, so that refinementsgenerally remain possible and reliabilitymay be improved. However, search for themeaning or possible causes of observed dif-ferences and similarities introduces a muchgreater level of uncertainty. Any answerrepresents a statement, not only about therelative likelihood of historical processesand events, but about the actual sequencein which changes occurred. Rarely canputative causes be confirmed directly; cer-tainly, the fossil record shows intrinsicuncertainty.
These generalizations are hardly new.Except for nomenclatorial issues (cf. Pat-terson, 1982) they tend to be generally
291
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

292 CARL GANS
No
SimilarityFIG. 1. Venn diagram to show three kinds of currentsimilarities likely to be exhibited by organisms andthe combined states that these may display. Note thatsimilarity of function is not an aspect generated bythe phenotype, but rather one that is permitted bythe phenotype.
accepted. Here, I would like to deal withthe operational standards that they implyfor any comparisons of biological systems.How do we actually deal with structuralcriteria? What does this imply about reli-ability? How widely should we sample?What do we do about "functions"? Whatevidence must we demand before we claimto have discovered similarities or demon-strated differences? Why is all of this pos-sibly interesting and certainly significant tostudies of the masticatory system?
O N COMPARISONS OF STRUCTURES
Comparative morphology is the toolapplied at the beginning of any descriptiveor functional study in biology. The rulesof comparative morphology have recentlybeen expressed by utilizing a set of Venndiagrams. As initially proposed for this useby Hailman (1976, modified versions pre-sented by Gans, 1974, and Hildebrand,1982), they combined currently observedand historical (patristic) similarities and thustwo discrete levels of reliability. Hence, Ihere modify the Venn diagram and use it
entirely for currently observed aspects ofstructures (Fig. 1). The figure suggests thatthe plane of the diagram includes all cur-rently possible states of comparison. Thismakes it easy to visualize a restriction ofthis array of theoretically possible statesand to compare various sets of similaritiesby means of such diagrams.
In the present example, there are threecomparisons. Objects that share similarityof "phenotypic structure" are enclosed inthe first circle. Those that share similarityof "phenotypic development" are enclosedin the second and those that share similar-ity of "function" enabled by the phenotypeare enclosed in the third. Structure meansappearance, shape, topographical relation,vascularization, innervation, and the mate-rial of which the tissue is composed. Devel-opment means ontogeny, the nature of theembryonic tissues, ontogenetic characterprecedence, timing and mechanism of ori-gin. It also should include similarity of thegenetic instruction, although this posessome questions that have only recentlybegun to be faced. (Certain structuralaspects, such as the innervation of musclesand the topography of blood vessels areintermediate between categories; althoughthe adult condition may represent struc-tural similarity, it implies an unusuallydirect reflection of development.) Func-tion means mechanism and physical andchemical activity. All of these aspects maybe viewed in the present.
Such a purely descriptive scheme makesit easy to document that certain subsets ofobjects (such as wing of bird and wing ofbat) share only similar function and notsimilar structure, and that other subsets ofobjects (such as humerus of man and ofchimpanzee) share similar structure andsimilar development. Some elements (man-dible of mouse and mandible of rat) shareall three presently compared attributes,whereas many groups of elements (limb ofarthropod and jaw of elephant) do not dis-play even a single shared attribute. Thisformulation involves three sets of attri-butes that are subject to observation, torefinement and to test. Finer resolution, ineach of the three kinds of such compari-sons expressed in Venn diagrams, always
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

COMPARISON IN MASTICATION 293
reduces the probability of being mistakenin a claim that a situation involves similar-ity or difference.
SOME DEFINITIONS
We now come to the causes of theobserved similarities. Perhaps the easiestcause to establish is that certain structuresfound in two or more species arose by con-tinuous phyletic evolution from structuresin a common ancestor, in other words thatthey are homologous. Structures that didnot arise in this fashion are consideredhomoplastic. The concept of analogyapplies to structures, homologous or not,the similarity of which derives from pastor presently shared function.
Three other terms deserve definition.Parallelism is similar structural change inhomologous structures in separate phyleticlines. Reversal is the return of a (homol-ogous) structure to an original state froma modified form. Convergence is modifi-cation of two homoplastic structures to asimilar appearance, presumably by theaction of convergent selection.
