Diatom derived dissolved organic matter as a driver of bacterial productivity: The role of nutrient limitation Romain Pete a,b, ⁎, Keith Davidson a , Mark C. Hart a , Tony Gutierrez a,c , Axel E.J. Miller a a Scottish Association for Marine Science, Scottish Marine Institute, Oban, PA37 1QA, Argyll, Scotland, UK b Laboratoire Ecolag, UMR 5119, Université Montpellier 2, Bat 24. Place Eugéne Bataillon, 34095 Montpellier cedex 5, France c Department of Environmental Sciences and Engineering, School of Public Health, CB 7431, University of North Carolina, Chapel Hill, NC 27599-7431, USA abstract article info Article history: Received 25 January 2010 Received in revised form 1 June 2010 Accepted 1 June 2010 Keywords: Bacterial production Diatoms DOM FISH Nutrient limitation Variable inorganic nutrient concentrations were used to generate silicate (Si) or nitrogen (N) limited conditions in cultures of the marine diatom Skeletonema costatum. Dissolved organic matter (DOM) harvested in the nutrient limited phase of these cultures was added to a natural bacterial community. Enhanced bacterial abundance and bacterial production were observed, in comparison to un-supplemented controls, when Si-limited (Si-DOM) rather than N-limited (N-DOM) diatom derived DOM or inorganic nutrients was added. This indicates that the bacterial population was limited by organic rather than inorganic resources but only Si-DOM had sufficient lability to alleviate this. Within the bacterial assemblage, a notable increase in the proportion of γ-Proteobacteria was evident on receipt of only Si-DOM. Assessment of the composition of the added DOM suggested that the observed dynamics were related to organic matter composition rather than molecular size as, within the Si- and N-DOM, the proportions of low and high molecular weight compounds were similar, but the polysaccharide and protein signatures were different. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Dissolved organic matter (DOM) is an important substrate for bacterial growth in aquatic ecosystems (Azam et al., 1994). DOM is mainly produced by phytoplankton (Carlson and Hansell, 2002), but may also result from riverine (Hedges et al., 1997) or atmospheric (Buatmenard et al., 1989) inputs, anthropogenic sources (Jickells, 1998), micro- and meso-zooplankton grazing (Strom et al., 1997) or viral lysis (Proctor and Fuhrman, 1992). Diatoms (Bacillariophyceae) are a particularly important component of the phytoplankton, contributing 20–25% of the global net marine primary production (Werner, 1977). The magnitude of the spring diatom bloom in coastal waters suggests that factors governing the quantity and quality of the DOM produced in the nutrient limited senescent phase of this event are likely to be of critical importance to the subsequent abundance, composition, and productivity of the bacterial communities. Nutrient stress has been shown to stimulate DOM excretion by phytoplankton (Maranon et al., 2005). Moreover, as such stress influences the chemical composition of phytoplankton (Obernos- terer and Herndl, 1995), it may influence the quality and quantity of DOM released (Puddu et al., 2003). For diatoms, Myklestad (1974, 1995) demonstrated that glucans (polymers of glucose) are common storage products, which provide a suite of bioavailable compounds (Hama et al., 2004). Glucans only accumulate in diatoms when growth is suboptimal (Myklestad, 1974), and are influenced by the form of nutrient limitation experienced by the population (Gilpin et al., 2004). For example, enhanced glucan concentrations are found under N-limitation. Under silicate limiting conditions, however, silicification processes within diatom cells are compromised leading to weaker cell walls and potentially “leaky” cells (Martin-Jézéquel et al., 2000), with the potential for enhanced DOM release. Hence, the form of nutrient limitation is likely to influence both the quantity and quality of the diatom produced DOM and, we hypothesise, the resultant productivity and composition of the bacterial populations utilising this organic matter. In Scottish and other temperate coastal waters, Skeletonema sp. are commonly the dominant spring bloom organisms (Tett, 1992). Hence, the period of their bloom cessation and decline might be expected to introduce significant DOM loading. The inorganic N:Si ratio within these waters is often close to one (Fehling et al., 2006) with both nutrients having the potential to be the limiting factor that arrests the spring diatom bloom. Our study, therefore, investigated the effect of N and Si-limitation on the release of DOM by the important diatom Skeletonema costatum and the influence of this material on the abundance, productivity and composition of a natural bacterial assemblage collected from coastal waters. Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26 ⁎ Corresponding author. Laboratoire Ecolag, UMR 5119, Université Montpellier 2, Bat 24. Place Eugéne Bataillon, 34095 Montpellier cedex 5, France. Tel.: + 33 4 67 14 47 32; fax: +33 4 67 14 37 19. E-mail address: [email protected] (R. Pete). 0022-0981/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.jembe.2010.06.002 Contents lists available at ScienceDirect Journal of Experimental Marine Biology and Ecology journal homepage: www.elsevier.com/locate/jembe

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
Contents lists available at ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r.com/ locate / jembe
Diatom derived dissolved organic matter as a driver of bacterial productivity:The role of nutrient limitation
Romain Pete a,b,⁎, Keith Davidson a, Mark C. Hart a, Tony Gutierrez a,c, Axel E.J. Miller a
a Scottish Association for Marine Science, Scottish Marine Institute, Oban, PA37 1QA, Argyll, Scotland, UKb Laboratoire Ecolag, UMR 5119, Université Montpellier 2, Bat 24. Place Eugéne Bataillon, 34095 Montpellier cedex 5, Francec Department of Environmental Sciences and Engineering, School of Public Health, CB 7431, University of North Carolina, Chapel Hill, NC 27599-7431, USA
⁎ Corresponding author. Laboratoire Ecolag, UMR 51124. Place Eugéne Bataillon, 34095 Montpellier cedex 5, Ffax: +33 4 67 14 37 19.
