Density loss and respiration rates in coarse woody debris of Pinus radiata, Eucalyptus regnans and Eucalyptus maculata Jens Mackensen a,1 , Ju ¨rgen Bauhus b, * a United Nations Environment Programme, Division of Policy Development and Law, P.O. Box 30552, Nairobi, Kenya b Australian National University, School of Resources, Environment and Society, Forestry Program and Cooperative Research Centre for Greenhouse Accounting, Canberra, ACT 0200, Australia Received 28 January 2002; received in revised form 6 September 2002; accepted 11 September 2002 Abstract This study compared field and laboratory decomposition rates of coarse woody debris (CWD) (. 10 cm diameter) from three tree species: Pinus radiata, Eucalyptus regnans, and Eucalyptus maculata. For this purpose, the density loss of logs on the ground sampled from chronosequences of sites following harvesting was determined using the water replacement technique. P. radiata logs were sampled 1, 2.5, 6, and 9 years following harvesting, and logs of E. regnans and E. maculata were collected from sites that were harvested 1, 3.5, 6.5, and 12 and 1.5, 6.5, and 11.5 years ago, respectively. In addition, the C/N ratio of wood was determined and current respiration rates of logs from these different age classes were measured through laboratory incubation. The times for loss of 95% of material (t 0.95 ) determined from density loss for these species were 24 years for P. radiata, 43 years for E. regnans, and 62 years for E. maculata. The decomposition rates of CWD derived from laboratory respiration were 6.1, 5.9 and 11.9 times higher than the decay rates from density loss in P. radiata, E. regnans, and E. maculata, respectively. This points to severe constraints of decomposition through adverse conditions in the field. The changes in respiration rates and C/N ratio with age of decaying logs indicated that the single component, negative exponential decay model could be applied satisfactorily only to P. radiata. In the case of the eucalypt species, substrate quality (expressed through respiration rates) declined in the oldest samples. This may be explained by the loss of rapidly decomposing sapwood and the retention of more decay-resistant heartwood. In these cases, a two-component model will be more suitable to describe the density loss of decaying wood. q 2003 Elsevier Science Ltd. All rights reserved. Keywords: Coarse woody debris; Wood density; Wood respiration; Decomposition 1. Introduction Recent estimates for CO 2 -emissions from land use changes and forestry activities are subject to large uncertainties (NGGIC, 1997a,b). These uncertainties have a range of sources, such as the area being cleared, carbon density of vegetation, fraction of cleared biomass which is burned, and the rate at which the remaining biomass decays. In the absence of better information, the release of carbon from land-use change and forestry activities through the decay of residual biomass is being calculated according to the Intergovernmental Panel on Climate Change (IPCC, 1997) default, which is equal to a linear decay of litter over 10 years. The IPCC default for decay of above- and below- ground litter appears to be a coarse estimate of actual carbon losses from decay. It does not consider the climatic and environmental variables controlling decay nor does it take into account other variables linked to decay such as size distribution of litter, density, chemical composition and the position in which litter is decaying. The problem of obtaining more reliable decay estimates lies in the long-term nature of the decay process, in particular of the coarse woody debris (CWD), which may constitute the majority of the residual biomass following land-use change and forestry activities. Few studies have determined the decay of individual pieces of wood over a long time (Arthur et al., 1993; Stone et al., 1998). To overcome this problem, many studies have assessed the decay of CWD from chronosequences, where the period for which wood was decaying was known (Grier, 1978; Graham 0038-0717/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved. PII: S0038-0717(02)00255-9 Soil Biology & Biochemistry 35 (2003) 177–186 www.elsevier.com/locate/soilbio 1 Tel.: þ254-2-62-4251; fax: þ 254-2-62-4324. * Corresponding author. Address: School of Resources, Environment and Society, Australian National University, Forestry Program, Canberra, ACT 0200, Australia. Tel.: þ 61-2-6125-2748; fax: þ61-2-6125-0746. E-mail addresses: [email protected] (J. Bauhus), jens. [email protected] (J. Mackensen).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Density loss and respiration rates in coarse woody debris of Pinus
radiata, Eucalyptus regnans and Eucalyptus maculata
Jens Mackensena,1, Jurgen Bauhusb,*
aUnited Nations Environment Programme, Division of Policy Development and Law, P.O. Box 30552, Nairobi, KenyabAustralian National University, School of Resources, Environment and Society, Forestry Program and Cooperative
Research Centre for Greenhouse Accounting, Canberra, ACT 0200, Australia
Received 28 January 2002; received in revised form 6 September 2002; accepted 11 September 2002
Abstract
This study compared field and laboratory decomposition rates of coarse woody debris (CWD) (.10 cm diameter) from three tree species:
Pinus radiata, Eucalyptus regnans, and Eucalyptus maculata. For this purpose, the density loss of logs on the ground sampled from
chronosequences of sites following harvesting was determined using the water replacement technique. P. radiata logs were sampled 1, 2.5, 6,
and 9 years following harvesting, and logs of E. regnans and E. maculata were collected from sites that were harvested 1, 3.5, 6.5, and 12 and
1.5, 6.5, and 11.5 years ago, respectively. In addition, the C/N ratio of wood was determined and current respiration rates of logs from these
different age classes were measured through laboratory incubation. The times for loss of 95% of material (t0.95) determined from density loss
for these species were 24 years for P. radiata, 43 years for E. regnans, and 62 years for E. maculata. The decomposition rates of CWD
derived from laboratory respiration were 6.1, 5.9 and 11.9 times higher than the decay rates from density loss in P. radiata, E. regnans, and E.
maculata, respectively. This points to severe constraints of decomposition through adverse conditions in the field. The changes in respiration
rates and C/N ratio with age of decaying logs indicated that the single component, negative exponential decay model could be applied
satisfactorily only to P. radiata. In the case of the eucalypt species, substrate quality (expressed through respiration rates) declined in the
oldest samples. This may be explained by the loss of rapidly decomposing sapwood and the retention of more decay-resistant heartwood. In
these cases, a two-component model will be more suitable to describe the density loss of decaying wood.
q 2003 Elsevier Science Ltd. All rights reserved.
Keywords: Coarse woody debris; Wood density; Wood respiration; Decomposition
1. Introduction
Recent estimates for CO2-emissions from land use
changes and forestry activities are subject to large
uncertainties (NGGIC, 1997a,b). These uncertainties have
a range of sources, such as the area being cleared, carbon
density of vegetation, fraction of cleared biomass which is
burned, and the rate at which the remaining biomass decays.
