*Laboratoire de Pharmacologie Expe ´rimentale, Universite ´ catholique de Louvain, Brussels, Belgium Neuroanatomy, Department of Neuroscience, Biomedical Center, Uppsala, Sweden Throughout the brain and the spinal cord, the grey matter is characterized by a high density of neuronal cells connected by dense networks of dendrites and short non-myelinated axons. White matter is mainly composed of longer myelin- ated axons that ensure communication between clusters of grey matter. Mainly recognized as protoplasmic astroglial cells (designed as type-1 astrocytes), grey matter astrocytes are thought to actively cooperate with neurons in synaptic networking. In contrast, white matter astrocytes, commonly adopting a fibrous morphology (also called type-2 astro- cytes), have been assigned a supportive role for the propagation of electrical impulses through long distance projecting axons (Miller and Raff 1984; Hertz and Zielke 2004). As white matter is largely devoid of typical post- synaptic targets, the local release of excitatory or inhibitory transmitters has received limited consideration. However, recent neurochemical studies focusing on the corpus callo- sum, a white matter bundle supporting inter-hemispheric communication, have revealed a substantial vesicular release of glutamate from unmyelinated axons that efficiently activates glutamatergic receptors detected in glial progenitors (Kukley et al. 2007; Ziskin et al. 2007). Possibly contributing Received July 30, 2008; revised manuscript received December 16, 2008; accepted December 22, 2008. Address correspondence and reprint requests to Emmanuel Hermans, Laboratoire de Pharmacologie Expe ´rimentale, Universite ´ catholique de Louvain, Brussels, Belgium. E-mail: [email protected] Abbreviations used: CHO, Chinese hamster ovary; CPCCOEt, 7-hydroxyiminocyclopropan[b]chromen-1a-carboxylic acid ethyl ester; Cy TM 3, cyanin 3; DAPI, 4¢,6-diamidino-2-phenylindole dilactate; DHK, dihydrokainic acid; DHPG, (S)-3,5-dihydroxyphenylglycine; FBS, foetal bovine serum; GFAP, glial fibrillary acidic protein; GLAST, glutamate– aspartate transporter; GLT, glutamate transporters; GS, glutamine synthetase; IPTG, isopropyl-b-D-thiogalactoside; L-SOS, L-serine O-sulphate potassium salt; LTHA, L-())-threo-3-hydroxyaspartic acid; mGlu5 receptor, type 5 metabotropic glutamate receptor; MPEP, 2-methyl-6-(phenylethynyl)-pyridine. Abstract While the astrocytic control of extracellular glutamate con- centration at synaptic contacts is well characterized, little is known regarding the clearance of glutamate along axon tracts, even though local excitotoxic damage has been reported. Therefore, we have compared glutamate handling in astrocyte cultures derived from white matter (corpus callosum) and grey matter tissues (cortical structures). These populations of as- trocytes showed clearly distinct phenotypes, adopting stellate or protoplasmic morphologies respectively. In addition, white matter astrocytes showed high densities of the intermediate filament proteins glial fibrillary acidic protein, vimentin and nestin. The glutamate–aspartate transporter and glutamate transporter-1, as well as glutamine synthetase, were found to be expressed at higher levels in white matter compared with grey matter astrocytes. Consistent with this aspartate uptake capacity was three to fourfold higher in white matter cells, and the use of specific inhibitors revealed a substantial activity of glutamate transporter-1, contrasting with grey matter cells where this transporter appeared poorly functional. In addition, expression of type 5 metabotropic glutamate receptors was considerably higher in white matter astrocytes where the agonist (S)-3,5-dihydroxyphenylglycine triggered a large re- lease of intracellular calcium. Differences in these astrocyte cultures were also observed when exposed to experimental conditions that trigger glial activation. This study highlights typical features of cultured astrocytes derived from white matter tissues, which appear constitutively adapted to handle excitotoxic insults. Moreover, the expression and activity of the astroglial components involved in the control of glutama- tergic transmission are reinforced when these cells are maintained under conditions mimicking a gliotic environment. Keywords: astroglial activation, glutamate transporter-1, glutamate–aspartate transporter, intermediate filaments, me- tabotropic glutamate 5 receptor, white matter astrocytes. J. Neurochem. (2009) 108, 1442–1452. JOURNAL OF NEUROCHEMISTRY | 2009 | 108 | 1442–1452 doi: 10.1111/j.1471-4159.2009.05889.x 1442 Journal Compilation ȑ 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452 ȑ 2009 The Authors

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

*Laboratoire de Pharmacologie Experimentale, Universite catholique de Louvain, Brussels, Belgium
�Neuroanatomy, Department of Neuroscience, Biomedical Center, Uppsala, Sweden
Throughout the brain and the spinal cord, the grey matter ischaracterized by a high density of neuronal cells connectedby dense networks of dendrites and short non-myelinatedaxons. White matter is mainly composed of longer myelin-ated axons that ensure communication between clusters ofgrey matter. Mainly recognized as protoplasmic astroglialcells (designed as type-1 astrocytes), grey matter astrocytesare thought to actively cooperate with neurons in synapticnetworking. In contrast, white matter astrocytes, commonlyadopting a fibrous morphology (also called type-2 astro-cytes), have been assigned a supportive role for thepropagation of electrical impulses through long distanceprojecting axons (Miller and Raff 1984; Hertz and Zielke2004). As white matter is largely devoid of typical post-synaptic targets, the local release of excitatory or inhibitorytransmitters has received limited consideration. However,recent neurochemical studies focusing on the corpus callo-sum, a white matter bundle supporting inter-hemispheric
communication, have revealed a substantial vesicular releaseof glutamate from unmyelinated axons that efficientlyactivates glutamatergic receptors detected in glial progenitors(Kukley et al. 2007; Ziskin et al. 2007). Possibly contributing
Received July 30, 2008; revised manuscript received December 16,2008; accepted December 22, 2008.Address correspondence and reprint requests to Emmanuel Hermans,
Laboratoire de Pharmacologie Experimentale, Universite catholique deLouvain, Brussels, Belgium. E-mail: [email protected] used: CHO, Chinese hamster ovary; CPCCOEt,
7-hydroxyiminocyclopropan[b]chromen-1a-carboxylic acid ethyl ester;CyTM3, cyanin 3; DAPI, 4¢,6-diamidino-2-phenylindole dilactate; DHK,dihydrokainic acid; DHPG, (S)-3,5-dihydroxyphenylglycine; FBS, foetalbovine serum; GFAP, glial fibrillary acidic protein; GLAST, glutamate–aspartate transporter; GLT, glutamate transporters; GS, glutaminesynthetase; IPTG, isopropyl-b-D-thiogalactoside; L-SOS, L-serineO-sulphate potassium salt; LTHA, L-())-threo-3-hydroxyaspartic acid;mGlu5 receptor, type 5 metabotropic glutamate receptor; MPEP,2-methyl-6-(phenylethynyl)-pyridine.
Abstract
While the astrocytic control of extracellular glutamate con-
centration at synaptic contacts is well characterized, little is
known regarding the clearance of glutamate along axon tracts,
even though local excitotoxic damage has been reported.
Therefore, we have compared glutamate handling in astrocyte
cultures derived from white matter (corpus callosum) and grey
matter tissues (cortical structures). These populations of as-
trocytes showed clearly distinct phenotypes, adopting stellate
or protoplasmic morphologies respectively. In addition, white
matter astrocytes showed high densities of the intermediate
filament proteins glial fibrillary acidic protein, vimentin and
nestin. The glutamate–aspartate transporter and glutamate
transporter-1, as well as glutamine synthetase, were found to
be expressed at higher levels in white matter compared with
grey matter astrocytes. Consistent with this aspartate uptake
capacity was three to fourfold higher in white matter cells, and
the use of specific inhibitors revealed a substantial activity of
glutamate transporter-1, contrasting with grey matter cells
where this transporter appeared poorly functional. In addition,
expression of type 5 metabotropic glutamate receptors was
considerably higher in white matter astrocytes where the
agonist (S)-3,5-dihydroxyphenylglycine triggered a large re-
lease of intracellular calcium. Differences in these astrocyte
cultures were also observed when exposed to experimental
conditions that trigger glial activation. This study highlights
typical features of cultured astrocytes derived from white
matter tissues, which appear constitutively adapted to handle
excitotoxic insults. Moreover, the expression and activity of
the astroglial components involved in the control of glutama-
tergic transmission are reinforced when these cells are
maintained under conditions mimicking a gliotic environment.
