of April 11, 2018. This information is current as Cure of Colitis T Cells during + CD25 + IL-10-Secreting CD4 and + CD25 + CD4 + Characterization of Foxp3 Jason D. Fontenot, Fred Ramsdell and Fiona Powrie Izcue, Claire Thompson, Andrea Fanger, Andrea Tannapfel, Holm H. Uhlig, Janine Coombes, Christian Mottet, Ana http://www.jimmunol.org/content/177/9/5852 doi: 10.4049/jimmunol.177.9.5852 2006; 177:5852-5860; ; J Immunol References http://www.jimmunol.org/content/177/9/5852.full#ref-list-1 , 22 of which you can access for free at: cites 45 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2006 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 11, 2018 http://www.jimmunol.org/ Downloaded from by guest on April 11, 2018 http://www.jimmunol.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

of April 11, 2018.This information is current as
Cure of Colitis T Cells during+CD25+IL-10-Secreting CD4
and+CD25+CD4+Characterization of Foxp3
Jason D. Fontenot, Fred Ramsdell and Fiona PowrieIzcue, Claire Thompson, Andrea Fanger, Andrea Tannapfel, Holm H. Uhlig, Janine Coombes, Christian Mottet, Ana
http://www.jimmunol.org/content/177/9/5852doi: 10.4049/jimmunol.177.9.5852
2006; 177:5852-5860; ;J Immunol
Referenceshttp://www.jimmunol.org/content/177/9/5852.full#ref-list-1
, 22 of which you can access for free at: cites 45 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Characterization of Foxp3�CD4�CD25� and IL-10-SecretingCD4�CD25� T Cells during Cure of Colitis1
Holm H. Uhlig,2*† Janine Coombes,2* Christian Mottet,* Ana Izcue,* Claire Thompson,*Andrea Fanger,† Andrea Tannapfel,‡ Jason D. Fontenot,§ Fred Ramsdell,¶ and Fiona Powrie3*
CD4�CD25� regulatory T cells can prevent and resolve intestinal inflammation in the murine T cell transfer model of colitis.Using Foxp3 as a marker of regulatory T cell activity, we now provide a comprehensive analysis of the in vivo distribution ofFoxp3�CD4�CD25� cells in wild-type mice, and during cure of experimental colitis. In both cases, Foxp3�CD4�CD25� cells werefound to accumulate in the colon and secondary lymphoid organs. Importantly, Foxp3� cells were present at increased density incolon samples from patients with ulcerative colitis or Crohn’s disease, suggesting similarities in the behavior of murine and humanregulatory cells under inflammatory conditions. Cure of murine colitis was dependent on the presence of IL-10, and IL-10-producing CD4�CD25� T cells were enriched within the colon during cure of colitis and also under steady state conditions. Ourdata indicate that although CD4�CD25� T cells expressing Foxp3 are present within both lymphoid organs and the colon, subsetsof IL-10-producing CD4�CD25� T cells are present mainly within the intestinal lamina propria suggesting compartmentalizationof the regulatory T cell response at effector sites. The Journal of Immunology, 2006, 177: 5852–5860.
I ncreasing evidence suggests that functionally specializedsubsets of CD4� T cells play a key role in the regulation ofimmune responses (1, 2). In particular, naturally occurring
CD4�CD25� regulatory T cells (TR)4 have been shown to preventboth T cell-mediated and innate immune pathology in a number ofdisease models (3–5). CD4�CD25� TR cells with similar proper-ties to those described in mice are also present in humans andimpaired function of these cells has been observed in patients withautoimmune disease (6–11). The transcription factor Foxp3 is dif-ferentially expressed by CD4�CD25� TR cells and plays a keyrole in their development and function (12–14). Accordingly, micelacking functional Foxp3 develop a multiorgan-inflammatory dis-ease (12, 15, 16). Similarly, loss of function mutations in FOXP3have been shown to be responsible for the human autoimmune andinflammatory disease, immune polyendocrine X-linked enteropa-thy, providing evidence that TR cells also contribute to immunehomeostasis in humans (17, 18).
The T cell transfer model of colitis provides a good system withwhich the mechanisms of TR function can be dissected. In this
model, transfer of naive CD4�CD45RBhigh T cells into immuno-deficient mice leads to a Th1-mediated colitis, while cotransfer ofCD4�CD25� TR cells can completely prevent disease (4). In ad-dition, recent studies in models of type 1 diabetes and inflamma-tory bowel disease (IBD) have shown that CD4�CD25� TR notonly prevent development of disease but can also actually reverseestablished inflammation (19–22). Thus, using the T cell transfermodel of colitis, we found that CD4�CD25� T cells can reversean established T cell-mediated inflammatory response in the intes-tinal mucosa by reducing the pathogenic T cell infiltrate, ultimatelyleading to restoration of normal intestinal architecture. It is likelythat the mechanisms by which TR can prevent or cure colitis differ.In prevention of colitis, TR must control activation of a predomi-nantly naive population of cells, while in cure of colitis, they mustact on Ag-experienced cells and an aggressive inflammatoryresponse.
The ability of CD4�CD25� TR cells to resolve established in-flammation in model systems raises the possibility that these cellsmay be useful as therapeutic agents for chronic inflammatory dis-eases in humans. With this in mind, it will be important to establishwhether the properties of TR cells determined through study ofprevention of colitis also apply to cure of colitis. Attempts to de-sign effective therapeutic strategies will also be aided by knowl-edge of the location and behavior of TR in the human IBDs.
We have previously shown that, during cure of experimentalcolitis, CD4�CD25� T cells proliferate and accumulate in themesenteric lymph nodes (MLN) and also in the colonic laminapropria (LP). At both sites, the progeny of CD4�CD25� T cellsare in direct contact with CD11c� dendritic cells, as well as ef-fector T cells (20). These findings suggest that regulation of anactive immune response by CD4�CD25� T cells occurs in thedraining lymph node, as well as at the site of inflammation. How-ever, these studies have been limited by the lack of specific mark-ers for naturally arising TR cells. Although useful, CD25 expres-sion does not uniquely identify TR cells, as the CD4�CD25� poolcan contain activated effector cells and not all TR cells expressCD25. Identification of Foxp3 as a more specific marker for TR
cells provides an opportunity to track the fate of TR cells in vivo
*Sir William Dunn School of Pathology, University of Oxford, Oxford, United King-dom; †Children’s Hospital and ‡Institute of Pathology, University of Leipzig, Leipzig,Germany; §Department of Immunology, University of Washington, Seattle, WA98195; and ¶Zymogenetics, Seattle, WA 98102
Received for publication February 22, 2006. Accepted for publication August 3, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Wellcome Trust (to F.P. and C.T.), the MedicalResearch Council (to J.C.), Deutsche Forschungsgemeinschaft Grant HU 128-2 (toH.H.U.), the University of Leipzig (to H.H.U., F1-49), the Swiss National Fund (toC.M.), training grants from the National Institutes of Health and the Cancer ResearchInstitute, and the Spanish Ministerio de Educacion y Ciencia (to A.I.).2 H.H.U. and J.C. contributed equally.3 Address correspondence and reprint requests to Dr. Fiona Powrie, Sir William DunnSchool of Pathology, University of Oxford, South Parks Road, Oxford OX1 3RE,U.K. E-mail address: [email protected] Abbreviations used in this paper: TR, regulatory T cell; IBD, inflammatorybowel disease; MLN, mesenteric lymph node; LP, lamina propria; CD, Crohn’sdisease; UC, ulcerative colitis; POD, peroxidase; MHC-II, MHC class II; DAPI,4�,6�-diamidino-2-phenylindole.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

(23, 24), although studies to date have primarily focused on ex-pression of Foxp3 mRNA.
