Ž . Molecular Brain Research 67 1999 296–302 Short communication Expression of glutamate transporters in the adult bovine corpus callosum Marıa Domercq, Carlos Matute ) ´ Departamento de Neurociencias, Facultad de Medicina y Odontologıa, UniÕersidad del Paıs Vasco, 48940 Leioa, Vizcaya, Spain ´ ´ Accepted 9 February 1999 Abstract The presence and distribution of the glutamate transporters, EAAC1, GLAST and GLT-1, were examined in the adult bovine corpus callosum by means of Western blotting and immunohistochemistry. We found GLAST to be expressed in oligodendrocytes and in some astrocytes. GLT-1 was located throughout the processes of fibrous astrocytes. In addition, EAAC1 was present in a small population of cells that are probably adult oligodendrocyte progenitor cells. q 1999 Elsevier Science B.V. All rights reserved. Keywords: Astrocyte; Oligodendrocyte; Progenitor cell; Glutamate homeostasis Glial cells are endowed with a variety of neurotrans- mitter receptors including those for the excitatory amino w x acid glutamate 29 . In white matter, tracts axons can signal to glial cells by releasing glutamate that may acti- vate receptors in nearby astrocytes and oligodendrocytes w x 24 . In addition to the physiological consequences that glial glutamate receptor activation may have, it has re- cently been demonstrated that excessive glutamate receptor w activation leads to excitotoxic oligodendroglial death 18– x 20 . Thus, alterations in glutamatergic homeostasis in white matter may have dramatic effects on the viability of oligo- dendrocytes. The physiological extracellular concentration of gluta- mate is regulated by the action of high-affinity, sodium-de- pendent glutamate transporters. At least five glutamate transporters have been distinguished on the basis of differ- ences in sequence, pharmacology, tissue distribution and w x channel-like properties 12 . Localization studies have indi- cated that EAAC1 is mainly present in glutamatergic w x w x neurons 11 , GLAST in cerebellar Bergmann glia 30 and wx GLT-1 in astroglia throughout the brain 4 . The expres- sion of EAAT4 and EAAT5 is largely restricted to cerebel- lar Purkinje cells and to cells in the retina, respectively w x 1,5 . According to these studies, EAAC1 has been re- garded as the major neuronal glutamate transporter whereas ) Corresponding author. Fax: q 34-94-4649266; E-mail: [email protected] GLAST and GLT-1 are viewed as the glial glutamate w x transporters 25 . However, the distribution of glutamate transporters in the rat brain has not yet been completely established and there is immunohistochemical evidence suggesting that this broad classification might not be definitive. For instance, CA3r4 pyramidal cells of the hippocampus were labeled with GLT-1 specific probes in w x in situ hybridization experiments 27 . Conversely, EAAC1 wx is present in glial cells of the subcortical white matter 3 w x and possibly in oligodendrocytes 13 . In order to characterize the glutamate transporters ex- pressed by glial cells in white matter regions we have analyzed the expression of EAAC1, GLAST and GLT-1, the most widely distributed glutamate transporters, in the adult bovine corpus callosum, a white matter area particu- larly well suited for morphological studies, which we have extensively characterized regarding cell phenotypic fea- w x tures and neurotransmitter receptors 8,9 . Adult bovine brains were obtained from the slaughter- house and samples of the corpus callosum were removed within 1 h after death. For Western blot experiments, these samples were cleared of associated gray matter tracts, e.g., the indusium griseum, in order to avoid contaminating the samples with material from neuron cell bodies. The tissue Ž . was homogenized in 20 mM Tris–HCl pH 7.4 , 10% sucrose, 1 mM EDTA, 5 mM EGTA, 1 mM phenylmethyl- sulfonic fluoride, 10 mM benzamidine, 20 mgrml leu- peptin and 0.1 mM pepstatin, and subsequently centrifuged Ž . 10 000 = g for 30 min and stored at y808C until use. Aliquots of samples were diluted with an electrophoresis 0169-328Xr99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved. Ž . PII: S0169-328X 99 00072-8

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ž .Molecular Brain Research 67 1999 296–302
Short communication
Expression of glutamate transporters in the adult bovine corpus callosum
Marıa Domercq, Carlos Matute )´Departamento de Neurociencias, Facultad de Medicina y Odontologıa, UniÕersidad del Paıs Vasco, 48940 Leioa, Vizcaya, Spain´ ´
Accepted 9 February 1999
Abstract
The presence and distribution of the glutamate transporters, EAAC1, GLAST and GLT-1, were examined in the adult bovine corpuscallosum by means of Western blotting and immunohistochemistry. We found GLAST to be expressed in oligodendrocytes and in someastrocytes. GLT-1 was located throughout the processes of fibrous astrocytes. In addition, EAAC1 was present in a small population ofcells that are probably adult oligodendrocyte progenitor cells. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Astrocyte; Oligodendrocyte; Progenitor cell; Glutamate homeostasis
Glial cells are endowed with a variety of neurotrans-mitter receptors including those for the excitatory amino
w xacid glutamate 29 . In white matter, tracts axons cansignal to glial cells by releasing glutamate that may acti-vate receptors in nearby astrocytes and oligodendrocytesw x24 . In addition to the physiological consequences thatglial glutamate receptor activation may have, it has re-cently been demonstrated that excessive glutamate receptor
wactivation leads to excitotoxic oligodendroglial death 18–x20 . Thus, alterations in glutamatergic homeostasis in white
matter may have dramatic effects on the viability of oligo-dendrocytes.
