Coral barium/calcium record of mid-Holocene upwelling activity in New Caledonia, South-West Pacific Lucien F. Montaggioni a, ⁎ , Florence Le Cornec b , Thierry Corrège b , Guy Cabioch c a Département des Sciences de la Terre et de I'Environnement, CNRS-FRE 2761, Université Provence,13331 Marseille Cedex 3, France b IRD, Institut de Recherche Pour le Développement, UR055 “Paléotropique”, 32 Avenue Varagnat, 93143 Bondy, France c IRD, Institut de Recherche Pour le Développement, UR055 “Paléotropique”, B.P. A5, 98848 Nouméa Cedex, New Caledonia Received 22 March 2005; received in revised form 2 November 2005; accepted 23 December 2005 Abstract A variety of inimical factors, i.e. sea surface temperature, nutrient input, nature of substrates, were invoked previously to explain the retardation in reef settlement in New Caledonia during the Holocene. In order to clarify the respective role played by temperature and nutrient level in the growth of fringing reefs along the southwest coast of the island, trace element ratios, i.e. Sr/Ca and Ba/Ca, were used, respectively, as a paleothermometer and as a tracer of nutrient load from modern and mid-Holocene (about 6000 calendar years old) massive Porites coral colonies. The results obtained from the modern coral record indicate that, near the outer barrier reef, upwelling activity is controlled by southeast winds blowing in summer. By contrast, along the southwest coastline, the fringing reefs are subjected to barium input mainly derived from weathering of barite-rich rocks cropping out in the adjacent mainland. Comparison between the fossil and modern coral Sr/Ca and Ba/Ca records shed light on three different aspects of the mid- Holocene climate and coral reef evolution in New Caledonia: (1) the variability of paleo-sea surface temperature (SST) seems to have been similar to the modern one, although the island probably experienced SST about 1 °C higher than at present; (2) upwelling activity appears to have been greater than at present. Barium being supplied from the open sea to coastal zones by strong-wind generated upwelling currents, in relation to cooling events; (3) the onset of reef development in the southwest areas of New Caledonia was delayed due to the stronger upwelling of nutrient-enriched waters during the early–middle Holocene. The flowing of upwelled waters over the southwest coastal areas began at 8200 calendar years BP, and ceased at the time when the outer barrier reef crest reached present-day sea level, about 5500 to 5000 calendar years ago. © 2006 Elsevier B.V. All rights reserved. Keywords: Trace elements; Barium; Strontium; Corals; Reefs; Paleoclimate; Modern; Mid-Holocene; New Caledonia; South Pacific 1. Introduction The growth history of reef-building corals is known to provide some of the richest archives of past climate and environments (for instance, see Montaggioni and Macintyre, 1991; Gagan et al., 2000; Quinn and Tudhope, 2000; Swart and Grottoli, 2003). During the last two decades, in association with the increasing development of subsurface drilling operations through reef sites, a great body of work has been produced to identify the main environmental factors that have controlled coral reef growth since the Last Glacial Maximum (Smith and Buddemeier, 1992; Hubbard, Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436 – 455 www.elsevier.com/locate/palaeo ⁎ Corresponding author. E-mail address: [email protected] (L.F. Montaggioni). 0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2005.12.018

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

alaeoecology 237 (2006) 436–455www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Coral barium/calcium record of mid-Holocene upwelling activity inNew Caledonia, South-West Pacific
Lucien F. Montaggioni a,⁎, Florence Le Cornec b, Thierry Corrège b, Guy Cabioch c
a Département des Sciences de la Terre et de I'Environnement, CNRS-FRE 2761, Université Provence,13331 Marseille Cedex 3, Franceb IRD, Institut de Recherche Pour le Développement, UR055 “Paléotropique”, 32 Avenue Varagnat, 93143 Bondy, France
c IRD, Institut de Recherche Pour le Développement, UR055 “Paléotropique”, B.P. A5, 98848 Nouméa Cedex, New Caledonia
Received 22 March 2005; received in revised form 2 November 2005; accepted 23 December 2005
Abstract
Avariety of inimical factors, i.e. sea surface temperature, nutrient input, nature of substrates, were invoked previously to explainthe retardation in reef settlement in New Caledonia during the Holocene. In order to clarify the respective role played bytemperature and nutrient level in the growth of fringing reefs along the southwest coast of the island, trace element ratios, i.e. Sr/Caand Ba/Ca, were used, respectively, as a paleothermometer and as a tracer of nutrient load from modern and mid-Holocene (about6000 calendar years old) massive Porites coral colonies. The results obtained from the modern coral record indicate that, near theouter barrier reef, upwelling activity is controlled by southeast winds blowing in summer. By contrast, along the southwestcoastline, the fringing reefs are subjected to barium input mainly derived from weathering of barite-rich rocks cropping out in theadjacent mainland.
Comparison between the fossil and modern coral Sr/Ca and Ba/Ca records shed light on three different aspects of the mid-Holocene climate and coral reef evolution in New Caledonia: (1) the variability of paleo-sea surface temperature (SST) seems tohave been similar to the modern one, although the island probably experienced SST about 1 °C higher than at present; (2)upwelling activity appears to have been greater than at present. Barium being supplied from the open sea to coastal zones bystrong-wind generated upwelling currents, in relation to cooling events; (3) the onset of reef development in the southwest areas ofNew Caledonia was delayed due to the stronger upwelling of nutrient-enriched waters during the early–middle Holocene. Theflowing of upwelled waters over the southwest coastal areas began at 8200 calendar years BP, and ceased at the time when the outerbarrier reef crest reached present-day sea level, about 5500 to 5000 calendar years ago.© 2006 Elsevier B.V. All rights reserved.
Keywords: Trace elements; Barium; Strontium; Corals; Reefs; Paleoclimate; Modern; Mid-Holocene; New Caledonia; South Pacific
1. Introduction
The growth history of reef-building corals is knownto provide some of the richest archives of past climateand environments (for instance, see Montaggioni and
⁎ Corresponding author.E-mail address: [email protected] (L.F. Montaggioni).
0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2005.12.018
Macintyre, 1991; Gagan et al., 2000; Quinn andTudhope, 2000; Swart and Grottoli, 2003). During thelast two decades, in association with the increasingdevelopment of subsurface drilling operations throughreef sites, a great body of work has been produced toidentify the main environmental factors that havecontrolled coral reef growth since the Last GlacialMaximum (Smith and Buddemeier, 1992; Hubbard,

437L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
1997; Kleypas, 1997; Montaggioni, 2000, 2005; Hal-lock, 2001; Blanchon and Montaggioni, 2003;McLaughlin et al., 2003).
One of the pivotal discoveries relates to the age ofinitiation of active modern reefs. Data from reef coresindicate that these reefs commenced to settle relativelycontemporaneously throughout the world; the timing ofinitiation was bracketed mostly within 10–8 ka(ka=1000 calendar years BP) (Montaggioni, 1988,2000). However, in New Caledonia, the age of Holocenereef initiation is younger than 8.3 ka for barrier reefs andthan 7.6 ka for fringing reefs (Cabioch, 1988, 2003;Cabioch et al., 1995; Castellaro, 1999). Furthermore,fringing reef development was initiated non-uniformlythroughout the region. Reef colonization has varied inage according to location. For instance, along the south-west coasts (e.g. Nouméa area), initial colonization offringing reefs did not operate prior to 7 ka, when sealevel approached its present-day position, while in thesouth-east parts of the island, fringing reef initiationoccurred as early as 7.6 ka (Cabioch et al., 1995). Such avariation in the timing of fringing reef initiation in NewCaledonia was explained in several ways (Cabioch etal., 1995; Cabioch, 2003). Retardation in reef settlementalong the south-west coasts may have resulted from: 1)sea surface temperature (SST) unconducive to coral andreef growth; 2) the absence of antecedent substratessuitable to be coral-colonized prior to 7 ka as suggestedby the earliest establishment of fringing reefs preferen-tially on karstified limestone; 3) poor quality, elevatednutrient and turbidity levels of coastal waters related toincreased runoff and weathering; 4) local delivery ofnutrients controlled by strong upwelling activity.Indeed, nutrient supply from terrestrial runoff or oceanicupwelling is considered to play a critical role asimportant as temperature in limiting coral reef develop-ment (Kinsey and Davies, 1979; Hallock and Schlager,1986; Hallock, 1988; Montaggioni et al., 1993;Montaggioni, 2005).
To identify the major driving factors of reef re-tardation during the Holocene, we need to constrain therelationship between sea level, SST, possible changingpatterns of ocean circulation and river discharge, andtheir combined influence on reef development. Toachieve this, a biproxy approach was conducted usingcoral skeleton geochemistry. Indeed, it is now wellestablished that the Sr/Ca content of coral carbonatescan provide a robust tool for gaining high-resolutionrecords of recent to past SST variability (Smith et al.,1979; Beck et al., 1992; de Villiers et al., 1994; Gagan etal., 2000; Marshall and McCulloch, 2002; Abram et al.,2003; Ayliffe et al., 2004). Similarly, barium in sedi-
mentary solid phases was considered to be a powerfulproxy for oceanographic processes related to produc-tivity along continental margins (Lea and Boyle, 1989;Elderfield, 1990; Dymond et al., 1992; McManus et al.,1998; Kasten et al., 2001; Klump et al., 2001; PrakashBabu et al., 2002) and for river discharge (Edmond etal., 1978; Carroll et al., 1993; Coffey et al., 1997;Moore, 1997; Fagel et al., 2004). More particularly,Ba/Ca ratios were successfully applied to corals toreconstruct rainfall, terrestrial runoff and riverine input(Hanor and Chan, 1977; Shen and Sanford, 1990;Anderegg et al., 1997; Tudhope et al., 1997; McCul-loch et al., 2003; Alibert et al., 2003), lateral advectionor vertical upwelling events (Lea et al., 1989; Shen etal., 1992; Tudhope et al., 1997; Fallon et al., 1999;Reuer et al., 2003; Sinclair and McCulloch, 2004).
Firstly, in order to reconstruct paleo-SST andcompare them with present-day SST, detailed analysesof Sr/Ca ratios were performed on mid-Holocenemassive corals and on modern colonies of the samespecies. Secondly, Ba/Ca ratios were measured from allof the collected coral samples. Our initial hypothesis isthat, in the southwest part of the New Caledonianlagoon, periods of high nutrient inputs would berecorded by corals growing in shallow waters as periodsof deposition of barium-enriched skeleton.
The aims of the present study are thus to differentiatebetween the effects of changes in SST and changes innutrient concentration in controlling fringing reefdevelopment in southwestern New Caledonia duringthe mid-Holocene, and to identify the sources of nutrientflux.