In the absence of better information, the release of carbon
from land-use change and forestry activities through the
decay of residual biomass is being calculated according to
the Intergovernmental Panel on Climate Change (IPCC,
1997) default, which is equal to a linear decay of litter over
10 years. The IPCC default for decay of above- and below-
ground litter appears to be a coarse estimate of actual carbon
losses from decay. It does not consider the climatic and
environmental variables controlling decay nor does it take
into account other variables linked to decay such as size
distribution of litter, density, chemical composition and the
position in which litter is decaying.
The problem of obtaining more reliable decay estimates
lies in the long-term nature of the decay process, in
particular of the coarse woody debris (CWD), which may
constitute the majority of the residual biomass following
land-use change and forestry activities. Few studies have
determined the decay of individual pieces of wood over a
long time (Arthur et al., 1993; Stone et al., 1998). To
overcome this problem, many studies have assessed the
decay of CWD from chronosequences, where the period for
which wood was decaying was known (Grier, 1978; Graham
0038-0717/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved.
PII: S0 03 8 -0 71 7 (0 2) 00 2 55 -9
Soil Biology & Biochemistry 35 (2003) 177–186
www.elsevier.com/locate/soilbio
1 Tel.: þ254-2-62-4251; fax: þ254-2-62-4324.
* Corresponding author. Address: School of Resources, Environment and
Society, Australian National University, Forestry Program, Canberra, ACT
0200, Australia. Tel.: þ61-2-6125-2748; fax: þ61-2-6125-0746.
E-mail addresses: [email protected] (J. Bauhus), jens.
[email protected] (J. Mackensen).

and Cromack, 1982; Fahey, 1983; Busse, 1994). A major
difficulty with this approach is in the assigning of
decomposition periods to certain logs, in particular where
dendrochronological approaches cannot be employed.
Decaying logs have to be traced back to certain harvesting
or disturbance events. In a particular stand, however,
different logs may have resulted from different events,
logs on the forest floor might have started to decay while
standing and alive, and they might not have decayed in this
position, e.g. on the ground or suspended, for the entire
period. In addition, it may be difficult to assign decaying
logs to tree species in mixed-species forest stands.
In this study we wanted to investigate how reliable decay
estimates of CWD were by assessing mass loss of three
different tree species sampled from chronosequences. For
this purpose we also determined the respiration rates and the
carbon-to-nitrogen ratios of the decaying wood. While the
C-to-N ratio can be regarded as a measure of the decay
status of litter, the respiration rate can be viewed as an
indicator of substrate availability to decay organisms, and as
such as a measure of substrate quality (Swift, 1977; Swift
et al., 1979). Nitrogen limits wood-decomposing micro-
organisms (Cowling, 1972). Therefore N is immobilised by
microbes during the wood decomposition process while C is
released through respiration (Harmon et al., 1986). As a
consequence the C-to-N ratio declines, and can thus be used
as an indicator of the decomposition stage within a given
wood type. We assumed that a comparison of mass loss
(wood density decrease), respiration rate, and C-to-N ratio
would permit the assessment of the reliability of mass loss
measurements. Respiration rates should also indicate the
applicability of constant decay factors in models of CWD
decomposition for these species.
2. Material and methods
2.1. Study sites
We measured decomposition rates of CWD using two
complementary methods: wood density decrease (method
A) and CO2 emission rate (method B). Both methods were
applied to samples of three tree species: Monterey pine
(Pinus radiata D. Don), Mountain Ash (Eucalyptus regnans
F. Muell.) and Spotted Gum (E. maculata Hook.). These
species represent important forest ecosystems in south-
eastern Australia. They also represent low-, medium- and
high-density wood species, respectively. Details about
sample area, climate and management systems are provided
in Table 1.
P. radiata samples from a chronosequence following
harvesting, including 1, 2.5, 6, and 9 years, were collected
from similar sites within the Australian Capital Territory.
The external cylindrical shape of log sections sampled was
intact. This was not representative of the sites from which
they were sampled, because it is a standard practice to crush
the woody residues following harvesting to facilitate site
preparation and planting. However, this mechanical frag-
mentation was not specifically addressed in our study.
E. regnans samples were collected from sites in the
Toolangi district, Victoria, that were clear-fell harvested 1,
3.5, 6.5, and 12 years ago. The regeneration practice in these
forests is clearfelling followed by a hot burn of logging
residues. Care was taken to avoid sampling charred
material. This could not be avoided in all cases. However,
the density of charred samples was not significantly
different from unburned samples.
E. maculata samples were collected from North Broo-
man State Forest, South Coast of NSW. The different sites
were harvested 1.5, 6.5, and 11.5 years ago. Sampling was
concentrated on pure patches of E. maculata in otherwise
mixed forests. Initially it was intended to sample older age
classes as well, but it was not possible to determine the
species with certainty in the field once the bark had fallen
off. For the ages that were sampled in this study, the logs
could be linked to stumps, that were either still carrying the
bark or had coppiced which allowed species identification.
2.2. Sample processing and determination of wood density
and respiration rate
Method A included sampling of cross-sections of CWD
of known age since clearing and determination of wood
density per disc. By establishing a chronosequence of sites
since disturbance, the decrease in wood density or weight
loss compared to initial density was used to calculate
decomposition rates. To reduce the variability of decay
stages within age-cohorts, samples were only collected from
logs that were positioned on the soil surface. Two cross
sections of approx. 5 cm width were sampled within a
distance of 0.3–1 m of each other per log. Tree disks were
cut using a chainsaw. Between 16 and 22 logs ranging in
diameter from 10 to 30 cm were sampled per species and
age class (Table 1).
Samples were stored in polyethylene bags for up to 5
days during fieldwork and were later stored cool (4 8C) until
further processing. Bark, mosses, fungal fruiting bodies and
soil were removed from samples prior to measurements.
Fresh weight and diameter of each disk was measured and
its volume was determined gravimetrically by water
displacement (Krankina and Harmon, 1995). Samples
were then oven-dried at 105 8C to a constant weight.
Water content and wood density were calculated
subsequently.
P. radiata samples of the two oldest age classes were
often fragile. Therefore one block of 25 cm in length was
sampled per log. To avoid disintegration this section was
wrapped with adhesive tape prior to cutting. For these
samples the volume was determined by measuring the exact
length and diameter at both ends, from which the cylindrical
volume could be calculated. Although this method neglects
decomposition due to fragmentation and biological
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186178

transformation, it is considered a standard method for
estimation of decomposition rates (Healey and Swift, 1971;
Christensen, 1984).
To measure respiration of wood samples (Method B),
five log sections per age class and species were collected for
incubation. These samples were log sections that were cut
out between the disks collected for method A. Incubation
samples had an average length of 30 cm and diameters
between 10 and 30 cm. Storage of samples was the same as
for the wood density samples. Water content and wood
density of these samples were derived from the accompany-
ing cross-section samples (see method A).
