Hypoxia-Dependent Regulation of Nonphagocytic NADPH Oxidase Subunit NOX4 in the Pulmonary Vasculature Manish Mittal, Markus Roth, Peter Ko ¨nig, Simone Hofmann, Eva Dony, Parag Goyal, Anne-Christin Selbitz, Ralph Theo Schermuly, Hossein Ardeschir Ghofrani, Grazyna Kwapiszewska, Wolfgang Kummer, Walter Klepetko, Mir Ali Reza Hoda, Ludger Fink, Jo ¨rg Ha ¨nze, Werner Seeger, Friedrich Grimminger, Harald H.H.W. Schmidt, Norbert Weissmann Abstract—Nonphagocytic NADPH oxidases have recently been suggested to play a major role in the regulation of physiological and pathophysiological processes, in particular, hypertrophy, remodeling, and angiogenesis in the systemic circulation. Moreover, NADPH oxidases have been suggested to serve as oxygen sensors in the lung. Chronic hypoxia induces vascular remodeling with medial hypertrophy leading to the development of pulmonary hypertension. We screened lung tissue for the expression of NADPH oxidase subunits. NOX1, NOXA1, NOXO1, p22phox, p47phox, p40phox, p67phox, NOX2, and NOX4 were present in mouse lung tissue. Comparing mice maintained for 21 days under hypoxic (10% O 2 ) or normoxic (21% O 2 ) conditions, an upregulation exclusively of NOX4 mRNA was observed under hypoxia in homogenized lung tissue, concomitant with increased levels in microdissected pulmonary arterial vessels. In situ hybridization and immunohistological staining for NOX4 in mouse lungs revealed a localization of NOX4 mRNA and protein predominantly in the media of small pulmonary arteries, with increased labeling intensities after chronic exposure to hypoxia. In isolated pulmonary arterial smooth muscle cells (PASMCs), NOX4 was localized primarily to the perinuclear space and its expression levels were increased after exposure to hypoxia. Treatment of PASMCs with siRNA directed against NOX4 decreased NOX4 mRNA levels and reduced PASMC proliferation as well as generation of reactive oxygen species. In lungs from patients with idiopathic pulmonary arterial hypertension (IPAH), expression levels of NOX4, which was localized in the vessel media, were 2.5-fold upregulated. These results support an important role for NOX4 in the vascular remodeling associated with development of pulmonary hypertension. (Circ Res. 2007;101:258-267.) Key Words: hypoxia hypoxic pulmonary vasoconstriction NADPH oxidase pulmonary hypertension vascular smooth muscle cell proliferation T he NADPH oxidases are superoxide-generating enzymes that release superoxide by electron transfer from NADPH to oxygen. The classical leukocyte NADPH oxidase plays an important role in host defense against bacterial and fungal pathogens. 1,2 This phagocytic type of NADPH oxidase consists of 2 membrane-bound subunits, gp91 phox and p22 phox which form the flavocytochrome b 558 complex, together with the cyctosolic subunits p40 phox , p47 phox , and p67 phox . Superox- ide production by this complex is induced by assembly of the cytosolic and membrane-bound subunits. Such an assembly can be induced by the phosphorylation of p47 phox . 3 Rac GTPases are also involved in this activation process. Re- cently, several additional isoforms of the membrane-bound subunit gp91 phox have been described. The first described homolog of gp91 phox , called mox1 (later NOX1), is primarily expressed in the colon and is suggested to be involved in mitogenic activity. 4 Additional homologs, including NOX3, NOX4 (Renox), NOX5, Duox1, and Duox2, were subse- quently described. 5–8 According to this new nomenclature, gp91 phox is synonymous with NOX2. It was suggested that each of these homologs can replace gp91 phox in the NADPH oxidase complex, and it has been demonstrated that these nonphagocytic NADPH oxidases release lower amounts of superoxide. 9,10 However, very recently 2 new isoforms of the cytosolic subunits p47 phox and p67 phox have been identified. These new subunits, NOXO1 and NOXA1, have been dem- onstrated to interact with NOX1 to generate significant amounts of superoxide without being activated by protein kinase C– dependent phosphorylation. 11,12 Isoforms of gp91 phox have been identified in different organs and cell Original received January 8, 2007; revision received April 30, 2007; accepted June 8, 2007. From the University of Giessen Lung Center (M.M., M.R., S.H., E.D., P.G., A.-C.S., R.T.S., H.A.G., J.H., S.W., F.G., N.W.), Medical Clinic II/V, the Department of Anatomy and Cell Biology (P.K., W.K.), and the Department of Pathology (G.K., L.F.), Justus-Liebig-University, Giessen, Germany; the Department of Cardiothoracic Surgery (W.K., M.A.R.H.), University of Vienna, Austria; and the Department of Pharmacology (H.H.H.W.S.), Monash University, Melbourne, Australia. Present address for P.K.: Institut fu ¨r Anatomie, Universita ¨t zu Lu ¨beck, Germany. Correspondence to Norbert Weissmann, PhD, University of Giessen Lung Center, Medical Clinic II/V, Klinikstr. 36, D-35392 Giessen, Germany. E-mail [email protected] © 2007 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.107.148015 258 by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from by guest on September 10, 2015 http://circres.ahajournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hypoxia-Dependent Regulation of Nonphagocytic NADPHOxidase Subunit NOX4 in the Pulmonary Vasculature
Manish Mittal, Markus Roth, Peter Konig, Simone Hofmann, Eva Dony, Parag Goyal,Anne-Christin Selbitz, Ralph Theo Schermuly, Hossein Ardeschir Ghofrani, Grazyna Kwapiszewska,Wolfgang Kummer, Walter Klepetko, Mir Ali Reza Hoda, Ludger Fink, Jorg Hanze, Werner Seeger,
Friedrich Grimminger, Harald H.H.W. Schmidt, Norbert Weissmann
Abstract—Nonphagocytic NADPH oxidases have recently been suggested to play a major role in the regulation ofphysiological and pathophysiological processes, in particular, hypertrophy, remodeling, and angiogenesis in thesystemic circulation. Moreover, NADPH oxidases have been suggested to serve as oxygen sensors in the lung. Chronichypoxia induces vascular remodeling with medial hypertrophy leading to the development of pulmonary hypertension.We screened lung tissue for the expression of NADPH oxidase subunits. NOX1, NOXA1, NOXO1, p22phox, p47phox,p40phox, p67phox, NOX2, and NOX4 were present in mouse lung tissue. Comparing mice maintained for 21 days underhypoxic (10% O2) or normoxic (21% O2) conditions, an upregulation exclusively of NOX4 mRNA was observed underhypoxia in homogenized lung tissue, concomitant with increased levels in microdissected pulmonary arterial vessels. Insitu hybridization and immunohistological staining for NOX4 in mouse lungs revealed a localization of NOX4 mRNAand protein predominantly in the media of small pulmonary arteries, with increased labeling intensities after chronicexposure to hypoxia. In isolated pulmonary arterial smooth muscle cells (PASMCs), NOX4 was localized primarily tothe perinuclear space and its expression levels were increased after exposure to hypoxia. Treatment of PASMCs withsiRNA directed against NOX4 decreased NOX4 mRNA levels and reduced PASMC proliferation as well as generationof reactive oxygen species. In lungs from patients with idiopathic pulmonary arterial hypertension (IPAH), expressionlevels of NOX4, which was localized in the vessel media, were 2.5-fold upregulated. These results support an importantrole for NOX4 in the vascular remodeling associated with development of pulmonary hypertension. (Circ Res.2007;101:258-267.)
Key Words: hypoxia � hypoxic pulmonary vasoconstriction � NADPH oxidase � pulmonary hypertension� vascular smooth muscle cell proliferation
The NADPH oxidases are superoxide-generating enzymesthat release superoxide by electron transfer from
NADPH to oxygen. The classical leukocyte NADPH oxidaseplays an important role in host defense against bacterial andfungal pathogens.1,2 This phagocytic type of NADPH oxidaseconsists of 2 membrane-bound subunits, gp91phox and p22phox
which form the flavocytochrome b558 complex, together withthe cyctosolic subunits p40phox, p47phox, and p67phox. Superox-ide production by this complex is induced by assembly of thecytosolic and membrane-bound subunits. Such an assemblycan be induced by the phosphorylation of p47phox.3 RacGTPases are also involved in this activation process. Re-cently, several additional isoforms of the membrane-boundsubunit gp91phox have been described. The first describedhomolog of gp91phox, called mox1 (later NOX1), is primarily
expressed in the colon and is suggested to be involved inmitogenic activity.4 Additional homologs, including NOX3,NOX4 (Renox), NOX5, Duox1, and Duox2, were subse-quently described.5–8 According to this new nomenclature,gp91phox is synonymous with NOX2. It was suggested thateach of these homologs can replace gp91phox in the NADPHoxidase complex, and it has been demonstrated that thesenonphagocytic NADPH oxidases release lower amounts ofsuperoxide.9,10 However, very recently 2 new isoforms of thecytosolic subunits p47phox and p67phox have been identified.These new subunits, NOXO1 and NOXA1, have been dem-onstrated to interact with NOX1 to generate significantamounts of superoxide without being activated by proteinkinase C–dependent phosphorylation.11,12 Isoforms ofgp91phox have been identified in different organs and cell
Original received January 8, 2007; revision received April 30, 2007; accepted June 8, 2007.From the University of Giessen Lung Center (M.M., M.R., S.H., E.D., P.G., A.-C.S., R.T.S., H.A.G., J.H., S.W., F.G., N.W.), Medical Clinic II/V, the
Department of Anatomy and Cell Biology (P.K., W.K.), and the Department of Pathology (G.K., L.F.), Justus-Liebig-University, Giessen, Germany; theDepartment of Cardiothoracic Surgery (W.K., M.A.R.H.), University of Vienna, Austria; and the Department of Pharmacology (H.H.H.W.S.), MonashUniversity, Melbourne, Australia. Present address for P.K.: Institut fur Anatomie, Universitat zu Lubeck, Germany.
Correspondence to Norbert Weissmann, PhD, University of Giessen Lung Center, Medical Clinic II/V, Klinikstr. 36, D-35392 Giessen, Germany.E-mail [email protected]
© 2007 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.107.148015
258 by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from
types, including the colon, kidney, uterus, testis, liver, vas-cular smooth muscle cells, fibroblasts, endothelial cells,pancreatic islets, and lymphocytes.1,3,9,13–15 The NOX2 ho-mologs have been suggested to be associated with thedevelopment of atherosclerosis, systemic hypertension, dia-betic vascular disease, and diseases of the brain.16 The 2vascular isoforms, NOX1 and NOX4, are thought to play arole in vascular pathology.17,18 However, with respect to thelung, relatively few reports of expression and regulation ofthe recently-identified new vascular NADPH oxidase sub-units exist. Hoidal and coworkers19 have demonstrated thatNOX4 is the predominant homolog in human airway and
pulmonary artery smooth muscle cells.20 In addition Hohler etal identified a low output NADPH oxidase in pulmonaryartery endothelial cells.21 Against this background, wescreened the lung for expression of the new NADPH oxidaseisoforms. We sought to assess the regulation of the variousisoforms, including the classical phagocytic NADPH oxidasesubunits by hypoxia because (1) NADPH oxidases recentlyhave been proposed as possible pulmonary oxygen sensorsfor the acute response to lung alveolar hypoxia (hypoxicpulmonary vasoconstriction), (2) reactive oxygen species arethought to play a role in the vascular remodeling that occursduring chronic alveolar hypoxia,22,23 and (3) the phagocytic
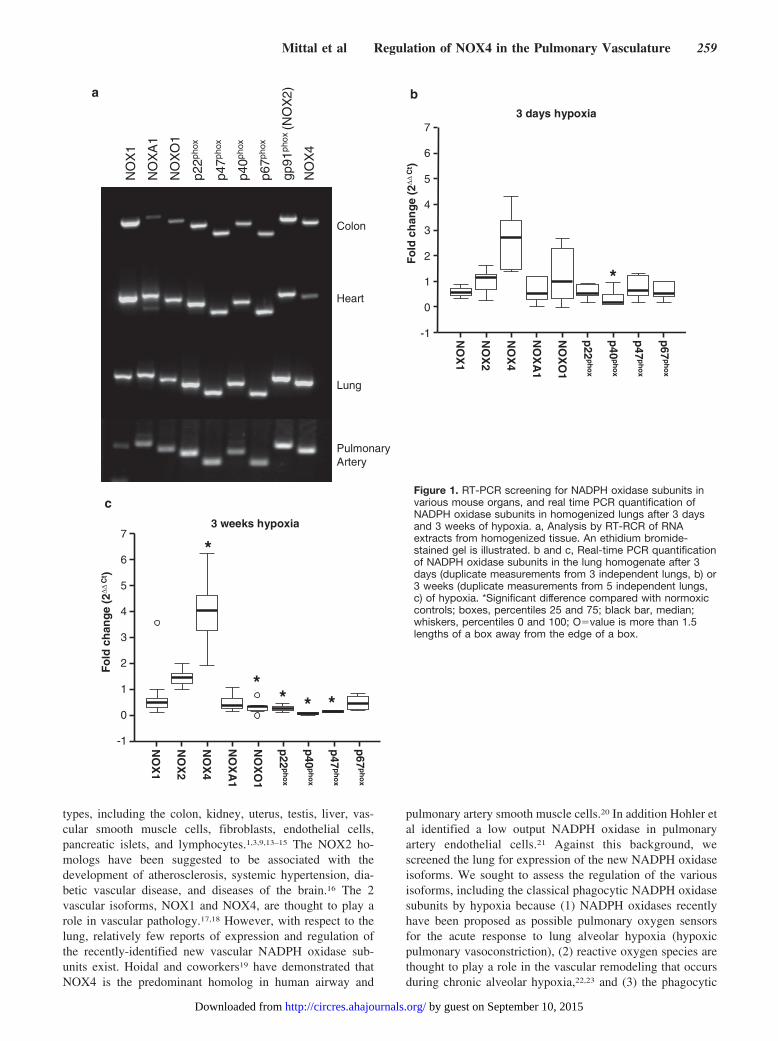
Figure 1. RT-PCR screening for NADPH oxidase subunits invarious mouse organs, and real time PCR quantification ofNADPH oxidase subunits in homogenized lungs after 3 daysand 3 weeks of hypoxia. a, Analysis by RT-RCR of RNAextracts from homogenized tissue. An ethidium bromide-stained gel is illustrated. b and c, Real-time PCR quantificationof NADPH oxidase subunits in the lung homogenate after 3days (duplicate measurements from 3 independent lungs, b) or3 weeks (duplicate measurements from 5 independent lungs,c) of hypoxia. *Significant difference compared with normoxiccontrols; boxes, percentiles 25 and 75; black bar, median;whiskers, percentiles 0 and 100; O�value is more than 1.5lengths of a box away from the edge of a box.
Mittal et al Regulation of NOX4 in the Pulmonary Vasculature 259
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