The physiological extracellular concentration of gluta-mate is regulated by the action of high-affinity, sodium-de-pendent glutamate transporters. At least five glutamatetransporters have been distinguished on the basis of differ-ences in sequence, pharmacology, tissue distribution and
w xchannel-like properties 12 . Localization studies have indi-cated that EAAC1 is mainly present in glutamatergic
w x w xneurons 11 , GLAST in cerebellar Bergmann glia 30 andw xGLT-1 in astroglia throughout the brain 4 . The expres-
sion of EAAT4 and EAAT5 is largely restricted to cerebel-lar Purkinje cells and to cells in the retina, respectivelyw x1,5 . According to these studies, EAAC1 has been re-garded as the major neuronal glutamate transporter whereas
) Corresponding author. Fax: q 34-94-4649266; E-mail:[email protected]
GLAST and GLT-1 are viewed as the glial glutamatew xtransporters 25 . However, the distribution of glutamate
transporters in the rat brain has not yet been completelyestablished and there is immunohistochemical evidencesuggesting that this broad classification might not bedefinitive. For instance, CA3r4 pyramidal cells of thehippocampus were labeled with GLT-1 specific probes in
w xin situ hybridization experiments 27 . Conversely, EAAC1w xis present in glial cells of the subcortical white matter 3
w xand possibly in oligodendrocytes 13 .In order to characterize the glutamate transporters ex-
pressed by glial cells in white matter regions we haveanalyzed the expression of EAAC1, GLAST and GLT-1,the most widely distributed glutamate transporters, in theadult bovine corpus callosum, a white matter area particu-larly well suited for morphological studies, which we haveextensively characterized regarding cell phenotypic fea-
w xtures and neurotransmitter receptors 8,9 .Adult bovine brains were obtained from the slaughter-
house and samples of the corpus callosum were removedwithin 1 h after death. For Western blot experiments, thesesamples were cleared of associated gray matter tracts, e.g.,the indusium griseum, in order to avoid contaminating thesamples with material from neuron cell bodies. The tissue
Ž .was homogenized in 20 mM Tris–HCl pH 7.4 , 10%sucrose, 1 mM EDTA, 5 mM EGTA, 1 mM phenylmethyl-sulfonic fluoride, 10 mM benzamidine, 20 mgrml leu-peptin and 0.1 mM pepstatin, and subsequently centrifugedŽ .10 000=g for 30 min and stored at y808C until use.Aliquots of samples were diluted with an electrophoresis
0169-328Xr99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0169-328X 99 00072-8
( )M. Domercq, C. MatuterMolecular Brain Research 67 1999 296–302 297
w xsample buffer 14 , boiled for 5 min, subjected to elec-trophoresis in 7.5% polyacrylamide gels containing SDSand electroblotted to polyvinylidene fluoride membranesŽ .Immobilon P, Millipore . After blocking non-specificbinding for 1 h with 10% non-fat powdered milk and 2%bovine serum albumin, the membranes were incubated for48 h at 48C with affinity purified polyclonal antibodies to
Ž w x.EAAC1 C-terminal antibody; 0.6 mgrml; 25 , GLASTŽ w x. ŽN-terminal antibody; 0.2 mgrml; 26 and GLT-1 C-
w x.terminal antibody; 37 ngrml; 25 in Tris-buffered salinecontaining 0.1% Tween-20 and 1% nonfat powdered milk.Antibody binding was detected with horseradish–per-
Ž .oxidase conjugated anti-rabbit antibody 1:1000, SigmaŽand an enhanced chemiluminescence substrate Super Sig-
.nal ULTRA, Pierce .Tissue fixation, sectioning and processing for conven-
tional immunoperoxidase histochemistry were carried outw xas described before 8 . Antibodies to glutamate trans-
porters were used at the following concentrations; C-Ž . Ž .EAAC1 66 ngrml , N-GLAST 40 ngrml and C-GLT-1
Ž .37 ngrml . In addition, the immunohistochemical resultsobtained with the antibody to the N-terminal of GLASTwere corroborated with different antibodies to N-terminal
Ž w x.and C-terminal epitopes; N-GLAST 0.5 mgrml; 15 ,Ž w x.A522 to aminoacid residues 522–541; 0.1 mgrml; 15
Ž .and C-GLAST 1:4000; Chemicon . A polyclonal antibodyw xto nestin was used at a 1:200 dilution 31 . Negative
controls in all immunostaining experiments included theomission of the primary antibody and yielded no labeling.
