The FASEB Journal • Research Communication Defects in subventricular zone pigmented epithelium-derived factor niche signaling in the senescence-accelerated mouse prone-8 Paola Castro-Garcia,* Mar´ ıa D´ ıaz-Moreno, † Carmen Gil-Gas,* Francisco J. Fern´ andez-G´ omez, ‡ Paloma Honrubia-G ´ omez,* Carmen Bel´ en ´ Alvarez-Sim ´ on,* Francisco S´ anchez-S´ anchez, §,{ Juan Carlos Castillo Cano, k Francisco Almeida, k Vicente Blanco, k Joaqu´ ın Jord´ an, ‡,{ Helena Mira, † and Carmen Ram´ ırez-Castillejo* ,1,2 *Laboratorio de C´ elulas Madre, Centro Regional de Investigaciones Biom´ edicas, Universidad de Castilla–La Mancha, Albacete, Spain; † Unidad Funcional de Investigaci ´ on en Enfermedades Cr´ onicas, Instituto de Salud Carlos III, Madrid, Spain; ‡ Grupo de Neurofarmacolog´ ıa, Departamento de Ciencias M´ edicas, § Area de Gen´ etica, Facultad de Medicina de Albacete, and { Instituto de Investigaci ´ on en Discapacidades Neurol ´ ogicas, Universidad de Castilla–La Mancha, Albacete, Spain; and k Departamento Estad´ ıstica, I. O. y Computaci´ on, Universidad de La Laguna, La Laguna, Canarias, Spain ABSTRACT We studied potential changes in the sub- ventricular zone (SVZ) stem cell niche of the senescence- accelerated mouse prone-8 (SAM-P8) aging model. Bromodeoxyuridine (BrdU) assays with longtime survival revealed a lower number of label-retaining stem cells in the SAM-P8 SVZ compared with the SAM-Resistant 1 (SAM- R1) control strain. We also found that in SAM-P8 niche signaling is attenuated and the stem cell pool is less re- sponsive to the self-renewal niche factor pigmented epithelium-derived factor (PEDF). Protein analysis dem- onstrated stable amounts of the PEDF ligand in the SAM- P8 SVZ niche; however, SAM-P8 stem cells present a significant expression decrease of patatin-like phospho- lipase domain containing 2, a receptor for PEDF (PNPLA2- PEDF) receptor, but not of laminin receptor (LR), a receptor for PEDF (LR-PEDF) receptor. We observed changes in self-renewal related genes (hairy and enhancer of split 1 (Hes1), hairy and enhancer of split 1 (Hes5), Sox2] and report that although these genes are down-regulated in SAM-P8, differentiation genes (Pax6) are up-regulated and neurogenesis is increased. Finally, sheltering mam- malian telomere complexes might be also involved given a down-regulation of telomeric repeat binding factor 1 (Terf1) expression was observed in SAM-P8 at young age periods. Differences between these 2 models, SAM-P8 and SAM-R1 controls, have been previously detected at more advanced ages. We now describe alterations in the PEDF signaling pathway and stem cell self-renewal at a very young age, which could be involved in the premature senes- cence observed in the SAM-P8 model. —Castro-Garcia, P., D´ ıaz-Moreno, M., Gil-Gas, C., Fern´ andez-G ´ omez, F. J., Honrubia-G ´ omez, P., ´ Alvarez-Sim ´ on, C. B., S´ anchez- S´ anchez, F., Cano. J. C. C., Almeida, F., Blanco. V., Jord´ an, J. Mira, H., Ram´ ırez-Castillejo, C. Defects in sub- ventricular zone pigmented epithelium-derived factor niche signaling in the senescence-accelerated mouse prone-8. FASEB J. 29, 1480–1492 (2015). www.fasebj.org Key Words: self-renewal • neural stem cell • PEDF • accelerated senescence • long-term LRC SAM-P8 PROVIDES A GOOD model for age-related pathologies (1, 2). SAM-P8 presents an early cognitive impairment, which affects memory and learning skills (3, 4), causes modifications in adenosine receptors (5), and an increase in the level of butyrylcholinesterase activity (6, 7). Recent data describe alterations in stem cell proliferation in the forebrain of SAM-P8 preceding age-related brain symp- toms (1, 2). SAM-P8 presents an early cognitive impair- ment, which affects memory and learning skills (3, 4), and causes modifications in adenosine receptors (5) and an increase in the level of butyrylcholinesterase activity (6, 7). Recent data describe alterations in stem cell proliferation in the forebrain of SAMP8 preceding age-related brain symptoms (8, 9). However, to date it remains unknown whether loss of the uncommitted and multipotency state of the neural stem cell (NSC) population in the adult subventricular zone (SVZ) are involved in the senescence- accelerating process. Abbreviations: BrdU, bromodeoxyuridine; BSA, bovine se- rum albumin; DCX, doublecortin; DIV, days in vitro, or days in culture; EBSS, Earle’s balanced salt solution; FBS, fetal bovine serum; FGF-2, Fibroblast growth factor 2; GFAP, glial fibrillary acidic protein; Hes1, hairy and enhancer of split 1; Hes5, hairy and enhancer of split 5; LR-PEDF, laminin receptor (LR), a receptor for PB, phosphate buffer; LRC, label-retaining cell; (continued on next page) 1 Current affiliation: Unidad Funcional de Investigaci ´ on de Enfermedades Cr ´ onicas (Instituto de Salud Carlos III)/Hos- pital 12 de Octubre, Madrid, Spain. 2 Correspondence: 12 October Hospital/UFIEC, ISCIII, Carretera Majadahonda-Pozuelo Km 2, 28220 Madrid, Spain. E-mail: [email protected] doi: 10.1096/fj.13-244442 This article includes supplemental data. Please visit http:// www.fasebj.org to obtain this information. 1480 0892-6638/15/0029-1480 © FASEB

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The FASEB Journal • Research Communication
Defects in subventricular zone pigmentedepithelium-derived factor niche signaling in thesenescence-accelerated mouse prone-8
Paola Castro-Garcia,* Marıa Dıaz-Moreno,† Carmen Gil-Gas,* Francisco J. Fernandez-Gomez,‡
Paloma Honrubia-Gomez,* Carmen Belen Alvarez-Simon,* Francisco Sanchez-Sanchez,§,{
Juan Carlos Castillo Cano,k Francisco Almeida,k Vicente Blanco,k Joaquın Jordan,‡,{
Helena Mira,† and Carmen Ramırez-Castillejo*,1,2
*Laboratorio de Celulas Madre, Centro Regional de Investigaciones Biomedicas, Universidad deCastilla–La Mancha, Albacete, Spain; †Unidad Funcional de Investigacion en Enfermedades Cronicas,Instituto de Salud Carlos III, Madrid, Spain; ‡Grupo de Neurofarmacologıa, Departamento de CienciasMedicas, §Area de Genetica, Facultad de Medicina de Albacete, and {Instituto de Investigacion enDiscapacidades Neurologicas, Universidad de Castilla–La Mancha, Albacete, Spain; and kDepartamentoEstadıstica, I. O. y Computacion, Universidad de La Laguna, La Laguna, Canarias, Spain
ABSTRACT We studied potential changes in the sub-ventricular zone (SVZ) stem cell niche of the senescence-accelerated mouse prone-8 (SAM-P8) aging model.Bromodeoxyuridine (BrdU) assays with longtime survivalrevealed a lowernumberof label-retaining stemcells in theSAM-P8 SVZ compared with the SAM-Resistant 1 (SAM-R1) control strain. We also found that in SAM-P8 nichesignaling is attenuated and the stem cell pool is less re-sponsive to the self-renewal niche factor pigmentedepithelium-derived factor (PEDF). Protein analysis dem-onstrated stable amounts of the PEDF ligand in the SAM-P8 SVZ niche; however, SAM-P8 stem cells presenta significant expression decrease of patatin-like phospho-lipase domain containing 2, a receptor for PEDF (PNPLA2-PEDF) receptor, but not of laminin receptor (LR), areceptor for PEDF (LR-PEDF) receptor. We observedchanges in self-renewal related genes (hairy and enhancerof split 1 (Hes1), hairy and enhancer of split 1 (Hes5), Sox2]and report that although these genes are down-regulatedin SAM-P8, differentiation genes (Pax6) are up-regulatedand neurogenesis is increased. Finally, sheltering mam-malian telomere complexes might be also involved givena down-regulation of telomeric repeat binding factor 1(Terf1) expression was observed in SAM-P8 at young ageperiods.Differencesbetween these2models, SAM-P8andSAM-R1 controls, have been previously detected at moreadvanced ages. We now describe alterations in the PEDFsignaling pathway and stem cell self-renewal at a veryyoung age, which could be involved in the premature senes-cence observed in the SAM-P8 model.—Castro-Garcia, P.,
Dıaz-Moreno, M., Gil-Gas, C., Fernandez-Gomez, F. J.,Honrubia-Gomez, P., Alvarez-Simon, C. B., Sanchez-Sanchez, F., Cano. J. C. C., Almeida, F., Blanco. V.,Jordan, J. Mira, H., Ramırez-Castillejo, C. Defects in sub-ventricular zone pigmented epithelium-derived factorniche signaling in the senescence-accelerated mouseprone-8. FASEB J. 29, 1480–1492 (2015). www.fasebj.org
Key Words: self-renewal • neural stem cell • PEDF • acceleratedsenescence • long-term LRC
SAM-P8 PROVIDES A GOODmodel for age-related pathologies(1, 2). SAM-P8 presents an early cognitive impairment,which affects memory and learning skills (3, 4), causesmodifications in adenosine receptors (5), and an increasein the level of butyrylcholinesterase activity (6, 7). Recentdata describe alterations in stem cell proliferation in theforebrain of SAM-P8 preceding age-related brain symp-toms (1, 2). SAM-P8 presents an early cognitive impair-ment, which affects memory and learning skills (3, 4), andcauses modifications in adenosine receptors (5) and anincrease in the level of butyrylcholinesterase activity (6, 7).Recent data describe alterations in stem cell proliferationin the forebrain of SAMP8 preceding age-related brainsymptoms (8, 9). However, to date it remains unknownwhether loss of the uncommitted and multipotency stateof the neural stem cell (NSC) population in the adultsubventricular zone (SVZ) are involved in the senescence-accelerating process.
Abbreviations: BrdU, bromodeoxyuridine; BSA, bovine se-rum albumin; DCX, doublecortin; DIV, days in vitro, or days inculture; EBSS, Earle’s balanced salt solution; FBS, fetal bovineserum; FGF-2, Fibroblast growth factor 2; GFAP, glial fibrillaryacidic protein; Hes1, hairy and enhancer of split 1; Hes5, hairyand enhancer of split 5; LR-PEDF, laminin receptor (LR),a receptor for PB, phosphate buffer; LRC, label-retaining cell;
(continued on next page)
1 Current affiliation: Unidad Funcional de Investigaci on deEnfermedades Cronicas (Instituto de Salud Carlos III)/Hos-pital 12 de Octubre, Madrid, Spain.
2 Correspondence: 12 October Hospital/UFIEC, ISCIII,Carretera Majadahonda-Pozuelo Km 2, 28220 Madrid, Spain.E-mail: [email protected]: 10.1096/fj.13-244442This article includes supplemental data. Please visit http://
www.fasebj.org to obtain this information.