2. Field setting
New Caledonia is located in the southwestern Pacificocean, between 20° and 23° S and 163° and 168° E, andis approximately 450 km long×50 km wide. It forms agroup of islands, aligned NW–SE and composed mainlyof sedimentary and volcanic formations ranging fromPermian to Quaternary in age (Paris, 1981). The mainisland (Grande-Terre) possesses a coastline fringed byreefs or mangroves and one of the larger barrier reeftracts in the world. The reef–lagoon system covers atotal area of 24,000 km2 (Fig. 1).The test area is situatedin the south-western lagoon (e.g. Nouméa region) thathas a mean depth of 17.5 m and a width varying from40 km in the south-east to around 5 km in the north-west.The barrier reef is cut by deep passes corresponding topaleorivers active during low sea level. The lagoonalareas close to embayments are expected to trapsignificant amount of lateritic material derived from
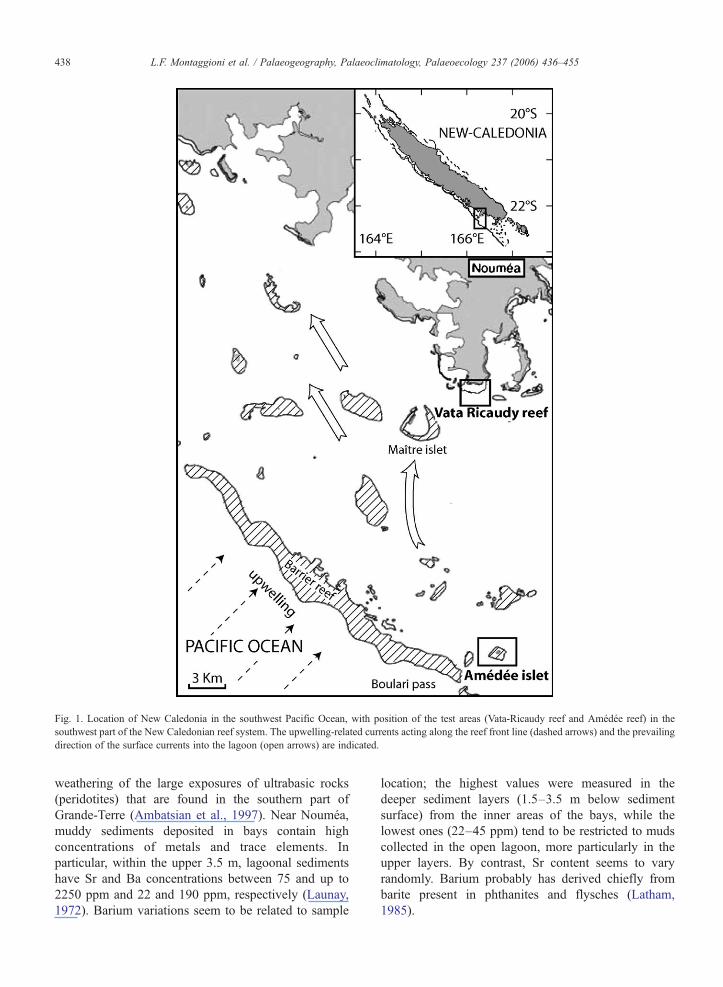
Fig. 1. Location of New Caledonia in the southwest Pacific Ocean, with position of the test areas (Vata-Ricaudy reef and Amédée reef) in thesouthwest part of the New Caledonian reef system. The upwelling-related currents acting along the reef front line (dashed arrows) and the prevailingdirection of the surface currents into the lagoon (open arrows) are indicated.
438 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
weathering of the large exposures of ultrabasic rocks(peridotites) that are found in the southern part ofGrande-Terre (Ambatsian et al., 1997). Near Nouméa,muddy sediments deposited in bays contain highconcentrations of metals and trace elements. Inparticular, within the upper 3.5 m, lagoonal sedimentshave Sr and Ba concentrations between 75 and up to2250 ppm and 22 and 190 ppm, respectively (Launay,1972). Barium variations seem to be related to sample
location; the highest values were measured in thedeeper sediment layers (1.5–3.5 m below sedimentsurface) from the inner areas of the bays, while thelowest ones (22–45 ppm) tend to be restricted to mudscollected in the open lagoon, more particularly in theupper layers. By contrast, Sr content seems to varyrandomly. Barium probably has derived chiefly frombarite present in phthanites and flysches (Latham,1985).

439L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
The general surface circulation pattern in this part ofthe Pacific Ocean can be described as a large-scaleanticyclonic gyre centred near 15°S (Donguy et al.,1970; Delcroix and Hénin, 1989). New Caledonia lies atthe south of the South Pacific Convergence Zone; itexperiences an oceanic tropical climate influenced byintertropical low pressures northwards and by thesubtropical anticyclonic belt southwards. This resultsin four distinct seasonal periods. The warm and wetseason (austral summer) lasts from mid-November tomid-April and is regularly affected by tropical depres-sions that can develop into cyclones. Cyclonic eventshave a frequency of 1–2 per year. The mid-April to mid-May period is transitional. The colder season (australwinter) is from mid-May to mid-September; it isperiodically subjected to polar disturbances that canresult in heavy rainfall. From mid-September to mid-November, the weather is the most stable and the leastrainy. Summer rainfall is typified by a high interannualvariability of about 30%, mainly related to tropicalcyclones. In the Nouméa region, annual rainfallaverages 1050 mm during rainy years and 500 mmduring dry ones. At Amédée Lighthouse, located closeto the southwest barrier reef rim, annual rainfallaverages 700 mm during rainy years, indicating thatrainfall is significantly higher on the mainland than inthe lagoon. The rivers flowing out from the Nouméaregion have average flood discharges of less than10 m3/s; during cyclonic events, maximum flow ratesof about 2000 m3/s can be reached for a few days(Launay, 1972; Météo France, 1995).The depletion ofrivers occurs generally from May to November andsometimes through to January in dry years. In thesouthernmost area, seasonal variability has a moderateeffect on river fluxes.
The two main parameters governing water circula-tion in the southwest New Caledonian lagoon are thewind-induced advection and the semi-diurnal tide.Swell-generated, cross-reef currents only have a localand limited control on water circulation on the back reefareas (Douillet et al., 2001; Pinazo et al., 2004). Thedominant currents are trade wind generated and arepropagated from east to west. As a result, the maindirection of the surface currents into the lagoon iswestward and constrained by the boundaries and shapeof the lagoon (Douillet et al., 2001). Finally, lagoonalwater masses move dominantly from the southeast to thenorthwest (Fig. 1), even if the tide induces varyingcurrent velocities during the tidal cycle that may beoriented differently from the wind-induced currents. Thewind effect is more marked in shallow areas, particularlybetween the barrier reef and Nouméa. A 30-year
instrumental record of sea surface temperatures indicatethat SST has averaged 23.7 °C at the Vata-Ricaudy site(seasonal mean variability=4.5–7 °C) and 23.4 °C atthe Amédée site (seasonal mean variability=4.5–8 °C).During the same time interval, sea surface salinities havevaried from 33.51 to 36.10 PSU (mean=35.11) at Vata-Ricaudy and from 34.80 to 36.50 PSU (mean=35.74) atthe Amédée site (Delcroix and Lenormand, 1997).
The occurrence of active seasonal upwelling eventsalong the west fore-slopes of New Caledonia wasevidenced from thermo-salinographer records andsatellite infrared images (Hénin and Cresswell, 2005).These upwellings, which are mostly evident in summer,are generated by strong southeasterly surges. Theyoperate within a 10 km wide band, seaward of thewestern outer reef line, extending down to about 100 m;they are more frequent along the southern half part of thewest barrier reef, but can extend to the north of theisland. The upwelled waters are usually 2–3 °C colderthan the surrounding waters. As a result, upwellingevents make the near-surface temperatures less than 23°C. During El Niño (La Niña) events, New Caledoniaexperiences negative (positive) SST anomalies andlower (higher) precipitation (Delcroix and Lenormand,1997).
3. Materials and methods
3.1. Location of samples
Three fossil Porites lobata coral colonies in growthposition, referred to as Colonies 1, 2 and 3, respectively,were selected at different levels of the Holocenesequence, between 0.30 and 4.20 m core depth, in oneof the cores (core NC 36 PV) that have penetrated theouter fringing reef flat at the Vata-Ricaudy site (Fig. 1).Complete details concerning the drilling operation,lithological analysis of the reef core and reconstructionof reef growth development are provided in Cabioch etal. (1995) and Cabioch (2003). These corals were datedby the conventional C-14 method. After calibrationusing the CALIB-4 software programme (Stuiver et al.,1998), the ages, expressed in calendar years, range from6090±90 years BP for Colony 1 to 6330±110 years BPfor Colony 3.
For comparison, two live specimens of Poriteslobata were collected from the fringing reef of Vata-Ricaudy, and from the reef surrounding the Amédéeislet, near the outer barrier reef rim. These sites werechosen to respectively investigate the effects of beingclose to, and far from, the mainland and riverinedischarge and to ascertain the regional reproducibility of

440 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
the Sr and Ba measurements. The Vata-Ricaudy Poritescolony was collected in July 1993, in less than 1 m waterdepth on the outer reef flat at a distance of about 300 moff the beach in Vata Bay. This site is about 20 km fromthe barrier reef and protected from oceanic swells byislets scattered in the lagoon; only the southeast part ofthe fringing reef front is exposed to prevailing winds andrelated strong waves. There are no major rivers withinNoumea. However, the Vata-Ricaudy reef is 15 kmsoutheast of the mouth of the Dumbéa River and 15 kmnorthwest of the mouth of the La Coulée River. Duringthe wet season, the surface currents in the Nouméalagoonal area sweep the turbid plumes from these riversnorthwestwards along the coast. During the relativelydry season, due to a drastic decrease in the riverdischarge, the plumes completely disappear. TheAmédée Porites colony was collected at a water depthof around 2 m on top of the inner part of the reef flat inDecember 1998. This site is 25 km south of Nouméa andabout 1 km from the outer barrier reef line and Boularipass. Given the distance and the direction of prevailingcurrents (Fig. 1), it appears not to be subjected to theinfluence of the river flood plumes penetrating the southlagoon area, but is well flushed by oceanic currentsflowing into the lagoon.
In addition to coral sampling, seawater was collectedonce a month at the Amédée site from April 1996 toMay 1998 and from January 2002 to April 2003. Waterwas taken at 1 m depth, well within the mixed layer.
3.2. Analytical methods
The modern and fossil coral colonies were cut into1-cm-thick slabs, X-rayed to determine growth axisposition, dissepiments orientation and width, andextensively cleaned in an ultrasonic bath prior tosampling, in order to remove organic matter. Onaverage, 12 subsamples per year were taken contin-uously along the main growth axis using a dental burrand a three-axis positioning system. The coral powderwas then dissolved in 2% spiked nitric acid. Ca, Srand Ba concentrations were measured with a VarianUltramass inductively coupled plasma mass spectrom-eter (ICP–MS) following the technique of Le Cornecand Corrège (1997). Reproducibility of the analyses isevaluated from a New Caledonian coral powder(labelled NC20; Corrège et al., 2000) used as astandard. Reproducibility (±2σ) is 0.05 mmol/mol forSr/Ca (0.7 °C) and 0.18 μmol/mol for Ba/Ca.
The seawater samples were filtered and diluted10 times before Sr, Ba and Ca were analyzed by ICP–MS. Standard solutions were also prepared in a 10 times
diluted seawater matrix. Note that analysis of Ca wasperformed only on the water samples collected duringthe years 2002 and 2003.
3.3. Calibration with sea surface temperatures (SST)
The calibration of Sr/Ca with SST designed to beapplied in the present study is based on modern Poritescollected at Amédée reef, the closest and bestdocumented site with in situ temperature recording.The calibration method employed is based on “extremevalues” as described by Cardinal et al. (2001); eachSr/Ca extreme is correlated with the correspondingmonthly mean SST. According to Cardinal et al.(2001), this procedure allows avoidance of approx-imations about intra-seasonal growth rate variationsand reduction of analytical noise by using measure-ments performed at and around the seasonal extremes;however, the precision expected cannot be signifi-cantly better than ±1 °C.
A chronology was established for both modern coralcolonies based on the dates of collection and the natureof the mean annual cycles in temperature-related Sr/Caratios. Time was assigned to the Sr/Ca depth series bypeak matching Sr/Ca minima with instrumentallydetermined SST maxima (and vice versa). The timeaxis is calculated by assuming that the annualtemperature maximum occurs in February and theminimum in August of each year. A visual comparisonbetween the density bands years and geochemical yearsconfirmed the accuracy of the age conversion. Similarly,the chronology of growth history for the three fossilcorals from Vata-Ricaudy reef was estimated on thebasis of the observed periodicities in Sr/Ca variations,coupled with the seasonal skeletal growth bands,assuming the extreme Sr/Ca values measured alongtheir depth profiles correspond to the seasonal extremes.
4. Results
4.1. Trace elements in near-surface waters
Sr concentrations in surface seawater samplescollected at the Amédée site range from 7.8 to 8.8 ppm(standard deviation (S.D.)=0.2 ppm). The valuesof Sr/Ca vary between 8.42 and 9.28 mmol/mol(S.D.=0.2).
