ORIGINAL ARTICLE Deficiency in the NADPH oxidase 4 predisposes towards diet-induced obesity Y Li 1,7,8 , S Mouche 1,8 , T Sajic 1 , C Veyrat-Durebex 2 , R Supale 1 , D Pierroz 2,3 , S Ferrari 2,3 , F Negro 4,5 , U Hasler 1 , E Feraille 1 , S Moll 5 , P Meda 1 , C Deffert 6 , X Montet 1 , K-H Krause 4,5 and I Szanto 1,3 OBJECTIVE: NADPH oxidase 4 (NOX4) is a reactive oxygen species (ROS) producing NADPH oxidase that regulates redox homeostasis in diverse insulin-sensitive cell types. In particular, NOX4-derived ROS is a key modulator of adipocyte differentiation and mediates insulin receptor signaling in mature adipocytes in vitro. Our study was aimed at investigating the role of NOX4 in adipose tissue differentiation, whole body metabolic homeostasis and insulin sensitivity in vivo. DESIGN: Mice with genetic ablation of NOX4 (NOX4-deficient mice) were subjected to chow or high-fat-containing diet for 12 weeks. Body weight gain, adiposity, insulin sensitivity, and adipose tissue and liver gene and protein expression were analyzed and compared with similarly treated wild-type mice. RESULTS: Here, we report that NOX4-deficient mice display latent adipose tissue accumulation and are susceptible to diet-induced obesity and early onset insulin resistance. Obesity results from accelerated adipocyte differentiation and hypertrophy, and an increase in whole body energy efficiency. Insulin resistance is associated with increased adipose tissue hypoxia, inflammation and adipocyte apoptosis. In the liver, more severe diet-induced steatosis was observed due to the lack of proper upregulation of mitochondrial fatty acid b-oxidation. CONCLUSION: These findings identify NOX4 as a regulator of metabolic homeostasis. Moreover, they indicate an anti-adipogenic role for NOX4 in vivo and reveal its function as a protector against the development of diet-induced obesity, insulin resistance and hepatosteatosis. International Journal of Obesity advance online publication, 20 March 2012; doi:10.1038/ijo.2011.279 Keywords: NOX4; adipocyte differentiation; high-fat diet INTRODUCTION NADPH oxidases are pro-oxidant enzymes whose main cellular function is the production of reactive oxygen species (ROS). The best renowned member of this family is the phagocyte NADPH oxidase (gp91 phox /NOX2) mediating host defense action. 1 NADPH oxidase 4 (NOX4), another recently cloned member of the NADPH oxidase family, was originally termed ‘Renox’ for its predominant expression in kidney. 2,3 NOX4 produces ROS in a constitutive manner and its regulation is achieved at the transcriptional and translational levels. 4,5 Physiological ROS production has a key role in the intracellular insulin signal transmission and exerts a protective effect against the onset of diet-induced insulin resistance. 6,7 NOX4 has been shown to convey a variety of cellular signaling events in different insulin- sensitive tissues, for example, hepatocytes, vascular smooth muscle cells, endothelial cells and fibroblasts reviewed in. 1 In particular, NOX4 has been shown to mediate insulin-induced glucose uptake in adipocytes. 6 Insulin is also a key modulator of adipocyte differentiation both in vitro and in vivo, and NOX4 is one of the genes whose expression is regulated during the course of this process. 8 - 11 Adipocyte differentiation and hypertrophy is closely linked to the onset of insulin resistance. 12 Elevated NOX4 levels have been observed in the adipose tissue in rodent models of obesity and in humans with extreme insulin resistance. 13,14 Despite the close link between insulin signaling, ROS produc- tion and NOX4 expression in adipocytes, the role of NOX4 in the regulation of metabolic homeostasis and the development of diet- induced obesity and insulin resistance in vivo has not yet been explored. Thus, our study was aimed at investigating the metabolic phenotype of mice with genetic ablation of NOX4 (NOX4-deficient mice) and studying their response to high-fat diet (HFD) challenge. Our data demonstrate that in the absence of NOX4, mice develop latent adipose tissue hypertrophy with the molecular signature of hypoxia and inflammation. Upon HFD feeding, NOX4-deficient mice were more sensitive to the development of diet-induced obesity, insulin resistance and liver steatosis. Taken together, these data suggest that NOX4-mediated constitutive ROS production has a key role in adipose tissue function, and decreased NOX4 activity sensitizes towards the development of metabolic disorders upon nutritional challenge. Received 11 August 2011; revised 1 December 2011; accepted 5 December 2011 1 Department of Cellular Physiology and Metabolism, University of Geneva, Geneva, Switzerland; 2 Department of Medical Specialties, University of Geneva, Geneva, Switzerland; 3 Department of Rehabilitation and Geriatrics, University of Geneva, Geneva, Switzerland; 4 Division of Gastroenterology and Hepatology, Geneva University Hospitals, Geneva, Switzerland; 5 Division of Clinical Pathology, Geneva University Hospitals, Geneva, Switzerland and 6 Department of Genetic and Laboratory Medicine, Geneva University Hospitals, Geneva, Switzerland. Correspondence: Dr I Szanto, Department of Rehabilitation and Geriatrics, Geneva University Hospitals, Geneva 1211, Switzerland. E-mail: [email protected] 7 Current address: Beijing Hospital, Beijing, P.R China 8 These authors contributed equally to this work. International Journal of Obesity (2012) 1 - 11 & 2012 Macmillan Publishers Limited All rights reserved 0307-0565/12 www.nature.com/ijo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Deficiency in the NADPH oxidase 4 predisposes towardsdiet-induced obesityY Li1,7,8, S Mouche1,8, T Sajic1, C Veyrat-Durebex2, R Supale1, D Pierroz2,3, S Ferrari2,3, F Negro4,5, U Hasler1, E Feraille1, S Moll5, P Meda1,C Deffert6, X Montet1, K-H Krause4,5 and I Szanto1,3
OBJECTIVE: NADPH oxidase 4 (NOX4) is a reactive oxygen species (ROS) producing NADPH oxidase that regulates redoxhomeostasis in diverse insulin-sensitive cell types. In particular, NOX4-derived ROS is a key modulator of adipocytedifferentiation and mediates insulin receptor signaling in mature adipocytes in vitro. Our study was aimed at investigating therole of NOX4 in adipose tissue differentiation, whole body metabolic homeostasis and insulin sensitivity in vivo.DESIGN: Mice with genetic ablation of NOX4 (NOX4-deficient mice) were subjected to chow or high-fat-containing diet for 12weeks. Body weight gain, adiposity, insulin sensitivity, and adipose tissue and liver gene and protein expression were analyzedand compared with similarly treated wild-type mice.RESULTS: Here, we report that NOX4-deficient mice display latent adipose tissue accumulation and are susceptible todiet-induced obesity and early onset insulin resistance. Obesity results from accelerated adipocyte differentiation andhypertrophy, and an increase in whole body energy efficiency. Insulin resistance is associated with increased adipose tissuehypoxia, inflammation and adipocyte apoptosis. In the liver, more severe diet-induced steatosis was observed due to the lackof proper upregulation of mitochondrial fatty acid b-oxidation.CONCLUSION: These findings identify NOX4 as a regulator of metabolic homeostasis. Moreover, they indicate ananti-adipogenic role for NOX4 in vivo and reveal its function as a protector against the development of diet-inducedobesity, insulin resistance and hepatosteatosis.
International Journal of Obesity advance online publication, 20 March 2012; doi:10.1038/ijo.2011.279
Keywords: NOX4; adipocyte differentiation; high-fat diet
INTRODUCTIONNADPH oxidases are pro-oxidant enzymes whose main cellularfunction is the production of reactive oxygen species (ROS). Thebest renowned member of this family is the phagocyte NADPHoxidase (gp91phox/NOX2) mediating host defense action.1
NADPH oxidase 4 (NOX4), another recently cloned member ofthe NADPH oxidase family, was originally termed ‘Renox’ for itspredominant expression in kidney.2,3 NOX4 produces ROS in aconstitutive manner and its regulation is achieved at thetranscriptional and translational levels.4,5 Physiological ROSproduction has a key role in the intracellular insulin signaltransmission and exerts a protective effect against the onset ofdiet-induced insulin resistance.6,7 NOX4 has been shown toconvey a variety of cellular signaling events in different insulin-sensitive tissues, for example, hepatocytes, vascular smoothmuscle cells, endothelial cells and fibroblasts reviewed in.1 Inparticular, NOX4 has been shown to mediate insulin-inducedglucose uptake in adipocytes.6 Insulin is also a key modulator ofadipocyte differentiation both in vitro and in vivo, and NOX4 is oneof the genes whose expression is regulated during the course of
this process.8 -- 11 Adipocyte differentiation and hypertrophy isclosely linked to the onset of insulin resistance.12 Elevated NOX4levels have been observed in the adipose tissue in rodent modelsof obesity and in humans with extreme insulin resistance.13,14
Despite the close link between insulin signaling, ROS produc-tion and NOX4 expression in adipocytes, the role of NOX4 in theregulation of metabolic homeostasis and the development of diet-induced obesity and insulin resistance in vivo has not yet beenexplored. Thus, our study was aimed at investigating themetabolic phenotype of mice with genetic ablation of NOX4(NOX4-deficient mice) and studying their response to high-fat diet(HFD) challenge. Our data demonstrate that in the absence ofNOX4, mice develop latent adipose tissue hypertrophy with themolecular signature of hypoxia and inflammation. Upon HFDfeeding, NOX4-deficient mice were more sensitive to thedevelopment of diet-induced obesity, insulin resistance and liversteatosis. Taken together, these data suggest that NOX4-mediatedconstitutive ROS production has a key role in adipose tissuefunction, and decreased NOX4 activity sensitizes towards thedevelopment of metabolic disorders upon nutritional challenge.