NADPH oxidase subunit NOX2 has recently been suggestedto play an important role in hypoxia-induced pulmonaryhypertension.24 The hypoxia-induced vascular remodelingprocess is characterized by hypertrophy and de novo muscu-larization of the vessel media, leading to a decrease invascular luminal area, increased vascular resistance, and thusdevelopment of pulmonary hypertension and right ventricularhypertrophy.25 In essence, we found that NOX4 is the onlysubunit prominently upregulated in pulmonary arterial ves-sels and in smooth muscle cells during chronic hypoxia, bothat the transcriptional and protein level. Cell culture experi-ments demonstrated a proproliferative activity of NOX4during hypoxia, because targeted knock-down of NOX4 withsiRNA suppressed pulmonary arterial smooth muscle cell(PASMC) proliferation. Most interestingly, NOX4 expres-sion was upregulated in the vessel media of lungs frompatients with idiopathic pulmonary hypertension (IPAH), incomparison to lungs from healthy donors, suggesting animportant role of this NADPH oxidase subunit in humanIPAH.
Materials and Methods
Chronic Hypoxia ExposureAll animal experiments were approved by local authorities. Mice(C57BL/6N) of either sex (Charles River Laboratories, Sulzfeld,Germany; 20 to 22 g) were exposed to normobaric hypoxia [inspira-tory O2 fraction (FiO2) 0.10] in a ventilated chamber for up to 3weeks as described previously.26
Mouse Lung Preparation for Laser AssistedMicrodissection and Right HeartHypertrophy AssessmentMouse lungs were prepared as described previously.27 For details seethe supplemental materials (available online at http://circres.ahajournals.org).
Laser-Assisted MicrodissectionLaser-microdissection was performed as described previously.27
RNA-Extraction and RT-PCRThe RNA was extracted from cells using guanidine thiocyanate-acidphenol (RNAzol B, WAK-Chemie, Germany) or with spin-columns
Figure 2. NOX4 and NOX2 mRNA quantification of microdissected pulmonary arteries by real time PCR during development of hypox-ia-induced pulmonary hypertension. a and b, Microdissected small pulmonary arteries (�100-�m diameter) from cryosections of mouselungs were used for the quantification of NOX4 and NOX2 mRNA. Mice were maintained under normoxic or hypoxic conditions for upto 21 days. The NOX2 and NOX4 mRNA levels were quantified by real-time PCR normalized to �2-microglobulin mRNA levels. c, DeltaCt values of NOX4 and NOX2 from microdissected mouse pulmonary arteries of normoxic mice. Values are duplicate measurements ofn�16 vessels from n�3 lungs each. d, Right ventricular hypertrophy after exposure of mice to chronic hypoxia. The right ventricle (RV)to left ventricle (LV) � septum (S) ratio was quantified from mouse hearts after exposure to chronic hypoxia (10% O2 for 3, 7, and 21days, respectively, n�5). *Significant differences as compared with normoxic controls.
260 Circulation Research August 3, 2007
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