Keywords: astroglial activation, glutamate transporter-1,
glutamate–aspartate transporter, intermediate filaments, me-
tabotropic glutamate 5 receptor, white matter astrocytes.
J. Neurochem. (2009) 108, 1442–1452.
JOURNAL OF NEUROCHEMISTRY | 2009 | 108 | 1442–1452 doi: 10.1111/j.1471-4159.2009.05889.x
1442 Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452� 2009 The Authors

to some excessive activation of glutamate receptors present inglial cells and neurons, this vesicular release along axonaltracts is thought to participate in the development ofexcitotoxic damage associated with white matter pathologies(for review, Matute et al. 2007). Under pathological condi-tions, the cellular adaptation of white matter glial cellsremains incompletely understood, and it is generally expectedthat type-2 astrocytes may be more vulnerable and degeneratemore rapidly than type-1 astrocytes (Miller et al. 1986). Thiscontrasts with surviving astrocytes of the grey matter whichcan exhibit considerable physiological changes when exposedto excitotoxic processes. These cells also show enhancedproliferative and migratory capacities, and form a glial scarthat impairs neuritic outgrowth from lesioned neurons (Chenand Swanson 2003). Nevertheless, it has also been reportedthat the activation of white matter astrocytes occurs earlier andmore intensively compared with grey matter astrocytes inexcitotoxic models of acute spinal cord injury (Gomes-Lealet al. 2004).
The concept of reactive gliosis and its relationship toneuroprotection against glutamate toxicity has recentlyreceived growing attention. In astrocytes, this adaptiveresponse is characterized by a strong up-regulation ofglutamate–aspartate transporter (GLAST) and glutamatetransporter-1 (GLT-1) expression (Anderson and Swanson2000), as well as an increased expression of glutaminesynthetase (GS), which is critical for glial metabolism ofglutamate (Chang et al. 2007). The glutamate uptake capacityof astrocytes is also indirectly influenced by the intracellularnetwork of intermediate filaments, as increased expression ofthe glial fibrillary acidic protein (GFAP) contributes to themembrane anchoring of GLAST in reactive astrocytes after ahypoxic insult (Sullivan et al. 2007). Indeed, reactive astro-cytes increase the thickness of their cell processes (Wil-helmsson et al. 2006), and are packed with intracellular glialfilaments such as GFAP, vimentin (Eng et al. 2000; Peknyand Nilsson 2005), and nestin (Eliasson et al. 1999; Tamagnoand Schiffer 2006; Korzhevskii et al. 2008). Finally, anotherkey protein that might participate in the control of glutamatetransmission by astrocytes is the type 5 metabotropicglutamate (mGlu5) receptor of which expression is increasedin reactive astrocytes (Aronica et al. 2000, 2001; Ferragutiet al. 2001). Also convincing is the observation that acute orsustained activation of mGlu5 receptors can specificallymodulate GLT-1 expression and activity in astrocytes(Aronica et al. 2003; Vermeiren et al. 2006).
Together, these data indicate that the regulation of GLTsand/or mGlu receptors is critical for the modulation of glialfunction during excitotoxic insults and raise questionsregarding the reactivity of astrocytes present in white matter.Therefore, we have compared the expression and activity ofGLT, GS and the mGlu5 receptor in primary culturesof astrocytes derived from corpus callosum or cortical tissuesof newborn rats. To mimic a gliotic process, astrocytes were
cultured in the presence of a combination of nutrients andgrowth factors promoting their in vitro activation (Yarowskyand Krueger 1989; Vermeiren et al. 2005a). In addition torevealing distinct structural properties, activated white andgrey matter astrocytes were found to exhibit markeddifferences in the astrocytic proteins that regulate glutamatetransmission in the CNS.
Materials and methods
MaterialsPercollTM/RediGradTM medium was obtained from GE Healthcare
(Uppsala, Sweden). CELLSTAR� standard flasks and dishes used
for cultured astrocytes were provided by Greiner Bio-one
(Wemmel, Belgium) and round glass coverslips with 12 or
15 mm2 diameters were from VWR International (Leuven,
Belgium). Poly-L-lysine, 4¢,6-diamidino-2-phenylindole dilactate
(DAPI), aspartate and L-serine O-sulphate potassium salt (L-SOS)
were provided by Sigma-Aldrich (Bornem, Belgium). Dihydrokai-
nic acid (DHK), L-())-threo-3-hydroxyaspartic acid (LTHA),
isopropyl-b-D-thiogalactoside (IPTG) (S)-3,5-dihydroxyphenylgly-cine (DHPG), 2-methyl-6-(phenylethynyl)-pyridine (MPEP) and 7-
hydroxyiminocyclopropan[b]chromen-1a-carboxylic acid ethyl es-
ter (CPCCOEt) were from Tocris (Bristol, UK). PerkinElmer NEN
(Zaventem, Belgium) provided D-[3H]-aspartate (specific activity of
23.9 Ci/mmol). Fura-2AM was supplied by Molecular Probes
(Leiden, Netherlands). The providers of primary and secondary
antibodies were Chemicon International Inc. (Hampshire, UK) for
the guinea pig anti-GLAST and anti-GLT-1 polyclonal and for the
mouse anti-nestin monoclonal antibodies, Dako Diagnostics
(Heverlee, Belgium) for the rabbit anti-GFAP polyclonal and the
mouse anti-vimentin monoclonal antibodies, Jackson Immunore-
search Laboratory (DePinte, Belgium) for the peroxidase-conju-
gated goat anti-guinea-pig IgG, the cyanin 3 (CyTM3)-conjugated
goat anti-rabbit IgG and the FITC-conjugated goat anti-mouse IgG
or anti-rabbit IgG, Sigma-Aldrich for the rabbit anti-actin and anti-
GS antibodies, the peroxidase-conjugated goat anti-rabbit and anti-
mouse IgG and Upstate (Lake Placid, NY, USA) for the rabbit
anti-mGlu5 receptor polyclonal antibody.
Culture of grey and white matter rat astrocytesThe animal procedures were conducted in strict adherence to the
European Community Council directive of 24 November 1986 (86-
609/EEC) and Decree of 20 October 1987 (87-848/EEC). Animals
were kept at constant temperature (23 ± 1�C) and relative humidity
(40–60%) on a 12 h light/dark cycle with access ad libitum to both
food and water. All procedures were performed as described
previously in details (Kozlova and Takenaga 2005) with some
modifications. Thus, primary cultures of astrocytes were prepared
from at least 10 postnatal (5-day old) Wistar rats. The middle part of
the corpus callosum and cortex areas were carefully isolated and
white and grey matter tissues were quickly placed separately in
30 cm2 Petri dish with Dulbecco’s modified Eagle’s medium
containing 4.5 g/L glucose, 25 mM HEPES and supplemented with
10% foetal bovine serum (FBS), 100 U/mL penicillin, 100 lg/mL
streptomycin and 6 mM glutamine (Invitrogen, Merelbeke Belgium).
Astroglial cells were carefully separated from oligodendrocytes or
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452
Glutamate handling in white matter astrocytes | 1443

microglia using discontinuous and cold PercollTM/RediGradTM
gradients (successively composed of 60% and 30% of PercollTM/
RediGradTM). After centrifugation at 2000 g for 10 min at 4�C, thegrey or white matter astrocytes enriched fractions were taken off the
gradients, generously washed and distributed in culture medium
containing 20% FBS at a density of 4 · 104 cells/cm2 (grey matter)
and 8 · 104 cells/cm2 (white matter) in 25 cm2 flasks. Commonly,
106 cortical cells and 0.5 · 106 callosal cells were collected from
one animal. The cultures were maintained for 2 weeks, decreasing
the concentration of FBS to 10% after the first week and then cells
were subcultured for next experiments. Cells were collected after
trypsinization (mortality never exceeding 5%) and distributed at
various densities either on poly-L-lysine coated surfaces for
immunocytochemical analysis, evaluation of D-[3H]-aspartate
uptake and measures of intracellular free calcium or into non-
coated 58 cm2 Petri dishes for immunoblotting experiments. After
adherence (48 h), the culture medium was renewed and for some
experiments, the G5 supplement (insulin, transferrin, selenite, biotin,
hydrocortisone, fibroblast growth factor and epidermal growth
factor; Invitrogen) was added during 7 days to trigger astrocyte
activation as previously reported (Vermeiren et al. 2005a). As the
G5 supplement also increases the cell proliferation, astrocytes were
plated at twofold lower densities for experiments performed in
presence of this additive.