1480 0892-6638/15/0029-1480 © FASEB

Neurogenesis takes place in the SVZ and other adultbrain regions such as the hippocampus. Stem cells partic-ipate in brain activities like memory formation and recon-struction of new neuronal connections. More specifically,the SVZ is an ultrastructure that generates different cellu-lar outputs at different times in life, for instance, severalneuronal types during midgestation, glia production atperinatal times, or olfactory bulb-destined neuron pro-duction following perinatal times (10). This area containsat least 3 kinds of progenitor cells: B, C, and A cells. B cellsare NSCs characterized by extended division cell cycles(11, 12), and theirprogenyare transitory amplifyingneuralprogenitorsorCcells. In contrast,C cells divide rapidly andgenerate polysialylated neuronal cell adhesion molecule+
and doublecortin (DCX)+ neuroblasts, also called A-cells,which migrate to the olfactory bulb and then differentiateinto interneurons (13, 14). B cells can be distinguishedfrom C and A cells based on the expression of the glialfibrillary acidic protein (GFAP) and the retention of thy-midine analogs such as BrdU.
The number of adult-generated neurons is related tocognitive performance in certain tasks, and a loss ofthese newborn neurons could participate in prematuresenescence. Self-renewal of stem cells involves bothproliferation and maintenance of an undifferentiatedstate in response to essential microenvironmental sig-nals (15). This regulation is key for the maintenance ofproliferative lifelongpersistentNSCs (16).Niche signalsare capable of sustaining a more undifferentiated phe-notype, which influences the postnatal neurogenicoutput. Deficiencies in this output may include pre-mature differentiation, loss of self-renewal capacity, andappearance of senescence symptoms (17). Multiplefactors have been involved in self-renewal and neuro-genesis, including the PEDF, the Notch pathway, the tran-scription factors SOX2 and PAX6, and (18, 19).
PEDF is a neurotrophic and neuroprotective factor withanti-angiogenic activity (20) and is the first soluble factorknown to specifically activate SVZ B cells. PEDF’s activitytoward NSCs indicates that it may contribute to the main-tenance of a pool of undifferentiated stem cells in neuro-genic niches, which is necessary to achieve a normal rate ofneurogenesis and also protects from neurodegeneration(15). The signaling pathway controlling how PEDF is af-fecting self-renewal is largely unknown; however, 2 differ-ent membrane proteins have been postulated as putativePEDF receptors. The first, PNPLA2, has phospholipaseactivity (21), the second, LR-PEDF, has anti-angiogenicactivity like PEDF protein (22). Moreover, PEDF inducesa noncanonical activation of the NF-kB pathway in SVZNSCs, leading to the dismissal of the transcriptional nu-clear receptor corepressor from specific Notch-responsivepromoters (23).
One important pathway to control stem cells is indeedthe Notch-dependent pathway, which is essential for NSCmaintenance (24, 25). Concerning SOX2 expression, workbyFavaro et al. (26)hasdemonstrated that the transcriptionfactor SOX2 is essential for adult hippocampal NSC main-tenance. Also, recently it has been found that activation ofthe Notch-signaling pathway, which leads to the transcrip-tional activationof recombination signal-bindingprotein Jktargets, enhances the activity of the SOX2 promoter andthe expression of SOX2 in adult hippocampal NSCs (27).
PAX6 is crucial formaintaining thepool of neural stem/progenitor cells. Interestingly, this factor can also promoteneuronal differentiation in context-dependent manners(28). PAX6 is a highly conserved transcription factor and iscrucial for the development of the CNS, eyes, nose, pan-creas, and pituitary gland (29, 30). PAX6 is expressed inspecific spatiotemporal patterns during mammalian braindevelopment and is thought to modify the expression ofgenes involved inbrainmodeling,neuronalmigration, andneural circuit establishment [reviewed elsewhere (31, 32)].
The sheltering protein family has been related with age-related diseases (33, 34). The human telomeric DNA-binding factor TERF1 (hTERF1) and its interactingproteins TIN2, tankyrase 1 and 2, and PINX1, have beenimplicated in the regulation of telomerase-dependenttelomere length maintenance. TERF1 on the other hand,may have another essential function unrelated to telomerelength regulation (35). All these factorsmay be involved inthe pathogenesis of the accelerated-senescent mice. Pos-sible deregulation in their functions could be responsiblefor the cognitive impairments observed during aging.
In this article, we demonstrate that SAM-P8 mice havea reduction in thenumberof long-term label-retainingcells(LRCs) in theSVZata veryearly age,withoutanydetectablealteration in the number of fast-dividing progenitor cells.We also found a marked decrease in the PNPLA2 PEDF-receptor levels both in SVZ tissue and inNSC cultures fromSAM-P8.Furthermore, endothelial-releasedPEDF inducedan increase in the number of renewing cell divisions inNSCs cultured fromSVZ in SAM-P8, although the responsewas clearly attenuated compared with NSCs from the nor-mal aging SAM-resistant 1 mouse strain (SAM-R1). Inconclusion, PEDF niche signals and self-renewal relatedsignalingaredecreased in theSAM-P8model.Wepostulatethat this event may contribute to the loss of the NSC pooland to theappearanceofpremature senescence symptoms.
MATERIALS AND METHODS
Animals
SAM-P8 and SAM-R1 (1), at 1 month of age and of indistinctgender, were obtained fromHarlan UK, Ltd. (Blackthorn, UnitedKingdom) through Harlan Interfauna Iberica (Barcelona,Spain). All of the animals werehoused andbredunder the samecontrolled conditions: temperature 20–25°C, humidity of50–60%, and a 12 hour light–dark cycle. These were treated inaccordance with the approval of the local ethics committee(University of Castilla-La Mancha).
Dissection and neurosphere cultures
Brains of different animals were dissected, and the regions con-taining the SVZ were isolated from each hemisphere and washed
NSC, neural stem cell; Pax6, paired box gene 6; PEDF, pig-mented epithelium-derived factor; PFA, paraformaldehyde;PNPLA2, patatin-like phospholipase domain containing 2,a receptor for PEDF; SAM-P8, senescence-accelerated mouseprone-8, a standard aging model; SAM-R1, senescence acceler-ated mouse resistant-1, a control strain for SAM-P8; Sox2, SRY-boxcontaining gene 2; SVZ, subventricular zone; Terf1, telomericrepeat binding factor 1
(continued from previous page)
SAM-P8 NEURAL STEM CELL SIGNALS 1481

in Earle’s balanced salt solution (EBSS; Gibco, Carlsbad, CA,USA). Tissues were transferred to EBSS containing 1.0 mg/ml pa-pain (Worthington Biochemical, Lakewood, NJ, USA), 0.5 mg/mlL-cysteine (Sigma-Aldrich, St. Louis, MO, USA) and 0.5 mg/mlEDTA (Sigma-Aldrich), and incubated for 30–60 minutes at37°C. Tissue was then rinsed in EBSS for 10 minutes, trans-ferred to DMEM/F12 medium (1:1 v/v; Life Technologies,Gaithersburg, MD, USA), and then carefully triturated witha flame-narrowed Pasteur pipette. Isolated cells were collectedby centrifugation, resuspended in DMEM/F12 medium con-taining 2 mM L-glutamine, 0.6% glucose, 9.6 g/ml putrescine,HEPES 5M, 6.3 ng/ml progesterone, 5.2 ng/ml sodium sele-nite, 0.025mg/ml insulin, sodiumbicarbonate0.12%0.1mg/ml,transferrin, 2 mg/ml heparin (sodium salt, grade II; Sigma-Aldrich), 1% penicillin/streptomycin (control medium) andsupplemented with epidermal growth factor and fibroblastgrowth factor 2 (FGF-2) (human recombinant, 10 and 20 ng/ml,respectively) and 4 mg/ml bovine serum albumin (BSA; com-plete medium). The final density of plating was 13 105cells percm2 (50,000 cells/ml), in the same medium as previously de-scribed (36). For each passage, spheres formed after 6–8 days invitro (DIV) were mechanically dissociated into a single-cell sus-pension and replated in complete medium. At each passage, thetotal number of viable cells was determined by Trypan Blue exclu-sion. Cells were grown in ahumidified cell incubator at 37°Cundera 5% CO2 atmosphere. For differentiation assays, neurosphereswere dissociated and plated onto matrigel with 10 ng/ml FGF2 for2 DIV followed by 5 DIV in 2% fetal bovine serum (FBS).
Counting and diameter measuring of neurospheres
Diameter measurement and cell counting of neurosphere cul-tures were performed after 5–7 days division. Thediameter of theneurospheres was measured from images of neurospheres usingthe program ImageJ (U.S. National Institutes of Health,Bethesda, MD, USA). The photographs were taken with a digitalcamera attached to a microscope (Carl Zeiss, Jena, Germany).The amount of neurospheres was determined by random displaycounting under a microscope (Carl Zeiss) using a310 objectivein 90 fields/flask. Results are expressed as mean6 SEM of at least3 independent experiments. Statistical analysis was performedusing thenonparametricMann–WhitneyU test toassessdifferencesbetween groups. P, 0.05 was considered statistically significant.
BrdU labeling
For BrdU labeling of proliferating cells in the adult brain, micereceived 7 intraperitoneal injections of 50 mg/kg body weightBrdU every 2 hours, and they were killed either 12 hours later or1 month after the last injection.
Immunohistofluoresence
Animals were perfused with 4% paraformaldehyde (PFA) in0.1 M phosphate buffer, pH 7.4 (PB), and then fixed in 4% PFAfor 15 min at room temperature. The brains were cryopreservedin 30% sucrose during 12 hours and cut into cryostat sections ofabout 12mm. For BrdUdetection, the slides were treated with 0.1NHCl for 15min, neutralized in 0.1M sodiumborate buffer (pH8.5), and incubated for 1hourat roomtemperaturewithblockingbuffer (10% FBS, 5% BSA, 0.2% Triton X-100). Incubation withmouse anti-BrdU (1:100; Dako, Glostrup, Denmark), mouseanti–b-tubulin (Tuj, Covance, 1:300), rabbit anti-GFAP (Dako, 1:300) and mouse anti-Sox2 (1:50; R&D, R&D Systems, Minneap-olis, MN, USA) antibodies was performed overnight in PB con-taining 10% FBS, 5% BSA, 0.2% Triton X-100 (blocking buffer).After washing, the slides were incubated with Cy3-conjugated
anti-mouse or anti-rabbit antibodies (Jackson ImmunoResearchLaboratories Inc., West Grove, PA, USA) at 1:2000 dilution inblocking buffer for 45 minutes, followed by extensive washing inPB. Then, the slides were counterstained with DAPI (500 ng/ml,Sigma) and mounted with Fluorsave (Calbiochem, La Jolla, CA,USA). Fluorescent signal was detected with a Zeiss microscope(LSM710-ConfoCor 3 System;Carl Zeiss), at excitation/emissionwavelengths of 470/505 and 350/405 (UV).
Thepercentageof cells thatwereBrdU-positivewasdeterminedfor each group by counting the number of labeled and unlabeledcells. Counting of BrdU and DAPI cells was done for 8 fields perwell (on average 900 counted cells per replica). Three replicas percondition in 2 independent experiments were analyzed.