Samples were kept moist and were incubated in airtight
bags (REXAM, one side metallised, biaxially orientated
polypropylene film) at constant temperature (25 8C) in
controlled-climate chambers. Samples were incubated for
three days and CO2 emissions were determined using soda
lime. This method is well established for soil respiration
measurements (Edwards, 1982). Dried soda lime (20 g) was
placed into the incubation bag with the sample. Before and
after incubation, the soda lime was dried at 105 8C and
weighed. The gain in weight after incubation equalled the
adsorbed amount of CO2, corrected for the chemically
released water. The factor applied to correct for the released
water was 1.69. From this the amount of C released per unit
of mass remaining was calculated.
The C and N concentrations in wood were determined
using the combustion furnace technique of an automated
C/N-analyser (LECO-CHN-2000). Prior to analysis the
samples were finely ground and oven-dried at 60 8C for
24 h.
2.3. Calculations and statistics
Various mathematical models have been used to describe
decomposition patterns. The single-exponential model (Eq.
(1)) is based on the assumption that the decomposition rate
is proportional to the amount of matter remaining. It is the
most common model used to describe decomposition
patterns (Jenny et al., 1949; Olson, 1963; Wieder and
Lang, 1982; Means et al., 1985; Harmon et al., 1986;
Lambert et al., 1980). Other models like the multiple-
exponential model, lag time model or linear model exist but
are less frequently used (for discussion see Mackensen and
Bauhus (1999)).
X ¼ X0 e2kt ð1Þ
where X is for e.g. present wood density, X0 is initial wood
density, k is decomposition constant, t is the time. This type
of model was fitted to the mass loss data.
Assuming an exponential decay model, the time to
decompose 50% (t0.5) or 95% (t0.95) of CWD can be
calculated according to Eqs. (2) and (3) (Olson, 1963):
t0:5 ¼ 2lnð0:5Þ=k ¼ 0:693=k ð2Þ
Table 1
Location, climate, stand history and number of samples (n ) for sampled stands of P. radiata, E. regnans and E. maculata
P. radiata
Sample region ACT Canberra, Pierces Creek and Tidbinbilla Forestry Districts (358250S,
1488550E)
Climate Median prec. 791 mm yr21, mean daily min-max temp. 7.3 8C/19.5 8C
Sampling date 21/6–24/6/1999
Number of samples Age-class 1 20 logs @ 2 discs per log ðn ¼ 40Þ
Age-class 2.5 20 logs @ 2 discs per log ðn ¼ 40Þ
Age-class 6 20 logs @ 1 block (25 cm) per log ðn ¼ 20Þ
Age-class 9 20 logs @ 1 block (25 cm) per log ðn ¼ 20Þ
E. regnans
Sample region Central Highlands, Victoria, Toolangi DNRE Forest District (378330S,
1458270E)
Climate Median prec. 1300–1500 mm yr21, mean daily min–max temp. 4 8C/23 8C
Sampling date 26/5–29/5/1999
Number of samples Age-class 1 20 logs @ 2 discs per log ðn ¼ 39Þ
Age-class 3.5 20 logs @ 2 discs per log ðn ¼ 33Þ
Age-class 6.5 20 logs @ 1 block per log ðn ¼ 62Þ
Age-class 12 20 logs @ 1 block per log ðn ¼ 37Þ
E. maculata
Sample region South Coast, New South Wales, North Brooman State Forest (358300S,
1508150E)
Climate Median prec. 1100 mm yr21, mean daily min–max temp. 11.2 8C/21.2 8C
Sampling date 28/6–30/6/1999
Number of samples Age-class 1.5 22 logs @ discs per log ðn ¼ 44Þ
Age-class 6.5 16 logs @ discs per log ðn ¼ 31Þ
Age-class 11.5 18 logs @ discs per log ðn ¼ 36Þ
Samples of P. radiata and E. regnans were collected on sites that had been clear-felled previously. E. maculata samples were from selectively logged sites,
with documented harvesting history.
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186 179
t0:95 ¼ 2lnð0:05Þ=k ¼ 2:996=k ð3Þ
To test for differences between respiration rates and C-to-N
ratios of wood samples of different age, analyses of variance
were performed. Welch’s ANOVA was used in case the
variances were unequal for the different age classes (SAS
Institute, 1996). Multiple linear regression analysis was
used to explain relations between different parameters
including respiration rates, age, size, density, moisture
content and C/N ratio. Only significant (P , 0.05)
regressions are being reported.
To compare the annual decay rates determined from
density loss with daily respiration rates determined in short
term incubations, the annual decay rates were divided by
365 to convert them to daily rates. No correction factor to
convert density loss to C loss was required for this
comparison, because the C concentration in CWD of all
species remained constant over the age classes. Thus C is a
constant proportion of both remaining and lost mass. Since
decomposition processes are temperature-dependent, differ-
ences in temperature between the field and the laboratory
also have to be considered. Burger and Pritchett (1984)
provided the following formula for this:
k2 ¼ k110{logQ10=½10=ðT22T1Þ�} ð4Þ
where k2 is the desired mineralisation rate constant for the
temperature T2 (in this case the average annual temperature
at field sites), and k1 is the known mineralisation rate
constant determined at the temperature T1. We used a Q10
of 2.53 for our calculations. This is based on the analysis of
the temperature effect on decay rates of CWD (Mackensen
et al., 2003). It means that the decay rate is accelerated by
this factor with an increase in temperature of 10 8C.
Unfortunately, we could not make any corrections for
differences in water content between the incubated
samples and logs undergoing drying in re-wetting cycles
in the field.