Received 11 August 2011; revised 1 December 2011; accepted 5 December 2011
1Department of Cellular Physiology and Metabolism, University of Geneva, Geneva, Switzerland; 2Department of Medical Specialties, University of Geneva, Geneva, Switzerland;3Department of Rehabilitation and Geriatrics, University of Geneva, Geneva, Switzerland; 4Division of Gastroenterology and Hepatology, Geneva University Hospitals, Geneva,Switzerland; 5Division of Clinical Pathology, Geneva University Hospitals, Geneva, Switzerland and 6Department of Genetic and Laboratory Medicine, Geneva University Hospitals,Geneva, Switzerland. Correspondence: Dr I Szanto, Department of Rehabilitation and Geriatrics, Geneva University Hospitals, Geneva 1211, Switzerland.E-mail: [email protected] address: Beijing Hospital, Beijing, P.R China8These authors contributed equally to this work.
International Journal of Obesity (2012) 1 -- 11& 2012 Macmillan Publishers Limited All rights reserved 0307-0565/12
www.nature.com/ijo

MATERIALS AND METHODSMice, diets and treatmentsNOX4-deficient mice were generated as described.15 Genotyping wascarried out by using the primers NOX4: forward 50-TCATGACAGTTGGGGACAAA3-0 ; NOX4: reverse 50-TTGAAAATTCAACACAAGTCTCC3-0 for wild type(WT) and the reverse primer NOX4 Neo R 50-AACGTCGTGACTGGGAAAAC-30
for the mutant allele. WT and NOX4-deficient mice were housed inconventional animal facility in individual cages under controlled tempera-ture (23 1C) and illumination (12-h light/12-h dark cycle). Ten- to eleven-week-old male mice were given either standard chow or HFD (60% ofenergy as fat, cat: 2127, Provimi Kliba AG, Kaiseraugst, Switzerland) for aperiod of 12 weeks. All mice were allowed ad libitum access to water. Micewere killed in the non-fasted state. The experimental protocol had beenaccepted by the Ethical Committee of the University of Geneva and theVeterinary Office of the Canton of Geneva.
Measurement of whole body fat and lean massWhole body fat and lean mass was measured by dual-energy X-rayabsorptiometry (PIXImus2, GE Lunar, Madison, WI, USA) as describedpreviously.16
Intraperitoneal glucose, insulin and pyruvate tolerance tests(ipGTT, ipITT and ipPTT)Mice were fasted overnight and were administered intraperitoneally with2 mg kg�1 glucose (ipGTT), 1 U kg�1 insulin (ipITT) or 2 g kg�1 sodiumpyruvate (ipPTT). Glucose levels were monitored during a period of 2 husing a glucometer (Accu-check, Roche Diagnostics GmbH, Mannheim,Germany).
Metabolic cage measurementsFood and water intake, energy expenditure, respiratory quotient (RQ) andlocomotor activity of mice were analyzed in a custom-made 12-cagecalorimetric system (LabMaster Multipurpose Screening System; TSESystems, Bad Homburg, Germany). The instrument consists of a combina-tion of highly sensitive feeding and drinking sensors for automated onlinemeasurement. The calorimetric system is an open-circuit system thatdetermines O2 consumption, CO2 production and RQ (RQ¼ VCO2/VO2,where V is volume). A photobeam-based activity monitoring systemdetects and records every ambulatory movement, including rearing andclimbing movements in every cage. Detection of animal location isperformed with infrared sensor pairs arranged in strips for horizontal andvertical (rearing) activity. The sensors for detection of movement operateefficiently in both light and dark phases, allowing continuous recording. Allof the parameters can be measured continuously and simultaneously in upto 12 animals. Data were collected for 48 h after a 5-day adaptation period.
Determination of body fat composition by micro-CTIn vivo micro-CT images of anesthetized mice (Nembutal, 50 mg kg�1 ip)were acquired on a Skyscan-1076 micro-CT (Skyscan, Aartselaar, Belgium)using an X-ray charge-coupled device camera with a cooled 1024� 1024-pixel 12-bit sensor (http://www.medecine.unige.ch/lafaculte/services/phenotypage). The camera was rotated through 1851, and images weremade every 1.41, achieving an in vivo resolution of 35mm. Reconstructionof acquired data was made through a cone-beam reconstruction methodto account for the conical geometry of the X-ray source. Afterreconstruction, fat deposition was manually segmented with Osirix andthe volume of the abdominal fat was calculated from the subcutaneous,intra-peritoneal, retroperitoneal and epididymal regions as described.17
Serum measurementsBlood was drawn from tail vein in non-fasted state or after overnight fooddeprivation. In both conditions mice had ad libitum access to water. Insulinand leptin levels were determined by ELISA analysis (Ulrasensitive MouseInsulin ELISA kit, Mercodia, Uppsala, Sweden and Mouse/Rat LeptinEnzyme Immunoassay Kit, Spi-bio, Montigny le Bretonneux, France).
Ketone body levels were determined by Autokit Total Ketone Bodies(Wako Chemicals, Neuss, Germany).
Histology and morphometric measurementsTissue samples were fixed in 4% paraformaldehyde, embedded in paraffin,and serial sections were subjected to standard hematoxylin/eosin, acidfuchsin orange G and periodic acid Schiff (PAS) stainings as specified.Sections were evaluated in a blind manner by an independent pathologist.White adipose tissue morphometric measurements were conducted onhematoxylin/eosin-stained duplicate sections derived from three differentlevels of tissue (six images per mouse). Images were obtained by AxiocamImaging Sytem (Carl Zeiss AG, Feldbach, Switzerland). Cell diameter wascalculated from computer-assisted image transformation using Meta-Morph Image Analysis software (Molecular Devices, Union City, CA, USA),as described in Supplementary Figure S6.
Liver lipid and glycogen contentLiver lipids were extracted by modified Folch method.18 In brief,50 -- 100 mg tissues were homogenized by Potter-Elvehjem Tissue Grinderwith Teflon Pestle in the solution of chloroform:methanol (2:1) and thelipids were extracted overnight. After the addition of 1/5 volume of 0.9%NaCl solution the mixture was centrifuged at 1000 r.p.m. The lower organic(lipid-containing) phase was dried and lipids were then resuspendedin 250mL of absolute ethanol and stored at �20 1C until analyzed.Triglyceride levels were determined by a commercial kit (RocheDiagnostics, Rotkrauz, Switzerland).
Liver glycogen was extracted according as described.19 In brief,50 -- 150 mg frozen tissue samples were digested in a suitable volume of30% KOH for 10 min at 100 1C. Digestion was completed by the addition of1/5 volume of 20% NaSO4. Macromolecules were precipitated by theaddition of two volumes of absolute ethanol and overnight incubation at�20 1C. Precipitates were washed with 70% ethanol and dried, and thenglycogen was hydrolyzed by digestion in 500ml of 4N H2SO4 for 10 min at100 1C. Samples were neutralized by the addition of an equivalent volumeof 4N NaOH and then deproteinized by equal volumes of Ba(OH)2 andZnSO4. Glucose was quantified using the GAGO-20 kit (Sigma-AldrichChemie GmbH, Buchs, Switzerland).
Islet insulin content measurementsFor determination of insulin content, pancreas were extracted for 24 h at4 1C in acid-ethanol and sonicated. Aliquots of the extracts were assayedby RIA using 125I-insulin as a tracer, a specific antibody to insulin (LincoResearch, St Charles, MO, USA) and rat insulin as standard.
RNA isolation and quantitative real-time PCRTotal RNA was prepared in TRIZOL Reagent (Invitrogen, Basel, Switzerland)and was purified by using the RNeasy Mini Kit (Qiagen, Hombrechtikon,Switzerland). cDNA was synthesized from 2 mg of DNA-free RNA bySuperscript II Reverse Transcriptase (Invitrogen). The primers used forquantitative real-time PCR are listed in the Supplementary Table S1. Real-time PCR reactions were carried out using Sybergreen in an ABI 7500instrument (Applied Biosystems, Rotkreuz, Switzerland). The results werequantified by the DCt method using cyclophillin A as the standard internalnon-variable gene to compensate for differences in RNA input andefficiency of cDNA synthesis. Results were expressed as arbitrary unitscompared to the average expression levels in WT mice.
Western blot analysisTissues were snap-frozen in liquid nitrogen immediately upon removal andwere stored at �80 1C till processing. Samples were homogenized in RIPAbuffer (150 mM NaCl, 1% NP40, 0.1% SDS, 0.5% deoxycholic sodium salt,50 mM Tris HCl, pH 7.4, 2 mM EDTA, 2 mM Na3VO4, 10 mM NaF, one tabletof Complete Inhibitor Cocktail (Roche Diagnostics) in 30 ml). Proteinconcentration was measured by bicinchoninic acid method (Pierce,Rockford, IL, USA). Lysates were dissolved in Laemmli buffer (10 mM
sodium phosphate, pH 7.0, 0.1% glycerol, 2% SDS, 100 mM DTT and a
NOX4 regulates adipogenesis in vivoY Li et al
2
International Journal of Obesity (2012) 1 -- 11 & 2012 Macmillan Publishers Limited
trace of BPB) and were resolved on a 5 -- 20% gradient polyacrylamide gel.Gels were transferred onto nitrocellulose membranes (GE Healthcare,Otelfingen, Switzerland) followed by incubation with the respectiveprimary antibodies. Antibodies are listed in Supplementary Table S2. Blotswere then washed and subsequently incubated with the applicablesecondary horseradish peroxidase-conjugated antibody (Sigma-Aldrich,Buch, Switzerland). Signals were revealed by enhanced chemilumines-cence and were recorded in ChemiDoc XRS system. Quantification of thedetected bands was performed by using the Quantity One program (Bio-Rad Laboratories, Reinach, Switzerland). Protein expression was related tothe amount of actin as a non-variable reference protein and was expressedas arbitrary units compared to the average expression in WT mice.
Statistical analysisData are expressed as mean±s.e.m. Results were analyzed by usingANOVA or Student’s t test, or in case of non-parametric data distribution byMann -- Whitney test using the Sigma Stat software (version 2.0---SPSS,Chicago, IL, USA). P-values less than 0.05 were considered significant.