DECISIONS ABOUT HOMOLOGY
The logic applied to this most importantdecision that two structures are homolo-gous, normally involves two steps, namelythe conclusion (demonstration) that thestructures and positions of two objects aresimilar in certain aspects, and the dem-onstration (assumption) that the structuresare also similar in the phylogenetically con-necting forms. Consequently, two homol-ogous structures are likely to represent acondition common to all members of thephyletic line that includes the species beingcompared. Hence, homology is an attrib-ute of synapomorphic characters. Althoughthese criteria are generally accepted, theway the issue is handled operationallyinvolves some short-cuts that are tolerableonly as long as their basis is understood.
Obviously, there is a hierarchy in the useof and requirement for the operational cri-teria used to induce homology, affectingthe assumptions one makes during a study.For instance, whenever we compare oneparticular characteristic between twospecies (or members of two other taxa) that
are otherwise remarkably similar, we tendto incorporate the observation of overallsimilarity of body plan (and assumption ofsimilarity in connecting forms) into deci-sions about the causes that have shaped theparticular structure. In short, we assumethat in two animals of high overall similar-ity, structural characteristics (such as sharedshape, topography and placement) may besufficient to document homology of yetanother structure examined. This assump-tion would, for instance, apply to compar-ison of major elements, such as the man-dibles, of two kinds of squirrels, elephantsor whales. The more probable the closerelation of the organisms, the less proof wetend to require for common ancestry.
The extent of proof we implicitly requireis also influenced by the generality of thepattern and by the topographic relation ofthe structures being examined. Conse-quently, there is likely to be little questionbut that femur, humerus and tibia of rhinosand antelopes are homologous, eventhough the evidence is entirely structural.However, there is much less certainty aboutthe homologies of digital elements as thesedemonstrate "adjacency" on only one side.
The fact that we use criteria of overallsimilarity to recognize the likelihood ofhomology, indeed the operational reduc-tion of the required level of proof, is aptto suggest circularity of argument. How-ever, if the character being examined isgenerally new, it should not be consideredrepetitious of those previously examined.In many other cases the test may be of asingle component in a series; if the overallchain is homologous, the evidence forhomology of any one link is presumablystrengthened.
What if the organisms bearing the struc-tures being compared do not share suchclose overall similarity or if their exactplacement is uncertain? What if the affin-ities are uncertain because the comparisonextends across families, orders, classes, oreven phyla? It then becomes necessary totest for at least two kinds of additionalinformation beyond structural similarity.The first test applies to similarity of devel-opment, whereas the second test demandsthat phylogenetically intermediate organ-
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

294 CARL GANS
isms share the homologous characteristic(Remane's [1956] criterion of continuity).
Similarity of development has histori-cally implied similarity of embryonicsources (ectoderm, neural crest) and moreor less structurally and temporally parallelmodifications from these. It must be con-sidered whenever the observed patterndoes not quite match other observations.The second advanced test for homologyasks about the state of the compared char-acter in phylogenetically intermediateorganisms. (Of course, it is implied whenwe include the systematic placement oforganisms in the operational decisionsmentioned above.) Appearance in inter-mediate species of characters, or at least oftransitional stages of characters providessupport for the idea that the phenotypicaspects are indeed homologous. Even if theobserved states are not structurally inter-mediate between two characters, they mustat least represent variations on a particularstructural plan. This kind of logic was crit-ical in the development of the theory ofthe vertebrate ear ossicles, although thephylogenetic comparison mainly examinedembryonic stages. Comparison only of thejaw joints of alligator and seal would notby itself have permitted the argument thatquadrate and incus were homologous.
An example of a combined approach isgiven by the orbitosphenoid of amphisbae-nians. This bone is generally much largerand tends to have a shape somewhat dif-ferent from that in lizards with which theseanimals have often been placed. Onceembryonic stages of amphisbaenians couldbe examined, they indicated that theirorbitosphenoid bone starts development inmembrane rather than cartilage (as it doesin lizards), so that the two kinds of orbi-tosphenoid are quite probably non-homol-ogous (Bellairs and Gans, 1983). However,this finding was not "surprising" as non-congruence in many other characteristicspreviously had cast doubt on amphisbae-nian affinities (Gans, 1978); indeed thedevelopmental study was undertakenbecause of the likelihood that the natureof cranial development might provideanother characteristic to test the affinity ofthese groups (Bellairs and Kamal, 1981).
The logic of such an analysis is similar
to that involved in including members bothof rodents and of lagomorphs in any studyof mammalian diversification; althoughthey have similar incisors, their other char-acters are sometimes dissimilar. Suchexamples suggest that information aboutpreviously discovered diversity will indi-cate the kinds of organisms for which a newcharacter should always be determined.The establishment of a framework forfuture test may well be the greatest benefitthat the phylogenetic scheme offers theexperimentalist.