Cure of colitis by CD4�CD25� T cells has also been shown tobe functionally dependent on IL-10, however, it is not knownwhether TR cells themselves are the important source of IL-10 andwhere it is produced (19). In this study, we have used anti-Foxp3Abs together with analysis of IL-10 secretion to further investigatethe functional and phenotypic characteristics of CD4�CD25� Tcells under physiological conditions and during resolution of in-testinal inflammation. Our results show that during cure of intes-tinal inflammation the majority of Foxp3� cells and IL-10-secret-ing cells derive from CD4�CD25� precursors. However, whereasFoxp3� cells are present in similar frequencies in both the sec-ondary lymphoid organs and LP of colitic animals, IL-10-produc-ing CD4�CD25� T cells selectively enrich within the colonic LP.In addition, we have extended these findings to analysis of humanIBD and show an accumulation of Foxp3�CD4�CD25� cells inthe inflamed intestine of patients with IBD. This suggests thatchronic intestinal inflammation in humans is not simply a conse-quence of a lack of TR cells at the inflammatory site.
Materials and MethodsMouse and human tissue samples
BALB/cJ, C57BL/6J, congenic C57BL/6.SJL.CD45, C57BL/6 recombi-nase-activating gene 1-deficient (rag1�/�), CB17 SCID, IL-10-knockout(IL-10�/�), and Foxp3�/� mice (12) were bred under specific pathogen-free conditions. All mice used were �6 wk old.
Colonic tissue sections from patients with Crohn’s disease (CD) andulcerative colitis (UC) as well as inflamed and noninflamed human colon,appendix, and tonsil tissue were obtained from the Department of Pathol-ogy and from the Department of Ear Nose and Throat diseases of theUniversity of Leipzig. All tissues were obtained for therapeutic and diag-nostic reasons. The studies were approved by the local ethical review com-mittee (HU242/04).
Cell purification and flow cytometry
CD4� T cell subsets were isolated from spleens as described (20). Essen-tially, erythrocyte-depleted spleen cell suspensions were enriched forCD4� cells by negative selection following incubation with anti-CD8(clone YTS169 (25)), anti-B220 (clone RA3-6B2 (26)), anti-CD11b (cloneTIB128; American Type Culture Collection (ATCC)) and anti-MHC-IIAbs (TIB120; ATCC). Ag-positive cells were depleted using sheep anti-rat-IgG Dynal beads (Dynal Biotech).
For MACS sorting of CD4�CD25� T cells, CD4�-enriched cells wereincubated with biotinylated anti-CD25 (clone 7D4), followed by strepta-vidin MACS beads, and sorted on an AutoMACS (Miltenyi Biotec).
For FACS sorting of CD4�CD45RBhigh and CD4�CD25� T cells,CD4�-enriched cells were stained with anti-CD45RB (clone 16A), anti-CD25 (clone 7D4), and anti-CD4 (clone H129.19; all BD Biosciences), andsorted on a MoFlo (DakoCytomation). The purity of MACS- and FACS-sorted cells was �90 and �99%, respectively. Because similar results wereobtained using MACS or FACS sorting, data were pooled.
Unless otherwise stated, all Abs were obtained from BD Biosciences.FACS analysis of Foxp3 expression was performed using the Foxp3
staining kit from eBioscience.
T cell transfer experiments
SCID and rag1�/� mice were injected i.p. with 4 � 105 syngeneicCD4�CD45RBhigh T cells. Mice developed colitis 3.5–4.5 wk posttransfer.Mice with clinical signs of disease were either sacrificed to assess theseverity of colitis or received 106 CD4�CD25� T cells i.p., or no treat-ment. A further group of mice received weekly injections of 0.5 mg ofanti-IL-10R Ab (clone 1B1.2). Mice were observed daily and weighedweekly. All animal experiments were performed according to the homeoffice guidelines and the United Kingdom Animals Scientific ProceduresAct of 1986.
Histology
Tissue sections were stained with H&E and colitis severity was gradedsemiquantitatively from 0 to 4, as outlined in Ref. 27.
Mouse tissue samples were snap-frozen and cryocut. Slides were ace-tone-fixed and blocked with donkey serum (Sigma-Aldrich). The endoge-nous peroxidase (POD) activity was inhibited using H2O2, sodium azide,and in some cases glucose oxidase. Foxp3 staining was performed usingrabbit polyclonal anti-mouse Foxp3 Abs (Celltech R&D) followed byFITC-conjugated donkey anti-rabbit IgG (Jackson ImmunoResearch Lab-oratories). The Foxp3 staining was combined either with staining for CD4,CD25, MHC class II (MHC-II) or CD45.1 and 4�,6�-diamidino-2-phenylin-dole (DAPI). Directly labeled anti-CD4-FITC (clone RM4-5; BD Bio-sciences) was used. Anti-CD25 (clone PC61) was detected using anti-mouse POD (Jackson ImmunoResearch Laboratories), and biotinylated anti-CD45.1 (A20; BD Biosciences) was detected using avidin-POD (VectorLaboratories). The POD activity was visualized by tyramide amplification(NEN Life Science Products) as described previously (20). Biotinylatedanti-MHC-II (I-A/I-E, clone 2G9) was detected using directly labeledstreptavidin fluorescence conjugates (BD Biosciences). Some tissue sec-tions were counterstained for 3–5 min with 0.1–1 �g/ml DAPI (SigmaAldrich). Fluorescence was detected using appropriate filter combinationsfor DAPI, FITC, Texas Red/Cy3, and Cy5.
Human formalin-fixed paraffin-embedded tissue samples were deparaf-finized, microwave pressure cooked to demask fixed Ags, and stained forCD3 (clone F7.2.38; DakoCytomation), and Foxp3 (rabbit polyclonal se-rum against human Foxp3; Celltech R&D) followed by anti-mouse or anti-rabbit POD-labeled Abs and routine POD staining. For costaining exper-iments, frozen tissue sections of human tonsil or appendix were cryocut,fixed in formalin, and incubated at 85°C in Ag retrieval buffer. After block-ing the endogenous POD activity, sections were stained using mouse anti-CD4 (clone 1F6; Serotec) and donkey anti-mouse-POD (Jackson Immu-noResearch Laboratories) followed by tyramide amplification. Aftersubsequent blocking of POD activity, Foxp3 was stained using polyclonalanti-Foxp3 followed by donkey anti-rabbit-POD (Jackson ImmunoRe-search Laboratories) and tyramide amplification. DAPI counterstainingwas performed to visualize the tissue structure and to ensure the nuclearlocalization of the Foxp3 signal. Similarly, costaining for IL-10, CD3, andFoxp3 was performed using rat anti-IL-10 (clone JES3-9D7; Caltag Lab-oratories), polyclonal anti-CD3 (A0452; DakoCytomation) and mouse anti-Foxp3 (clones 105/221D/D3 and 236A/E7; Abcam). Sections were subse-quently incubated with appropriate donkey POD-labeled Abs (JacksonImmunoResearch Laboratories) followed by tyramide amplification.
Quantitative real-time PCR analysis
Cells were FACS sorted to �99% purity and RNA extracted using theRNeasy minikit (Qiagen), including a DNase digestion step. cDNA wastranscribed with Superscript II (Invitrogen Life Technologies) and used asa template for real-time quantitative PCR. A multiplex reaction for CD3�and Foxp3 was preformed using the chromo4 (MJ Research) machine andresults expressed relative to an arbitrary standard of CD4� T cell cDNA.Foxp3 expression was then normalized to the internal standard, CD3�. Thefollowing primers and TaqMan probe sequences were used: CD3� probeVIC-ACATAGGCACCATATCCGGCTTTATCTTCG-TAMRA; CD3�5�-TTACAGAATGTGTGAAAACTGCATTG; CD3� 3�-CACCAAGAGCAAGGAAGAAGATG; Foxp3 probe FAM-ATCCTACCCACTGCTGGCAAATGGAGTC-TAMRA; Foxp3 5�-CCCAGGAAAGACAGCAACCTT; Foxp3 3�-TTCTCACAACCAGGCCACTTG.