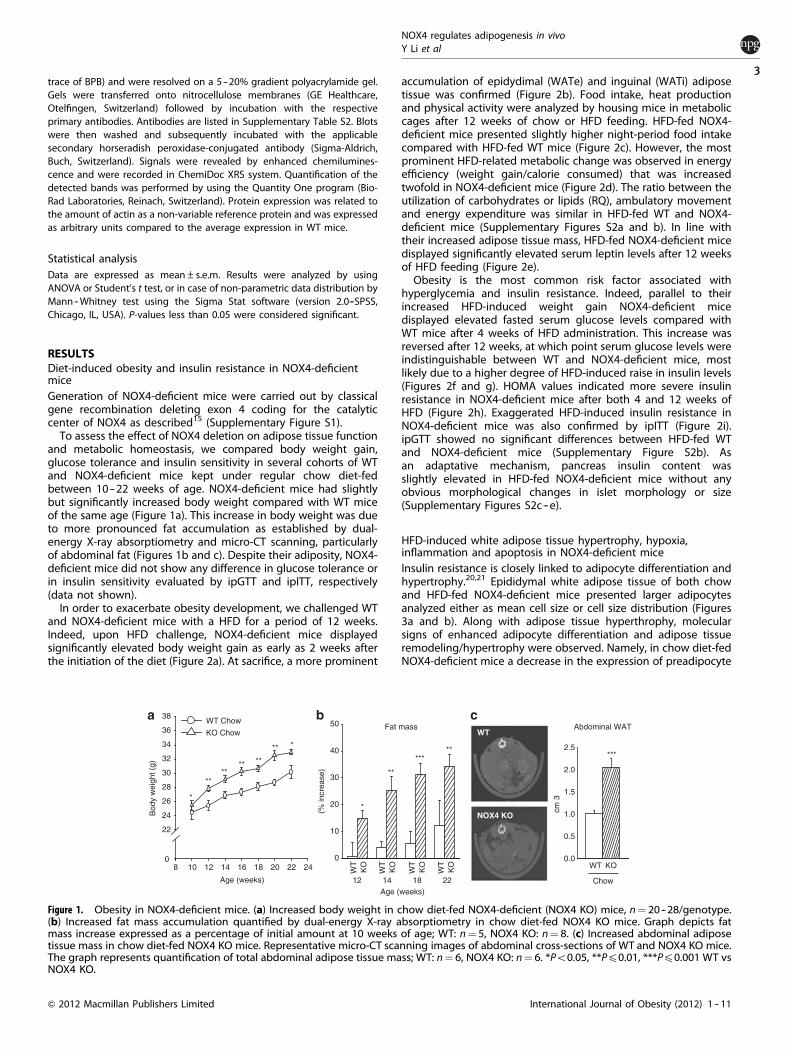
RESULTSDiet-induced obesity and insulin resistance in NOX4-deficientmiceGeneration of NOX4-deficient mice were carried out by classicalgene recombination deleting exon 4 coding for the catalyticcenter of NOX4 as described15 (Supplementary Figure S1).
To assess the effect of NOX4 deletion on adipose tissue functionand metabolic homeostasis, we compared body weight gain,glucose tolerance and insulin sensitivity in several cohorts of WTand NOX4-deficient mice kept under regular chow diet-fedbetween 10 -- 22 weeks of age. NOX4-deficient mice had slightlybut significantly increased body weight compared with WT miceof the same age (Figure 1a). This increase in body weight was dueto more pronounced fat accumulation as established by dual-energy X-ray absorptiometry and micro-CT scanning, particularlyof abdominal fat (Figures 1b and c). Despite their adiposity, NOX4-deficient mice did not show any difference in glucose tolerance orin insulin sensitivity evaluated by ipGTT and ipITT, respectively(data not shown).
In order to exacerbate obesity development, we challenged WTand NOX4-deficient mice with a HFD for a period of 12 weeks.Indeed, upon HFD challenge, NOX4-deficient mice displayedsignificantly elevated body weight gain as early as 2 weeks afterthe initiation of the diet (Figure 2a). At sacrifice, a more prominent
accumulation of epidydimal (WATe) and inguinal (WATi) adiposetissue was confirmed (Figure 2b). Food intake, heat productionand physical activity were analyzed by housing mice in metaboliccages after 12 weeks of chow or HFD feeding. HFD-fed NOX4-deficient mice presented slightly higher night-period food intakecompared with HFD-fed WT mice (Figure 2c). However, the mostprominent HFD-related metabolic change was observed in energyefficiency (weight gain/calorie consumed) that was increasedtwofold in NOX4-deficient mice (Figure 2d). The ratio between theutilization of carbohydrates or lipids (RQ), ambulatory movementand energy expenditure was similar in HFD-fed WT and NOX4-deficient mice (Supplementary Figures S2a and b). In line withtheir increased adipose tissue mass, HFD-fed NOX4-deficient micedisplayed significantly elevated serum leptin levels after 12 weeksof HFD feeding (Figure 2e).
Obesity is the most common risk factor associated withhyperglycemia and insulin resistance. Indeed, parallel to theirincreased HFD-induced weight gain NOX4-deficient micedisplayed elevated fasted serum glucose levels compared withWT mice after 4 weeks of HFD administration. This increase wasreversed after 12 weeks, at which point serum glucose levels wereindistinguishable between WT and NOX4-deficient mice, mostlikely due to a higher degree of HFD-induced raise in insulin levels(Figures 2f and g). HOMA values indicated more severe insulinresistance in NOX4-deficient mice after both 4 and 12 weeks ofHFD (Figure 2h). Exaggerated HFD-induced insulin resistance inNOX4-deficient mice was also confirmed by ipITT (Figure 2i).ipGTT showed no significant differences between HFD-fed WTand NOX4-deficient mice (Supplementary Figure S2b). Asan adaptative mechanism, pancreas insulin content wasslightly elevated in HFD-fed NOX4-deficient mice without anyobvious morphological changes in islet morphology or size(Supplementary Figures S2c -- e).
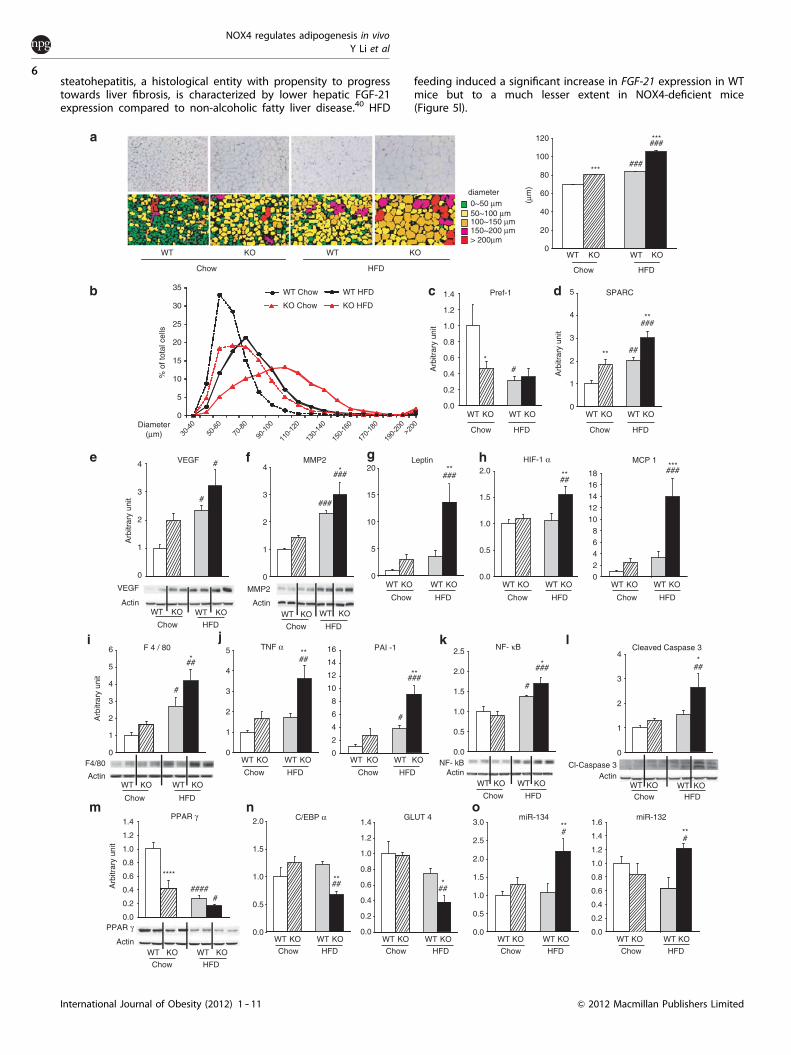
HFD-induced white adipose tissue hypertrophy, hypoxia,inflammation and apoptosis in NOX4-deficient miceInsulin resistance is closely linked to adipocyte differentiation andhypertrophy.20,21 Epididymal white adipose tissue of both chowand HFD-fed NOX4-deficient mice presented larger adipocytesanalyzed either as mean cell size or cell size distribution (Figures3a and b). Along with adipose tissue hyperthrophy, molecularsigns of enhanced adipocyte differentiation and adipose tissueremodeling/hypertrophy were observed. Namely, in chow diet-fedNOX4-deficient mice a decrease in the expression of preadipocyte
Fat mass
(% in
crea
se) **
***
WT
NOX4 KO
Abdominal WAT
cm 3
Chow
WT KO0.0
0.5
1.0
1.5
2.0
2.5***
12
WT Chow
KO Chow
Bod
y w
eigh
t (g)
Age (weeks)
****
*
** **
**
Age (weeks)
0
10
20
30
40
50
**
8 10 12 14 16 18 20 22 240
22
24
26
28
30
32
34
36
38
*
*
KO
WT
KO
WT
KO
WT
KO
WT
221814
Figure 1. Obesity in NOX4-deficient mice. (a) Increased body weight in chow diet-fed NOX4-deficient (NOX4 KO) mice, n¼ 20--28/genotype.(b) Increased fat mass accumulation quantified by dual-energy X-ray absorptiometry in chow diet-fed NOX4 KO mice. Graph depicts fatmass increase expressed as a percentage of initial amount at 10 weeks of age; WT: n¼ 5, NOX4 KO: n¼ 8. (c) Increased abdominal adiposetissue mass in chow diet-fed NOX4 KO mice. Representative micro-CT scanning images of abdominal cross-sections of WT and NOX4 KO mice.The graph represents quantification of total abdominal adipose tissue mass; WT: n¼ 6, NOX4 KO: n¼ 6. *Po0.05, **Pp0.01, ***Pp0.001 WT vsNOX4 KO.