Simpson (1961, p. 78) defined homologyin a treatise on taxonomy, listing only thosepossibly homologous characters that sharesimilar structure. However, it often tendsto be left unstated, that only the evidencefor, not the theoretical existence of,homology requires similarity. There mustbe many situations in which homologousaspects have in evolutionary time becomedissimilar in structure, function and devel-opment. Homology often can be demon-strated even though, for instance, appear-ance and function have become markedlymodified from the ancestral state. Thereare even cases in which we see changes inthe genetico-developmental route by whichhomologous structures are formed. Selec-tion may affect each of the three criteriaby itself. Sometimes, more than one cri-terion is modified; however, modificationof all three remains possible. Of course,the demonstration that two structures, dis-similar by all three of the present criteria,were indeed homologous, might be diffi-cult and would have to rest on the dem-onstration of transitions in intermediateforms. Indeed, the argument of Haas andSimpson (1946) explicitly, and in my viewcorrectly, places the burden of proof uponthe argument for homology; this does havethe corollary that some homologous struc-tures will not be recognized as such.
These then are the operational ap-proaches for deciding when morphologicalsimilarity might be due to homology. Thesuggested standards indicate the desirabil-ity of a broad comparison utilizing a widerange of species. As long as the comparisonremains restricted to a single pair of species,it is impossible to decide that some partic-ular character shown in these two organ-
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

COMPARISON IN MASTICATION 295
isms is due to common ancestry rather thanto accident or function; indeed, this is sim-ilar to an attempt to resolve a set of simul-taneous equations of a number fewer thanthat of the unknown variables. Only a com-bination of phylogenetic and morpholog-ical criteria permits one to argue whetheror not particular aspects of two or moreotherwise dissimilar organisms are homol-ogous; i.e., more than two species have tobe sampled. Whatever the criteria, docu-mentation that two elements are homolo-gous permits a special kind of biologicalcomparison.
DECISIONS ABOUT HOMOPLASY
What about systems that are phenotyp-ically similar but not homologous (Fig. 2)?Categories of non-homologous similarityof phenotype have been lumped as homo-plasy (Wiley, 1981, p. 122; see definitionsin Northcutt, 1984). Homoplastic similar-ity may be due to random events or to func-tional shaping by adaptation to a commonenvironment. For our purposes, it is suf-ficient to note that certain aspects are nothomologous and leave it at that.
Function may have affected structures atdifferent times; it is necessary to separatecurrent performance of very similar func-tions and past functional influences thatthen generated structural similarity. Asseen above, there is the complication thatmembers of two phyletic lines in which aparticular structure had already becomemodified in different directions may havesecondarily become more similar becauseboth ancestors retained a genetico-devel-opmental mechanism that respondedequivalently to a common function. Thus,similar response of two lines of organismsto a particular environment (i.e., cooler cli-mate), might produce similar phenotypesas long as the underlying genetic mecha-nisms changed in parallel (i.e., leading toheavier fur and changed hair structure).Such parallel modification must haveoccurred often in tooth shape. Parallelismsare sometimes considered to represent akind of homoplasy (analogy); however, itseems more appropriate to consider themcases of homology.
Consequently, the idea of homoplasy,including a mixture of historical and cur-
rently adaptive and random processes (andeven mistaken assessments), represents anoperational morass that is logically difficultto deal with. The set of concepts is com-plicated by the fact that analogy or func-tional shaping of organisms being com-pared may not only have involvedstructures that have similar genetic back-grounds (i.e., homologous ones), but thosewith quite different backgrounds (i.e., theanalogous portion of the homoplastic ones).Mastication is a functional process; it is bestto consider the nature of functional simi-larity, initially independently of the geneticbackground of the component structures.
WHY STUDY FUNCTIONS?
Before discussing function we shouldrecognize that the word has at least twomeanings leading to some intrinsic confu-sion. Bock and von Wahlert (1965) rede-fined the word "function" to subsume anyphysical or chemical actions that the com-ponents of an organism's phenotype mightperform or exert. They also introducedthe term "biological role," applying thisterm to the adaptively significant subset ofthe functionally possible. (The term "bio-logical role" is unwieldy, and many still usethe word function with two different mean-ings. I here propose substitution of theshorter term "role" for the identical con-cept.) Most functional anatomy properlysets this aside, initially describing functionas the mechanically (physico-chemically)possible and relegating consideration ofrole to a separate study. However, the con-cept of role introduces a set of extremelyimportant ideas. Only the adaptively sig-nificant aspects are viewed by natural selec-tion and potentially affect the selectiveadvantage of the organism. Functions perse are of interest mainly in the utilizationof excessive constructions or potential pro-toadaptations for the rare cases in whichthey permit a particular individual toimprove its fitness.