Barium concentrations measured in surface seawaterat the Amédée site range from 4.3 to 6.7 ppb ([mean±S.D.] 4.77±0.1 ppb) (Fig. 2), equivalent to 31.46 and48.92 nmol Ba/kg seawater (34.85±0.72 nmol/kg).Higher values of Ba (up to 5.0 ppb) were recorded in
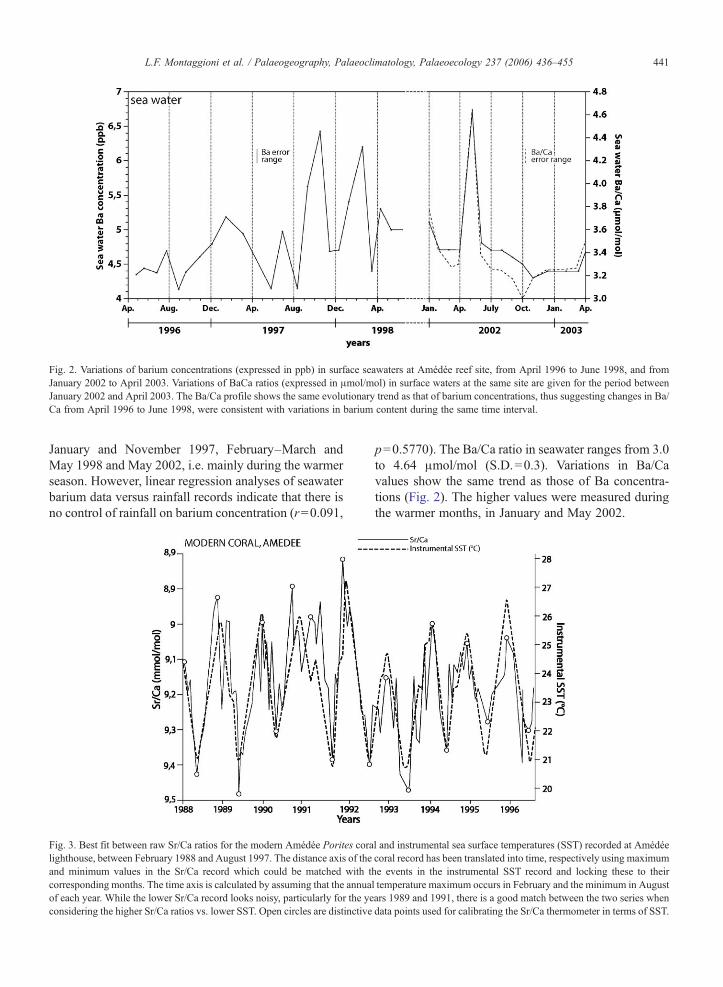
Fig. 2. Variations of barium concentrations (expressed in ppb) in surface seawaters at Amédée reef site, from April 1996 to June 1998, and fromJanuary 2002 to April 2003. Variations of BaCa ratios (expressed in μmol/mol) in surface waters at the same site are given for the period betweenJanuary 2002 and April 2003. The Ba/Ca profile shows the same evolutionary trend as that of barium concentrations, thus suggesting changes in Ba/Ca from April 1996 to June 1998, were consistent with variations in barium content during the same time interval.
441L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
January and November 1997, February–March andMay 1998 and May 2002, i.e. mainly during the warmerseason. However, linear regression analyses of seawaterbarium data versus rainfall records indicate that there isno control of rainfall on barium concentration (r=0.091,
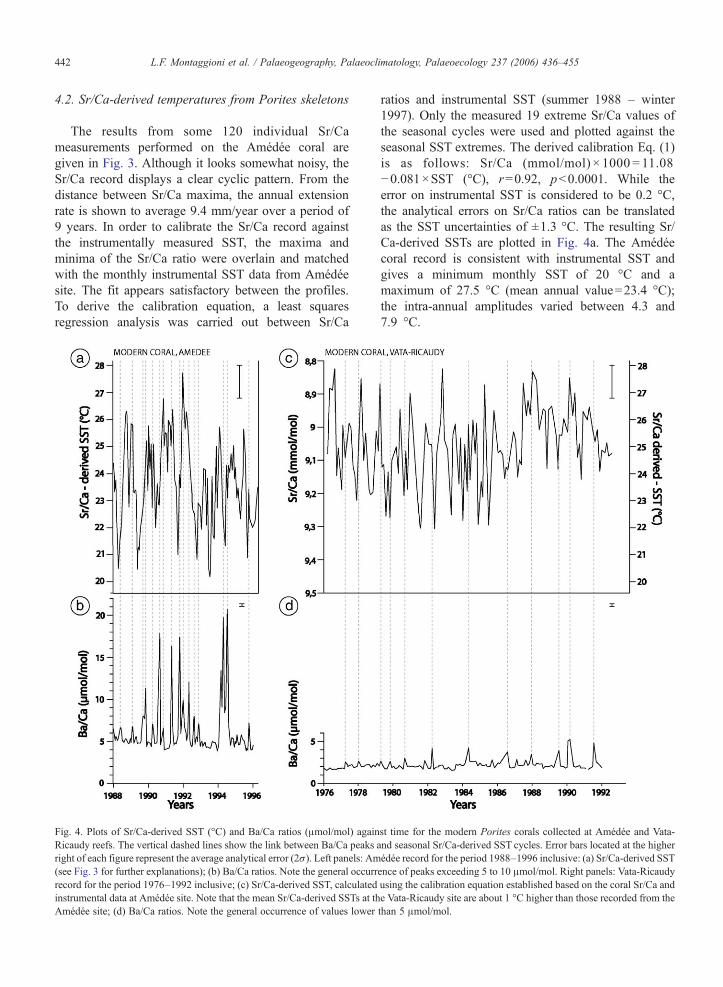
Fig. 3. Best fit between raw Sr/Ca ratios for the modern Amédée Porites coralighthouse, between February 1988 and August 1997. The distance axis of theand minimum values in the Sr/Ca record which could be matched with tcorresponding months. The time axis is calculated by assuming that the annuaof each year. While the lower Sr/Ca record looks noisy, particularly for the yeconsidering the higher Sr/Ca ratios vs. lower SST. Open circles are distinctive
p=0.5770). The Ba/Ca ratio in seawater ranges from 3.0to 4.64 μmol/mol (S.D.=0.3). Variations in Ba/Cavalues show the same trend as those of Ba concentra-tions (Fig. 2). The higher values were measured duringthe warmer months, in January and May 2002.
l and instrumental sea surface temperatures (SST) recorded at Amédéecoral record has been translated into time, respectively using maximumhe events in the instrumental SST record and locking these to theirl temperature maximum occurs in February and the minimum in Augustars 1989 and 1991, there is a good match between the two series whendata points used for calibrating the Sr/Ca thermometer in terms of SST.
442 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
4.2. Sr/Ca-derived temperatures from Porites skeletons
The results from some 120 individual Sr/Cameasurements performed on the Amédée coral aregiven in Fig. 3. Although it looks somewhat noisy, theSr/Ca record displays a clear cyclic pattern. From thedistance between Sr/Ca maxima, the annual extensionrate is shown to average 9.4 mm/year over a period of9 years. In order to calibrate the Sr/Ca record againstthe instrumentally measured SST, the maxima andminima of the Sr/Ca ratio were overlain and matchedwith the monthly instrumental SST data from Amédéesite. The fit appears satisfactory between the profiles.To derive the calibration equation, a least squaresregression analysis was carried out between Sr/Ca
Fig. 4. Plots of Sr/Ca-derived SST (°C) and Ba/Ca ratios (μmol/mol) againRicaudy reefs. The vertical dashed lines show the link between Ba/Ca peaksright of each figure represent the average analytical error (2σ). Left panels: Am(see Fig. 3 for further explanations); (b) Ba/Ca ratios. Note the general occurrrecord for the period 1976–1992 inclusive; (c) Sr/Ca-derived SST, calculatedinstrumental data at Amédée site. Note that the mean Sr/Ca-derived SSTs at thAmédée site; (d) Ba/Ca ratios. Note the general occurrence of values lower
ratios and instrumental SST (summer 1988 – winter1997). Only the measured 19 extreme Sr/Ca values ofthe seasonal cycles were used and plotted against theseasonal SST extremes. The derived calibration Eq. (1)is as follows: Sr/Ca (mmol/mol) × 1000 = 11.08−0.081×SST (°C), r=0.92, pb0.0001. While theerror on instrumental SST is considered to be 0.2 °C,the analytical errors on Sr/Ca ratios can be translatedas the SST uncertainties of ±1.3 °C. The resulting Sr/Ca-derived SSTs are plotted in Fig. 4a. The Amédéecoral record is consistent with instrumental SST andgives a minimum monthly SST of 20 °C and amaximum of 27.5 °C (mean annual value=23.4 °C);the intra-annual amplitudes varied between 4.3 and7.9 °C.
st time for the modern Porites corals collected at Amédée and Vata-and seasonal Sr/Ca-derived SST cycles. Error bars located at the higherédée record for the period 1988–1996 inclusive: (a) Sr/Ca-derived SSTence of peaks exceeding 5 to 10 μmol/mol. Right panels: Vata-Ricaudyusing the calibration equation established based on the coral Sr/Ca ande Vata-Ricaudy site are about 1 °C higher than those recorded from thethan 5 μmol/mol.
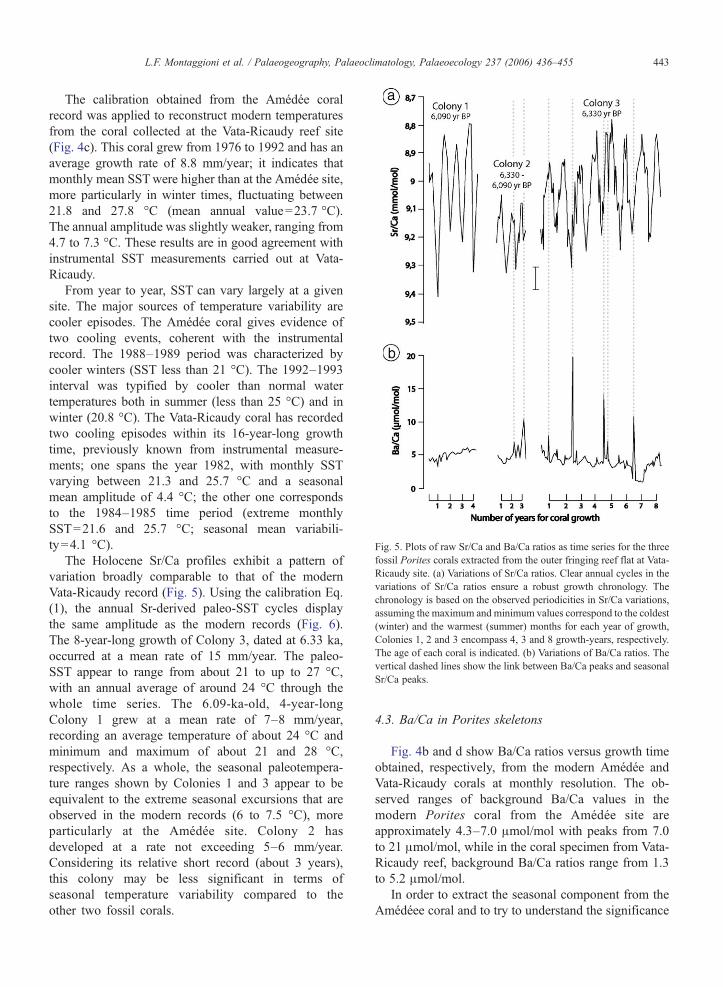
Fig. 5. Plots of raw Sr/Ca and Ba/Ca ratios as time series for the threefossil Porites corals extracted from the outer fringing reef flat at Vata-Ricaudy site. (a) Variations of Sr/Ca ratios. Clear annual cycles in thevariations of Sr/Ca ratios ensure a robust growth chronology. Thechronology is based on the observed periodicities in Sr/Ca variations,assuming the maximum and minimum values correspond to the coldest(winter) and the warmest (summer) months for each year of growth,Colonies 1, 2 and 3 encompass 4, 3 and 8 growth-years, respectively.The age of each coral is indicated. (b) Variations of Ba/Ca ratios. Thevertical dashed lines show the link between Ba/Ca peaks and seasonalSr/Ca peaks.