E-mail address: [email protected] (R. Pet
0022-0981/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.jembe.2010.06.002
a b s t r a c t
a r t i c l e i n f oArticle history:Received 25 January 2010Received in revised form 1 June 2010Accepted 1 June 2010
Keywords:Bacterial productionDiatomsDOMFISHNutrient limitation
Variable inorganic nutrient concentrations were used to generate silicate (Si) or nitrogen (N) limitedconditions in cultures of the marine diatom Skeletonema costatum. Dissolved organic matter (DOM)harvested in the nutrient limited phase of these cultures was added to a natural bacterial community.Enhanced bacterial abundance and bacterial production were observed, in comparison to un-supplementedcontrols, when Si-limited (Si-DOM) rather than N-limited (N-DOM) diatom derived DOM or inorganicnutrients was added. This indicates that the bacterial population was limited by organic rather thaninorganic resources but only Si-DOM had sufficient lability to alleviate this. Within the bacterial assemblage,a notable increase in the proportion of γ-Proteobacteria was evident on receipt of only Si-DOM. Assessmentof the composition of the added DOM suggested that the observed dynamics were related to organic mattercomposition rather than molecular size as, within the Si- and N-DOM, the proportions of low and highmolecular weight compounds were similar, but the polysaccharide and protein signatures were different.
9, Université Montpellier 2, Batrance. Tel.: +33 4 67 14 47 32;
e).
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Dissolved organic matter (DOM) is an important substrate forbacterial growth in aquatic ecosystems (Azam et al., 1994). DOM ismainly produced by phytoplankton (Carlson and Hansell, 2002), butmay also result from riverine (Hedges et al., 1997) or atmospheric(Buatmenard et al., 1989) inputs, anthropogenic sources (Jickells,1998), micro- and meso-zooplankton grazing (Strom et al., 1997) orviral lysis (Proctor and Fuhrman, 1992).
Diatoms (Bacillariophyceae) are a particularly important componentof the phytoplankton, contributing 20–25% of the global net marineprimary production (Werner, 1977). The magnitude of the springdiatom bloom in coastal waters suggests that factors governing thequantity and quality of the DOM produced in the nutrient limitedsenescent phase of this event are likely to be of critical importance to thesubsequent abundance, composition, and productivity of the bacterialcommunities.
Nutrient stress has been shown to stimulate DOM excretion byphytoplankton (Maranon et al., 2005). Moreover, as such stressinfluences the chemical composition of phytoplankton (Obernos-terer and Herndl, 1995), it may influence the quality and quantity of
DOM released (Puddu et al., 2003). For diatoms, Myklestad (1974,1995) demonstrated that glucans (polymers of glucose) are commonstorage products, which provide a suite of bioavailable compounds(Hama et al., 2004). Glucans only accumulate in diatoms whengrowth is suboptimal (Myklestad, 1974), and are influenced by theform of nutrient limitation experienced by the population (Gilpinet al., 2004). For example, enhanced glucan concentrations are foundunder N-limitation. Under silicate limiting conditions, however,silicification processes within diatom cells are compromised leadingto weaker cell walls and potentially “leaky” cells (Martin-Jézéquelet al., 2000), with the potential for enhanced DOM release. Hence, theform of nutrient limitation is likely to influence both the quantity andquality of the diatom produced DOM and, we hypothesise, theresultant productivity and composition of the bacterial populationsutilising this organic matter.
In Scottish and other temperate coastal waters, Skeletonema sp. arecommonly the dominant spring bloom organisms (Tett, 1992). Hence,the period of their bloom cessation and decline might be expected tointroduce significant DOM loading. The inorganic N:Si ratio withinthese waters is often close to one (Fehling et al., 2006) with bothnutrients having the potential to be the limiting factor that arrests thespring diatom bloom. Our study, therefore, investigated the effect of Nand Si-limitation on the release of DOM by the important diatomSkeletonema costatum and the influence of this material on theabundance, productivity and composition of a natural bacterialassemblage collected from coastal waters.

21R. Pete et al. / Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
2. Methods
The experiments were conducted in two stages. Initially, culturesof S. costatumwere grown to a post exponential phase under either Nor Si-limitation, with DOM produced collected by ultrafiltration.
In stage 2, this harvested DOM was added to freshly collectedsamples of natural marine bacteria. The subsequent changes inbacterial abundance, production and community composition thatoccurred in response to the added DOM were measured.
To quantify and characterise the DOM harvested in stage 1 foraddition in stage 2, the produced DOM material was analysed byHTCO (high temperature catalytic oxidation) and HPLC (high-performance liquid chromatography).
2.1. Stage 1: S. costatum cultures: generation and characterisation ofDOM
Axenic cultures of S. costatum, previously isolated from LochCreran, Scotland (CCAP, 1077/9), were grown in 10 l carboyscontaining autoclaved filtered seawater (8 l) with a slightly modifiedf/2 medium, with EDTA excluded to avoid an additional source ofdissolved organic carbon (DOC). Cells were maintained at 15 °C undera 12/12 light/dark photo-cycle (cool-white daylight spectrum tubes)and a photon flux density of 160 μmol m−2 s−1. All equipment wereboth acid washed and steam-sterilised (121 ° C; 15 min) prior to use.All experiments were conducted in duplicate. After inoculation theculture vessels remained closed systems with samples being removedby siphon into sterile syringes. All sampling was conducted in a class 2laminar flow cabinet. Lack of bacterial contamination was confirmedby fluorescence microscope enumeration of DAPI stained cells, asdescribed below.
To generate DOM from N and Si-limited phytoplankton, differentinitial concentrations of inorganic nitrate and silicate were used. ForN-limitation, a N:Si ratio of 1:3 (40 μM and 120 μM, respectively) wasused, whereas for Si-limitation, the N:Si ratio used was 3:1 (120 μMand 40 μM, respectively). Phosphorus and other trace elements wereadded in excess.