(RNeasy, Qiagen, Germany). For details see the supplementalmaterials.
Real-Time PCRRelative quantification of the NADPH oxidase subunits was doneusing ABI prism 7700 detection system (Applied Biosystem, Weit-erstadt, Germany). For details please refer to the supplementalmaterials.
In Situ HybridizationFor a detailed description of the in situ hybridization protocol pleaserefer to the supplemental materials.
Immunohistochemistry for Mouse Lung SectionsImmunohistochemistry was performed as described previously.26
For details see the supplemental materials.
Immunohistochemistry for NOX4 and NOX2 inHuman Lung SectionsLung tissue samples from healthy individuals and from patients withIPAH were formalin-fixed, paraffin-embedded, and cut into 3-�msections. The immunostaining of the human lung sections wasperformed with a custom-made rabbit anti-human NOX4 polyclonalantibody29 or rabbit anti-human NOX2 polyclonal antibody (Upstate,Germany) as previously described.26,28
Western Blot of NOX4 in Frozen HumanLung TissueFor the detection of NOX4 by Western blot, a custom-madepolyclonal anti-NOX4 antibody raised in rabbits was used.28 Fordetails see the online supplemental materials.
Cell CultureSmooth muscle cells from human and murine pulmonary arterieswere isolated and cultured as described previously.29,30 For theinvestigation of the effect of hypoxia on NOX4 mRNA levels, cellswere either exposed to 1% O2 (hypoxia) or to 21% O2 (normoxia).
Immunocytochemistry of Murine PASMCsIsolated PASMCs were cultured on chamber slides, treated asindicated, fixed in ice cold acetone and methanol (1:1), and blocked
with 3% (m/v) BSA in PBS for 1 hour, followed by overnightincubation with an anti-NOX4 antibody (1:25) diluted in 3% (m/v)BSA in PBS.28 Indirect immunofluorescence was obtained byincubation with a Cy3-conjugated anti-goat antibody (Dako, Den-mark) diluted 1:100 in PBS for 90 minutes. Nuclear counterstainingwas performed with Hoechst-33258 (1:10 000 dilution in PBS;Invitrogen, Karlsruhe, Germany) for 10 minutes.
RNA Interference and Proliferation AssayA detailed description of the siRNA transfection and the proliferationassay is available in the supplemental materials.
StatisticsValues are given as mean�SEM if not indicated differently. Forstatistical analysis a Student t test was used for comparison of 2groups. For more than 2 groups, ANOVA with LSD posthoc test wasperformed. A probability value of less then 0.05 was consideredsignificant. Empirical assessment of NOX4 immunoreactive vesselswas performed in blinded fashion. Two conditions were evaluatedfor assessment of NOX4-immunoreactive vessels: first, the numberof NOX4-immunoreactive vessels different between the groups, andsecond, the mean diameter of NOX4-immunoreactive vessels differ-ent between the groups. Statistical analysis was performed by anonparametric variance analysis (Kruskal-Wallis test). If the proba-bility value in that test was �0.05, a comparison of the groupsbetween each other was performed using a Mann-Whitney test,where P�0.05 was regarded as significant. Comparison of groupswas stopped after P�0.05 to prevent �-inflation.
ResultsTo investigate the role of NADPH oxidases in the developmentof hypoxia-induced pulmonary hypertension, expression of theNADPH-oxidase subunits NOX1, NOX2, NOX4, p22phox,p40phox, p47phox, p67phox, as well as NOXO1 and NOXA1,was assessed by RT-PCR in different mouse tissues. As evidentfrom Figure 1a, all subunits were detected in the colon, heart,lung, and pulmonary arteries. In pulmonary arteries p22phox,p47phox, NOX2, and NOX4 appeared to be more prominentlyexpressed, as compared with the other subunits investigated(Figure 1a). Focusing on the hypoxic regulation of NADPH
Figure 3. Localization of NOX4 in mouse lung sections by nonisotopic in situ hybridization (NISH). a and b, Hybridization of the NOX4antisense probe to mouse lung cryosections (green fluorescence). c and d, The same sections stained with a Cy3-labeled antibodydirected against �-smooth muscle actin (SMA, red fluorescence). e and f, Overlay of the images a and b depicting predominant colo-calization of NOX4 transcripts with SMA in the smooth muscle cell layer of the pulmonary artery (yellow fluorescence). B indicatesbronchus; PA, pulmonary artery.
Mittal et al Regulation of NOX4 in the Pulmonary Vasculature 261
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

oxidase subunits in mouse lungs, NOX4 was the only subunitsignificantly upregulated in homogenized tissue over the time-course of exposure to chronic hypoxia (Figure 1b and 1c). Incontrast NOX2, NOXA1, and p67phox were not significantlyregulated and NOX1 as well as the other cytosolic NADPHoxidases exhibited an overall downregulation after 21 days ofchronic hypoxia (Figure 1b and 1c). Considering that NOX4 wasprominently upregulated and NOX2 was previously suggested
to play an important role in hypoxia-induced pulmonary hyper-tension,24 we next investigated the expression of NOX4 andNOX2 in microdissected small pulmonary arterial vessels(�100 �m diameter), the major site of pulmonary vascularremodeling in chronic hypoxia by real-time PCR (Figure 2athrough 2c). Comparing these vessels from animals exposed tonormoxic (21% O2) and chronic hypoxic (10% O2) conditionsfor up to 3, 7, and 21 days, it was observed that NOX4 mRNA
Figure 4. NOX4 immunostaining comparing mice maintained either under normoxic (21% O2) or hypoxic (10% O2) conditions. a, In nor-moxic mice, NOX4 immunoreactivity was not detected in the majority of vessels (arrows). b, Under hypoxia, the majority of vesselswere immunoreactive for NOX4 (arrows). Note the slightly autofluorescent bronchial epithelium (arrowhead). c and d, Labeling of NOX4and �-smooth muscle actin (SMA), confocal image. A subgroup of SMA-immunoreactive cells in the vessel wall was also NOX4 immu-noreactive (arrows), as evident for the merged images c and d (e). f, Percentage of NOX4 immunoreactive vessels in normoxic miceand mice exposed to chronic hypoxia (10% O2) for up to 21 days. The mean value for every animal is shown in the box plot. g, Meandiameter of NOX4 immunoreactive vessels during development of hypoxia-induced pulmonary hypertension. The mean value for everyanimal is shown in the box plot; n�number of animals examined per experimental condition, *P�0.05; boxes, percentiles 25 and 75,black bar, median; whiskers, percentiles 0 and 100; O�value is more than 1.5 lengths of a box away from the edge of a box.
262 Circulation Research August 3, 2007
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

expression was upregulated in the pulmonary arteries over thecourse of exposure to hypoxia, with the highest elevation after 3weeks (Figure 2a). In contrast to NOX4, no regulation of NOX2was observed (Figure 2b). Under normoxic conditions NOX2mRNA levels were not different from those of NOX4 (Figure2c). The hypoxic upregulation of NOX4 paralleled thedevelopment of pulmonary hypertension in mice induced bychronic hypoxia. The ratio of the right to the left ventricularmass was 0.27�0.01 in mice maintained under normoxicconditions, and increased to 0.28�0.03, 0.31�0.02, and0.37�0.01 (n�5 each) after 3, 7, and 21 days of hypoxia,respectively (Figure 2d). In situ hybridization demonstratedNOX4 mRNA expression in different cell types with prom-inent presence in the vessel media, as confirmed by itscolocalization with �-smooth muscle actin (Figure 3a through3f). Nonvascular area that stained positive for NOX4 mRNA(Figure 3) comprises bronchial smooth muscle cells and mayinclude alveolar type II cells. In this regard we detectedNOX4 transcripts in isolated type II cells from the mouse(supplemental Figure I). Our observations that NOX4 mRNAwas the predominant NOX mRNA present in the vesselmedia was also confirmed on the protein level (Figure 4).NOX4 immunoreactivity was observed in a subset of cells ofthe medial wall of the pulmonary artery, as well as in somesmaller pulmonary arteries (Figure 4a through 4e). Afterexposure to chronic hypoxia, the number of NOX4-positivevessels was significantly increased after 3 days of exposure tohypoxia (Figure 4f). The number of small NOX4 immunopo-sitive vessels was also significantly increased after 7 and 21days of hypoxia (Figure 4g), indicating that the newly-formedsmaller vessels were also NOX4-immunoreactive. At thesub-cellular level, NOX4 protein exhibited a predominantlyperinuclear localization in mouse PASMCs with increasedintensity after 48 hours of hypoxic incubation (Figure 5).
Histological staining of human lung sections from healthydonors and from patients with idiopathic pulmonary arterial
hypertension (IPAH) confirmed NOX4 expression in thevessel media of the pulmonary arteries (Figure 6a through6d). In contrast to NOX4 we found that NOX2 was primarilyexpressed in the endothelial layer of the human pulmonaryarteries (Figure 6e and 6f). Western blot analysis revealed asignificant (P�0.001) 2.5-fold higher NOX4 protein level inlungs from IPAH patients compared with healthy donor lungs(Figure 7a, full blot and specificity of the NOX4 antibody seesupplemental Figure IIa). In addition, NOX4 transcriptsquantified by real-time PCR were increased in human donorPASMCs from passage 3 exposed to hypoxia for 24 hour,compared with normoxic controls (Figure 7b). To confirm afunctional role for NOX4 in cell proliferation, we demon-strated that siRNA directed against human NOX4 signifi-cantly reduced the NOX4 mRNA level (Figure 8a andsuppressed the proliferation of human passage 3 PASMCs(Figure 8b) correlating with a decrease of reactive oxygenspecies (ROS) generation (Figure 8c). Reduced proliferationof human PASMCs after siNOX4 treatment was additionallyconfirmed by cell counting (supplemental Figure III). Aspreviously reported for systemic and PASMCs the NOX4levels decreased with higher passages (supplemental FigureIVa).20,31 However, siRNA against NOX4 decreased cell pro-liferation of both passage 3 and passage 5 cells with higherefficacy in passage 5 cells (see supplemental Figure IVb).
DiscussionNo extensive analysis of the expression of the NADPH-oxidase subunits, particularly nonphagocytic NADPH oxi-dase subunits, in the lung and their regulation in hypoxia hasbeen performed to date. Thus, in screening for the expressionof the NADPH oxidase subunits NOX1, NOX2, NOX4,p22phox, p40phox, p47phox, p67phox, NOXA1, and NOXO1 indifferent mouse organs, it was observed that all of thesesubunits could be detected by RT-PCR in lung tissue with asimilar signal intensity compared with the colon and heart
Figure 5. Cellular NOX4 localization inmouse PASMCs by immunofluorescence.The NOX4 immunostaining of isolatedmurine PASMCs revealed a perinuclearlocalization, and a slight increase of NOX4immunoreactivity when PASMCs wereincubated for 48 hours under hypoxicconditions, compared with cells incubatedunder normoxic conditions. MurinePASMCs stained with an anti-NOX4 anti-body (a) and with an additional nuclearmarker (b) (Hoechst). NOX4 immunoreac-tivity of murine PASMCs after 48 hoursunder normoxic (21% O2; c), comparedwith hypoxic (1% O2; d), conditions.
Mittal et al Regulation of NOX4 in the Pulmonary Vasculature 263
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