NOX4 regulates adipogenesis in vivoY Li et al
3
International Journal of Obesity (2012) 1 -- 11& 2012 Macmillan Publishers Limited
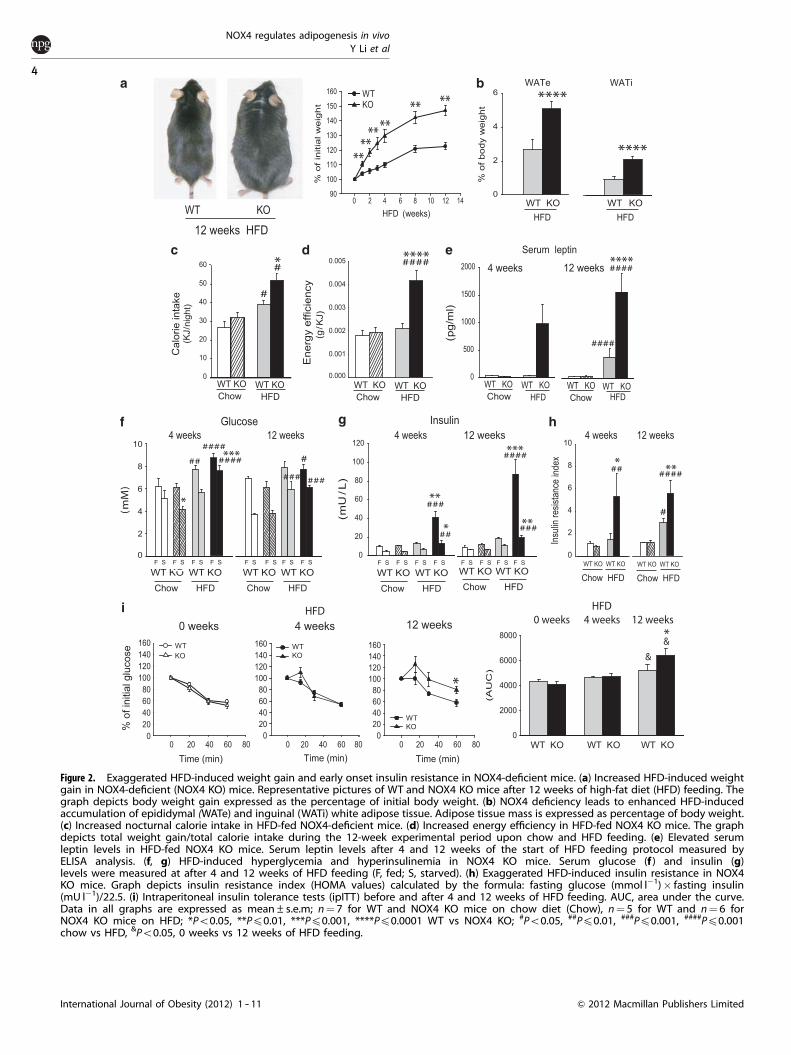
Figure 2. Exaggerated HFD-induced weight gain and early onset insulin resistance in NOX4-deficient mice. (a) Increased HFD-induced weightgain in NOX4-deficient (NOX4 KO) mice. Representative pictures of WT and NOX4 KO mice after 12 weeks of high-fat diet (HFD) feeding. Thegraph depicts body weight gain expressed as the percentage of initial body weight. (b) NOX4 deficiency leads to enhanced HFD-inducedaccumulation of epididymal (WATe) and inguinal (WATi) white adipose tissue. Adipose tissue mass is expressed as percentage of body weight.(c) Increased nocturnal calorie intake in HFD-fed NOX4-deficient mice. (d) Increased energy efficiency in HFD-fed NOX4 KO mice. The graphdepicts total weight gain/total calorie intake during the 12-week experimental period upon chow and HFD feeding. (e) Elevated serumleptin levels in HFD-fed NOX4 KO mice. Serum leptin levels after 4 and 12 weeks of the start of HFD feeding protocol measured byELISA analysis. (f, g) HFD-induced hyperglycemia and hyperinsulinemia in NOX4 KO mice. Serum glucose (f ) and insulin (g)levels were measured at after 4 and 12 weeks of HFD feeding (F, fed; S, starved). (h) Exaggerated HFD-induced insulin resistance in NOX4KO mice. Graph depicts insulin resistance index (HOMA values) calculated by the formula: fasting glucose (mmol l�1)� fasting insulin(mU l�1)/22.5. (i) Intraperitoneal insulin tolerance tests (ipITT) before and after 4 and 12 weeks of HFD feeding. AUC, area under the curve.Data in all graphs are expressed as mean±s.e.m; n¼ 7 for WT and NOX4 KO mice on chow diet (Chow), n¼ 5 for WT and n¼ 6 forNOX4 KO mice on HFD; *Po0.05, **Pp0.01, ***Pp0.001, ****Pp0.0001 WT vs NOX4 KO; #Po0.05, ##Pp0.01, ###Pp0.001, ####Pp0.001chow vs HFD, &Po0.05, 0 weeks vs 12 weeks of HFD feeding.
NOX4 regulates adipogenesis in vivoY Li et al
4
International Journal of Obesity (2012) 1 -- 11 & 2012 Macmillan Publishers Limited

factor 1 (Pref-1) and an increase in the expressions of SPARC(secreted protein acidic cystein-rich) and vascular endothelialgrowth factor (VEGF) were noted (Figures 3c -- e). HFD regimeinduced exaggerated elevation in the expressions of SPARC andmetalloproteinase 2 (MMP-2), and higher mRNA expression of theadipocyte differentiation marker leptin in NOX4-deficient mice(Figures 3f and g).
Hypertrophic adipose tissue is characterized by insufficientblood supply relative to the increased demand from hypertrophicadipocytes triggering the onset of hypoxia, followed bymacrophage infiltration and an elevation in inflammatory cytokineproduction.12,22 -- 25 In line with their hypertrophic adiposetissue, HFD-fed NOX4-deficient mice showed increased mRNAexpressions of hypoxia-inducible factor 1 (HIF-1a), macrophagechemoattractant protein-1 (MCP-1), the macrophage signatureprotein F4/80 and the pro-inflammatory factors TNFa and PAI-1(Figures 3h -- j). Accordingly, increased protein level of NF-kB, themaster transcriptional regulator of inflammatory response, wasobserved (Figure 3k). Adipose tissue inflammation triggersadipocyte apoptosis through activation of the effector proteasecaspase 3.26 HFD-fed NOX4-deficient mice presented increasedamount of the activated (cleaved) form of caspase 3 (Figure 3l).Peroxisome proliferator-activated receptor g (PPARg) is a majorregulator of adipogenesis and adipose tissue insulin sensitivity.27
PPARg protein levels were reduced in chow diet-fed NOX4-deficient mice compared with WT mice. HFD feeding led to adecrease in PPARg expression in both WT and NOX4-deficientmice that diminished the difference between genotypes(P¼ 0.013 Student’s t-test) (Figure 3m). CCAAT-enhancer-bindingprotein alpha (C/EBPa) cooperates with PPARg to promote adiposetissue differentiation and to induce transcription of the insulin-sensitive glucose transporter 4 (GLUT4).28,29 Both C/EBPa andGLUT4 expressions were downregulated in HFD-fed NOX4-deficient mice compared with WT mice (Figure 3n).
Obesity and the development of metabolic syndrome areassociated with differential regulation of several adipose tissuemicroRNAs. Namely, miR-134 is related to insulin sensitivity andchronic hyperglycemia, whereas miR-132 is linked to obesity-induced adipose tissue inflammation in humans.30 HFD-fed NOX4-deficient mice showed increased miR-134 and miR-132 expressioncompared with WT mice on the same regime (Figure 3o).Upregulation of miR-143 was associated with HFD-inducedobesity, and miR-21 has been shown to regulate adipogenicdifferentiation of mesenchymal stem cells.31,32 NOX4-deficientmice displayed similar miR-143 and miR-21 levels to WT miceregardless of the diet (data not shown).