To determine when the stationary phase had been reached,samples for chlorophyll a (chl a) determination were collected dailyfrom the culture vessels. The samples were filtered onto 25 mm glassfibre filters (type A/E, Pall Corporation) and stored frozen prior toanalysis by fluorometry after extraction with 90% acetone. To identifythe point at which nutrient limitation (N or Si) had been reached,parallel determinations for dissolved nitrate, nitrite and ammonium(DIN), silicate (Si) and phosphate (P) were measured using a LachatQuickchem 8000 FI nutrient analyzer (Hach Lange, Colorado, USA).
2.1.1. Biomass determinationS. costatum cells were viewed using an inverted light microscope
(Axiovert 200, Zeiss) mounted with a digital camera (Axiocam, Zeiss)coupled to an image analysis software (Axiovision, Zeiss). Cell volumewas determined from measurements of cell length and width(Davidson et al., 2002). Live cell volume was obtained using aconversion factor of 1.33 from measurements made on Lugol fixedcells (Montagnes et al., 1994). The algorithms of Montagnes et al.(1994) and Menden-Deuer and Lessard (2000) were used to estimatecarbon biomass from volume, which were found to give identicalresults.
2.1.2. DOM harvest, ultrafiltration, quantification and characterisationDOM from the S. costatum cultures was harvested during the
nutrient limited stationary phase (after 14 days growth in bothnutrient regimes) by gravity filtration using pre-ashed GF/F filters(Whatman 0.4 μm nominal porosity) to remove particulate materialand transparent exopolymers (TEP). The resultant filtrate, pooledfrom duplicated cultures and referred to as the DOM fraction, was
sub-divided for further processing and analytical determinations, aswell as for use in the stage 2 experiments (see below).
A volume of the collected DOMwas subjected to tangential filtration(Schleicher & Schuell) using a 1 kDa membrane. DOC and dissolvedorganic nitrogen (DON) concentrations in the low molecular weight(LMW b1 kDa) and high molecular weight (HMW N1 kDa) fractionswere determined following treatment with 85% orthophosphoric acidof 10 ml samples. Samples were flame sealed in precombustedglass ampules prior to analysis by HTCO with coupled infraredand chemiluminescence detection (Alvarez-Salgado and Miller, 1998).
Polysaccharides were extracted from the DOM fraction by addingtwo volumes of cold ethanol and extensively dialyzing the precipi-tated material with deionised water (ELGA Ultrapure system, VeoliaWater, UK) prior to freeze drying. The polysaccharide extracts wereanalysed by high-performance liquid chromatography using anAgilent 1100 chromatograph (Agilent Technologies) equipped witha refractometer and diode-array UV detector. A PL gel filtrationchromatography column (Polymer Laboratories; 7.5×300 mm) wasused at 30 °C. The mobile phase was 0.1 M NaNO3 with a flow rate0.6 ml min−1. Dextran standards of Mr range 50–1400 kDa (Sigma)were used for molecular weight estimation.
The polysaccharide extracts were analysed for total carbohydratecontent following the method of Dubois et al. (1956) with glucose asstandard. Total protein concentrations were determined using theBCA protein assay kit (Sigma, St. Louis, MO) with bovine serumalbumin as the standard. Assays were performed on 1 to 4 mg ml−1
solutions of the polysaccharide extracts derived from Si- and N-limited phytoplankton.
2.2. Stage 2: DOM addition experiments
Seawater was collected from 10 m with a Niskin bottle from theScottish west coast fjord, Loch Creran (station C5, 56° 32.05′ N, 05°19.40′ W), then pre-screened with a 160-μm mesh net to removemacro-zooplankton, and the sample stored in a Nalgene carboy duringits rapid (b2 h) return to the laboratory. Water was further screenedby gravity filtration through GF/F filters to reduce the nanoflagellatedensity using our previous protocols (Lønborg et al., 2009a, 2010).
100 ml of the DOM fraction generated from both N- and Si-limitedS. costatum cultures in stage 1 was added (separately) to duplicate 1 lbottles containing 9 parts of the natural bacterial community.Inorganic nutrients as nitrate, silicate and phosphate were addedwith the DOM to prevent inadvertent inorganic limitation (10 μM,10 μM, and 1 μM, respectively) of bacterial growth. To provide controlcomparisons, two additional treatments received inorganic nutrientsonly (no DOM) or no nutrient addition. In the following section,treatments are referred to as (1) “Si-DOM”—in receipt of DOM fromSi-limited phytoplankton, (2) “N-DOM”—in receipt of DOM from N-limited phytoplankton, (3) “Inorganic”—in receipt of inorganicnutrients alone and (4) “control”—no additions made. All experimen-tal and sampling equipment were acid cleaned and rinsed withdeionised water to prevent any organic C or N contamination. Flaskswere incubated at 15 °C for 72 h and sampled every 24 h, after gentleshaking, for bacterial abundance (BA), bacterial production (BP), andbacterial community composition. Contamination by heterotrophic(HNF) and phototrophic (PNAN) nanoflagellates was assessed at thesame frequency. Inorganic and organic nutrient concentrations weredetermined at t=0 and t=48.
