Diversity of nitrogen-fixing cyanobacteria under various ecosystems of Thailand: I. Morphology, physiology and genetic diversity Neung Teaumroong 1 , Sasidhorn Innok 1 , Somporn Chunleuchanon 2 and Nantakorn Boonkerd 1, * 1 School of Biotechnology, Institute of Agricultural Technology, Suranaree University of Technology, Nakhon Ratchasima 30000, Thailand 2 Department of Soil Sciences, Faculty of Agriculture, Chiang Mai University, Chiangmai 50202, Thailand *Author for correspondence: Tel.: +66-44-224751, Fax: +66-44-224750, E-mail: [email protected] Received 28 December 2001; accepted 7 May 2002 Keywords: Biodiversity, Cyanobacteria, DNA fingerprint, N 2 -fixation Summary The diversity among 853 isolates of nitrogen-fixing cyanobacteria obtained from soil samples collected from different ecosystems including mountainous, forest and cultivated areas in the central, northern and northeastern regions of Thailand was examined. Most isolates showed slow growth rate and had filamentous, heterocystous cells. The percentage of heterocysts in the filaments of different isolates varied from 8.3 to 9.6. Only a few strains showed high nitrogen-fixing potential, while most of the strains exhibited low capacity for nitrogen fixation. Anabaena and Nostoc were the dominant genera among these isolates. One hundred and two isolates were randomly selected from this diverse collection to determine the extent of genetic diversity on the basis of DNA fingerprinting using the PCR method. Based on the PCR products obtained by using a combination of three primers, all strains could be distinguished from one another. When a subset of 45 isolates of Nostoc and a subset of 44 isolates of Anabaena were further analysed by PCR, a wide range of diversity was observed within each of these genera. Introduction Cyanobacteria have inhabited much of the surface of the earth for billions of years (Mazel et al. 1990). They are an ancient group of prokaryotic microorganisms exhibiting the general characteristics of Gram-negative bacteria (Rai 1990). They were presumably the first oxygen-evolving photosynthetic organisms during the Precambrian era and are thought to be responsible for the transition of the atmosphere of the earth from its primordial anoxygenic state to the current atmospheric condition (Fogg et al. 1973). They can be found in almost all environments, including freshwater, seawater, non-acidic hot springs and deserts. Some cyanobacteria also have the ability to fix atmospheric nitrogen, yet relatively little is known about ecology of natural populations and the diversity of nitrogen-fixing cyano- bacteria. One of the difficulties in examining natural popula- tions is that nitrogen-fixing microbial communities could be complex mixtures of bacteria and cyanobacte- ria. Problems with identification of nitrogen-fixing cyanobacteria in nature include distinguishing different species, which rely upon identifying morphological, developmental and biochemical characteristics (Hiroki et al. 1998). Considerable expertise is required to iden- tify cyanobacteria at the species level, although in some cases it is not difficult to identify the genera. Recently, molecular genetic approaches have been used for detec- tion and characterization of cyanobacteria (Porath & Zehr 1994), identification and phylogenetic analysis of toxigenic cyanobacteria (Neilan 1995) and phenotypic and genotypic comparison of symbiotic and free-living cyanobacteria from a single field site (West & Adams 1997). The DNA fingerprinting methods using the PCR reaction, such as randomly amplified polymorphic DNA (RAPD) are extensively used nowadays to classify and group isolates. DNA amplification fingerprinting (DAF) is able to group the Azolla symbiotic cyanobacteria (Eskew et al. 1993). Furthermore, enterobacterial repeti- tive intergenic consensus (ERIC) and repetitive extra- genic palindromic (REP) sequences have been used as primers for fingerprinting axenic cyanobacterial cultures (Lupski & Weinstock 1992). Moreover, short tandemly repeated repetitive sequences (STRR) and highly itera- ted palindromic sequences (HIP1) had been shown to distinguish symbiotic Nostoc and Anabaena in cycads (Rasmussen & Svenning 1998) and many cyanobacteria like Synechococcus sp. and some toxin-producing cyanobacteria, respectively (Robinson et al. 1995; Rouhiainen et al. 1995). On the other hand, to deter- mine the diversification and confirm the N 2 -fixing World Journal of Microbiology & Biotechnology 18: 673–682, 2002. 673 ȑ 2002 Kluwer Academic Publishers. Printed in the Netherlands.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Diversity of nitrogen-fixing cyanobacteria under various ecosystems of Thailand:
I. Morphology, physiology and genetic diversity
Neung Teaumroong1, Sasidhorn Innok1, Somporn Chunleuchanon2 and Nantakorn Boonkerd1,*1School of Biotechnology, Institute of Agricultural Technology, Suranaree University of Technology, NakhonRatchasima 30000, Thailand2Department of Soil Sciences, Faculty of Agriculture, Chiang Mai University, Chiangmai 50202, Thailand*Author for correspondence: Tel.: +66-44-224751, Fax: +66-44-224750, E-mail: [email protected]
Received 28 December 2001; accepted 7 May 2002
Keywords: Biodiversity, Cyanobacteria, DNA fingerprint, N2-fixation
Summary
The diversity among 853 isolates of nitrogen-fixing cyanobacteria obtained from soil samples collected fromdifferent ecosystems including mountainous, forest and cultivated areas in the central, northern and northeasternregions of Thailand was examined. Most isolates showed slow growth rate and had filamentous, heterocystous cells.The percentage of heterocysts in the filaments of different isolates varied from 8.3 to 9.6. Only a few strains showedhigh nitrogen-fixing potential, while most of the strains exhibited low capacity for nitrogen fixation. Anabaena andNostoc were the dominant genera among these isolates. One hundred and two isolates were randomly selected fromthis diverse collection to determine the extent of genetic diversity on the basis of DNA fingerprinting using the PCRmethod. Based on the PCR products obtained by using a combination of three primers, all strains could bedistinguished from one another. When a subset of 45 isolates of Nostoc and a subset of 44 isolates of Anabaena werefurther analysed by PCR, a wide range of diversity was observed within each of these genera.