Culture of the CHO-lac-mGlu5a cellsChinese hamster ovary (CHO) cells stably expressing the human
mGlu5a receptor under the control of the inducible Lac Switch-II
system (Stratagene, La Jolla, CA, USA), denoted CHO-lac-mGlu5a,
were maintained as described previously (Atkinson et al. 2006). Formaximal induction of mGlu5a receptor expression in CHO cells, the
medium was replaced with fresh culture medium containing
100 lM IPTG 18 h before experiments.
Analysis of astrocyte morphology and immunocytochemicaldetection of GFAP, nestin and actinAstrocytes from grey or white matter were seeded at, respectively,
2.5 and 5 · 103 cells/cm2 either in non-coated six-well plates for
morphological analysis or on coated 12 mm2 coverslips for immu-
nocytochemical detection. Cells were fixed (ethanol 95%) and
incubated with either primary rabbit anti-GFAP antibody (1 : 1500)
or successively rabbit anti-actin and mouse anti-nestin (1 : 1000) and
afterwards with either anti-rabbit FITC-conjugated IgG (1 : 500) or
successively anti-rabbit CyTM3-conjugated IgG (1 : 250) and anti-
mouse FITC-conjugated IgG (1 : 500). Nuclei were stained with the
nuclear dye DAPI (1/5000). Cells morphology observed by phase
contrast microscopy and immunolabellings were examined using an
Olympus IX70 inverted microscope coupled to a charged coupled
device camera (T.I.L.L. Photonics, Martinsried, Germany). Excita-
tion light (400, 488 and 540 nm for DAPI, FITC and CyTM3
respectively) was obtained from a xenon lamp coupled to a
monochromator (T.I.L.L. Photonics). Digital images were acquired
using appropriate filters and combined using the TILLVISION
software version 3.3 (T.I.L.L. Photonics).
Western blottingGrey and white matter astrocytes plated at, respectively, 5 and
10 · 103 cells/cm2 on non-coated 58 cm2 Petri dishes were rinsed
and scrapped in phosphate-buffered saline (137 mM NaCl, 21 mM
NaHPO4, 29 mM KH2PO4 and 1.2 mM KCl, pH 7.4). After
centrifugation at 8000 g for 10 min, total proteins extracts in the cell
pellet were solubilized in the solubilization buffer (10 mM Tris–
HCl, pH 7.4, 20 mM CHAPS, 0.5 mM EDTA, 30 mM dithiothre-
itol, 0.5 mM phenylmethylsulphonyl fluoride and 1 lM protease
inhibitor cocktail). After measuring the protein concentration, each
sample (25 lg for vimentin, 40 lg for GS, 50 lg for GFAP, nestin,
GLAST and mGlu5 receptor and 75 lg for GLT-1) was diluted into
the loading buffer (125 mM Tris–HCl, pH 7.4, 50 mM dithiothre-
itol, 4% sodium dodecyl sulphate, 20% glycerol and 0.01%
bromophenol blue, pH 6.8) and stored at )20�C. After thawing,
the samples were boiled for 5 min and electrophoresed through a
10% (7.5% for nestin and mGluR5 detection) sodium dodecyl
sulphate–polyacrylamide gel according to Laemmli and transferred
to nitrocellulose membranes with a Bio-Rad minitransblot electro-
phoretic transfer cell (Bio-Rad, Hercules, CA, USA). To avoid non-
specific immunodetection, membranes were incubated for 30 min in
Tris-buffered saline (50 mM Tris–HCl and 150 mM NaCl, pH 8.1)
containing 0.05% Tween 20 and 5% non-fat milk. Immunoprobing
was then performed overnight at 4�C using primary antibodies
recognizing GFAP (1 : 1500), nestin (1 : 1000), vimentin
(1 : 2000), GLAST and GLT-1 (1 : 2500), GS (1 : 500), mGlu5
receptor (1 : 1200) and actin (1 : 750) and subsequently for 1 h at
22�C using peroxidase-conjugated secondary goat anti-rabbit
(1 : 3000), anti-mouse (1 : 2000) and anti-guinea-pig IgG
(1 : 5000). The immunoreactive proteins were detected with
enhanced chemiluminescence reagents and densitometric analysis
of the signal was performed using the Multimodal Imaging system
of Kodak Image Station 2000 MM (Eastman Kodak Company, New
Haven, CT, USA).
Measurement of aspartate transport activityUptake assays were performed as previously described (Goursaud
et al. 2008) with some modifications. Astrocytes were seeded on
coated 12-well plates at 5 · 103 cells/cm2 (for grey matter) or
10 · 103 cells/cm2 (for white matter) and grown for 7 days. When
indicated, inhibitors of GLT were added 6 min before the addition
of the substrate D-[3H]-aspartate (50 nM), to final concentrations of
100 lM for LTHA (Ki < 50 lM) and DHK (Ki = 23 ± 6 lM) or
200 lM for L-SOS (Ki for GLAST = 107 ± 8 lM) in accordance
with previous investigations (Arriza et al. 1994). For saturation
assays, D-[3H]-aspartate (50 nM) was diluted with unlabelled L-
aspartate to achieve final concentrations of substrate ranging from
10 to 300 lM. The uptake (stopped after 6 min), was linear as
previously observed (Kimmich et al. 2001) and the concentration
of substrate in the medium was not significantly modified during
this incubation time. The specific activity of the GLT (expressed as
the uptake velocity per mg of protein) was estimated after
subtracting the data obtained using the non-selective GLT inhibitor
LTHA.
Measurement of intracellular calcium concentration [Ca2+]i insingle cellsChanges in [Ca2+]i were measured in single cells, seeded at low
densities (2.5 and 5 · 103 cells/cm2 for, respectively, grey and
white matter astrocytes) on coated 15 mm2 coverslips. Cells were
loaded with the intracellular calcium indicator Fura-2AM (2 lM
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452� 2009 The Authors
1444 | S. Goursaud et al.

for 45 min at 20�C) and the calcium measurements were
performed as previously detailed (Vermeiren et al. 2005b). When
specified, astrocytes were perfused with 50 lM DHPG for 30 s in
presence or not of the antagonists CPCCOEt (50 lM) or MPEP
(0.5 lM) applied 30 s before the agonist. Loaded cells were
excited for 50 ms alternatively at 340 and 380 nm (1 Hz excitation
lights) and emitted fluorescence was monitored at 510 nm using a
charged coupled device camera coupled to an inverted Olympus
IX70 microscope (T.I.L.L. Photonics). Fluorescence intensities
from single cell excited at the two wavelengths were recorded
separately and corrected for the background using the software
TILLVISION v3.3.
Statistical analysisThe uptake kinetic parameters Km (affinity) and Vmax (maximal
velocity) were obtained by non-linear analysis of experimental data
using GRAPHPAD Prism version 3.02 (GRAPHPAD Software, San
Diego, CA, USA). Data are expressed as mean ± SEM and
significant differences between control and samples treated with
various drugs determined by one-way ANOVA followed by the
Bonferroni’s test for multiple comparisons using GRAPHPAD Prism
version 3.02. Values of p < 0.05 were considered as statistically
significant (*,#p < 0.05; **,##p < 0.01; ***,###p < 0.001). Symbols
*, ** and *** were used for analyses between naive astrocytes from
grey and white matter, whereas #, ## and ### indicated significant
differences between naive and G5-treated astrocytes either from
grey or white matter.