(Figure 1a). In 2 recent investigations, elevated expression ofNOX1 and NOX4 was demonstrated for the colon and kidneyby Northern blotting.4,32 In a previous study, we identifiedNOX1 in homogenized rabbit lungs and in PASMCs.33
Recently, NOX4 was suggested to be the predominant NOX2homolog in human airway PASMCs,19,20 and Liu and col-leagues provided evidence that the phagocytic NADPH oxi-dase subunit NOX2 plays an important role in the develop-ment of hypoxia-induced pulmonary hypertension.24
Investigations into the recently identified new isoforms ofphagocytic NADPH oxidase subunits in the lung is ofinterest, because NADPH oxidases have been proposed aspossible pulmonary oxygen sensors.19,22 Suliman and co-workers supported a possible role for NOX4 in the context ofoxygen sensing in the mouse kidney, demonstrating inducedexpression of the renal-specific NADPH oxidase (NOX4)under hypoxic conditions.34
With respect to the lung vasculature, oxygen sensing isimportant under circumstances of acute hypoxia (lastingseconds to minutes) as well as of chronic hypoxia (lastingdays to months and years). Acute alveolar hypoxia inducesconstriction of pulmonary arterial vessels, which is an essen-tial mechanism to adapt perfusion to ventilation, and thus tooptimize pulmonary gas exchange.22,35 Recently we havedemonstrated that a nonphagocytic NADPH oxidase mayplay an important role in the acute hypoxic response of thepulmonary arteries in the lung.23
In contrast, chronic alveolar hypoxia induces remodelingof the pulmonary vasculature, characterized by hypertrophyof the vessel media, and thus a narrowing of the vascularlumen. This leads to an increased pulmonary vascular resis-tance, pulmonary hypertension, and ultimately resulting inright heart failure. For both acute and chronic alveolarhypoxia, a possible role for reactive oxygen species has been
Figure 6. Localization of NOX4 and NOX2 in human donor and IPAH lungs. The NOX4 immunostaining of human donor (a, c) and IPAH(b, d) lung sections revealed a localization of NOX4 predominantly in the medial layer of pulmonary arteries. In contrast, NOX2 immuno-staining was localized mainly in the endothelium of human donor (e) and IPAH (f) lung sections.
264 Circulation Research August 3, 2007
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

widely discussed19,22,35,36 and the phagocytic NADPH oxi-dase NOX2 has been demonstrated in a knockout mousemodel to be of major importance for the development ofhypoxia-induced pulmonary hypertension. With respect to anadditional role for nonphagocytic NADPH oxidase subunitsin the pathophysiology of hypoxia-induced pulmonary hyper-tension, the present study focused on NOX4, because: (1)NOX4 is the only nonphagocytic NADPH oxidase subunitprominently expressed in pulmonary arteries, (2) NOX 4 wasthe only NADPH oxidase subunit upregulated in chronichypoxia in homogenized mouse lung tissue (Figure 1b and1c), (3) NOX4 acts as an oxygen sensor to regulate TASK-1activity in HEK 293 cells, and (4) it was recently suggestedthat this subunit may contribute to pathophysiologicalchanges in the systemic vasculature and in the pulmonaryarteries.1,10,20,37 Moreover, we compared the regulation ofNOX4 to that of NOX2 considering the recent findings by
Liu et al.24 As remodeling of small pulmonary arteries isthought to be the major cause of the increase in vascularresistance occurring during chronic hypoxia, we focused onthe hypoxia-dependent regulation of NOX4 and NOX2mRNA in these vessels of the murine pulmonary vasculature.Our analysis revealed that NOX4, in contrast to NOX2, iselevated in the pulmonary vasculature by chronic hypoxia:upregulation of NOX4 but not of NOX2 occurred in thepulmonary arteries within 21 days of exposure to hypoxia, asdemonstrated by quantitative PCR of microdissected vessels.Moreover, in situ hybridization revealed that NOX4 tran-scripts were localized to the pulmonary artery smooth musclelayer. The hypoxia-dependent increase in NOX4 expressionlevels in the pulmonary vasculature correlated well with thedevelopment of pulmonary hypertension,38 and was corrob-orated further at the protein level: (1) NOX4-immunoreactivity was detected in the pulmonary vasculature
β
Figure 7. Detection of NOX4 by Western blotin human donor and IPAH lungs and hypoxia-induced upregulation of NOX4 in isolatedPASMCs. a, Western blots of human IPAHlungs (n�4) compared with healthy donor lungs(n�6) revealed a 2.5-fold upregulation of NOX4expression in human IPAH lungs (specific bandat 64 kDa). The NOX4 was normalized to�-actin. The full blot as well as the specificity ofthe antibody is shown in supplemental FigureIIa. *Significant difference as compared withdonor lungs. b, Isolated human PASMCs weremaintained under hypoxic (1% O2) or normoxic(21% O2) conditions for 24 hours. The NOX4mRNA levels were quantified by real-time PCRand standardized to �2-microglobulin mRNAlevels. A significant increase in NOX4 mRNAwas observed after 24 hours of hypoxic vsnormoxic treatment (*). Data are derived fromduplicate measurement of n�3 independentcell preparations.
Figure 8. Suppression of human PASMC proliferation and reactive oxygen species generation by siRNA directed against NOX4. a,Quantification of NOX4 in siNOX4-transfected human PASMCs showing significant downregulation of NOX4 as compared with scram-bled control. b, The normoxic proliferation of PASMCs was investigated using 3H-thymidine. c, Reactive oxygen species quantificationby dihydroethidium fluorescence in scrambled and NOX4 siRNA transfected human PASMCs. *Significant differences between NOX4siRNA and scrambled siRNA experiments. Data are derived from duplicate cell isolations of n�5 independent lungs. cpm indicatescounts per minute.
Mittal et al Regulation of NOX4 in the Pulmonary Vasculature 265
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