Double immunofluorescent labeling experiments werecarried out at 48C. Sections were incubated with antibody
Ž .to glutamate transporters EAAC1 0.66 mgrml , GLASTŽ . Ž .0.4 mgrml or GLT-1 0.37 mgrml and with one anti-body to the following glial markers, glial fibrillary acidic
Žprotein GFAP; monoclonal, 0.2 mgrml; Boehringer. Ž . X XMannheim , GFAP polyclonal, 20 mgrml; Dako , 2 ,3 -
X Žcyclicnucleotide-3 -phosphodiesterase CNPase; 1:100;. ŽSigma , glutathione-S-transferase Yp subunit GST-Yp;
.1:500; Biotrin, Ireland and platelet derived growth factorŽ .receptor-alpha PDGFR-a; 0.5 mgrml; Santa Cruz . Pri-
mary antibody binding was viewed with a fluoresceinŽisothiocyanate-conjugated goat anti-rabbit IgG Alexae
.488; 1:200; Molecular Probes and with biotinylated anti-Ž .bodies to mouse or rat IgGs 1:200; Vector followed by a
streptavidin–tetramethyl rhodamine isothiocyanate-Ž .conjugate 1:100; Chemicon . When the two antibodies
used in double labeling experiments were from the samespecies, tissue sections were incubated with 8% goat serumafter completing immunohistochemistry with the first pri-mary antibody, and then the same procedure stated abovewas repeated using the second primary antibody. Negativecontrols in all the immunostaining experiments includedthe omission of one of the primary antibodies and yield nolabeling, indicating the absence of secondary antibodycross-reactivity.
A quantitative estimate of the number of cells express-ing each of the glutamate transporters examined was ob-tained from double immunofluorescence stained sectionstaken from two to three experiments, by counting thenumber of labeled cells in 12–15 randomly selected fieldsper section. The percentage of cells bearing the trans-porters within each cell class was calculated using as areference the number obtained by counting GFAPq astro-cytes, CNPaseq oligodendrocytes and GST-Ypq andPDGFR-aq oligodendrocyte precursors.
The specificity of the antibodies employed to glutamatew xtransporters has been analyzed previously 25,26 . Accord-
ingly, we show here that antibodies to the glutamate
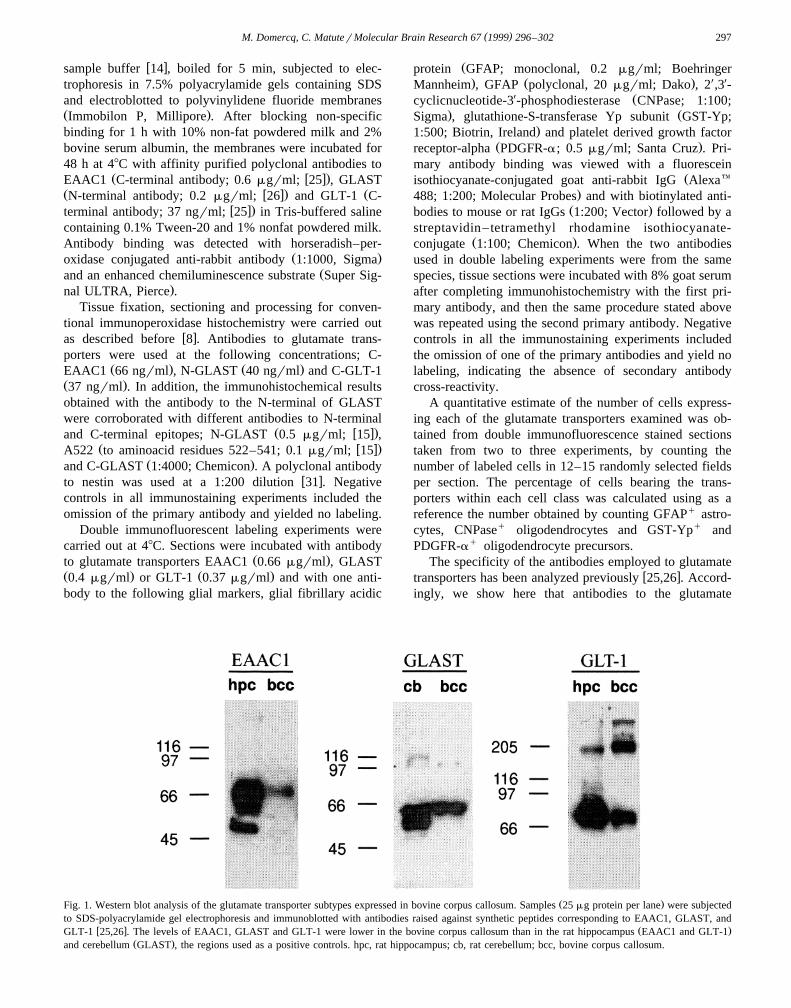
Ž .Fig. 1. Western blot analysis of the glutamate transporter subtypes expressed in bovine corpus callosum. Samples 25 mg protein per lane were subjectedto SDS-polyacrylamide gel electrophoresis and immunoblotted with antibodies raised against synthetic peptides corresponding to EAAC1, GLAST, and
w x Ž .GLT-1 25,26 . The levels of EAAC1, GLAST and GLT-1 were lower in the bovine corpus callosum than in the rat hippocampus EAAC1 and GLT-1Ž .and cerebellum GLAST , the regions used as a positive controls. hpc, rat hippocampus; cb, rat cerebellum; bcc, bovine corpus callosum.