Intracellular cytokine staining
Cell suspensions were prepared from spleen, MLN, and the LP. For prep-aration of LP cell suspensions, colons were cut into 0.5-cm pieces andincubated in RPMI 1640 containing 10% heat-inactivated FCS and 5 mMEDTA for 15 min to remove epithelial cells. This step was performed in ashaking incubator at 37°C, and was repeated a further two times, with allsupernatant being discarded. The remaining tissue was digested twice usingRPMI 1640 containing 10% FCS, 15 mM HEPES, and 0.3 mg/ml Type IICollagenase/Dispase (Worthington Biochemical) for 1 h in a 37°C shakingincubator. LP cells were collected and layered on a Percoll gradient (Am-ersham Biosciences) and centrifuged at 600 � g for 20 min. The lympho-cyte-enriched population was recovered from the 40 to 75% interface. Cellsuspensions were stimulated with 100 ng/ml PMA and 1 �g/ml ionomycinfor 4 h in the presence of 20 �g/ml brefeldin A. After this in vitro stim-ulation, cells were stained for CD4 (clone H129.19) and CD45.1 (cloneA20) followed by fixation in 2% paraformaldehyde and permeabilization in0.5% saponin. This was followed by staining for intracellular IL-10 (cloneJES5-16E3), IFN-� (clone XMG1.2), or appropriate isotype controls(clones A95.1 and R3-34, respectively). For IL-10 and Foxp3 staining,cells were fixed in eBioscience Fix/Perm buffer after the in vitro stimula-tion. This was followed by permeabilization in 0.5% saponin and staining
5853The Journal of Immunology
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
for IL-10 as indicated above. Finally, the cells were permeabilized in eBio-science buffer and stained for Foxp3 according to the manufacturer’s in-structions. Cells were analyzed using a FACSCalibur or FACSort (BDBiosciences).
Statistics
Two-tailed Mann-Whitney U test was performed using GraphPad Prism3.00 (GraphPad). Values of p � 0.05 were regarded as significant. Data arepresented as mean � SD.
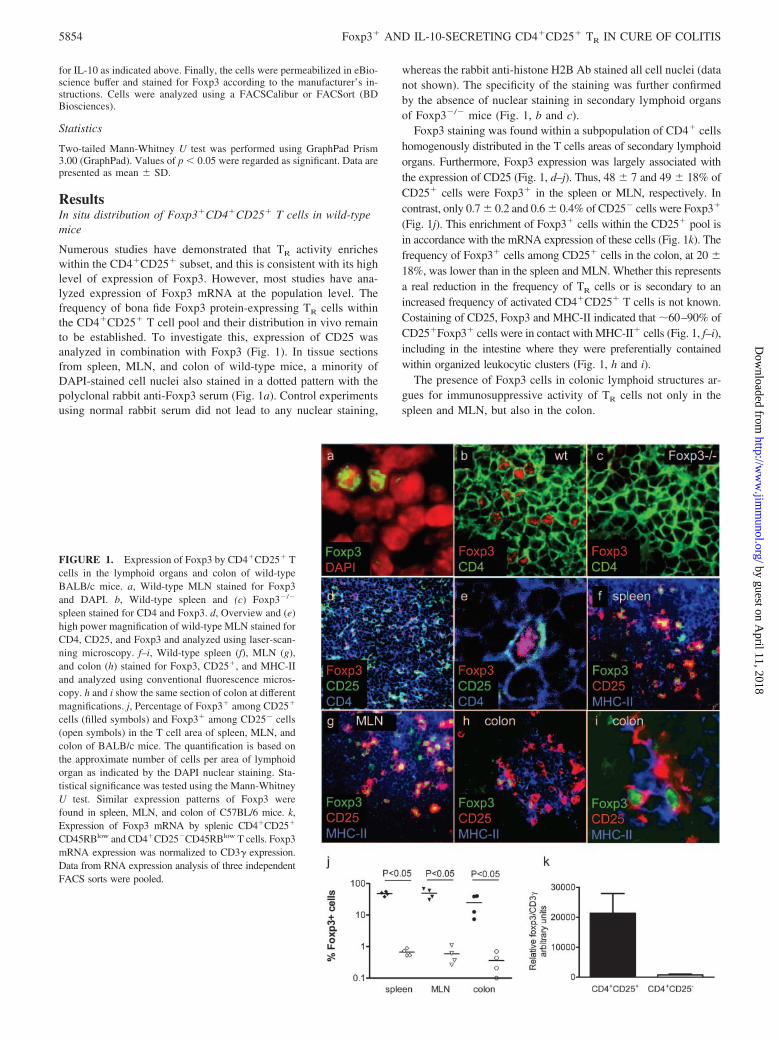
ResultsIn situ distribution of Foxp3�CD4�CD25� T cells in wild-typemice
Numerous studies have demonstrated that TR activity enricheswithin the CD4�CD25� subset, and this is consistent with its highlevel of expression of Foxp3. However, most studies have ana-lyzed expression of Foxp3 mRNA at the population level. Thefrequency of bona fide Foxp3 protein-expressing TR cells withinthe CD4�CD25� T cell pool and their distribution in vivo remainto be established. To investigate this, expression of CD25 wasanalyzed in combination with Foxp3 (Fig. 1). In tissue sectionsfrom spleen, MLN, and colon of wild-type mice, a minority ofDAPI-stained cell nuclei also stained in a dotted pattern with thepolyclonal rabbit anti-Foxp3 serum (Fig. 1a). Control experimentsusing normal rabbit serum did not lead to any nuclear staining,
whereas the rabbit anti-histone H2B Ab stained all cell nuclei (datanot shown). The specificity of the staining was further confirmedby the absence of nuclear staining in secondary lymphoid organsof Foxp3�/� mice (Fig. 1, b and c).
Foxp3 staining was found within a subpopulation of CD4� cellshomogenously distributed in the T cells areas of secondary lymphoidorgans. Furthermore, Foxp3 expression was largely associated withthe expression of CD25 (Fig. 1, d–j). Thus, 48 � 7 and 49 � 18% ofCD25� cells were Foxp3� in the spleen or MLN, respectively. Incontrast, only 0.7 � 0.2 and 0.6 � 0.4% of CD25� cells were Foxp3�
(Fig. 1j). This enrichment of Foxp3� cells within the CD25� pool isin accordance with the mRNA expression of these cells (Fig. 1k). Thefrequency of Foxp3� cells among CD25� cells in the colon, at 20 �18%, was lower than in the spleen and MLN. Whether this representsa real reduction in the frequency of TR cells or is secondary to anincreased frequency of activated CD4�CD25� T cells is not known.Costaining of CD25, Foxp3 and MHC-II indicated that �60–90% ofCD25�Foxp3� cells were in contact with MHC-II� cells (Fig. 1, f–i),including in the intestine where they were preferentially containedwithin organized leukocytic clusters (Fig. 1, h and i).
The presence of Foxp3 cells in colonic lymphoid structures ar-gues for immunosuppressive activity of TR cells not only in thespleen and MLN, but also in the colon.
FIGURE 1. Expression of Foxp3 by CD4�CD25� Tcells in the lymphoid organs and colon of wild-typeBALB/c mice. a, Wild-type MLN stained for Foxp3and DAPI. b, Wild-type spleen and (c) Foxp3�/�
spleen stained for CD4 and Foxp3. d, Overview and (e)high power magnification of wild-type MLN stained forCD4, CD25, and Foxp3 and analyzed using laser-scan-ning microscopy. f–i, Wild-type spleen (f), MLN (g),and colon (h) stained for Foxp3, CD25�, and MHC-IIand analyzed using conventional fluorescence micros-copy. h and i show the same section of colon at differentmagnifications. j, Percentage of Foxp3� among CD25�
cells (filled symbols) and Foxp3� among CD25� cells(open symbols) in the T cell area of spleen, MLN, andcolon of BALB/c mice. The quantification is based onthe approximate number of cells per area of lymphoidorgan as indicated by the DAPI nuclear staining. Sta-tistical significance was tested using the Mann-WhitneyU test. Similar expression patterns of Foxp3 werefound in spleen, MLN, and colon of C57BL/6 mice. k,Expression of Foxp3 mRNA by splenic CD4�CD25�
CD45RBlow and CD4�CD25�CD45RBlow T cells. Foxp3mRNA expression was normalized to CD3� expression.Data from RNA expression analysis of three independentFACS sorts were pooled.