by immunostaining, and (2) the percentage of NOX4 immu-noreactive vessels was strongly increased by chronic alveolarhypoxia with an increase in the number of NOX4-positivesmall vessels. The upregulation at the protein level (Figure4f) preceded the regulation on the mRNA level (Figure 2a).This suggests that NOX4 can be regulated on both the mRNAand the protein level. It was also evident from immunohisto-chemical data that the upregulation of NOX4 occurs in thevessel media, further supporting the idea that NOX4 contrib-utes to the pathophysiological process of hypoxia-inducedvascular remodeling in the lung, which is triggered byROS-dependent smooth muscle cell proliferation. In line withthis observation and the recent finding of Sturrock et al, thesilencing of NOX4 by siRNA reduced human PASMCproliferation as well as ROS generation.20 A possible role forNOX4 in the pathogenesis of pulmonary hypertension ingeneral was supported by the fact that NOX4 is upregulatedin the vessel media of lung sections from patients with IPAH,compared with healthy donor lungs. The perinuclear local-ization of NOX4 in the PASMCs supports the notion of thepresence of the protein in the endoplasmic reticulum (ER), asrecently demonstrated in microvascular endothelial cells byPetry and coworkers.39 The presence of NOX4 in the ERfurther suggests an important role of NOX4 in maintainingthe redox potential and Ca2�-homeostasis in PASMCs.40
The findings of Liu et al that NOX2 is essential fordevelopment of hypoxia-induced pulmonary hypertension,together with the fact that we as well as Liu et al were unableto detect regulation of NOX2 in pulmonary arteries byhypoxia, are suggestive that NOX2 and NOX4 play a differ-ential role in the development of hypoxia-induced pulmonaryhypertension. Hypothetically, endothelial ROS generation byNOX2 may stimulate NOX4 upregulation in the vesselmedia, which would be important for hypoxia-dependentPASMC proliferation. In line with this argumentation is thedetection primarily of NOX2 in pulmonary vascular endothe-lial cells in our study, as well as 2 recent reports demonstrat-ing a ROS-dependent upregulation of NOX4 in cardiaccells.41,42
The fact that NOX4 is upregulated in the vessel media inboth hypoxia-induced pulmonary hypertension and in humanIPAH may be explained by distinct or common regulators ofNOX4. With regard to the latter it has been shown thatTGF-� can upregulate NOX4 in human PASMCs,20 thathypoxia can increase TGF-� in PASMCs,43 and that interfer-ence with TGF-� blocks hypoxia-induced vascular remodel-ing.44 Interestingly, it has been shown that TGF-� can viceversa be regulated by ROS.45 Thus, hypoxia-induced andhuman IPAH may share some common pathophysiologicalmechanisms with regard to NOX4.
In conclusion, we demonstrate in the present study that allmajor subunits of the phagocytic as well as nonphagocyticNADPH-oxidase subunits are expressed in the lung. Further-more, NOX4 was found to be upregulated in the pulmonaryvasculature, both in chronic hypoxic pulmonary hypertensionas well as in human IPAH. The correlation of NOX4expression with the development of pulmonary hypertensionsuggests a contribution of NOX4 to the development of thisdisease. With respect to the upregulation of NOX4 in IPAH
patients, a functional interference with NOX4 may offer anew therapeutic approach for the treatment of this disease.
AcknowledgmentsThe authors thank Dr Rory Morty for linguistic editing of themanuscript and Karin Quanz, Carmen Homberger, IngridBreitenborn-Muller, Christiane Hild, and Marcel Zoremba for excel-lent technical assistance.
Sources of FundingThis work was supported by the Deutsche ForschungsgemeinschaftSFB 547, projects B7, C1, and C7, the European CommissionContract No LSHM-CT-2005�018725, PULMOTENSION, the Na-tional & Medical Research Council of Australia, and the NationalHeart Foundation (Australia).
DisclosuresNone.
References1. Cai H, Griendling KK, Harrison DG. The vascular NAD(P)H oxidases as
therapeutic targets in cardiovascular diseases. Trends PharmacolSci. 2003;24:471–478.
2. Babior BM, Lambeth JD, Nauseef W. The neutrophil NADPH oxidase.Arch Biochem Biophys. 2002;397:342–344.
3. Bokoch GM, Knaus UG. NADPH oxidases: not just for leukocytesanymore! Trends Biochem Sci. 2003;28:502–508.
4. Suh YA, Arnold RS, Lassegue B, Shi J, Xu X, Sorescu D, Chung AB,Griendling KK, Lambeth JD. Cell transformation by the superoxide-generating oxidase Mox1. Nature. 1999;401:79–82.
5. Cheng G, Cao Z, Xu X, van Meir EG, Lambeth JD. Homologs ofgp91phox: cloning and tissue expression of Nox3, Nox4, and Nox5.Gene. 2001;269:131–140.
6. Lambeth JD, Cheng G, Arnold RS, Edens WA. Novel homologs ofgp91phox. Trends Biochem Sci. 2000;25:459–461.
7. Geiszt M, Kopp JB, Varnai P, Leto TL. Identification of renox, anNAD(P)H oxidase in kidney. Proc Natl Acad Sci U S A. 2000;97:8010–8014.
8. Geiszt M. NADPH oxidases: new kids on the block. Cardiovasc Res.2006;71:289–299.
9. Griendling KK, Sorescu D, Ushio-Fukai M. NAD(P)H oxidase: role incardiovascular biology and disease. Circ Res. 2000;86:494–501.
10. Lassegue B, Clempus RE. Vascular NAD(P)H oxidases: specific features,expression, and regulation. Am J Physiol Regul Integr Comp Physiol.2003;285:R277–R297.
11. Banfi B, Clark RA, Steger K, Krause KH. Two novel proteins activatesuperoxide generation by the NADPH oxidase NOX1. J Biol Chem.2003;278:3510–3513.
12. Geiszt M, Lekstrom K, Witta J, Leto TL. Proteins homologous top47phox and p67phox support superoxide production by NAD(P)Hoxidase 1 in colon epithelial cells. J Biol Chem. 2003;278:20006–20012.
13. Uchizono Y, Takeya R, Iwase M, Sasaki N, Oku M, Imoto H, Iida M,Sumimoto H. Expression of isoforms of NADPH oxidase components inrat pancreatic islets. Life Sci. 2006;80:133–139.
14. Miller AA, Drummond GR, Schmidt HH, Sobey CG. NADPH oxidaseactivity and function are profoundly greater in cerebral versus systemicarteries. Circ Res. 2005;97:1055–1062.
15. Miller AA, Drummond GR, Mast AE, Schmidt HHHW, Sobey CG.Effect of gender on NADPH-oxidase activity, expression and function inthe cerebral circulation: role of estrogen. Stroke. In press.
16. Cai H, Harrison DG. Endothelial dysfunction in cardiovascular diseases:the role of oxidant stress. Circ Res. 2000;87:840–844.
17. Sorescu D, Weiss D, Lassegue B, Clempus RE, Szocs K, Sorescu GP,Valppu L, Quinn MT, Lambeth JD, Vega JD, Taylor WR, Griendling KK.Superoxide production and expression of nox family proteins in humanatherosclerosis. Circulation. 2002;105:1429–1435.
18. Brandes RP. Role of NADPH oxidases in the control of vascular geneexpression. Antioxid Redox Signal. 2003;5:803–811.
19. Hoidal JR, Brar SS, Sturrock AB, Sanders KA, Dinger B, Fidone S,Kennedy TP. The role of endogenous NADPH oxidases in airway andpulmonary vascular smooth muscle function. Antioxid Redox Signal.2003;5:751–758.
266 Circulation Research August 3, 2007
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