( )M. Domercq, C. MatuterMolecular Brain Research 67 1999 296–302298
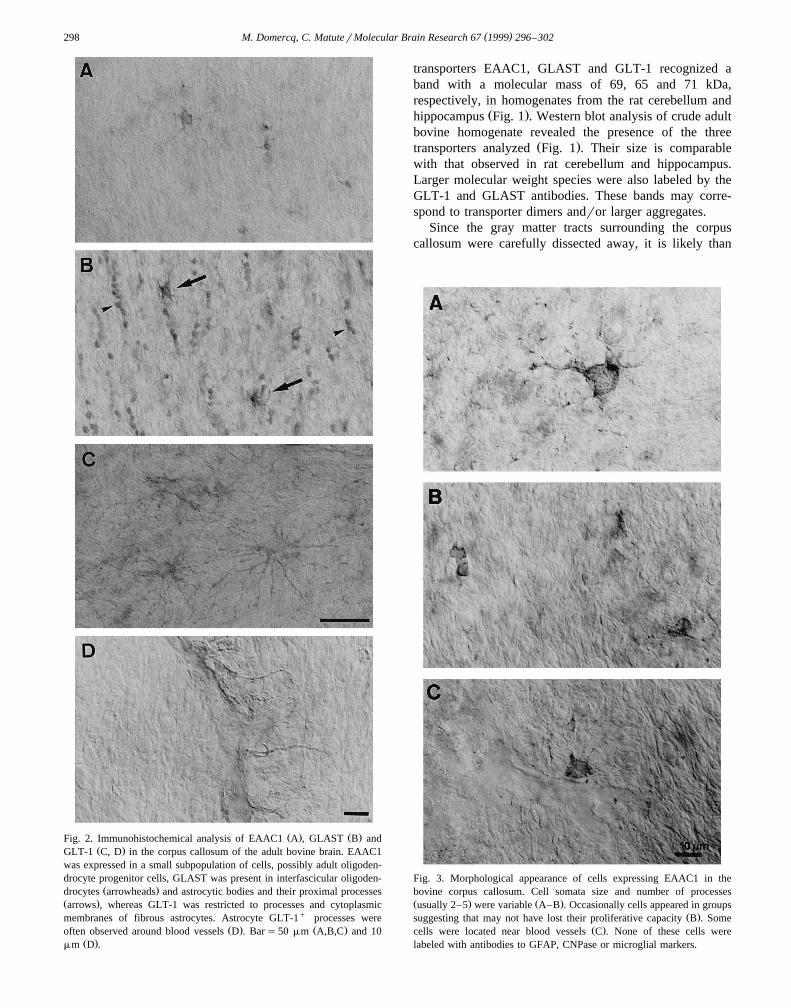
Ž . Ž .Fig. 2. Immunohistochemical analysis of EAAC1 A , GLAST B andŽ .GLT-1 C, D in the corpus callosum of the adult bovine brain. EAAC1
was expressed in a small subpopulation of cells, possibly adult oligoden-drocyte progenitor cells, GLAST was present in interfascicular oligoden-
Ž .drocytes arrowheads and astrocytic bodies and their proximal processesŽ .arrows , whereas GLT-1 was restricted to processes and cytoplasmicmembranes of fibrous astrocytes. Astrocyte GLT-1q processes were
Ž . Ž .often observed around blood vessels D . Bars50 mm A,B,C and 10Ž .mm D .
transporters EAAC1, GLAST and GLT-1 recognized aband with a molecular mass of 69, 65 and 71 kDa,respectively, in homogenates from the rat cerebellum and
Ž .hippocampus Fig. 1 . Western blot analysis of crude adultbovine homogenate revealed the presence of the three
Ž .transporters analyzed Fig. 1 . Their size is comparablewith that observed in rat cerebellum and hippocampus.Larger molecular weight species were also labeled by theGLT-1 and GLAST antibodies. These bands may corre-spond to transporter dimers andror larger aggregates.
Since the gray matter tracts surrounding the corpuscallosum were carefully dissected away, it is likely than
Fig. 3. Morphological appearance of cells expressing EAAC1 in thebovine corpus callosum. Cell somata size and number of processesŽ . Ž .usually 2–5 were variable A–B . Occasionally cells appeared in groups
Ž .suggesting that may not have lost their proliferative capacity B . SomeŽ .cells were located near blood vessels C . None of these cells were
labeled with antibodies to GFAP, CNPase or microglial markers.