3. Results
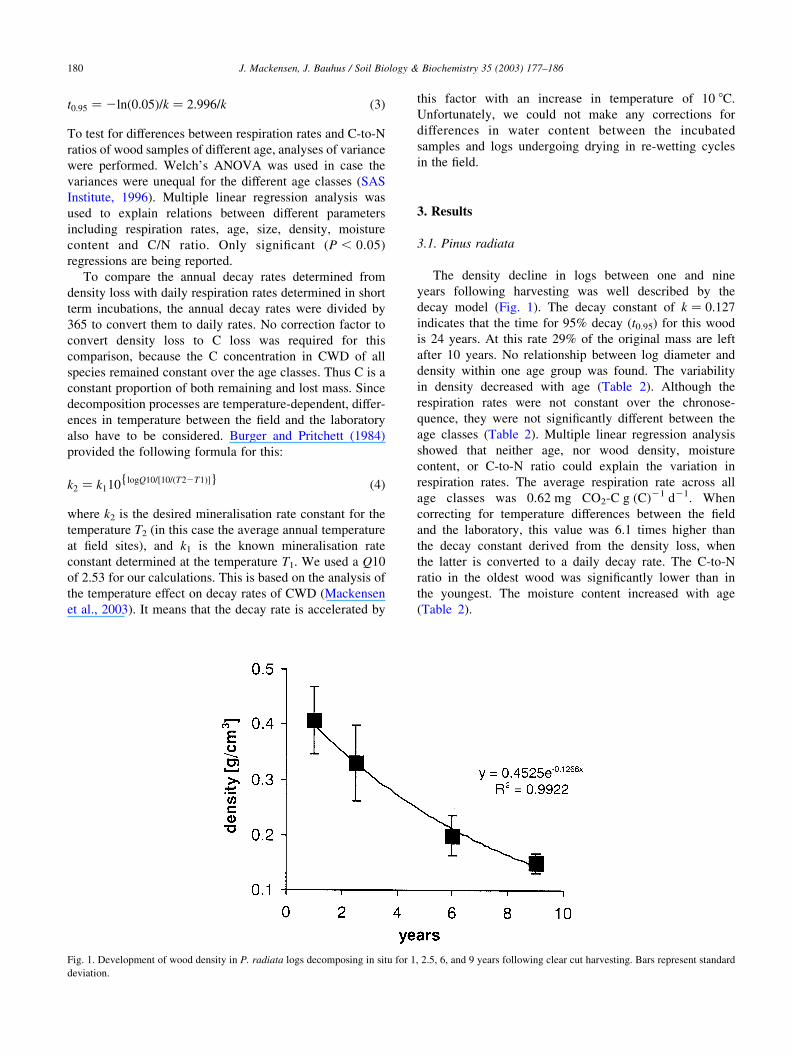
3.1. Pinus radiata
The density decline in logs between one and nine
years following harvesting was well described by the
decay model (Fig. 1). The decay constant of k ¼ 0:127
indicates that the time for 95% decay (t0.95) for this wood
is 24 years. At this rate 29% of the original mass are left
after 10 years. No relationship between log diameter and
density within one age group was found. The variability
in density decreased with age (Table 2). Although the
respiration rates were not constant over the chronose-
quence, they were not significantly different between the
age classes (Table 2). Multiple linear regression analysis
showed that neither age, nor wood density, moisture
content, or C-to-N ratio could explain the variation in
respiration rates. The average respiration rate across all
age classes was 0.62 mg CO2-C g (C)21 d21. When
correcting for temperature differences between the field
and the laboratory, this value was 6.1 times higher than
the decay constant derived from the density loss, when
the latter is converted to a daily decay rate. The C-to-N
ratio in the oldest wood was significantly lower than in
the youngest. The moisture content increased with age
(Table 2).
Fig. 1. Development of wood density in P. radiata logs decomposing in situ for 1, 2.5, 6, and 9 years following clear cut harvesting. Bars represent standard
deviation.
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186180
3.2. Eucalyptus regnans
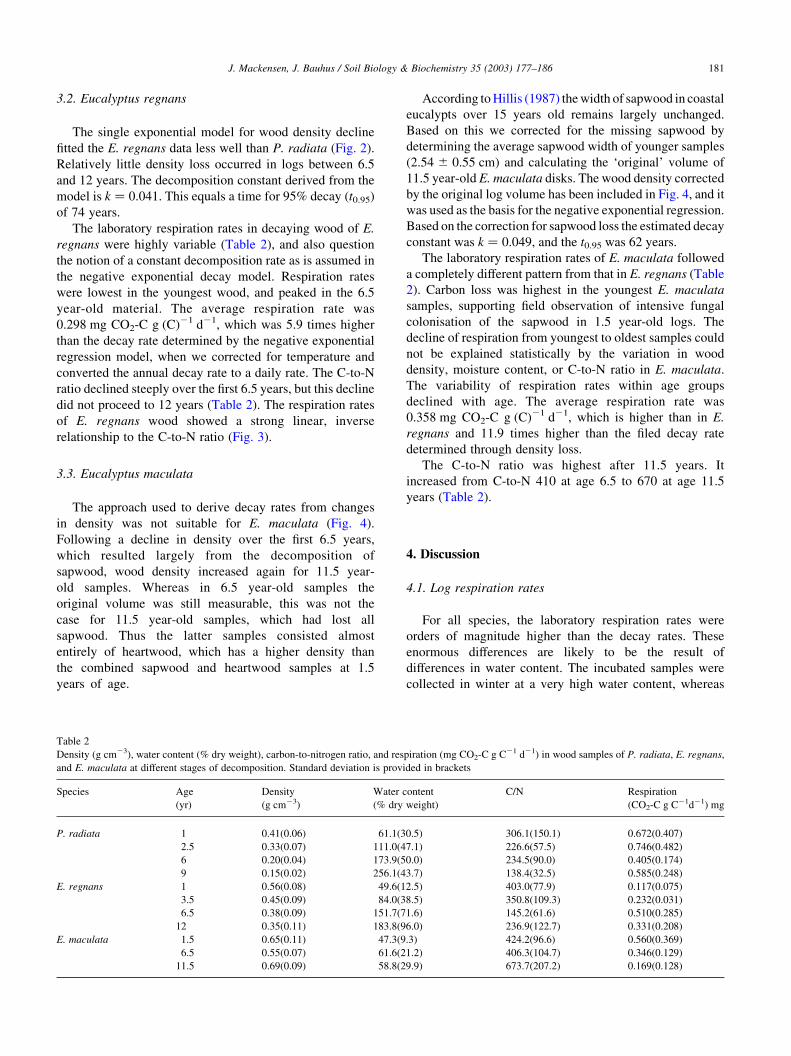
The single exponential model for wood density decline
fitted the E. regnans data less well than P. radiata (Fig. 2).
Relatively little density loss occurred in logs between 6.5
and 12 years. The decomposition constant derived from the
model is k ¼ 0:041: This equals a time for 95% decay (t0.95)
of 74 years.