443L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
The calibration obtained from the Amédée coralrecord was applied to reconstruct modern temperaturesfrom the coral collected at the Vata-Ricaudy reef site(Fig. 4c). This coral grew from 1976 to 1992 and has anaverage growth rate of 8.8 mm/year; it indicates thatmonthly mean SSTwere higher than at the Amédée site,more particularly in winter times, fluctuating between21.8 and 27.8 °C (mean annual value=23.7 °C).The annual amplitude was slightly weaker, ranging from4.7 to 7.3 °C. These results are in good agreement withinstrumental SST measurements carried out at Vata-Ricaudy.
From year to year, SST can vary largely at a givensite. The major sources of temperature variability arecooler episodes. The Amédée coral gives evidence oftwo cooling events, coherent with the instrumentalrecord. The 1988–1989 period was characterized bycooler winters (SST less than 21 °C). The 1992–1993interval was typified by cooler than normal watertemperatures both in summer (less than 25 °C) and inwinter (20.8 °C). The Vata-Ricaudy coral has recordedtwo cooling episodes within its 16-year-long growthtime, previously known from instrumental measure-ments; one spans the year 1982, with monthly SSTvarying between 21.3 and 25.7 °C and a seasonalmean amplitude of 4.4 °C; the other one correspondsto the 1984–1985 time period (extreme monthlySST=21.6 and 25.7 °C; seasonal mean variabili-ty=4.1 °C).
The Holocene Sr/Ca profiles exhibit a pattern ofvariation broadly comparable to that of the modernVata-Ricaudy record (Fig. 5). Using the calibration Eq.(1), the annual Sr-derived paleo-SST cycles displaythe same amplitude as the modern records (Fig. 6).The 8-year-long growth of Colony 3, dated at 6.33 ka,occurred at a mean rate of 15 mm/year. The paleo-SST appear to range from about 21 to up to 27 °C,with an annual average of around 24 °C through thewhole time series. The 6.09-ka-old, 4-year-longColony 1 grew at a mean rate of 7–8 mm/year,recording an average temperature of about 24 °C andminimum and maximum of about 21 and 28 °C,respectively. As a whole, the seasonal paleotempera-ture ranges shown by Colonies 1 and 3 appear to beequivalent to the extreme seasonal excursions that areobserved in the modern records (6 to 7.5 °C), moreparticularly at the Amédée site. Colony 2 hasdeveloped at a rate not exceeding 5–6 mm/year.Considering its relative short record (about 3 years),this colony may be less significant in terms ofseasonal temperature variability compared to theother two fossil corals.
4.3. Ba/Ca in Porites skeletons
Fig. 4b and d show Ba/Ca ratios versus growth timeobtained, respectively, from the modern Amédée andVata-Ricaudy corals at monthly resolution. The ob-served ranges of background Ba/Ca values in themodern Porites coral from the Amédée site areapproximately 4.3–7.0 μmol/mol with peaks from 7.0to 21 μmol/mol, while in the coral specimen from Vata-Ricaudy reef, background Ba/Ca ratios range from 1.3to 5.2 μmol/mol.
In order to extract the seasonal component from theAmédéee coral and to try to understand the significance
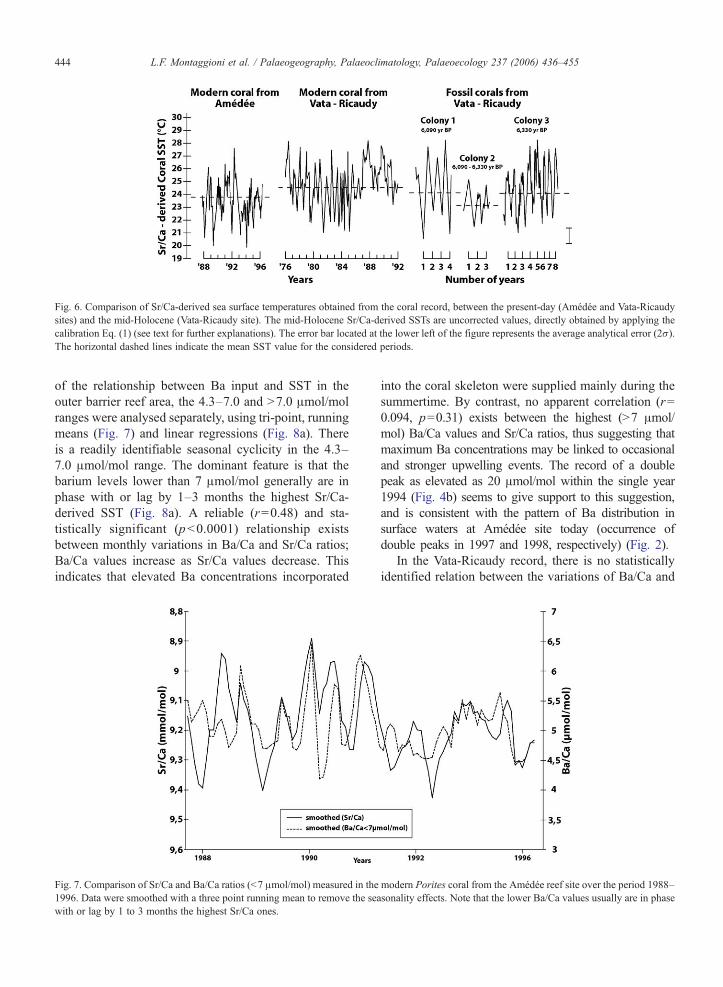
Fig. 6. Comparison of Sr/Ca-derived sea surface temperatures obtained from the coral record, between the present-day (Amédée and Vata-Ricaudysites) and the mid-Holocene (Vata-Ricaudy site). The mid-Holocene Sr/Ca-derived SSTs are uncorrected values, directly obtained by applying thecalibration Eq. (1) (see text for further explanations). The error bar located at the lower left of the figure represents the average analytical error (2σ).The horizontal dashed lines indicate the mean SST value for the considered periods.
444 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
of the relationship between Ba input and SST in theouter barrier reef area, the 4.3–7.0 and N7.0 μmol/molranges were analysed separately, using tri-point, runningmeans (Fig. 7) and linear regressions (Fig. 8a). Thereis a readily identifiable seasonal cyclicity in the 4.3–7.0 μmol/mol range. The dominant feature is that thebarium levels lower than 7 μmol/mol generally are inphase with or lag by 1–3 months the highest Sr/Ca-derived SST (Fig. 8a). A reliable (r=0.48) and sta-tistically significant (pb0.0001) relationship existsbetween monthly variations in Ba/Ca and Sr/Ca ratios;Ba/Ca values increase as Sr/Ca values decrease. Thisindicates that elevated Ba concentrations incorporated
Fig. 7. Comparison of Sr/Ca and Ba/Ca ratios (b7 μmol/mol) measured in the1996. Data were smoothed with a three point running mean to remove the sewith or lag by 1 to 3 months the highest Sr/Ca ones.
into the coral skeleton were supplied mainly during thesummertime. By contrast, no apparent correlation (r=0.094, p=0.31) exists between the highest (N7 μmol/mol) Ba/Ca values and Sr/Ca ratios, thus suggesting thatmaximum Ba concentrations may be linked to occasionaland stronger upwelling events. The record of a doublepeak as elevated as 20 μmol/mol within the single year1994 (Fig. 4b) seems to give support to this suggestion,and is consistent with the pattern of Ba distribution insurface waters at Amédée site today (occurrence ofdouble peaks in 1997 and 1998, respectively) (Fig. 2).
In the Vata-Ricaudy record, there is no statisticallyidentified relation between the variations of Ba/Ca and
modern Porites coral from the Amédée reef site over the period 1988–asonality effects. Note that the lower Ba/Ca values usually are in phase
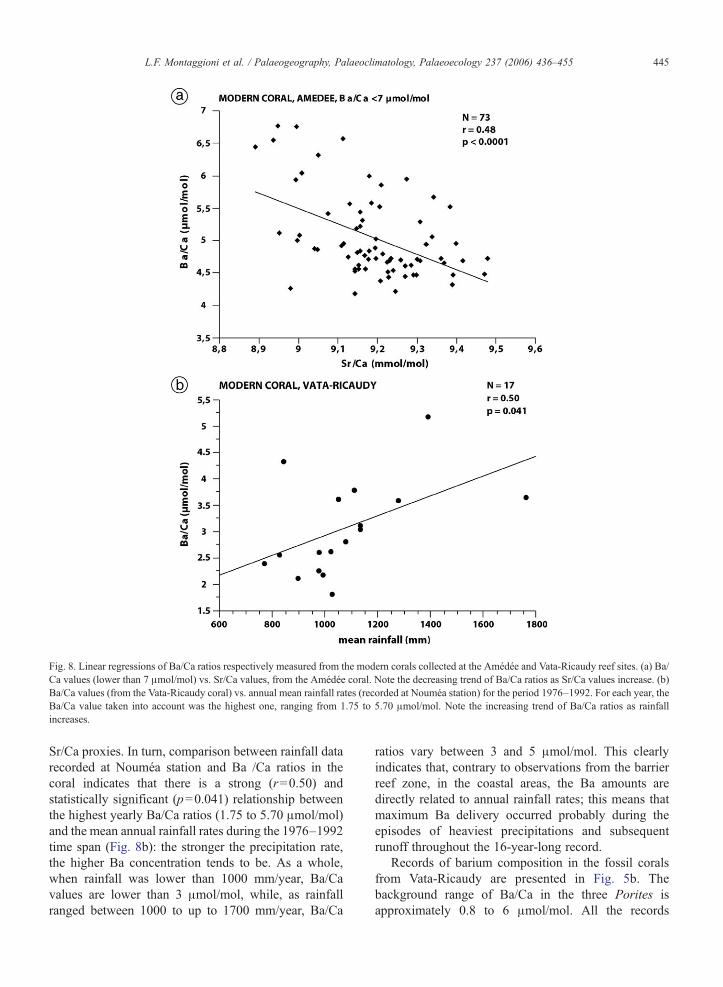
Fig. 8. Linear regressions of Ba/Ca ratios respectively measured from the modern corals collected at the Amédée and Vata-Ricaudy reef sites. (a) Ba/Ca values (lower than 7 μmol/mol) vs. Sr/Ca values, from the Amédée coral. Note the decreasing trend of Ba/Ca ratios as Sr/Ca values increase. (b)Ba/Ca values (from the Vata-Ricaudy coral) vs. annual mean rainfall rates (recorded at Nouméa station) for the period 1976–1992. For each year, theBa/Ca value taken into account was the highest one, ranging from 1.75 to 5.70 μmol/mol. Note the increasing trend of Ba/Ca ratios as rainfallincreases.
445L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
Sr/Ca proxies. In turn, comparison between rainfall datarecorded at Nouméa station and Ba /Ca ratios in thecoral indicates that there is a strong (r=0.50) andstatistically significant (p=0.041) relationship betweenthe highest yearly Ba/Ca ratios (1.75 to 5.70 μmol/mol)and the mean annual rainfall rates during the 1976–1992time span (Fig. 8b): the stronger the precipitation rate,the higher Ba concentration tends to be. As a whole,when rainfall was lower than 1000 mm/year, Ba/Cavalues are lower than 3 μmol/mol, while, as rainfallranged between 1000 to up to 1700 mm/year, Ba/Ca
ratios vary between 3 and 5 μmol/mol. This clearlyindicates that, contrary to observations from the barrierreef zone, in the coastal areas, the Ba amounts aredirectly related to annual rainfall rates; this means thatmaximum Ba delivery occurred probably during theepisodes of heaviest precipitations and subsequentrunoff throughout the 16-year-long record.