2.3. Enumeration of bacterial abundance, production, communitycomposition, and heterotrophic, phototrophic and nanoflagellateabundance
BP was determined from [methyl-3H] thymidine incorporationfollowing the protocol outlined in Davidson et al. (2007). Carbonequivalents were calculated using 30.2 fg C cell−1 determined for
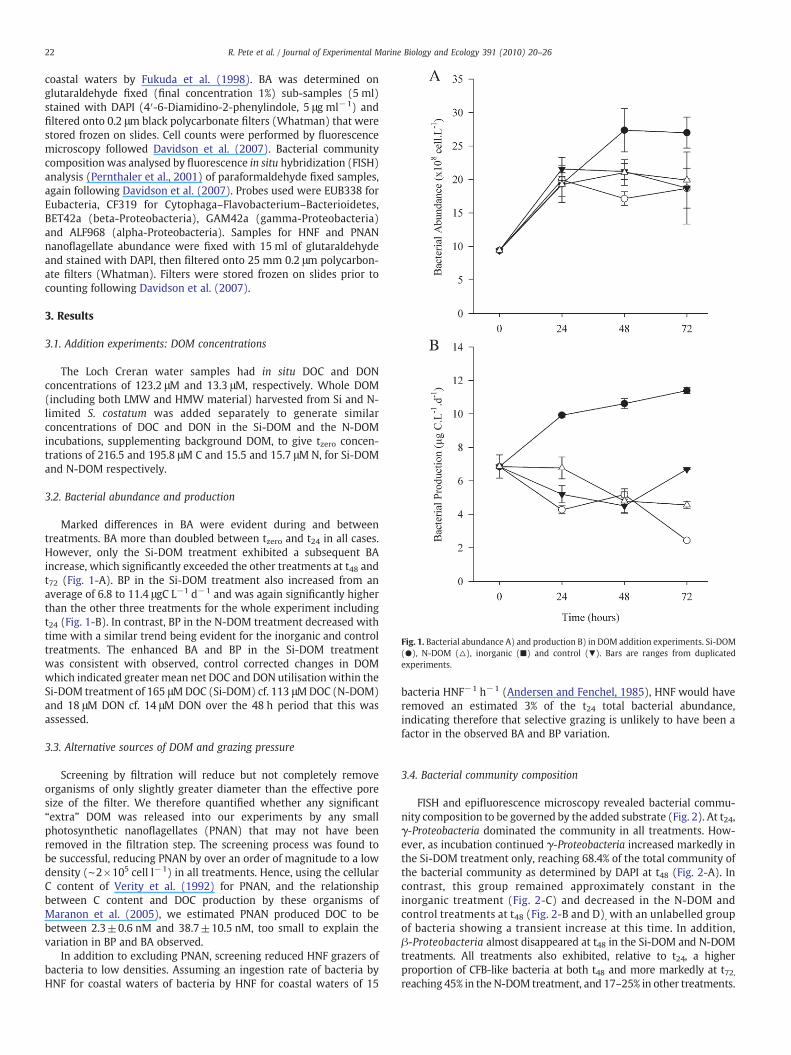
Fig. 1. Bacterial abundance A) and production B) in DOM addition experiments. Si-DOM(●), N-DOM (▵), inorganic (■) and control (▾). Bars are ranges from duplicatedexperiments.
22 R. Pete et al. / Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
coastal waters by Fukuda et al. (1998). BA was determined onglutaraldehyde fixed (final concentration 1%) sub-samples (5 ml)stained with DAPI (4′-6-Diamidino-2-phenylindole, 5 μg ml−1) andfiltered onto 0.2 μm black polycarbonate filters (Whatman) that werestored frozen on slides. Cell counts were performed by fluorescencemicroscopy followed Davidson et al. (2007). Bacterial communitycomposition was analysed by fluorescence in situ hybridization (FISH)analysis (Pernthaler et al., 2001) of paraformaldehyde fixed samples,again following Davidson et al. (2007). Probes used were EUB338 forEubacteria, CF319 for Cytophaga–Flavobacterium–Bacterioidetes,BET42a (beta-Proteobacteria), GAM42a (gamma-Proteobacteria)and ALF968 (alpha-Proteobacteria). Samples for HNF and PNANnanoflagellate abundance were fixed with 15 ml of glutaraldehydeand stained with DAPI, then filtered onto 25 mm 0.2 μm polycarbon-ate filters (Whatman). Filters were stored frozen on slides prior tocounting following Davidson et al. (2007).
3. Results
3.1. Addition experiments: DOM concentrations
The Loch Creran water samples had in situ DOC and DONconcentrations of 123.2 μM and 13.3 μM, respectively. Whole DOM(including both LMW and HMW material) harvested from Si and N-limited S. costatum was added separately to generate similarconcentrations of DOC and DON in the Si-DOM and the N-DOMincubations, supplementing background DOM, to give tzero concen-trations of 216.5 and 195.8 μM C and 15.5 and 15.7 μMN, for Si-DOMand N-DOM respectively.
3.2. Bacterial abundance and production
Marked differences in BA were evident during and betweentreatments. BA more than doubled between tzero and t24 in all cases.However, only the Si-DOM treatment exhibited a subsequent BAincrease, which significantly exceeded the other treatments at t48 andt72 (Fig. 1-A). BP in the Si-DOM treatment also increased from anaverage of 6.8 to 11.4 μgC L−1 d−1 and was again significantly higherthan the other three treatments for the whole experiment includingt24 (Fig. 1-B). In contrast, BP in the N-DOM treatment decreased withtime with a similar trend being evident for the inorganic and controltreatments. The enhanced BA and BP in the Si-DOM treatmentwas consistent with observed, control corrected changes in DOMwhich indicated greater mean net DOC and DON utilisation within theSi-DOM treatment of 165 μMDOC (Si-DOM) cf. 113 μMDOC (N-DOM)and 18 μM DON cf. 14 μM DON over the 48 h period that this wasassessed.
3.3. Alternative sources of DOM and grazing pressure
Screening by filtration will reduce but not completely removeorganisms of only slightly greater diameter than the effective poresize of the filter. We therefore quantified whether any significant“extra” DOM was released into our experiments by any smallphotosynthetic nanoflagellates (PNAN) that may not have beenremoved in the filtration step. The screening process was found tobe successful, reducing PNAN by over an order of magnitude to a lowdensity (∼2×105 cell l−1) in all treatments. Hence, using the cellularC content of Verity et al. (1992) for PNAN, and the relationshipbetween C content and DOC production by these organisms ofMaranon et al. (2005), we estimated PNAN produced DOC to bebetween 2.3±0.6 nM and 38.7±10.5 nM, too small to explain thevariation in BP and BA observed.
