CYANOBACTERIA IN ANTARCTICA: ECOLOGY, PHYSIOLOGYAND COLD ADAPTATION K.D. PANDEY, S.P. SHUKLA, P.N. SHUKLA, D.D.GIRI, J.S. SINGH, P. SINGH and A.K. KASHYAP ! Centre of Advance Study in Botany, Banaras Hindu University, Varanasi 221005, India Fax: +91 542 231 7074; E-mail: [email protected] Received August 12, 2004; Accepted September 10, 2004 Abstract - Cyanobacterial species composition of fresh water and terrestrial ecosystems and chemical environment of water in Schirmacher Oasis in Continental Antarctica was investigated. Over 35 species of cyanobacteria were recorded. Diazotrophic species both heterocystous and unicellular contributed more than half to the count except in lake ecosystem. The species composition varied among the fresh water as well as terrestrial ecosystems. The physico-chemical analyses of water revealed its poor nurient content which might have supported the growth of diazotrophic cyanobacteria in an Antarctic environment. Among the cyanobacteria Oscillatoria, Phormidium and Nostoc commune were the dominant flora in most of the habitats. The physiological characteristics of isolated cyanobacteria strains indicated that N 2 -fixation, nitrate uptake, nitrate-reduction, ammonium-uptake, GS-transferase activity and photosynthesis was unaffected at low temperature (5C) which indicated low temperature adaptation for Antarctic cyanobacteria. This phenomenon was not evident in different strains of tropical origin. The temperature optima for N 2 -fixation for the different Antarctic cyanobacterial strains was in the range of 15-25C, nearly 10C lower than their respective reference strains of tropical origin. Similar results were obtained for cyanobacteria-moss association. The low endergonic activation energy exhibited by the above metabolic activities supported the view that cyanobacteria were adapted to Antarctic ecosystem. Key words: Antarctic cyanobacteria, ecology, physiology, temperature gradient, cold adaptation INTRODUCTION Since the first expedition (1895) to Antarctica, several studies have revealed the presence of microbial life forms on Ice surface near the south pole (9), 3.5 km deep Vostok ice covered lake (40) and on the ice cover of permanent lake ice (41). Antarctic ecosystems are dominated by prokaryotic microorganisms and in particular by photosynthetically active cyanobacteria. Being oxyphototrophic prokaryote, cyanobacteria are ubiquitous in all the major ecosystems of the world including Antarctica. Since Antarctica provides a harsh environment for the growth and survival of organisms, this group of organisms occupy specific niches in the Oasis and their distribution is very restricted. (23). Previous studies (7,33,35) indicate that cyanobacteria are most dominant component of flora in the regions of Antarctica that are ice- free (Oasis) during the polar summer. Also, the ability of cyanobacterial communities to fix atmospheric N 2 has been well documented. However, reports available on N 2 - fixation have not observed the species composition or diversity while those who determined species diversity did not examine the rates of N 2 -fixation in cyanobacterial mats or cyanobacteria-moss association. Cyanobacteria provide adequate amount of fixed carbon via photosynthesis to drive well developed ecosystem (54). Since the diazotrophic cyanobacteria which are the major component of the system (15,27) provide organic carbon and nitrogen to the Antarctic ecosystem, their presence will support both carbon and nitrogen cycle and in their absence biodiversity of microorganisms is generally compromised. Phototrophy, heterotrophy and diazotrophy may occur simultaneously in aggregates of microbial communities. Temperature, light energy, liquid water, nitrogen, phosphorus and trace metals are some of important factors which significantly regulate the processes in Antarctic ecosystem. To complete the C, N and P cycling, a close spatial proximity of phototrophs and heterotrophs is essential. They must exist in a highly efficient manner to survive in extreme cold conditions of Antarctica. Cyanobacteria in Antarctic ecosystem are adapted to the environment in terms of temperature, freezing and thawing cycle, photoprotection, light acquisition or photosynthesis, low humidity and prolonged period of 575 Cellular and Molecular Biology TM 50 (5), 575-584 ISSN 1165-158X DOI 10.1170/T547 2004 Cell. Mol. Biol . TM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

CYANOBACTERIA IN ANTARCTICA: ECOLOGY, PHYSIOLOGYAND COLD ADAPTATION
K.D. PANDEY, S.P. SHUKLA, P.N. SHUKLA, D.D.GIRI, J.S. SINGH, P. SINGH and A.K. KASHYAP!
Centre of Advance Study in Botany, Banaras Hindu University, Varanasi 221005, IndiaFax: +91 542 231 7074; E-mail: [email protected]
Received August 12, 2004; Accepted September 10, 2004
Abstract - Cyanobacterial species composition of fresh water and terrestrial ecosystems and chemical environment of water inSchirmacher Oasis in Continental Antarctica was investigated. Over 35 species of cyanobacteria were recorded. Diazotrophic speciesboth heterocystous and unicellular contributed more than half to the count except in lake ecosystem. The species composition variedamong the fresh water as well as terrestrial ecosystems. The physico-chemical analyses of water revealed its poor nurient contentwhich might have supported the growth of diazotrophic cyanobacteria in an Antarctic environment. Among the cyanobacteriaOscillatoria, Phormidium and Nostoc commune were the dominant flora in most of the habitats. The physiological characteristics ofisolated cyanobacteria strains indicated that N2-fixation, nitrate uptake, nitrate-reduction, ammonium-uptake, GS-transferase activityand photosynthesis was unaffected at low temperature (5°C) which indicated low temperature adaptation for Antarctic cyanobacteria.This phenomenon was not evident in different strains of tropical origin. The temperature optima for N2-fixation for the differentAntarctic cyanobacterial strains was in the range of 15-25°C, nearly 10°C lower than their respective reference strains of tropicalorigin. Similar results were obtained for cyanobacteria-moss association. The low endergonic activation energy exhibited by the abovemetabolic activities supported the view that cyanobacteria were adapted to Antarctic ecosystem.
Key words: Antarctic cyanobacteria, ecology, physiology, temperature gradient, cold adaptation
INTRODUCTION
Since the first expedition (1895) to Antarctica, severalstudies have revealed the presence of microbial life formson Ice surface near the south pole (9), 3.5 km deep Vostokice covered lake (40) and on the ice cover of permanentlake ice (41). Antarctic ecosystems are dominated byprokaryotic microorganisms and in particular byphotosynthetically active cyanobacteria. Beingoxyphototrophic prokaryote, cyanobacteria are ubiquitousin all the major ecosystems of the world includingAntarctica. Since Antarctica provides a harsh environmentfor the growth and survival of organisms, this group oforganisms occupy specific niches in the Oasis and theirdistribution is very restricted. (23). Previous studies(7,33,35) indicate that cyanobacteria are most dominantcomponent of flora in the regions of Antarctica that are ice-free (Oasis) during the polar summer. Also, the ability ofcyanobacterial communities to fix atmospheric N2 hasbeen well documented. However, reports available on N2-fixation have not observed the species composition ordiversity while those who determined species diversity did
not examine the rates of N2-fixation in cyanobacterial matsor cyanobacteria-moss association. Cyanobacteria provideadequate amount of fixed carbon via photosynthesis todrive well developed ecosystem (54). Since thediazotrophic cyanobacteria which are the majorcomponent of the system (15,27) provide organic carbonand nitrogen to the Antarctic ecosystem, their presence willsupport both carbon and nitrogen cycle and in their absencebiodiversity of microorganisms is generally compromised.Phototrophy, heterotrophy and diazotrophy may occursimultaneously in aggregates of microbial communities.Temperature, light energy, liquid water, nitrogen,phosphorus and trace metals are some of important factorswhich significantly regulate the processes in Antarcticecosystem. To complete the C, N and P cycling, a closespatial proximity of phototrophs and heterotrophs isessential. They must exist in a highly efficient manner tosurvive in extreme cold conditions of Antarctica.