Records of barium composition in the fossil coralsfrom Vata-Ricaudy are presented in Fig. 5b. Thebackground range of Ba/Ca in the three Porites isapproximately 0.8 to 6 μmol/mol. All the records
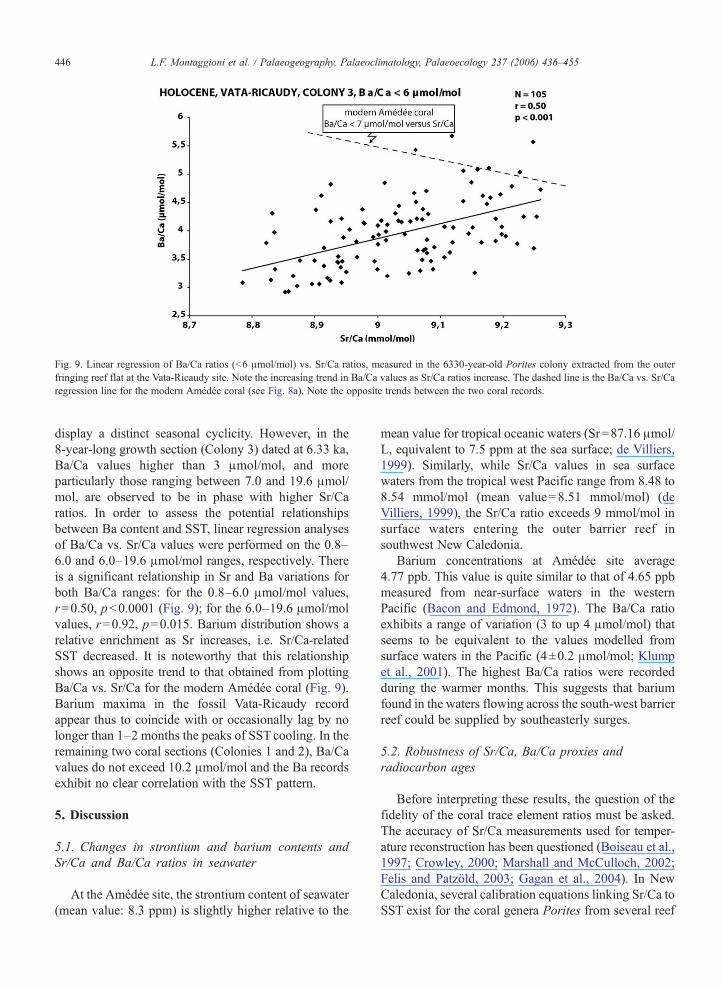
Fig. 9. Linear regression of Ba/Ca ratios (b6 μmol/mol) vs. Sr/Ca ratios, measured in the 6330-year-old Porites colony extracted from the outerfringing reef flat at the Vata-Ricaudy site. Note the increasing trend in Ba/Ca values as Sr/Ca ratios increase. The dashed line is the Ba/Ca vs. Sr/Caregression line for the modern Amédée coral (see Fig. 8a). Note the opposite trends between the two coral records.
446 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
display a distinct seasonal cyclicity. However, in the8-year-long growth section (Colony 3) dated at 6.33 ka,Ba/Ca values higher than 3 μmol/mol, and moreparticularly those ranging between 7.0 and 19.6 μmol/mol, are observed to be in phase with higher Sr/Caratios. In order to assess the potential relationshipsbetween Ba content and SST, linear regression analysesof Ba/Ca vs. Sr/Ca values were performed on the 0.8–6.0 and 6.0–19.6 μmol/mol ranges, respectively. Thereis a significant relationship in Sr and Ba variations forboth Ba/Ca ranges: for the 0.8–6.0 μmol/mol values,r=0.50, pb0.0001 (Fig. 9); for the 6.0–19.6 μmol/molvalues, r=0.92, p=0.015. Barium distribution shows arelative enrichment as Sr increases, i.e. Sr/Ca-relatedSST decreased. It is noteworthy that this relationshipshows an opposite trend to that obtained from plottingBa/Ca vs. Sr/Ca for the modern Amédée coral (Fig. 9).Barium maxima in the fossil Vata-Ricaudy recordappear thus to coincide with or occasionally lag by nolonger than 1–2 months the peaks of SST cooling. In theremaining two coral sections (Colonies 1 and 2), Ba/Cavalues do not exceed 10.2 μmol/mol and the Ba recordsexhibit no clear correlation with the SST pattern.
5. Discussion
5.1. Changes in strontium and barium contents andSr/Ca and Ba/Ca ratios in seawater
At the Amédée site, the strontium content of seawater(mean value: 8.3 ppm) is slightly higher relative to the
mean value for tropical oceanic waters (Sr=87.16 μmol/L, equivalent to 7.5 ppm at the sea surface; de Villiers,1999). Similarly, while Sr/Ca values in sea surfacewaters from the tropical west Pacific range from 8.48 to8.54 mmol/mol (mean value=8.51 mmol/mol) (deVilliers, 1999), the Sr/Ca ratio exceeds 9 mmol/mol insurface waters entering the outer barrier reef insouthwest New Caledonia.
Barium concentrations at Amédée site average4.77 ppb. This value is quite similar to that of 4.65 ppbmeasured from near-surface waters in the westernPacific (Bacon and Edmond, 1972). The Ba/Ca ratioexhibits a range of variation (3 to up 4 μmol/mol) thatseems to be equivalent to the values modelled fromsurface waters in the Pacific (4±0.2 μmol/mol; Klumpet al., 2001). The highest Ba/Ca ratios were recordedduring the warmer months. This suggests that bariumfound in the waters flowing across the south-west barrierreef could be supplied by southeasterly surges.
5.2. Robustness of Sr/Ca, Ba/Ca proxies andradiocarbon ages
Before interpreting these results, the question of thefidelity of the coral trace element ratios must be asked.The accuracy of Sr/Ca measurements used for temper-ature reconstruction has been questioned (Boiseau et al.,1997; Crowley, 2000; Marshall and McCulloch, 2002;Felis and Patzöld, 2003; Gagan et al., 2004). In NewCaledonia, several calibration equations linking Sr/Ca toSST exist for the coral genera Porites from several reef

447L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
sites (Beck et al., 1992; Castellaro, 1999; Corrège et al.,2000; Quinn and Sampson, 2002). The coral Sr/Ca–SSTrelationship obtained in the present study provided aslope value of 0.081±0.008 that is close to slope valuesaveraging 0.0597±0.005 to 0.0795±0.010 reportedfrom previous studies conducted from Porites coralsin New Caledonia and in other tropical areas, usingdifferent methods (de Villiers et al., 1994; Heiss et al.,1996; McCulloch et al., 1996; Gagan et al., 2000; Wei etal., 2000; Marshall and McCulloch, 2002; Felis et al.,2004). Potential errors in Sr/Ca-reconstructed SST maybe generated by the presence of secondary (abiogenic)aragonite needles deposited within the skeletal cavityspaces during early marine diagenesis. Such analteration of coral skeletons can result in significantchanges in the initial chemical composition of aragoniteand elemental ratios (Enmar et al., 2000; Ribaud-Laurenti et al., 2001; Müller et al., 2001; Cohen andHart, 2004; Gagan et al., 2004). The abiotic aragonite istypified by a significantly higher Sr/Ca ratio than theaverage ratio in primary aragonite. As a result, Sr/Ca-derived paleo-SST can be shifted by 2 to 5 °C towardcooler temperature, compared to those actually experi-enced by the living corals. In order to detect the effect ofearly marine diagenesis on strontium abundance in theNew Caledonian coral skeletons under study, Castellaro(1999) evaluated the quantity of secondary aragonitepresent within both modern Porites collected at theAmédée reef site and fossil Colonies 1 to 3. All of thecoral samples were analysed petrographically. The massof secondary aragonite was determined using mercuryporosimetry and measurements of biogenic/secondaryaragonite ratios from SEM images. On the basis ofseveral diagnostic features (e.g. calcification centres,dissepiments), the original ultrastructures of the moderncorals appeared to be well preserved, and no secondaryinfilling was observed within skeletal cavities. Bycontrast, the amount of precipitated secondary aragoniteinto the fossil corals averages 8.8%, 10.2% and 11.8%of the total pore volume in Colonies 3, 2 and 1,respectively. This results mainly in a deviation of Sr/Caratios (by 0.07±0.03 μmol/mol) toward higher values.Applying the calibration Eq. (1) to the diageneticallyaltered Sr/Ca ratios lowers the apparent SSTs by 0.50 to1.20 °C. Another potential source of error in evaluatingcoral Sr/Ca-derived SST is related to reef coring.Penetrating all of the three fossil Porites colonies in avertical growth direction was highly unlikely. Since fora given massive colony Sr/Ca ratios measured from theshaded sides may be significantly higher than thoseobtained from the principal growth axis (de Villiers etal., 1994; Gagan et al., 2000), the Sr/Ca record for fossil
corals may tend to yield colder artefacts. Therefore, theSr/Ca-derived SSTs obtained from the fossil coralsprobably underestimate the real mid-Holocene SST byabout 1 °C.
In their recent analysis of coral skeleton geochem-istry as a tool for reconstructing sea surface conditions,based on a modern Porites sampled from offshoreAmédée islet, Quinn and Sampson (2002) noted that theBa/Ca records in New Caledonia display large excur-sions. The authors pointed out that the Ba/Ca valueswould not be easily interpreted in terms of environmen-tal variability, since extensive cleaning of organic matterwas not conducted on the Porites colony they studied.Cleaning procedures appear to be of prime importancefor eliminating pre-existing surface or occluded organicmatter or detrital contamination from coral specimens,since, as emphasized by Tudhope et al. (1996) andAnderegg et al. (1997), contamination can cause bariumlevels to greatly increase. Elevated levels of Ba/Ca mayreflect the incorporation of barium into the skeleton insome non-lattice bound phase (i.e. organic matter orpolyp tissues) (Hart et al., 1996; Tudhope et al., 1996),that is liable to decay with time (Tudhope et al., 1996). Iftrue, the higher amounts of Ba/Ca in the fossil NewCaledonian corals cannot be attributed to Ba retentionwithin organic compounds, due to the rapid decay oforganic matter through fossilization; they probably arestrictly contained within aragonite crystals. Given thesimilarity of Ba/Ca concentrations in the fossil coralsfrom Vata-Ricaudy and in the modern one fromAmédée, one can assume that, in the latter, barium istrapped mostly in the carbonate lattice. Uniformly lowconcentrations of barium (less than 2 to 5 μmol/mol),comparable to those measured in the modern Poritesfrom Vata-Ricaudy, are common in the modern coralrecord and are regarded as “noise background levels”(Tudhope et al., 1996, 1997; Anderegg et al., 1997;Fallon et al., 1999; Quinn and Sampson, 2002; Alibert etal., 2003; Reuer et al., 2003). The Ba/Ca levels in themodern Vata-Ricaudy coral must therefore be mostlyattributed to a lattice-bound rather than organicdeposition.
An additional potential error in estimating the Ba/Cavalues from the fossil corals may be related to the factthat these corals contain secondary aragonite. Given thatbarium seems to dominantly be lattice-bound, earlydiagenetic process may have caused a change in the bulkBa/Ca ratio within fossil corals. The mode of incorpo-ration of Ba into the coral aragonite is still debated (seeSinclair, 2005) and the possible effect of inorganicaragonite precipitation on Ba concentration is, to ourknowledge, not documented. However, on the basis of

448 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
previous studies on the behaviour of lattice-bound traceelements (i.e. strontium, uranium) during early diagen-esis, it could be suggested that the barium concentrationin secondary aragonite filling the skeketal cavities ishigher than in biogenic aragonite. For instance, thestrontium content in inorganic aragonite deposited in theHolocene corals from Vata-Ricaudy reef and from Eilat(Red Sea), respectively, is about 9% and 11% higherthan that of biogenic aragonite (Castellaro, 1999; Enmaret al., 2000). By contrast, Holocene corals can exhibituranium content in inorganic aragonite 60% higher thanin biogenic aragonite (Lazar et al., 2004). If such aconcentration difference between the two aragonitetypes is applied to barium distribution in the fossil Vata-Ricaudy corals, the bulk barium concentration could notbe higher than 6%, compared to that of the moderncoral, since the amount of secondary aragonite averages10%. Assuming the maximum Ba/Ca value in coralsfrom Vata-Ricaudy site is about 5 μmol/mol, theincrease in the Ba/Ca ratios within the fossil coralscould not exceed about 0.3 μmol/mol. Accordingly, anypotential enrichment in Ba content linked to marinediagenesis would not explain the general occurrence ofhigh Ba/Ca peaks in the Holocene record. Furthermore,growth rates of individual corals control the amount ofbarium incorporated, through differences in kineticelement fractionation (Pingitore et al., 1989; Reuer etal., 2003). On one hand, although the growth rates of thetwo modern corals from Vata-Ricaudy and Amédée sitesare of equal magnitude (8.8 to 9.4 mm/year), the Ba/Canoise levels are quite different (2 compared to 5 μmol/mol). On the other hand, while the extension rates of thefossil corals have varied largely, from 5 to 15 mm/year,their background levels are quite similar, averagingaround 3.5 μmol/mol. These figures are compatible withthe idea that variations in Ba/Ca ratios do not dependupon the kinetic partitioning of barium into aragonite,but actually express strong environmental controls onbarium uptake. Furthermore, Reuer et al. (2003)addressed the possible influence of anthropogenicelemental artefacts on coastal sedimentation. In south-west New Caledonia, the studies by Launay (1972) andAmbatsian et al. (1997) showed that no lagoonal deposithas apparently undergone significant in situ metalenrichment that could be related to extensive humanactivity, such as mining.