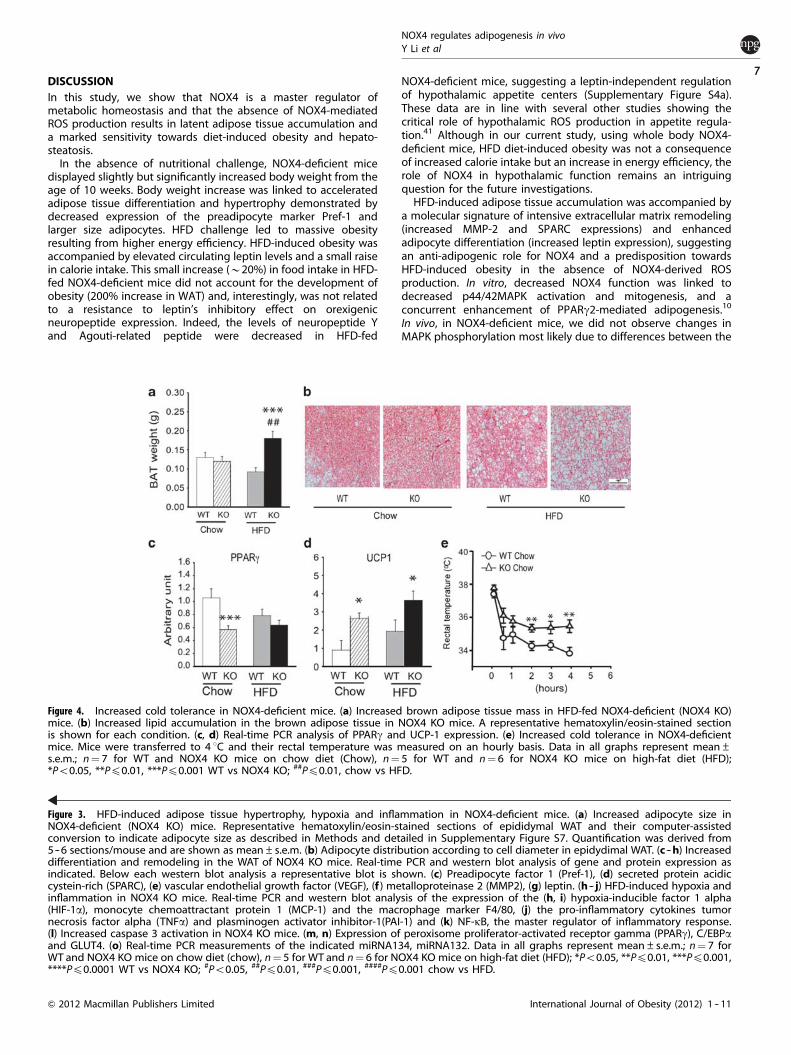
Increased cold tolerance in NOX4-deficient miceBrown adipose tissue increases energy expenditure and is animportant organ in the regulation of metabolic homeostasis.NOX4-deficient mice developed higher amount of brown adiposetissue under HFD regime parallel to a more significant lipidaccumulation in this tissue (Figures 4a and b). Chow diet-fedNOX4-deficient mice displayed attenuated PPARg mRNA levelsthat were similar to the levels observed in WT mice upon HFD-feeding (Figure 4c). Uncoupling protein-1 (UCP-1) is a brownadipose tissue-specific mitochondrial protein inducing non-shivering thermogenesis, thus increasing energy expenditure.NOX4-deficient mice had increased UCP-1 expression under bothchow and HFD regime (Figure 4d). When control, chow diet-fedmice were challenged by cold exposure at 4 1C, NOX4-deficientmice maintained their body temperature more efficiently than WTmice (Figure 4e).
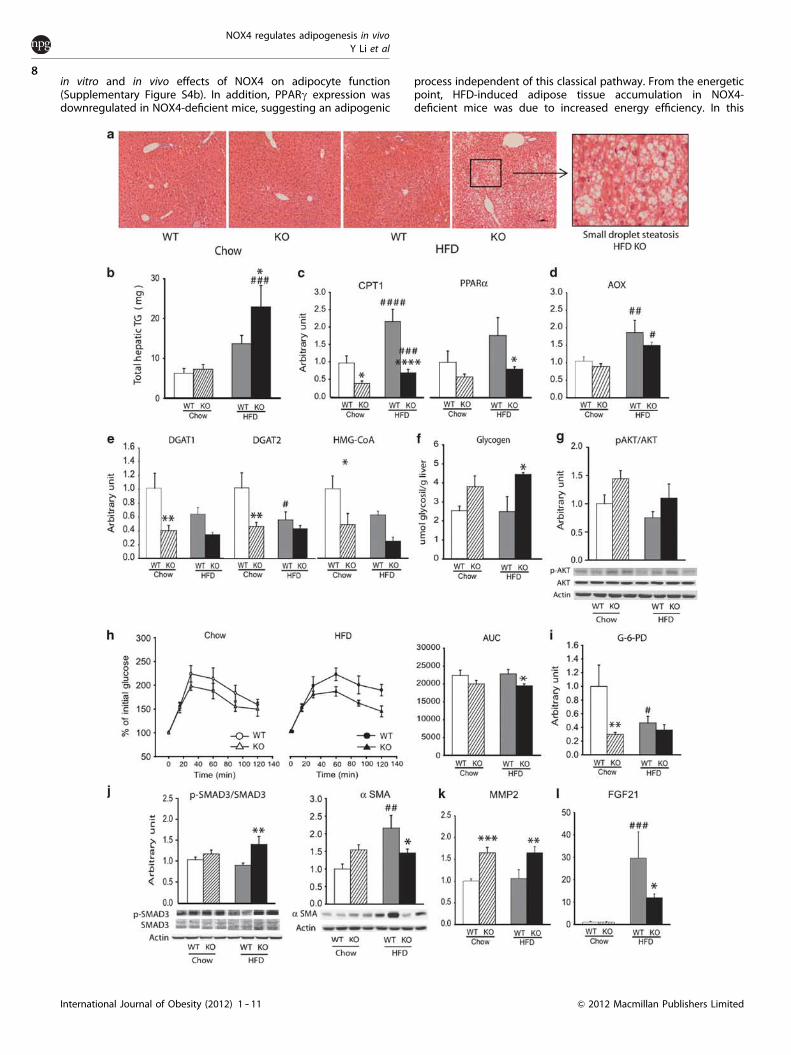
HFD-induced liver steatosis in NOX4-deficient miceObesity is associated with the occurrence of non-alcoholicfatty liver disease, characterized by intrahepatic triglyceride
accumulation. In line with their adiposity, HFD-fed NOX4-deficientmice displayed elevated hepatic triglyceride storage in the form ofsmall lipid droplets (Figures 5a and b). Accordingly, liver weightwas increased in HFD-fed NOX4-deficient mice compared withHFD-fed WT mice (1.51±0.24gr and 1.76±0.33gr, WT vs NOX4-deficient mice, respectively, P¼ 0.041, Student’s t test). Steatosisdevelops when the rate of fatty acid (FA) input (uptake andsynthesis) is greater than the rate of FA output (secretion andoxidation). HFD-fed NOX4-deficient mice displayed comparablelevels of the FA transporter CD36 (FA uptake), a decrease in thelevels of the acetyl-coA carboxylase (ACC) (the rate-limitingenzyme of FA synthesis) and equal amounts of the microsomaltriglyceride transfer protein (MTTP) (triglyceride export), suggest-ing that these processes do not have a crucial role in the onset ofexacerbated hepatosteatosis (Supplementary Figures S3a -- c). Bycontrast, a role for decreased mitochondrial FA b-oxidation wasimplied by the lack of diet-induced upregulation of the expressionof the rate-limiting enzyme carnitin-palmitoyl transport protein 1(CPT-1). CPT-1 expression is regulated by the transcription factorPPARa, and impaired PPARa activity leads to increased fat storagein liver.33,34 PPARa levels were decreased in HFD-fed NOX4-deficient mice (Figure 5c). No alteration was observed in theexpression of acyl-coA oxidase (AOX), the rate-limiting enzyme ofperoxisomal FA oxidation (Figure 5d). Serum ketone bodyconcentrations were elevated in NOX4-deficient mice alreadyafter 2 weeks of HFD feeding (218±9.21 and 337±13.9mM for WTand for NOX4-deficient mice, respectively, Student’s t-testPo0.001). Liver triglyceride synthesis is mediated by diglycerideacyltransferases 1 and 2 (DGAT1 and 2). 3-Hydroxy-3-methylglu-taryl-coenzyme A synthase (HMG-CoA synthase) is the rate-limiting enzyme of cholesterol biosynthesis. Both DGAT 1/2 andHMG-CoA synthase expressions were downregulated in chow diet-fed NOX4-deficient mice reaching similar levels than in HFD-fedWT mice (Figure 5e).
Liver is a major site of glucose homeostasis regulationthrough its glycogen storage and neoglucogenic capacity.Liver glycogen content was elevated in HFD-fed NOX4-deficient mice (Figure 5f). AKT phosphorylation tended to beelevated, though this increase did not reach statistical significance(Figure 5g). Neoglucogenesis was decreased in HFD-fedNOX4-deficient mice, as shown by lower glucose levels inresponse to intraperitoneal injection of pyruvate, the majorsubstrate of glucose synthesis (ipPTT) (Figure 5h). Expressionof the neoglucogenic enzymes glucose-6 phosphatase (G-6-Pase)and phospho-enol-carboxykinase (PEPCK) showed a slightbut statistically non-significant decrease, pointing towardsa regulation at the level of enzyme activity (SupplementaryFigure S3d).
NOX4 possesses constitutive NADPH oxidase activity transform-ing NADPH into NADPþ .4 Glucose-6-phosphate-1-dehydrogenase(G-6-PD) is the rate-limiting enzyme of the alternative glucoseutilization pathway (pentose-phosphate shunt) that is regulated inresponse to alterations in the cellular NADPH/NADPþ ratio. Theexpression of G-6-PD was significantly decreased in chow diet-fedNOX4-deficient mice reaching levels that were comparable toHFD-fed WT mice (Figure 5i).
Liver metabolic disturbances may lead to the activation ofhepatic stellate cells triggering Smad3-mediated cytoskeletalre-organization and the induction of a-smooth muscle actin (a-SMA)expression.35 Along with increased liver lipid deposition HFD-fedNOX4-deficient mice presented higher degree of Smad3 phos-phorylation (activation) but failed to upregulate a-SMA proteinlevels (Figure 5j). Activated hepatic stellate cells synthesize matrixMMP-2 to degrade normal extracellular matrix.36 MMP2 expressionwas increased in the livers of NOX4-deficient mice upon bothchow and HFD administration (Figure 5k). Liver steatosis isreversed by fibroblast growth factor-21 (FGF-21) in differentrodent models of insulin resistance.37 -- 39 Human non-alcoholic
NOX4 regulates adipogenesis in vivoY Li et al
5
International Journal of Obesity (2012) 1 -- 11& 2012 Macmillan Publishers Limited
steatohepatitis, a histological entity with propensity to progresstowards liver fibrosis, is characterized by lower hepatic FGF-21expression compared to non-alcoholic fatty liver disease.40 HFD
feeding induced a significant increase in FGF-21 expression in WTmice but to a much lesser extent in NOX4-deficient mice(Figure 5l).