Cyanobacteria in Antarctic ecosystem are adapted tothe environment in terms of temperature, freezing andthawing cycle, photoprotection, light acquisition orphotosynthesis, low humidity and prolonged period of
575
Cellular and Molecular BiologyTM 50 (5), 575-584 ISSN 1165-158XDOI 10.1170/T547 2004 Cell. Mol. Biol.TM

darkness (26). Nevertheless, only few cyanobacteria can beconsidered true psychrophiles and are mostly associatedwith polar fresh water ecosystem (16). Their higher growthrate, N2-fixation and photosynthetic optima at about 20°C,make them psychrotolerent. Nadeau and Castenholz (26)isolated true psychrophilic Oscillatorian cyanobacteriafrom a pond on the Ross Ice shelf, Antarctica. Geneticanalysis of the strains revealed that psychrotolerant formsare more closely related to temperate cyanobacteria andtrue psychrophiles probably evolved in polar environment.Cyanobacteria while growing in Antarctica have theunique ability to maintain the vital process ofphotosynthesis at freezing temperature (-5°C) (1). Theearlier reports have established that nitrogenase, a coldlabile enzyme responsible for atmospheric nitrogenfixation, remains active at �7°C (11). The temperatureminima and maxima in Antarctica and tropics are ofentirely different magnitudes, a different physiological setup is assumed in Antarctic and tropical strains ofcyanobacteria.
In the present communication, an attempt has beenmade to investigate the diversity of cyanobacteria occuringin different fresh water ecosystems and on soils and rocksof Schirmacher Oasis in Continental Antarctica. Nutrientstatus of fresh water streams, ponds and lakes wasdetermined to correlate the nutrient status with theiroccurrence. Further, physiological studies like N2-fixation,nitrate and ammonia uptake and their metabolism wereconducted in natural and isolated strains to explore thephysiological basis of cold adaptation.
MATERIALS AND METHODS
Description of siteSchirmacher Oasis (70° 46�04� - 70° 44� 21� S; 11° 49�54� - 11° 26�
03� E), Dronning Maud land is about 70 km. south of Princess Astridcoast. The Oasis is nearly 30 km long, oriented in East-West directionwith maximum width of 3 km and an area of 20 km2. The elevation ofOasis ranges between 0-228 meter. It is surrounded by 100 km wide ice-shelf in the north of the Oasis and 100 m thick and about 50 km widebasin glaciers in the south (19,35).
Identification and enumeration of cyanobacteriaCyanobacterial samples were collected in sterile polythene bags and
specimen tubes using a variety of clean implements. They wereexamined microscopically within 24 hr of collection. At least 20-25 slidesof each sample were examined. Identification based on cell dimensionsand morphological characteristics, was made with the help of literature(3,12,17,43).
Isolation of cyanobacterial strainsThe cyanobacterial natural samples, directly transferred on BG-11
(43) nutrient agar slants, were incubated photoautotrophically in GreenHouse of Indian station, Maitri for adaptation. Green house temperatureranges within 10-15°C and light intensity was about 500 µ Einstein m-2sec-1.The slants were brought to the laboratory at Banaras Hindu University,Varanasi (India). Clones were isolated following the standardmicrobiological techniques and were maintained in BG-11 medium
either in combined inorganic nitrogen supplement or in its absencedepending upon diazotrophic nature of the strains. The axenic cloneswere grown in an incubator at 20°C and light-intensity of 50 µ E m-2sec-1.
Cyanobacterial abundance relationshipRelative frequency (RF) and relative density (RD) of a species was
determined as:Number of samples in which species are present
RF = x 100Total number of samples
Number of individuals of a species in microscopic fieldRD = x 100
Total number of individuals in microscopic field
where a microscopic field represents an area of 1.31 mm2.The average relative frequency and density [(RFD= RF + RD)/2] at
a site was calculated and maximum value was considered at 100%. Allthe species of that site were divided into 4 classes: 0-25 (taxon present),26-50 (common constituent), 51-75 (dominant) and 76-100 (important)similar to those of Priddle and Belcher (39).
The similarity matrix, SJ (coefficient of Jaccard Sj=Nab/(Nab + Na+ Nb) (22), represents the ratio of shared and unique taxa in sites A andB and Na and Nb are the numbers of taxa restricted to sites A and B,respectively.
Water analysisWater samples from various aquatic ecosystems were collected in a
sterile plastic container and stored at 4°C during the stay andtransportation. The pH was measured by pH meter, chloride wasdetermined by argentometric method and sulphate was estimatedgravimetrically after precipitation in HCl as barium sulphate (42).Ammonium was estimated by using Nessler�s reagent (8). Nitrate wasdetermined by Brucine-sulphuric acid method (28) and phosphate wasmeasured as orthophosphate (14).
Moisture content of moss samplesThe fresh weight of mass samples (one cm3 core) collected from
different sites was estimated during 6 hr of collection. They were ovendried at 80°C till constant weight and moisture content was determined.
Temperature regulation and activation energyThe activation energy (Ea) was calculated from the Arrhenius plot
using the formula∆ log VT
Ea = 2.303 R - - - - - - - - - -∆ 1 / T
where VT represents the velocity of enzymatic reaction at temperature t,T is the absolute temperature (oK) and R is the gas constant.
To obtain the desired temperature, cyanobacterial cells wereincubated in a water bath made up of perpex sheet and connected to watercirculator (Haake, F-3, Germany). Light was provided with coolfluorescent tubes at the intensity of 50 µ E m-2 sec-1 on the surface ofculture vessel (34). The cells were pre exposed to a temperature for 5-6hr before performing the analyses.
Estimation of chlorophyll-αChlorophyll-α was extracted in 80% acetone (80: 20%, v/v) and
quantitative estimation was made by measuring absorbance and usingspecific absorption coefficient (α) as described by Myers and Kratz (25).
Estimation of proteinProtein content of cyanobacterial samples was determined by the
method of Lowry et al. (24) as modified by Herbert Phipps and Strange(18) using lysozyme as standard.
576 K.D. Pandey et al.

Antarctic cyanobacteria-cold adaptation 577
Uptake of nutrientsUptake of nitrate/ammonium was determined using exponential
phase diazotrophic cells. The cultures were harvested (10,000 g; 10 min),washed with buffer and re-suspended in 50 ml phosphate buffer (0.02 MpH 7.0) with a final density of 200 µg protein ml-1 and incubated in light.The uptake was initiated by addition of KNO3/NH4Cl (100 µM and 50µM, respectively). The disappearance of nitrate/ammonia ions from theassay medium was determined colorimetrically and the uptake rates werecalculated from the linear portion of the curve (50).