( )M. Domercq, C. MatuterMolecular Brain Research 67 1999 296–302 299
the Western blot immunostaining represents transporterslocalized to glial cells or axons. To confirm this idea andto determine the phenotype of the cells expressing thethree glutamate transporters examined, we carried out im-munohistochemical experiments on adult bovine corpuscallosum. Thus, antibodies to EAAC1, GLAST and GLT-1strongly stained morphologically different types of glial
Ž .cells in the corpus callosum Fig. 2 .The expression of EAAC1 transporter was detected in
some cells with no particular orientation, a variable num-Ž .ber of branches between 2 and 5 and different soma sizes
Ž .Figs. 2A and 3 . We studied in more detail the morphol-ogy of EAAC1q cells by drawing with camera lucida arepresentative sample of them and comparing their mor-phology with that of cells expressing the filament protein
Ž . qnestin Fig. 4 . The EAAC1 cells exhibited a simplemorphology which resembled that of a subpopulation ofnestinq cells, a marker of immature cells in adult rat
w xsubcortical white matter 10,31 . To characterize furtherthe phenotype of the immature cells expressing EAAC1,we carried out double immunolabeling experiments usingpolyclonal antibodies to GST-Yp and to PDGFR-a , two
w xmarkers of immature oligodendrocytes 10,21 . Thus, wefound that all cells expressing EAAC1 were GST-Ypq or
q Ž .PDGFR-a Fig. 5 . In addition, we estimated that thepercentage of GST-Ypq and PDGFR-aq cells bearing thetransporter was 46"3% and 41"3%, respectively. Incontrast, EAAC1 was not detected in GFAPq astrocytes,confirming the absence of cross-reactivity between both
Žsecondary antibodies in the procedure employed data not.shown . All these features suggest that in adult bovine
corpus callosum EAAC1 was located in a subpopulation ofimmature oligodendroglial cells, probably adult oligoden-
drocyte progenitor cells, a quiescent population known tow xbe present in the mature CNS 6,7 . Accordingly, previous
reports have described the expression of EAAC1 in thecorpus callosum and suggested that the transporter may be
w xpresent in oligodendrocytes 13 . In addition, co-localiza-tion studies in sequential sections have demonstrated thatmost EAAC1q cells in subcortical white matter are GFAPy
w x3 .The GLAST transporter was expressed in small rounded
cells with no visible processes, frequently grouped intoŽshort rows of cells oriented parallel to axon fibers Fig.
.2B , a feature which is characteristic of interfascicularw xoligodendrocytes 23 . This idea was confirmed by double
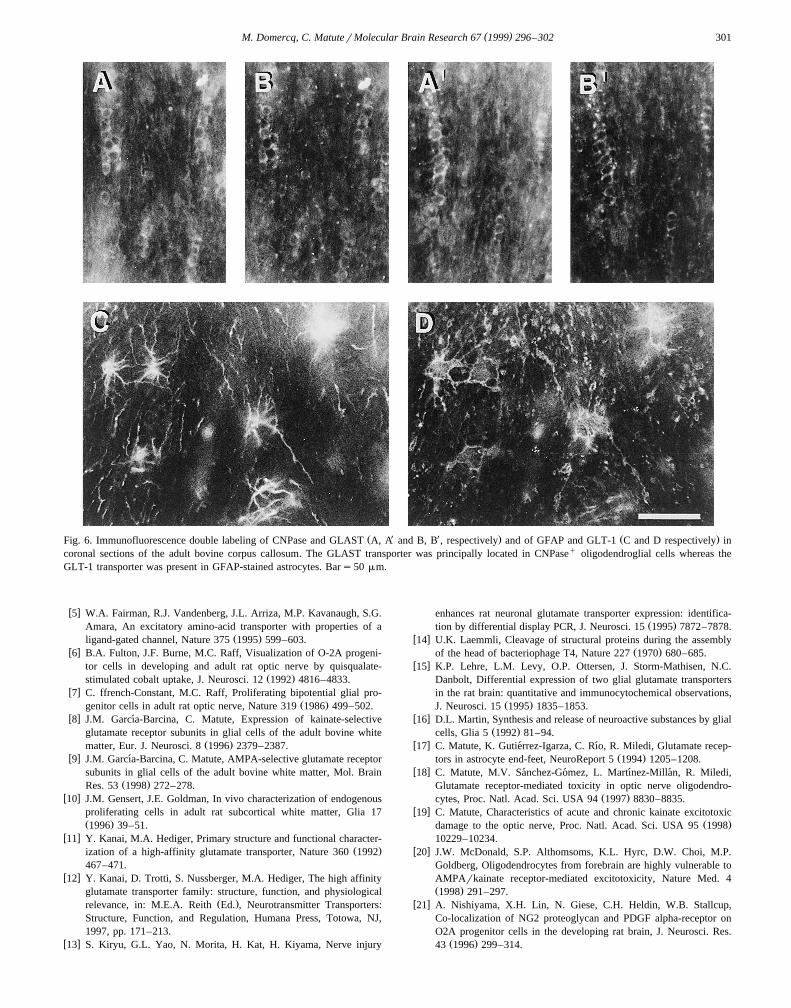
label immunohistochemical experiments using antibodiesto GLAST and to CNPase, an oligodendrocyte markerŽ X X.Fig. 6A, B and A, B . The percentage of oligodendro-cytes expressing GLAST was 71"3%. In addition, wealso observed immunolabeling with antibodies to GLASTin a minor subpopulation of cells with the typical stellateappearance of astrocytes. In the latter cells, both cell
Ž .bodies and proximal process were stained Fig. 2B . Athigher magnification, some fiber and punctuate-like im-munostaining parallel to neuronal axons was observedwhich may correspond to glutamate transporters in axons
Ž .or at Ranvier nodes data not shown . However, confirma-tion of this will require analysis by electron microscopy.The GLAST transporter has been described previously inastrocytes in the rat corpus callosum but not in oligoden-
w xdrocytes 15,28 . In order to confirm our results, we per-formed immunohistochemistry with an additional antibody
w xto the N-terminal of GLAST 15 and with two additionalŽ w xantibodies to C-terminal epitopes antibody A522 15 and.C-terminal antibody from Chemicon . Similar results were
Fig. 4. Camera lucida drawings of EAAC1q cells and nestinq cells. Although some of the cells appear quite complex in morphology, they are still muchsimpler than multi-branched astrocytes or microglial cells and can be clearly distinguished from interfascicular oligodendrocytes. A morphologicallyheterogeneous population of cells was labeled by nestin antibodies, possibly reflecting different developmental stages. Note the similar appearance ofEAAC1q cells and many of the nestin-expressing cells. Bars50 mm.