5854 Foxp3� AND IL-10-SECRETING CD4�CD25� TR IN CURE OF COLITIS
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

In situ distribution of Foxp3�CD4�CD25� T cells during cureof colitis
We next investigated the distribution of Foxp3� cells followingcure of colitis by transfer of CD4�CD25� T cells. Colitis wasinduced in immune-deficient (C57BL/6J Rag�/�) mice by transferof wild-type (CD45.2�) CD4�CD45RBhigh T cells. After devel-opment of the first signs of colitis, indicated by weight loss anddiarrhea, a group of mice received a second transfer of congenicCD45.1�CD4�CD25� T cells. As previously described (20) thissecond transfer led to increased weight and resolution of colitis(Fig. 2). At the time of transfer, �90% of CD25� T cells ex-pressed Foxp3 (data not shown).
Two weeks after transfer of CD4�CD25� T cells, when colitiswas still present, �50% of the CD45.1� progeny of theCD4�CD25� T cells expressed Foxp3 irrespective of whetherspleen, MLN, or colon were analyzed (Fig. 2a and data notshown). Our data suggest that Foxp3�CD4�CD25� regulatorycells migrate not only to lymphoid organs but also into the in-flamed LP. Following resolution of colitis, 5–10 wk after the sec-ondary transfer of CD4�CD25� T cells, Foxp3 expression wasstill primarily restricted to the CD25� progeny and still detectablewithin �50% of these cells (Fig. 2b). Less than 2% of theCD4�CD45RBhigh progeny expressed Foxp3 at this time (n 4,data not shown), compared with �1% of donor CD45RBhigh Tcells directly ex vivo (n 3, data not shown). Furthermore, insimilar experiments performed in mice on the BALB/c back-ground, we again saw no increase in the percentage of Foxp3�
cells among the CD45RBhigh progeny. This suggests that cure ofcolitis in this model situation is not associated with the inductionof Foxp3 in the progeny of CD4�CD45RBhigh T cells.
Presence of Foxp3�IL-10� cells in the LP of the human colon
The T cell transfer model of colitis reflects several features of thehuman IBDs, UC and CD. However, the human diseases presentdistinct histological morphology and are of heterogeneous patho-genicity. Consequently, we investigated the distribution of Foxp3�
cells in human tissue. Within lymphoid organs, as well as in theappendix, Foxp3 expression was confined to the nuclei of CD4�
and CD3� cells (Fig. 3, a and e) and present predominantly in theT cell areas. In tissue samples from noninflamed human colons,and in patients with intestinal inflammation, nuclear Foxp3-posi-tive cells of lymphocyte morphology were present with the highestdensity in lymphoid follicles, although there were also scattered
cells within the LP (Fig. 3b). The total number of Foxp3� cellswas greater in inflamed tissue than in normal controls (Fig. 3c).This in part reflected the increase in CD3� cells and lymphoidfollicles in the inflamed colon (Fig. 3, c and d). There was noincrease in the actual density of Foxp3� cells in the T cell areas ininflamed vs uninflamed samples. By contrast there was a higherdensity of Foxp3� cells in the inflamed LP compared with con-trols. Together, the data suggest that inflammation is driving theaccumulation of regulatory cells, particularly in effector sites.
To analyze whether the accumulation of Foxp3� cells in in-flamed IBD LP is a specific feature of CD or UC or is due to ageneral accumulation of Foxp3� cells in inflamed intestinal tissue,we compared the density of Foxp3� cells in colon sections ofpatients with IBD with those with diverticulitis, pseudomembra-nous colitis, or CMV-induced colitis (CMV) (Fig. 3, b–d). In allcases we found an increased density of Foxp3� cells indicatingthat the accumulation of Foxp3� cells is driven by inflammation inthe intestine and is not a specific feature of IBD. The presence ofFoxp3� cells in patients with IBD argues against a simple lack ofFoxp3� TR as a cause of the intestinal inflammation, but for in-effective activity of these cells, or nonresponsiveness to their ac-tivity, under conditions of chronic intestinal inflammation.
To investigate whether there is an association of Foxp3 positivecells with IL-10 secretion within the human intestine, we stainedhuman appendix for IL-10 and Foxp3. IL-10-producing cells werefound in high density within the germinal center as well as in thesubepithelial dome area (Fig. 3e). We show the presence of IL-10-positive CD3�Foxp3� TR cells as well CD3�Foxp3� cells thatare IL-10 negative but are in close contact with IL-10-positiveCD3�Foxp3� cells (Fig. 3e). This indicates that some Foxp3�
cells secrete IL-10 within the intestine but that these cells are fre-quently in close contact with IL-10-producing non-T cells(CD3�Foxp3� cells).
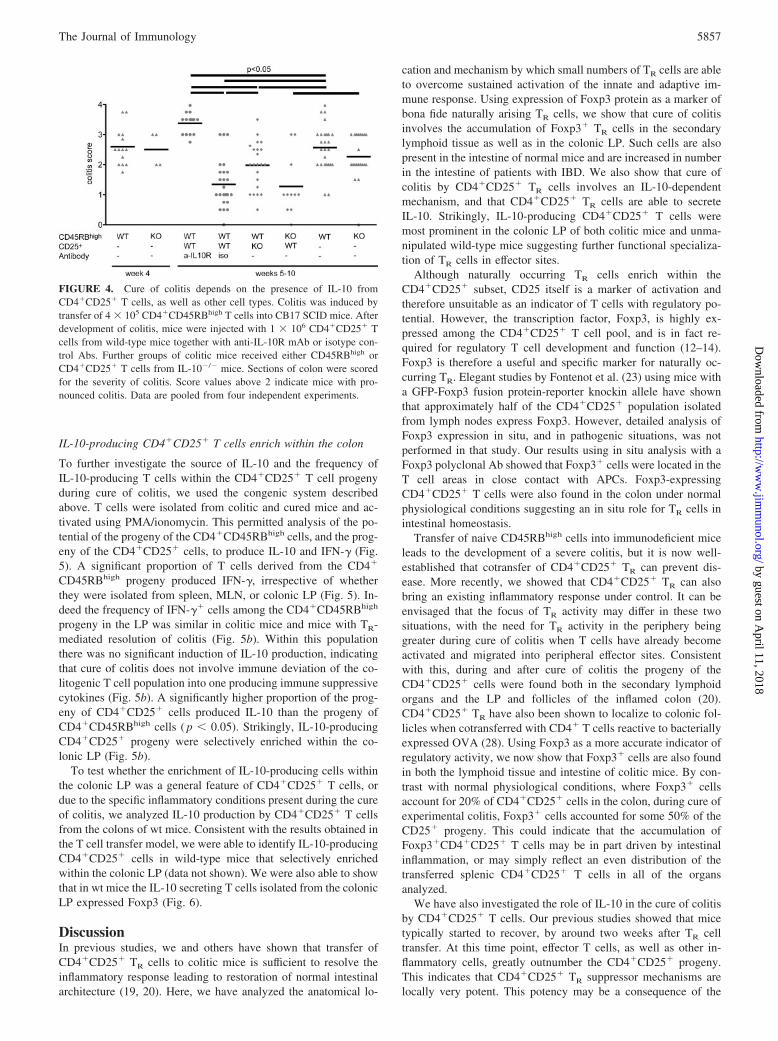
Cure of colitis is dependent on IL-10
We have described the presence of Foxp3�CD4�CD25� T cells inboth the secondary lymphoid organs and colonic LP during cure ofexperimental colitis, suggesting that regulation occurs at both sites.We next decided to further investigate the mechanism by whichCD4�CD25� T cells cure colitis. Again, we transferred CD4�
CD25� cells into animals with established T cell-induced colitis.However, if the transfer of CD4�CD25� cells was combined
FIGURE 2. Presence of Foxp3�CD4�CD25� Tcells in the colons of mice with colitis and mice curedof colitis. rag-1�/� mice were injected with 4 � 105
wild-type CD45.2� CD4�CD45RBhigh T cells. Fourweeks after the T cell transfer, wasting disease and co-litis were evident, and a second transfer of 1 � 106
congenic CD45.1�CD4�CD25� T cells was per-formed. Sections of colon taken either 2 (a) or 10 (b)wk after the second T cell transfer were stained withH&E, Abs to CD4 and CD45.1, or Abs to Foxp3 andCD45.1. DAPI counterstaining was performed to visu-alize the tissue structure. E indicates epithelium. Thepresented micrographs are representative of three tofive mice analyzed.