Enzyme assay� Nitrogenase: In vitro assay of nitrogenase (C2H2-reduction) was
performed according to Schödlhorn and Burris (44) as described byPandey and Singh (32) in Venoject tubes (Terumomedical, USA) sealedwith air tight rubber septum. The analysis was performed in a gaschromatograph (Tracor, 540 USA) connected to a 3392 A HewlettPackard integrator filled with parapak-R column. The enzyme activitywas expressed as nmol C2H4 mg-1 protein min-1 or nmol C2H4 µg-1Chl a_hr-1.
The moss samples were incubated in an incubator in light at atemperature of 20°C. Dried cyanobacterial flakes of N. commune werewetted with sterile double distilled water and were incubated in sameway. Sample of moss community (1 cm3 core) or 100 mg moistcyanobacterial flakes was placed in Venoject tubes and C2H2-reductionwas performed.
� Nitrate reductase: The nitrate reductase activity was determined asdescribed by Pandey and Singh (32). The crude enzyme of the cells wasextracted in 50 mM Tris-HCl buffer (pH. 7.5) containing sucrose (250mM), EDTA (1 mM) and MgCl2 (5 mM). The enzyme was mixed withreaction mixture and incubated at desired temperature for 30 min. Thenitrite formed was determined by the method of Snell and Snell (51).
Glutamine synthetase transferase activityThe glutamine synthetase (GS) assay was performed to measure
transferase activity of GS following the method of Shapiro and Stadtman(46). The enzyme activity was quantified by measuring the productformed (γ-glutamine hydroxymate) and has been expressed at µmol γ-glutamyl hydroxymate mg-1 protein min-1.
Photosynthetic PS-II activity (O2 evolution)The O2 evolution rate was measured using Clark type electrode as
described by Prasad and Kashyap (38). The temperature was regulated inwater bath. The light intensity at the surface of reaction vessel wasmaintained at ~20 Watt m-2 with the help of halogen lamp (8 V, 50 W.Kaondo, Japan) during the measurement. The culture was continuouslystirred on a magnetic stirrer (600 rpm).
Salinity treatmentThe buffered culture medium (HEPES, 20 mM, pH 7.5) was
supplemented with various concentrations of NaCl (50-300 mM) tocreate different levels of salinity. The concentration of NaCl for inhibitionof 50% activity of the enzyme was considered as IC-50.
RESULTS
Ecology of cyanobacteria in Schirmacher OasisThe fresh water ecosystems, lakes, ponds and streams
and terrestrial habitats, soil, and rock of the Oasis differedin species composition of cyanobacteria. Two types of meltwater streams were encountered in the ice free region, theglacier bound streams and snow drift stream. The glacierbound streams originate from the glacier during polar
summer and carry melt water from glacier to the lakes. Thewater flow is rapid depending upon the elevation of boldersand rocks. The snow drift melt water streams are shorter inthe length and carry less water. Glacier melt water stream(SEM) flows east of Maitri (Indian station) and carrieswater to the Priyadarshani lake. SEM supported the growthof a maximum number of cyanobacteria compared to otherstreams of the Oasis. The visual observations disclosed thatupper stream supported the least growth of cyanobacteriadominated by Oscillatoria and Phormidium. The growthof cyanobacteria was abundant and readily visible on therocks, bolders and on the bottom soil of the middle regionsof the streams. The flora of this region was mostlydominated by N2-fixing species Gloeocapsa, Nostoc andCalothrix. The lower stream closely resembles the middlestream in terms of the cyanobacterial composition.However, the region also supported the growth of non N2-fixers and green filamentous algae.
The richness of cyanobacteria in snow drift melt waterstream was comparable to the stream SEM. Stream NScarried water to Shivling lake and has several channelsbefore its terminating end. The contribution of diazotrophicspecies both heterocystous and aerobic unicellular N2-fixers in both the streams was more than 50% of the countsand their dominance was greatest in the middle region ofstream (Table 1).
Oasis supported large number of lakes, which areapparent only during polar summer and in the winter thelakes change into icefields. Priyadarsini lake supported thegrowth of 17 species of cyanobacteria. Most of the specieswere non N2-fixer and the diazotrophic species contributed1/3rd of total species count. The cyanobacteria formed thedominant constituent only in lake as this water body alsosupported good growth of a number of green algae anddiatoms. Two types of ponds are found in the Oasis � dueto glacier melt water at its edge and seepage wateraccumulated from melting of snow of rocks, soils and hills.They are comparatively smaller and the water is turbid.One of the pond selected near the Russian stationsupported growth of 13 cyanobacterial species and thecontribution of N2-fixing species was about 70% (Table 1).
Among the several moss sites visited in the Oasis, themoss sample collected near the melt water streamsupported growth of the maximum number ofcyanobacterial species. The contribution of diazotrophicspecies growing in association with moss at this site wasnearly 70%. Stigonema minutum was an importantconstituent of the moss community followed by Nostoccommune and Gloeocapsa species. A Nunatak, nearly 35km south to the Maitri was 78 meter high as measured inthe month of January. Moss growing on it supportedgrowth of cyanobacterial species. This site was mostpeculiar in that it remains ice covered during most part ofthe year and only its top portion is exposed for a brief

Table 1 Distribution pattern of cyanobacteria in streams, lakes, epiphytic on moss communities and on soils and quartz rocks inSchirmacher Oasis and a nearby Nunatak, Antarctica
Cyanobacteria Stream Lake In Association with Moss Soil and Rock
SEM NS Priyadarshini Pond Near on Soil Quartz Stream Nunatak Rock
Unicellular:1. Synechococcus aeruginosus Nag. ∆ ∆ ∆2. Chroococcus aeruginosus Nag. x3. C. pallidus Nag. x x4. C. minutus (Kütz.) Nag. ∆ x5. Gloeocapsa kuetzingiana Nag. ∆ ∆ •6. G. ralfsiana (Harve.) Kürtz. • ∆ ∆ ∆ x7. G. magna (Breb.) Kürtz. ∆ x ∆8. Gloecocapsa sp. ∆ x o o9. Aphanothece nidulans Richter. ∆ ∆ ∆10. A. muscicola (Menegh) Wille. x11. Chaemosiphon subglobosus (Rostaf.) Lemm. ∆
Filamentous non-heterocystous:12. Lyngbya aestuarii Liebm. ex Gomont ∆ ∆ ∆ •13. L. attenuata Fritsch. ∆ o14. Oscillatoria limosa Ag. ex Gomont. o ∆ x15. O. kuetlizii Lemm. • ∆16. O. limnetica Lemm. • ∆ • o x17. O. agardhii Gomont. • x18. O. tenuis Ag. ex Gomont. x • o x19. Phormidium fragile Gomont ∆ ∆ • x20. P. frigidum Fritsch. o o o o • ∆21. P. autumnale (Ag.) Gomont. ∆22. Plectonema sp. ∆ x ∆23. Schizothrix sp. I ∆ x24. Schizothrix sp. II x
Heterocystous:
25. Nostoc commune Vaucher ex Born et Flah o • ∆ o o • o
26. N. sphaericum Vaucher ex. Born et Flah ∆ ∆ ∆ •
27. Nostoc. sp. x ∆ x •
28. N. punctiforme (Kürtz.) Hariot. •
29. Anabaena sp. ∆ • x
30. Calothrix gracilis Fritsch ∆ ∆ ∆
31. C. parietina Thureter Born. at Flah. ∆ ∆ ∆ x x ∆
32. C. brevissima West, G.S x •
33. Tolypothrix conglutinata Borzi. ∆ x
34. Stigonema minutum (Ag.) Born. at Flah. ∆ • •
35. Scytonema sp. ∆ ∆
Total number of species occurrence 18 10 17 16 13 6 15 4
o: Dominant species of the community; •: Important species; ∆: Dominant but not as above; x: Just present