( )M. Domercq, C. MatuterMolecular Brain Research 67 1999 296–302300
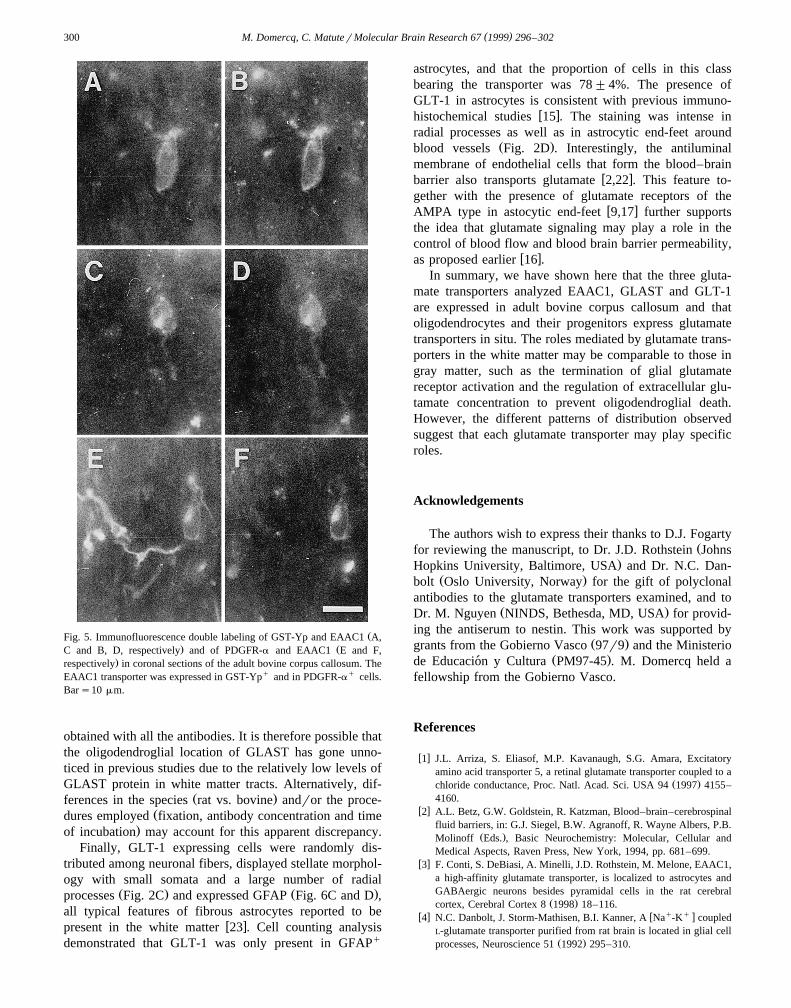
ŽFig. 5. Immunofluorescence double labeling of GST-Yp and EAAC1 A,. ŽC and B, D, respectively and of PDGFR-a and EAAC1 E and F,
.respectively in coronal sections of the adult bovine corpus callosum. TheEAAC1 transporter was expressed in GST-Ypq and in PDGFR-aq cells.Bars10 mm.