5855The Journal of Immunology
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

with injection of a neutralizing anti-IL-10R mAb, mice contin-ued to lose weight and remained colitic (Fig. 4). Many differentcell types have been shown to be able to produce IL-10. Toinvestigate whether the IL-10 needs to be secreted by theCD4�CD25� T cell progeny a third group of colitic mice re-ceived CD4�CD25� T cells from IL-10�/� mice. Interestingly,although CD4�CD25�IL-10�/� T cells were less effective atcontrolling intestinal inflammation than wild-type CD4�
CD25� cells they did lead to some amelioration of disease (Fig.4). The data demonstrates that the presence of IL-10 is requiredfor treatment of colitis, and also suggests that functionallyrelevant IL-10 may derive from both TR and non-TR sources.However, wild-type CD4�CD25� cells were equally effectiveat curing colitis induced by wild-type or IL-10� /�
CD4�CD45RBhigh cells, suggesting that IL-10 production byCD4�CD45RBhigh progeny is not critical for cure of colitis.
FIGURE 3. Presence of Foxp3-positive cells in the mucosa and intestinal lymphoid follicles of patients with IBD. Colon samples from normal controlsand patients with ulcerative colitis (UC), Crohn’s disease (CD), non-IBD inflammations of the colon (diverticulitis (D), pseudomembranous colitis (PC),and CMV-induced colitis (CMV)) were analyzed for CD3, CD4, and Foxp3. Sections from tonsil were analyzed for CD4 and Foxp3. a, Costaining for CD4and Foxp3 on human tonsil. Overview (�100) and high-power magnification. b, CD3 and Foxp3 staining on colon tissue. c, Density of CD3� and Foxp3�
cells within colon. Numbers of cells were quantified per area at �400 magnification. Each data point represents one patient. d, Density of Foxp3� cellswithin colonic LP, as well as T and B cell areas of mucosa-associated lymphoid tissue. Each data point represents one patient. e, Costaining of Foxp3 andIL-10 on human appendix tissue. Left, Costaining of Foxp3, IL-10, and DAPI showing IL-10-positive cells within the germinal center (GC) and subep-ithelial dome (SED) close to the epithelium (E). Detail of Foxp3�IL-10� cells within the subepithelial dome area are also shown. Right, Costaining ofFoxp3, IL-10, and CD3. Detail shows IL-10�CD3�Foxp3� cells as well CD3�Foxp3� cells that are in close contact with IL-10�CD3�Foxp3� cells.
5856 Foxp3� AND IL-10-SECRETING CD4�CD25� TR IN CURE OF COLITIS
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
IL-10-producing CD4�CD25� T cells enrich within the colon
To further investigate the source of IL-10 and the frequency ofIL-10-producing T cells within the CD4�CD25� T cell progenyduring cure of colitis, we used the congenic system describedabove. T cells were isolated from colitic and cured mice and ac-tivated using PMA/ionomycin. This permitted analysis of the po-tential of the progeny of the CD4�CD45RBhigh cells, and the prog-eny of the CD4�CD25� cells, to produce IL-10 and IFN-� (Fig.5). A significant proportion of T cells derived from the CD4�
CD45RBhigh progeny produced IFN-�, irrespective of whetherthey were isolated from spleen, MLN, or colonic LP (Fig. 5). In-deed the frequency of IFN-�� cells among the CD4�CD45RBhigh
progeny in the LP was similar in colitic mice and mice with TR-mediated resolution of colitis (Fig. 5b). Within this populationthere was no significant induction of IL-10 production, indicatingthat cure of colitis does not involve immune deviation of the co-litogenic T cell population into one producing immune suppressivecytokines (Fig. 5b). A significantly higher proportion of the prog-eny of CD4�CD25� cells produced IL-10 than the progeny ofCD4�CD45RBhigh cells ( p � 0.05). Strikingly, IL-10-producingCD4�CD25� progeny were selectively enriched within the co-lonic LP (Fig. 5b).
To test whether the enrichment of IL-10-producing cells withinthe colonic LP was a general feature of CD4�CD25� T cells, ordue to the specific inflammatory conditions present during the cureof colitis, we analyzed IL-10 production by CD4�CD25� T cellsfrom the colons of wt mice. Consistent with the results obtained inthe T cell transfer model, we were able to identify IL-10-producingCD4�CD25� cells in wild-type mice that selectively enrichedwithin the colonic LP (data not shown). We were also able to showthat in wt mice the IL-10 secreting T cells isolated from the colonicLP expressed Foxp3 (Fig. 6).
DiscussionIn previous studies, we and others have shown that transfer ofCD4�CD25� TR cells to colitic mice is sufficient to resolve theinflammatory response leading to restoration of normal intestinalarchitecture (19, 20). Here, we have analyzed the anatomical lo-
cation and mechanism by which small numbers of TR cells are ableto overcome sustained activation of the innate and adaptive im-mune response. Using expression of Foxp3 protein as a marker ofbona fide naturally arising TR cells, we show that cure of colitisinvolves the accumulation of Foxp3� TR cells in the secondarylymphoid tissue as well as in the colonic LP. Such cells are alsopresent in the intestine of normal mice and are increased in numberin the intestine of patients with IBD. We also show that cure ofcolitis by CD4�CD25� TR cells involves an IL-10-dependentmechanism, and that CD4�CD25� TR cells are able to secreteIL-10. Strikingly, IL-10-producing CD4�CD25� T cells weremost prominent in the colonic LP of both colitic mice and unma-nipulated wild-type mice suggesting further functional specializa-tion of TR cells in effector sites.
Although naturally occurring TR cells enrich within theCD4�CD25� subset, CD25 itself is a marker of activation andtherefore unsuitable as an indicator of T cells with regulatory po-tential. However, the transcription factor, Foxp3, is highly ex-pressed among the CD4�CD25� T cell pool, and is in fact re-quired for regulatory T cell development and function (12–14).Foxp3 is therefore a useful and specific marker for naturally oc-curring TR. Elegant studies by Fontenot et al. (23) using mice witha GFP-Foxp3 fusion protein-reporter knockin allele have shownthat approximately half of the CD4�CD25� population isolatedfrom lymph nodes express Foxp3. However, detailed analysis ofFoxp3 expression in situ, and in pathogenic situations, was notperformed in that study. Our results using in situ analysis with aFoxp3 polyclonal Ab showed that Foxp3� cells were located in theT cell areas in close contact with APCs. Foxp3-expressingCD4�CD25� T cells were also found in the colon under normalphysiological conditions suggesting an in situ role for TR cells inintestinal homeostasis.