Any biological study must have somepurpose; in other words there should besomething that we expect to learn by car-rying it out. For example, the reason thatwe have considered possible homologies haspresumably been that these tell us some-thing about the affinities among the ani-
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022
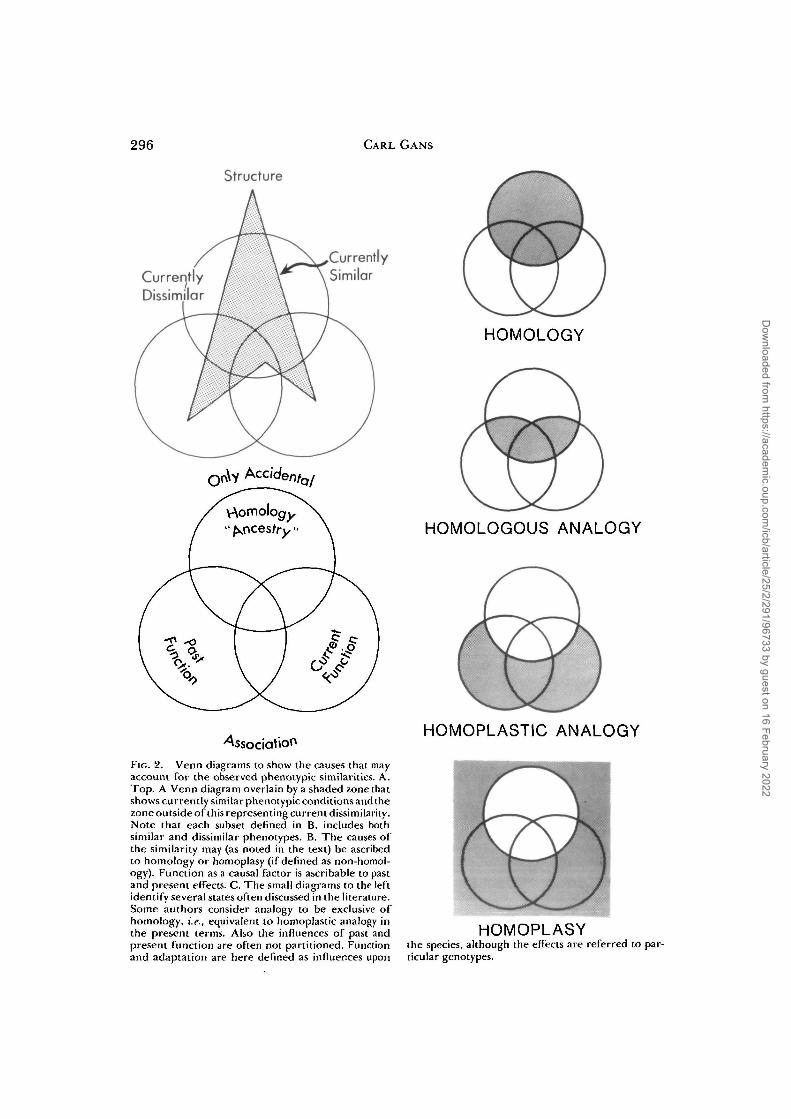
S96 CARL GANS
Structure
CurrentlyDissimilar
CurrentlySimilar
Fie. 2. Venn diagrams to show the causes that mayaccount for the observed phenotypic similarities. A.Top. A Venn diagram overlain by a shaded zone thatshows currently similar phenotypic conditions and thezone outside of this representing current dissimilarity.Note that each subset defined in B. includes bothsimilar and dissimilar phenotypes. B. The causes ofthe similarity may (as noted in the text) be ascribedto homology or homoplasy (if defined as non-homol-ogy). Function as a causal factor is ascribable to pastand present effects. C. The small diagrams to the leftidentify several states often discussed in the literature.Some authors consider analogy to be exclusive ofhomology, i.e., equivalent to homoplastic analogy inthe present terms. Also the influences of past andpresent function are often not partitioned. Functionand adaptation are here defined as influences upon
HOMOLOGY
HOMOLOGOUS ANALOGY
HOMOPLASTIC ANALOGY
HOMOPLASYthe species, although the effects are referred to par-ticular genotypes.