The laboratory respiration rates in decaying wood of E.
regnans were highly variable (Table 2), and also question
the notion of a constant decomposition rate as is assumed in
the negative exponential decay model. Respiration rates
were lowest in the youngest wood, and peaked in the 6.5
year-old material. The average respiration rate was
0.298 mg CO2-C g (C)21 d21, which was 5.9 times higher
than the decay rate determined by the negative exponential
regression model, when we corrected for temperature and
converted the annual decay rate to a daily rate. The C-to-N
ratio declined steeply over the first 6.5 years, but this decline
did not proceed to 12 years (Table 2). The respiration rates
of E. regnans wood showed a strong linear, inverse
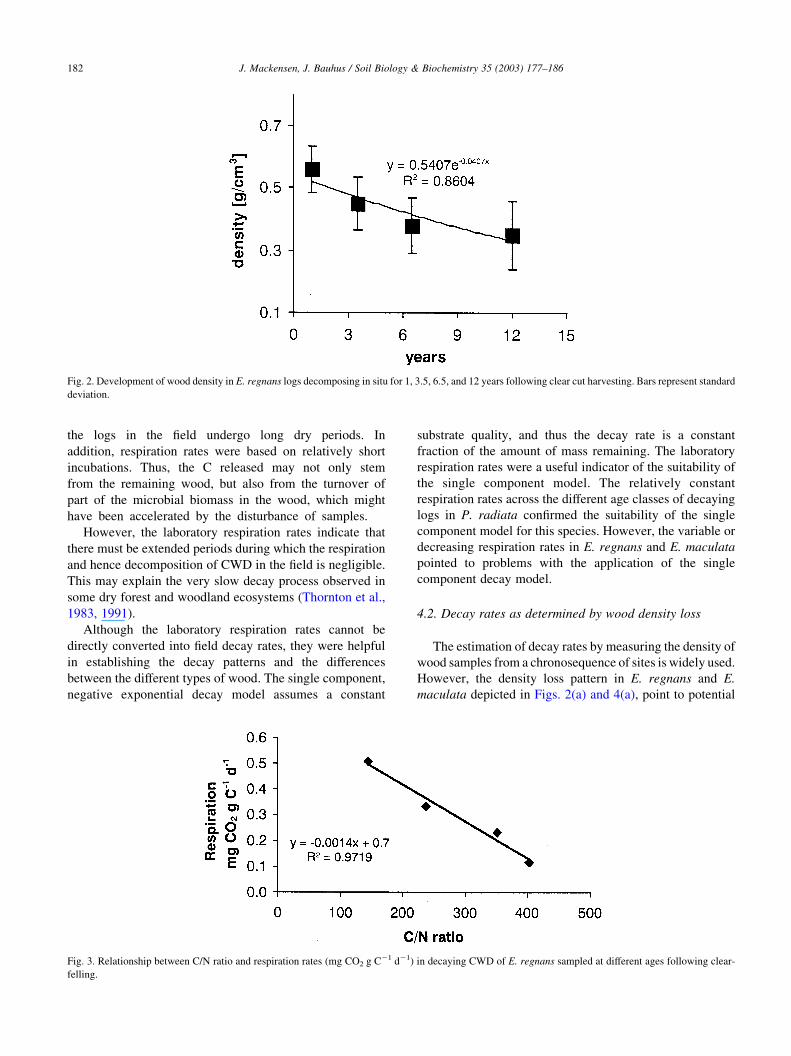
relationship to the C-to-N ratio (Fig. 3).
3.3. Eucalyptus maculata
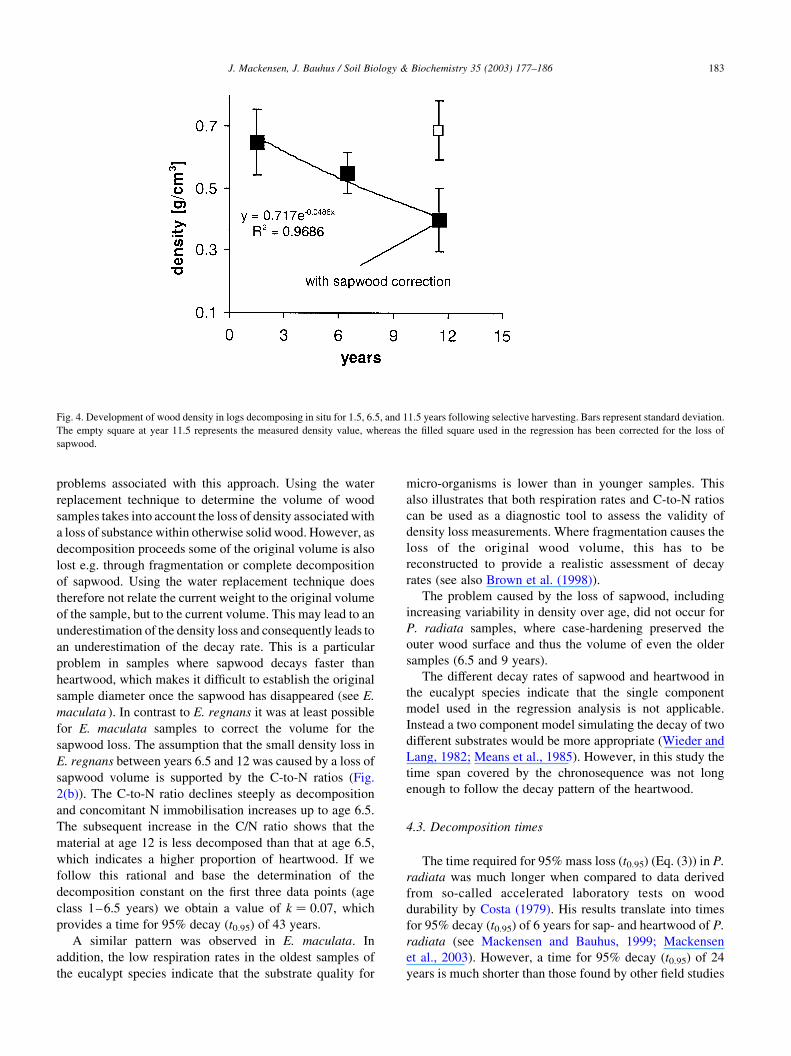
The approach used to derive decay rates from changes
in density was not suitable for E. maculata (Fig. 4).
Following a decline in density over the first 6.5 years,
which resulted largely from the decomposition of
sapwood, wood density increased again for 11.5 year-
old samples. Whereas in 6.5 year-old samples the
original volume was still measurable, this was not the
case for 11.5 year-old samples, which had lost all
sapwood. Thus the latter samples consisted almost
entirely of heartwood, which has a higher density than
the combined sapwood and heartwood samples at 1.5
years of age.
According to Hillis (1987) the width of sapwood in coastal
eucalypts over 15 years old remains largely unchanged.
Based on this we corrected for the missing sapwood by
determining the average sapwood width of younger samples
(2.54 ^ 0.55 cm) and calculating the ‘original’ volume of
11.5 year-old E. maculata disks. The wood density corrected
by the original log volume has been included in Fig. 4, and it
was used as the basis for the negative exponential regression.
Based on the correction for sapwood loss the estimated decay
constant was k ¼ 0:049; and the t0.95 was 62 years.
The laboratory respiration rates of E. maculata followed
a completely different pattern from that in E. regnans (Table
2). Carbon loss was highest in the youngest E. maculata
samples, supporting field observation of intensive fungal
colonisation of the sapwood in 1.5 year-old logs. The
decline of respiration from youngest to oldest samples could
not be explained statistically by the variation in wood
density, moisture content, or C-to-N ratio in E. maculata.