578 K.D. Pandey et al.
period from 15 Dec. to 15 Jan. Almost all thecyanobacterial species recorded at Nunatak were N2-fixersand N. commune was the important constituent of thecommunity (Table 1).
Schirmacher Oasis does not have any true soil. Soil inthe Oasis is of weathered rock and is still in "Soilformation" stage. The soil, low in nutrients pool coloniseda wide range of cyanobacteria on its moist surface. Thecontribution of N2-fixer was about 40% among the 15species recorded and N. commune was importantconstituent of the soil. The soil near moss community,supported the growth of non N2-fixers indicating itsnitrogen and organic rich nature. The quartz rocks althoughnon-abundant were ubiquitous in the Oasis. Theysupported minimum species diversity with a littlecyanobacterial activity in the form of patches on thesurface or between the ridges and curves (Table 1).
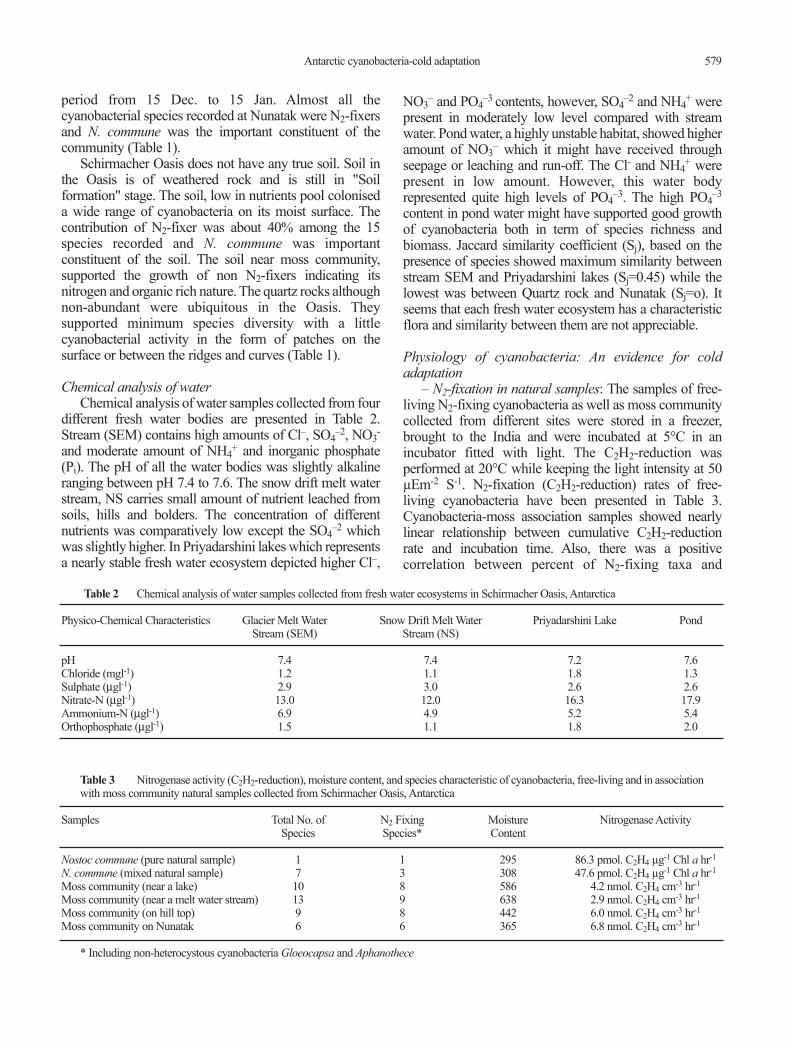
Chemical analysis of waterChemical analysis of water samples collected from four
different fresh water bodies are presented in Table 2.Stream (SEM) contains high amounts of Cl�, SO4
�2, NO3-
and moderate amount of NH4+ and inorganic phosphate
(Pi). The pH of all the water bodies was slightly alkalineranging between pH 7.4 to 7.6. The snow drift melt waterstream, NS carries small amount of nutrient leached fromsoils, hills and bolders. The concentration of differentnutrients was comparatively low except the SO4
�2 whichwas slightly higher. In Priyadarshini lakes which representsa nearly stable fresh water ecosystem depicted higher Cl�,
NO3� and PO4
�3 contents, however, SO4�2 and NH4
+ werepresent in moderately low level compared with streamwater. Pond water, a highly unstable habitat, showed higheramount of NO3
� which it might have received throughseepage or leaching and run-off. The Cl- and NH4
+ werepresent in low amount. However, this water bodyrepresented quite high levels of PO4
�3. The high PO4�3
content in pond water might have supported good growthof cyanobacteria both in term of species richness andbiomass. Jaccard similarity coefficient (Sj), based on thepresence of species showed maximum similarity betweenstream SEM and Priyadarshini lakes (Sj=0.45) while thelowest was between Quartz rock and Nunatak (Sj=o). Itseems that each fresh water ecosystem has a characteristicflora and similarity between them are not appreciable.