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

COMPARISON IN MASTICATION 297
mals studied. The establishment of homol-ogies ultimately provides a natural logic tothe nature of animal structures and theirsequential changes in history.
What shall be the aims of functional (asopposed to structural) comparison? Thefirst aim of these might be the determi-nation of functional usage patterns ofhomologous structures. A set of roles rep-resents a kind of natural history that tellsus something about diversity, not just oforganisms, but of their movement andaction patterns. Documentation of the rolesa structure allows in Recent species gen-erates a catalogue of such behaviors. Thiscatalogue may be used in turn to generatea set of hypotheses regarding changes ofrole in time; presumably the evolutionarysequence of functions and roles shouldreflect the sequence of adaptive factors thatgenerated the changes observed in thephenotypes.
The process of comparing sets of struc-tures and functions requires certain cave-ats. If two related organisms currently showdissimilar functions both of which areenabled by homologous but slightly dissim-ilar phenotypes, we might presumably askhow the changes in function (and presum-ably in role) relate to changes in the homol-ogous structures thus utilized. However,such "comparisons" should never be madein a facile fashion nor based upon the sit-uation in only two species. First of all, thereis a distinct difference between the assump-tion that current similarity of structurereflects current similarity of role and theassumption that the current similar struc-ture derives from a formerly similar role.In either case it is necessary to establish thefunctional patterns throughout the life his-tory of the organisms. The null hypothesismust be that the homologous characterswere modified by factors extraneous to thestudy and this hypothesis must be elimi-nated before any association is accepted.Secondly, the examination of adaptiveeffects on genetically very similar back-grounds risks confusion as both functionand ancestry would generate similarity.
The association of different functionswith homologous structures, includingthose in the CNS, has led to the descriptionof what may be called homologous actions
in the nervous system and homologousbehavior in ethology and physiology. Thisset of terms is likely to raise the hackles ofanyone raised in the old tradition in whichhomology referred to structure and anal-ogy to structures shaped by function. How-ever, if it is strictly applied the combinationdoes have merit in that it describes statesnot otherwise defined. Homologous func-tions (in ethology, physiology) then may beused to refer to present functions that areenabled by homologous structures andhomoplastic functions to those enabled bynon-homologous ones.
Actually, homoplastic functions helpgenerate the purest test for the adaptiveaction of physico-chemically determinedpressures of the environment. The extentto which multiple species deriving fromdisparate lineages show similarity afterexposure to seemingly similar environ-mental pressures should indicate the mag-nitude of these pressures. The restrictionof the similarity to the role-associatedaspects then provides a built-in test. Thewings of birds and bats are used for flying.The tails of some fishes and the wings ofcertain butterflies present the impressionof a vertebrate eye and may lead to a startlereaction in a potential predator. The sixthgill pouch and the hind gut each haveformed vascularized diverticulae that letdifferent species of fishes absorb oxygenfrom air. Many more examples can be cho-sen, involving the endocrines, water bal-ance, and temperature control, indeed,much of what is referred to as ecophysiol-ogy represents a search for various kindsof analogous conditions. Although we maynot wish to admit it, this aspect justifies thefunding for much of our research, theassumption being that the underlyingphysiological principles to be discovered inother animals, will also have bearing on thenature of man.
The discernment of underlying chemi-cal and physical constraints may be a majorreason for the study of similar, but non-homologous aspects of phenotypes. How-ever, there remain two problems. The firstis assurance that the aspects are indeed non-homologous, i.e., that we are not lookingat parallelisms (discussed above, Ridley,1983). The second is the major problem
MERNER-PFEIFFER LIBRARYTENNESSEE WESLEYAN COLLEGE
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

298 CARL GANS
of recognizing constraints. In theory, theirtightness may bear on the degree of match-ing or optimization between structure and"demand." In practice, we recognize theexistence of "factors of safety," themomentarily excessive constructions thatkeep structures congruent with function,but generally more than match thedemands of the latter at any moment. Inview of our uncertainty about the occur-rence of rare demands, we need to samplebroadly and attempt to extract the com-mon denominator from the slightly differ-ent adaptive demands that have shapedmany related species (Gans, 1983).
These then are some concerns that mustbe kept in mind whenever functional com-parisons are considered. Let us now moveto the operational standards for functionalcomparisons. I discuss these here, as thesuccess of both morphological and func-tional comparisons will be no better thanthe techniques employed. As the topic ofour symposium is mastication, I shall selectmy examples from the analysis of move-ment; however, optical, biophysical andbiochemical processes might be moreimportant in the study of the sensory ordigestive systems.