PEDF experiments
The effect of PEDF protein was assayed by adding conditionedmediumcontaining 200 ng/ml of recombinant PEDF (expressedusing a pcDNA3.1 vector transfected in 293T cells) to the differ-ent cell suspensions as previously described (36). The design ofthe experiments using PEDF was as follows. First, after disaggre-gation of the tissue, cells were plated with or without PEDF.Second, after 2 to 3 days in culture the neurospheres werecentrifuged and then disaggregated by gently pipetting up anddown (70 times) until a unicellular suspension was obtained.Third, the cells were plated at a density of 100,000 cells/ml, withandwithoutPEDF. Indoses-responseassays25, 50,and100ng/mlof PEDF recombinant protein were tested.
RNA extraction and cDNA synthesis
RNA was isolated using RNeasy MicroKit (Qiagen, Germantown,MD, USA). of RNA (1 mg) was reverse transcribed into cDNAusing random primers and SuperScript II RT reverse transcrip-tase (Kit RevertAid, TM Fermentas; Thermo Scientific,Waltham,MA,USA) in the presence of first strand buffer (50mMTris-HCl,75 mM KCl, 3 mM Mg2Cl), 5 mM DTT, and 0.25 mM each ofdeoxyribonucleotide triphosphates (AmershamBiosciences,Bucks,United Kingdom) for 2 hours at 42°C.
Quantitative real-time PCR
Real-time quantitative PCR (RT-qPCR)was performed accordingto themanufacturer’s protocol usingKit SYBRPremixExTaqTM(Takara, Tokyo, Japan) Gene Expression Assays, reference fluo-rescence ROX II, and an ABI Prism 7500 Sequence DetectionSystem (Applied Biosystems, Foster City, CA, USA). cDNA, pre-pared as described above, was used at a concentration of 0.2 mM.The gene expression assays used are described in Table 1. Genesusedwere:PEDF andPEDFprotein described receptors (PNPLA2(21) label as PEDF-receptor and LR-PEDF (22) in Table 1). Also, 2genes involves inPEDFphosphorylationwereused.These5geneswere selected to study the implicationof PEDFproteinpathway inself-renewal and premature senescence process. Finally, 3 genesrelated with self-renewal process were selected: Hes1, Hes5, andSRY-box containing gene 2 (Sox2), and also a gene related totelomeres senescence, Terf1, and a gene involved in the differ-entiation process, as Pax6. All of them selected to study the pos-sible implication of self-renewal anddifferentiation process in thepremature senescence. Ribosomal phosphoprotein P0 (RPLP0)genewas used as reference because is oneof themost suited genefor normalization purposes in brain murine model (37).
DNA Sequencing
Genomic DNA from SAM-R1 and SAM-P8 brain tissue wasisolated using the E.Z.N.A. Tissue DNA Kit (Omega Bio-Tek,
1482 Vol. 29 April 2015 CASTRO-GARCIA ET AL.The FASEB Journal x www.fasebj.org

Doraville, GA, USA) following the manufacturer’s instruc-tions. The translated region (exons I to VII) of PEDF wasamplified (exon I to IV; exon III to V; exon VI and VII) usingDNA Polymerase (Biotools, Madrid, Spain). The used oligo-nucleotide primers are listed in Table 2. For the PEDF-receptor, the complete DNA sequence of Genbank entryNC_000073, including nucleotides 1-6062 (specifying exons Ito IX), was amplified. PCR conditions were: 30 seconds at 94°C,followed by 35 cycles of 30 seconds at 60°C, and 30 seconds at72°C. PCR products were purified using the PCR Cleanup Kit(Millipore, Billerica, MA, USA). Sequencing reactions wereperformed using the Seq BigDye V3.1 sequencing kit (AppliedBiosystems) with primers detailed in Table 2, and then puri-fied with the Sequencing Reaction Cleanup Kit (Millipore).Analyses were performed with an ABI Prism 3100 automaticsequencer (Applied Biosystems), and the following NCBIReference Sequences were used to compare each sequence:Genbank NC_000077.5 for PEDF and Genbank NC_000073.5for PEDF-receptor.
Protein extraction, PAGE, and Western blot analysis
For protein extraction cells were homogenized in RIPA lysisbuffer (20 nM sodium phosphate buffer, 150 nM NaCl, 5 mMEDTA, 1% Triton X-100, 1 mM sodium orthovanadate, 1 mMNaF, 1 mM phenylmethylsulfonyl fluoride, 10 mg/ml aprotinin,10mg/ml leupeptin), andprotein concentrationwas determinedby Bio-Rad DC protein assay. SDS-PAGE was performed usingthe Mini-PROTEAN III gel electrophoresis system (Bio-Rad).Samples were incubated with loading buffer containing 100 mMb-mercaptoethanol at 95°C for 5 minutes. After electrophoresisgels were transferred onto Hybond ECL nitrocellulose mem-branes (Amersham). Membranes were blocked in Tris-buffersalinewith 0.2%Tween and5%skimmilk. Recombinantproteinswere immunodetectedwith eithermousemonoclonal anti-PEDF,clone 10F12.2 (Millipore) or anti-PNPLA2 (Sigma-Aldrich) di-luted at 1:500, or b-actin (clone AC15, 1:5000; Sigma-Aldrich). Ahorseradish peroxidase–conjugated antibody against mouse IgG(Sigma-Aldrich) was diluted at 1:10,000. Chemiluminescence wasperformed with Chemiluminescent HRP Substrate ImmobilonWestern blot reagent (Millipore).
Quantifications
Positive cells in the immunoassays were counted in 8 fields perslide (average of 900 counted cells per replicate) using the com-putational platform described next. Counting and evaluation ofdata were performed by a person blinded as to treatment or ge-notype of mice.
Computational platform
A Web services computing platform has been developed to ad-dress the cell-countingproblem.The software tool allows a remoteautomatic processing of the cell images using aWeb interface. Toachieve this facility, we have integrated the cell image-processingsoftware, LLCECO (38) in a web services platform. LLCECO isa classification and cell counting software joint developed byHighPerformance Computing Group, Universidad de La Laguna andStem Cell Molecular and Cellular Laboratory, University of Cas-tilla La-Mancha. TheWeb Services platform used to integrate thissoftware is OpenCF (39). LLCECO has been developed fromscratch for this project and designed as a tool to be exploited froma computing Web service approach. The tool allows not only cellcounting based on a set of parameters but also postprocessing ofthe data obtained (graph generation, spreadsheets, etc.). Agraphical interface has been added to facilitate the submission ofcomputational process with image sets, as well as Web services forthe execution of different tasks from a Web services client. Sup-plemental Figure 1 shows an example of this platform counting.
Statistics
We have analyzed 3 replicates per condition in 3 independent ex-periments in all assays. Mann–Whitney U test (40) test was applied.
RESULTS
NSC numbers are affected in the SVZ of SAM-P8
In this study, we employed 2 different BrdU administra-tion and sacrifice schemes to determine the proliferation
TABLE 1. Primers for gene expression
Gene Forward Reverse
Reverse transcriptase PCRPEDF cDNAa 59-CGGAATTCATGCAGGCCCTGGTGCTACTCC-39 59-GCGGGATCCGGGCCCCTGGGGTCCAGAA-39RT-qPCRPEDF-receptorb 59-CTGAAGCTGAGTTATGAAGGCGC-39 59-GGTTAAGGTGATAGTCCAGCGC-39
PEDFa 59-GAGCTACATCTTCTTCCTGCC-39 59-CCAGTAATCTTGCTGAAGTCGG-39Hesa 59-GCAGCATAGAGCAGCTGAAG-39 59-GAAGGCTTTGCTGTGTTCA-39Hes5a 59-GCAGCATAGAGCAGCTGAAG-39 59-GAAGGCTTTGCTGTGTTCA-39Sox2a 59-ATGCACAACTCGGAGATCAG-39 59-TATAATCC GGGTGCTCCTTC-39Pax6b 59-ACTTGGACGGGAACTGACAC-39 59-CACCACACCTGTCTCCTCCT-39LR-PEDF b 59-ATTGTTGCCATCGAGAATCC-39 59-CAGCATCCACCACATCAGAC-39CKII b 59-ATGTGGAGCTTGGGTTGTATG-39 59-ATGCTGGTTTTCACTGTGGAC-39PKAb 59-ATCTGGAAGTTCCGGTTCCT-39 59-TAAAAGTTGTCCCCGTCGTC-39Terf1b 59-AACAGCGCAGAGGCTATTATTCATGG-39 59-GTTTGTCAGTTCCTTTTCAATTG-39P0b 59-TGACTTGGTTGCTTTGGCGGGATTAG-39 59-GCACTTTCGCTTTCTGGAGG GTGTC-39b-Actinb 59-CCGGGACCTGACAGACTACCT-39 59-GCCATCTCCTGCTCGAAGTCTA-39
rrPEDF-receptorb,c 59-GGCTGCCAGACTCAATGAGG-39 59-GGACATCAGGCAGCCACTCC-39
PEDF cDNA was obtained by RT-PCR (Kit RevertAid TM Fermentas). The data generated by real-time PCR were plotted as the Rnfluorescence signal versus the cycle number, reference fluorescence ROX II and an ABI Prism 7500 Sequence Detection System Primer sequencesfor different genes. All genes were identified by GenBank accession number for the gene sequences used to design the primers. The primers weredesigned using Primer 3 program or Primer Express Software (Applied Biosystems), except the following. aPEDF, Hes1, Hes5, and Sox2 wereselected from Ramirez-Castillejo et al. (36). bPrimers designed with program Primers Express. cPrimers used in Supplemental Fig. 1.