Transfer of naive CD45RBhigh cells into immunodeficient miceleads to the development of a severe colitis, but it is now well-established that cotransfer of CD4�CD25� TR can prevent dis-ease. More recently, we showed that CD4�CD25� TR can alsobring an existing inflammatory response under control. It can beenvisaged that the focus of TR activity may differ in these twosituations, with the need for TR activity in the periphery beinggreater during cure of colitis when T cells have already becomeactivated and migrated into peripheral effector sites. Consistentwith this, during and after cure of colitis the progeny of theCD4�CD25� cells were found both in the secondary lymphoidorgans and the LP and follicles of the inflamed colon (20).CD4�CD25� TR have also been shown to localize to colonic fol-licles when cotransferred with CD4� T cells reactive to bacteriallyexpressed OVA (28). Using Foxp3 as a more accurate indicator ofregulatory activity, we now show that Foxp3� cells are also foundin both the lymphoid tissue and intestine of colitic mice. By con-trast with normal physiological conditions, where Foxp3� cellsaccount for 20% of CD4�CD25� cells in the colon, during cure ofexperimental colitis, Foxp3� cells accounted for some 50% of theCD25� progeny. This could indicate that the accumulation ofFoxp3�CD4�CD25� T cells may be in part driven by intestinalinflammation, or may simply reflect an even distribution of thetransferred splenic CD4�CD25� T cells in all of the organsanalyzed.
We have also investigated the role of IL-10 in the cure of colitisby CD4�CD25� T cells. Our previous studies showed that micetypically started to recover, by around two weeks after TR celltransfer. At this time point, effector T cells, as well as other in-flammatory cells, greatly outnumber the CD4�CD25� progeny.This indicates that CD4�CD25� TR suppressor mechanisms arelocally very potent. This potency may be a consequence of the
FIGURE 4. Cure of colitis depends on the presence of IL-10 fromCD4�CD25� T cells, as well as other cell types. Colitis was induced bytransfer of 4 � 105 CD4�CD45RBhigh T cells into CB17 SCID mice. Afterdevelopment of colitis, mice were injected with 1 � 106 CD4�CD25� Tcells from wild-type mice together with anti-IL-10R mAb or isotype con-trol Abs. Further groups of colitic mice received either CD45RBhigh orCD4�CD25� T cells from IL-10�/� mice. Sections of colon were scoredfor the severity of colitis. Score values above 2 indicate mice with pro-nounced colitis. Data are pooled from four independent experiments.
5857The Journal of Immunology
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

secretion of high concentrations of immunosuppressive cytokines,or instruction of effector cells to produce such cytokines. Indeed,previous studies have demonstrated the necessity of IL-10 for thecure of T cell-induced colitis, and of CD4�CD25� T cell-derivedIL-10 for inhibition of colonic inflammation and dysplasia (19,29). Our own results confirm IL-10 is required for the resolution ofestablished colitis, and furthermore demonstrate that a proportionof the CD4�CD25� cell progeny in the colons of treated miceproduced IL-10. Similarly, recent studies have demonstrated thepresence of IL-10-producing CD4�CD25� T cells in prediabeticand Leishmania major lesions, and in the CNS in EAE (30–32).However, we also show that IL-10�/� CD4�CD25� T cells are,although less potent, able to cure colitis. It would appear that al-though TR are a major source of the IL-10 required for cure ofcolitis, other cell types, possibly under the instruction of TR, canmake an important contribution. Indeed, experiments investigatingthe protective role of IL-10 in schistosomiasis have shown that
both innate immune cells and T cells contribute to IL-10 produc-tion (33). Many different cell types, including epithelial cells, mac-rophages, DC, and other T cells subsets, have been shown to pro-duce IL-10. For example, naive T cells can differentiate into IL-10producers with regulatory function both in vitro and in vivo (2).Importantly, such cells are also able to cure colitis in the T celltransfer model (21). However, we do not consider the naive T cellpopulation to be an essential source of IL-10 in our model. This isbased on the finding that colitis induced by IL-10�/�CD45RBhigh
T cells was efficiently cured by CD4�CD25�TR cells.Foxp3� TR cells can also arise from naive progenitors in vivo
following chronic exposure to low doses of Ag (34) or in vitrowhen activated in the presence of TGF-� (24, 35–37). However,we found no evidence that resolution of intestinal inflammation, atleast in the T cell transfer model, was associated with significantimmune deviation of CD4�CD45RBhigh progeny into eitherFoxp3�- or IL-10-producing TR cells. Cure of colitis was also not
FIGURE 5. Accumulation of the IL-10-producingprogeny of CD4�CD25� T cells within the colon. rag-1�/� mice were injected with 4 � 105 wild-typeCD45.2� CD4�CD45RBhigh T cells. After develop-ment of colitis, mice received a second transfer of 1 �106 congenic CD45.1� CD4�CD25� T cells. After 4wk, the ability of CD4�CD45RBhigh (n 6) andCD4�CD25� (n 6) progeny to produce IL-10 andIFN-� was analyzed. Lymphocytes were prepared fromspleen, MLN, and the colonic LP and stimulated withPMA/ionomycin and brefeldin A. The results of twoindependent experiments are pooled. a, RepresentativeFACS plot showing preferential IL-10 production byLP CD4�CD25� T cell progeny (CD45.1�) and pref-erential IFN-� production by the CD4�CD45RBhigh Tcell progeny (CD45.1�). b, Production of IL-10 andIFN-� by CD4�CD25� T cell progeny and comparisonof IL-10 and IFN-� production by CD4�CD45RBhigh Tcell progeny in the presence or absence of CD4�
CD25� T cells. Data are shown for spleen, MLN, andLP. Significance was tested using the Mann-Whitney Utest; n.s., not significant.
5858 Foxp3� AND IL-10-SECRETING CD4�CD25� TR IN CURE OF COLITIS
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

dependent on a reduction in the proportion of IFN-�-secreting cellsamong the CD4�CD45RBhigh progeny, although, as reported pre-viously, the total number of CD4� T cells present in the colon wasgreatly reduced (20).
We have previously demonstrated that IL-10 is not required forthe prevention of colitis by CD4�CD25� TR. However, when thewhole CD4�CD45RBlow fraction is transferred instead of CD4�
CD25� TR, prevention of colitis becomes dependent on IL-10. Inthis situation, TR-derived IL-10 appears to be essential for thecontrol of colitogenic cells contained within the Ag-experiencedCD45RBlow pool, but not for control of naive T cells (27). Thefinding that IL-10 is essential for cure of colitis, where a largepopulation of Ag-experienced cells are present, is consistentwith this.
We have shown that the IL-10-producing subset of CD25� se-lectively enriches within the colon of cured mice. The population
of TR used in our model was isolated from the spleen and as suchexpressed only low levels of IL-10 upon transfer. It is thereforepossible that the gut environment conditioned the TR to furtherdifferentiate into IL-10-producing cells. Alternatively, the accumu-lation of IL-10� TR in the colon could be attributed to the pref-erential expansion and homing of a minor subset of IL-10 produc-ers contained within the transferred population. The inability ofCD4�CD25� cells isolated from peripheral lymphoid organs tosecrete IL-10 in vitro has led to the notion that naturally arisingFoxp3� TR cells function via cytokine independent mechanisms(38). However, our finding that IL-10�Foxp3� cells are enrichedwithin tissue sites illustrates the problems with extrapolating ob-servations in vitro to predicted function in vivo. The identificationof IL-10-producing CD4�CD25� T cells in the normal colon sug-gests a constitutive role for those cells in the prevention of intes-tinal inflammatory responses. Our data is supported by the earlierobservation by Cong et al. (39) that bacterially reactive IL-10-secreting CD4� T cells are present in the colons of normal mice.However, that study did not establish whether such bacterially re-active cells belonged to the naturally occurring Foxp3� TR pool, oracquired regulatory activity in the periphery. We now show thatIL-10-secreting CD4�CD25� TR in the normal colon are con-tained within the Foxp3� subset. It is however likely that undercertain conditions, other IL-10-producing TR cells that do not ex-press Foxp3 may be induced (40).