Introduction
Cyanobacteria have inhabited much of the surface ofthe earth for billions of years (Mazel et al. 1990). Theyare an ancient group of prokaryotic microorganismsexhibiting the general characteristics of Gram-negativebacteria (Rai 1990). They were presumably the firstoxygen-evolving photosynthetic organisms during thePrecambrian era and are thought to be responsible forthe transition of the atmosphere of the earth from itsprimordial anoxygenic state to the current atmosphericcondition (Fogg et al. 1973). They can be found inalmost all environments, including freshwater, seawater,non-acidic hot springs and deserts. Some cyanobacteriaalso have the ability to fix atmospheric nitrogen, yetrelatively little is known about ecology of naturalpopulations and the diversity of nitrogen-fixing cyano-bacteria.One of the difficulties in examining natural popula-
tions is that nitrogen-fixing microbial communitiescould be complex mixtures of bacteria and cyanobacte-ria. Problems with identification of nitrogen-fixingcyanobacteria in nature include distinguishing differentspecies, which rely upon identifying morphological,developmental and biochemical characteristics (Hirokiet al. 1998). Considerable expertise is required to iden-
tify cyanobacteria at the species level, although in somecases it is not difficult to identify the genera. Recently,molecular genetic approaches have been used for detec-tion and characterization of cyanobacteria (Porath &Zehr 1994), identification and phylogenetic analysis oftoxigenic cyanobacteria (Neilan 1995) and phenotypicand genotypic comparison of symbiotic and free-livingcyanobacteria from a single field site (West & Adams1997). The DNA fingerprinting methods using the PCRreaction, such as randomly amplified polymorphic DNA(RAPD) are extensively used nowadays to classify andgroup isolates. DNA amplification fingerprinting (DAF)is able to group the Azolla symbiotic cyanobacteria(Eskew et al. 1993). Furthermore, enterobacterial repeti-tive intergenic consensus (ERIC) and repetitive extra-genic palindromic (REP) sequences have been used asprimers for fingerprinting axenic cyanobacterial cultures(Lupski & Weinstock 1992). Moreover, short tandemlyrepeated repetitive sequences (STRR) and highly itera-ted palindromic sequences (HIP1) had been shown todistinguish symbiotic Nostoc and Anabaena in cycads(Rasmussen & Svenning 1998) and many cyanobacterialike Synechococcus sp. and some toxin-producingcyanobacteria, respectively (Robinson et al. 1995;Rouhiainen et al. 1995). On the other hand, to deter-mine the diversification and confirm the N2-fixing
World Journal of Microbiology & Biotechnology 18: 673–682, 2002. 673� 2002 Kluwer Academic Publishers. Printed in the Netherlands.

capability on the basis of genotypic and phenotypicproperties, nifH profiles and acetylene reduction assay(ARA) have been extensively conducted.Since the ecological and agricultural importance of
many cyanobacteria depends on their ability to fixnitrogen, which determines their suitability for use asbiofertilizers instead of chemical fertilizers, this workwas aimed at identifying indigenous nitrogen-fixingcyanobacteria based on morphology and N2-fixingefficiency, and investigating their genetic diversity byusing molecular genetic methods.
Materials and methods
Cyanobacterial strain enumeration and growthmeasurement
The cyanobacterial strains used as reference strainswere: Nostoc sp., Anabaena cylindrica, Hapalosiphon sp.DASH 05101, Calothrix sp. DASH 02101 and Scyto-nema sp. which were obtained from the Department ofSoil Sciences, Faculty of Agriculture, Chiangmai Uni-versity, Thailand. Soil samples from mountain areas,flat areas of agricultural cultivation (field crop cultiva-tion, rice cultivation, rice in rotation with other crops)and uncultivated areas from northern, Central andnortheastern parts of Thailand were chosen as samplingsites during the study period (1997–1999). Fifty gramsof soil samples were mixed and shaken in 450 mldistilled water, then 1 ml was diluted by the 10-foldserial dilution technique. Each dilution was cultured inBG110 liquid medium (without sole nitrogen source)(Richmond 1986) and incubated under a 12 h/12 hlight/dark cycle with an average light irradiance of400 lE/s1/m2 about 25–30 days. The population num-ber in the growth tubes was determined according tothe standard MPN method (Grant et al. 1985). Thegrowth of cyanobacterial strains was determined bymeans of dry weight after 12 days of cultivation. Cellpellets were filtered through Whatman no. 42 filterpaper and then rinsed with deionized water and dried at80 �C for 2 h before being kept overnight in a des-iccator. To enumerate the number of special cell typessuch as akinete, heterocyst or vegetative cells, the cellpellet was treated by homogenizer for making randomdispersion prior to counting in a Petroff–Haussercounting chamber.
Identification of cyanobacteria
Colony-forming characteristics of each cyanobacterialisolate were observed and recorded as the data for thenext analyses using a 400· magnification microscope.The results of cell morphology such as colony form,colour, size of vegetative cell, heterocysts and theirlocalization, and akinete cells were interpreted followingDesikachary (1985) and Rippka (1988).
Acetylene reduction assay and measurementof chlorophyll-a
The 50 ml tubes containing cyanobacterial cultures with25 ml of BG110 liquid medium were incubated at 25 �Cunder a 12 h/12 h light/dark cycle. After 30 days ofcultivation, tubes were plugged with a double septumbefore 10% of the head space air volume was replacedwith acetylene gas. The incubation proceeded at 25 �Cfor 1 h under 400 lE/s1/m2 light intensity. One millilitreof gas mixture was withdrawn and analysed by gaschromatograph (GC) equipped with a Porapak Ncolumn (pack column no. 530-5014, Auto System XL,Perkin Elmer, USA). Ethylene production per tube perhour was determined by comparing the peak height withthat of the known amount of standard ethylene.Efficiency of N2-fixing cyanobacteria was comparedwith the amount of chlorophyll-a extracted according tothe method of Wintermans & Demots (1965).