SAM-P8 NEURAL STEM CELL SIGNALS 1483

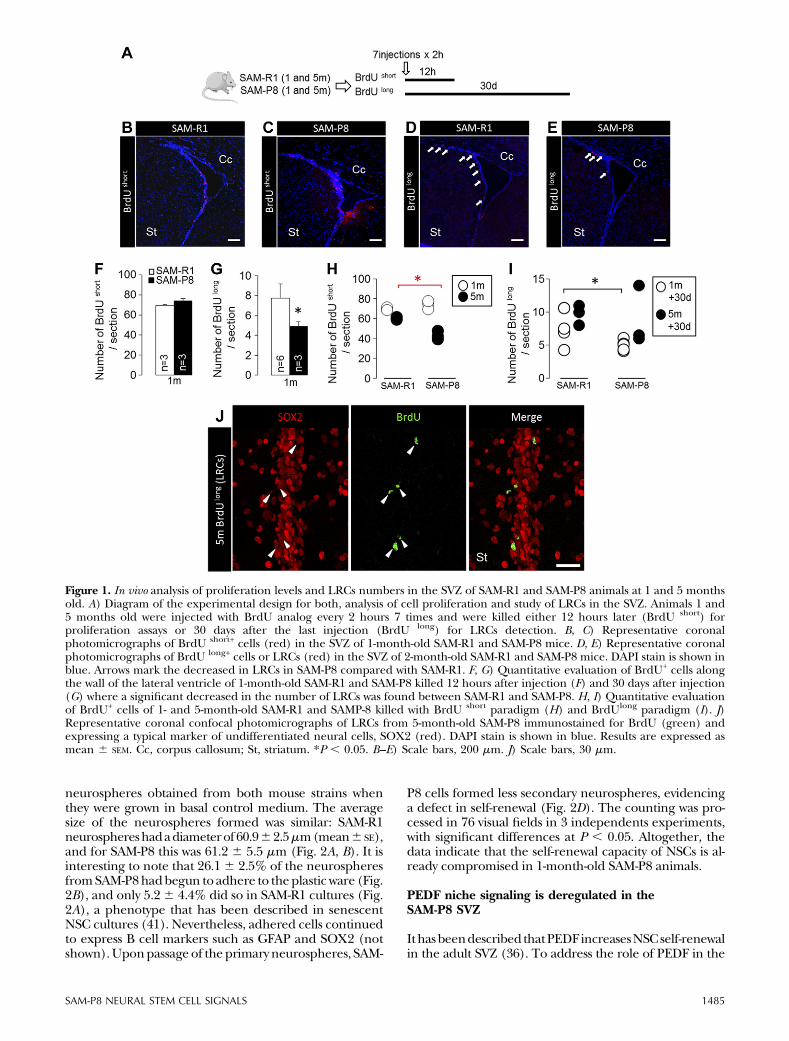
of SWCs and progenitors in the SVZ from young SAM-P8and SAM-R1 animals, as well as to localize long-term cy-cling stem cell populations (LRCs).One-month-oldmicereceived 7 intraperitoneal injections of BrdU every 2hours and were killed either 12 hours later (BrdUShort)or 30 days after the last injection (BrdULong). Immuno-fluorescence analysis using antibodies against BrdU areshown in Fig. 1. Shortly after BrdU administration, SAM-P8 animals showed a slight tendency to a higher numberof BrdUShort positive cells in the SVZ (P = 0.06, Fig. 1A–C,F), pointing to an incipient increase in overall SVZ pro-liferation that resulted in higher numbers of BrdU+DCX+
newly born neurons reaching the olfactory bulb 1 monthlater (Supplemental Fig. 1). This is in line with a recentstudy showing that proliferation in the SVZof SAM-R1 andSAM-P8 is similar in very young animals, transientlyincreases at the age of 2 months and abruptly declines atlater stages (8). In accordance with the aforementionedstudy, in 5-month-old SAM-P8 animals injected withBrdU and killed 12 hours later, the number of BrdUShort-positive cells was significantly diminished compared bothwith younger animals of the same strain and with SAM-R1animals of equivalent age (Fig. 1H). Despite such pro-liferation and neurogenic profile, we found a significantdecrease in long-term retaining BrdULong cells in SAM-P8compared with SAM-R1 animals (Fig. 1D, E,G, I). Of note,the majority of the SVZ-LRCs expressed SOX2 (Fig. 1J;88.6 6 7.3% in SAM-R1 and 83.6 6 4.3% in SAM-P8
animals of BrdUL cells at 30 days colabeled for SOX2),indicating SVZ-LRCs were not locally differentiated stemcell progeny that could retain the label due to cell cycleexit. Despite the difficulty, we were able to unequivocallyassign a GFAP+ process to approximately one third of theBrdUL+SOX2+ SVZ cells, supporting their B cell identity.Thus, in young SAM-P8 mice the SVZ already has about25% less label-retaining NSCs as compared with SAM-R1(P, 0.05, Fig. 1G). The reduced number of LRCs couldnot be attributed to an increase in apoptosis, because wedetected no differences in the presence of apoptoticfigures or cleaved caspase-3 cells between both micestrains (not shown). Even with that reduction, overallproliferation and neurogenesis are enhanced, pointingto compensatory mechanisms coming into play.
NSC self-renewal is compromised in SAM-P8
Next, we focused on the early ages to unravel whetherNSC cultures grown in vitro were also altered. Neuro-spheres were isolated from the SVZ of 1-month-old ani-mals and plausible differences between the NSC andprogenitor cell populations of SAM-P8 and SAM-R1wereanalyzed. After 5 days in vitro, SAM-P8 cultures containedsignificantly lower numbers of primary neurospheresthan those obtained from SAM-R1 (Fig. 2A–C). Inter-estingly, we found no differences in proliferation between
TABLE 2. Primers for gene sequencing
Name Forward Name Reverse
PEDF-receptor1a,b
59-AGGATCAGGCAGCTAAAACCC-39 PEDF-receptor 791a,b 59-CGTAGATGTGAGTGGCGTTGGC-39
PEDF-receptor694a,b
59-GACCAAGTGGAACATCTCATTCG-39 PEDF-receptor 1475a,b 59-GCTGGCTACAGCTGTCAATCAC-39
PEDF-receptor1329a,b,c
59-GGCTCTGCCGCTACAAACTGC-39 PEDF-receptor 2106a,b 59-CAATAGTACGGTGGCCAGAGAGAGG-39
PEDF-receptor1977a,b
59-AGAACAATGACAGGTTGCCTACTG-39 PEDF-receptor 2755a,b,c 59-CTTTAGTAGACAGCCACGGATGG-39
PEDF-receptor2667a,b
59-GAAGCAGGTGCCAACATTATTG-39 PEDF-receptor 3442a,b 59-GACACTGTGATGGTATTCTTCAGC-39
PEDF-receptor3244a,b
59-GGTGCCCATCTCAACTCC-39 PEDF-receptor 4036a,b 59-GCTTCCTCTGCATCCTCTTCC-39
PEDF-receptor3848a,b
59-GCTACAGAGATGGACTTCGATTCC-39 PEDF-receptor 4642a,b 59-CAATTTCCTCTTGGCCCTCATCACC-39
PEDF-receptor4573a,bc
59-CTGCCTGATGTCCCTGAAGATATCC-39 PEDF-receptor 5351a,b,c 59-GACTGACCCTGCACTGAAAAGG-39
PEDF-receptor5234a,b
59-CCTTTCACCAGCCACTCACTATGC-39 PEDF-receptor 6014a,b 59-GGCAGTGGAGCATGTGACAACC-39
PEDF-receptor5380a,b
59-CTCCACATCTGCTGCTGCTCC-39 PEDF-receptor 6063a,b 59-TGGCGGGAAGCAGGTATGG-39
PEDF1a 59-ATGCAGGCCCTGGTGCTA-39 PEDF3056a 59-CACGCCAAGGAGAAGGAT-39PEDF2313b 59-TCACCAACCCTGACATCC-39 PEDF4086b 59-ATCAGAGTCCAAGCCGTATC-39PEDF6779a 59-GGAAGGACTGCCAGTAACCA-39 PEDF7861a 59-GGGGGTTCTGTAGAGCACTG-39PEDF1370b 59-ACAGTTCAGGTCTGGCAAGC-39 PEDF300b 59-TAAAGCCCACAATCCAGGTC-39PEDF2775b 59-GGAAGATGCTGAGCAGGAAG-39 PEDF2517b 59-AACTTCACGCCAGGCATAAT-39PEDF3046b 59-TGCCCTCAGCATCCTTCT-39PEDF3784b 59-ATGGAGCCTATGGTCTGTGC-39
The translated region (exons I to VII) of PEDF was amplified (exon I to IV; exon III to V; exon VI and VII). Complete PEDF-receptor genewas amplified and sequenced (Gen Bank GeneID 66853). The following NCBI Reference Sequence was used to compare each sequence: GenBank NC_000077.5 for PEDF and Gen Bank NC_000073.5 for PNPLA2. aPrimers were used also for gene expression analysis. bPrimers designedfor gene amplification with program Primers Express. cPrimers used are shown in Supplemental Fig. 1G–I.
1484 Vol. 29 April 2015 CASTRO-GARCIA ET AL.The FASEB Journal x www.fasebj.org
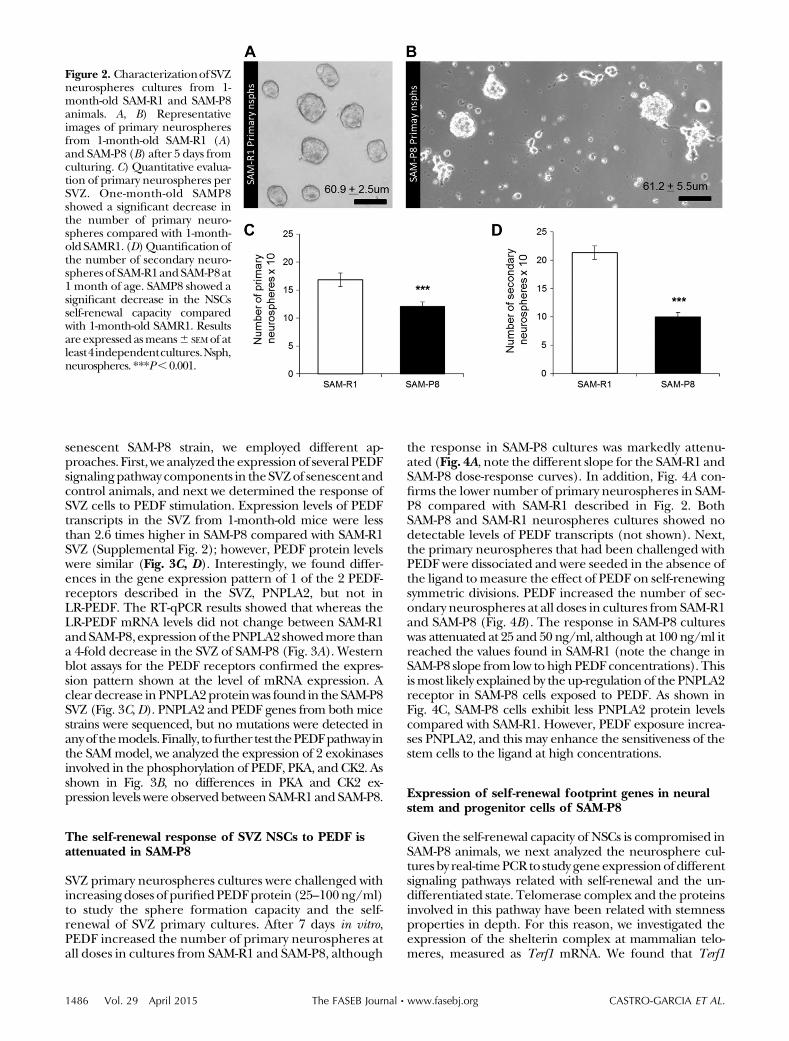
neurospheres obtained from both mouse strains whenthey were grown in basal control medium. The averagesize of the neurospheres formed was similar: SAM-R1neurosphereshadadiameterof60.962.5mm(mean6 SE),and for SAM-P8 this was 61.2 6 5.5 mm (Fig. 2A, B). It isinteresting to note that 26.16 2.5% of the neurospheresfromSAM-P8hadbegun to adhere to theplastic ware (Fig.2B), and only 5.26 4.4% did so in SAM-R1 cultures (Fig.2A), a phenotype that has been described in senescentNSC cultures (41). Nevertheless, adhered cells continuedto express B cell markers such as GFAP and SOX2 (notshown).Uponpassage of the primary neurospheres, SAM-
P8 cells formed less secondary neurospheres, evidencinga defect in self-renewal (Fig. 2D). The counting was pro-cessed in 76 visual fields in 3 independents experiments,with significant differences at P , 0.05. Altogether, thedata indicate that the self-renewal capacity of NSCs is al-ready compromised in 1-month-old SAM-P8 animals.