In addition to excluding PNAN, screening reduced HNF grazers ofbacteria to low densities. Assuming an ingestion rate of bacteria byHNF for coastal waters of bacteria by HNF for coastal waters of 15
bacteria HNF−1 h−1 (Andersen and Fenchel, 1985), HNF would haveremoved an estimated 3% of the t24 total bacterial abundance,indicating therefore that selective grazing is unlikely to have been afactor in the observed BA and BP variation.
3.4. Bacterial community composition
FISH and epifluorescence microscopy revealed bacterial commu-nity composition to be governed by the added substrate (Fig. 2). At t24,γ-Proteobacteria dominated the community in all treatments. How-ever, as incubation continued γ-Proteobacteria increased markedly inthe Si-DOM treatment only, reaching 68.4% of the total community ofthe bacterial community as determined by DAPI at t48 (Fig. 2-A). Incontrast, this group remained approximately constant in theinorganic treatment (Fig. 2-C) and decreased in the N-DOM andcontrol treatments at t48 (Fig. 2-B and D), with an unlabelled groupof bacteria showing a transient increase at this time. In addition,β-Proteobacteria almost disappeared at t48 in the Si-DOM and N-DOMtreatments. All treatments also exhibited, relative to t24, a higherproportion of CFB-like bacteria at both t48 and more markedly at t72,reaching 45% in the N-DOM treatment, and 17–25% in other treatments.

Fig. 2. Time course of bacterial taxonomic composition with DOM added to the Creran seawater (A) Si-DOM , (B) N-DOM, (C) inorganic, and (D) control, represented as percentage oftotal DAPI count. Legend is for α-Proteobacteria (black), γ-Proteobacteria (light vertical), β-Proteobacteria (light upward diagonal), Cytophaga–Flavobacterium–Bacterioidetes(outlined diamonds), Eubacteria (white) and undetected bacteria (grey).
23R. Pete et al. / Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
While γ-Proteobacteria remained the dominant group in all fourtreatments, community compositions at t72 was different from thatfound at t24 displaying a succession fromγ-Proteobacteriadomination toa balance between γ-Proteobacteria, CFB group and α-Proteobacteria.
3.5. Characterisation of the DOM produced by S. costatum
When the S. costatum cultures were harvested during the stationaryphase, DOC concentrations were 1055.6±9.6 μM and 849.3±3.5 μMfor Si- and N-limited cultures, respectively. DON concentrations were35.2±0.6 μM and 37.1±0.4 μM for Si- and N-limited cultures,respectively. If we normalise this liberated DOC and DON to cell carboncontent at peak yield, we find similar quantities of DOC (0.143 cf.0.125 mg C (mg cell C)−1) andDON(0.02 cf. 0.016 mgN(mg cell C)−1)to be generated per unit biomass in the two treatment conditions.
The relative proportion of LMW and HMW compounds within thisDOM was also similar in both nutrient regimes, with the majority ofproduced DOC composed of LMW compounds—i.e. 72% and 68% for Si-and N-limited cultures, respectively. A similar pattern was observedfor DON, with 59% and 69% of the DOM comprised of LMWcompounds for Si- and N-limited cultures, respectively.
Notwithstanding the similarities in the bulk composition mea-surements above, chromatography of DOM extracts revealed differ-ences in their chemical and molecular weight composition.Approximately 57.5% and 95% of the DOC produced by S. costatumin Si- and N-limited conditions, respectively, is of polysaccharidecomposition. As shown in Fig. 3-A, N-DOM contained two highmolecular weight components, as defined by two refractive index (RI)signals that eluted at 10.5 and 13.6 min. Using a plot of log Mr ofstandards versus retention time, the relative molecular mass of thesepeaks were estimated to be N1,400,000 and 330,000 Da respectively.Since the first peak eluted in the void volume (i.e. size-exclusion limitof the column; 1,400,000 Da) this peak could represent one or morehigh molecular weight species, the mass of which were unresolvableunder the conditions used. However, chromatography of the Si-DOM(Fig. 3-B) revealed a marked diminution of the peak signal denotingthe void volume (i.e. N1,400,000 Da MW species). No other peaksignals, with longer retention times, were identified in both the N- andSi-DOM.
Chemical analysis indicated that for both theN- and Si-DOM, only∼2%of these fractions were composed of a carbohydrate component.Total protein analysis revealed that the polysaccharide extract fromthe Si-DOM contained low yet discernible quantities of protein

Fig. 3. HPLC of DOMextracts fromA)N- and B) Si-limited S. costatum cultures on a PL-GFCcolumn. The figures represent a snapshot of theHPLC chromatogramwhere peaks specificof polysaccharides components were observed: no peaks were detected at longer elutiontimes. RI, refractive index.
24 R. Pete et al. / Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
(8.7 mg g−1 of total extract). No protein, however, was detected inthe N-DOM.
4. Discussion
S. costatum was selected as the source of DOM for this study asSkeletonema sp. frequently dominate the phytoplankton biomassduring the spring bloom in Loch Creran, and hence may be the singlemost important source of autochthonous DOM in these and othercoastal waters. The particular strain studied had previously beenisolated from Loch Creran, from where the bacterial population, towhich the DOM generated was added, was obtained.
4.1. Effect of DOM type on bacterial dynamics
The relative proportion of LMW and HMW DOM has beensuggested to be an index of its biological availability, with the LMWmaterial being expected to be more labile (Saunders, 1976; Keil andKirchman, 1999) although other authors (Amon et al., 2001) havedemonstrated that HMW compounds may also be more bioreactiveand available to bacteria. Although the same amount of total DOM (asa combination of LMW and HMW materials) was added in ourexperiments, we cannot comment on the relative bioavailability of thetwo fractions. However, the enhanced BP observed on receipt of Si-DOM suggests that Si-limitation induces the production of DOM fromS. costatum that is bacteriologically more labile compared to DOMformed during growth of this diatom under N-limitation, amendmentwith solely inorganic N and Si, or under nutrient starvation conditions.