The main implicit assumption in deriving C-14 datesfrom Holocene coral skeletons is that the system hasremained closed over time (Yokoyama and Esat, 2004).However, early marine diagenesis of corals may be apossible cause of alteration in radiocarbon ages. Theprecipitation of secondary aragonite results from the
circulation of fluids saturated with respect to CaCO3
through the pores of original skeletons. These fluidsmay have a C-14 signature that is affected by thereservoir age in particular in reef areas, where surfacewaters are fed by upwelled deeper waters (Müller et al.,2004). Petrographical studies of Porites and othermassive coral colonies indicate that secondary infillingis a rapid process that can occur obviously within thelifetime of the corals (Spiro, 1971; Gvirtzman andFriedman, 1977; Enmar et al., 2000). Concerning theHolocene corals from Vata-Ricaudy, it is not possible toassume with any certainty that inorganic aragonite hasdeposited close in age to the primary skeleton; thedeposition may have operated over a period of hundredsor thousands of years.
The accuracy of the radiocarbon ages will depend onthe quantity and on the time of deposition of inorganicaragonite. Cohen and Hart (2004) analysed the effect ofmarine diagenesis on the radiocarbon dating of an about13,000-year-old Porites coral that had been coredthrough the outer barrier reef at Tahiti. Comparing theages obtained, respectively, from pristine and heavilyinfilled (containing up to 50% secondary aragonite)parts of the same coral chunk, Cohen and Hart (2004)indicated that the difference (about 100 years) was notstatistically significant, given the error range on the ages(±70 and ±65 years, respectively) and other sources oferrors such as short term variability of the reservoir age.We therefore consider that, whatever time it took forsecondary aragonite to precipitate within the fossilcorals from Vata-Ricaudy reef, the potential agedistortion is probably included within the measurementsince the percentage of aragonite overgrowths does notexceed 10% of total skeletal volume and the error on theages ranges between 90 and 100 years. Moreover,during the preparation of the coral samples to beradiocarbon-dated, most of the secondary aragonite hasbeen routinely removed due to mechanical cleaning.This reinforces the assumption on the reliability of theages obtained.
5.3. Mid-Holocene warming
Comparison of the modern and fossil Sr/Ca-derivedSST records from southwest New Caledonia empha-sized the fact that, during the mid-Holocene, thesummers probably were about 1 °C warmer than thepresent ones, while the seasonal variability seems tohave been comparable, at least episodically (Fig. 6).These results are is in good agreement with previousdata from other tropical west Pacific areas; on theAustralian Great Barrier Reef (Gagan et al., 1998;
449L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
Gagan et al., 2002) and in Vanuatu (Gagan et al., 2000),SST culminated with an optimum 1 °C above modernvalues at about 6.0 ka. This climatic warming, that isoften referred to as the Holocene Climatic Optimum, isalso recorded in many tropical parts of the world (Rimbuet al., 2004). However, SST captured by Colony 2 mayreflect an apparent 2°C cooling. The shortness of therecord precludes any interpretation on its cause. Inparticular, it is not possible to specify whether thecooling is linked to interannual and/or interdecadalclimate variations or caused by a volcanic eruption.Alternatively, the sample from Colony 2 could comefrom the shaded side of the coral, which has been shownto have anomalously high Sr/Ca (and hence cooler SST;see de Villiers et al., 1994). Whatever the significance ofthe higher Sr/Ca values from Colony 2, and based on themean paleo-SST obtained, it is obvious that, along thesouthwest coast of New Caledonia, the retardation inreef initiation cannot be ascribed directly to possibleabrupt cooling episodes during the mid-Holoceneclimatic optimum, contrary to the reef demise event at7.0 ka reported from the north South China Sea (Yu etal., 2004).
5.4. Sources of barium
Measurements of barium concentrations and Ba/Caratios in seawater from the barrier reef zone at Amédéestation, indicate that Ba is supplied to the reef mainly inthe summer time, and, thus, strongly suggest that Baderives from upwelling induced by the south-easterlies.By contrast, on the modern fringing reef at Vata-Ricaudy, the barium distribution in corals directly
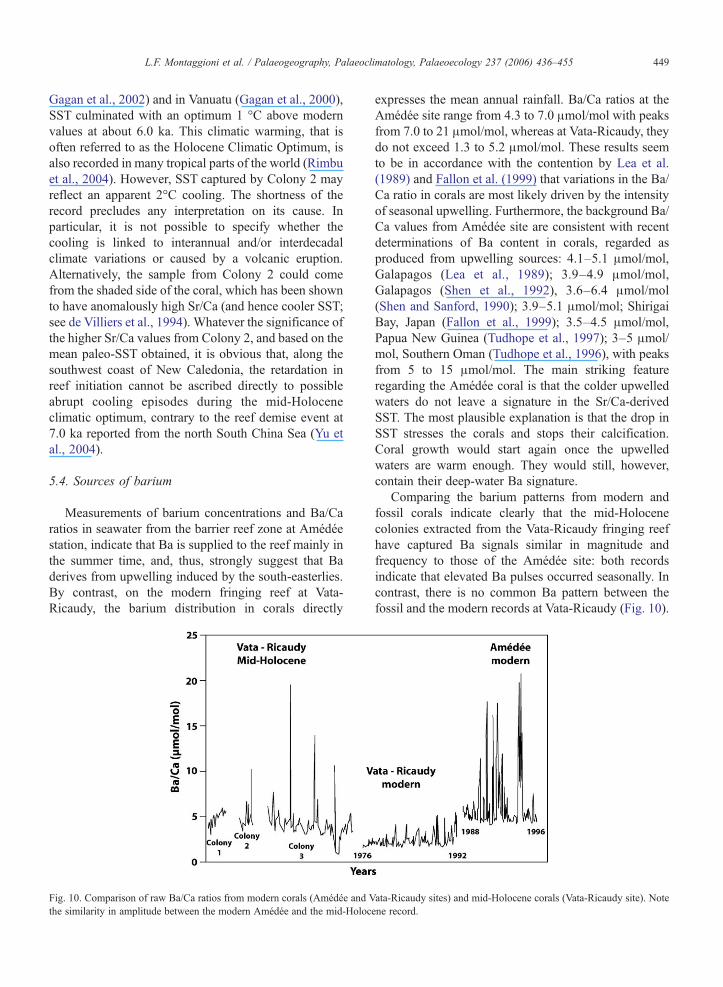
Fig. 10. Comparison of raw Ba/Ca ratios from modern corals (Amédée and Vthe similarity in amplitude between the modern Amédée and the mid-Holoc
expresses the mean annual rainfall. Ba/Ca ratios at theAmédée site range from 4.3 to 7.0 μmol/mol with peaksfrom 7.0 to 21 μmol/mol, whereas at Vata-Ricaudy, theydo not exceed 1.3 to 5.2 μmol/mol. These results seemto be in accordance with the contention by Lea et al.(1989) and Fallon et al. (1999) that variations in the Ba/Ca ratio in corals are most likely driven by the intensityof seasonal upwelling. Furthermore, the background Ba/Ca values from Amédée site are consistent with recentdeterminations of Ba content in corals, regarded asproduced from upwelling sources: 4.1–5.1 μmol/mol,Galapagos (Lea et al., 1989); 3.9–4.9 μmol/mol,Galapagos (Shen et al., 1992), 3.6–6.4 μmol/mol(Shen and Sanford, 1990); 3.9–5.1 μmol/mol; ShirigaiBay, Japan (Fallon et al., 1999); 3.5–4.5 μmol/mol,Papua New Guinea (Tudhope et al., 1997); 3–5 μmol/mol, Southern Oman (Tudhope et al., 1996), with peaksfrom 5 to 15 μmol/mol. The main striking featureregarding the Amédée coral is that the colder upwelledwaters do not leave a signature in the Sr/Ca-derivedSST. The most plausible explanation is that the drop inSST stresses the corals and stops their calcification.Coral growth would start again once the upwelledwaters are warm enough. They would still, however,contain their deep-water Ba signature.
Comparing the barium patterns from modern andfossil corals indicate clearly that the mid-Holocenecolonies extracted from the Vata-Ricaudy fringing reefhave captured Ba signals similar in magnitude andfrequency to those of the Amédée site: both recordsindicate that elevated Ba pulses occurred seasonally. Incontrast, there is no common Ba pattern between thefossil and the modern records at Vata-Ricaudy (Fig. 10).
ata-Ricaudy sites) and mid-Holocene corals (Vata-Ricaudy site). Noteene record.
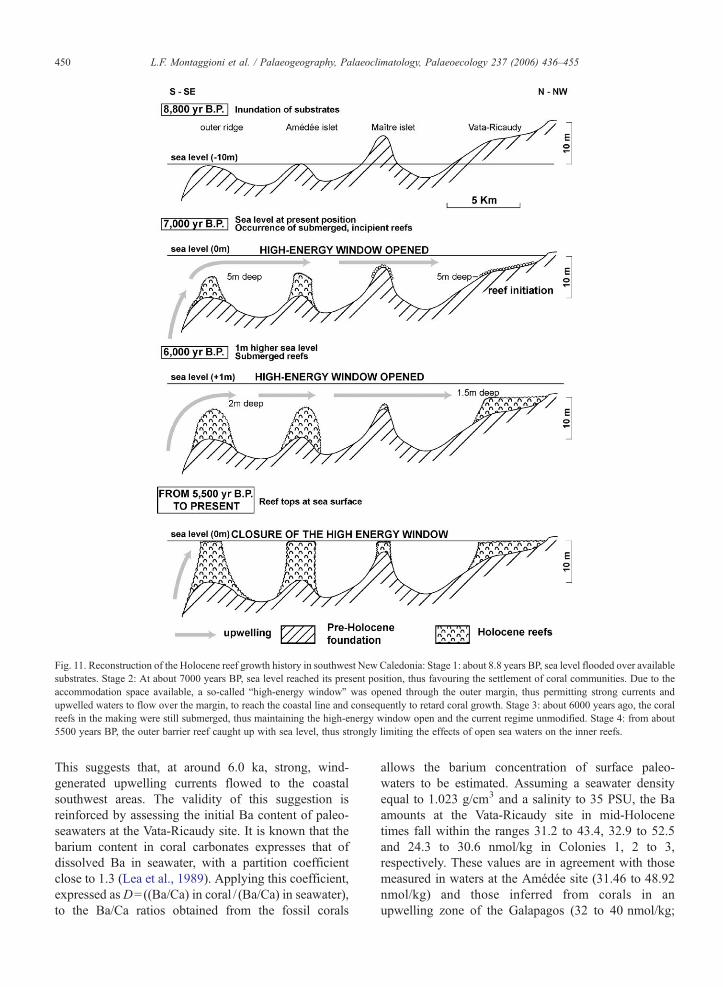
Fig. 11. Reconstruction of the Holocene reef growth history in southwest New Caledonia: Stage 1: about 8.8 years BP, sea level flooded over availablesubstrates. Stage 2: At about 7000 years BP, sea level reached its present position, thus favouring the settlement of coral communities. Due to theaccommodation space available, a so-called “high-energy window” was opened through the outer margin, thus permitting strong currents andupwelled waters to flow over the margin, to reach the coastal line and consequently to retard coral growth. Stage 3: about 6000 years ago, the coralreefs in the making were still submerged, thus maintaining the high-energy window open and the current regime unmodified. Stage 4: from about5500 years BP, the outer barrier reef caught up with sea level, thus strongly limiting the effects of open sea waters on the inner reefs.