PEDF niche signaling is deregulated in theSAM-P8 SVZ
Ithasbeendescribed thatPEDFincreasesNSCself-renewalin the adult SVZ (36). To address the role of PEDF in the
Figure 1. In vivo analysis of proliferation levels and LRCs numbers in the SVZ of SAM-R1 and SAM-P8 animals at 1 and 5 monthsold. A) Diagram of the experimental design for both, analysis of cell proliferation and study of LRCs in the SVZ. Animals 1 and5 months old were injected with BrdU analog every 2 hours 7 times and were killed either 12 hours later (BrdU short) forproliferation assays or 30 days after the last injection (BrdU long) for LRCs detection. B, C) Representative coronalphotomicrographs of BrdU short+ cells (red) in the SVZ of 1-month-old SAM-R1 and SAM-P8 mice. D, E) Representative coronalphotomicrographs of BrdU long+ cells or LRCs (red) in the SVZ of 2-month-old SAM-R1 and SAM-P8 mice. DAPI stain is shown inblue. Arrows mark the decreased in LRCs in SAM-P8 compared with SAM-R1. F, G) Quantitative evaluation of BrdU+ cells alongthe wall of the lateral ventricle of 1-month-old SAM-R1 and SAM-P8 killed 12 hours after injection (F) and 30 days after injection(G) where a significant decreased in the number of LRCs was found between SAM-R1 and SAM-P8. H, I) Quantitative evaluationof BrdU+ cells of 1- and 5-month-old SAM-R1 and SAMP-8 killed with BrdU short paradigm (H) and BrdUlong paradigm (I). J)Representative coronal confocal photomicrographs of LRCs from 5-month-old SAM-P8 immunostained for BrdU (green) andexpressing a typical marker of undifferentiated neural cells, SOX2 (red). DAPI stain is shown in blue. Results are expressed asmean 6 SEM. Cc, corpus callosum; St, striatum. *P , 0.05. B–E) Scale bars, 200 mm. J) Scale bars, 30 mm.
SAM-P8 NEURAL STEM CELL SIGNALS 1485
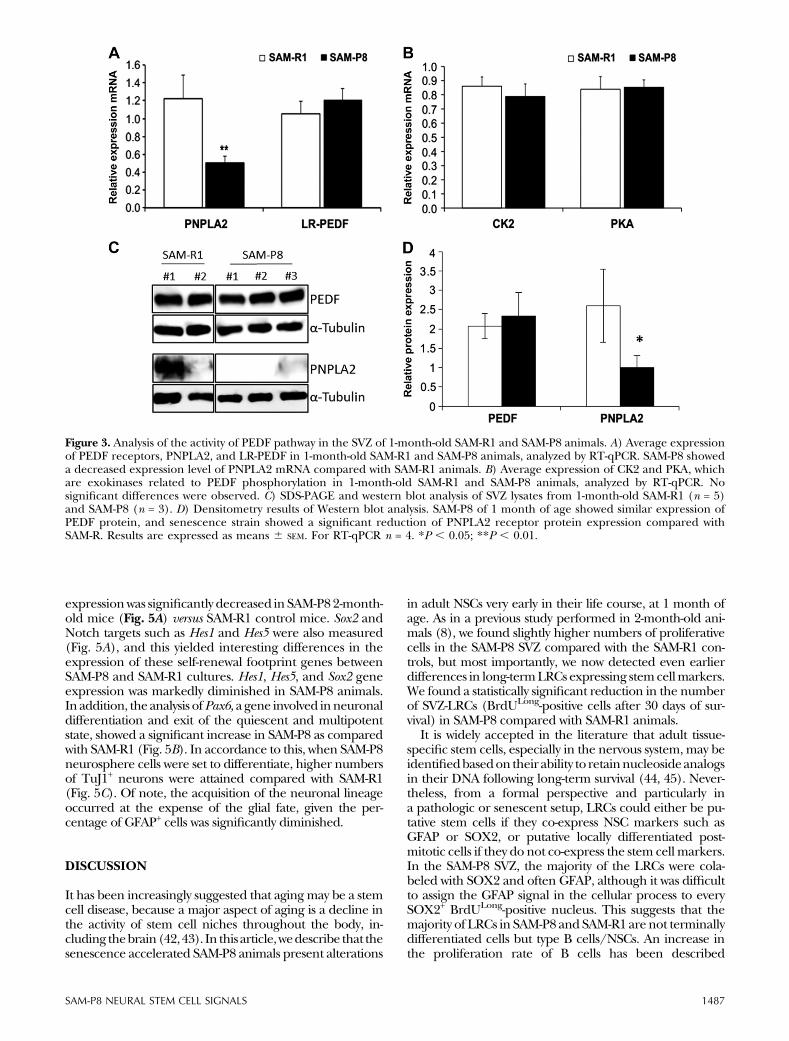
senescent SAM-P8 strain, we employed different ap-proaches. First,we analyzed the expressionof several PEDFsignalingpathway components in theSVZof senescent andcontrol animals, and next we determined the response ofSVZ cells to PEDF stimulation. Expression levels of PEDFtranscripts in the SVZ from 1-month-old mice were lessthan 2.6 times higher in SAM-P8 compared with SAM-R1SVZ (Supplemental Fig. 2); however, PEDF protein levelswere similar (Fig. 3C, D). Interestingly, we found differ-ences in the gene expression pattern of 1 of the 2 PEDF-receptors described in the SVZ, PNPLA2, but not inLR-PEDF. The RT-qPCR results showed that whereas theLR-PEDF mRNA levels did not change between SAM-R1andSAM-P8, expressionof thePNPLA2 showedmore thana 4-fold decrease in the SVZ of SAM-P8 (Fig. 3A). Westernblot assays for the PEDF receptors confirmed the expres-sion pattern shown at the level of mRNA expression. Aclear decrease in PNPLA2proteinwas found in theSAM-P8SVZ (Fig. 3C, D). PNPLA2 and PEDF genes from both micestrains were sequenced, but no mutations were detected inanyof themodels.Finally, to further test thePEDFpathway inthe SAMmodel, we analyzed the expression of 2 exokinasesinvolved in the phosphorylation of PEDF, PKA, and CK2. Asshown in Fig. 3B, no differences in PKA and CK2 ex-pression levels were observed between SAM-R1 and SAM-P8.
The self-renewal response of SVZ NSCs to PEDF isattenuated in SAM-P8
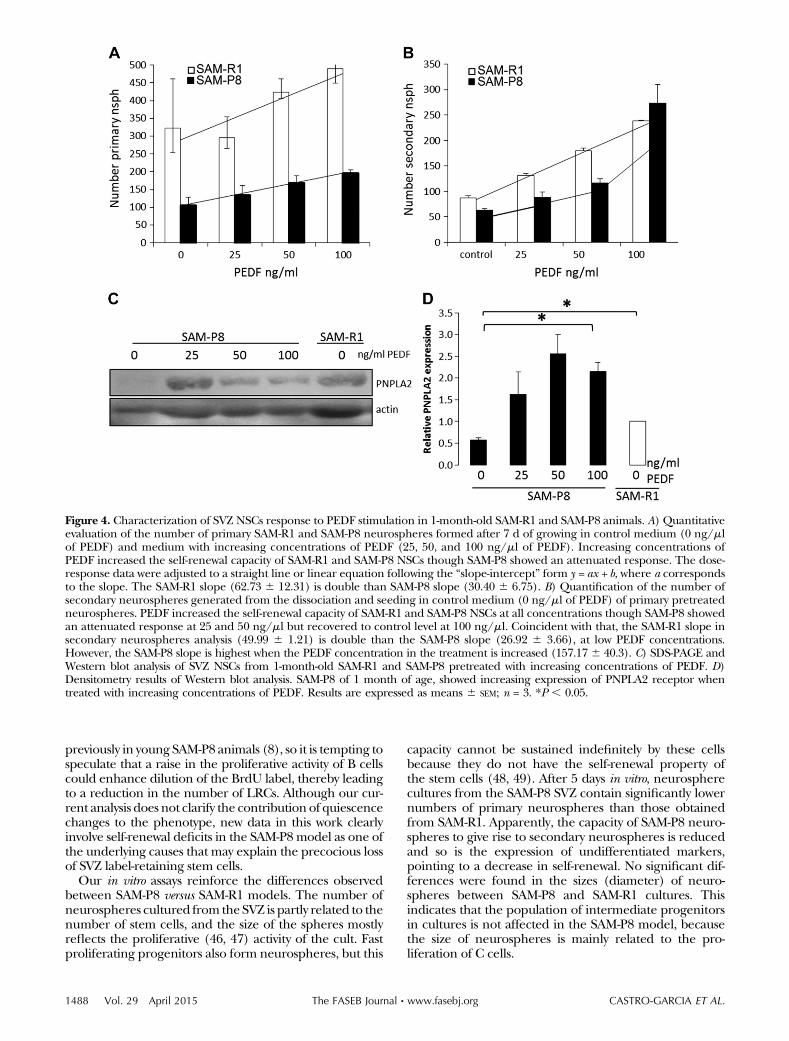
SVZ primary neurospheres cultures were challenged withincreasingdoses of purifiedPEDFprotein (25–100ng/ml)to study the sphere formation capacity and the self-renewal of SVZ primary cultures. After 7 days in vitro,PEDF increased the number of primary neurospheres atall doses in cultures from SAM-R1 and SAM-P8, although
the response in SAM-P8 cultures was markedly attenu-ated (Fig. 4A, note the different slope for the SAM-R1 andSAM-P8 dose-response curves). In addition, Fig. 4A con-firms the lower number of primary neurospheres in SAM-P8 compared with SAM-R1 described in Fig. 2. BothSAM-P8 and SAM-R1 neurospheres cultures showed nodetectable levels of PEDF transcripts (not shown). Next,the primary neurospheres that had been challenged withPEDF were dissociated and were seeded in the absence ofthe ligand tomeasure the effect of PEDF on self-renewingsymmetric divisions. PEDF increased the number of sec-ondary neurospheres at all doses in cultures from SAM-R1and SAM-P8 (Fig. 4B). The response in SAM-P8 cultureswas attenuated at 25 and 50 ng/ml, although at 100 ng/ml itreached the values found in SAM-R1 (note the change inSAM-P8 slope from low to highPEDF concentrations). Thisismost likely explainedby theup-regulation of the PNPLA2receptor in SAM-P8 cells exposed to PEDF. As shown inFig. 4C, SAM-P8 cells exhibit less PNPLA2 protein levelscompared with SAM-R1. However, PEDF exposure increa-ses PNPLA2, and this may enhance the sensitiveness of thestem cells to the ligand at high concentrations.