Human IBDs consist of two dominant disease subtypes. CD islargely Th1 in nature (41), while IL-5-, and IL-13-producing T orNKT cells are found in UC (42). Although there is evidence for Tcell-mediated immune dysregulation in human IBD, there is lim-ited evidence that disease results from a primary lack or dysfunc-tion of TR cells. However, clear evidence that a lack of function-ally sufficient TR cells can lead to chronic intestinal inflammationin humans does come from the study of patients with immunepolyendocrine X-linked enteropathy syndrome (17, 18). CD4�
CD25� Foxp3 mRNA expressing T cells have recently been iso-lated from human intestinal tissue samples, and CD4�CD25� Tcells from the colons of IBD patients have been shown to be sup-pressive in vitro (43, 44). Our data show that Foxp3-positive cellsare present within the noninflamed LP, and are also present inincreased numbers in patients with acute UC and CD. The increasein Foxp3� cell density was seen largely in the LP, rather than inthe T cell areas of isolated lymphoid follicles. This can be ex-plained by considering the T cell areas of the isolated lymphoidfollicles to be sites of induction of immune responses, analogous tothose in the lymph node. Therefore, during an active inflammatoryresponse, TR may be expected to be preferentially recruited to thesite of inflammation. A recent study also found an increase inCD25�Foxp3� cells in the colon of UC and CD patients, althoughthis increase was modest compared with that seen in diverticulitis(45). In our study, a similar increase in Foxp3� cells is seen in UC,CD, and diverticulitis, but also in CMV colitis and pseudomem-branous colitis, strongly suggesting that chronic intestinal inflam-mation is not simply a consequence of the absence of Foxp3�
CD4� TR cells at the site of inflammation. Whether the TR cellspresent have, at least in some patients, impaired intrinsic immu-nosuppressive activity, or whether the inflamed tissue environmentis resistant to immunosuppression by TR cells remains to be in-vestigated. In any case, our data on human Foxp3� cells supportsthe hypothesis that intestinal immune regulation in mice and hu-mans may share common features. Consequently, further investi-gation of the mechanisms by which TR can cure experimental co-litis may give us insight into how regulatory T cell function mightbe enhanced in human IBD.
FIGURE 6. IL-10 secretion by Foxp3� cells in the LP of wild-typemice. Lamina propria and spleen cell suspensions were restimulated withPMA/ionomycin as described in Materials and Methods. a, Representativedot plots showing Foxp3 expression against IL-10 or an isotype control inspleen and LP of B6 SJL CD45 congenic mice. Plots are gated on CD4�
cells. The numbers indicate the percentage of IL-10� cells in the Foxp3�
or Foxp3� populations. b, Percentage of IL-10-secreting cells in theFoxp3� (filled symbols) and Foxp3� (open symbols) populations fromspleen (Sp) and LP CD4� lymphocytes (LPL) from B6 SJL CD45 con-genic mice. Each symbol represents data from an individual mouse. In allcases, the isotype control stained �1% of total cells. Similar results wereobtained in an experiment using BALB/c mice.
5859The Journal of Immunology
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

AcknowledgmentsWe thank animal facility personnel, Liz Darley, Sandy Laue, and MartinaFuegenschuh for histology, Nigel Rust for cell sorting, and Kevin Maloyfor critical reading of the manuscript.
DisclosuresThe authors have no financial conflict of interest.
References1. Maloy, K. J., and F. Powrie. 2001. Regulatory T cells in the control of immune
pathology. Nat. Immunol. 2: 816–822.2. O’Garra, A., P. L. Vieira, P. Vieira, and A. E. Goldfeld. 2004. IL-10-producing
and naturally occurring CD4� Tregs: limiting collateral damage. J. Clin. Invest.114: 1372–1378.
3. Sakaguchi, S. 2004. Naturally arising CD4� regulatory T cells for immunologicself-tolerance and negative control of immune responses. Annu. Rev. Immunol.22: 531–562.
4. Coombes, J. L., N. J. Robinson, K. J. Maloy, H. H. Uhlig, and F. Powrie. 2005.Regulatory T cells and intestinal homeostasis. Immunol. Rev. 204: 184–194.
5. Read, S., V. Malmstrom, and F. Powrie. 2000. Cytotoxic T lymphocyte-associ-ated antigen 4 plays an essential role in the function of CD25�CD4� regulatorycells that control intestinal inflammation. J. Exp. Med. 192: 295–302.
6. Kriegel, M. A., T. Lohmann, C. Gabler, N. Blank, J. R. Kalden, andH. M. Lorenz. 2004. Defective suppressor function of human CD4�CD25� reg-ulatory T cells in autoimmune polyglandular syndrome type II. J. Exp. Med. 199:1285–1291.
7. Viglietta, V., C. Baecher-Allan, H. L. Weiner, and D. A. Hafler. 2004. Loss offunctional suppression by CD4�CD25� regulatory T cells in patients with mul-tiple sclerosis. J. Exp. Med. 199: 971–979.
8. Baecher-Allan, C., V. Viglietta, and D. A. Hafler. 2004. Human CD4�CD25�
regulatory T cells. Semin. Immunol. 16: 89–98.9. Kukreja, A., G. Cost, J. Marker, C. Zhang, Z. Sun, K. Lin-Su, S. Ten, M. Sanz,
M. Exley, B. Wilson, et al. 2002. Multiple immuno-regulatory defects in type-1diabetes. J. Clin. Invest. 109: 131–140.
10. Ehrenstein, M. R., J. G. Evans, A. Singh, S. Moore, G. Warnes, D. A. Isenberg,and C. Mauri. 2004. Compromised function of regulatory T cells in rheumatoidarthritis and reversal by anti-TNF� therapy. J. Exp. Med. 200: 277–285.
11. Bacchetta, R., L. Passerini, E. Gambineri, M. Dai, S. E. Allan, L. Perroni,F. Dagna-Bricarelli, C. Sartirana, S. Matthes-Martin, A. Lawitschka, et al. 2006.Defective regulatory and effector T cell functions in patients with FOXP3 mu-tations. J. Clin. Invest. 116: 1713–1722.
12. Fontenot, J. D., M. A. Gavin, and A. Y. Rudensky. 2003. Foxp3 programs thedevelopment and function of CD4�CD25� regulatory T cells. Nat. Immunol. 4:330–336.
13. Hori, S., T. Nomura, and S. Sakaguchi. 2003. Control of regulatory T Cell de-velopment by the transcription factor FOXP3. Science 299: 1057–1061.
14. Khattri, R., T. Cox, S. A. Yasayko, and F. Ramsdell. 2003. An essential role forScurfin in CD4�CD25� T regulatory cells. Nat. Immunol. 4: 337–342.
15. Godfrey, V. L., J. E. Wilkinson, and L. B. Russell. 1991. X-linked lymphore-ticular disease in the scurfy (sf) mutant mouse. Am. J. Pathol. 138: 1379–1387.
16. Brunkow, M. E., E. W. Jeffery, K. A. Hjerrild, B. Paeper, L. B. Clark,S. A. Yasayko, J. E. Wilkinson, D. Galas, S. F. Ziegler, and F. Ramsdell. 2001.Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatallymphoproliferative disorder of the scurfy mouse. Nat. Genet. 27: 68–73.
17. Wildin, R. S., F. Ramsdell, J. Peake, F. Faravelli, J. L. Casanova, N. Buist,E. Levy-Lahad, M. Mazzella, O. Goulet, L. Perroni, et al. 2001. X-linked neo-natal diabetes mellitus, enteropathy and endocrinopathy syndrome is the humanequivalent of mouse scurfy. Nat. Genet. 27: 18–20.