450 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
This suggests that, at around 6.0 ka, strong, wind-generated upwelling currents flowed to the coastalsouthwest areas. The validity of this suggestion isreinforced by assessing the initial Ba content of paleo-seawaters at the Vata-Ricaudy site. It is known that thebarium content in coral carbonates expresses that ofdissolved Ba in seawater, with a partition coefficientclose to 1.3 (Lea et al., 1989). Applying this coefficient,expressed asD=((Ba/Ca) in coral / (Ba/Ca) in seawater),to the Ba/Ca ratios obtained from the fossil corals
allows the barium concentration of surface paleo-waters to be estimated. Assuming a seawater densityequal to 1.023 g/cm3 and a salinity to 35 PSU, the Baamounts at the Vata-Ricaudy site in mid-Holocenetimes fall within the ranges 31.2 to 43.4, 32.9 to 52.5and 24.3 to 30.6 nmol/kg in Colonies 1, 2 to 3,respectively. These values are in agreement with thosemeasured in waters at the Amédée site (31.46 to 48.92nmol/kg) and those inferred from corals in anupwelling zone of the Galapagos (32 to 40 nmol/kg;

451L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
Lea et al., 1989; Shen et al., 1992). However, there is asignificant offset in the time of occurrence of Ba pulsesrecorded in the fossil corals at Vata-Ricaudy site and inthe modern coral at the Amédée site. Whereas, in theouter barrier reef record, the observed Ba enrichmenthappens during the normal warm season, in the mid-Holocene corals, elevated barium content appear to havebeen coupled with the colder season or, at least, withanomalously cool seasonal flows. The source of bariumincorporated at the Vata-Ricaudy site during the phase ofreef accretion cannot be linked to river runoff or rainfall,since the potential Ba delivery operates mainly duringthe summer rainy season. The best explanation for suchinterannual variations in barium content therefore seemsto be the upwelling hypothesis. If correct, this impliesthat winter conditions at 6 ka BP in New Caledonia weresimilar to those encountered today during El Niñoevents, when stronger southearly winds cool surfacewaters (Delcroix and Lenormand, 1997) and have thepotential to generate coastal upwellings.
5.5. Reconstruction of the Holocene reef growth historyin southwest New Caledonia
It is notable that both Holocene barrier and fringingreefs from southwest New Caledonia have developedfollowing a catching-up mode (i.e. reef growth was ableto catch up with sea level after its stabilization;Neumann and Macintyre, 1985; Davies and Montag-gioini, 1985). The barrier reef began to develop atapproximately 8.2 ka from substrates at present situatedat about 10 m deep, and caught up with sea level notprior to about 5 ka (Cabioch et al., 1999; Cabioch,2003). By contrast, the adjacent fringing reefs started togrow at 7–6.5 ka from substrates located today at adepth of about 5 m, i.e. at the time when sea levelreached its present position; these reefs caught up fasterwith sea level at about 6–5.5 ka (Cabioch et al., 1995).Assuming that sea level reached the present-day 10 and5 m depth lines by about 8.8 and 8.3 ka successively(Cabioch et al., 1995; Castellaro, 1999), time lags of 600and 1300 to 1800 years have separated the inundation ofsubstrates from initial colonization of barrier andfringing reefs, respectively (Fig. 11). This indicatesthat the relatively long delay in fringing reef initiationwas not a function of sea-level position, but must havebeen a response to water quality. From 8.3 to 5.5 ka, aphase of high-wave energy was likely to have occurredbefore the outer barrier reef formed close to present-daysea surface, since the water depth range over theincipient reefs was as great as 5–7 m. As long as this“high-energy window” (in the sense of Hopley, 1984)
has remained open, the water circulation regime onto theshelf has been dominated by waves and strong tidalcurrents flowing over coral communities from the outermargin shoreward; these currents, strengthened byactive trade winds, have probably helped incursions ofnutrient-rich upwelled waters to the innermost parts ofthe shelf, thus disturbing reef colonization, particularlyalong the coastal areas. The maintenance of the high-energy window opening has been favoured by a 1 mhigher sea level position relative to the present, fromabout 6.5 to 5.5 ka (Cabioch et al., 1995; Castellaro,1999). The relative similarity between the bariumpatterns recognized in the modern coral from Amédéeand the fossil ones from Vata-Ricaudy lends support tothe idea of the direct control of wind-driven upwellingon fringing reef development in the considered region.From about 5.5–5 ka, the window progressively closedas the barrier reef crests reached the sea level stabilizednear its present-day position and became sufficientlyextended laterally. Consequently, the barrier linestopped direct exchanges between the open sea andthe inner lagoon, and the present-day circulation regimeestablished. Significant changes in the rates of verticalreef accretion through time provide further support forthe validity of this evolutive scheme. Drill holespenetrating the Amédée reef (i.e. just behind the barrierreef) reveal that, from 8.2 to about 6.5 ka, during theepisode of active high-energy window, the reef grewvertically at rates not exceeding 2.5 to 5.1 mm/year,while from 6.5 ka, it developed at rates close to 10 mm/year, as the high-energy window began to close.Similarly, from 6.3 ka until its reef crest reached thesea surface, the fringing reef at Vata-Ricaudy accretedupward at the mean rate of 12 mm/year (Castellaro,1999). Such decreases in vertical reef accretion mayhave been caused by a reduction in coral skeletogenesiscontrolled by strong nutrient inputs (Marubini andDavies, 1996).
6. Conclusions
Analysis of the Sr and Ba concentrations of modernand mid-Holocene Porites corals on the southwestlagoon of New Caledonia provides a new insight intoseasonal and inter-annual environmental variability in aregion situated at the southernmost limit of the WesternPacific Warm Pool and near the southernmost limit ofreef growth. The main conclusions can be summarizedas follows:
(1) Inputs from upwelled waters are found today asthe main source of barium into reefal waters of

452 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
southwest New Caledonia, but the Ba content ofthe coral skeletons do not show a simple responseto the intensity of upwelling events. An additionalsource was identified specifically in the shallowcoastal areas where barium exhibits a clear rainfallresponse that could be linked to weathering ofbarite-bearing siliceous rocks cropping out in theNouméa area. In both types of record, there is nostrong cyclicity of the Ba signal, despite the factthat Ba incorporation in corals is controlled bycyclic phenomena, i.e. seasonal upwelling orannual rainfall. Thus, the present study confirmsthe potential of coral Ba/Ca as an indicator ofseasonal upwelling of nutrient and bariumenriched waters to sea surface, and demonstratesthat the comparative analysis of Ba/Ca ratios ofmodern and fossil coral skeletons in a given reefalregion can be used to improve our knowledge ofthe paleoceanography of coastal areas in tropicalseas during the Holocene, and particularly the roleof wind-driven current regimes.
(2) At about 6 ka, sea surface temperatures along thesouthwest coast of the island are estimated to havebeen about 1 °C higher, relative to the present.However, at that time, New Caledonia may haveexperienced occasional brief cooling events.These cannot be considered to have played asignificant role in the retardation in fringing reefdevelopment along the southwest New Caledo-nian coast, during the Holocene climatic opti-mum. Nutrient supply from upwelling appears tohave been the major factor controlling theretardation in growth of these fringing reefs.Although initial coral recruitment occurred about8.2 ka in southwest New Caledonia, the reef crestsof both the outer barrier and fringing reefs reachedthe stabilized sea surface not prior to 6–5 ka. Atthe same time as the barrier reef crest caught upwith stabilized sea level, the effects of inimicalupwelled waters on the inner shelf were stopped,and fringing reef growth was able to compensatefor the accommodation space.
(3) The knowledge of the state of preservation ofboth modern and fossil corals from southwestNew Caledonian reef tracts permitted identifica-tion of the magnitude of geochemical changesand, thus, assessment of the fidelity of the Sr/Caand Ba/Ca proxies and radiocarbon dating forpaleoenvironmental reconstruction. However,despite previous successes in the use of coralproxies for tracking climatic and oceanographicparameters back in time, several issues con-
cerning the fidelity remain to be resolved,particularly the impact of early marine alterationon the distribution of barium in live andHolocene coral skeletons. Similarly, someuncertainties remain about the behaviour ofradiocarbon during early diagenetic processes.
Acknowledgements
This research was supported by the French NationalProgramme on Coastal Environment (PNEC), Grant no.001 140708, sponsored by CNRS, IFREMER and IRD.The authors wish to thank the technicians who havehelped them in field investigations and laboratoryexperiments, especially those from IRD Centre atNouméa (New Caledonia) and from IRD Centre atBondy (France). The question of the effect of earlymarine diagenesis on radiocarbon dating has benefitedfrom discussions with Edouard Bard, Anne Cohen,Anne Mûller and Yusuke Yokoyama. The thoroughreviews of Mike Gagan, a second anonymous reviewerand the careful handling and suggestions of one of theEditors have greatly improved the manuscript.
References
Abram, N.J., Gagan, M.K., McCulloch, M.T., Chappell, J., Hantoro,W.S., 2003. Coral reef death during the 1997 Indian Ocean dipolelinked to Indonesian wildfires. Science 301 (5635), 952–955.
Alibert, C., Kinsley, L., Fallon, S.J., McCulloch, M.T., Berkelmans,R., McAllister, F., 2003. Source of trace element variability inGreat Barrier Reef corals affected by the Burdekin flood plumes.Geochimica et Cosmochimica Acta 67 (2), 231–246.
Ambatsian, P., Fernex, F., Bernat, M., Parron, C., Lecolle, J., 1997.High metal inputs to closed seas: the New Caledonian lagoon.Journal of Geochemical Exploration 59, 59–74.
Anderegg, D., Dodge, R.E., Swart, P.K., Fisher, F., 1997. Bariumchronologies from south Florida reef corals–environmentalimplications. Proceedings Eighth International Coral Reef Sympo-sium, Panama, 2, 1725–1730.
Ayliffe, L.K., Bird, M.I., Gagan, M.K., Isdale, P.J., Scott-Gagan, H.,Parker, B., Griffin, D., Nongkas, M., McCulloch, M.T., 2004.Geochemistry of coral from Papua NewGuinea as proxy for ENSOocean–atmosphere interactions in the Pacific Warm Pool.Continental Shelf Research 24, 2343–2356.
Bacon, M.P., Edmond, J.M., 1972. Barium at GEOSECS III in thesouth-western Pacific. Earth and Planetary Science Letters 16,66–74.
Beck, J.W., Edwards, R.L., Ito, E., Taylor, F.W., Récy, J., Rougerie, F.,Joannot, P., Henin, C., 1992. Sea-surface temperature from coralskeletal strontium–calcium ratios. Science 257, 644–647.
Blanchon, P., Montaggioni, L.F., 2003. Impact of rapid sea level andclimate change on late Quaternary reef development. SedimentaryGeology 159 (1–2), 1–3.
Boiseau, M., Cornu, H., Turpin, L., Juillet-Leclerc, A., 1997. Mesuresconjointes des rapports Sr/Ca et d18O effectuées sur Acroporanobilis et Porites lutea: le paléothermomètre Sr/Ca est-il toujours

453L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
fiable? Comptes Rendus de l'Académie des Sciences, Paris,Sciences de la Terre et des Planètes 325, 747–752.
Cabioch, G., 1988. Récifs frangeants de Nouvelle-Calédonie (Pacifi-que sud-ouest). Structure interne et influences de l' eustatisme et dela néotectonique. Publications de l'Université de Provence, Aix-en-Provence. 291 pp.