Results
Cell morphology and expression of intermediate filamentsin white and grey matter astrocytesConventional protocols for cortical astrocyte cultures gener-ally give rise to dense proliferating cultures where cells fromthe grey matter are largely predominant, restricting thebiochemical studies of astrocytes of white matter origin. Tospecifically examine the properties of astrocytes from bothgrey and white matter, an optimized procedure was followedto derive enriched cultures from cortical layers of forebrainhemispheres, or from the corpus callosum respectively(Kozlova and Takenaga 2005). Immunocytochemical analy-sis revealed that the majority of cells (at least 95%) obtainedfrom newborn rat corpus callosum or cortical structures wereGFAP-positive, confirming the purity of the astrocytecultures (Fig. 1a and b). Daily microscopic observation ofthe culture monolayers and evaluation of protein content atdifferent stages suggested that callosal astrocytes show lower
(a) (c)
(b) (d)
Fig. 1 Morphology and detection of GFAP and nestin expression in
grey and white matter astrocytes. Grey matter (G) and white matter
(W) astrocytes were, respectively, isolated from cortical and callosal
tissues from 5-day-old rats. After a 2 weeks proliferation step, cultures
were maintained in the absence (a and c) or in the presence (b and d)
of a growth factor supplement for 7 days. Morphology was examined
by phase contrast microscopy (a and b, left panels) while GFAP (a and
b, right panels), actin and nestin (c and d, respectively, left and middle
panels) were visualized after immunocytochemical staining with spe-
cific antibodies. GFAP and nestin appear in green (FITC) whereas
actin and cell nuclei appear, respectively, in red (CyTM3) and blue
(DNA staining with DAPI). Right panels of (c and d) show merged
images combining nestin and actin detection (scale bar, 20 lm).
These panels were typically representative of three independent
immunocytochemical analyses.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452
Glutamate handling in white matter astrocytes | 1445

proliferation rates compared with cortical astrocytes (data notshown). This observation is consistent with an earlier reportconcerning newborn rat optic nerve astrocyte cultures (Raffet al. 1983). Consequently, to facilitate the comparison ofbiochemical data, the two cell populations were plated atdifferent densities in all experiments.
Phase contrast microscopy provided evidence of consi-derable morphological differences between grey and whitematter astrocytes. After being maintained under identicalstandard culture conditions, grey matter astrocytes had largecytoplasmic regions with laminar processes and showedmoderate GFAP immunoreactivity (typically protoplasmic ortype-1 astrocytes) (Fig. 1a). In contrast, white matter astro-cytes exhibited a reduced cytoplasmic volume with smallernuclei and longer and branched processes showing intenseGFAP immunostaining (commonly referred to as fibrous ortype-2 astrocytes) (Fig. 1a).
The enrichment of the culture medium with a number ofgrowth factors and nutrients (G5) has previously been shownto switch astrocyte cultures toward an activated phenotype.Exposing the cells for 7 consecutive days to this supplementinduced important morphological changes in both corticaland callosal astroglial cultures and tended to enhance GFAPimmunostaining in grey matter astrocytes (Fig. 1b). Indeed,the latter showed a reduction in cytoplasmic volume anddeveloped fibrous membrane extensions. In addition, acti-vated white matter astrocytes exhibited increased length andbranching of fibrous processes, establishing a dense, con-necting network between cells. Immunocytochemical detec-tion of the other intermediate filament protein vimentin didnot reveal noticeable differences between cortical and callosalastrocytes and no change was observed after exposure to thegrowth factor supplement (data not shown). In contrast, cleardifferences between the two cell cultures were evident whenthe intensity and pattern of nestin-immunoreactivity wasanalysed. In grey matter astrocytes, nestin staining wasweakly detected close to nuclei of cells maintained undercontrol conditions (Fig. 1c), but was much more pronouncedafter exposure to the G5 supplement, mainly in the cytoplasmand to a lesser extent in some membrane extensions (Fig. 1d).In contrast, the entire population of white matter astrocytesexposed or not to the G5 supplement stained positively fornestin, exhibiting a homogenous and intense co-localizationwith actin immunoreactivity throughout the cytoplasm and incell extensions (Fig. 1c and d).
Consistently, immunoblotting studies revealed that understandard culture conditions the intermediate filaments wereexpressed at higher density in astrocytes isolated from whitematter (Fig. 2a). In particular, expression of nestin andvimentin appeared, respectively, 4.8- and 2.7-fold higher(p < 0.001) and GFAP detection was 1.6-fold (p < 0.05)increased in astrocytes from corpus callosum compared withthose from cortical structures (Fig. 2b). The addition of thegrowth factor supplement increased (by twofold) the expres-
sion of these three proteins in greymatter astrocytes, but had nosignificant effect in activatedwhitematter astrocytes (Fig. 2b).
Expression and functional properties of glutamatetransporters in white and grey matter astrocytesQuantitative differences in the expression of the main GLTGLAST and GLT-1 in the cortical and callosal astroglialcultures were first assessed in immunoblotting studies(Fig. 3a). Thus, when grown under standard conditions,both GLT were more abundant in white matter astrocytes(threefold higher when compared with grey matter astro-cytes, p < 0.001) (Fig. 3b). In addition, a higher expression
(a)
(b)
Fig. 2 Immunoblotting analysis of GFAP, vimentin and nestin
expression in grey and white matter astrocytes. Panel (a) illustrates
immunodetection of the three intermediate filaments and actin in
protein extracts from grey matter (G) or white matter (W) astrocytes.
Protein samples from cell grown in the absence (control) or in the
presence of a growth factors supplement (GF) were separated by
sodium dodecyl sulphate–polyacrylamide gel electrophoresis and
probed with anti-GFAP, anti-vimentin, anti-nestin and anti-actin anti-
bodies. Data shown are from a typical experiment performed four
times independently. Panel (b) shows quantitative data from densito-
metric analysis of GFAP, vimentin or nestin immunoreactive signals
normalized to actin, obtained in four independent experiments (mean
with SEM; mean signals from naive astrocytes extract were arbitrarily
set to the unit) (*,#p < 0.05; ##p < 0.01; ***,###p < 0.001).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452� 2009 The Authors
1446 | S. Goursaud et al.

of the key glial glutamate-metabolizing enzyme GS was alsoobserved in white matter cells (1.8-fold higher, p < 0.001).The exposure to the growth factor supplement significantlyincreased the expression of these proteins in grey matterastrocytes (2.0-fold for GLAST, p < 0.05; 2.3-fold for GLT-1, p < 0.05; and 2.8-fold for GS, p < 0.001). In white matterastrocytes, the expression of GLT-1 and GS was strongly up-regulated in the presence of the G5 supplement (2.5- and 4.4-fold increases, respectively, p < 0.001) while expression ofGLAST was not significantly altered.
The involvement of GLAST and GLT-1 was examined byevaluating the specific uptake of D-[3H]-aspartate (Fig. 4) inthe presence of selective pharmacological inhibitors. In theseexperiments, the substrate D-[3H]-aspartate was used at atracer concentration (50 nM) to ensure an efficient compe-tition between the substrate and DHK (selective GLT-1inhibitor) or L-SOS (GLAST inhibitor). Consistent withimmunoblotting studies, the specific uptake appeared 3.4-fold higher in white matter astrocytes when compared withgrey matter astrocytes grown under standard conditions(Fig. 4a). In the former, uptake involved both GLAST andGLT-1, as inhibition was observed with both L-SOS (60%inhibition, p < 0.001) and DHK (40% inhibition p < 0.001).In contrast, the substrate uptake measured in grey matterastrocytes appeared exclusively dependent on GLAST, asessentially 100% blockade was obtained in the presence ofL-SOS, and DHK was without significant effect (Fig. 4a).Exposure to the growth factor supplement resulted in a
doubling of the specific D-[3H]-aspartate uptake capacity inboth grey and white matter astrocytes (Fig. 4b). The uptakevalues obtained in the presence of the inhibitors revealed thatthe increased activity induced by the G5 supplement in greymatter astrocytes was mainly the result of an induction ofGLT-1. In contrast, the increased uptake in the white matterastrocytes was due to an increased activity of GLAST.
The activity of the GLT was first characterized bymeasuring the specific uptake of isotopic dilutions of D-[3H]-aspartate (dilutions with L-aspartate) to derive theuptake kinetic parameters (Fig. 4a and b). Analysis ofsaturation isotherms revealed that when grown in standardculture conditions, white matter astrocytes showed a higherVmax value (40.17 ± 1.79 nmol/min/mg protein) and a lowerKm value (45.91 ± 6.94 lM) when compared with greymatter astrocytes (Vmax and Km values of 31.31 ± 3.72 nmol/min/mg protein and 132.80 ± 35.08 lM respectively). Theexposure to the G5 supplement induced an increase of Vmax
in both models (45.70 ± 4.40 and 67.02 ± 3.02 nmol/min/mg protein for grey and white matter astrocytes respectively).Besides, activation with growth factors caused a decreasedaffinity for aspartate in cultured cortical glial cells (Km valueof 99.20 ± 23.86 lM) whereas no modification was detectedfor cultured callosal astrocytes (Km values of 48.96 ±7.31 lM). Overall, these differences in Vmax values areconsistent with a higher density of GLT in cultures derivedfrom the white matter. In addition, in accordance with ahigher affinity of aspartate for GLT-1 when compared with
(a) (b)
Fig. 3 Immunoblotting analysis of GLAST, GLT-1 and GS expression
in grey and white matter astrocytes. Panel (a) illustrates immunode-
tection of the two glutamate transporters, the GS and actin in protein
extracts from grey matter (GM) or white matter (WM) astrocytes.