The variability of respiration rates within age groups
declined with age. The average respiration rate was
0.358 mg CO2-C g (C)21 d21, which is higher than in E.
regnans and 11.9 times higher than the filed decay rate
determined through density loss.
The C-to-N ratio was highest after 11.5 years. It
increased from C-to-N 410 at age 6.5 to 670 at age 11.5
years (Table 2).
4. Discussion
4.1. Log respiration rates
For all species, the laboratory respiration rates were
orders of magnitude higher than the decay rates. These
enormous differences are likely to be the result of
differences in water content. The incubated samples were
collected in winter at a very high water content, whereas
Table 2
Density (g cm23), water content (% dry weight), carbon-to-nitrogen ratio, and respiration (mg CO2-C g C21 d21) in wood samples of P. radiata, E. regnans,
and E. maculata at different stages of decomposition. Standard deviation is provided in brackets
Species Age
(yr)
Density
(g cm23)
Water content
(% dry weight)
C/N Respiration
(CO2-C g C21d21) mg
P. radiata 1 0.41(0.06) 61.1(30.5) 306.1(150.1) 0.672(0.407)
2.5 0.33(0.07) 111.0(47.1) 226.6(57.5) 0.746(0.482)
6 0.20(0.04) 173.9(50.0) 234.5(90.0) 0.405(0.174)
9 0.15(0.02) 256.1(43.7) 138.4(32.5) 0.585(0.248)
E. regnans 1 0.56(0.08) 49.6(12.5) 403.0(77.9) 0.117(0.075)
3.5 0.45(0.09) 84.0(38.5) 350.8(109.3) 0.232(0.031)
6.5 0.38(0.09) 151.7(71.6) 145.2(61.6) 0.510(0.285)
12 0.35(0.11) 183.8(96.0) 236.9(122.7) 0.331(0.208)
E. maculata 1.5 0.65(0.11) 47.3(9.3) 424.2(96.6) 0.560(0.369)
6.5 0.55(0.07) 61.6(21.2) 406.3(104.7) 0.346(0.129)
11.5 0.69(0.09) 58.8(29.9) 673.7(207.2) 0.169(0.128)
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186 181
the logs in the field undergo long dry periods. In
addition, respiration rates were based on relatively short
incubations. Thus, the C released may not only stem
from the remaining wood, but also from the turnover of
part of the microbial biomass in the wood, which might
have been accelerated by the disturbance of samples.
However, the laboratory respiration rates indicate that
there must be extended periods during which the respiration
and hence decomposition of CWD in the field is negligible.
This may explain the very slow decay process observed in
some dry forest and woodland ecosystems (Thornton et al.,
1983, 1991).
Although the laboratory respiration rates cannot be
directly converted into field decay rates, they were helpful
in establishing the decay patterns and the differences
between the different types of wood. The single component,
negative exponential decay model assumes a constant
substrate quality, and thus the decay rate is a constant
fraction of the amount of mass remaining. The laboratory
respiration rates were a useful indicator of the suitability of
the single component model. The relatively constant
respiration rates across the different age classes of decaying
logs in P. radiata confirmed the suitability of the single
component model for this species. However, the variable or
decreasing respiration rates in E. regnans and E. maculata
pointed to problems with the application of the single
component decay model.
4.2. Decay rates as determined by wood density loss
The estimation of decay rates by measuring the density of
wood samples from a chronosequence of sites is widely used.
However, the density loss pattern in E. regnans and E.
maculata depicted in Figs. 2(a) and 4(a), point to potential
Fig. 2. Development of wood density in E. regnans logs decomposing in situ for 1, 3.5, 6.5, and 12 years following clear cut harvesting. Bars represent standard
deviation.
Fig. 3. Relationship between C/N ratio and respiration rates (mg CO2 g C21 d21) in decaying CWD of E. regnans sampled at different ages following clear-
felling.
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186182
problems associated with this approach. Using the water
replacement technique to determine the volume of wood
samples takes into account the loss of density associated with
a loss of substance within otherwise solid wood. However, as
decomposition proceeds some of the original volume is also
lost e.g. through fragmentation or complete decomposition
of sapwood. Using the water replacement technique does
therefore not relate the current weight to the original volume
of the sample, but to the current volume. This may lead to an
underestimation of the density loss and consequently leads to
an underestimation of the decay rate. This is a particular
problem in samples where sapwood decays faster than
heartwood, which makes it difficult to establish the original
sample diameter once the sapwood has disappeared (see E.
maculata ). In contrast to E. regnans it was at least possible
for E. maculata samples to correct the volume for the
sapwood loss. The assumption that the small density loss in
E. regnans between years 6.5 and 12 was caused by a loss of
sapwood volume is supported by the C-to-N ratios (Fig.
2(b)). The C-to-N ratio declines steeply as decomposition
and concomitant N immobilisation increases up to age 6.5.
The subsequent increase in the C/N ratio shows that the
material at age 12 is less decomposed than that at age 6.5,
which indicates a higher proportion of heartwood. If we
follow this rational and base the determination of the
decomposition constant on the first three data points (age
class 1–6.5 years) we obtain a value of k ¼ 0:07; which
provides a time for 95% decay (t0.95) of 43 years.
A similar pattern was observed in E. maculata. In
addition, the low respiration rates in the oldest samples of
the eucalypt species indicate that the substrate quality for
micro-organisms is lower than in younger samples. This
also illustrates that both respiration rates and C-to-N ratios
can be used as a diagnostic tool to assess the validity of
density loss measurements. Where fragmentation causes the
loss of the original wood volume, this has to be
reconstructed to provide a realistic assessment of decay
rates (see also Brown et al. (1998)).
The problem caused by the loss of sapwood, including
increasing variability in density over age, did not occur for
P. radiata samples, where case-hardening preserved the
outer wood surface and thus the volume of even the older
samples (6.5 and 9 years).
The different decay rates of sapwood and heartwood in
the eucalypt species indicate that the single component
model used in the regression analysis is not applicable.
Instead a two component model simulating the decay of two
different substrates would be more appropriate (Wieder and
Lang, 1982; Means et al., 1985). However, in this study the
time span covered by the chronosequence was not long
enough to follow the decay pattern of the heartwood.
4.3. Decomposition times
The time required for 95% mass loss (t0.95) (Eq. (3)) in P.
radiata was much longer when compared to data derived
from so-called accelerated laboratory tests on wood
durability by Costa (1979). His results translate into times
for 95% decay (t0.95) of 6 years for sap- and heartwood of P.
radiata (see Mackensen and Bauhus, 1999; Mackensen
et al., 2003). However, a time for 95% decay (t0.95) of 24
years is much shorter than those found by other field studies
Fig. 4. Development of wood density in logs decomposing in situ for 1.5, 6.5, and 11.5 years following selective harvesting. Bars represent standard deviation.