20. Sturrock A, Cahill B, Norman K, Huecksteadt TP, Hill K, Sanders K,Karwande SV, Stringham JC, Bull DA, Gleich M, Kennedy TP, HoidalJR. Transforming growth factor-beta1 induces Nox4 NAD(P)H oxidaseand reactive oxygen species-dependent proliferation in human pulmonaryartery smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. 2006;290:L661–L673.
21. Hohler B, Holzapfel B, Kummer W. NADPH oxidase subunits andsuperoxide production in porcine pulmonary artery endothelial cells.Histochem Cell Biol. 2000;114:29–37.
22. Weissmann N, Sommer N, Schermuly RT, Ghofrani HA, Seeger W,Grimminger F. Oxygen sensors in hypoxic pulmonary vasoconstriction.Cardiovasc Res. 2006;71:620–629.
23. Weissmann N, Zeller S, Schafer RU, Turowski C, Ay M, Quanz K,Ghofrani HA, Schermuly RT, Fink L, Seeger W, Grimminger F. Impactof mitochondria and NADPH oxidases on acute and sustained hypoxicpulmonary vasoconstriction. Am J Respir Cell Mol Biol. 2006;34:505–513.
24. Liu JQ, Zelko IN, Erbynn EM, Sham JS, Folz RJ. Hypoxic pulmonaryhypertension: role of superoxide and NADPH oxidase (gp91phox). Am JPhysiol Lung Cell Mol Physiol. 2006;290:L2–10.
25. Jeffery TK, Wanstall JC. Pulmonary vascular remodeling: a target fortherapeutic intervention in pulmonary hypertension. Pharmacol Ther.2001;92:1–20.
26. Schermuly RT, Dony E, Ghofrani HA, Pullamsetti S, Savai R, Roth M,Sydykov A, Lai YJ, Weissmann N, Seeger W, Grimminger F. Reversal ofexperimental pulmonary hypertension by PDGF inhibition. J Clin Invest.2005;115:2811–2821.
27. Fink L, Kohlhoff S, Stein MM, Hanze J, Weissmann N, Rose F,Akkayagil E, Manz D, Grimminger F, Seeger W, Bohle RM. cDNA arrayhybridization after laser-assisted microdissection from nonneoplastictissue. Am J Pathol. 2002;160:81–90.
28. Goyal P, Weissmann N, Rose F, Grimminger F, Schafers HJ, Seeger W,Hanze J. Identification of novel Nox4 splice variants with impact on ROSlevels in A549 cells. Biochem Biophys Res Commun. 2005;329:32–39.
29. Rose F, Grimminger F, Appel J, Heller M, Pies V, Weissmann N, Fink L,Schmidt S, Krick S, Camenisch G, Gassmann M, Seeger W, HanzeJ. Hypoxic pulmonary artery fibroblasts trigger proliferation of vascularsmooth muscle cells: role of hypoxia-inducible transcription factors.FASEB J. 2002;16:1660–1661.
30. Weissmann N, Dietrich A, Fuchs B, Kalwa H, Ay M, Dumitrascu R,Olschewski A, Storch U, Mederos YS, Ghofrani HA, Schermuly RT,Pinkenburg O, Seeger W, Grimminger F, Gudermann T. Classicaltransient receptor potential channel 6 (TRPC6) is essential for hypoxicpulmonary vasoconstriction and alveolar gas exchange. Proc Natl AcadSci U S A. 2006;103:19093–19098.
31. Clempus RE, Sorescu D, Dikalova AE, Pounkova L, Jo P, Sorescu GP,Schmidt HH, Lassegue B, Griendling KK. Nox4 is required for maintenance
of the differentiated vascular smooth muscle cell phenotype. ArteriosclerThromb Vasc Biol. 2007;27:42–48.
32. Shiose A, Kuroda J, Tsuruya K, Hirai M, Hirakata H, Naito S, Hattori M,Sakaki Y, Sumimoto H. A novel superoxide-producing NAD(P)Hoxidase in kidney. J Biol Chem. 2001;276:1417–1423.
33. Weissmann N, Tadic A, Hanze J, Rose F, Winterhalder S, Nollen M,Schermuly RT, Ghofrani HA, Seeger W, Grimminger F. Hypoxic vaso-constriction in intact lungs: a role for NADPH oxidase-derived H(2)O(2)?Am J Physiol Lung Cell Mol Physiol. 2000;279:L683–L690.
34. Suliman HB, Ali M, Piantadosi CA. Superoxide dismutase-3 promotesfull expression of the EPO response to hypoxia. Blood. 2004;104:43–50.
35. Stenmark KR, Fagan KA, Frid MG. Hypoxia-induced pulmonaryvascular remodeling: cellular and molecular mechanisms. Circ Res. 2006;99:675–691.
36. Sylvester JT. Hypoxic pulmonary vasoconstriction: a radical view. CircRes. 2001;22;88:1228–1230.
37. Lee YM, Kim BJ, Chun YS, So I, Choi H, Kim MS, Park JW. NOX4 as anoxygen sensor to regulate TASK-1 activity. Cell Signal. 2006;18:499–507.
38. Weissmann N, Manz D, Buchspies D, Keller S, Mehling T, VoswinckelR, Quanz K, Ghofrani HA, Schermuly RT, Fink L, Seeger W, GassmannM, Grimminger F. Congenital erythropoietin over-expression causes“anti-pulmonary hypertensive” structural and functional changes in mice,both in normoxia and hypoxia. Thromb Haemost. 2005;94:630–638.
39. Petry A, Djordjevic T, Weitnauer M, Kietzmann T, Hess J, Gorlach A.NOX2 and NOX4 mediate proliferative response in endothelial cells.Antioxid Redox Signal. 2006;8:1473–1484.
40. Gorlach A, Klappa P, Kietzmann T. The endoplasmic reticulum: folding,calcium homeostasis, signaling, and redox control. Antioxid RedoxSignal. 2006;8:1391–1418.
41. Djordjevic T, Pogrebniak A, BelAiba RS, Bonello S, Wotzlaw C, AckerH, Hess J, Gorlach A. The expression of the NADPH oxidase subunitp22phox is regulated by a redox-sensitive pathway in endothelial cells.Free Radic Biol Med. 2005;38:616–630.
42. Buggisch M, Ateghang B, Ruhe C, Strobel C, Lange S, Wartenberg M,Sauer H. Stimulation of ES-cell-derived cardiomyogenesis and neonatalcardiac cell proliferation by reactive oxygen species and NADPHoxidase. J Cell Sci. 2007;120:885–894.
43. Jiang Y, Dai A, Li Q, Hu R. Hypoxia induces transforming growthfactor-beta1 gene expression in the pulmonary artery of rats via hypox-ia-inducible factor-1alpha. Acta Biochim Biophys Sin (Shanghai). 2007;39:73–80.
44. Chen YF, Feng JA, Li P, Xing D, Zhang Y, Serra R, Ambalavanan N,Majid-Hassan E, Oparil S. Dominant negative mutation of the TGF-betareceptor blocks hypoxia-induced pulmonary vascular remodeling. J ApplPhysiol. 2006;100:564–571.
45. Jackson IL, Chen L, Batinic-Haberle I, Vujaskovic Z. Superoxide dis-mutase mimetic reduces hypoxia-induced O2*-, TGF-beta, and VEGFproduction by macrophages. Free Radic Res. 2007;41:8–14.
Mittal et al Regulation of NOX4 in the Pulmonary Vasculature 267
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/1
EXPANDED MATERIALS AND METHODS
Mouse lung preparation for laser assisted microdissection and right heart hypertrophy assessment.
After 3, 7 and 21 days of hypoxia, the animals were euthanized by intraperitoneal injection of a lethal
ketamine and xylazine dose, and the lungs were flushed with Krebs Henseleit buffer (125.0 mM NaCl,
4.3 mM KCl, 1.1 mM KH2PO4, 2.4 mM CaCl2, 1.3 mM MgCl2 and 13.32 mM glucose) through a
catheter in the pulmonary artery at a pressure of 20 cm H2O at room temperature. The buffer was pre-
equilibrated with a gas mixture of 3%O2, 5.3%CO2, balanced N2. NaHCO3 was adjusted to result in a
pH of 7.4. During perfusion of the lungs, the buffer was allowed to drain freely from a catheter in the
left ventricle. Once the effluent was clear of blood, lungs were dissected from the thoracic cavity and
immediatley frozen in liquid nitrogen for mRNA analysis.
For immunohistochemistry, in situ hybridization and laser assisted microdissection, 800 µl pre-
warmed TissueTek® (Sakura Finetek, Zoeterwoude, The Netherlands) was instilled into the airways
via a tracheal cannula. After ligation of the trachea, the lungs were excised and immediately snap
frozen in melting isopentane. Preparation of the hypoxic animals was performed continuously in a
hypoxic environment (10% O2). Lungs from normoxic animals (control) were prepared accordingly
under normoxic conditions. The right ventricular wall (RV) was separated from the left ventricle plus
septum (LV+S) to calculate the ratio of RV/(LV+S) of the dehydrated heart tissue. For analysis of
colon, heart, and pulmonary arteries, these tissues and organs were removed immediately after the
lung had been flushed.
RT-PCR
For reverse transcription (RT) of extracted RNA, 1 µg of total RNA was denatured at 65 °C for 5 min.
After cooling on ice, the following components were added to the samples: 5 µl of 5× RT buffer, 2 µl
of 10 mM deoxynucleotide mixture, 1 µl of random-hexamer primer, 0.5 µl of 0.1 M dithiothreitol,
1 µl Moloney murine leukemia virus reverse transcriptase (MMLV) (Gibco-BRL, Karlsruhe,
Germany) and volume for adjustment to 20 µl. After 10 min of incubation at room temperature, and

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/2
60 min at 39 °C, reverse transcriptase was inactivated by heating the mixture to 95 °C. For PCR
reactions, 4 µl of 10 µM forward primer, 4 µl of 10 µM reverse primer, 10 µl of 10× PCR buffer, 61 µl
of water and 0.5 µl of hot-start-Taq-polymerase (Qiagen, Hilden, Germany) were added to 20 µl of the
cDNA. The thermal cycler profile consisted of an initial incubation at 95°C for 15 min, followed by
40 cycles of 95 °C 30 s, 62 °C, 30 s and 72 °C, 60 s, and a final extension at 72 °C for 10 min. For the
sequences of primers used in RT PCR see Online Table I.
Real-time PCR
All primers used in the real time PCR experiments were intron-spanning. The Ct values of NOX4
were normalized to the endogenous control B2M (β2-microglobulin) in both mouse and human
samples (1). For cDNA synthesis reagents and incubation time were applied as described before (2).
The fold change 2∆∆Ct was calculated as described previously (3).For the sequences primers used in real
time PCR please see Online Table II.
Riboprobes for in situ hybridization
Single-stranded digoxigenin (DIG)-labeled riboprobes for non-isotopic in situ hybridization were
generated by the in vitro transcription method. The template for the generation of single-stranded
RNA probes was amplified by nested PCR. Briefly, 1 µg of the purified PCR-amplified template
harbouring T3 and T7 RNA polymerase promoter sequences was mixed with 2 µl digoxigenin-11-
uridine triphosphate (Roche, Mannheim, Germany), 4 µl of 5× transcription buffer (Promega,
Mannheim,Germany), 1 µl of RNasin (Peqlab, Erlangen, Germany) and 2 µl of T3 or T7 Phage
polymerase (Promega, Mannheim, Germany) in a total reaction volume of 20 µl. The reaction mixture
was incubated at 37 °C for 2 h. The RNA probes were purified with a Qiagen PCR purification kit
(Qiagen, Hilden, Germany).