0
5
10
15
20
WT
Chow HFD
0
20
40
60
80
100
120
Chow
0.0
0.2
0.4
0.6
Arb
itrar
y un
it
Arb
itrar
y un
it
Arb
itrar
y un
it
Arb
itrar
y un
it
Arb
itrar
y un
it
0.8
1.0
1.2
1.4
MMP2
Actin
***###
###***
*#
###**
###
###*
Diameter(μm)
WT Chow WT HFD
KO HFDKO Chow
Chow HFD
0
1
2
3
4
0
1
2
3
4
VEGF
Actin
#
#
Pref-1
VEGF
Chow HFD
0
1
2
3
4
5 SPARC
##**
**###
Chow HFD
Chow HFD
MMP2 Leptin
Chow HFD
0
5
10
15
20
% o
f tot
al c
ells
25
30
35
Actin
0
1
2
3
4
5
6 F 4 / 80
#
##*
F4/80
Chow HFD
0
2
4
6
8
10
12
14
16
0
1
2
3
4
5
#
**
##**
Chow HFD
TNF α
###
PAI -1
Chow HFD
0.0
0.5
1.0
1.5
2.0
2.5 NF- κB
NF- kB
#
*
Actin
Chow HFD
###
0
1
2
3
4Cleaved Caspase 3
Cl-Caspase 3Actin
##*
Chow HFD
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
#####
PPAR γ
****
Chow HFD
PPAR γ
Actin
C/EBP α
0.0
0.5
1.0
1.5
2.0
Chow HFD
##**
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4GLUT 4
##*
Chow HFD
0.0
0.5
1.0
1.5
2.0HIF-1 α
##**
Chow HFD
02468
1012141618
***###
MCP 1
Chow HFD
Chow HFD
#**
miR-132
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Chow HFD
miR-134
#**
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
> 200μm
KOWTKO
diameter0~50 μm50~100 μm100~150 μm150~200 μm
(μm
)
KOWTKOWT
HFD
KOWTKOWT KOWTKOWT
KOWT KOWT KOWT KOWT
KOWT KOWT KOWT KOWT KOWTKOWT
KOWT KOWT
KOWTKOWT KOWT
KOWT KOWT
KOWT
KOWT KOWT
KOWT KOWT
KOWT KOWT KOWT KOWT KOWT KOWT KOWT KOWT
NOX4 regulates adipogenesis in vivoY Li et al
6
International Journal of Obesity (2012) 1 -- 11 & 2012 Macmillan Publishers Limited
DISCUSSIONIn this study, we show that NOX4 is a master regulator ofmetabolic homeostasis and that the absence of NOX4-mediatedROS production results in latent adipose tissue accumulation anda marked sensitivity towards diet-induced obesity and hepato-steatosis.
In the absence of nutritional challenge, NOX4-deficient micedisplayed slightly but significantly increased body weight from theage of 10 weeks. Body weight increase was linked to acceleratedadipose tissue differentiation and hypertrophy demonstrated bydecreased expression of the preadipocyte marker Pref-1 andlarger size adipocytes. HFD challenge led to massive obesityresulting from higher energy efficiency. HFD-induced obesity wasaccompanied by elevated circulating leptin levels and a small raisein calorie intake. This small increase (B20%) in food intake in HFD-fed NOX4-deficient mice did not account for the development ofobesity (200% increase in WAT) and, interestingly, was not relatedto a resistance to leptin’s inhibitory effect on orexigenicneuropeptide expression. Indeed, the levels of neuropeptide Yand Agouti-related peptide were decreased in HFD-fed
NOX4-deficient mice, suggesting a leptin-independent regulationof hypothalamic appetite centers (Supplementary Figure S4a).These data are in line with several other studies showing thecritical role of hypothalamic ROS production in appetite regula-tion.41 Although in our current study, using whole body NOX4-deficient mice, HFD diet-induced obesity was not a consequenceof increased calorie intake but an increase in energy efficiency, therole of NOX4 in hypothalamic function remains an intriguingquestion for the future investigations.
HFD-induced adipose tissue accumulation was accompanied bya molecular signature of intensive extracellular matrix remodeling(increased MMP-2 and SPARC expressions) and enhancedadipocyte differentiation (increased leptin expression), suggestingan anti-adipogenic role for NOX4 and a predisposition towardsHFD-induced obesity in the absence of NOX4-derived ROSproduction. In vitro, decreased NOX4 function was linked todecreased p44/42MAPK activation and mitogenesis, and aconcurrent enhancement of PPARg2-mediated adipogenesis.10
In vivo, in NOX4-deficient mice, we did not observe changes inMAPK phosphorylation most likely due to differences between the
Figure 3. HFD-induced adipose tissue hypertrophy, hypoxia and inflammation in NOX4-deficient mice. (a) Increased adipocyte size inNOX4-deficient (NOX4 KO) mice. Representative hematoxylin/eosin-stained sections of epididymal WAT and their computer-assistedconversion to indicate adipocyte size as described in Methods and detailed in Supplementary Figure S7. Quantification was derived from5--6 sections/mouse and are shown as mean±s.e.m. (b) Adipocyte distribution according to cell diameter in epidydimal WAT. (c --h) Increaseddifferentiation and remodeling in the WAT of NOX4 KO mice. Real-time PCR and western blot analysis of gene and protein expression asindicated. Below each western blot analysis a representative blot is shown. (c) Preadipocyte factor 1 (Pref-1), (d) secreted protein acidiccystein-rich (SPARC), (e) vascular endothelial growth factor (VEGF), (f ) metalloproteinase 2 (MMP2), (g) leptin. (h -- j) HFD-induced hypoxia andinflammation in NOX4 KO mice. Real-time PCR and western blot analysis of the expression of the (h, i) hypoxia-inducible factor 1 alpha(HIF-1a), monocyte chemoattractant protein 1 (MCP-1) and the macrophage marker F4/80, (j) the pro-inflammatory cytokines tumornecrosis factor alpha (TNFa) and plasminogen activator inhibitor-1(PAI-1) and (k) NF-kB, the master regulator of inflammatory response.(l) Increased caspase 3 activation in NOX4 KO mice. (m, n) Expression of peroxisome proliferator-activated receptor gamma (PPARg), C/EBPaand GLUT4. (o) Real-time PCR measurements of the indicated miRNA134, miRNA132. Data in all graphs represent mean±s.e.m.; n¼ 7 forWT and NOX4 KO mice on chow diet (chow), n¼ 5 for WT and n¼ 6 for NOX4 KO mice on high-fat diet (HFD); *Po0.05, **Pp0.01, ***Pp0.001,****Pp0.0001 WT vs NOX4 KO; #Po0.05, ##Pp0.01, ###Pp0.001, ####Pp0.001 chow vs HFD.
Figure 4. Increased cold tolerance in NOX4-deficient mice. (a) Increased brown adipose tissue mass in HFD-fed NOX4-deficient (NOX4 KO)mice. (b) Increased lipid accumulation in the brown adipose tissue in NOX4 KO mice. A representative hematoxylin/eosin-stained sectionis shown for each condition. (c, d) Real-time PCR analysis of PPARg and UCP-1 expression. (e) Increased cold tolerance in NOX4-deficientmice. Mice were transferred to 4 1C and their rectal temperature was measured on an hourly basis. Data in all graphs represent mean±s.e.m.; n¼ 7 for WT and NOX4 KO mice on chow diet (Chow), n¼ 5 for WT and n¼ 6 for NOX4 KO mice on high-fat diet (HFD);*Po0.05, **Pp0.01, ***Pp0.001 WT vs NOX4 KO; ##Pp0.01, chow vs HFD.
NOX4 regulates adipogenesis in vivoY Li et al
7
International Journal of Obesity (2012) 1 -- 11& 2012 Macmillan Publishers Limited
in vitro and in vivo effects of NOX4 on adipocyte function(Supplementary Figure S4b). In addition, PPARg expression wasdownregulated in NOX4-deficient mice, suggesting an adipogenic
process independent of this classical pathway. From the energeticpoint, HFD-induced adipose tissue accumulation in NOX4-deficient mice was due to increased energy efficiency. In this
NOX4 regulates adipogenesis in vivoY Li et al
8
International Journal of Obesity (2012) 1 -- 11 & 2012 Macmillan Publishers Limited

respect, elevated expressions of the mitochondrial uncouplingproteins UCP-2 (white adipose tissue) and UCP-1 (brown adiposetissue) could be regarded as compensatory mechanisms toenhance mitochondrial function and increase energy expenditure.Taken together, these data imply that the lack of NOX4-mediatedROS production leads to increased adipocyte differentiation andenhanced triglyceride storage independent of the classicaladipogenic (PPARg) and mitochondrial energy producing(UCP-1/UCP-2) pathways.
The abrupt increase in adipose tissue mass in NOX4-deficientmice brought on by HFD challenge provoked the development of‘obese’ adipose tissue, characterized by hypoxia, macrophagerecruitment and the production of inflammatory cytokinestriggering adipocyte apoptosis and the development of insulinresistance.22,42,43 Indeed, NOX4-deficient mice presented elevatedexpressions of HIF-1a and NF-kB, the master regulators of cellularhypoxic and inflammatory responses, and their target genes TNFa,MCP-1, PAI-1 and VEGF along with enhanced activation ofcaspase3, the executional factor of apoptosis. The onset of wholebody insulin resistance was evidenced by increased HOMA valuesalready at very early time points (4 weeks) of HFD feeding.Enhanced NOX4 expression and NOX4-derived ROS productionhave been suggested to contribute to HFD-induced adipose tissueoxidative stress and insulin resistance in a mouse model ofobesity.13 In addition, an upregulation of NOX4 in the adiposetissue of obese Pima Indians compared with lean subjects wasdetected.44 Our study confirmed upregulation of adipose tissueNOX4 expression upon HFD feeding (Supplementary Figure S5).However, in view of the results of our current study NOX4upregulation should be regarded as a defensive mechanism tolimit adipose tissue expansion.