In order to identify differences between the N- and Si-DOM thatmight explain its differential effect on BP, we used chemical andchromatographic procedures to characterise its composition. Wefound that polysaccharides composed a significant component of the
DOM from both N- and Si-limited S. costatum. Notably, theircontribution to the total DOM concentration was highest in the N-DOM where molecules ≥1400 kDa formed a major component.Considering that N-DOM exerted no significant effect to BP, we caninfer that it was either not readily available or refractory to biologicaldegradation, and thereby served as a poor substrate to the bacterialcommunity. Phytoplanktonmay continue to fix C for some time underN-limitation (Flynn et al., 1993; Davidson et al., 1999) leading to theproduction of large polysaccharide molecules (Myklestad, 1995).However, as the total organic matter content of the LMW and HMWpools were similar in both Si- and N-limited treatments, we wouldexpect other compounds (unidentified in our analysis) or physico-chemical differences between the DOM fractions to have influencedthe enhanced production of the bacterial community treated with theSi-DOM.
Further analysis of the N- and Si-DOM fractions showed thatproteins were only detected in the latter (8.7 mg g−1 of polysaccha-ride extract in the Si-DOM fraction). This may be explained by thepossible release of proteins outside of the diatom cells due to theformation of leaky frustules from low levels of silification under Si-limitation conditions (Martin-Jézéquel et al., 2000). N-limitation ofdiatom growth can lead to an increase in glucan production coupled tolow levels of protein synthesis (Myklestad, 1995), thereby explainingthe absence of any detected protein in the N-DOM fraction. C:N ratiosof the new DOM produced were 30 and 23 for Si-DOM and N-DOMrespectively, indicating a lack of N relative to C and consistent with thevery low concentrations of proteins in the DOM.
Collectively, our results suggest that the chemical composition ofS. costatum-derived DOM, rather than its molecular size, confers agreater influence upon the growth of bacterial assemblages. By natureof their refractory nature, large and generally more complex DOM canlimit bacterial growth, as has been reported for exopolysaccharidesproduced by micro-algae (Aluwihare and Repeta, 1999) and bacteria(Ogawa et al., 2001). While this organic matter “quality” hypothesis isconsistent with the treatment specific differences in polysaccharidesand proteins in our study, it is important to note that these polymerextracts were highly refractory to chemical analysis, with ∼97% beingunaccounted for in both conditions. This feature is not uncommon,and has been attributed to uronic acids (Bejar et al., 1996) orglycosidic linkages of hexosamines (Biermann, 1988), which canconfer polysaccharides with a high resistance to chemical degradation.Whether the high chemical refractory quality of these extractscorrelateswith their susceptibility to biological degradationwill requirefurther analysis. However, the differences in the characteristics of theDOM between treatments are consistent with the bacterioplanktondynamics observed in the Si- and N-DOM treatments, as discussedbelow.
4.2. Changes in bulk bacterial properties following DOM additions
The similarity in BA and BP between inorganic and controltreatments indicates that the addition of inorganic nutrients did notenhance bacterial growth, and hence, inorganic nutrients were notlimiting the community. Since the Si-DOM addition was the onlytreatment that enhanced the BP and BA in comparison to tzero, andconsidering that both the Si- and N-DOM treatments weresupplemented with inorganic nutrients, one may conclude thatbacterial metabolism was limited by the availability of organicmatter, and to which the Si-DOM was sufficiently labile to alleviatethis limitation. While bacterial respiration (which was not assessed)could conceivably have utilised any highly available compounds thatwere present in the N-DOM, the trends observed in BA and BP arestill consistent with the characterisation of the DOM above inindicating that there were greater quantities of biologically availablecompounds within the Si-DOM. Based on the change in DOC con-centrations, and assuming a conversion factor of 30.2 fg C cell−1

25R. Pete et al. / Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
(Fukuda et al., 1998), bacterial growth efficiency (BGE) wasestimated to be 10.4% and 6.4% in the Si-DOM and N-DOMexperiments, respectively. These values are consistent with previousestimates for coastal waters of 7 to 61% (DelGiorgio and Cole, 1998),suggesting that C is mainly respired.
4.3. Effect on bacterial community composition
The initial increase in dominance of γ-Proteobacteria in alltreatments was consistent with their ability to respond rapidly tonutrient additions (Puddu et al., 2003) and suggests that this group hasan initial competitive advantage when DOM exudates from Si-limiteddiatoms are available. Increased BP in the Si-DOM treatment at this (t24)time is almost certainly attributable to the γ-Proteobacteria rapidlyutilising labile components of the Si-DOM. The lack of a parallel increasein BA, in another treatment than Si-DOM, suggests that C demandfulfilled an increase in cell activity (andpotentially biomass) rather thancell division. Puddu et al. (2003) observed a similar trend when abacterial assemblage was amended with DOM produced in balancedgrowth by the diatom Cylindrotheca closterium.
Subsequently rapid changes of thebacterial community compositionoccurred, with increases in the relative and absolute abundances of theα-Proteobacteria and CFB group in all treatments, coinciding with adecrease in the γ-Proteobacteria. This is potentially due to the morelabile components of the DOM becoming depleted. Members of theCFB group and α-Proteobacteria are adept at utilising more complexmacromolecules and exudates from algal productivity (Rooney-Vargaet al., 2005) and are also abundant in media rich in particulate organicmatter (Delong et al., 1993). Hence, these groups may be activelyutilising the HMW or more complex DOM. Kirchman et al. (2004)described a similar evolution of bacterial properties with modificationsof DOM, suggesting that DOM may dictate modifications to therelative abundance of bacterial subgroups, which in turn, affect theDOM pool.