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/3
Combined approach of non isotopic in situ hybridization and immunofluorescence on mouse lung
sections
The non-isotopic in situ hybridization (NISH) was performed on 8 µm thick TissueTek®-embedded
mouse lung cryostat sections. The sections were heated at 55 °C for 15 min followed by transferring
the sections in to 2× SSC buffer for 30 min at 70 °C, diethylpyrocarbonate-treated water for 1 min,
Proteinase K (5 µg/ml) for 10 min at room temperature, 0.2% (m/v) glycine in PBS solution (for the
inactivation of Proteinase K) for 30 s, PBS for 30 s, freshly prepared cold 4% (m/v) paraformaldehyde
for 20 min, PBS for 5 min, 0.1 M acetylated triethanolamine (0.5 ml of acetic anhydride/200 ml of
triethanolamine) on a shaking platform for 10 min, PBS for 3 min, followed by dehydration of slides
by passing through 70% (v/v), 80% (v/v) and 90% (v) ethanol (each for 2 min). The slides were then
pre-hybridized with 2× Prehyb solution (1 M NaCl, 0.02 M Tris (pH 7.5), 2× Denhardt’s reagent,
2 mM EDTA, 10 mg/ml salmon sperm DNA, 0.2 mg/ml yeast tRNA) at 55 °C for 2-3 h in a
humidified chamber, followed by hybridization with a denatured antisense NOX4 probe in
2× hybridization solution (1 M NaCl, 0.02 M Tris (pH7.5), 2× Denhardt’s reagent, 2 mM EDTA, 2 g
dextran sulphate, 0.2 mg/ml yeast tRNA) at 55 °C for overnight. The following day, slides were
washed from low to very highly stringent conditions as follows: on shaking platform 2× SSC for 1 h at
room temperature, 0.1× SSC at 60 °C and finally to pre-heated 0.1x SSC (at 60 °C), followed by
incubation at room temperature. The sections were treated with blocking buffer (2% Blocking reagent
(Roche, Mannheim, Germany), 0.1% (m/v) BSA, 0.1 M Tris (pH 7.5), 5 M NaCl) for 30 min at room
temperature, followed by incubation with a peroxidase-labeled anti-DIG antibody (Roche, Mannheim,
Germany) in 1:20 dilution for 2 h at room temperature. After antibody incubation, sections were
washed in TBT buffer (50 mM, 1 M Tris-HCl (pH 7.5), 150 mM NaCl and 0.1% Triton X-100; 3 ×
15 min). The fluorescent substrate Alexa fluor 488 tyramide (Molecular Probes, Invitrogen, Karlsruhe,
Germany) at a dilution of 1:60 in amplification buffer was applied to the sections for 2 h.
Subsequently, sections were washed (3 × 20 min) in PBT buffer (PBS, 0.1% (v/v) Tween 20) and
incubated with a mouse monoclonal Cy3-labeled α-smooth muscle actin antibody (Sigma, Hamburg,
Germany) at 1:500 in PBS, for 1 h. The sections were washed (3 × 3 min) in PBS and subsequently

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/4
incubated with Hoechst-33258 (1:10.000 in PBS; Invitrogen, Karlsruhe, Germany) for 10 min, washed
and mounted in carbonate-buffered glycerol (pH-8.6).
Type II cell isolation
Mouse alveolar type II cells were isolated as described previously (4).
Immunohistochemistry for mouse lung sections
Cryosections (10 µm) were fixed in ice-cold acetone for 10 min and unspecific binding of primary
antibodies was blocked by incubation with 50% (v/v) heat-inactivated normal swine serum in
phosphate buffered saline (9.1 mM dibasic sodium phosphate, 1.7 mM monobasic sodium phosphate
and 150 mM NaCl, pH 7.4 (PBS)) with double the salt concentration (PBS+S) for 1 h. Incubation was
performed overnight with an affinity-purified rabbit polyclonal anti-NOX4 antibody diluted 1:200,
together with a mouse monoclonal FITC-conjugated α-smooth muscle actin antibody (clone 1A4,
1:500; Sigma, Deisenhofen, Germany) in PBS+S (5). After washing in PBS, the sections were
incubated with a Cy3-conjugated donkey anti-rabbit antibody (Chemicon, Hofheim, Germany) diluted
1:2000 in PBS+S for 1 h, and after a final wash step, the sections were mounted with carbonate-
buffered glycerol, pH 8.6. To test specificity, the NOX4 antibody was pre-incubated for 1 h with the
corresponding peptide antigen (20 µg/ml). All labeling reported was specific as judged from its
absence in sections incubated with pre-adsorbed antibody. Immunoreactivity was evaluated using an
Axioplan 2 imaging epifluorescence microscope (Zeiss, Göttingen, Germany) equipped with a
Axiocam digital camera, Axiovision software (Zeiss, Göttingen, Germany), and with appropriate filter
sets for Cy3 and FITC or with a TCS-SP2 AOBS confocal laser scanning microscope (Leica,
Heidelberg, Germany).
Quantitative analysis of mouse lung sections
Examination of sections was performed blinded. For every animal, 50 α-smooth muscle actin-
immunoreactive vessels were evaluated for anti-NOX4-immunoreactive cells in the vessel wall, and

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/5
the vessel diameter was measured using the Axiovision software. To restrict analysis to smaller blood
vessels, vessels with a diameter larger than 60 µm were not examined.
Western blot of NOX4 in frozen human lung tissue
Protein extracts were prepared from frozen lung tissues from healthy human donor lungs and lungs
from patients with IPAH in RIPA buffer containing 1 mM sodium vanadate, Protease-Inhibitor Mix
complete (Roche, Mannheim, Germany) and 0.1 mM PMSF. Equivalent amounts of protein were
resolved on 10% SDS polyacrylamide gels. Proteins were transferred to polyvinylidene fluoride
(PVDF) membranes (Pall Corporation, Dreieich, Germany) by semi-dry electro-blotting. Nonspecific
antibody binding was blocked by incubation in 6% (m/v) non-fat dry milk powder in T-TBS (20 mM
Tris-Cl, pH 7.5, 150 mM NaCl, 0.1% (v/v) Tween 20) at room temperature for 1 h. Incubation with a
1:5000 diluted anti-NOX4 primary antibody was performed at 4 °C overnight. After washing the
membranes in T-TBS buffer, specific immunoreactive signals were detected by enhanced
chemiluminescence (ECL, Amersham, Freiburg, Germany) using a secondary antibody coupled to
horseradish-peroxidase.
RNA interference and proliferation assay
Human pulmonary arterial smooth muscle cells (10,000 cells per well) from passage 3 were cultured
in 48-well tissue culture plates, and used for the RNA interference and proliferation assays.
Transfection of NOX4 siRNA was performed in low-serum and antibiotic-free medium (1% (v/v) FCS
in DMEM). The medium was changed at least 4 h before transfection. Approximately 100 nM of
NOX4 siRNA (5’-GGUACAGCUGGAUGUUGAC-3’) (Eurogentec, Seraing, Belgium) was
transfected using 1 µl X-tremeGENE siRNA Transfection Reagent (Roche, Mannheim, Germany) per
cm2 of the well. Both siRNA and transfection reagent were diluted in OPTI-MEM medium (Gibco,
Karlsruhe, Germany). For controls, a FITC-labelled, scrambled siRNA (Invitrogen, Karlsruhe,
Germany) was employed. After five hours of transfection, the medium was changed to low-serum
medium containing antibiotics (1% (v/v) FCS, 1% (m/v) penicillin and streptomycin in DMEM and
incubated overnight for cellular synchronisation. The following day, the cells were stimulated with

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/6
smooth muscle cell medium (Promocell, Heidelberg, Germany) supplemented with medium
containing 5% (v/v) FCS and antibiotics for 20 h, followed by 3H-thymidine (Amersham, Munich,
Germany) for 4 h. Subsequently, cells were washed 3× with ice-cold PBS and lysed with NaOH on a
shaker for 4 h. The radioactivity in the lysate was measured by liquid scintillation counting (Rotiszint®
eco plus, Roth, Germany) with a Packard liquid scintillation counter. For determination of cell
numbers cells/ml were counted after synchronization for 72 h.
ROS measurement and quantification
ROS in human PASMC were measured using superoxide sensitive dye DHE (Dihydroethidium,
Invitrogen). Briefly, the cells were grown on chamber slides, transfected with scrambled and NOX4
siRNA and incubated in normoxic and hypoxic chambers for 24 h. After 24 h the cells were incubated
with 5 µM of DHE for 15 min in normoxic and hypoxic chamber. Subsequently the cells were washed
in PBS, fixed in acetone and methanol mixture (1:1) for 10 min and stained with nuclear stain
Hoechst-33258. The cells were visualized under a fluorescent microscope (excitation: 514 nm;
emission: 560 nm). A total of eight images was captured from each group in a blinded fashion and
were quantified using Image J software (U.S. National Institutes of Health, Bethesda, MD).
Online Table I: Nucleotide sequences of primers used for Revese Transcriptase-Polymerase Chain Reaction (RT
PCR)
Species Primer Orientation Sequence Accession Number
Mice NOX1 Sense 5´-ACAGAGGAGAGCTTGGGTGA-3´ NM_172203
Antisense 5´-CCCAACCAGTACAGCCACTT-3´
NOX2 Sense 5´-GCTTGTGGCTGTGATAAGCA-3´ NM_007807
Antisense 5´-CCACACAGGAAAACGCCTAT-3´
NOX4 Sense 5´-GCATCTGCATCTGTCCTGAA-3´ NM_015760
Antisense 5´-ACCACCTGAAACATGCAACA-3´