Expression of self-renewal footprint genes in neuralstem and progenitor cells of SAM-P8
Given the self-renewal capacity of NSCs is compromised inSAM-P8 animals, we next analyzed the neurosphere cul-tures by real-timePCRto study geneexpressionofdifferentsignaling pathways related with self-renewal and the un-differentiated state. Telomerase complex and the proteinsinvolved in this pathway have been related with stemnessproperties in depth. For this reason, we investigated theexpression of the shelterin complex at mammalian telo-meres, measured as Terf1 mRNA. We found that Terf1
Figure 2. CharacterizationofSVZneurospheres cultures from 1-month-old SAM-R1 and SAM-P8animals. A, B) Representativeimages of primary neurospheresfrom 1-month-old SAM-R1 (A)and SAM-P8 (B) after 5 days fromculturing. C) Quantitative evalua-tion of primary neurospheres perSVZ. One-month-old SAMP8showed a significant decrease inthe number of primary neuro-spheres compared with 1-month-old SAMR1. (D)Quantificationofthe number of secondary neuro-spheres of SAM-R1andSAM-P8at1 month of age. SAMP8 showed asignificant decrease in the NSCsself-renewal capacity comparedwith 1-month-old SAMR1. Resultsare expressed asmeans6 SEMof atleast4independentcultures.Nsph,neurospheres. ***P,0.001.
1486 Vol. 29 April 2015 CASTRO-GARCIA ET AL.The FASEB Journal x www.fasebj.org
expressionwas significantly decreased in SAM-P8 2-month-old mice (Fig. 5A) versus SAM-R1 control mice. Sox2 andNotch targets such as Hes1 and Hes5 were also measured(Fig. 5A), and this yielded interesting differences in theexpression of these self-renewal footprint genes betweenSAM-P8 and SAM-R1 cultures. Hes1, Hes5, and Sox2 geneexpression was markedly diminished in SAM-P8 animals.In addition, the analysis ofPax6, agene involved inneuronaldifferentiation and exit of the quiescent and multipotentstate, showed a significant increase in SAM-P8 as comparedwith SAM-R1 (Fig. 5B). In accordance to this, when SAM-P8neurosphere cells were set to differentiate, higher numbersof TuJ1+ neurons were attained compared with SAM-R1(Fig. 5C). Of note, the acquisition of the neuronal lineageoccurred at the expense of the glial fate, given the per-centage of GFAP+ cells was significantly diminished.
DISCUSSION
It has been increasingly suggested that agingmay be a stemcell disease, because a major aspect of aging is a decline inthe activity of stem cell niches throughout the body, in-cluding thebrain(42,43). In thisarticle,wedescribe that thesenescence accelerated SAM-P8 animals present alterations
in adult NSCs very early in their life course, at 1 month ofage. As in a previous study performed in 2-month-old ani-mals (8), we found slightly higher numbers of proliferativecells in the SAM-P8 SVZ compared with the SAM-R1 con-trols, but most importantly, we now detected even earlierdifferences in long-termLRCs expressing stemcellmarkers.We found a statistically significant reduction in the numberof SVZ-LRCs (BrdULong-positive cells after 30 days of sur-vival) in SAM-P8 compared with SAM-R1 animals.
It is widely accepted in the literature that adult tissue-specific stem cells, especially in the nervous system, may beidentifiedbasedon their ability to retainnucleosideanalogsin their DNA following long-term survival (44, 45). Never-theless, from a formal perspective and particularly ina pathologic or senescent setup, LRCs could either be pu-tative stem cells if they co-express NSC markers such asGFAP or SOX2, or putative locally differentiated post-mitotic cells if they do not co-express the stem cell markers.In the SAM-P8 SVZ, the majority of the LRCs were cola-beled with SOX2 and often GFAP, although it was difficultto assign the GFAP signal in the cellular process to everySOX2+ BrdULong-positive nucleus. This suggests that themajority of LRCs in SAM-P8 and SAM-R1 are not terminallydifferentiated cells but type B cells/NSCs. An increase inthe proliferation rate of B cells has been described
Figure 3. Analysis of the activity of PEDF pathway in the SVZ of 1-month-old SAM-R1 and SAM-P8 animals. A) Average expressionof PEDF receptors, PNPLA2, and LR-PEDF in 1-month-old SAM-R1 and SAM-P8 animals, analyzed by RT-qPCR. SAM-P8 showeda decreased expression level of PNPLA2 mRNA compared with SAM-R1 animals. B) Average expression of CK2 and PKA, whichare exokinases related to PEDF phosphorylation in 1-month-old SAM-R1 and SAM-P8 animals, analyzed by RT-qPCR. Nosignificant differences were observed. C) SDS-PAGE and western blot analysis of SVZ lysates from 1-month-old SAM-R1 (n = 5)and SAM-P8 (n = 3). D) Densitometry results of Western blot analysis. SAM-P8 of 1 month of age showed similar expression ofPEDF protein, and senescence strain showed a significant reduction of PNPLA2 receptor protein expression compared withSAM-R. Results are expressed as means 6 SEM. For RT-qPCR n = 4. *P , 0.05; **P , 0.01.
SAM-P8 NEURAL STEM CELL SIGNALS 1487
previously in young SAM-P8 animals (8), so it is tempting tospeculate that a raise in the proliferative activity of B cellscould enhance dilution of the BrdU label, thereby leadingto a reduction in the number of LRCs. Although our cur-rent analysis does not clarify the contribution of quiescencechanges to the phenotype, new data in this work clearlyinvolve self-renewal deficits in the SAM-P8 model as one ofthe underlying causes that may explain the precocious lossof SVZ label-retaining stem cells.
Our in vitro assays reinforce the differences observedbetween SAM-P8 versus SAM-R1 models. The number ofneurospheres cultured from theSVZ is partly related to thenumber of stem cells, and the size of the spheres mostlyreflects the proliferative (46, 47) activity of the cult. Fastproliferating progenitors also form neurospheres, but this
capacity cannot be sustained indefinitely by these cellsbecause they do not have the self-renewal property ofthe stem cells (48, 49). After 5 days in vitro, neurospherecultures from the SAM-P8 SVZ contain significantly lowernumbers of primary neurospheres than those obtainedfrom SAM-R1. Apparently, the capacity of SAM-P8 neuro-spheres to give rise to secondary neurospheres is reducedand so is the expression of undifferentiated markers,pointing to a decrease in self-renewal. No significant dif-ferences were found in the sizes (diameter) of neuro-spheres between SAM-P8 and SAM-R1 cultures. Thisindicates that the population of intermediate progenitorsin cultures is not affected in the SAM-P8 model, becausethe size of neurospheres is mainly related to the pro-liferation of C cells.
Figure 4. Characterization of SVZ NSCs response to PEDF stimulation in 1-month-old SAM-R1 and SAM-P8 animals. A) Quantitativeevaluation of the number of primary SAM-R1 and SAM-P8 neurospheres formed after 7 d of growing in control medium (0 ng/mlof PEDF) and medium with increasing concentrations of PEDF (25, 50, and 100 ng/ml of PEDF). Increasing concentrations ofPEDF increased the self-renewal capacity of SAM-R1 and SAM-P8 NSCs though SAM-P8 showed an attenuated response. The dose-response data were adjusted to a straight line or linear equation following the “slope-intercept” form y = ax + b, where a correspondsto the slope. The SAM-R1 slope (62.73 6 12.31) is double than SAM-P8 slope (30.40 6 6.75). B) Quantification of the number ofsecondary neurospheres generated from the dissociation and seeding in control medium (0 ng/ml of PEDF) of primary pretreatedneurospheres. PEDF increased the self-renewal capacity of SAM-R1 and SAM-P8 NSCs at all concentrations though SAM-P8 showedan attenuated response at 25 and 50 ng/ml but recovered to control level at 100 ng/ml. Coincident with that, the SAM-R1 slope insecondary neurospheres analysis (49.99 6 1.21) is double than the SAM-P8 slope (26.92 6 3.66), at low PEDF concentrations.However, the SAM-P8 slope is highest when the PEDF concentration in the treatment is increased (157.17 6 40.3). C) SDS-PAGE andWestern blot analysis of SVZ NSCs from 1-month-old SAM-R1 and SAM-P8 pretreated with increasing concentrations of PEDF. D)Densitometry results of Western blot analysis. SAM-P8 of 1 month of age, showed increasing expression of PNPLA2 receptor whentreated with increasing concentrations of PEDF. Results are expressed as means 6 SEM; n = 3. *P , 0.05.
1488 Vol. 29 April 2015 CASTRO-GARCIA ET AL.The FASEB Journal x www.fasebj.org
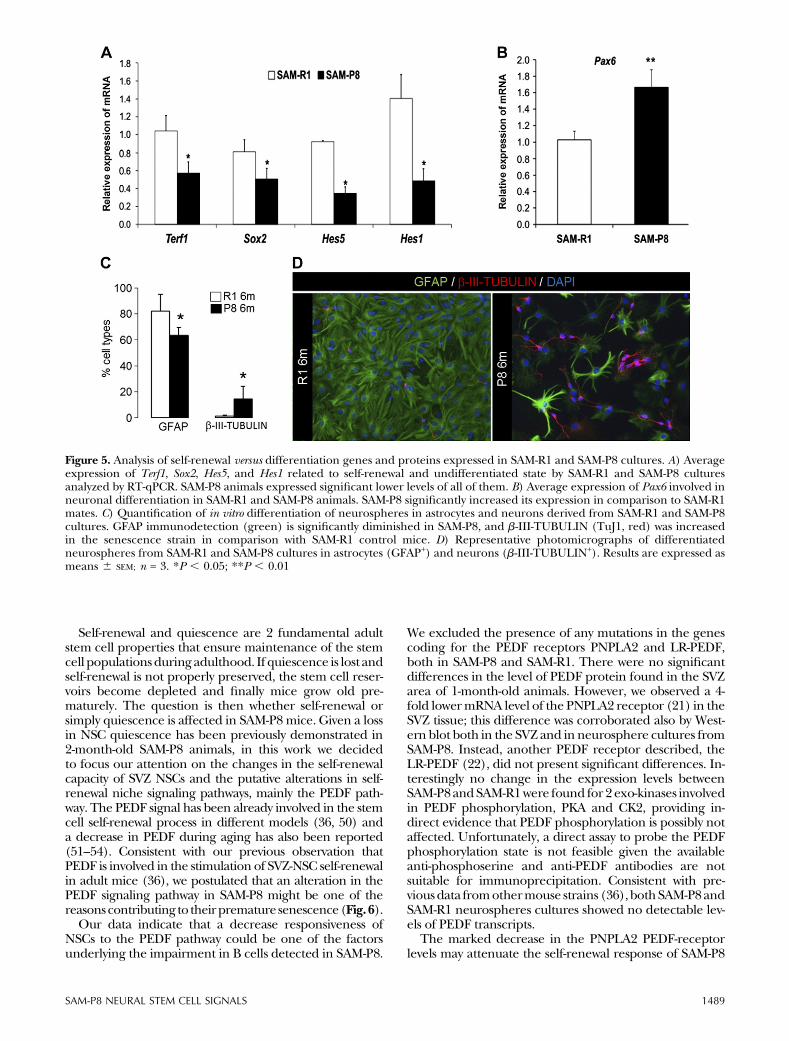
Self-renewal and quiescence are 2 fundamental adultstem cell properties that ensure maintenance of the stemcell populationsduring adulthood. If quiescence is lost andself-renewal is not properly preserved, the stem cell reser-voirs become depleted and finally mice grow old pre-maturely. The question is then whether self-renewal orsimply quiescence is affected in SAM-P8 mice. Given a lossin NSC quiescence has been previously demonstrated in2-month-old SAM-P8 animals, in this work we decidedto focus our attention on the changes in the self-renewalcapacity of SVZ NSCs and the putative alterations in self-renewal niche signaling pathways, mainly the PEDF path-way. The PEDF signal has been already involved in the stemcell self-renewal process in different models (36, 50) anda decrease in PEDF during aging has also been reported(51–54). Consistent with our previous observation thatPEDF is involved in the stimulation of SVZ-NSC self-renewalin adult mice (36), we postulated that an alteration in thePEDF signaling pathway in SAM-P8 might be one of thereasonscontributing to theirpremature senescence(Fig.6).