HFD-induced adipose tissue accumulation in NOX4-deficientmice was accompanied by exaggerated hepatosteatosis, one ofthe major obesity-related complications. Excessive liver triglycer-ide deposition was associated with a failure to upregulate theexpression of CPT-1, the rate-limiting enzyme of liver mitochon-drial FA oxidation. Decreased FA oxidation is an important factorin the development of liver steatosis.45,46 In the absence ofappropriate CPT-1 activity, FA oxidation is incomplete, leading tothe formation of ketone bodies. Indeed, NOX4-deficient micepresented higher ketone body levels after already 2 weeks of HFDregime. A sovereign role for NOX4-induced ROS production in liverlipid accumulation was also suggested by the compensatorydownregulation of the triglyceride synthesis enzymes DGAT1 andDGAT2 observed in chow-diet fed NOX4-deficient mice that wereundistinguishable from the levels detected in HFD-fed WT mice.NOX4 is a key mediator of TGF-b action in hepatocytes.47 TGF-bproduction is elevated in steatotic livers triggering fibrosisthrough upregulation of a-SMA production by activated hepaticstellate cells.36 In line with these data, NOX4-deficient mice failedto upregulate a-SMA expression in response to HFD feeding. Inaddition, expression of MMP-2 was elevated compared to HFD-fedWT mice, suggesting more intensive extracellular matrix break-down and a defense against fibrotic tissue formation. These dataare in agreement with studies demonstrating that diminishedNOX4 function protects against the development of fibrosis in the
lung.15,48 It is of importance that non-alcoholic fatty liver diseasewas suggested to be an independent risk factor for cardiovasculardisease, though some controversial issues still remained unre-solved.49 -- 51 NOX4 has been demonstrated to have a decisive rolein different cardiovascular pathologies.52,53 Thus, it is plausible tospeculate that NOX4 might constitute a common factor betweenthese two obesity-related pathologies, though complete explora-tion of this possible link will require further studies.
In a generalized way, our data demonstrated that the loss ofNOX4 leads to latent molecular alterations in several organs,specifically in adipose tissue and liver. These changes do notmanifest in an obvious phenotype if mice are kept in regularconditions but constitute a predisposition that can be broughtinto light by stress events for example, HFD feeding. Thus,mutations disabling NOX4 function could represent a state ofimbalance in adipocyte redox homeostasis that would lead to apredisposition towards diet-induced obesity and insulin resis-tance. Along this line, upregulation of NOX4 expression in obeseadipose tissue reported by others13,14 should be regarded as acounterregulatory mechanism to prevent adipose tissue hyper-trophy and the onset of insulin resistance. Of potentialsignificance is that in humans quantitative trait loci in the relationbetween obesity and diabetes was described in the chromosomeregion 11q14 -- 24, which also contains NOX4 (11q 14.2 -- 21).54 Anoverview of the predisposing role of NOX4-deficiency in thedevelopment of HFD-induced obesity and insulin resistance isdepicted in Figure 6.
In summary, this study was aimed at examining the con-sequences of the loss of NOX4 function on the regulation of wholebody metabolic and energy homeostasis. The results demonstratethat the lack of NOX4 function triggers a complex metabolicphenotype with adipose tissue hypertrophy and confers sensitivitytowards HFD-induced obesity and insulin resistance. These datasupport recent observations that adequate ROS production is avital component for the maintenance of whole body homeostasis
Figure 5. Hepatosteatosis in NOX4-deficient mice. (a) Microsteatosis in NOX4-deficient (NOX4 KO) HFD mice. Representative hematoxylin/eosin-stained sections of liver. (b) Increased liver triglyceride content in NOX4 KO mice. (c --e) Decreased fatty acid oxidation in NOX4-deficient mice. Real-time PCR and western blot analysis of liver lipid enzymes. Carnitine palmitoyl transferase 1 (CPT1), peroxisomeproliferator-activated receptor-alpha (PPARa), acyl-coA oxidase (AOX), diacyl-glycerol transferase 1 and 2 (DGAT1, DGAT2), hydroxymethyl-glutaryl coA (HMG-coA). (f ) Increased hepatic glycogen content in NOX4 KO mice. (g) Changes in AKT phosphorylation. (h) Decreasedpyruvate sensitivity in HFD-fed NOX4 KO mice. AUC, area under curve. (i) Real-time PCR analysis glucose-6-phosphate dehydrogenase(G-6-PD). (j -- l) Western blot and real-time PCR analysis of the proteins and genes indicated. a-Smooth muscle actin (a SMA), matrixmetalloproteinase 2 (MMP2), fibroblast growth factor 21 (FGF-21). Data in all graphs represent mean±s.e.m.; n¼ 7 for WT and NOX4 KOmice on chow diet (chow), n¼ 5 for WT and n¼ 6 for NOX4 KO mice on high-fat diet (HFD); *Po0.05, **Pp0.01, ***Pp0.001, ****Pp0.0001 WTvs NOX4 KO; #Po0.05, ##Pp0.01, ###Pp0.001, ###Pp0.001 chow vs HFD.
NOX4 deficiency
Adipocyte differentiation and hypertrophy
WAT accumulation
Hypoxia Macrophageinfiltration
Inflammatorycytokines
Insulin resistanceHepatosteatosis
ApoptosisOxidative
stress
HFD
+
+
+
Figure 6. NOX4 deficiency predisposes towards HFD-inducedobesity and insulin resistance. NOX4 deficiency results in increasedadipocyte differentiation and hypertrophy, leading to WATaccumulation, hypoxia and inflammation in vivo. These alterationsare latent but can be brought into light by nutritional challengeresulting in overt obesity, insulin resistance and hepatosteatosis.
NOX4 regulates adipogenesis in vivoY Li et al
9
International Journal of Obesity (2012) 1 -- 11& 2012 Macmillan Publishers Limited

and insulin sensitivity but also add a new dimension by definingNOX4 as a primary ROS source regulating adipose tissue functionin this context.
CONFLICT OF INTERESTThe authors declare no conflict of interest.
ACKNOWLEDGEMENTSThis work was supported by The Swiss National Science Foundation (Grant No.3100A0-122327) to IS, 3100A0-103725 to KHK and 310030-141162 to PM. We thankF Rohner-Jeanrenaud and C Wollheim (University of Geneva) for helpful discussionsand critical reading of the manuscript. We are grateful to C Chapponnier (Universityof Geneva) for providing the a-SMA antibody. We thank our collaborators at theUniversity of Geneva: Olivier Plastre and Dorothee Rigo for the excellent technicalassistance, Sergei Startcsik for the computer-assisted analysis of adipose tissue,Patrick Descombes and Didier Chollet for the miRNA analysis and AnneliseWohlwend for help with histological evaluation.
REFERENCES1 Bedard K, Krause KH. The NOX family of ROS-generating NADPH oxidases:
physiology and pathophysiology. Physiol Rev 2007; 87: 245 -- 313.2 Geiszt M, Kopp JB, Varnai P, Leto TL. Identification of renox, an NAD(P)H oxidase in
kidney. Proc Natl Acad Sci USA 2000; 97: 8010 -- 8014.3 Shiose A, Kuroda J, Tsuruya K, Hirai M, Hirakata H, Naito S et al. A novel
superoxide-producing NAD(P)H oxidase in kidney. J Biol Chem 2001; 276:1417 -- 1423.
4 Serrander L, Cartier L, Bedard K, Banfi B, Lardy B, Plastre O et al. NOX4 activity isdetermined by mRNA levels and reveals a unique pattern of ROS generation.Biochem J 2007; 406: 105 -- 114.
5 Meng D, Lv DD, Fang J. Insulin-like growth factor-I induces reactive oxygenspecies production and cell migration through Nox4 and Rac1 in vascular smoothmuscle cells. Cardiovasc Res 2008; 80: 299 -- 308.
6 Mahadev K, Motoshima H, Wu X, Ruddy JM, Arnold RS, Cheng G et al. TheNAD(P)H oxidase homolog Nox4 modulates insulin-stimulated generation ofH2O2 and plays an integral role in insulin signal transduction. Mol Cell Biol 2004;24: 1844 -- 1854.
7 Loh K, Deng H, Fukushima A, Cai X, Boivin B, Galic S et al. Reactive oxygen speciesenhance insulin sensitivity. Cell Metab 2009; 10: 260 -- 272.
8 Entingh AJ, Taniguchi CM, Kahn CR. Bi-directional regulation of brown fatadipogenesis by the insulin receptor. J Biol Chem 2003; 278: 33377 -- 33383.
9 Bluher M, Michael MD, Peroni OD, Ueki K, Carter N, Kahn BB et al. Adipose tissueselective insulin receptor knockout protects against obesity and obesity-relatedglucose intolerance. Dev Cell 2002; 3: 25 -- 38.
10 Schroder K, Wandzioch K, Helmcke I, Brandes RP. Nox4 acts as a switch betweendifferentiation and proliferation in preadipocytes. Arterioscler Thromb Vasc Biol2009; 29: 239 -- 245.
11 Mouche S, Mkaddem SB, Wang W, Katic M, Tseng YH, Carnesecchi S et al. Reducedexpression of the NADPH oxidase NOX4 is a hallmark of adipocyte differentiation.Biochim Biophys Acta 2007; 1773: 1015 -- 1027.
12 Goossens GH. The role of adipose tissue dysfunction in the pathogenesis ofobesity-related insulin resistance. Physiol Behav 2008; 94: 206 -- 218.
13 Furukawa S, Fujita T, Shimabukuro M, Iwaki M, Yamada Y, Nakajima Y et al.Increased oxidative stress in obesity and its impact on metabolic syndrome. J ClinInvest 2004; 114: 1752 -- 1761.
14 Park HS, Jin DK, Shin SM, Jang MK, Longo N, Park JW et al. Impaired generation ofreactive oxygen species in leprechaunism through downregulation of Nox4.Diabetes 2005; 54: 3175 -- 3181.
15 Carnesecchi S, Deffert C, Donati Y, Basset O, Hinz B, Preynat-Seauve O et al. A keyrole for NOX4 in epithelial cell death during development of lung fibrosis. AntioxidRedox Signal 15: 607 -- 619.