STANDARDS IN FUNCTIONAL COMPARISON
The clearly dissimilar tails of the teleostsand mammals called "dolphins" produceoscillatory movements that propel theseanimals through the water. Structuralcomparison of placement and architectureof the caudal fin disclosed an obvioushomoplasy, which is expressed in the direc-tion of its power stroke. What measure-ments must be made to permit tests of thesimilarity of function? Which tests must beapplied before one can claim that theobservations are reliable? What level ofprobability is required to confirm or falsifyconclusions?
In simplest terms, comparison of movingsystems may involve two levels, the kine-matic (movements) and the dynamic(forces). Description of the kinematic levelwill have to vary, depending on the com-plexity of the action. For simple mechan-ically constrained systems, kinematicdescription need only characterize the
movement (in three-dimensional space) ofa single point on each element, for instanceof the tip of the jaw or of the cusp of atooth for the mandible of a cat. However,for movements with more degrees of free-dom and for multi-jointed systems such asthe jaws of snakes, we must specify themovement of multiple points and condi-tion of intermediate components. Forinstance, one might describe the excursionof the fingers by characterizing only thedisplacement of the tips of each digit orone might also include information aboutthe position of each knuckle. The positionof intermediate elements may indeedbecome important in motion of the hyoidand tongue relative to those of the sym-physis and palate. In comparing move-ments, it is also critical to determinewhether the similarity is affected by theirvelocity or rate. Finally, one must establishwhether the activity has componentsbeyond the immediate ones and take theseinto account. Examples are the activity ofthe neck muscles in positioning food andperhaps in aiding the driving activity of theteeth.
Next one approaches dynamic aspects.There is the consideration of energy stor-age due to inertial or other effects, beyondwhich one asks about the nature and acti-vation pattern of the muscles driving thesystem. Muscle masses are far from homo-geneous and seemingly isomorphic onesmay contain drastically different popula-tions of motor units. Do these start to actat the same time and do they act for anequal portion of the movement cycle? Whatis the physiology and recruitment patternof their motor units? In short, how doesforce generation wax and wane? Also, whatare the levels of control and how do higherlevels of the nervous system recruit motorunits?
A physiologist's approach to functionwould examine the same movements andforces but interpret their actions in termsof the effects that the animal generates.Previously, the nature of the action, per-haps the force and stress levels that theanimal generates and can apply to the preyare analyzed. The physiological analysisconcentrates on the functional effects, for
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

COMPARISON IN MASTICATION 299
instance on the image of a mimicked pat-tern produced by a pigment array, thesprint speed and endurance produced by alocomotor system and the kinds of foodreduction achieved in mastication. Com-parison often shifts to functions and roles.Certainly these may be difficult to deter-mine; they are rarely obvious and need notinvolve mechanical or energetic factors.However, as selection is concerned withthis portion of function, the way environ-mental constraints affect structure shouldbe most obviously observed here. A phys-iological approach to mastication mayignore jaw movements and only comparewhether the food is cut or ripped, shred-ded or crushed, ground wet or dry.
It is useful also to consider experimen-tation. Not only should we examine animalresponse to the most obvious tasks but alsoto obscure ones, even to those that seem-ingly exceed the demands of their ecolog-ical range. The capacity to move by side-winding is in the repertoire of many snakesthat seemingly never do it in nature. Whatis known about the animal's normal food;how does it respond to aberrant food types?We should also consider the interaction ofthis step with the preceding and followingones; for instance how (and whether) doesthe presence or absence of rumination andfecal recycling affect mastication and mas-ticatory structures?
In characterizing any static and dynamicaspects of function, we should be able toapply standards of reproducibility approx-imating those of the physical sciences.Movements can be quantified, particle size(or surface area exposed to chemical diges-tion) may be measured, forces may berecorded. Not only quantification of activ-ity but the presentation of statistics aboutvariation is essential for meaningful com-parison.
Functional aspects are particularly proneto behavioral differences. Such differencesintroduce a kind of variability that is oftenset aside; for instance, students tend toreport the typical and most elegant resultsand ignore the less easily standardized (nottoo clean) events. It is precisely these thatmay give us important information aboutvariability (and the unexpected) and may
mask both behavioral and genetic differ-ences. The literature of animal behaviorcontains a body of statistical approaches tosuch comparison.