Physiology of cyanobacteria: An evidence for coldadaptation
� N2-fixation in natural samples: The samples of free-living N2-fixing cyanobacteria as well as moss communitycollected from different sites were stored in a freezer,brought to the India and were incubated at 5°C in anincubator fitted with light. The C2H2-reduction wasperformed at 20°C while keeping the light intensity at 50µEm-2 S-1. N2-fixation (C2H2-reduction) rates of free-living cyanobacteria have been presented in Table 3.Cyanobacteria-moss association samples showed nearlylinear relationship between cumulative C2H2-reductionrate and incubation time. Also, there was a positivecorrelation between percent of N2-fixing taxa and
Antarctic cyanobacteria-cold adaptation 579
Table 2 Chemical analysis of water samples collected from fresh water ecosystems in Schirmacher Oasis, Antarctica
Physico-Chemical Characteristics Glacier Melt Water Snow Drift Melt Water Priyadarshini Lake PondStream (SEM) Stream (NS)
pH 7.4 7.4 7.2 7.6Chloride (mgl-1) 1.2 1.1 1.8 1.3Sulphate (µgl-1) 2.9 3.0 2.6 2.6Nitrate-N (µgl-1) 13.0 12.0 16.3 17.9Ammonium-N (µgl-1) 6.9 4.9 5.2 5.4Orthophosphate (µgl-1) 1.5 1.1 1.8 2.0
Table 3 Nitrogenase activity (C2H2-reduction), moisture content, and species characteristic of cyanobacteria, free-living and in associationwith moss community natural samples collected from Schirmacher Oasis, Antarctica
Samples Total No. of N2 Fixing Moisture Nitrogenase ActivitySpecies Species* Content
Nostoc commune (pure natural sample) 1 1 295 86.3 pmol. C2H4 µg-1 Chl a hr-1
N. commune (mixed natural sample) 7 3 308 47.6 pmol. C2H4 µg-1 Chl a hr-1
Moss community (near a lake) 10 8 586 4.2 nmol. C2H4 cm-3 hr-1
Moss community (near a melt water stream) 13 9 638 2.9 nmol. C2H4 cm-3 hr-1
Moss community (on hill top) 9 8 442 6.0 nmol. C2H4 cm-3 hr-1
Moss community on Nunatak 6 6 365 6.8 nmol. C2H4 cm-3 hr-1
* Including non-heterocystous cyanobacteria Gloeocapsa and Aphanothece

580 K.D. Pandey et al.
cumulative C2H2-reduction rates (r= �0.462 + 0.009 x, r =0.96, p<0.01).
N. commune collected from soil near the pond waterwas nearly pure and consisted of single species. Theflattened thallus with 93% moisture content showed poorN2-fixation. Another sample of N. commune which wasintermingled with three other N2-fixing and four non N2-fixing species exhibited still poor rate of N2-fixation in thenatural sample. The moss community growing near aglacier melt water stream supported the growth of 13cyanobacteria species, among them nine werediazotrophic. The minimum of six species, all diazotrophicwere observed in moss community growing on Nunatak,while the moss community growing near a lake and on hilltop supported 10 and nine species, respectively. Althoughthe number of cyanobacteria taxa associated with mosscommunities was low, the relative proportion of N2-fixingspecies was quite high ranging between 70-100%. Themoss communities collected from different habitats haveexperienced different levels of freezing and thawing duringpolar summer and winter cycle. The water content variedsignificantly among the sites. The moss sample collectedfrom Nunatak with almost all the N2-fixing speciesreduced acetylene at a highest rate whereas the mosssample of stream showed lowest rate of N2-fixation.
� N2-fixation by pure axenic Antarctic isolates ofcyanobacteria: The measurement of N2-fixation and asearch for pure population of cyanobacteria under naturalAntarctic environment is tedious. It was therefore, desiredto isolate some of the strains under pure axenic culture inthe laboratory and to investigate N2-fixation of the strainsas influenced by temperature gradient. Number of studieson N2-fixation by natural samples and pure isolates ofcyanobacteria revealed that temperature optima ofAntarctic cyanobacteria for enzyme activity generallyremain lower than the temperate or tropical strains. It is,therefore, desired to take a similar strain of tropicalcyanobacteria as a reference to compare nitrogenaseactivity.
Antarctic (Ant.) strain of N. commune when compared
with tropical (Trop.) strain revealed that: a) the highest rateof nitrogenase activity was at 25°C while the temperatureoptima of tropical strain was 35°C; b) N2-fixation rate ofAnt. strain was low (76%) in comparison to Trop. strain,and c) nitrogenase activity continued at lower temperatureof 5°C in Ant. strain which was not evident in the Trop.strain. The activation energy derived from Arrehinius plotof temperature dependent nitrogenase activity was also lowin Ant. strain compared to Trop. strain. An Antarctic strainof Anabaena from Priyadarshini lake of Schirmacher Oasisalso performed N2-fixation at low temperature of 5°C asobserved in Ant. strain of N. commune which was notevident for Trop. strain of Anabaena. The Ant. strain of thiscyanobacterium showed its highest nitrogenase activity at15°C, a temperature 10°C lower than the temperatureoptima of Trop. strain. In continuation to this study N2-fixation was performed in a unicellular non-heterocystousaerobic diazotroph Gloeocapsa strain from mosscommunity. Unlike the above two strains the N2-fixationrate was comparable to that of Gloeocapsa species strainfrom a tropical rice field. The temperature optima of theAntarctic Gloeocapsa for N2-fixation was 20°C whereastropical species expressed its maximum activity at 30°C.However, activation energy was similar in both the strainsof Gloeocapsa (Table 4). The Antarctic strain ofGloeocapsa was able to fix nitrogen in the dark and sucroseas a carbon source promoted C2H2-reduction rate in theabsence of light.
Nitrate uptake and nitrate reductase activityNitrate transported across the cyanobacterial membrane
requires energy and reductant which is thought to bederived from photosynthesis. Nitrate uptake measured inTrop. and Ant. strains of Anabaena under stimulatedlaboratory conditions revealed that uptake of NO3
- ion waswell evident at lower temperature of 5°C and was mostsusceptible to thermal inactivation. Tropical strain ofAnabaena did not transport nitrate ion below 10°C andmaximum activity was shown at higher temperature. Theuptake of the nitrate ion by Ant. strain was comparativelylow (6.3 nM µg-1 protein ml-1) than the Trop. strain (6.5 nM
Table 4 Nitrogenase activity, temperature optima and activation energy in Antarctic strains of cyanobacteria and their respective tropicalreference strains
Cyanobacteria Nitrogenase Activity Temp. Optima (oC) Activation Energy(C2 H2-Reduction) (kJmol-1)
Antarctic Strains Tropical Strains Antarctic Strains Tropical Strains Antarctic Strains Tropical Strains
N. commune 6.6* 8.8* 25 35 33 52Anabaena sp. 15.2** 32** 15 25 32.4 56.3Gloeocapsa sp. 3.6* 4.0* 20 30 47.5 48.5
* nmol C2H4. µg-1 Chl a hr-1; ** nmol C2H4. mg-1 protein min-1

µg-1 protein ml-1) measured at their respective temperatureoptima. The temperature influenced nitrate reductase (NR)activity in both the strain, differed significantly withrespect to the thermal response. It is interesting to knowthat both, uptake and reduction of the nitrate ions in Ant.strain was initiated below 5°C which suggested a differentphysiological set up in Antarctic cyanobacteria. The NRactivity was nearly 81% in Ant. strain compared to Trop.strain and also, the activation energy was significantly low(64% of the Trop. strain).
NH4+ uptake and GS-transferase
NH4+ the most reduced form of nitrogen source enters
the cell via a well defined transport system in the Ant.strain of Anabaena at a rate of 9.4 nmol mg-1 protein min-
1 at its temperature optima of 20°C. Tropical strain at 30°C(temperature optima) transported NH4
+ ion at a slightlyhigher (9.6 nM mg-1 protein min-1) rate. The activationenergy of NH4
+ transport in Trop. strain was about ninepercent higher than the Ant. isolate (38.3 kJmol-1). TheGS-transferase activity determined in diazotrophic culturesof Ant. strain at 5°C was not evident in Trop. strain ofAnabaena.Ant. isolate exhibited nearly 13% lower activityof this enzyme compared to Trop. strain (0.99 µmol γ-glutamate hydroxylate mg-1 min-1) measured on theirrespective temperature optima of 20 and 30°C,respectively. However, there was wide difference of about70% higher activation energy in Trop. strain compared toAnt. strain of cyanobacteria (32.4 kJmol-1).