obtained with all the antibodies. It is therefore possible thatthe oligodendroglial location of GLAST has gone unno-ticed in previous studies due to the relatively low levels ofGLAST protein in white matter tracts. Alternatively, dif-
Ž .ferences in the species rat vs. bovine andror the proce-Ždures employed fixation, antibody concentration and time
.of incubation may account for this apparent discrepancy.Finally, GLT-1 expressing cells were randomly dis-
tributed among neuronal fibers, displayed stellate morphol-ogy with small somata and a large number of radial
Ž . Ž .processes Fig. 2C and expressed GFAP Fig. 6C and D ,all typical features of fibrous astrocytes reported to be
w xpresent in the white matter 23 . Cell counting analysisdemonstrated that GLT-1 was only present in GFAPq
astrocytes, and that the proportion of cells in this classbearing the transporter was 78"4%. The presence ofGLT-1 in astrocytes is consistent with previous immuno-
w xhistochemical studies 15 . The staining was intense inradial processes as well as in astrocytic end-feet around
Ž .blood vessels Fig. 2D . Interestingly, the antiluminalmembrane of endothelial cells that form the blood–brain
w xbarrier also transports glutamate 2,22 . This feature to-gether with the presence of glutamate receptors of the
w xAMPA type in astocytic end-feet 9,17 further supportsthe idea that glutamate signaling may play a role in thecontrol of blood flow and blood brain barrier permeability,
w xas proposed earlier 16 .In summary, we have shown here that the three gluta-
mate transporters analyzed EAAC1, GLAST and GLT-1are expressed in adult bovine corpus callosum and thatoligodendrocytes and their progenitors express glutamatetransporters in situ. The roles mediated by glutamate trans-porters in the white matter may be comparable to those ingray matter, such as the termination of glial glutamatereceptor activation and the regulation of extracellular glu-tamate concentration to prevent oligodendroglial death.However, the different patterns of distribution observedsuggest that each glutamate transporter may play specificroles.
Acknowledgements
The authors wish to express their thanks to D.J. FogartyŽfor reviewing the manuscript, to Dr. J.D. Rothstein Johns
.Hopkins University, Baltimore, USA and Dr. N.C. Dan-Ž .bolt Oslo University, Norway for the gift of polyclonal
antibodies to the glutamate transporters examined, and toŽ .Dr. M. Nguyen NINDS, Bethesda, MD, USA for provid-
ing the antiserum to nestin. This work was supported byŽ .grants from the Gobierno Vasco 97r9 and the Ministerio
Ž .de Educacion y Cultura PM97-45 . M. Domercq held a´fellowship from the Gobierno Vasco.
References
w x1 J.L. Arriza, S. Eliasof, M.P. Kavanaugh, S.G. Amara, Excitatoryamino acid transporter 5, a retinal glutamate transporter coupled to a
Ž .chloride conductance, Proc. Natl. Acad. Sci. USA 94 1997 4155–4160.
w x2 A.L. Betz, G.W. Goldstein, R. Katzman, Blood–brain–cerebrospinalfluid barriers, in: G.J. Siegel, B.W. Agranoff, R. Wayne Albers, P.B.
Ž .Molinoff Eds. , Basic Neurochemistry: Molecular, Cellular andMedical Aspects, Raven Press, New York, 1994, pp. 681–699.
w x3 F. Conti, S. DeBiasi, A. Minelli, J.D. Rothstein, M. Melone, EAAC1,a high-affinity glutamate transporter, is localized to astrocytes andGABAergic neurons besides pyramidal cells in the rat cerebral
Ž .cortex, Cerebral Cortex 8 1998 18–116.w x w q q x4 N.C. Danbolt, J. Storm-Mathisen, B.I. Kanner, A Na -K coupled
L-glutamate transporter purified from rat brain is located in glial cellŽ .processes, Neuroscience 51 1992 295–310.
( )M. Domercq, C. MatuterMolecular Brain Research 67 1999 296–302 301
Ž X X . Ž .Fig. 6. Immunofluorescence double labeling of CNPase and GLAST A, A and B, B , respectively and of GFAP and GLT-1 C and D respectively incoronal sections of the adult bovine corpus callosum. The GLAST transporter was principally located in CNPaseq oligodendroglial cells whereas theGLT-1 transporter was present in GFAP-stained astrocytes. Bars50 mm.
w x5 W.A. Fairman, R.J. Vandenberg, J.L. Arriza, M.P. Kavanaugh, S.G.Amara, An excitatory amino-acid transporter with properties of a
Ž .ligand-gated channel, Nature 375 1995 599–603.w x6 B.A. Fulton, J.F. Burne, M.C. Raff, Visualization of O-2A progeni-
tor cells in developing and adult rat optic nerve by quisqualate-Ž .stimulated cobalt uptake, J. Neurosci. 12 1992 4816–4833.
w x7 C. ffrench-Constant, M.C. Raff, Proliferating bipotential glial pro-Ž .genitor cells in adult rat optic nerve, Nature 319 1986 499–502.
w x8 J.M. Garcıa-Barcina, C. Matute, Expression of kainate-selective´glutamate receptor subunits in glial cells of the adult bovine white
Ž .matter, Eur. J. Neurosci. 8 1996 2379–2387.w x9 J.M. Garcıa-Barcina, C. Matute, AMPA-selective glutamate receptor´
subunits in glial cells of the adult bovine white matter, Mol. BrainŽ .Res. 53 1998 272–278.
w x10 J.M. Gensert, J.E. Goldman, In vivo characterization of endogenousproliferating cells in adult rat subcortical white matter, Glia 17Ž .1996 39–51.
w x11 Y. Kanai, M.A. Hediger, Primary structure and functional character-Ž .ization of a high-affinity glutamate transporter, Nature 360 1992
467–471.w x12 Y. Kanai, D. Trotti, S. Nussberger, M.A. Hediger, The high affinity
glutamate transporter family: structure, function, and physiologicalŽ .relevance, in: M.E.A. Reith Ed. , Neurotransmitter Transporters:
Structure, Function, and Regulation, Humana Press, Totowa, NJ,1997, pp. 171–213.