Our data indicate that a decrease responsiveness ofNSCs to the PEDF pathway could be one of the factorsunderlying the impairment in B cells detected in SAM-P8.
We excluded the presence of any mutations in the genescoding for the PEDF receptors PNPLA2 and LR-PEDF,both in SAM-P8 and SAM-R1. There were no significantdifferences in the level of PEDF protein found in the SVZarea of 1-month-old animals. However, we observed a 4-fold lowermRNA level of the PNPLA2 receptor (21) in theSVZ tissue; this difference was corroborated also by West-ern blot both in the SVZ and in neurosphere cultures fromSAM-P8. Instead, another PEDF receptor described, theLR-PEDF (22), did not present significant differences. In-terestingly no change in the expression levels betweenSAM-P8andSAM-R1were found for 2 exo-kinases involvedin PEDF phosphorylation, PKA and CK2, providing in-direct evidence that PEDF phosphorylation is possibly notaffected. Unfortunately, a direct assay to probe the PEDFphosphorylation state is not feasible given the availableanti-phosphoserine and anti-PEDF antibodies are notsuitable for immunoprecipitation. Consistent with pre-viousdata fromothermouse strains (36),bothSAM-P8andSAM-R1 neurospheres cultures showed no detectable lev-els of PEDF transcripts.
The marked decrease in the PNPLA2 PEDF-receptorlevels may attenuate the self-renewal response of SAM-P8
Figure 5. Analysis of self-renewal versus differentiation genes and proteins expressed in SAM-R1 and SAM-P8 cultures. A) Averageexpression of Terf1, Sox2, Hes5, and Hes1 related to self-renewal and undifferentiated state by SAM-R1 and SAM-P8 culturesanalyzed by RT-qPCR. SAM-P8 animals expressed significant lower levels of all of them. B) Average expression of Pax6 involved inneuronal differentiation in SAM-R1 and SAM-P8 animals. SAM-P8 significantly increased its expression in comparison to SAM-R1mates. C) Quantification of in vitro differentiation of neurospheres in astrocytes and neurons derived from SAM-R1 and SAM-P8cultures. GFAP immunodetection (green) is significantly diminished in SAM-P8, and b-III-TUBULIN (TuJ1, red) was increasedin the senescence strain in comparison with SAM-R1 control mice. D) Representative photomicrographs of differentiatedneurospheres from SAM-R1 and SAM-P8 cultures in astrocytes (GFAP+) and neurons (b-III-TUBULIN+). Results are expressed asmeans 6 SEM; n = 3. *P , 0.05; **P , 0.01
SAM-P8 NEURAL STEM CELL SIGNALS 1489

NSCs to PEDF in the SVZ niche. At least in vitro, PEDF wasless effective at promoting self-renewing symmetric divi-sions in SAM-P8 NSCs compared with SAM-R1 NSCs. Wealso performed gene expression analysis of several genessuch as the Notch targetsHes1andHes5, as well as the Sox2gene, which have been related with stem cell maintenanceand self-renewal previously (23, 36, 55). Of note, Hes1,Hes5, and Sox2 gene expression was down regulated inSAM-P8 animals. Interestingly, it has been described thatNotch signaling increases Sox2 promoter activity and Sox2expression in adult NSCs (27), and PEDFenhancesNotch-dependent transcription in cells with low Notch signaling,thereby subverting the output of an asymmetric NSC di-vision to theproductionof 2highly self-renewingcells (23).In our premature senescence model, the down-regulationof the stem cell–related genes could compromise themaintenance of normal stem cell numbers in the SVZ.These 2 phenomena (decreased sensitivity to the PEDFself-renewal signal and decreased expression of stemnessgenes) could be the origin of the premature senescenceprocess found in theSAM-P8model. Inaddition, analysis ofTrf1mRNA levels indicated that expressionof the shelterincomplex at mammalian telomeres was significantly de-creased in SAM-P8. This down-regulation in the shelterinexpression is detrimental to the protective effect oftelomere complex and could also be related with the
premature senescence phenotype of this mouse (56–58).TRF1 protein expression, which is maintained duringdevelopment (59), decreases during aging (60). Incontrast, higher expression of Pax6, a neurogenesis-controlling factor, was found in the SAM-P8 SVZ. Con-sistent with Pax6 functions in proliferation inhibitionand maturation promotion, SAM-P8 neurospheres cul-tures readily differentiated into the neuronal lineage atthe expense of the glial fate. This may contribute at leastin part to the increase in newly born neurons reachingthe SAM-P8 olfactory bulb.
In conclusion, PEDF niche signals and self-renewal re-lated genes are decreased in the SAM-P8 model. Wepostulate that this event limits stem cell maintenance inSAM-P8andmay contribute to the loss of theNSCpool andto the appearance of premature senescence symptoms.Further studies are necessary to ascertain whether this ir-reversibly affects the intrinsic regenerative capacity ofNSCsin vivo. If not, we anticipate that the activation of theremaining stem cell population could still result in an im-provement of the alterations that have been reported inthis senescent mouse model.
The authors thank Rafael Hortiguela for technicalsupport. This work was supported by grants from theAsociacion Española Contra el Cancer, the program of theInstituto de Salud Carlos III (Fondo de InvestigacionSanitaria PI081746), and Ministerio de Sanidad y Consumo(04005-00), the Consejerıa de Sanidad from Junta deComunidades de Castilla-La Mancha (PI2007/55); GrantSAF2008-05143-C03-1 from Comision Interministerial deCiencia y Tecnologıa to J.J. and Grant PI12/101from theSpanish Ministerio de Economıa y Competitividad, Fondode Investigacion Sanitaria and S2010/BMD-2336 from Comu-nidad de Madrid to H.M. C.R.C. was investigator of the“Ramon y Cajal Programme” from the Spanish “Ministerio deEducacion y Ciencia”. P.C.G. has a Consejo Nacionalde Ciencia y Tecnologıa fellowship for foreign study (MexicoCity, Mexico); ID Scholarship 205933. M.D.M. is a recipient ofa predoctoral fellowship (FPU) from the Spanish Ministeriode Educacion. The authors declare that no competingfinancial interests exist and also declare no sources of fundingand no contracts relating to this research.
REFERENCES
1. Takeda, T., Hosokawa, M., and Higuchi, K. (1997) Senescence-accelerated mouse (SAM): a novel murine model of senescence.Exp. Gerontol. 32, 105–109
2. Takeda, T., Matsushita, T., Kurozumi, M., Takemura, K.,Higuchi, K., and Hosokawa, M. (1997) Pathobiology of thesenescence-accelerated mouse (SAM). Exp. Gerontol. 32, 117–127
3. Flood, J. F., and Morley, J. E. (1998) Learning and memory in theSAMP8 mouse. Neurosci. Biobehav. Rev. 22, 1–20
4. Tomobe, K., Isobe, M., Okuma, Y., Kitamura, K., Oketani, Y., andNomura, Y. (2005) Genetic analysis of learning and memorydeficits in senescence-accelerated mouse (SAM). Physiol. Behav.84, 505–510
5. Castillo, C. A., Albasanz, J. L., Leon, D., Jordan, J., Pallas, M.,Camins, A., and Martın, M. (2009) Age-related expression ofadenosine receptors in brain from the senescence-acceleratedmouse. Exp. Gerontol. 44, 453–461
6. Fernandez-Gomez, F. J., Muñoz-Delgado, E., Montenegro, M. F.,Campoy, F. J., Vidal, C. J., and Jordan, J. (2008) The level ofbutyrylcholinesterase activity increases and the content of themRNA remains unaffected in brain of senescence-acceleratedmouse SAMP8. Chem. Biol. Interact. 175, 332–335
Figure 6. Hypothesis of PEDF receptor deficit in SAM-P8animals and the effect over the self-renewal capacity of thestrain. One-month-old SAM-R1 and SAM-P8 animals expressadequate levels of active PEDF protein and the exokinasesCK2 and PKA, involved in the phosphorylation of PEDF,which points to a normal production and signaling of PEDFnot only in SAM-R1 but also in SAM-P8. However, althoughSAM-R1 control animals present similar levels of PNPLA2 andLR-PEDF receptors, SAM-P8 animals show a significant re-duction of PNPLA2 receptor. This point to a defect in thereception of PEDF signaling, which leads to a lower self-renewal capacity of the SVZ NSCs of SAM-P8 mice.