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/7
p22phox Sense 5´-TGGACGTTTCACACAGTGGT-3´ NM_007806
Antisense 5´-CTCCTCTTCACCCTCACTCG-3´
p47phox Sense 5´-GCCCAAAGATGGCAAGAATA-3´ NM_010876
Antisense 5´-ATGACCTCAATGGCTTCACC-3´
p67phox Sense 5´-CTTCAACATAGGCTGCGTGA-3´ NM_010877
Antisense 5´-CTTCATGTTGGTTGCCAATG-3´
p40phox Sense 5´-AAGGGTGTGTCTCCACAAGG-3´ NM_008677
Antisense 5´-TCCCACATCCTCATCTGACA-3´
NOXO1 Sense 5´-TTCACGGCTTCTTTGTACCC-3´ NM_027988
Antisense 5´-TTATACCTGCACAGCCACCA-3´
NOXA1 Sense 5´-CATTCCTGATGACCACAACG-3´ NM_172204
Antisense 5´-AGTCCAAATCCTCCGGTCTT-3´
Online Table II: Nucleotide sequences of primers used for quantitative real time PCR
Species Primer Orientation Sequence Accession Number
Mice NOX1 Sense 5´-TGGCTAAATCCCATCCAGTC-3´ NM_172203
Antisense 5´-CCCAAGCTCTCCTCTGTTTG-3´
NOX2 Sense 5´-TCGCTGGAAACCCTCCTATG-3´ NM_007807
Antisense 5´-GGATACCTTGGGGCACTTGA-3´

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/8
NOX4 Sense 5´-ACTTTTCATTGGGCGTCCTC-3´ NM_015760
Antisense 5´-AGAACTGGGTCCACAGCAGA-3´
p22phox Sense 5´-CAGATCGAGTGGGCCATGT-3´ NM_007806
Antisense 5´-AGCACACCTGCAGCGATAGA-3´
p47phox Sense 5´-GTCCCTGCATCCTATCTGGA-3´ NM_010876
Antisense 5´-TATCTCCTCCCCAGCCTTCT-3´
p67phox Sense 5´-CAGACCCAAAACCCCAGAAA-3´ NM_010877
Antisense 5´-AGGGTGAATCCGAAGCTCAA-3´
p40phox Sense 5´-TTTGAGCAGCTTCCAGACGA-3´ NM_008677
Antisense 5´-GGTGAAAGGGCTGTTCTTGC-3´
NOXO1 Sense 5´-TTCCTGATGCTCCATTGCTG-3´ NM_027988
Antisense 5´-GGTTGGGATAAGGGCTCCTC-3´
NOXA1 Sense 5´-AGCTGCAGAGGTTCCAGGAG NM_172204
Antisense 5´-GATGTCTTGAGCCCCCTCTG
B2M Sense 5´-AGCCCAAGACCGTCTACTGG-3´ NM_009735
Antisense 5´-TTCTTTCTGCGTGCATAAATTG-3´
Human NOX4 Sense 5´-GGTTAAACACCTCTGCCTGTTC-3´ NM_016931
Antisense 5´-CTTGGAACCTTCTGTGATCCTC-3´
B2M Sense 5´-CTGTGCTCGCGCTACTCTCT-3´ NM_004048
Antisense 5´-CTTCAATGTCGGATGGATGAA-3´

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/9
SUPPLEMENTARY FIGURES
Supplementary figure 1
RT-PCR of isolated alveolar type II cells.
RT-PCR revealed the presence of NOX4 mRNA transcripts in mouse alveolar type II cells.
Supplementary figure 2
Specificity of the NOX4 antibody.
a) Full Western-blot for NOX4 in human donor and IPAH lungs
Western-blots of human IPAH lungs (n=4) compared to healthy donor lungs (n=6) revealed a 2.5-fold
up-regulation of NOX4 expression in human IPAH lungs (specific band at 64 kDa). NOX4 was
normalized to β-actin.
b) Down-regulation of NOX4 protein by siRNA directed against NOX4 as recognized by the NOX4
antibody. PASMC were transfected with siRNA directed against NOX4 or with scrambled siRNA
65 h prior to western-blotting. β-actin was used as loading control.
Supplementary figure 3
Suppression of human pulmonary arterial smooth muscle cell proliferation by siRNA directed
against NOX4.
Passage 3 isolated human human pulmonary arterial smooth muscle cells (PASMC) were either
transfected with 100 nM NOX4 siRNA or scrambled siRNA as control. The normoxic proliferation of
PASMC was investigated by changes in cell numbers. Data are derived from duplicate cell isolations
from n=3 independent lungs.
Supplementary figure 4
a) Comparison of NOX4 mRNA transcript levels between different passages in human PASMC.
NOX4 mRNA transcript levels were quantified by real-time PCR for cell passage 3, 4 and 5.Data are
from duplicate measurements of at least 3 different cell isolations. (∆Ct= CtB2M- CtNOX4).

Online Supplement Mittal et al. Hypoxia-dependent regulation of non-phagocytic NADPH…
R1/10
b) Cell proliferation and its suppression by siRNA directed against NOX4 in human PASMC from
passage 5. Isolated human human pulmonary arterial smooth muscle cells (PASMC) were either
transfected with 100 nM NOX4 siRNA or scrambled siRNA as control. Data are from n=3-5 cell
isolations.
REFERENCES
(1) Ord JJ, Streeter EH, Roberts IS, Cranston D, Harris AL. Comparison of hypoxia
transcriptome in vitro with in vivo gene expression in human bladder cancer. Br J
Cancer. 2005;93:346-54.
(2) Fink L, Seeger W, Ermert L, Hanze J, Stahl U, Grimminger F, Kummer W, Bohle RM.
Real-time quantitative RT-PCR after laser-assisted cell picking. Nat Med. 1998;4:1329-
33.
(3) Fink L, Kohlhoff S, Stein MM, Hanze J, Weissmann N, Rose F, Akkayagil E, Manz D,
Grimminger F, Seeger W, Bohle RM. cDNA array hybridization after laser-assisted
microdissection from nonneoplastic tissue. Am J Pathol. 2002;160:81-90.
(4) Herold S, von WW, Steinmueller M, Pleschka S, Kuziel WA, Mack M, Srivastava M,
Seeger W, Maus UA, Lohmeyer J. Alveolar epithelial cells direct monocyte
transepithelial migration upon influenza virus infection: impact of chemokines and
adhesion molecules. J Immunol. 2006;177:1817-24.
(5) Wingler K, Wunsch S, Kreutz R, Rothermund L, Paul M, Schmidt HH. Upregulation of
the vascular NAD(P)H-oxidase isoforms Nox1 and Nox4 by the renin-angiotensin
system in vitro and in vivo. Free Radic Biol Med. 2001;31:1456-64.

NOX4
Supplementary Figure 1

donor IPAH
NOX4
Supplementary Figure 2a

ß-actin
scrambledsiRNA
PASMC normoxia
NOX4
NOX4siRNA
NOX4siRNA
un-transfected
Supplementary Figure 2b

0
20
40
60
80
100
120
scrambledsiRNA
NOX4 siRNA
Supplementary Figure 3
Cel
lnum
bers
afte
rpro
lifer
atio
n(%
)

Supplementary Figure 4
a)
0
1
2
3
4
5
Passage 3 Passage 4 Passage 5
ΔC
t
0
1000
2000
3000
4000
5000
6000
7000
Thym
idin
ein
corp
orat
ion
[cpm
]
b)
*
scrambledsiRNA
NOX4 siRNA
Passage 5

WeissmannHänze, Werner Seeger, Friedrich Grimminger, Harald H.H.W. Schmidt and Norbert Kwapiszewska, Wolfgang Kummer, Walter Klepetko, Mir Ali Reza Hoda, Ludger Fink, Jörg
Anne-Christin Selbitz, Ralph Theo Schermuly, Hossein Ardeschir Ghofrani, Grazyna Manish Mittal, Markus Roth, Peter König, Simone Hofmann, Eva Dony, Parag Goyal,
Pulmonary VasculatureHypoxia-Dependent Regulation of Nonphagocytic NADPH Oxidase Subunit NOX4 in the
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2007 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/CIRCRESAHA.107.1480152007;101:258-267; originally published online June 21, 2007;Circ Res.
http://circres.ahajournals.org/content/101/3/258World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2007/06/21/CIRCRESAHA.107.148015.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on September 10, 2015http://circres.ahajournals.org/Downloaded from
Related Documents