DNA preparation from cyanobacterial isolates
Cyanobacterial strains were cultured in 100 ml Erlen-meyer flask containing 25 ml of BG110 liquid medium.Cell pellets were harvested by centrifugation at9000 rev/min for 10 min, resuspended with 3 ml ofextraction buffer (100 mM Tris–HCl [pH 8.0], 250 mMNaCl, 100 mM EDTA and 0.4% 2-mercaptoethanol),then 1 mg lysozyme/1 ml TE buffer (10 mM Tris–HCl[pH 8.0] and 1 mM EDTA) was added and incubated at37 �C for 1 h with occasional shaking. Two hundredmicrolitre of 1% (w/v) Sarcosyl was added, vortexedand extracted twice with an equal volume of phenolsolution. The aqueous phase was transferred into ananother microcentrifuge tube, an equal volume ofCTAB stock (2% CTAB in 1.4 M NaCl) was addedbefore being incubated at 65 �C for 30 min and thenextracted with equal volume of chloroform/isoamylalcohol (24:1, v/v). DNA was precipitated overnightwith 2.0 volumes of cold isopropanol. The solutionswere centrifuged before being washed with 70% ethanol,then dried and the DNA pellet was resuspended in TEbuffer containing 1/10 RNase A. The DNA solution wasincubated at 55 �C for 10 min prior to further analysis.
Oligonucleotide primers and PCR amplification
Standard PCR reactions were carried out in a finalvolume of 50 ll which included 50 ng of sample DNA,200 lM of each of dATP, dCTP, dGTP and dTTP, 1–2 pmol of each specific primer, 1.5 mM MgCl2 and2.5 U of Taq polymerase (Promega, Madison, W1).Thermal cycling was done as follows for 30 cycles:denaturation for 1 min at 93 �C, primer annealing at50 �C for 1 min and extension at 70 �C for 1.5 min. Theforward and reverse primers used for nifH amplificationwere primer 1:5¢-GGAATTCCTGYGAYCCNAARGCNGA-3¢ and primer 2:5¢-CGATCCGDNGCCATCA-TYTCNCC-3¢ (Y was T or C; N was A, C, G or T; R
674 N. Boonkerd et al.

was A or G and D; A, G or T), respectively (Porath &Zehr 1994), while those for STRR fragment (Rasmussen& Svenning 1998) were 5¢-CCARTCCCCARTCCCC-3¢. The cycles were: 1 cycle at 95 �C for 6 min; 30 cyclesof 94 �C for 1 min, 56 �C for 1 min and 65 �C for 5 min;1 cycle at 65 �C for 16 min and final step at 4 �C.Another set of DNA amplification fingerprint primerswas originally used to amplify Anabaena sp. in Azolla sp.DNA (Eskew et al. 1993). Sequences [5¢–3¢] wereGCTGGTGG (DAF 8.7b) and GTGACGTAGG(DAF 10.6e). Thermal cycling was done for 30 cyclesof 30 s at 95 �C, 1 min at 30 �C and 1 min at 72 �C,followed by a final extension period of 10 min at 72 �Cand final step at 4 �C. The PCR reagents were purchasedfrom GibcoBRT�, Brazil. All amplification conditionswere performed in PCR Sprint Temperature CyclingSystem (Hybaid Limited, UK). After the amplification,15 ll aliquots of the PCR products were resolved by gelelectrophoresis at 80 V/cm in 2.0% agarose gel stainedwith the ethidium bromide.
PCR amplification on intact filaments
The method was partially modified from Rasmussen &Svenning (1998). A single cyanobacterial colony waspelleted by centrifugation and washed twice in TEbuffer. The pellet was suspended in an appropriatevolume of TE buffer to ensure that at least a fewfilaments or cells were present in the 1–2 ll which wasused directly as a template for PCR as described above.All PCRs were compared with the results from large-scale (25 ml of culture) DNA extraction.
Phylogenetic analysis
A dendrogram was constructed from the similaritymatrix by the unweighted pair group method witharithmetic mean (UPGMA). In order to test thegoodness-of-fit of cluster analysis, cophenetic valuematrices were calculated and compared with the originalsimilarity metrics that were UPGMA clustered by usingthe NTSYS-pc package (version 1.8; Exeter Software,Setauket, N.Y.).
Results
Morphological diversity and identification
A total of 853 N2-fixing cyanobacterial strains wereobtained on the basis of growth under N2-free cultiva-tion. All isolates were found to be heterocystous andfilamentous. Anabaena sp. and Nostoc sp. were the mostdominant indigenous N2-fixing strains in every part ofThailand. The second most common cyanobacterialspecies in these soils were Hapalosiphon sp. and Scyto-nema sp. The rest of identified strains were Stigonemasp., Tolypothrix sp., Fischerella sp., Nodularia sp.,Anabaenopsis sp. and Cylindrospermum sp. Interestingly,
both Stigonema sp. and Cylindrospermum sp. werefound only in the northeastern part of the country.However, identification using conventional classificationwas unable to clarify all strains (Table 1). Only 11genera were classified while 132 out of 853 isolates stillremain to be identified. The size of colonies on solidmedium varied from 0.25 to more than 1.0 cm. Manyforms of colonies such as circular, irregular, filamen-tous, hairy and gas-forming were also found. Elevationsof colonies were flat and convex. The edge characteris-tics were entire, enrose, filamentous and curled. Exam-ples of colony-forming characteristics and morphologyused for identification are depicted in Figure 1.Among the Nostoc sp. strains, the morphology of
heterocysts was variable; 50% were spherical, 44% wereellipsoidal and 6% were in quadrate form. The mor-phology of the Anabaena sp. group was similar to that ofNostoc sp. except that some strains developed gas-forming colonies. The morphology of heterocysts wasmostly ellipsoidal and trichome, and they were arrangedin formal filaments. In contrast, Nostoc sp. wereirregular filamentous including heterocyst cells whichwere unaggregated on filament. In the putative Ana-baenopsis sp. group, all the strains were brownish incolour and filaments were spirally coiled. For theNodularia sp. group, colonies on solid medium weredistinct from Nostoc sp. and Anabaena sp. due to thepresence of hairy colonies, quadrate form heterocysts,and relatively wide and short vegetative cells. Thegrowth of the branching group was slower than othercyanobacteria and some of these strains had hairycolonies.Morphological characteristics of all isolates, the
percentage numbers of akinete and percentage numberof heterocyst/vegetative cell were investigated (Table 2).The range of percentage akinete cells was divided intothree levels; high range (6–9 akinete cells/ml), middlerange (4–6 akinete cells/ml) and low range (2–4 akinetecells/ml). The percentage of akinetes among all strains inThailand indicated that the cyanobacterial population
Table 1. Summary of soil N2-fixing cyanobateria isolated from three
parts of Thailand.