1490 Vol. 29 April 2015 CASTRO-GARCIA ET AL.The FASEB Journal x www.fasebj.org

7. Fernandez-Gomez, F. J., Muñoz-Delgado, E., Montenegro,M. F., Campoy, F. J., Vidal, C. J., and Jordan, J. (2010)Cholinesterase activity in brain of senescence-accelerated-resistant mouse SAMR1 and its variation in brain ofsenescence-accelerated-prone mouse SAMP8. J. Neurosci. Res.88, 155–166
8. Dıaz-Moreno, M., Hortiguela, R., Gonçalves, A.,Garcıa-Carpio, I., Manich, G., Garcıa-Bermudez, E.,Moreno-Estelles, M., Eguiluz, C., Vilaplana, J., Pelegrı, C.,Vilar, M., and Mira, H. (2013) Ab increases neural stem cellactivity in senescence-accelerated SAMP8 mice. Neurobiol.Aging 34, 2623–2638
9. Gang, B., Yue, C., Han, N., Xue, H., Li, B., Sun, L., Li, X., andZhao, Q. (2011) Limited hippocampal neurogenesis inSAMP8 mouse model of Alzheimer’s disease. Brain Res. 1389,183–193
10. Lois, C., and Alvarez-Buylla, A. (1993) Proliferatingsubventricular zone cells in the adult mammalian forebraincan differentiate into neurons and glia. Proc. Natl. Acad. Sci. USA90, 2074–2077
11. Alvarez-Buylla, A., and Lim, D. A. (2004) For the long run:maintaining germinal niches in the adult brain. Neuron 41,683–686
12. Alvarez-Buylla, A., Seri, B., and Doetsch, F. (2002) Identificationof neural stem cells in the adult vertebrate brain. Brain Res. Bull.57, 751–758
13. Doetsch, F., Caille, I., Lim, D. A., Garcıa-Verdugo, J. M., andAlvarez-Buylla, A. (1999) Subventricular zone astrocytesare neural stem cells in the adult mammalian brain. Cell 97,703–716
14. Doetsch, F., Garcıa-Verdugo, J. M., and Alvarez-Buylla, A. (1997)Cellular composition and three-dimensional organization of thesubventricular germinal zone in the adult mammalian brain.J. Neurosci. 17, 5046–5061
15. Palmer, T. D., Takahashi, J., and Gage, F. H. (1997) The adult rathippocampus contains primordial neural stem cells. Mol. Cell.Neurosci. 8, 389–404
16. Doetsch, F. (2003) A niche for adult neural stem cells. Curr. Opin.Genet. Dev. 13, 543–550
17. Roobrouck, V. D., Ulloa-Montoya, F., and Verfaillie, C. M. (2008)Self-renewal and differentiation capacity of young and aged stemcells. Exp. Cell Res. 314, 1937–1944
18. Batista, L. F. (2014) Telomere biology in stem cells andreprogramming. Prog. Mol. Biol. Transl. Sci. 125, 67–88
19. Gomez-Lopez, S., Wiskow, O., Favaro, R., Nicolis, S. K., Price,D. J., Pollard, S. M., and Smith, A. (2011) Sox2 and Pax6maintain the proliferative and developmental potential ofgliogenic neural stem cells in vitro. Glia 59, 1588–1599
20. Barnstable, C. J., and Tombran-Tink, J. (2004) Neuroprotectiveand antiangiogenic actions of PEDF in the eye: moleculartargets and therapeutic potential. Prog. Retin. Eye Res. 23,561–577
21. Notari, L., Baladron, V., Aroca-Aguilar, J. D., Balko, N., Heredia,R., Meyer, C., Notario, P. M., Saravanamuthu, S., Nueda, M. L.,Sanchez-Sanchez, F., Escribano, J., Laborda, J., and Becerra, S. P.(2006) Identification of a lipase-linked cell membrane receptorfor pigment epithelium-derived factor. J. Biol. Chem. 281,38022–38037
22. Bernard, A., Gao-Li, J., Franco, C. A., Bouceba, T., Huet, A., andLi, Z. (2009) Laminin receptor involvement in the anti-angiogenic activity of pigment epithelium-derived factor. J. Biol.Chem. 284, 10480–10490
23. Andreu-Agullo, C., Morante-Redolat, J. M., Delgado, A. C., andFariñas, I. (2009) Vascular niche factor PEDF modulates Notch-dependent stemness in the adult subependymal zone. Nat. Neu-rosci. 12, 1514–1523
24. Androutsellis-Theotokis, A., Leker, R. R., Soldner, F.,Hoeppner, D. J., Ravin, R., Poser, S. W., Rueger, M. A., Bae,S. K., Kittappa, R., and McKay, R. D. (2006) Notch signallingregulates stem cell numbers in vitro and in vivo. Nature 442,823–826
25. Hitoshi, S., Alexson, T., Tropepe, V., Donoviel, D., Elia, A. J.,Nye, J. S., Conlon, R. A., Mak, T. W., Bernstein, A., andvan der Kooy, D. (2002) Notch pathway molecules are essentialfor the maintenance, but not the generation, of mammalianneural stem cells. Genes Dev. 16, 846–858
26. Favaro, R., Valotta, M., Ferri, A. L., Latorre, E., Mariani, J.,Giachino, C., Lancini, C., Tosetti, V., Ottolenghi, S., Taylor, V.,and Nicolis, S. K. (2009) Hippocampal development and neuralstem cell maintenance require Sox2-dependent regulation ofShh. Nat. Neurosci. 12, 1248–1256
27. Ehm, O., Goritz, C., Covic, M., Schaffner, I., Schwarz, T. J.,Karaca, E., Kempkes, B., Kremmer, E., Pfrieger, F. W., Espinosa,L., Bigas, A., Giachino, C., Taylor, V., Frisen, J., and Lie, D. C.(2010) RBPJkappa-dependent signaling is essential for long-termmaintenance of neural stem cells in the adult hippocampus.J. Neurosci. 30, 13794–13807
28. Kohwi, M., Osumi, N., Rubenstein, J. L., and Alvarez-Buylla, A.(2005) Pax6 is required for making specific subpopulations ofgranule and periglomerular neurons in the olfactory bulb.J. Neurosci. 25, 6997–7003
29. Hanson, I., and Van Heyningen, V. (1995) Pax6: more thanmeets the eye. Trends Genet. 11, 268–272
30. Kioussi, C., O’Connell, S., St-Onge, L., Treier, M., Gleiberman,A. S., Gruss, P., and Rosenfeld, M. G. (1999) Pax6 is essential forestablishing ventral-dorsal cell boundaries in pituitary gland de-velopment. Proc. Natl. Acad. Sci. USA 96, 14378–14382
31. Manuel, M., and Price, D. J. (2005) Role of Pax6 in forebrainregionalization. Brain Res. Bull. 66, 387–393
32. Osumi, N. (2001) The role of Pax6 in brain patterning. Tohoku J.Exp. Med. 193, 163–174
33. Kuimov, A. N. (2004) Polypeptide components of telomerenucleoprotein complex. Biochemistry Mosc. 69, 117–129
34. Martınez, P., Thanasoula, M., Muñoz, P., Liao, C., Tejera, A.,McNees, C., Flores, J. M., Fernandez-Capetillo, O., Tarsounas,M., and Blasco, M. A. (2009) Increased telomere fragility andfusions resulting from TRF1 deficiency lead to degenerativepathologies and increased cancer in mice. Genes Dev. 23,2060–2075
35. Sevigny, J. J., Ryan, J. M., van Dyck, C. H., Peng, Y., Lines,C. R., and Nessly, M. L.; MK-677 Protocol 30 Study Group.(2008) Growth hormone secretagogue MK-677: no clinicaleffect on AD progression in a randomized trial. Neurology 71,1702–1708
36. Ramırez-Castillejo, C., Sanchez-Sanchez, F., Andreu-Agullo, C.,Ferron, S. R., Aroca-Aguilar, J. D., Sanchez, P., Mira, H.,Escribano, J., and Fariñas, I. (2006) Pigment epithelium-derived factor is a niche signal for neural stem cell renewal.Nat. Neurosci. 9, 331–339
37. Pernot, F., Dorandeu, F., Beaup, C., and Peinnequin, A. (2010)Selection of reference genes for real-time quantitative reversetranscription-polymerase chain reaction in hippocampal struc-ture in a murine model of temporal lobe epilepsy with focalseizures. J. Neurosci. Res. 88, 1000–1008
38. Castillo, J. C., Almeida, F., Blanco, V., and Ramirez-Castillejo, C.(2014) Web services based platform for the cell counting prob-lem. 1st Workshop on Applications of Parallel Computation in Industryand Engineering; Euro-Par 2014: 20th International European Con-ference on Parallel Processing.
39. Santos, A., Almeida, F., Blanco, V., and Castillo, J. C.(2007) Lightweight web services for high performance com-puting. European Conference on Software Architecture 4758,225–236
40. Conover, W. J., and Iman, R. I. (1981) Rank transformations asa bridge between parametric and nonparametric statistics. Am.Stat. 35, 124–129
41. Porlan, E., Morante-Redolat, J. M., Marques-Torrejon, M. A.,Andreu-Agullo, C., Carneiro, C., Gomez-Ibarlucea, E., Soto,A., Vidal, A., Ferron, S. R., and Fariñas, I. (2013) Transcriptionalrepression of Bmp2 by p21(Waf1/Cip1) links quiescenceto neural stem cell maintenance. Nat. Neurosci. 16,1567–1575
42. Carlson, M. E., and Conboy, I. M. (2007) Loss of stem cellregenerative capacity within aged niches. Aging Cell 6, 371–382
43. Chambers, S. M., Shaw, C. A., Gatza, C., Fisk, C. J., Donehower,L. A., and Goodell, M. A. (2007) Aging hematopoietic stem cellsdecline in function and exhibit epigenetic dysregulation. PLoSBiol. 5, e201
44. Morshead, C. M., Craig, C. G., and van der Kooy, D. (1998) Invivo clonal analyses reveal the properties of endogenous neuralstem cell proliferation in the adult mammalian forebrain.Development 125, 2251–2261
SAM-P8 NEURAL STEM CELL SIGNALS 1491

45. Morshead, C. M., Reynolds, B. A., Craig, C. G., McBurney, M. W.,Staines, W. A., Morassutti, D., Weiss, S., and van der Kooy, D.(1994) Neural stem cells in the adult mammalian forebrain:a relatively quiescent subpopulation of subependymal cells.Neuron 13, 1071–1082
46. Kallos, M. S., Behie, L. A., and Vescovi, A. L. (1999) Extendedserial passaging of mammalian neural stem cells in suspensionbioreactors. Biotechnol. Bioeng. 65, 589–599
47. Weiss, S., Reynolds, B. A., Vescovi, A. L., Morshead, C., Craig,C. G., and van der Kooy, D. (1996) Is there a neural stemcell in the mammalian forebrain? Trends Neurosci. 19,387–393
48. Galli, R., Gritti, A., Bonfanti, L., and Vescovi, A. L. (2003) Neuralstem cells: an overview. Circ. Res. 92, 598–608
49. Gritti, A., Bonfanti, L., Doetsch, F., Caille, I., Alvarez-Buylla, A.,Lim, D. A., Galli, R., Verdugo, J. M., Herrera, D. G., andVescovi, A. L. (2002) Multipotent neural stem cells reside intothe rostral extension and olfactory bulb of adult rodents.J. Neurosci. 22, 437–445
50. Cao, W., Tombran-Tink, J., Chen, W., Mrazek, D., Elias, R., andMcGinnis, J. F. (1999) Pigment epithelium-derived factor protectscultured retinal neurons against hydrogen peroxide-induced celldeath. J. Neurosci. Res. 57, 789–800
51. Kojima, T., Nakahama, K., Yamamoto, K., Uematsu, H., andMorita, I. (2006) Age- and cell cycle-dependent changes in EPC-1/PEDF promoter activity in human diploid fibroblast-like(HDF) cells. Mol. Cell. Biochem. 293, 63–69
52. Smith, C. P., and Steinle, J. J. (2007) Changes in growth factorexpression in normal aging of the rat retina. Exp. Eye Res. 85,817–824
53. Steinle, J. J., Sharma, S., and Chin, V. C. (2008) Normal aginginvolves altered expression of growth factors in the rat choroid.J. Gerontol. A Biol. Sci. Med. Sci. 63, 135–140
54. Van Kirk, C. A., VanGuilder, H. D., Young, M., Farley, J. A.,Sonntag, W. E., and Freeman, W. M. (2011) Age-related alter-ations in retinal neurovascular and inflammatory transcripts.Mol. Vis. 17, 1261–1274
55. Pumiglia, K., and Temple, S. (2006) PEDF: bridging neurovascularinteractions in the stem cell niche. Nat. Neurosci. 9, 299–300
56. Donate, L. E., and Blasco, M. A. (2011) Telomeres in cancer andageing. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 76–84
57. Martınez, P., and Blasco, M. A. (2010) Role of shelterin in cancerand aging. Aging Cell 9, 653–666
58. Martınez, P., and Blasco, M. A. (2011) Telomeric and extra-telomeric roles for telomerase and the telomere-binding pro-teins. Nat. Rev. Cancer 11, 161–176
59. Klapper, W., Shin, T., and Mattson, M. P. (2001) Differentialregulation of telomerase activity and TERT expression duringbrain development in mice. J. Neurosci. Res. 64, 252–260
60. Blasco, M. A. (2014) Targetting telomeres in cancer. InternationalSymposium: Advanced Oncological Therapies. Ramon Areces Fundation.
Received for publication January 6, 2014.Accepted for publication December 5, 2014.
1492 Vol. 29 April 2015 CASTRO-GARCIA ET AL.The FASEB Journal x www.fasebj.org
Related Documents