The empty square at year 11.5 represents the measured density value, whereas the filled square used in the regression has been corrected for the loss of
sapwood.
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186 183

for similar species, e.g. 61 years for Pinus pinaster in West
Australia (Brown et al., 1996) and North American species
ranging from 50 to 200 years (Harmon, 1982; Foster and
Lang, 1982; Fahey, 1983; Barber and VanLear, 1984; Alban
and Pastor, 1993; Busse, 1994; Krankina and Harmon,
1995).
A time for 95% decay (t0.95) of 43–74 years for E.
regnans appears to be long when compared to results of
Accelerated Laboratory Tests for durability studies by Costa
(1979) reporting a t0.95 of only 7 years. According to
Thornton et al. (1997) E. regnans and P. radiata both
belong to durability class 4 (low durability). Durability
classes were determined from long-term field exposure of
small wood samples that were half-buried in soil. Transfer-
ring weight loss as obtained in durability studies (e.g.
Thornton et al. 1997) into time for 95% decay would
translate durability class 4 into a t0.95 of 11 years
(Mackensen and Bauhus, 1999). The much longer decay
periods found in this field study compared to durability and
laboratory studies confirm that decomposition depends on a
wide variety of factors including microclimate and decom-
poser communities.
The estimated t0.95 for E. maculata (durability class 2) of
62 years fell between the times calculated for the
comparable Eucalyptus diversicolor (durability class 2–3,
Thornton et al., 1997). Brown et al. (1996) calculated a t0.95
of 17 years for samples of Eucalyptus diversicolor including
bark from decomposition rates in a 5 year long study, while
for the 2 year long study by O’Connell (1997) calculated
t0.95 values ranged from 65 to 136 years for wood samples.
4.4. General limitations of both methods
Our sampling was concentrated on logs that were
positioned on the surface. However, it cannot be ruled out
that some of these logs may have been initially suspended
and only later fell to the ground (e.g. Næsset, 1999), and
thus will not have undergone decomposition with ground
contact for the entire period following harvesting. Although
sampling was only of stem or crown sections with clear
cutting marks, it cannot be ruled out that trees might have
been dead prior to felling or that, in the selection logging
system, the logs on the ground might have originated from
previous operations such as pole thinning.
Modification of decay through charring of logs as a
result of slash burning after logging or prescribed fires
and wildfires has not yet been determined. As charcoal is
highly resistant to decay, it is usually assumed that decay
of charred logs is reduced (see Mackensen and Bauhus
(1999)). However, our observations in the field could not
confirm this. The surface of logs is never entirely
charred, and the cracks in the charcoal allow entry of
fungal spores, if not already present before the logs were
charred. Thus we found that decomposition processes had
continued beneath a charred surface. This aspect deserves
further study.
The above considerations point to the limitations of the
chronosequence approach. They also indicate the necessity
to establish long-term experiments that allow a controlled
investigation of the different factors that may influence
CWD decomposition.
We applied a single-component, negative exponential
decay model to derive the decay rates for different woods
from density loss. However, the results for E. regnans and
E. maculata indicate that a two-component decay model,
accounting for the differences in decay resistance between
the sapwood and the heartwood, may be more appropriate.
For this study, we considered the data basis as insufficient to
apply a two-component decay model. In this case the decay
was initially faster, when sapwood was decomposing, and
then slower. However, the opposite has also been observed
(Grier, 1978; Næsset, 1999) and was interpreted as a lag
period resulting from a slow colonisation of the log with
decomposer organisms. To determine decay rates with a
greater degree of confidence, long-term studies including
logs of known volume and spanning over periods allowing
substantial decay of both sap and heartwood would be
required.
4.5. Significance of decay rates for estimating CO2
release from CWD
To determine the CO2 release from the decay of CWD
as a result of land use change and forestry activities in a
reference year, it is important to know how far back in
time estimates of the input of CWD into the decaying
pool have to go. Using the IPCC default of a 10 year
decay period of litter, the inputs would have to be
calculated for the preceding 10 years. However, the field
decay rates in this study indicate that the input of wood
into a decaying pool might have to be calculated for a
significantly longer period prior to a reference year, for
which the CO2 emission is to be established. If the input
to that pool was constant over time, the period is not
important. However, if through changes in harvesting and
land clearing activities the inputs were highly variable,
an accurate determination of the period over which input
of CWD into a decaying pool is determined will be
important.
5. Conclusions
It can be concluded that an assumed decay period of 10
years (IPCC default) is a considerable overestimation of the
rate of CWD-decomposition. Despite the variety of abiotic
and biotic parameters influencing the rate of decay, we
assume that the time for most CWD to disappear (t0.95)
exceeds 25–30 years in most cases. For many durable
species a t0.95 of more then 50 years is realistic. Instead of
assuming a linear decay of all material within 10 years, an
exponential decay function with rate constants according to
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186184

the durability class of species should be used. For species
with considerable variation in the decay resistance of sap-
and heartwood, two component decay models will be more
appropriate.
Our study has demonstrated that the combination of
different methods, including current respiration rates and
chemical characterisation of the decomposition stage (C-to-
N ratio), can assist the development of a realistic picture of
wood decay.
Acknowledgements
This study was financially supported by the Australian
Greenhouse Office. We thank Mick Morley (DNRE
Victoria), Aidan Flanagan (ACT Forests) and Ian Barnes
(State Forests of NSW) for their co-operation and assistance
with the selection of suitable sampling sites. We thank John
Kane, Sanjiva de Silva, and Scobie Mackay for their
enthusiastic help with sampling, analysis and data proces-
sing as well as Nina Stahl and Emily Backen for collecting
and managing the references. We also acknowledge the
support by Gregor Christie and Paul Smart (CSIRO,
Division of material Science and Technology), who
generously donated the foil used for incubation bags. Two
anonymous reviewers provided helpful comments on a
previous version of the manuscript.
References
Alban, D.H., Pastor, J., 1993. Decomposition of aspen, spruce, and pine
boles on two sites in Minnesota. Canadian Journal of Forest Research
23, 1744–1749.
Arthur, M.A., Tritton, L.M., Fahey, T.J., 1993. Dead bole mass and nutrient
remaining 23 years after clear-felling of a northern hardwood forest.
Canadian Journal of Forest Research 23, 1298–1305.
Barber, B.L., VanLear, D.H., 1984. Weight loss and nutrient dynamics in
decomposing woody loblolly pine logging slash. Soil Science Society
of America Journal 48, 906–910.
Brown, S., Mo, J., McPherson, J.K., Bell, D.T., 1996. Decomposition of
woody debris in Western Australian forests. Canadian Journal of Forest
Research 26, 954–966.