Photosynthetic activityThe influence of temperature on photosynthetic (PS-II)
activity of Ant. and Trop. strains of Anabaena wasmeasured after exposing them to a temp. of 5-40°C. Therates PS-II activity in Ant. and Trop. strain were 216 and148 µmol O2 mg-1 Chl a hr-1at their respective temperatureoptima of 20°C and 30°C. In contrast to trop. strain, the
Antarctic cyanobacteria-cold adaptation 581
Table 5 Effect of salinity* on the enzymes of nitrogenmetabolism in Antarctic and tropical strains of Anabaena
Enzymes NaCl Antarctic TropicalConcentration Strains Strains
Nitrogenase Control (- NaCl) 15.2 32IC-50 17.1 17.3
Nitrate reductase Control 6.2 8.6IC-50 3.2 4.2
GS-transferase Control 0.88 0.99IC-50 0.44 0.52
*IC-50 values of sodium chloride for nitrogenase, NR and GStransferase activities in Antarctic and tropical strain ofAnabaena was about 200 and 150 mmol., respectively.
higher level of PS-II activity in Ant. strain was aninteresting observation and it might play an important rolein the biomass production of cyanobacteria under coldstress.
Effect of salinity (NaCl) on nitrogen metabolism The activities of major nitrogen metabolism enzymes �
nitrogenase, nitrate reductase and GS-transferase � werecomparatively higher in Trop. strain of Anabaena ascompared to Ant. strain of Anabaena at their respectivetemperature optima in the absence of NaCl. Thenitrogenase activity in trop. strain was more than two timeshigher in the absence of NaCl. However, the activity of thisenzyme at their respective IC-50 level and temperatureoptima was quite comparable in both strains. The resultsrevealed a strong requirement of NaCl for the nitrogenfixation by the Ant. strain because of its adaptation to growat higher salinity level in the lakes of Oasis. Salinitysuppressed the nitrate reductase activity in both the strains,as about 50% reduction in the activity of this enzyme wasevident at their respective IC-50 level of NaCl salinity. Asimilar trend of reduction in GS-Transferase activity inboth the strains was also recorded at their respective K-50level. The results revealed a differential response for NaClsalinity by the different enzymes of nitrogen metabolism inAnt. strain (Table 5).
DISCUSSION
In view of the considerable number of summer meltwater streams, lakes and ponds in ice free areas (Oasis),most supporting rich growth of cyanobacteria, theirbiology deserves more detailed investigations. In constrastto Penninsular Antarctica, continental areas supportedrestricted growth of cyanobacteria in Oasis, however, theyare the most dominant component of the flora. The totalnumber of cyanobacteria species recorded in Oasis wasconsiderably lower than those reported from other regionsof Antarctica (6,20,36,37). The reason for such adiscrepancy is not clear, but it could be due to poor growthof moss species in Oasis, unaccessible sites due to difficultterrain and limited period of visits restricted to polarsummer only. Nevertheless, contribution of diazotrophicspecies (more than half of total number of cyanobacteriarecorded at any sites) revealed the nutrient depletedconditions of fresh water and terrestrial ecosystems inOasis. Cyanobacteria have a great potential to survive inharsh climatic conditions of Antarctica, since they have thepotentiality to fix carbon (present in the form of CO2) andmolecular dinitrogen besides mobilizing inorganicphosphate (PO4
�3) from organic phosphates (55).Environmental factors regulating phototrophy(Photosynthesis), heterotrophy and diazotrophic (N2-fixation) are the intensity of radiant energy, liquid water, N,

P and trace elements (31). The metabolic exchangesexisting between phototrophs and heterotrophs in ice-freeregion appear to be a limiting factor and to accomplish thismutual phenomenon the microbes must exist in a highlyco-operative and efficient manner to survive the extremecold of Antarctic environment.
The microbes Azotobacter, methanogenic andmethanotrophic bacteria nitrifying and denitrifyingbacteria, along with the cyanobacteria in free living statewere encountered in bottom layer soil of aquatic ecosystemand on the weathered rock soils. The presence of otherheterotrophs include Micrococcus, Planococcus andPseudomonas sp. (48,49). Cyanobacteria may form a closeassemblage with moss species and with fungi in lichen.Thus a temporal association exists between phototrophsand heterotrophs in Schirmacher Oasis. In majority of thecases cyanobacterial activity were associated with theaggregates and the presence and dominance of individualspecies was rare except N. commune, a well knowncyanobacteria tolerant to stress in most of the extremeconditions.
At the bottom of the Priyadarshini lake in theSchirmocher Oasis, Antarctica, 90 cm thick cyanobacterialbiomass, the thickest microbial biofilm to be found innatural environment, was dominated by Oscillatoriancyanobacteria (21). Mats constituted mainly ofOscillatoria, Phormidium and N. commune similar to thoseobserved by Ellis-Evans and Bayliss (13) for Antarcticstreams (47,53). One of the most striking phenomenon wasthat Chaemosiphon sp. grew on the small rocks ofrestricted area in a SEM stream in Schirmacher Oasis,which is not evident in other aquatic ecosystems.Oscillatorian cyanobacteria are often found as a majorelement of cyanobacterial diversity in the lakes of Oasis asrecorded in Dry Valley lakes (55). Bliss and Gold (2)documented the role of N2-fixation by cyanobacteriawhich permits the colonization of other microorganismsand higher plants on the soils and exposed moraines. Ascompared to well studied microbial communities in SignyIsland (60°S) (4,5), little is known about the microbialcolonization of soils and rocks in Schirmacher Oasis(70°S). The biodiversity of cyanobacteria in polar region isof considerable debate and uncertainty (54) due totaxonomic criteria available in the literatures. Humaninterference and sample collection by scientists are themajor threat for biodiversity of cyanobacteria inSchirmacher Oasis.
The cyanobacteria colonising lakes and streams may beconsidered as the "life support system" of Antarctic polardesert. In McMurdo Dry Valley, heterotrophic bacteriawere attached to the soil particles and associated withcyanobacterial aggregates (30). When melting of the iceduring polar summer is localized, O2 consumptionassociated with the cyanobacterial aggregate is sufficient to
create reduced microsomes. These microsomes may be thepotential site for O2 sensitive process such as N2-fixation(29).
The heterotrophic populations most widely investigatedin Sydney ice land, was so low that Christie (10)considered their contribution to nitrogen fixation and N-cycle as insignificant. Similar findings have been alsoreported in Schirmacher Oasis (48,49). Pandey, Kashyapand Gupta (33) also did not observe any rate of C2H2-reduction by aerobic or anaerobic heterotrophs associatedwith moss communities collected from Oasis. However,the unicellular phototrophic diazotroph Gloeocapsaspecies showed nitrogenase activity in the dark which wasstimulated by the addition of organic carbon source,sucrose (34). These observations imply that heterotrophicN2-fixation does prevail in Oasis but mostly by thephototrophic organisms.