Protein samples from cell grown in the absence (control) or in the
presence of a growth factors supplement (GF) were separated by
sodium dodecyl sulphate–polyacrylamide gel electrophoresis and
probed with anti-GLAST, anti-GLT-1, anti-GS and anti-actin antibod-
ies. Data shown are from a typical experiment performed five times
independently. Panel (b) shows quantitative data from densitometric
analysis of GLAST, GLT-1 or GS immunoreactive signals normalized
to actin, obtained in five independent experiments (mean with SEM;
mean signals from naive astrocytes extract were arbitrarily set to the
unit) (#p < 0.05 and ***,###p < 0.001).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452
Glutamate handling in white matter astrocytes | 1447

GLAST, the involvement of GLT-1 demonstrated here abovein white matter astrocytes or in grey matter cells exposed tothe growth factor supplement was correlated with lowerexperimental Km values.
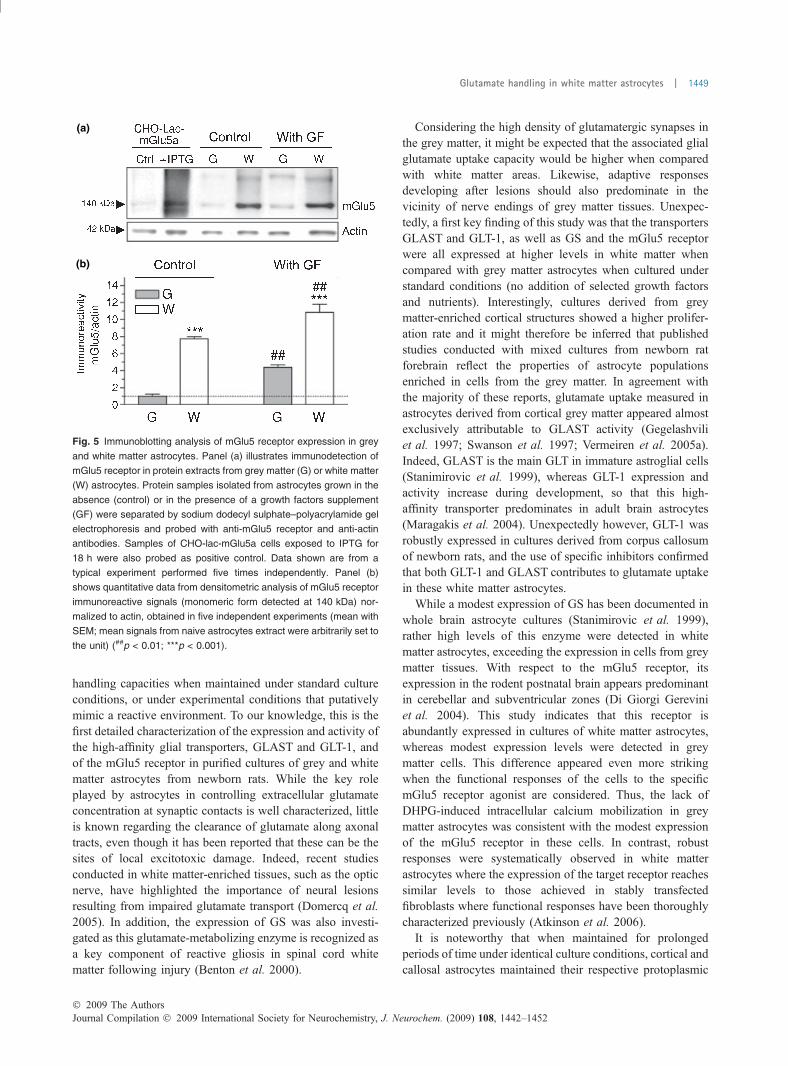
Differential regulation of mGlu5 receptor by growthfactors in grey and white matter astrocytesPreliminary studies revealed that among the group I mGlureceptors only the mGlu5a receptor subtype was expressedunder physiological conditions in astroglial cells (Ferragutiet al. 2001; Vermeiren et al. 2005b). In the present report,quantitative immunoblotting analysis of astrocytes culturedin the absence of growth factors indicated that the astroglialmGlu5 receptor is detectable in cultures from cortex, but ishighly expressed in cells from the corpus callosum (7.7-folddifference, p < 0.001). In these white matter astrocytes, theimmunoreactive signal was similar to that observed intransfected fibroblasts (CHO cells) expressing the recombi-nant mGlu5a receptor (IPTG-inducible expression), usedhere as a positive control (Fig. 5a and b). In the presence ofgrowth factors, consistent with previous studies conducted inconventional cultures of astrocytes (Vermeiren et al. 2005b),a robust increase (4.4-fold) in the expression of the mGlu5receptor was observed in cultures of grey matter astrocytes(p < 0.01). Such treatment resulted in only a modest increase(1.4-fold) in the expression of this receptor in white matterastrocytes (Fig. 5a and b).
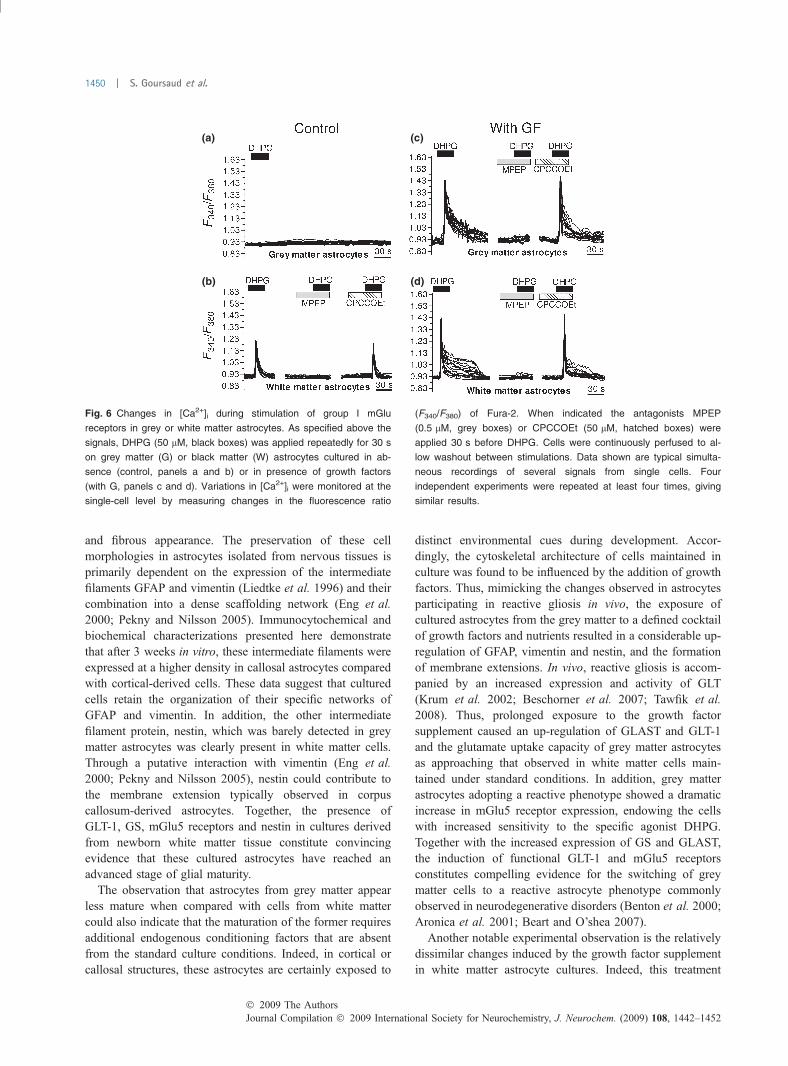
These differences in the expression levels of mGlu5receptors were correlated with distinct functional responses.Single cell imaging and analysis of fluorescence ratios inFura-2 loaded cells allowed us to determine the proportion ofastrocytes responding to brief stimulations with the group ImGlu receptor agonist DHPG. In cells grown under standardconditions, DHPG (50 lM) caused an immediate, transientincrease in [Ca2+]i in the majority of white matter astrocytes(Fig. 6b), whereas no responding cells were detected in thecultures isolated from grey matter tissues (Fig. 6a). Incontrast, a robust response was systematically observed inthese grey matter astrocytes when grown in the presence ofthe growth factor supplement (Fig. 6c). For white matterastrocytes, exposure to this treatment resulted in a relativeincrease in the amplitude of [Ca2+]i responses. In addition, alarge proportion (�60%) of white matter astrocytes showed apeak and plateau response on exposure to the agonist(Fig. 6d), likely reflecting increased expression of the targetreceptor. In both astrocyte populations, the responses toDHPG were totally inhibited in the presence of the mGlu5receptor antagonist MPEP, but not by CPCCOEt, a selectiveantagonist of mGlu1 receptors.