Genera Number of isolated
Northern Central Northeastern
Anabaena sp. 123 98 101
Nostoc sp. 77 90 75
Scytonema sp. 10 10 13
Stigonema sp. – – 5
Calothrix sp. 3 4 3
Tolypothrix sp. 3 – 3
Hapalosiphon sp. 12 24 21
Fischerella sp. 3 5 3
Nodularia sp. 3 4 5
Anabaenopsis sp. 3 4 5
Cylindrospermum sp. – – 1
Unidentified 40 45 47
Total 277 284 282
Cyanobacterial diversity (I) 675
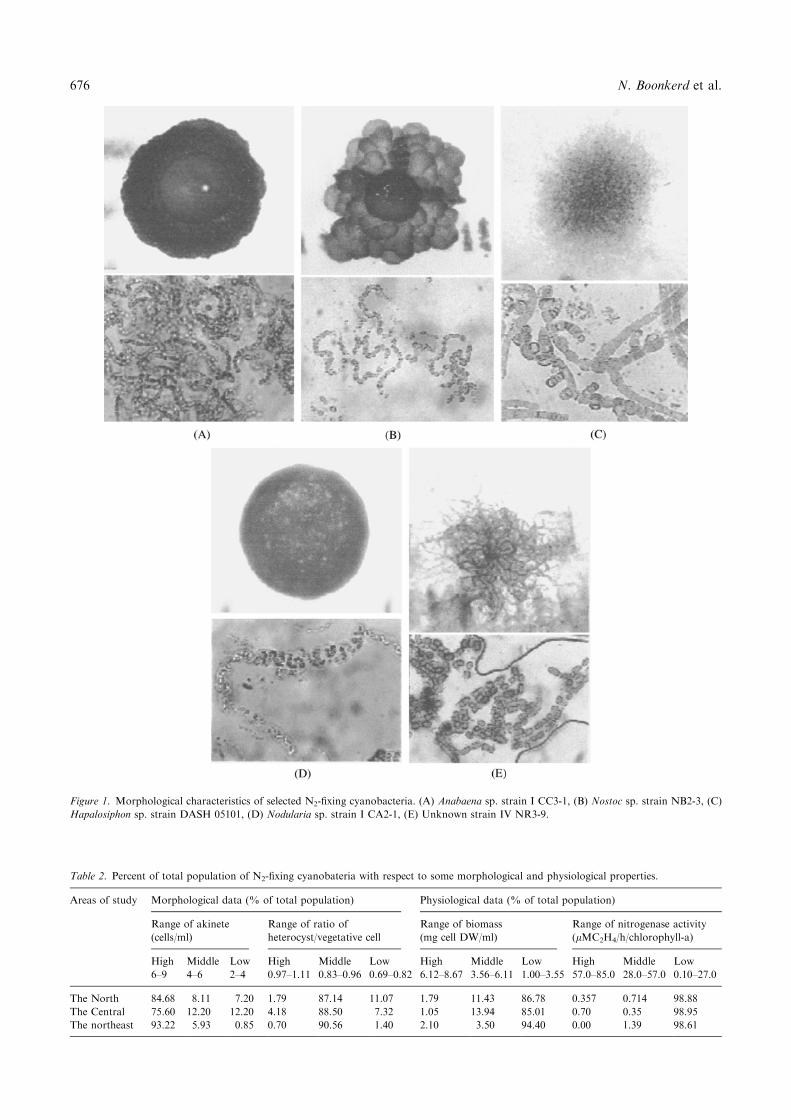
Figure 1. Morphological characteristics of selected N2-fixing cyanobacteria. (A) Anabaena sp. strain I CC3-1, (B) Nostoc sp. strain NB2-3, (C)
Hapalosiphon sp. strain DASH 05101, (D) Nodularia sp. strain I CA2-1, (E) Unknown strain IV NR3-9.
Table 2. Percent of total population of N2-fixing cyanobateria with respect to some morphological and physiological properties.
Areas of study Morphological data (% of total population) Physiological data (% of total population)
Range of akinete
(cells/ml)
Range of ratio of
heterocyst/vegetative cell
Range of biomass
(mg cell DW/ml)
Range of nitrogenase activity
(lMC2H4/h/chlorophyll-a)
High Middle Low High Middle Low High Middle Low High Middle Low
6–9 4–6 2–4 0.97–1.11 0.83–0.96 0.69–0.82 6.12–8.67 3.56–6.11 1.00–3.55 57.0–85.0 28.0–57.0 0.10–27.0
The North 84.68 8.11 7.20 1.79 87.14 11.07 1.79 11.43 86.78 0.357 0.714 98.88
The Central 75.60 12.20 12.20 4.18 88.50 7.32 1.05 13.94 85.01 0.70 0.35 98.95
The northeast 93.22 5.93 0.85 0.70 90.56 1.40 2.10 3.50 94.40 0.00 1.39 98.61
676 N. Boonkerd et al.

(76–93% of total population) contained high amountsof akinete cells/ml and most of the strains in this groupbelonged to the genus Anabaena. To investigate the ratiobetween heterocysts and vegetative cells, the ratioheterocyst/vegetative cells could be divided into threelevels; high (0.97–1.11), moderate (0.83–0.96) and low(0.69–0.82). The results showed that most of isolates hadmoderate range of heterocyst/vegetative cells ratio (63–87% of total population). Only a few isolates had a highratio of heterocyst/vegetative cells, which most likelybelong to only two genera Anabaena and Nostoc, exceptfor a few isolates of Hapalosiphon sp. and unidentifiedisolates from the middle part of Thailand.