Several studies on natural samples in field and isolatedstrains in laboratory revealed that low ambienttemperatures may not completely inhibit metabolism andgrowth of Antarctic cyanobacteria. Photosynthesis andN2-fixation increased up to 15-25°C (54) and isolatedclones were unable to grow at temperatures less than orequal to 5°C which implies that these organismsoriginated from warmer temperate latitudes (45). Thevarious metabolic functions as photosynthesis, N2-fixation, NO3
� uptake and reduction, NH4+ uptake and its
further metabolism at 5°C was not considerable in tropicalstrains. Also the temperature optima for various metabolicfunctions was in the range of 15-25°C, nearly 10°C lessthan their respective Trop. strain. It may be concluded thatcyanobacteria colonizing in Schirmacher Oasis areadapted to lower temperature for metabolic functions.However, a detailed genomic analysis and biochemistry ofcold tolerance is necessary to reach any acceptableconclusion. Most of isolated strains of AntarcticOscillatorian cyanobacteria have temperature optima forgrowth in the range of 15-35°C with a insignificantgrowth at 5°C (52). Although, these strains were notadapted genetically to low temperatures, they were likelyto be tolerant to the variable temperature regimes of theirenvironment.
The energy of activation for different metabolicfunctions in isolated Antarctic strains was considerably lowwhich reflect the lesser endergonic nature of themetabolism in Antarctic cyanobacteria. This seems to be anoutcome of constant selection pressure imposed by thecold stress of the Antarctic environment.
Acknowledgments � We thank Head, Department of Botany, BanarasHindu University, Varanasi for providing laboratory facilities; Secretary,Department of Ocean Development (DOD), Govt. of India, for logisticsupport during Scientific Expeditions to Antarctica, University GrantsCommission (UGC) and Council of Scientific and Industrial Research(CSIR), New Delhi for financial support.
582 K.D. Pandey et al.

REFERENCES
1. Becker, E.M., Physiological studies on Antarctic Prasiola crispaand Nostoc commune at low temperatures. Polar Biol. 1982, 1: 99-104.
2. Bliss, I.S. and Gold, W.G., The patterning of plant communities andedaphic factors along the high Arctic coastline implications forsuccession. Can. J. Bot. 1994, 72: 1095-1107.
3. Bourrelly, P., Les Algues d�can douce. Initiation a la system atique3. Les Algues bleues et rouges. Boubee and Cie, Paris, 1970, p. 572.
4. Broady, P.A., Diversity, distribution and dispersal of Antarcticterrestrial algae. Biodiver. Conserv. 1996, 5: 1307-1335.
5. Broady, P.A., Taxonomy and ecology of algae in a fresh waterstream in Taylor Valley, Victoria Land, Antarctica. Arch. Hydrobiol.Suppl. 1982, 63: 331-349.
6. Broady, P.A., The Signy Island terrestrial reference sites. IX. Theecology of the algae of site 2, a moss carpet. Br. Antarct. Sury. Bull.1979, 47: 13-29.
7. Broady, P.A., Broad scale patterns in the distribution of aquatic andterrestrial vegetation at three ice free regions on Ross Island,Antarctica. Hydrobiologia 1989, 172: 77-95.
8. Burris, R.H. and Wilson, P.Q., Methods for measurement ofnitrogen fixation. In: Methods in Enzymology, Colovick, S.P. andKaplan, N.O. (eds.), Acad. Press, London, 1957, 4: 355-366.
9. Carpenter, E.J., Lin, S. and Capone, D.G., Bacterial activity in SouthPole snow. Appl. Environ. Microbiol. 2000, 66: 4514-4517.
10. Christie, P., Nitrogen in two contrasting Antarctic bryophytecommunities. J. Ecol. 1987, 75: 93.
11. Davey, A. and Merchant, H.T., Seasonal variation in nitrogenfixation by Nostoc commune Vaucher at the vest-fold hills,Antarctica. Phycologia 1989, 22: 377-385.
12. Desikachary, T.V., Cyanophyta, Indian Council of Agril. Research,New Delhi, India, 1959, p. 686.
13. Ellis-Evan, J.C. and Bayliss, P.R., Biologically active microgradientin cyanobacterial mats of Antarctic lakes and streams. Verh. Intl.Vereina Limnol. 1993, 25: 948-952.
14. Fiske, C.H. and Subbarow, Y., Colorimetric determination ofphosphorous. J. Biol. Chem. 1925, 66: 375-400.
15. Fogg, G.E. and Stewart, W.D.P., In situ determination of biologicalnitrogen fixation in Antarctica. Br. Antarct. Sury. Bull. 1968, 15: 39-46.
16. Fritsen, C.H. and Priscu, J.C., Cyanobacterial assemblages inpermanent ice covers of Antarctic lakes: distribution, growth rateand temperature response of photosynthesis. J. Phycol. 1998, 34:587-597.
17. Geitler, L., In: Kryptogamen Flora von Deutschland, Osterreichunder Schweiz, Robenhorst, L. (ed.), Akademische VerlagGesellschaft, Leipzig, 1932, p. 14.
18. Herbert, D., Phipps, P.I. and Strange, R.E., Chemical analysis ofmicrobial cells. In: Methods in Microbiology, Norris, J.R. andRibbons, D.W. (eds.), Acad. Press, London, 1971, Vol. VB, pp. 209-344.
19. Hermichen, W.D., Kowski, P. and Stranch, G., The isotopeglaciological situation in the surroundings of SchirmacherOasis/Dronning Maud Land. A first overview. ZFT-Mittelungen1984, 89: 87-102.
20. Howard-Williams, C., Vincent, C.L., Broady, P.A. and Vincent,W.F., Antarctic stream ecosystems: variability in environmentalproperties and algal community structure. Int. Rev. Ges. Hydrobiol.1986, 71: 511-544.
21. Ingole, B.S. and Parulekar, A.H., Limnology of Priyadarshini lakeSchirmacher Oasis, Antarct. Polar Res. 1990, 26: 13-17.
22. Jaccard, P., Nouvelles recherches sur la distribution florale. Bull.Vaudoise Sci. 1908, 44: 223-270.
23. Kashyap, A.K., Gupta, R.K. and Pandey, K.D., Check list ofcyanobacteria occuring in Schirmacher oasis, Antarctica. J.Scientific Res. 1998, 47: 171-179.
24. Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J., Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem. 1951,193: 265-275.
25. Myers, I. and Kratz, W.A., Relation between pigment content andphotosynthetic characteristics in a blue-green alga. J. Gen. Physiol.1955, 39: 11-12.
26. Nadeau, T. and Castenholz, R.W., Evolutionary relationships ofcultivated Antarctic Oscillatorians (cyanobacteria). J. Phycol. 2001,37: 650-654.
27. Nakatsubo, T. and Ino, Y., Nitrogen cycling in an Antarcticecosystem. Estimation of the amount of nitrogen fixation in a mosscommunity on East Ongul Island: Ecol. Res. 1987, 2: 31-40.