5. Conclusion
Literature often reports the production of DOM by phytoplanktonunder one limiting factor or none. Although more experimentalwork is needed to fully characterise this DOM and to confirm theseresults under different nutrient conditions, our results demonstrat-ed that the quality of the diatom produced DOM (from N or Si-limitation) can significantly affect the productivity and compositionof natural bacterial communities, and potentially have importantconsequences for microbial ecosystem dynamics. Further studieswill also be required to determine the role of phytoplankton derivedDOM in the food web as this material may provide a source ofnutrition for, as well as bacteria, other phytoplankton potentiallyincluding harmful species (Stolte et al., 2002; Davidson and Bresnan,2009). Furthermore, potentially taxonomically different groups ofattached bacteria on the surface of phytoplankton may utilise, oralter the generated DOM (Znachor and Nedoma, 2010). AlternativeDOM sources, such as those from heterotropic grazers (Lønborg etal., 2009b) further complicating the picture. Finally, potentiallyselective (Montagnes et al., 2008) grazing on bacteria by heterotro-phic nanoflagellates may influence the trophic transfer of organicmatter within the food web.
Acknowledgements
This work was funded by a SAMS research studentship (RP) and aSAMS Oceans 2025 research funding (KD). We thank E. Mitchell forexperimental assistance, and R. J. Leakey and D. Green for usefuldiscussions. [SS]
References
Aluwihare, L.I., Repeta, D.J., 1999. A comparison of the chemical characteristics ofoceanic DOM and extracellular DOM produced bymarine algae. Mar. Ecol. Prog. Ser.186, 105–117.
Alvarez-Salgado, X.A.,Miller, A.E.J., 1998. Simultaneous determination of dissolved organiccarbon and total dissolved nitrogen in seawater by high temperature catalyticoxidation: conditions for precise shipboard measurements. Mar. Chem. 62, 325–333.
Amon, R.M.W., Fitznar, H.P., Benner, R., 2001. Linkages among the bioreactivity,chemical composition, and diagenetic state of marine dissolved organic matter.Limnol. Oceanogr. 46, 287–297.
Andersen, P., Fenchel, T., 1985. Bacterivory by microheterotrophic flagellates inseawater samples. Limnol. Oceanogr. 30, 198–202.
Azam, F., Smith, D.C., Steward, G.F., Hagstrom, A., 1994. Bacteria–organic-mattercoupling and its significance for oceanic carbon cycling. Micro. Ecol. 28, 167–179.
Bejar, V., Calvo, C., Moliz, J., Diaz-Martinez, F., Quesada, F., Quesada, E., 1996. Effect ofgrowth conditions on the rheological properties and chemical composition ofVolcaniella eurihalina exopolysaccharide. Appl. Bioch. Biotech. 59, 77–86.
Biermann, C.J., 1988. Hydrolysis and other cleavages of glycosidic linkages inpolysaccharides. Adv. Carbohydr. Chem. Biochem. 46, 251–271.
Buatmenard, P., Davies, J., Remoudaki, E., Miquel, J.C., Bergametti, G., Lambert, C.E., Ezat,U., Quetel, C., Larosa, J., Fowler, S.W., 1989. Non-steady state biological removal ofatmospheric particles from the Mediterranean surface waters. Nature 340,131–134.
Carlson, C.A., Hansell, D.A., 2002. Production and removal processes. In: Carlson, C.A.,Hansell, D.A. (Eds.), Biogeochemistry of Marine Dissolved Organic Matter.Academic Press, pp. 91–151.
Davidson, K., Bresnan, E., 2009. Shellfish toxicity in UK waters: a threat to humanhealth? Environ. Health 8 (Suppl 1), S12, doi 10.1186/1476-069X-8-S1-S12.
Davidson, K., Wood, G., John, E.H., Flynn, K.J., 1999. An investigation of non steady statealgal growth. I. An experimental model ecosystem. J. Plank. Res. 21, 811–837.
Davidson, K., Roberts, E.C., Gilpin, L.C., 2002. The relationship between carbon andbiovolume in marine microbial mesocosms under different nutrient regimes. Eur. J.Phyco. 37, 501–507.
Davidson, K., Gilpin, L.C., Hart, M.C., Fouilland, E., Mitchell, E., Calleja, I.A., Laurent, C.,Miller, A.E.J., Leakey, R.J.G., 2007. The influence of the balance of inorganic andorganic nitrogen on the trophic dynamics of microbial food webs. Limnol.Oceanogr. 52, 2147–2163.
DelGiorgio, P.A., Cole, J.J., 1998. Bacterial growth efficiency in natural aquatic systems.Annu. Rev. Eco. Sys. 29, 503–541.
Delong, E.F., Franks, D.G., Alldredge, A.L., 1993. Phylogenetic diversity of aggregate-attached vs free-living marine bacterial assemblages. Limnol. Oceanogr. 38,924–934.
DuBois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, F., 1956. Colorimetricmethod fordetermination of sugars and related substances. Anal. Chem. Vol. 28 (3), 350–356.
Fehling, J., Davidson, K., Bolch, C., Tett, P., 2006. Seasonality of Pseudo-nitzschiaspp. (Bacillariophyceae) in western Scottish waters. Mar. Ecol. Prog. Ser. 323,91–105.
Flynn, K.J., Davidson, K., Leftley, J.W., 1993. Carbon–nitrogen relations during batchgrowth of Nannochloropsis oculata (Eustigmatophyceae) under alternating lightand dark. J. App. Phycol. 5, 465–475.
Fukuda, R., Ogawa, H., Nagata, T., Koike, I., 1998. Direct determination of carbon andnitrogen contents of natural bacterial assemblages in marine environments. Appl.Env. Micro. 64, 3352–3358.
Gilpin, L.C., Davidson, K., Roberts, E., 2004. The influence of changes in nitrogen: siliconratios on diatom growth dynamics. J. Sea Res. 51, 21–35.
Hama, T., Yanagi, K.,Hama, J., 2004.Decrease inmolecularweightofphotosynthetic productsof marine phytoplankton during early diagenesis. Limnol. Oceanogr. 49, 471–481.
Hedges, J.I., Keil, R.G., Benner, R., 1997. What happens to terrestrial organic matter inthe ocean? Org. Geochem. 27, 195–212.