Physiological diversity: growth rate and N2-fixingcapability
To determine the growth rate of all N2-fixing cyano-bacterial isolates, biomass determination was used. Thegrowth rate was divided into three ranges; slow growthrate (1.00–3.55 mg cell dry weight/ml/12 days), mediumgrowth rate (3.56–6.11 mg cell dry weight/ml/12 days)and high growth rate (6.12–8.67 mg cell dry weight/ml/12 days). Most isolates (85–95%) showed a slow growthrate while very few isolates had a high growth rate. Thegroups with high growth rate were identified as Anaba-ena and Nostoc, while other genera and some uniden-tified species were slow growers. To confirm anddetermine the capability and efficiency of N2-fixingcyanobacterial strains, the ARA for nitrogenase wascarried out. The efficiency of N2-fixation was conductedon the basis of single isolates and incubations wereperformed at 25 �C under a 12 h/12 h light/dark cyclefor 30 days. Results obtained from this study could begrouped into three ranges of N2-fixing efficiency; lowefficiency (0.10–27.0 lM C2H4/mg of chlorophyll-a/h),moderate (28.0–57.0 lM C2H4/mg of chlorophyll-a/h)and high efficiency (57.0–85.0 lM C2H4/mg of chloro-phyll-a/h). From this study, the groups with high N2-fixing ability mostly belong to the genera Nostoc andAnabaena. None of the strains from the northeasternpart of Thailand showed high N2-fixation efficiency.
nifH gene profile analysis
To further characterize these N2-fixing cyanobacteria, agroup of 102 randomly selected isolates comprising 45isolates of Nostoc sp., 44 isolates of Anabaena sp., fiveisolates of Anabaenopsis sp., three isolates of Nodulariasp. and five isolates of an unidentified branching group,were used for PCR amplification of nifH. Two differentsets of degenerate oligonucleotide primers were used forthe PCR amplification of the nifH gene fragment.Primer 1 was located at positions 318 to 335 in referenceto the Anabaena sp. strain PCC 7120 nifH sequence.Primer 2 was located in positions 659–674 of theopposite strand. Amplification of DNA samples from107 cyanobacterial isolates (including five referencestrains) produced one to six amplification products,
depending on the strains. The Hapalosiphon sp. DASH05101 yielded a single 330 bp major band. Two bandproducts (830 and 330 bp) were generally observed withthe Calothrix sp. DASH 02101, as in Scytonema sp., aswell as, Anabaena cylindrica, but Nostoc sp. gave fivebands in size 830, 446, 330, 208 and 182 bp. This mainPCR product was found in size about 330 bp fragment(data not shown) which agreed with the 359 bp expectedsize based on available sequences. The additional PCRproducts might be generated from some unrelatedsequences as vnfH (91% identical) or anfH (63%identical) (Porath & Zehr 1994).The cyanobacterial strains were distinguished into 77