Discussion
The aim of this study was to address the question of whethercallosal and cortical astrocytes exhibit distinct glutamate
(a) (c)
(b) (d)
Fig. 4 Characterization of aspartate uptake in grey and white matter
astrocytes. The velocity of [3H]-aspartate uptake was measured in
naive cells (control, panels a and c) or in cells exposed for 7 days to a
supplement of growth factors (GF; panels b and d). Saturation of
aspartate uptake velocity (panels a and b) was obtained with dilutions
of D-[3H]-aspartate (50 nM) with unlabelled L-aspartate to achieve final
concentrations of substrate ranging from 10 to 300 lM. When indi-
cated (panels c and d), the GLT-1- and GLAST-dependent D-[3H]-
aspartate (50 nM) uptake were evaluated in the presence of the
specific pharmacological inhibitors DHK (100 lM) or L-SOS (200 lM)
respectively. Data shown are the mean with SEM from, respectively,
three or four independent experiments performed in quadruplicate
(#p < 0.05; ***,###p < 0.001).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452� 2009 The Authors
1448 | S. Goursaud et al.
handling capacities when maintained under standard cultureconditions, or under experimental conditions that putativelymimic a reactive environment. To our knowledge, this is thefirst detailed characterization of the expression and activity ofthe high-affinity glial transporters, GLAST and GLT-1, andof the mGlu5 receptor in purified cultures of grey and whitematter astrocytes from newborn rats. While the key roleplayed by astrocytes in controlling extracellular glutamateconcentration at synaptic contacts is well characterized, littleis known regarding the clearance of glutamate along axonaltracts, even though it has been reported that these can be thesites of local excitotoxic damage. Indeed, recent studiesconducted in white matter-enriched tissues, such as the opticnerve, have highlighted the importance of neural lesionsresulting from impaired glutamate transport (Domercq et al.2005). In addition, the expression of GS was also investi-gated as this glutamate-metabolizing enzyme is recognized asa key component of reactive gliosis in spinal cord whitematter following injury (Benton et al. 2000).
Considering the high density of glutamatergic synapses inthe grey matter, it might be expected that the associated glialglutamate uptake capacity would be higher when comparedwith white matter areas. Likewise, adaptive responsesdeveloping after lesions should also predominate in thevicinity of nerve endings of grey matter tissues. Unexpec-tedly, a first key finding of this study was that the transportersGLAST and GLT-1, as well as GS and the mGlu5 receptorwere all expressed at higher levels in white matter whencompared with grey matter astrocytes when cultured understandard conditions (no addition of selected growth factorsand nutrients). Interestingly, cultures derived from greymatter-enriched cortical structures showed a higher prolifer-ation rate and it might therefore be inferred that publishedstudies conducted with mixed cultures from newborn ratforebrain reflect the properties of astrocyte populationsenriched in cells from the grey matter. In agreement withthe majority of these reports, glutamate uptake measured inastrocytes derived from cortical grey matter appeared almostexclusively attributable to GLAST activity (Gegelashviliet al. 1997; Swanson et al. 1997; Vermeiren et al. 2005a).Indeed, GLAST is the main GLT in immature astroglial cells(Stanimirovic et al. 1999), whereas GLT-1 expression andactivity increase during development, so that this high-affinity transporter predominates in adult brain astrocytes(Maragakis et al. 2004). Unexpectedly however, GLT-1 wasrobustly expressed in cultures derived from corpus callosumof newborn rats, and the use of specific inhibitors confirmedthat both GLT-1 and GLAST contributes to glutamate uptakein these white matter astrocytes.
While a modest expression of GS has been documented inwhole brain astrocyte cultures (Stanimirovic et al. 1999),rather high levels of this enzyme were detected in whitematter astrocytes, exceeding the expression in cells from greymatter tissues. With respect to the mGlu5 receptor, itsexpression in the rodent postnatal brain appears predominantin cerebellar and subventricular zones (Di Giorgi Gereviniet al. 2004). This study indicates that this receptor isabundantly expressed in cultures of white matter astrocytes,whereas modest expression levels were detected in greymatter cells. This difference appeared even more strikingwhen the functional responses of the cells to the specificmGlu5 receptor agonist are considered. Thus, the lack ofDHPG-induced intracellular calcium mobilization in greymatter astrocytes was consistent with the modest expressionof the mGlu5 receptor in these cells. In contrast, robustresponses were systematically observed in white matterastrocytes where the expression of the target receptor reachessimilar levels to those achieved in stably transfectedfibroblasts where functional responses have been thoroughlycharacterized previously (Atkinson et al. 2006).
It is noteworthy that when maintained for prolongedperiods of time under identical culture conditions, cortical andcallosal astrocytes maintained their respective protoplasmic
(a)
(b)
Fig. 5 Immunoblotting analysis of mGlu5 receptor expression in grey
and white matter astrocytes. Panel (a) illustrates immunodetection of
mGlu5 receptor in protein extracts from grey matter (G) or white matter
(W) astrocytes. Protein samples isolated from astrocytes grown in the
absence (control) or in the presence of a growth factors supplement
(GF) were separated by sodium dodecyl sulphate–polyacrylamide gel
electrophoresis and probed with anti-mGlu5 receptor and anti-actin
antibodies. Samples of CHO-lac-mGlu5a cells exposed to IPTG for
18 h were also probed as positive control. Data shown are from a
typical experiment performed five times independently. Panel (b)
shows quantitative data from densitometric analysis of mGlu5 receptor
immunoreactive signals (monomeric form detected at 140 kDa) nor-
malized to actin, obtained in five independent experiments (mean with
SEM; mean signals from naive astrocytes extract were arbitrarily set to
the unit) (##p < 0.01; ***p < 0.001).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452
Glutamate handling in white matter astrocytes | 1449
and fibrous appearance. The preservation of these cellmorphologies in astrocytes isolated from nervous tissues isprimarily dependent on the expression of the intermediatefilaments GFAP and vimentin (Liedtke et al. 1996) and theircombination into a dense scaffolding network (Eng et al.2000; Pekny and Nilsson 2005). Immunocytochemical andbiochemical characterizations presented here demonstratethat after 3 weeks in vitro, these intermediate filaments wereexpressed at a higher density in callosal astrocytes comparedwith cortical-derived cells. These data suggest that culturedcells retain the organization of their specific networks ofGFAP and vimentin. In addition, the other intermediatefilament protein, nestin, which was barely detected in greymatter astrocytes was clearly present in white matter cells.Through a putative interaction with vimentin (Eng et al.2000; Pekny and Nilsson 2005), nestin could contribute tothe membrane extension typically observed in corpuscallosum-derived astrocytes. Together, the presence ofGLT-1, GS, mGlu5 receptors and nestin in cultures derivedfrom newborn white matter tissue constitute convincingevidence that these cultured astrocytes have reached anadvanced stage of glial maturity.