28. Nicholas, D.J.D. and Nason, A., Determination of nitrate and nitrite.In: Methods in Enzymology, Colowick, S.P. and Kaplan, N.O.(eds.), Acad. Press, London, 1957, pp. 981-984.
29. Olson, J.B., Steppe, T.F., Litaker, R.W. and Paerl, H.W., N2-fixingmicrobial consortia associated with the ice cover of lake Bonney,Antarctica. Microbial Ecol. 1998, 36: 231-238.
30. Paerl, H.W. and Pinckney, J.L., Ice aggregates as a microbial habitatin Lake Bonney, dry valley lakes, Antarctica: Nutrient-rich micro-ozones in an oligotrophic ecosystem. Antarct. J. USA 1996, 31: 220-222.
31. Paerl, H.W. and Priscu, J.C., Microbial phototrophic, heterotrophicand diazotrophic activities associated with aggregates in thepermanent ice cover of lake Bonney, Antarctica. Microbiol Ecol.1998, 36: 221-230.
32. Pandey, K.D. and Singh, P.K., Isolation and characterization ofnitrate reductase mutants and regulation of nitrate reductase andnitrogenase in the cyanobacterium Nostoc muscorum. Mol. Gen.Genet. 1984, 195: 180-185.
33. Pandey, K.D., Kashyap, A.K. and Gupta, R.K., Nitrogen fixation bycyanobacteria associate with moss communities in SchirmacherOasis. Antarctica. Israel J. Bot. 1992, 41: 187-198.
34. Pandey, K.D., Kashyap, A.K. and Gupta, R.K., Nitrogen fixation bynon-heterocystous cyanobacteria in an Antarctic ecosystem. IsraelJ. Plant Sci., 2000, 48: 267-270.
35. Pandey, K.D., Kashyap, A.K. and Gupta, R.K., Nutrient Status,algal and cyanobacterial flora of six fresh water streams ofSchirmacher Oasis, Antarctica. Hydrobiology 1995, 299: 83-91.
36. Pankow Von, H., Haendel, D., Richter, W. and Wand, U.,Algologische Beobachtungen in der Schirmacher andUntereseeoase (Dronning-Maud-Land, Ostantarktika). Arch.Protistenkd. 1987, 134: 59-82.
37. Parker, B.C., Samsell, G.L. and Prescott, G.W., Freshwater algae ofthe Antarctic Peninsula. I. Systematics and ecology in the U.S.Palmer Station area. In: Antarctic Terrestrial Biology, Llano, G.(ed.), Antarctic Research Series, American Geophysical Union,Washington DC, 1972, 20: 69-81.
38. Prasad, P. and Kashyap, A.K., Ammonium transport in thealkalophilic diazotrophic cyanobacterium Nostoc calcicola:influence of phosphate limitation and metabolic inhibitors. J. PlantPhysiol. 1991, 138: 244-247.
39. Priddle, J. and Belcher, J.H., An annotated list of benthic algae(excluding diatoms) from freshwater lakes on Signy Island. Br.Antarct. Surv. Bull. 1982, 57: 41-53.
40. Priscu, J.C., Adams, E.E., Lyons, W.B., Voytek, M.A., Mogk, D.W.,Brown, R.L., McKay, C.P., Takacs, C.D., Welch, K.A., Wolf, C.F.,Kirstein, J.D. and Avci, R., Geomicrobiology of sub-glacial iceabove Vostok Station. Science 1999, 286: 2141-2144.
41. Priscu, J.C., Ecosystem dynamics in a polar desert: The McMurdoDry Valleys, Antarctica. Antarct. Res. Series 1998, 72: 1-369.
42. Rand, M.C., Greenberg, A.E., Taras, M.J. and Franson, M.E.,
Antarctic cyanobacteria-cold adaptation 583

584 K.D. Pandey et al.
Standard methods for the examination of water and waste water.American Public Health Association, Amm. Water Works Assoc.,Water Pollut Contr. Fed., p. 1193.
43. Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M. and Stanier,R.Y., Genetic assignments, strain histories and properties of purecultures of cyanobacteria. J. Gen. Microbiol. 1979, 111: 1-61.
44. Schollhorn, R. and Burris, R.H., Acetylene as competitive inhibitorof N2-fixation. Proc. Natl. Acad. Sci. USA 1967, 58: 213-216.
45. Seaburg, K.G., Parker, B.C., Wharton, R.A. Jr. and Simmons, G.M.,Temperature-growth response of algal isolates from Antarctic Oasislakes. J. Phycol. 1981, 17: 353-360.
46. Shapiro, B.M. and Stadtman, E.R., The regulation of glutaminesynthetase in microorganisms. Annu. Rev. Microbiol. 1970, 24: 501-524.
47. Sheath, R.G. and Muller, K.M., Distribution of stream macroalgaein four high Arctic drainage basins. Arctic. 1997, 50: 355-364.
48. Shivaji, S., Rao, N.S., Saishree, L., Seth, V., Reddy, G.S.N. andBhargava, P.M., Isolation and identification of Pseudomonas spp.from Schirmacher Oasis, Antarctica. Appl. Environ. Microbiol.1989, 55: 767-770.
49. Shivaji, S., Rao, N.S., Saishree, L., Seth, V., Reddy, G.S.N. andBhargava, P.M., Isolation and identification of Micrococcus roseusand Planococcus spp. from Schirmacher Oasis, Antarctica. J. Biol.Sci. 1988, 13: 401-414.
50. Shukla, S.P., Pandey, K.D. and Kashyap, A.K., Nitrogen fixation,ammo. transport and glutamine synthetase activity in an AntarcticCyanobacterium Anabaena sp.: Influence of temp. J. Plant Physiol.1997, 150: 351-354.
51. Snell, F.D. and Snell, C.T., Nitrite by sulphanilamide and N-(1-naphthyl) ethylene diamine hydrochloride. In: ColorimetricMethods of Analysis, D. Van Nostrand, New Jersey, New York,USA, 1949, 2: 804-805.
52. Tang, E.P.Y.,Tremblay, R. and Vincent, W.F., Cyanobacterialdominance of polar freshwater ecosystems: Are high latitude-mat-formers adapted to low temperature? J. Phycol. 1997, 33: 171-181.
53. Vezina, S. and Vincent, W.F., Arctic cyanobacteria and limnologicalproperties of their environment: Bylot Island: Northwest Territories,Canada (73°N, 80°W). Polar Biol. 1997, 17: 523-534.
54. Vincent, W.F., Cyanobacterial dominance in the polar regions. In:The Ecology of Cyanobacteria: Their Diversity in Time and Space,Whitton, B.A. and Potts, M. (eds.), Kluwer, The Netherlands, 2000,pp. 321-340.
55. Vincent, W.F. and James, M.R., Biodiversity in extreme aquaticenvironments: lakes ponds and streams of the Ross Sea Sector,Antarctic. Biodiver. Conserv. 1996, 55: 1451-1471.
56. Whitton, B.A., Soils and rice fields. In: The Ecology ofCyanobacteria: Their Diversity in Time and Space, Whitton, B.A.and Potts, M. (eds.), Kluwer, The Netherlands, 2000, pp. 233-255.
Related Documents