different groups according to their patterns of PCR, andonly four strains, I CF3-22 (Nostoc sp.), I CD1-1(Nostoc sp.), I NM3-1-3 (Anabaena sp.) and III NCR4-1(Anabaenopsis sp.), did not contain the 330 bp majorband. When nifH PCR products were used to constructa dendrogram based on the UPGMA algolithm, theresults divided the isolates into four main groups(Figure 2). Every genus of cyanobacteria seemed toscatter in the main cluster. Most isolates that containedPCR products in size of about 290, 330 and 380 bpcould fix N2 with high efficiency, thus, nifH fragmentmight be used as prediction tool for indicating theefficiency of nitrogen fixation among the various groupsof cyanobacteria. However, further nifH fragmentsanalyses by Southern blot hybridization and sequencingtechnique are needed.
Combination of PCR products from three primers
Application of the primer from the STRR in the PCR tothe cyanobacterial isolates yielded multiple distinctDNA products ranging in size from approximately132–4000 bp (data not shown) which could be classifiedinto 97 different groups (include reference strains).When two sets of random primers, DAF 8.7b andDAF 10.6e were used separately in PCR amplification,the multiple distinct DNA products ranged in size fromapproximately 178–3000 bp and 178–4000 bp, respec-tively (data not shown). Only DNAs from 78 cyano-bacterial isolates were able to be amplified by using theDAF 8.7b primer and 94 cyanobacterial isolates byDAF 10.6e primer. The PCR products obtained fromthe 78 isolates using the DAF 8.7b primer werecategorized into 71 different groups, while the use ofthe DAF 10.6e primer distinguished 94 cyanobacterialisolates into 75 groups. Each primer used in this studyindicated a different ability to discriminate the cyano-bacterial DNA templates. Therefore, PCR productsfrom using STRR, DAF 8.7b and DAF 10.6e werecombined to generate the most clearly distinguisheddendrogram. Reproducibility of the dendrogram wasachieved from combined PCR products which couldclearly distinguish all cyanobacterial strains (Figure 3).The dendrogram in Figure 3, can be separated into
two main clusters. The first major clusters indicated thatmost isolates belong to these clusters with high percent
Cyanobacterial diversity (I) 677
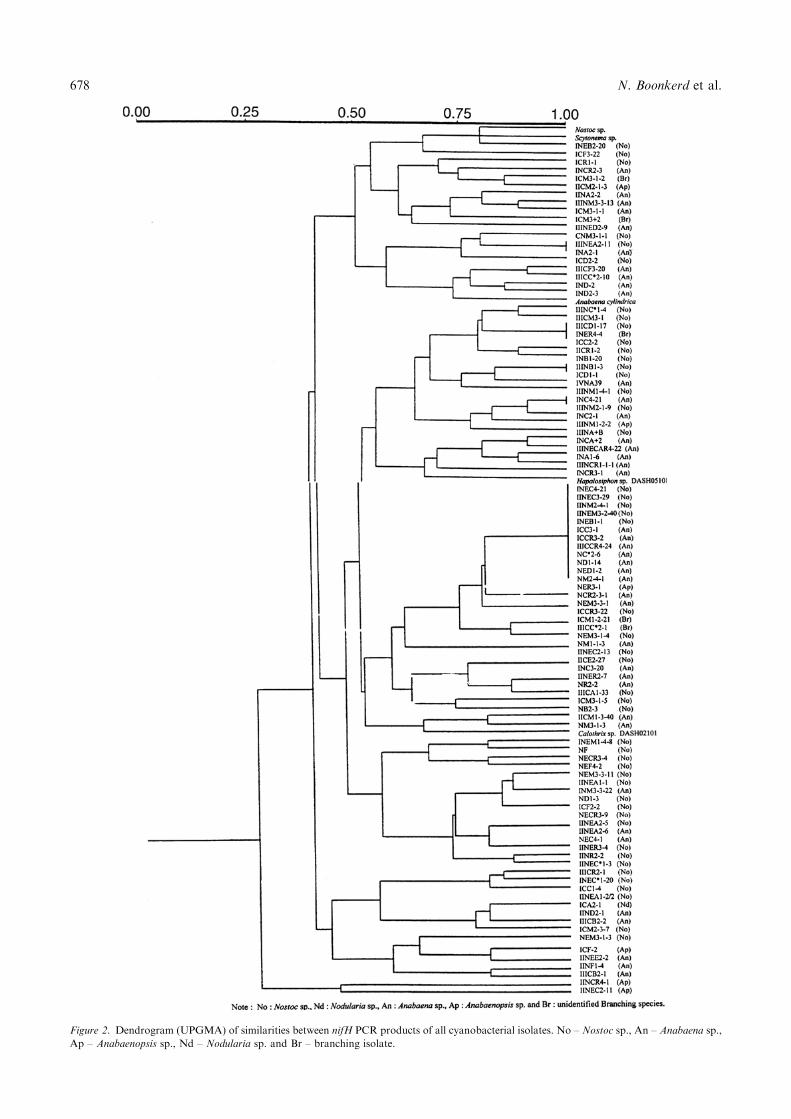
Figure 2. Dendrogram (UPGMA) of similarities between nifH PCR products of all cyanobacterial isolates. No – Nostoc sp., An – Anabaena sp.,
Ap – Anabaenopsis sp., Nd – Nodularia sp. and Br – branching isolate.
678 N. Boonkerd et al.
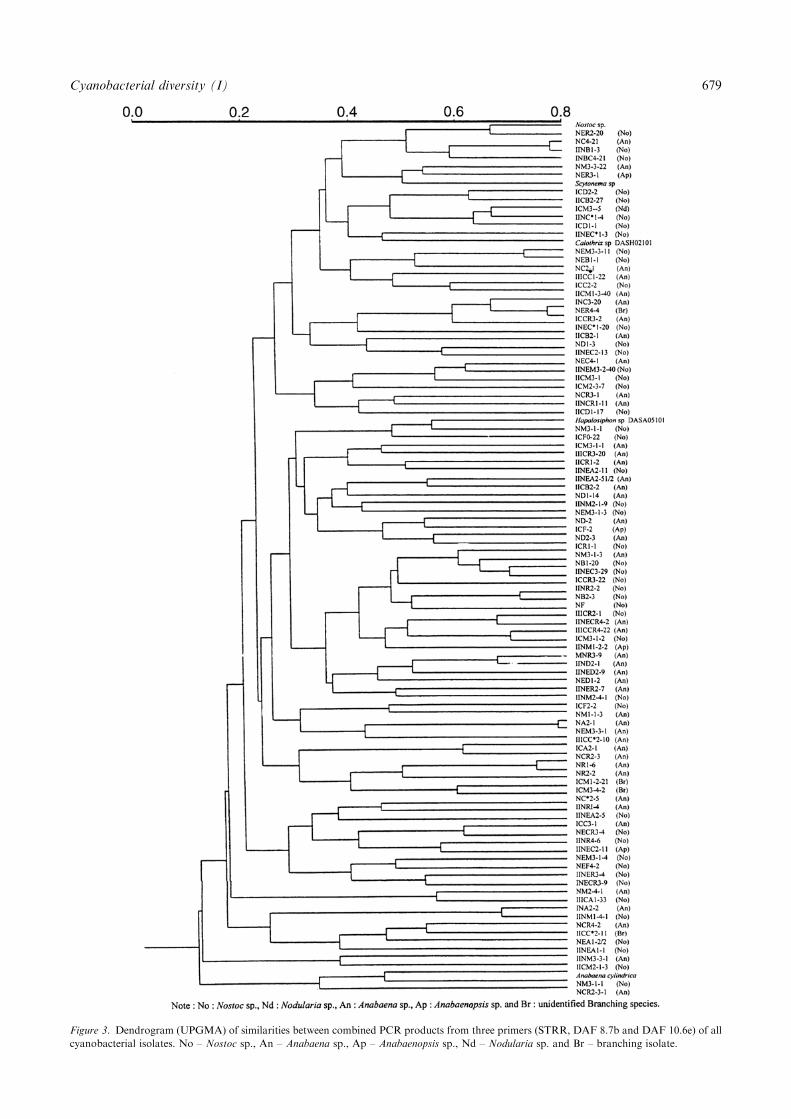
Figure 3. Dendrogram (UPGMA) of similarities between combined PCR products from three primers (STRR, DAF 8.7b and DAF 10.6e) of all
cyanobacterial isolates. No – Nostoc sp., An – Anabaena sp., Ap – Anabaenopsis sp., Nd – Nodularia sp. and Br – branching isolate.