The observation that astrocytes from grey matter appearless mature when compared with cells from white mattercould also indicate that the maturation of the former requiresadditional endogenous conditioning factors that are absentfrom the standard culture conditions. Indeed, in cortical orcallosal structures, these astrocytes are certainly exposed to
distinct environmental cues during development. Accor-dingly, the cytoskeletal architecture of cells maintained inculture was found to be influenced by the addition of growthfactors. Thus, mimicking the changes observed in astrocytesparticipating in reactive gliosis in vivo, the exposure ofcultured astrocytes from the grey matter to a defined cocktailof growth factors and nutrients resulted in a considerable up-regulation of GFAP, vimentin and nestin, and the formationof membrane extensions. In vivo, reactive gliosis is accom-panied by an increased expression and activity of GLT(Krum et al. 2002; Beschorner et al. 2007; Tawfik et al.2008). Thus, prolonged exposure to the growth factorsupplement caused an up-regulation of GLAST and GLT-1and the glutamate uptake capacity of grey matter astrocytesas approaching that observed in white matter cells main-tained under standard conditions. In addition, grey matterastrocytes adopting a reactive phenotype showed a dramaticincrease in mGlu5 receptor expression, endowing the cellswith increased sensitivity to the specific agonist DHPG.Together with the increased expression of GS and GLAST,the induction of functional GLT-1 and mGlu5 receptorsconstitutes compelling evidence for the switching of greymatter cells to a reactive astrocyte phenotype commonlyobserved in neurodegenerative disorders (Benton et al. 2000;Aronica et al. 2001; Beart and O’shea 2007).
Another notable experimental observation is the relativelydissimilar changes induced by the growth factor supplementin white matter astrocyte cultures. Indeed, this treatment
(a) (c)
(b) (d)
Fig. 6 Changes in [Ca2+]i during stimulation of group I mGlu
receptors in grey or white matter astrocytes. As specified above the
signals, DHPG (50 lM, black boxes) was applied repeatedly for 30 s
on grey matter (G) or black matter (W) astrocytes cultured in ab-
sence (control, panels a and b) or in presence of growth factors
(with G, panels c and d). Variations in [Ca2+]i were monitored at the
single-cell level by measuring changes in the fluorescence ratio
(F340/F380) of Fura-2. When indicated the antagonists MPEP
(0.5 lM, grey boxes) or CPCCOEt (50 lM, hatched boxes) were
applied 30 s before DHPG. Cells were continuously perfused to al-
low washout between stimulations. Data shown are typical simulta-
neous recordings of several signals from single cells. Four
independent experiments were repeated at least four times, giving
similar results.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452� 2009 The Authors
1450 | S. Goursaud et al.

tended to reinforce their stellate morphology with longer andmore abundant membrane extensions, confirming that bothpopulations of astrocytes can adopt common features ofgliotic reactivity. Nevertheless, the density of intermediatefilaments in white matter cells was not substantially influ-enced by the growth factors, suggesting that morphologicalchanges more likely reflect alteration in their cytoplasmiclocalization and/or local rearrangements (Bushong et al.2004). Intriguingly, the use of pharmacological inhibitorsrevealed that the increased substrate uptake capacity inducedby growth factors in white matter cells was primarilysupported by an increased activity of GLAST. Thus, up-regulation of GLT-1 expression appeared to mainly give riseto non-functional transporter molecules. On the other hand,the increased GLAST-dependent substrate uptake contrastswith the lack of significant change in the expression of thistransporter. Recent studies have reported the importance ofintermediate filaments in the control of GLAST trafficking(Sullivan et al. 2007), supporting the idea that increasedsubstrate uptake in white matter astrocytes may be related tochanges in cell morphology rather than de novo GLASTexpression.
In this study, a number of new observations indicating thatcultured astrocytes initially isolated from grey and whitematter tissues show distinct properties, in particular withrespect to their morphology and their glutamate-handlingcapacity. Even though cultures were derived from differentbrain structures, it is remarkable that neurochemical diffe-rences were preserved despite being maintained underidentical in vitro conditions for more than 20 days. Thissuggests that like neuronal cells, which are frequentlycultured from specific brain regions, glial cells may alsoshown distinct phenotypes that are deeply engraved at earlystages of animal’s development. In addition, our datahighlight the typical features of white matter astrocyteswhich appear constitutively well adapted to face excitotoxicinsults. Indeed, these cells show high expression levels offunctional GLT and glutamate-metabolizing enzymes. Inaddition, these white matter cells express functional mGlu5receptors which have been previously shown to positivelymodulate GLT expression and function (Aronica et al. 2000;Vermeiren et al. 2005b). Even though these properties can befurther reinforced under defined culture conditions mimic-king a gliotic environment, our data indicate that, in compa-rison to grey matter cells, native white matter astrocytesshare common properties with reactive astrocytes. As severalneurodegenerative diseases have been shown to affectneuronal functions along axonal tracts, the study of excito-toxic processes in white matter tissues has recently receivedgrowing attention (Baltan et al. 2008). In addition, gluta-mate-mediated excitotoxic insults to oligodendrocytes in thewhite matter appear to play a critical role in several disordersof the CNS (for review, Matute et al. 2007). This studydemonstrates that glial cells in these tissues show unique
molecular and cellular properties and therefore certainlydeserve further consideration for the development of futureneuroprotective treatments.
Acknowledgements
We thank A. Lebbe, R. Lenaert and T. Timmerman for their
excellent technical assistance. This work was supported by the
National Fund for Scientific Research (F.N.R.S., Belgium, Conven-
tion FRSM 3.4560.07 and credit au chercheur 1.5.109.09.F), by the
Belgian Queen Elisabeth Medical Foundation (Belgium), by the
DIANE research program of Belgium’s Walloon region (DGTRE)
and by the Swedish Research Council (project No. 5420 and 20716).
EH is Research Director of the F.N.R.S.
References
Anderson C. M. and Swanson R. A. (2000) Astrocyte glutamate trans-port: review of properties, regulation, and physiological functions.Glia 32, 1–14.
Aronica E., van Vliet E. A., Mayboroda O. A., Troost D., da Silva F. H.and Gorter J. A. (2000) Upregulation of metabotropic glutamatereceptor subtype mGluR3 and mGluR5 in reactive astrocytes in arat model of mesial temporal lobe epilepsy. Eur. J. Neurosci. 12,2333–2344.
Aronica E., Catania M. V., Geurts J., Yankaya B. and Troost D. (2001)Immunohistochemical localization of group I and II metabotropicglutamate receptors in control and amyotrophic lateral sclerosishuman spinal cord: upregulation in reactive astrocytes. Neurosci-ence 105, 509–520.
Aronica E., Gorter J. A., Ijlst-Keizers H., Rozemuller A. J., Yankaya B.,Leenstra S. and Troost D. (2003) Expression and functional role ofmGluR3 and mGluR5 in human astrocytes and glioma cells:opposite regulation of glutamate transporter proteins. Eur. J.Neurosci. 17, 2106–2118.
Arriza J. L., Fairman W. A., Wadiche J. I., Murdoch G. H., KavanaughM. P. and Amara S. G. (1994) Functional comparisons of threeglutamate transporter subtypes cloned from human motor cortex.J. Neurosci. 14, 5559–5569.
Atkinson P. J., Young K. W., Ennion S. J., Kew J. N., Nahorski S. R. andChalliss R. A. (2006) Altered expression of G(q/11alpha) proteinshapes mGlu1 and mGlu5 receptor-mediated single cell inositol1,4,5-trisphosphate and Ca(2+) signaling. Mol. Pharmacol. 69,174–184.
Baltan S., Besancon E. F., Mbow B., Ye Z., Hamner M. A. and RansomB. R. (2008) White matter vulnerability to ischemic injury in-creases with age because of enhanced excitotoxicity. J. Neurosci.28, 1479–1489.
Beart P. M. and O’shea R. D. (2007) Transporters for L-glutamate: anupdate on their molecular pharmacology and pathologicalinvolvement. Br. J. Pharmacol. 150, 5–17.
Benton R. L., Ross C. D. and Miller K. E. (2000) Glutamine synthetaseactivities in spinal white and gray matter 7 days following spinalcord injury in rats. Neurosci. Lett. 291, 1–4.
Beschorner R., Simon P., Schauer N., Mittelbronn M., Schluesener H. J.,Trautmann K., Dietz K. and Meyermann R. (2007) Reactive as-trocytes and activated microglial cells express EAAT1, but notEAAT2, reflecting a neuroprotective potential following ischaemia.Histopathology 50, 897–910.
Bushong E. A., Martone M. E. and Ellisman M. H. (2004) Maturation ofastrocyte morphology and the establishment of astrocyte domains
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452
Glutamate handling in white matter astrocytes | 1451

during postnatal hippocampal development. Int. J. Dev. Neurosci.22, 73–86.