Brown, P.M., Shepperd, W.D., Mata, S.A., McClain, D.L., 1998. Longevity
of windthrown logs in a subalpine forest of central Colorado. Canadian
Journal of Forest Research 28, 932–936.
Burger, J., Pritchett, W., 1984. Effects of clearfelling and site preparation
on nitrogen mineralisation in a Southern pine stand. Soil Science
Society of America Journal 48, 1432–1437.
Busse, M.D., 1994. Downed bole-wood decomposition in lodgepole pine
forests of Central Oregon. Soil Science Society of America Journal 58
(1), 221–227.
Christensen, O., 1984. The states of decay of woody litter by relative
density. Oikos 42, 211–219.
Costa, E.W.B.da, 1979. Comparative decay resistance of Australian
timbers in accelerated laboratory tests. Australian Forest Research 9,
119–135.
Cowling, E.B., 1972. Nitrogen in forest trees and its role in wood
deterioration. Abstracts of Uppsala Dissertations in Science.
Edwards, N.T., 1982. The use of soda-lime for measuring respiration rates
in terrestrial systems. Pedobiologia 23, 321–330.
Fahey, T.J., 1983. Nutrient dynamics of aboveground detritus in lodgepole
pine (Pinus contorta ssp. latifolia) ecosystems, south-eastern Wyoming.
Ecological Monographs 53, 51–72.
Foster, J.R., Lang, G.E., 1982. Decomposition of red spruce and balsam fir
boles in the White Mountains of New Hamshire. Canadian Journal of
Forest Research 12, 617–626.
Graham, R.L., Cromack, K.C., 1982. Mass, nutrient content, and decay rate
of dead boles in rain forests of Olympic National Park. Canadian
Journal of Forest Research 12, 511–521.
Grier, C.C., 1978. A Tsuga heterophylla–Picea sitchensis ecosystem of
coastal Oregon: decomposition and nutrient balances of fallen logs.
Canadian Journal of Forest Research 8, 198–206.
Harmon, M.E., 1982. Decomposition of standing dead trees in the southern
Appalachian Mountains. Oecologia 52, 214–215.
Harmon, M.E., Franklin, J.F., Swanson, F.J., Sollins, P., Gregory, S.V.,
Lattin, J.D., Anderson, N.H., Cline, S.P., Aumen, N.G., Sedell, J.R.,
Lienkaemper, G.W., Cromack, J.R., Cummins, K.W., 1986. Ecology of
coarse woody debris in temperate ecosystems. Advances in Ecological
Research 15, 133–302.
Healey, I.N., Swift, M.J., 1971. Aspects of the accumulation and
decomposition of wood in the litter layer of a coppiced beech-oak
woodland. Annual Zoological Ecology Anim./hors serie, 417–430.
Hillis, W.E., 1987. Heartwood and Tree Exudates, Springer Series in Wood
Science, Springer, Berlin, 287 pp.
IPCC, 1997. Revised 1996 IPCC Guidelines for National Greenhouse Gas
Inventories, Reference Manual, Intergovernmental Panel on Climate
Change, vol. 3. Meteorological Office, Bracknell, UK.
Jenny, H., Gessel, S.P., Bingham, F.T., 1949. Comparative study of
decomposition of organic matter in temperate and tropical regions. Soil
Science 68, 419–432.
Krankina, O.N., Harmon, M.E., 1995. Dynamics of the dead wood carbon
pool in north-western Russian boreal forests. Water, Air and Soil
Pollution 82, 227–238.
Lambert, R.L., Lang, G.E., Reiners, W.A., 1980. Loss of mass and chemical
change in decaying boles of a subalpine balsam fir forest. Ecology 61,
1460–1473.
Mackensen, J., Bauhus, J., 1999. The decay of coarse woody debris.
National Carbon Accounting System, Technical Report No. 6,
Australian Greenhouse Office, Commonwealth of Australia, Canberra,
40 pp.
Mackensen, J., Bauhus, J., Webber, E., 2003. Decomposition rates of coarse
woody debris—a review with particular emphasis on Australian tree
species. Australian Journal of Bot. in press.
Means, J.E., Cromack, K.J., MacMillan, P.C., 1985. Comparison of
decomposition models using wood density of Douglas-fir logs.
Canadian Journal of Forest Research 15, 1092–1098.
Næsset, E., 1999. Decomposition rate constants of Picea abies logs in
southeastern Norway. Canadian Journal Forest Research 29,
372–381.
NGGIC (National Greenhouse Gas Inventory Committee), 1997a. Land
Use Change and Forestry. Workbook for Carbon Dioxide from the
Biosphere. Workbook 4.2, Revision 2.
NGGIC (National Greenhouse Gas Inventory Committee), 1997b. Land
Use Change and Forestry Sector 1988–1995, based on Workbook 4.2.
O’Connell, A.M., 1997. Decomposition of slash residues in thinned
regrowth eucalypt forest in Western Australia. Journal of Applied
Ecology 34, 111–122.
Olson, J.S., 1963. Energy storage and the balance of producers and
decomposers in ecological systems. Ecology 44 (2), 322–331.
SAS Institute, 1996. JMPw Start Statistics, Duxbury Press, Belmont.
Stone, J.N., MacKinnon, A., Parminter, J.V., Lertzman, K.P., 1998.
Coarse woody debris decomposition documented over 65 years on
southern Vancouver Island. Canadian Journal of Forest Research 28,
788–793.
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186 185

Swift, M.J., 1977. The ecology of wood decomposition. Science
Programme 64, 175–199.
Swift, M.J., Heal, O.W., Anderson, J.M., 1979. Decomposition in
Terrestrial Ecosystems, University of California Press, Berkeley, pp.
354, 354 pp.
Thornton, J.D., Walters, N.E.M., Saunders, I.W., 1983. An in-ground
natural durability field test of Australian timbers and exotic reference
species I. Progress report after more than 10 years exposure. Material
und Organismen 18, 27–49.
Thornton, J.D., Johnson, G.C., Nguyen, N., 1991. An in-ground natural
durability field test of Autralian timbers and exotic reference species.
Material und Organismen 26, 1–17.
Thornton, J.D., Johnson, G.C., Nguyen, N., 1997. Revised CSIRO Natural
Durability Classification—In-Groud Durability Ratings for Mature
Outer heartwood, CSIRO Forestry and Forest Products, Melbourne, 2 pp.
Wieder, R.K., Lang, G.E., 1982. A critique of the analytical methods used
in examining decomposition data obtained from litter bags. Ecology 63,
1636–1642.
J. Mackensen, J. Bauhus / Soil Biology & Biochemistry 35 (2003) 177–186186
Related Documents