Cyanobacterial diversity (I) 679

similarity. Several genera of cyanobacteria were dis-persed in these clusters and moreover, no relationshipscould be seen between closely related groups of PCRpatterns and identification results obtained from con-ventional classification. Cyanobacterial strains that didnot show correlation between DNA pattern and phe-notypic characteristics were NC4-21 (Anabaena sp.), IINB1-3 (Nostoc sp.), III C3-20 (Anabaena sp.) andNER44 (branching group), which were nearly identicalto each other and only diverged at a similarity level ofabout 0.05. Some Anabaena strains such as NEM3-3-1and III CC2-10 or Anabaena strain NCR2-3 and NR1-6shared both the similarity of phenotypic characters andsimilarity level in range of 0.05–0.075. Another clusterwas excluded and it presented high similarity levelapproximately 0.4% as A. cylindrica, Nostoc sp. strainNM3-1-1 and Anabaena sp. NCR2-3-1.Since Nostoc sp. and Anabaena sp. were the major
indigenous species found in Thai soil, the diversityamong each of them was further elucidated. When thePCR products generated from three sets of primers werecombined for a group of Nostoc sp., most of the strainsshowed low degrees of similarity among themselves.Only two strains were discriminated from the groupamong genus Nostoc, III NEM1-4-3 and III NM1-4-1(Figure 4). On the other hand, when the Anabaena sp.group was analysed in the same way as in Nostoc sp.,high diversity was also found in this genus. The strainswhich demonstrate more closely related isolates such asI NEY3-3-1 and III CC2-10 shared approximately 80%similarity. Only one strain, I NCR2-3-1 was excludedfrom the main groups of genus Anabaena (Figure 5).
Discussion
Our results indicated that the nitrogen-fixing cyanobac-teria of genera such as Alphanothece, Gloeothece, Syn-echococcus, Democarpa, Xenococcus, Myxosarcina,
Chroococcidipiosis and Pleurocapsa were not found inthis study. These results indicate that the nitrogen-fixingcyanobacterial populations in Thailand are differentfrom those reported in India (Kaplan et al. 1986;Ventakaraman 1986). The unicellular cyanobacteria arealso found in the rice fields of India, whereas in severalcountries in South Asia and Southeast Asia, filamentousforms including Anabaena and Nostoc (Watanabe et al.1951; Konishi & Seino 1961; Jutono 1973) are mainlyfound. The results obtained from this study suggest thatthe dominant species of N2-fixing cyanobacteria in Thaisoil were Anabaena and Nostoc. This also stronglysupports several investigations in various countries suchas India (Ventakaraman 1979), Australia (Bunt 1996),Morocco (Renaut et al. 1975) and in the rice fields ofValencia, Spain (Quesada et al. 1998). However, thisstudy used only one condition for isolation, cultivationand determination of N2-fixing efficiency. Therefore,using other methods or conditions might result in
Figure 4. Dendrogram (UPGMA) of similarities between combined PCR products from three primers (STRR, DAF 8.7b and DAF 10.6e) of all
Nostoc sp. isolates.
Figure 5. Dendrogram (UPGMA) of similarities between combined
PCR products from three primers (STRR, DAF 8.7b and DAF 10.6e)
of all Anabaena sp. isolates.
680 N. Boonkerd et al.

additional cyanobacterial genera for which ASM medi-um is not appropriate, for example Anacystic nidulans,while other media such as BG110 or Z8 are morefavourable (Rippka 1988). To determine the otherecophysiological characteristics such as akinetes andheterocysts, some results implied that these morpho-physiological traits are correlated to agro-ecosystems.Akinetes are produced as resting-spores under phos-phorus-limited condition (Desikachary 1959). The high-est number of akinetes from N2-fixing cyanobacteria inthis study was isolated from the northeastern region,which reflects the poor soil condition in this part ofThailand. The middle range percent ratio of heterocyst/vegetative cells was the predominant characteristic ofN2-fixing cyanobacterial isolates from all areas. Thisalso implied that most Thai soils contain moderateamounts of organic nitrogen, since heterocyst formationis induced when vegetative cells are in conditions of lownitrogen. For further selecting the appropriate N2-fixingcyanobacterial strain as a biological fertilizer, the straincontaining high nitrogenase activity (high heterocystcells) and fast growth rate should be emphasized.Prior to analysing the genetic diversity, PCR products
obtained from large-scale DNA extraction and a fewintact filaments were compared. The results showedindistinguishable PCR products from both methods.Thus, application of intact cells for PCR represents avery efficient as mentioned by Rasmussen & Svenning(1998).In an attempt to analyse the genetic diversity among
strains, a group of random primers were employed inthis study. Each primer of STRR, DAF 8.7b and DAF10.6e did not show the efficiency to discriminate allstrains. When the combination of PCR products genera-ted from these primers was analysed, all strains wereclearly differentiated. This shows that the N2-fixingcyanobacteria in Thai soil are diverse. In addition, thecorrelation between DNA pattern and genus establish-ment in each habitat was not found in this study.However, a small relationship between informationobtained from conventional identification and DNAfingerprint technique was observed. This circumstancewas similar to N2-fixing cyanobacteria TrichodesmiumNIBB 1067 and Oscillatoria PCC 7515, which per-formed as close relatives, presenting almost 95% of 16SrRNA sequence similarity (Wilmotte et al. 1994). There-fore, both phenotypic and genotypic characteristicsshould be taken into consideration.Phylogenetic analysis clearly distinguished all cyano-
bacterial strains even at intraspecies level. This showed awide range of diversity among the N2-fixing cyanobac-teria in Thai soil. A database from these results wouldbe very fruitful since it has never been investigatedbefore in Thailand. Thus agricultural managementshould be concerned with appropriate strategies toconserve the microbial diversity. Some cyanobacterialisolates identified in this study will be suitable foragricultural application such as utilizing N2-fixing cy-
anobacterial inoculum as biofertilizer in sustainableagriculture. The use of cyanobacterial biofertilizerinstead of chemical fertilizer may be an intriguing wayto promote environmentally friendly practices in inten-sive agriculture.
Acknowledgements
This work was fully supported by The Biodiversity/Research and Training Program (BRT240001-540073)and Suranaree University of Technology.