w x13 S. Kiryu, G.L. Yao, N. Morita, H. Kat, H. Kiyama, Nerve injury
enhances rat neuronal glutamate transporter expression: identifica-Ž .tion by differential display PCR, J. Neurosci. 15 1995 7872–7878.
w x14 U.K. Laemmli, Cleavage of structural proteins during the assemblyŽ .of the head of bacteriophage T4, Nature 227 1970 680–685.
w x15 K.P. Lehre, L.M. Levy, O.P. Ottersen, J. Storm-Mathisen, N.C.Danbolt, Differential expression of two glial glutamate transportersin the rat brain: quantitative and immunocytochemical observations,
Ž .J. Neurosci. 15 1995 1835–1853.w x16 D.L. Martin, Synthesis and release of neuroactive substances by glial
Ž .cells, Glia 5 1992 81–94.w x17 C. Matute, K. Gutierrez-Igarza, C. Rıo, R. Miledi, Glutamate recep-´ ´
Ž .tors in astrocyte end-feet, NeuroReport 5 1994 1205–1208.w x18 C. Matute, M.V. Sanchez-Gomez, L. Martınez-Millan, R. Miledi,´ ´ ´ ´
Glutamate receptor-mediated toxicity in optic nerve oligodendro-Ž .cytes, Proc. Natl. Acad. Sci. USA 94 1997 8830–8835.
w x19 C. Matute, Characteristics of acute and chronic kainate excitotoxicŽ .damage to the optic nerve, Proc. Natl. Acad. Sci. USA 95 1998
10229–10234.w x20 J.W. McDonald, S.P. Althomsoms, K.L. Hyrc, D.W. Choi, M.P.
Goldberg, Oligodendrocytes from forebrain are highly vulnerable toAMPArkainate receptor-mediated excitotoxicity, Nature Med. 4Ž .1998 291–297.
w x21 A. Nishiyama, X.H. Lin, N. Giese, C.H. Heldin, W.B. Stallcup,Co-localization of NG2 proteoglycan and PDGF alpha-receptor onO2A progenitor cells in the developing rat brain, J. Neurosci. Res.
Ž .43 1996 299–314.

( )M. Domercq, C. MatuterMolecular Brain Research 67 1999 296–302302
w x22 W.H. Oldendorf, J. Szabo, Amino acid assignment to one of threeŽ .blood–brain barrier amino acid carriers, Am. J. Physiol. 230 1976
94–98.w x23 A. Peters, S.L. Palay, H.D. Webster, The neuroglial cells, in: A.
Ž .Peters, S.L. Palay, H.D. Webster Eds. , The Fine Structure of theNervous System, Oxford Univ. Press, New York, 1991, pp. 273–311.
w x24 B.R. Ransom, R.K. Orkand, Glial–neuronal interactions in non-syn-aptic areas of the brain: studies in the optic nerve, Trends Neurosci.
Ž .19 1996 352–358.w x25 J.D. Rothstein, L. Martin, A.I. Levey, M. Dykes-Hoberg, L. Jin, D.
Wu, N. Nash, R.W. Kunel, Localization of neuronal and glialŽ .glutamate transporters, Neuron 13 1994 713–725.
w x26 J.D. Rothstein, M. Dykes-Hoberg, C. Pardo, L.A. Bristol, L. Jin,R.W. Kuncl, Y. Kanai, M.A. Hediger, Y. Wang, J.P. Schielke, D.F.Welty, Knockout of glutamate transporters reveals a major role forastroglial transport in excitotoxicity and clearance of glutamate,
Ž .Neuron 16 1996 675–686.w x27 A. Schmitt, E. Asan, B. Puschel, Th. Jons, P. Kugler, Expression of¨ ¨
the glutamate transporter GLT1 in neural cells of the rat centralnervous system: non-radioactive in situ hybridization and compara-
Ž .tive immunocytochemistry, Neuroscience 71 1996 989–1004.w x28 A. Schmitt, E. Asan, B. Puschel, P. Kugler, Cellular and regional¨
distribution of the glutamate transporter GLAST in the CNS of rats:nonradioactive in situ hybridization and comparative immunocyto-
Ž .chemistry, J. Neurosci. 17 1997 1–10.w x29 C. Steinhauser, V. Gallo, News on glutamate receptors in glial cells,¨
Ž .Trends Neurosci. 19 1996 339–345.w x30 T. Storck, S. Schulte, K. Hofmann, W. Stoffel, Structure, expres-
sion, and functional analysis of a Naq-dependent glutamateraspar-Ž .tate transporter from rat brain, Proc. Natl. Acad. Sci. USA 89 1992
10955–10959.w x31 T. Tohyama, V.M.-L. Lee, L.B. Rorke, M. Marvin, R.D.G. McKay,
J.Q. Trojanowski, Nestin expression in embryonic human neuroep-ithelium and in human neuroepithelial tumor cells, Lab. Invest. 66Ž .1992 303–313.
Related Documents