The current component of behaviorcharacterizes the way the animal respondsat the time of the experiment. If it is suf-ficiently hungry or thirsty, the animal willpresumably eat and drink, but so will a con-demned prisoner. However, such behaviorsometimes proves to be different frombehavior that would be observed wheneverorganisms act under more natural condi-tions. They may be induced to act morenormally either by isolating them so thatthe experiment proceeds by remote con-trol or by setting up a training programthat reduces psychological stress by con-ditioning them to associate the investigatorwith particularly desirable food items.
Furthermore, animal behavior cannot beunderstood only from a current perspec-tive; there is ample evidence that eventsearly in the life of the individual, such asimprinting and the acquisition of searchimages, may permanently change the fre-quency with which an organism performsparticular actions or may affect the waysuch actions are performed. One suspectsthat this behavioral plasticity is particularlyimportant in the feeding system, in whichthere must have been long-term selectionfor utilizing formerly "aberrant" but sud-denly common food types by samplingbeyond the strictly defined. Here again, weneed careful and perhaps statisticalapproaches to understand the nature andmeaning of functional variability and to seethe limits of behavioral/structural roles.
STANDARDS IN ELECTROMYOGRAPHICALCOMPARISON
Electromyography is becoming a com-mon approach in many studies of masti-cation. This suggests that it provides a goodexample for the kinds of comparisonsapplied to the data generated by a partic-ular technique. Also, it lets us consider whathas to be stated to permit evaluation of thecorrectness of this (or any other) approach.Obviously, mere notification that a seriesof muscles acts more or less at the sametime without description of technique or
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

300 CARL GANS
indication of variability is no longer suffi-cient. We need to know about the aspectsthat may have affected the signal and mayin turn affect biological conclusions.
The first series of caveats ought reallyto be assumed as good practice, but per-haps are still worth restating. The shape,size and position of electrodes used inmacro-myographic analysis affects the sig-nals; hence, they should be standardizedand these aspects reported as part of thedocumentation. Placement of electrodes inthe muscles should be documented relativeto the several levels of topographical land-marks (tendinous, sheets, varied attach-ments). Settings of filters and levels ofamplification affect the resulting signal;they should be given. Indeed the absolutevoltage represents a good indicator of thegeneral properties of the electrodes used(quite apart from the fact that changes involtage level may remind an investigatorduring the experiment that the electrodemay have moved within the animal).
Tape recorder properties, chart recorderresponse rates and computer sampling ratesare all significant and must be noted. Dem-onstration of an unretouched EMG recordgives an immediate indicator of signal tonoise ratio and the level of artefact; itsinclusion in papers is well worth the costof arguing with an editor. Finally one hopesfor some documentation of the fact thatthe signal indeed derives from the musclebeing studied.
Assuming that all of the above "house-keeping" procedures have been docu-mented, what can be compared? For whichaspects does similarity make sense? Assum-ing that the signals have not been clipped,the onset and cut-off of activity remain theeasiest and often very useful items. Thestart and stop of a signal may be comparedto that of a second one "it fires some sec-onds after," "firing terminates some sec-onds before." A better test may be the cor-relation of the signal with some portion ofa cyclic event, by statements such as "firingof A follows that of B by an interval equalto x per cent of its burst duration or y percent of its interburst duration." The latterstatement forces one to deal with changesin activation with changes in rate. Both
kinds of comparisons may be applied todifferences among muscles, differenceswithin reduction sequences, differencesamong food types and differences amongspecies.
Another level of comparison asks aboutthe change in recruitment pattern, indeedthe rise and fall of EMG activity. This maybe recorded as the area-under-the-curvewhich apparently has some documentedlevel of correlation with the activity pat-terns within the motoneurons. The halfdozen described methods of signal quan-tification differ in their effects and it is use-ful to note which was used. Some integra-tors generate artefacts under particularcircumstances; hence statements that therecords were "integrated" may not allowthe experiment to be repeated. However,if the rise and fall of signals are to be com-pared they should be quantified, "duringthe first x per cent of the activity interval,the amplitude peaked," "the median,mode, semi-interquartile interval was
There are also specialized measures, suchas counts of zero crossings and inflectionpoints, and measures of spike amplitudes,that often disclose some interesting andsurprising things about the way particularmuscles are recruited. Whatever the mea-sures, the description must be stated interms that permit repetition and compar-
ison.RATIONALE
We have now seen that the search forhomology has substantial utility in estab-lishing phylogenetic patterns and that itprovides a basis for studying the putativefunctional changes that occurred in his-tory. It should also have become clear thatfunctional comparisons, per se, have themerit of permitting analysis of adaptation;indeed this may be mirrored better in ahomoplastic analogy than in a homologousone.