References
Bunt, J.S. 1996 Nitrogen fixing blue green algae in Australian rice soils.
Nature 192, 479–480.
Desikachary, T.V. 1959 Cyanophyta. Indian Council of agricultural
research, New Delhi.
Desikachary, T.V. 1985 Cyanophyta. University Botany laboratory,
Madras.
Eskew, D.M., Anolles, G.C., Bassam, B.J. & Gresshoff, P.M. 1993
DNA amplification fingerprint of the Azolla-Anabaena symbiosis.
Plant Molecular Biology 21, 363–373.
Fogg, G.E., Stewart, W.D.P., Fay, P. & Walsby, A.E. 1973 The Blue-
Green Algae. 459 pp. London and New York: Academic Press.
ISBN 0-12-261650-2.
Grant, I.F., Roger, P.A. & Watanabe, I. 1985 Effect of grazer
regulation and algal inoculation on photodependent nitrogen
fixation in a wetland rice field. Biology and Fertility of Soil 1, 61–72.
Hiroki, M., Shimizu, A., Li, R., Watanabe, M. & Watanabe, M.M.
1998 Development of a database system useful for identification of
Anabaena spp. (cyanobacteria). Phycological Research 46, 85–93.
Jutono, D.K.K. 1973 Blue green algae in rice soils of Jogiakarta,
Central Java. Soil Biology and Biochemistry 5, 91–96.
Kaplan, D., Richmond, A.E., Dubinsky, Z. & Aaronson, S. 1986 Algal
Nutrition. InHandbook of Microalgal Mass Culture. ed Richmond,
A. pp. 147–198. Roca Raton, Florida: CRC Press Inc. ISBN
0-84933240-0.
Konishi, C. & Seino, K. 1961 Studies on the maintenance of soil
fertility of paddy field in nature (in Japanese, English summary).
Hokuriku Agricultural Experimental Station Bulletin 2, 41–136.
Lupski, J.R. & Weinstock, G.M. 1992 Short, interspersed repetitive
DNA sequences in prokaryotic genomes. Journal of Bacteriology
174, 4525–4529.
Mazel, D., Houmard, J., Castets, A.M. & Taodeau de Marsac, N. 1990
Highly repetitive DNA sequences in cyanobacterial genomes.
Journal of Bacteriology 172, 2755–2761.
Neilan, B.A. 1995 A study of the algae from paddy soils of Ballia and
Ghazipur districts of Uttar Pradesh, India. I. Cultural and
ecological considerations. Nova Hedwigia 9, 299–334.
Porath, J.B. & Zehr, J.P. 1994 Detection and characterization of
cyanobacteria nifH genes. Applied and Environmental Microbiology
60, 880–887.
Quesada, A., Nieva, M., Leganes, F., Ucha, A., Martin, M., Prosperi,
C. & Fernandez-Valiente, E. 1998 Acclimation of cyanobacterial
communities in rice fields and response of the nitrogenase activity
to light regime. Microbial Ecology 35, 147–155.
Rai, A.N. 1990 In Handbook of Symbiotic Cyanobacteria, ed Rai, A.N.
Boca Raton, Florida: CRC Press. ISBN 0-84933275-3.
Rasmussen, U. & Svenning, M.M. 1998 Fingerprinting of cyanobac-
teria based on PCR with primers derived from short and long
tandemly repeated repetitive sequences. Applied and Environmental
Microbiology 64, 265–272.
Renant, J., Sasson, A., Pearson, H.W. & Stewart, W.P.D. 1975
Nitrogen fixing algae in Morocco. In Nitrogen Fixation by Free-
Cyanobacterial diversity (I) 681

Living Microorganism. ed Stewart, W.P.D. pp. 229–246. Cam-
bridge: Cambridge University Press. ISBN 0-521-20708-8.
Richmond, A. 1986 CRC Handbook of Microalgal Mass Culture. Boca,
Raton, Florida, USA: CRC Press Inc. ISBN 0-84933240-0.
Rippka, R. 1988 Recognition and identification of cyanobacteria.
Methods in Enzymology 167, 28–67.
Robinson, N.J., Robinson, P.J., Gupta, A., Bleasby, A.J., Whitton,
B.A. & Morby, A.P. 1995 Singular over-representation of an
octameric palindrome, HIP 1, in DNA from many cyanobacteria.
Nucleic Acids Research 23, 729–735.
Rouhiainen, L.R., Sivonen, K., Buikema, W.J. & Haselkorn, R. 1995
Characterization of toxin-producing cyanobacteria by using an
oligonucleotide probe containing a tandemly repeated heptamer.
Journal of Bacteriology 177, 6021–6026.
Ventakaraman, G.S. 1979 Algal inoculation in rice fields. In Nitrogen
and Rice. pp. 311–324. Loss Bonos, Laguna, Philippines: IRRI.
Ventakaraman, G.S. 1986 Blue-green algae as biofertilizer. In Hand-
book of Microalgal Mass Culture. ed Richmond, A. pp. 455–472.
Boca Raton, Florida: CRC Press Inc. ISBN 0-84933240-0.
Watanabe, A., Nishigaka, S. & Konishi, C. 1951 Effect of nitrogen-
fixing blue-green algae on the growth of rice plants. Nature 168,
748–749.
West, N.J. & Adams, D.G. 1997 Phenotypic and genotypic compari-
son of symbiotic and free-living cyanobacteria from a single field
site. Applied and Environmental Microbiology 63, 4479–4484.
Wintermans, J.F.G.M. & Demots, A. 1965 Spectrophotometric
characteristics of chlorophyll a, b and their pheophytins in
ethanol. Biochemica et Biophysica Acta 109, 440–453.
Wilmotte, A., Neefs, J.M. & Wachter, R De. 1994 Evolutionary
affiliation of the marine nitrogen-fixing cyanobacterium Trichodes-
mium sp. strain NIBB 1067, derived by 16S ribosomal RNA
sequence analysis. Microbiology 140, 2159–2164.
682 N. Boonkerd et al.
Related Documents