16 Somm E, Henrichot E, Pernin A, Juge-Aubry CE, Muzzin P, Dayer JM et al.Decreased fat mass in interleukin-1 receptor antagonist-deficient mice: impact onadipogenesis, food intake, and energy expenditure. Diabetes 2005; 54:3503 -- 3509.
17 Veyrat-Durebex C, Montet X, Vinciguerra M, Gjinovci A, Meda P, Foti M et al. TheLou/C rat: a model of spontaneous food restriction associated with improvedinsulin sensitivity and decreased lipid storage in adipose tissue. Am J PhysiolEndocrinol Metab 2009; 296: E1120 -- E1132.
18 Baran K, Preston E, Wilks D, Cooney GJ, Kraegen EW, Sainsbury A. Chronic centralmelanocortin-4 receptor antagonism and central neuropeptide-Y infusion in ratsproduce increased adiposity by divergent pathways. Diabetes 2002; 51: 152 -- 158.
19 Crosson SM, Khan A, Printen J, Pessin JE, Saltiel AR. PTG gene deletion causesimpaired glycogen synthesis and developmental insulin resistance. J Clin Invest2003; 111: 1423 -- 1432.
20 Skurk T, Alberti-Huber C, Herder C, Hauner H. Relationship between adipocyte size andadipokine expression and secretion. J Clin Endocrinol Metab 2007; 92: 1023 -- 1033.
21 Johnson JA, Trasino SE, Ferrante Jr AW, Vasselli JR. Prolonged decrease ofadipocyte size after rosiglitazone treatment in high- and low-fat-fed rats. Obesity(Silver Spring) 2007; 15: 2653 -- 2663.
22 Hosogai N, Fukuhara A, Oshima K, Miyata Y, Tanaka S, Segawa K et al. Adiposetissue hypoxia in obesity and its impact on adipocytokine dysregulation. Diabetes2007; 56: 901 -- 911.
23 Ye J, Gao Z, Yin J, He Q. Hypoxia is a potential risk factor for chronic inflammationand adiponectin reduction in adipose tissue of ob/ob and dietary obese mice. AmJ Physiol Endocrinol Metab 2007; 293: E1118 -- E1128.
24 Wood IS, de Heredia FP, Wang B, Trayhurn P. Cellular hypoxia and adipose tissuedysfunction in obesity. Proc Nutr Soc 2009; 68: 370 -- 377.
25 Lee MJ, Wu Y, Fried SK. Adipose tissue remodeling in pathophysiology of obesity.Curr Opin Clin Nutr Metab Care 2010; 13: 371 -- 376.
26 Alkhouri N, Gornicka A, Berk MP, Thapaliya S, Dixon LJ, Kashyap S et al. Adipocyteapoptosis, a link between obesity, insulin resistance, and hepatic steatosis. J BiolChem 2010; 285: 3428 -- 3438.
27 Anghel SI, Wahli W. Fat poetry: a kingdom for PPAR gamma. Cell Res 2007; 17:486 -- 511.
28 Armoni M, Kritz N, Harel C, Bar-Yoseph F, Chen H, Quon MJ et al. Peroxisomeproliferator-activated receptor-gamma represses GLUT4 promoter activity inprimary adipocytes, and rosiglitazone alleviates this effect. J Biol Chem 2003; 278:30614 -- 30623.
29 Stephens JM, Pekala PH. Transcriptional repression of the GLUT4 and C/EBP genesin 3T3-L1 adipocytes by tumor necrosis factor-alpha. J Biol Chem 1991; 266:21839 -- 21845.
30 Kloting N, Berthold S, Kovacs P, Schon MR, Fasshauer M, Ruschke K et al. MicroRNAexpression in human omental and subcutaneous adipose tissue. PLoS One 2009; 4: e4699.
31 Takanabe R, Ono K, Abe Y, Takaya T, Horie T, Wada H et al. Up-regulatedexpression of microRNA-143 in association with obesity in adipose tissue of micefed high-fat diet. Biochem Biophys Res Commun 2008; 376: 728 -- 732.
32 Kim YJ, Hwang SJ, Bae YC, Jung JS. MiR-21 regulates adipogenic differentiationthrough the modulation of TGF-beta signaling in mesenchymal stem cells derivedfrom human adipose tissue. Stem Cells 2009; 27: 3093 -- 3102.
33 Sharma MR, Polavarapu R, Roseman D, Patel V, Eaton E, Kishor PB et al.Transcriptional networks in a rat model for nonalcoholic fatty liver disease: amicroarray analysis. Exp Mol Pathol 2006; 81: 202 -- 210.
34 Reddy JK, Rao MS. Lipid metabolism and liver inflammation. I. Hepatic fatty aciduptake: possible role in steatosis. Am J Physiol Gastrointest Liver Physiol 2006; 290:G852 -- G858.
35 Uemura M, Swenson ES, Gaca MD, Giordano FJ, Reiss M, Wells RG. Smad2 andSmad3 play different roles in rat hepatic stellate cell function and alpha-smoothmuscle actin organization. Mol Biol Cell 2005; 16: 4214 -- 4224.
36 Brenner DA. Molecular pathogenesis of liver fibrosis. Trans Am Clin Climatol Assoc2009; 120: 361 -- 368.
37 Xu J, Lloyd DJ, Hale C, Stanislaus S, Chen M, Sivits G et al. Fibroblast growth factor21 reverses hepatic steatosis, increases energy expenditure, and improves insulinsensitivity in diet-induced obese mice. Diabetes 2009; 58: 250 -- 259.
38 Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Micanovic R, Galbreath EJet al. FGF-21 as a novel metabolic regulator. J Clin Invest 2005; 115: 1627 -- 1635.
39 Kharitonenkov A, Wroblewski VJ, Koester A, Chen YF, Clutinger CK, Tigno XT et al.The metabolic state of diabetic monkeys is regulated by fibroblast growthfactor-21. Endocrinology 2007; 148: 774 -- 781.
40 Dushay J, Chui PC, Gopalakrishnan GS, Varela-Rey M, Crawley M, Fisher FM et al.Increased fibroblast growth factor 21 in obesity and nonalcoholic fatty liverdisease. Gastroenterology 2010; 139: 456 -- 463.
41 Horvath TL, Andrews ZB, Diano S. Fuel utilization by hypothalamic neurons: rolesfor ROS. Trends Endocrinol Metab 2009; 20: 78 -- 87.
42 Schenk S, Saberi M, Olefsky JM. Insulin sensitivity: modulation by nutrients andinflammation. J Clin Invest 2008; 118: 2992 -- 3002.
43 Trayhurn P, Wang B, Wood IS. Hypoxia and the endocrine and signalling role ofwhite adipose tissue. Arch Physiol Biochem 2008; 114: 267 -- 276.
44 Lee YH, Nair S, Rousseau E, Allison DB, Page GP, Tataranni PA et al. Microarrayprofiling of isolated abdominal subcutaneous adipocytes from obese vs non-obese Pima Indians: increased expression of inflammation-related genes.Diabetologia 2005; 48: 1776 -- 1783.
45 Bradbury MW. Lipid metabolism and liver inflammation. I. Hepatic fatty aciduptake: possible role in steatosis. Am J Physiol Gastrointest Liver Physiol 2006; 290:G194 -- G198.
46 Hijona E, Hijona L, Arenas JI, Bujanda L. Inflammatory mediators of hepaticsteatosis. Mediators Inflamm 2010; 2010: 837419.
NOX4 regulates adipogenesis in vivoY Li et al
10
International Journal of Obesity (2012) 1 -- 11 & 2012 Macmillan Publishers Limited

47 Carmona-Cuenca I, Roncero C, Sancho P, Caja L, Fausto N, Fernandez M et al.Upregulation of the NADPH oxidase NOX4 by TGF-beta in hepatocytes is requiredfor its pro-apoptotic activity. J Hepatol 2008; 49: 965 -- 976.
48 Hecker L, Vittal R, Jones T, Jagirdar R, Luckhardt TR, Horowitz JC et al. NADPHoxidase-4 mediates myofibroblast activation and fibrogenic responses to lunginjury. Nat Med 2009; 15: 1077 -- 1081.
49 Ghouri N, Preiss D, Sattar N. Liver enzymes, nonalcoholic fatty liver disease, andincident cardiovascular disease: a narrative review and clinical perspective ofprospective data. Hepatology 2010; 52: 1156 -- 1161.
50 Dunn W, Xu R, Wingard DL, Rogers C, Angulo P, Younossi ZM et al. Suspectednonalcoholic fatty liver disease and mortality risk in a population-based cohortstudy. Am J Gastroenterol 2008; 103: 2263 -- 2271.
51 McKimmie RL, Daniel KR, Carr JJ, Bowden DW, Freedman BI, RegisterTC et al. Hepatic steatosis and subclinical cardiovascular disease in a cohortenriched for type 2 diabetes: the Diabetes Heart Study. Am J Gastroenterol 2008;103: 3029 -- 3035.
52 Rivera J, Sobey CG, Walduck AK, Drummond GR. Nox isoforms in vascularpathophysiology: insights from transgenic and knockout mouse models. RedoxRep 2010; 15: 50 -- 63.
53 Nabeebaccus A, Zhang M, Shah AM. NADPH oxidases and cardiac remodelling.Heart Fail Rev 2011; 16: 5 -- 12.
54 van Tilburg JH, Sandkuijl LA, Strengman E, Pearson PL, van Haeften TW,Wijmenga C. Variance-component analysis of obesity in type 2 diabetes confirmsloci on chromosomes 1q and 11q. Obes Res 2003; 11: 1290 -- 1294.
Supplementary Information accompanies the paper on International Journal of Obesity website (http://www.nature.com/ijo)
NOX4 regulates adipogenesis in vivoY Li et al
11
International Journal of Obesity (2012) 1 -- 11& 2012 Macmillan Publishers Limited
Related Documents