Which comparisons prove to be signifi-cant in the end? The most important mes-sage derives from animal diversity andindicates that the test itself will have toprovide the answer. Observation may indi-cate how often which movements are used.Observation may indicate when move-
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

COMPARISON IN MASTICATION 301
merits are stereotyped, indeed analyses ofvariance provide excellent guides to levelsof control. Experiment may indicate thecloseness to which the stereotyped aspectsrepresent mechanical constraint or match-ing to roles rather than merely to function.The possibility that certain movements oractions are possible does not indicate thatthey are indeed important to the animals.Not all movements of which an animal iscapable are actually used a significant num-ber of times, aid the animal in its survivaland contribute to its fitness. The extent towhich events occur will differ amongrelated species even among members of apopulation. Consequently, it is impossibleto obtain it by inspection of the phenotypeand we must always format observations toelicit precisely this information.
Perhaps the most important componentof any comparative study is a judicious mixof preconceived notions and their experi-mental test. Subdivision of activities intophases and association of such phases withrole-associated aspects remains an impor-tant task. However, once some regularitiesare discovered they should not be assumedfor the next half-dozen animals and foodtypes but always tested against organismicvariability. Our task is to recognize howclosely the generalizations fit the functionsthat are observed.
ACKNOWLEDGMENTSThanks are due to G. Estabrook, G. C.
Gorniak and R. G. Northcutt for com-ments on the manuscript. The studies uponwhich these conclusions have been basedwere funded by NIH and NSF, mostrecently under NIH NS17017-2, DHEW-PHS-G-SROL DE 05112-03 and NSF DEB8121229.
REFERENCES
Bellairs, A. d'A. and C. Cans. 1983. The reinter-pretation of the amphisbaenian orbitosphenoid.Nature 302(5905):243-244.
Bellairs, A. d'A. and A. M. Kamal. 1981. The chon-drocranium and the development of the skull inRecent reptiles. In C. Gans and T. S. Parsons(eds.), Biology of the Reptilia. Vol. 11, pp. 1-265.Academic Press, Ltd., London.
Bock, W.J. and G. von Wahlert. 1965. Adaptationand the form-function complex. Evolution 19(3):269-299.
Gans, C. 1974. Biomechanics: Approach to vertebratebiology. J. P. Lippincott, Philadelphia; 1981. TheUniversity of Michigan Press.
Cans, C. 1978. The characteristics and affinities ofthe Amphisbaenia. Trans. Zool. Soc. London 34:347-416.
Gans, C. 1983. On the fallacy of perfection. In R. R.Fay and G. Gourevitch (eds.), Perspectives on mod-ern auditory research: Papers in honor ofE. G. Wever,(8): 101 — 114. Amphora Press, Groton, Connect-icut.
Haas, O. and G. G. Simpson. 1946. Analysis of somephylogenetic terms, with attempts at redefinition.Proc. Amer. Phil. Soc. 90:319-349.
Hailman.J. P. 1976. Uses of the comparative studyof behavior. In R. B. Masterton, W. Hodos, andH. Jerison (eds.), Evolution, brain and behavior.Persistent problems, pp. 13-22. Lawrence ErlbaumAssociates, Hillsdale, New Jersey.
Hildebrand, M. 1982. Analysis of vertebrate structure,2nd ed. John Wiley & Sons, Inc., New York.
Northcutt, R. G. 1984. Evolution of the vertebratecentral nervous system: Patterns and processes.Amer. Zool. 24:701-716.
Patterson, C. 1982. Morphological characters andhomology. In K. A. Joysey and A. E. Friday (eds.),Problems of phylogenetic reconstruction. SystematicsAssociation Special Volume 21, pp. 21-74. Aca-demic Press, London.
Remane, A. 1956. Die Grundlagen des naturlichen Sys-tems der vergleichenden Anatoime und Phylogenetik.Geest und Portik K. G., Leipzig.
Ridley, M. 1983. The explanation of organic diversity.The comparative method and adaptations for mating.Clarendon Press, Oxford.
Simpson, G. G. 1961. Principles of animal taxonomy.Columbia Univ. Press, New York.
Wiley, E. O. 1981. Phylogenelics. John Wiley & Sons,Inc., New York.
Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022

Dow
nloaded from https://academ
ic.oup.com/icb/article/25/2/291/96733 by guest on 16 February 2022
Related Documents