Chang M. L., Wu C. H., Jiang-Shieh Y. F., Shieh J. Y. and Wen C. Y.(2007) Reactive changes of retinal astrocytes and Muller glial cellsin kainate-induced neuroexcitotoxicity. J. Anat. 210, 54–65.
Chen Y. and Swanson R. A. (2003) Astrocytes and brain injury. J. Cereb.Blood Flow Metab. 23, 137–149.
Di Giorgi Gerevini V. D., Caruso A., Cappuccio I., Ricci V. L., RomeoS., Della R. C., Gradini R., Melchiorri D. and Nicoletti F. (2004)The mGlu5 metabotropic glutamate receptor is expressed in zonesof active neurogenesis of the embryonic and postnatal brain. BrainRes. Dev. Brain Res. 150, 17–22.
Domercq M., Etxebarria E., Perez-Samartin A. and Matute C. (2005)Excitotoxic oligodendrocyte death and axonal damage induced byglutamate transporter inhibition. Glia 52, 36–46.
Eliasson C., Sahlgren C., Berthold C. H., Stakeberg J., Celis J. E.,Betsholtz C., Eriksson J. E. and Pekny M. (1999) Intermediatefilament protein partnership in astrocytes. J. Biol. Chem. 274,23996–24006.
Eng L. F., Ghirnikar R. S. and Lee Y. L. (2000) Glial fibrillary acidicprotein: GFAP-thirty-one years (1969–2000). Neurochem. Res. 25,1439–1451.
Ferraguti F., Corti C., Valerio E., Mion S. and Xuereb J. (2001) Acti-vated astrocytes in areas of kainate-induced neuronal injury upre-gulate the expression of the metabotropic glutamate receptors 2/3and 5. Exp. Brain Res. 137, 1–11.
Gegelashvili G., Danbolt N. C. and Schousboe A. (1997) Neuronalsoluble factors differentially regulate the expression of the GLT1and GLAST glutamate transporters in cultured astroglia. J. Neu-rochem. 69, 2612–2615.
Gomes-Leal W., Corkill D. J., Freire M. A., Picanco-Diniz C. W. andPerry V. H. (2004) Astrocytosis, microglia activation, oligoden-drocyte degeneration, and pyknosis following acute spinal cordinjury. Exp. Neurol. 190, 456–467.
Goursaud S., Maloteaux J. M. and Hermans E. (2008) Activation of VIP/PACAP type 2 receptor by the peptide histidine isoleucine in as-trocytes influences GLAST-mediated glutamate uptake. J. Neuro-chem. 105, 1165–1175.
Hertz L. and Zielke H. R. (2004) Astrocytic control of glutamatergicactivity: astrocytes as stars of the show. Trends Neurosci. 27, 735–743.
Kimmich G. A., Roussie J., Manglapus M. and Randles J. (2001)Characterization of Na+-coupled glutamate/aspartate transport by arat brain astrocyte line expressing GLAST and EAAC1. J. Membr.Biol. 182, 17–30.
Korzhevskii D. E., Lentsman M. V., Gilyarov A. V., Kirik O. V. andVlasov T. D. (2008) Induction of nestin synthesis in rat brain cellsby ischemic damage. Neurosci. Behav. Physiol. 38, 139–143.
Kozlova E. N. and Takenaga K. (2005) A procedure for culturing as-trocytes from white matter and the application of the siRNAtechnique for silencing the expression of their specific marker,S100A4. Brain Res. Brain Res. Protoc. 15, 59–65.
Krum J. M., Phillips T. M. and Rosenstein J. M. (2002) Changes inastroglial GLT-1 expression after neural transplantation or stabwounds. Exp. Neurol. 174, 137–149.
Kukley M., Capetillo-Zarate E. and Dietrich D. (2007) Vesicularglutamate release from axons in white matter. Nat. Neurosci. 10,311–320.
Liedtke W., Edelmann W., Bieri P. L., Chiu F. C., Cowan N. J.,Kucherlapati R. and Raine C. S. (1996) GFAP is necessary for theintegrity of CNS white matter architecture and long-term mainte-nance of myelination. Neuron 17, 607–615.
Maragakis N. J., Dietrich J., Wong V., Xue H., Mayer-Proschel M., RaoM. S. and Rothstein J. D. (2004) Glutamate transporter expressionand function in human glial progenitors. Glia 45, 133–143.
Matute C., Alberdi E., Domercq M., Sanchez-Gomez M. V., Perez-Samartin A., Rodriguez-Antiguedad A. and Perez-Cerda F. (2007)Excitotoxic damage to white matter. J. Anat. 210, 693–702.
Miller R. H. and Raff M. C. (1984) Fibrous and protoplasmic astrocytesare biochemically and developmentally distinct. J. Neurosci. 4,585–592.
Miller R. H., Abney E. R., Davis S., Ffrench-Constant C., Lindsay R.,Patel R., Stone J. and Raff M. C. (1986) Is reactive gliosis aproperty of a distinct subpopulation of astrocytes? J. Neurosci. 6,22–29.
Pekny M. and Nilsson M. (2005) Astrocyte activation and reactivegliosis. Glia 50, 427–434.
Raff M. C., Abney E. R., Cohen J., Lindsay R. and Noble M. (1983)Two types of astrocytes in cultures of developing rat white matter:differences in morphology, surface gangliosides, and growthcharacteristics. J. Neurosci. 3, 1289–1300.
Stanimirovic D. B., Ball R., Small D. L. and Muruganandam A. (1999)Developmental regulation of glutamate transporters and glutaminesynthetase activity in astrocyte cultures differentiated in vitro.Int. J. Dev. Neurosci. 17, 173–184.
Sullivan S. M., Lee A., Bjorkman S. T., Miller S. M., Sullivan R. K.,Poronnik P., Colditz P. B. and Pow D. V. (2007) Cytoskeletalanchoring of GLAST determines susceptibility to brain damage: anidentified role for GFAP. J. Biol. Chem. 282, 29414–29423.
Swanson R. A., Liu J., Miller J. W., Rothstein J. D., Farrell K., Stein B.A. and Longuemare M. C. (1997) Neuronal regulation of glutamatetransporter subtype expression in astrocytes. J. Neurosci. 17, 932–940.
Tamagno I. and Schiffer D. (2006) Nestin expression in reactive astro-cytes of human pathology. J. Neurooncol. 80, 227–233.
Tawfik V. L., Regan M. R., Haenggeli C., Lacroix-Fralish M. L., Nutile-McMenemy N., Perez N., Rothstein J. D. and DeLeo J. A. (2008)Propentofylline-induced astrocyte modulation leads to alterationsin glial glutamate promoter activation following spinal nervetransection. Neuroscience 152, 1086–1092.
Vermeiren C., Najimi M., Maloteaux J. M. and Hermans E. (2005a)Molecular and functional characterisation of glutamate transportersin rat cortical astrocytes exposed to a defined combination of growthfactors during in vitro differentiation. Neurochem. Int. 46, 137–147.
Vermeiren C., Najimi M., Vanhoutte N., Tilleux S., de Hemptinne I.,Maloteaux J. M. and Hermans E. (2005b) Acute up-regulation ofglutamate uptake mediated by mGluR5a in reactive astrocytes.J. Neurochem. 94, 405–416.
Vermeiren C., de Hemptinne I., Vanhoutte N., Tilleux S., MaloteauxJ. M. and Hermans E. (2006) Loss of metabotropic glutamatereceptor-mediated regulation of glutamate transport in chemicallyactivated astrocytes in a rat model of amyotrophic lateral sclerosis.J. Neurochem. 96, 719–731.
Wilhelmsson U., Bushong E. A., Price D. L., Smarr B. L., Phung V.,Terada M., Ellisman M. H. and Pekny M. (2006) Redefining theconcept of reactive astrocytes as cells that remain within theirunique domains upon reaction to injury. Proc. Natl Acad. Sci. USA103, 17513–17518.
Yarowsky P. J. and Krueger B. K. (1989) Development of saxitoxin-sensitive and insensitive sodium channels in cultured neonatal ratastrocytes. J. Neurosci. 9, 1055–1061.
Ziskin J. L., Nishiyama A., Rubio M., Fukaya M. and Bergles D. E.(2007) Vesicular release of glutamate from unmyelinated axons inwhite matter. Nat. Neurosci. 10, 321–330.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 108, 1442–1452� 2009 The Authors
1452 | S. Goursaud et al.
Related Documents