18. Bennett, C. L., J. Christie, F. Ramsdell, M. E. Brunkow, P. J. Ferguson,L. Whitesell, T. E. Kelly, F. T. Saulsbury, P. F. Chance, and H. D. Ochs. 2001.The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syn-drome (IPEX) is caused by mutations of FOXP3. Nat. Genet. 27: 20–21.
19. Liu, H., B. Hu, D. Xu, and F. Y. Liew. 2003. CD4�CD25� regulatory T cellscure murine colitis: the role of IL-10, TGF-�, and CTLA4. J. Immunol. 171:5012–5017.
20. Mottet, C., H. H. Uhlig, and F. Powrie. 2003. Cutting edge: cure of colitis byCD4�CD25� regulatory T cells. J. Immunol. 170: 3939–3943.
21. Foussat, A., F. Cottrez, V. Brun, N. Fournier, J. P. Breittmayer, and H. Groux.2003. A comparative study between T regulatory type 1 and CD4�CD25� T cellsin the control of inflammation. J. Immunol. 171: 5018–5026.
22. Tang, Q., K. J. Henriksen, M. Bi, E. B. Finger, G. Szot, J. Ye, E. L. Masteller,H. McDevitt, M. Bonyhadi, and J. A. Bluestone. 2004. In vitro-expanded antigen-specific regulatory T cells suppress autoimmune diabetes. J. Exp. Med. 199:1455–1465.
23. Fontenot, J. D., J. P. Rasmussen, L. M. Williams, J. L. Dooley, A. G. Farr, andA. Y. Rudensky. 2005. Regulatory T cell lineage specification by the forkheadtranscription factor foxp3. Immunity 22: 329–341.
24. Wan, Y. Y., and R. A. Flavell. 2005. Identifying Foxp3-expressing suppressor Tcells with a bicistronic reporter. Proc. Natl. Acad. Sci. USA 102: 5126–5131.
25. Cobbold, S. P., A. Jayasuriya, A. Nash, T. D. Prospero, and H. Waldmann. 1984.Therapy with monoclonal antibodies by elimination of T-cell subsets in vivo.Nature 312: 548–551.
26. Coffman, R. L. 1982. Surface antigen expression and immunoglobulin gene re-arrangement during mouse pre-B cell development. Immunol. Rev. 69: 5–23.
27. Asseman, C., S. Read, and F. Powrie. 2003. Colitogenic Th1 cells are present inthe antigen-experienced T cell pool in normal mice: control by CD4� regulatoryT cells and IL-10. J. Immunol. 171: 971–978.
28. Watanabe, T., M. Yamori, T. Kita, T. Chiba, and Y. Wakatsuki. 2005.CD4�CD25� T cells regulate colonic localization of CD4 T cells reactive to amicrobial antigen. Inflamm. Bowel Dis. 11: 541–550.
29. Erdman, S. E., V. P. Rao, T. Poutahidis, M. M. Ihrig, Z. Ge, Y. Feng,M. Tomczak, A. B. Rogers, B. H. Horwitz, and J. G. Fox. 2003. CD4�CD25�
regulatory lymphocytes require interleukin 10 to interrupt colon carcinogenesis inmice. Cancer Res. 63: 6042–6050.
30. McGeachy, M. J., L. A. Stephens, and S. M. Anderton. 2005. Natural recoveryand protection from autoimmune encephalomyelitis: contribution ofCD4�CD25� regulatory cells within the central nervous system. J. Immunol.175: 3025–3032.
31. Herman, A. E., G. J. Freeman, D. Mathis, and C. Benoist. 2004. CD4�CD25� Tregulatory cells dependent on ICOS promote regulation of effector cells in theprediabetic lesion. J. Exp. Med. 199: 1479–1489.
32. Belkaid, Y., C. A. Piccirillo, S. Mendez, E. M. Shevach, and D. L. Sacks. 2002.CD4�CD25� regulatory T cells control Leishmania major persistence and im-munity. Nature 420: 502–507.
33. Hesse, M., C. A. Piccirillo, Y. Belkaid, J. Prufer, M. Mentink-Kane, M. Leusink,A. W. Cheever, E. M. Shevach, and T. A. Wynn. 2004. The pathogenesis ofschistosomiasis is controlled by cooperating IL-10-producing innate effector andregulatory T cells. J. Immunol. 172: 3157–3166.
34. Apostolou, I., and H. von Boehmer. 2004. In vivo instruction of suppressor com-mitment in naive T cells. J. Exp. Med. 199: 1401–1408.
35. Cobbold, S. P., R. Castejon, E. Adams, D. Zelenika, L. Graca, S. Humm, andH. Waldmann. 2004. Induction of foxP3� regulatory T cells in the periphery ofT cell receptor transgenic mice tolerized to transplants. J. Immunol. 172:6003–6010.
36. Chen, W., W. Jin, N. Hardegen, K. J. Lei, L. Li, N. Marinos, G. McGrady, andS. M. Wahl. 2003. Conversion of peripheral CD4�CD25� naive T cells toCD4�CD25� regulatory T cells by TGF-� induction of transcription factorFoxp3. J. Exp. Med. 198: 1875–1886.
37. Fantini, M. C., C. Becker, G. Monteleone, F. Pallone, P. R. Galle, andM. F. Neurath. 2004. Cutting edge: TGF-� induces a regulatory phenotype inCD4�CD25� T cells through Foxp3 induction and down-regulation of Smad7.J. Immunol. 172: 5149–5153.
38. Bluestone, J. A., and A. K. Abbas. 2003. Natural versus adaptive regulatory Tcells. Nat. Rev. Immunol. 3: 253–257.
39. Cong, Y., C. T. Weaver, A. Lazenby, and C. O. Elson. 2002. Bacterial-reactiveT regulatory cells inhibit pathogenic immune responses to the enteric flora. J. Im-munol. 169: 6112–6119.
40. Vieira, P. L., J. R. Christensen, S. Minaee, E. J. O’Neill, F. J. Barrat, A. Boonstra,T. Barthlott, B. Stockinger, D. C. Wraith, and A. O’Garra. 2004. IL-10-secretingregulatory T cells do not express Foxp3 but have comparable regulatory functionto naturally occurring CD4�CD25� regulatory T cells. J. Immunol. 172:5986–5993.
41. Bouma, G., and W. Strober. 2003. The immunological and genetic basis of in-flammatory bowel disease. Nat. Rev. Immunol. 3: 521–533.
42. Fuss, I. J., F. Heller, M. Boirivant, F. Leon, M. Yoshida, S. Fichtner-Feigl,Z. Yang, M. Exley, A. Kitani, R. S. Blumberg, et al. 2004. Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response inulcerative colitis. J. Clin. Invest. 113: 1490–1497.
43. Makita, S., T. Kanai, S. Oshima, K. Uraushihara, T. Totsuka, T. Sawada,T. Nakamura, K. Koganei, T. Fukushima, and M. Watanabe. 2004.CD4�CD25bright T cells in human intestinal lamina propria as regulatory cells.J. Immunol. 173: 3119–3130.
44. Kelsen, J., J. Agnholt, H. J. Hoffmann, J. L. Romer, C. L. Hvas, andJ. F. Dahlerup. 2005. FoxP3�CD4�CD25� T cells with regulatory properties canbe cultured from colonic mucosa of patients with Crohn’s disease. Clin. Exp.Immunol. 141: 549–557.
45. Maul, J., C. Loddenkemper, P. Mundt, E. Berg, T. Giese, A. Stallmach, M. Zeitz,and R. Duchmann. 2005. Peripheral and intestinal regulatory CD4� CD25high Tcells in inflammatory bowel disease. Gastroenterology 128: 1868–1878.
5860 Foxp3� AND IL-10-SECRETING CD4�CD25� TR IN CURE OF COLITIS
by guest on April 11, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Related Documents