Jickells, T.D., 1998. Nutrient biogeochemistry of the coastal zone. Science 281, 217–222.Keil, R.G., Kirchman, D.L., 1999. Utilization of dissolved protein and amino acids in the
northern Sargasso Sea. Aquat. Micro. Ecol. 18, 293–300.Kirchman, D.L., Dittel, A.I., Findlay, S.E.G., Fischer, D., 2004. Changes in bacterial activity
and community structure in response to dissolved organic matter in the HudsonRiver, New York. Aquat. Micro. Ecol. 35, 243–257.
Lønborg, C., Davidson, K., Álvarez-Salgado, X.A., Miller, A.E.J., 2009a. Bioavailability andbacterial degradation rates of dissolved organic matter in a temperate coastal areaduring an annual cycle. Mar. Chem. 113, 219–226.
Lønborg, C., Álvarez-Salgado, X.A., Davidson, K., Miller, A.E.J., 2009b. Production ofbioavailable and refractory dissolved organic matter by coastal heterotrophicmicrobial populations. Est. Coast. Shelf. Sci. 82, 682–688.
Lønborg, C., Álvarez-Salgado, X.A., Davidson, K., Martínez-García, S.A., Teira, E., 2010.Assessing the microbial bioavailability and degradation rate constants of dissolvedorganic matter by fluorescence spectroscopy in the coastal upwelling system of theRia de Vigo. Mar. Chem. 119, 121–129.
Maranon, E., Cermeno, P., Perez, V., 2005. Continuity in the photosynthetic productionof dissolved organic carbon from eutrophic to oligotrophic waters. Mar. Ecol. Prog.Ser. 299, 7–17.
Martin-Jézéquel, V., Hildebrand, M., Brzezinski, M.A., 2000. Silicon metabolism indiatoms: implications for growth. J. Phycol. 36, 821–840.
Menden-Deuer, S., Lessard, E.J., 2000. Carbon to volume relationships for dinoflagel-lates, diatoms, and other protist plankton. Limnol. Oceanog. 45, 569–579.
Montagnes, D.J.S., Berges, J.A., Harrison, P.J., Taylor, F.J.R., 1994. Estimating carbon,nitrogen, protein, and chlorophyll-a from volume in marine-phytoplankton.Limnol. Oceanog. 39, 1044–1060.

26 R. Pete et al. / Journal of Experimental Marine Biology and Ecology 391 (2010) 20–26
Montagnes, D.J.S., Barbosa, A.B., Boenigk, J., Davidson, K., Jürgens, K., Macek, M., Parry, J.,Roberts, E.C., Šimek, K., 2008. Selective feeding behaviour of free-living protists:views on, and avenues for, continued study. Aquat. Microb. Ecol. 53, 83–98.
Myklestad, S., 1974. Production of carbohydrates by marine planktonic diatoms.1.Comparison of 9 different species in culture. J. Exp. Mar. Biol. Ecol. 15, 261–274.
Myklestad, S., 1995. Release of extracellular products by phytoplankton with specialemphaisis on polysaccharides. Sci. Total Environ. 165, 155–164.
Obernosterer, I., Herndl, G.J., 1995. Phytoplankton extracellular release and bacterial-growth—dependence on the inorganic N–P ratio. Mar. Ecol. Prog. Ser. 116, 247–257.
Ogawa, H., Amagai, Y., Koike, I., Kaiser, K., Benner, R., 2001. Production of refractorydissolved organic matter by bacteria. Science 292, 917–920.
Pernthaler, J., Glockner, F.O., Schonhuber, W., Amann, R., 2001. Fluorescence in situhybridization (FISH) with rRNA-targeted oligonucleotide probes. Methods inMicrobiology, vol. 30, pp. 207–226.
Proctor, L.M., Fuhrman, J.A., 1992. Mortality of marine bacteria in response toenrichments of the virus size fraction from seawater. Mar. Ecol. Prog. Ser. 87,283–293.
Puddu, A., Zoppini, A., Fazi, S., Rosati, M., Amalfitano, S., Magaletti, E., 2003. Bacterialuptake of DOM released from P-limited phytoplankton. FEMS Micro. Ecol. 46,257–268.
Rooney-Varga, J.N., Giewat, M.W., Savin, M.C., Sood, S., LeGresley, M., Martin, J.L., 2005.Links between phytoplankton and bacterial community dynamics in a coastalmarine environment. Microb. Ecol. 49, 163–175.
Saunders, G., 1976. Decomposition in fresh waters. In: MacFadyen, J.A. (Ed.), The Role ofTerrestrial and Aquatic Organisms in Decomposition Processes. Blackwell, pp. 341–374.
Stolte, W., Panosso, R., Gisselson, L.-Â., Granéli, E., 2002. Utilization efficiency of nitrogenassociated with riverine dissolved organic carbon (N1 kDa) by two toxin-producingphytoplankton species. Aquat. Microb. Ecol. 29, 97–105.
Strom, S.L., Benner, R., Ziegler, S., Dagg, M.J., 1997. Planktonic grazers are a potentiallyimportant source of marine dissolved organic carbon. Limnol. Oceanogr. 42, 1364–1374.
Tett, P., 1992. The ecology of plankton in Scottish coastal waters. Proc. R. Soc. EdinburghSect B- Biol. Sci. 100, 27–54.
Verity, P.G., Robertson, C.Y., Tronzo, C.R., Andrews, M.G., Nelson, J.R., Sieracki, M.E.,1992. Relationships between cell-volume and the carbon and nitrogen-content ofmarine photosynthetic nanoplankton. Limnol. Oceanogr. 37, 1434–1446.
Werner, D., 1977. Contributions of the study of diatoms to biology. In: Werner, D. (Ed.),The Biology of Diatoms. Blackwell, Oxford, pp. 1–3.
Znachor, P., Nedoma, J., 2010. Importance of dissolved organic carbon for phytoplank-ton nutrition in a eutrophic reservoir. J. Plank. Res. 32, 367–376.
Related Documents