Cabioch, G., 2003. Post-glacial reef development in the South-WestPacific: case studies from New Caledonia and Vanuatu. Sedimen-tary Geology 159 (1–2), 49–59.
Cabioch, G., Montaggioni, L.F., Faure, G., 1995. Holocene initiationand development of New Caledonian fringing reefs, SW Pacific.Coral Reefs 14, 131–140.
Cabioch, G., Corrège, Th., Turpin, L., Castellaro, Ch., Récy, J., 1999.Development patterns of fringing and barrier reefs in NewCaledonia (Southwest Pacific). Oceanologica Acta 22, 567–578.
Cardinal, D., Hamelin, B., Bard, E., Pätzold, J., 2001. Sr/Ca, U/Ca andδ18O records in recent massive corals from Bermuda: relationshipswith sea surface temperature. Chemical Geology 176, 213–233.
Carroll, J., Falkner, K.K., Brown, E.T., Moore, W.S., 1993. The role ofthe Ganges–Brahmaputra mixing zone in supplying barium and226Ra to the Bay of Bengal. Geochimica et Cosmochimica Acta 57,2981–2990.
Castellaro, C., 1999. Reconstitutions paléoclimatiques et paléoenvir-onnementales à l'Holocène et au Pléistocène terminal en Nouvelle-Calédonie et aux Seychelles (région Indo-Pacifique): l'enregistre-ment par les coraux. Unpublished PhD thesis, University ofProvence. 210 pp.
Coffey, M., Dehairs, F., Collette, O., Luther, G., Church, T., Jickells,T., 1997. The behaviour of dissolved barium in estuaries.Estuarine, Coastal and Shelf Science 45, 113–121.
Cohen, A.L., Hart, S.R., 2004. Deglacial sea surface temperatures ofthe western tropical Pacific: a new look at old coral. Paleoceano-graphy 19, A4031, doi:10.1029/2004PA001084.
Corrège, T., Delcroix, T., Récy, J., Beck, W., Cabioch, G., Le Cornec,F., 2000. Evidence for stronger El Niño–Southern Oscillation(ENSO) events in a mid-Holocene massive coral. Paleoceano-graphy 15 (4), 465–470.
Crowley, T.J., 2000. Causes of climate change over the past 1000years. Science 289, 270–277.
Davies, P.J., Montaggioini, L.F., 1985. Reef growth and sea levelchange: the environmental signature. Proceedings Fifth Interna-tional Coral Reef Congress, Tahiti 3, 477–511.
Delcroix, T., Hénin, C., 1989. Mechanisms of subsurface thermalstructure and subsurface thermohaline variability in the south-western tropical Pacific during 1975–85. Journal of MarineResearch 47, 777–812.
Delcroix, T., Lenormand, O., 1997. ENSO signals in the vicinity ofNew Caledonia, south-western Pacific. Oceanologica Acta 20,481–491.
de Villiers, S., 1999. Seawater Sr and Sr/Ca variability in the Atlanticand Pacific Oceans. Earth and Planetary Science Letters 171,623–634.
de Villiers, S., Shen, G.T., Nelson, B.K., 1994. The Sr/Ca–temperaturerelationship in coralline aragonite: influence of variability in (Sr/Ca) sea water and skeletal growth parameters. Geochimica etCosmochimica Acta 58, 197–208.
Donguy, J.R., Oudot, C., Rougerie, F., 1970. Circulation superficielleet subsuperficielle en Mer de Corail et à 170°E. Cahiers-ORSTOM. Série Océanographie VIII (1), 1–20.
Douillet, P., Ouillon, S., Cordier, E., 2001. A numerical model for finesuspended sediment transport in the south-west lagoon of NewCaledonia. Coral Reefs 20, 361–372.
Dymond, J., Suess, E., Lyle, M., 1992. Barium in deep-sea sediment: ageochemical proxies for paleoproductivity. Paleoceanography 7,163–181.
Edmond, J.M., Boyle, E.A., Drummond, D., Grant, B., Gislick, T.,1978. Desorption of barium in the plume of the Zaire (Congo)River. Netherlands Journal of Sea Research 12, 324–328.
Elderfield, H., 1990. Tracers of ocean paleoproductivity andpaleochemistry: an introduction. Paleoceanography 5, 711–717.
Enmar, R., Stein, M., Bar-Matthews, Sass, E., Katz, A., Lazar, B.,2000. Diagenesis in live corals from the Gulf of Aqaba: I. Theeffect on paleo-oceanography tracers. Geochimica et Cosmochi-mica Acta 64, 3123–3132.
Fagel, N., Dehairs, F., Peinert, R., Antia, A., André, L., 2004.Reconstructing export production at the NE Atlantic margin:potential and limits of the Ba proxy. Marine Geology 204,11–25.
Fallon, S.J., McCulloch, M.T., van Woesik, R., Sinclair, D.J., 1999.Corals at their latitudinal limits: laser ablation trace elementsystematic in Porites from Shirigai Bay, Japan. Earth and PlanetaryScience Letters 172, 221–238.
Felis, T., Patzöld, J., 2003. Climate records from corals. In: Wefer, G.,Lamy, F., Mantoura, F. (Eds.), Marine Science Frontiers forEurope. Sprnger-Verlag, Berlin, pp. 11–27.
Felis, T., Lohmann, G., Kuhnert, H., Lorenz, S.J., Scholz, D., Pätzold,J., Al-Rousan, S.A., Al-Moghrabi, S.M., 2004. Increased season-ality in Middle East temperatures during the last interglacialperiod. Nature 429, 164–168.
Gagan, M.K., Ayliffe, L.K., Hopley, D., Cali, J.A., Mortimer, G.E.,Chappell, J., McCulloch, M.T., Head, M.J., 1998. Temperature andsurface-ocean water balance of the mid-Holocene tropical westernPacific. Science 279, 1014–1018.
Gagan, M.K., Ayliffe, L.K., Beck, J.W., Cole, J.E., Druffel, E.R.M.,Dunbar, R.B., Schrag, D.P., 2000. New views of tropicalpaleoclimates from corals. Quaternary Science Reviews 19, 45–64.
Gagan, M.K., Ayliffe, L.K., Opdyke, B.N., Hopley, D., Scott-Gagan,H., Cowley, J., 2002. Coral oxygen isotope evidence for recentgroundwater fluxes to the Australian Great Barrier Reef.Geophysical Research Letters 29 (20), 1982. doi:10.1029/2002GL015336.
Gagan, M.K., Hendy, E.J., Haberle, S.G., Hantoro, W.S., 2004. Post-glacial evolution of the Indo-Pacific Warm Pool and El Niño–Southern Oscillation. Quaternary International 118–119, 127–143.
Gvirtzman, G., Friedman, G.M., 1977. Sequence of progressivediagenesis in coral reefs. In: Frost, S.H., Weiss, M.P., Saunders, J.B. (Eds.), Reefs and Related Carbonates–Ecology and Sedimen-tology. Studies in Geology, vol. 4. American Association ofPetroleum Geologists, Tulsa, pp. 357–380.
Hallock, P., 1988. The role of nutrient availability in bioerosion:consequence to carbonate buildups. Palaeogeography, Palaeocli-matology, Palaeoecology 63, 275–291.
Hallock, P., 2001. Coral reefs, carbonate sediments, nutrient and globalchange. In: Stanley, G.D. (Ed.), The History and Sedimentology ofAncient Reef Systems. Kluwer Academy-Plenum Publisher, New-York, pp. 387–427.
Hallock, P., Schlager, W., 1986. Nutrient excess and the demise ofcoral reefs and carbonate platforms. Palaios 1, 389–398.
Hanor, J.S., Chan, L.-H., 1977. Non-conservative behavior of bariumduring mixing of Mississippi River and Gulf of Mexico waters.Earth and Planetary Science Letters 37, 242–250.
Hart, R.S., Cohen, A.L., Ramsay, P., 1996. Microscale analysis of Sr/Ca and Ba/Ca in Porites. Proceedings Eighth International CoralReef Symposium, Panama 2, 1707–1712.

454 L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
Heiss, G.A., Camoin, G.F., Eisenhauer, A., Wischow, D., Dullo, W.C. ,Hansen, B., 1996. Stable isotope and Sr/Ca – signals in corals fromthe Indian Ocean. Proceedings Eighth International Coral ReefSymposium, Panama 2, 1713–1718.
Hénin, C., Cresswell, G.R., 2005. Upwelling along the western barrierreef of New Caledonia. Marine and Freshwater Research 56,1005–1010.
Hopley, D., 1984. The Holocene “high-energy window” on the CentralGreat Barrier Reef. In: Thom, B.G. (Ed.), Coastal Geomorphology inAustralia. Academic Press, Sydney, pp. 303–340.
Hubbard, D.K., 1997. Reefs as dynamic systems. In: Birkeland, C.(Ed.), Life and Death of Coral Reefs. Chapman & Hall, New-York,pp. 43–67.
Kasten, S., Haese, R.R., Zabel, M., Rühlemann, C., Schulz, H.D.,2001. Barium peaks at glacial terminations in sediments of theequatorial Atlantic Ocean–relicts of deglacial productivity pulses?Chemical Geology 175, 635–651.
Kinsey, D.W., Davies, P.J., 1979. Effects of elevated nitrogen andphosphorous on coral reef growth. Limnology and Oceanography24, 935–940.
Kleypas, J., 1997. Modelled estimates of global reef habitat andcarbonate production since the Last Glacial Maximum. Paleocea-nography 12, 533–545.
Klump, J., Hebbeln, D., Wefer, G., 2001. High concentrations ofbiogenic barium in Pacific sediments after termination: 1. A signalof changes in productivity and deep water chemistry. MarineGeology 177, 1–11.
Latham, M., 1985. Altération et pédogenèse sur roches ultra-basiquesen Nouvelle-Calédonie. Unpublished D.Sc thesis, Dijon Univer-sity. 331 pp.
Launay, J., 1972. La sédimentation en baie de Dumbéa (côte ouest–Nouvelle Calédonie). Cahiers-ORSTOM. Série Océanographie IV(1), 25–51.
Lazar, B., Enmar, R., Schossberger, M., Bar-Mattews, M., Halicz, L.,Stein, M., 2004. Diagenetic effects on the distribution of uraniumin live and Holocene corals from the Gulf of Aqaba. Geochimica etCosmochimica Acta 68, 4583–4593.
Lea, D.W., Boyle, E.A., 1989. Barium content of benthicforaminifera controlled by bottom water composition. Nature338, 751–753.
Lea, D.W., Shen, G.T., Boyle, E.A., 1989. Coralline barium recordstemporal variability in Equatorial Pacific upwelling. Nature 340,373–376.
Le Cornec, F., Corrège, T., 1997. Determination of uranium to calciumand strontium to calcium ratios in corals by inductively coupledplasma mass spectrometry. Journal of Analytical Atomic Spec-trometry 12, 969–973.
Marshall, J.F., McCulloch, T.M., 2002. An assessment of the Sr/Caratio in shallow water hermatypic corals as a proxy for sea surfacetemperature. Geochimica et Cosmochimica Acta 66 (18),3263–3280.
Marubini, S., Davies, P.S., 1996. Nitrate increases zooxanthellaepopulation density and reduces skeletogenesis in corals. MarineBiology 127, 319–328.
McCulloch, M.T., Mortimer, G.E., Esat, T., Xianhua, L., Pillans, B.,Chappell, J., 1996. High resolution windows into early Holoceneclimate: Sr/Ca coral records from the Huon Peninsula. Earth andPlanetary Science Letters 138, 169–178.
McCulloch, M.T., Fallon, S., Wyndham, T., Hendy, E., Lough, J.,Barnes, D., 2003. Coral record of increased sediment flux to theinner Great Barrier Reef since European settlement. Nature 421,727–730.
McLaughlin, C.J., Smith, C.A., Buddemeier, R.W., Bartley, J.D.,Maxwell, B.A., 2003. Rivers, runoff, and reefs. Global andPlanetary Change 39, 191–199.
McManus, J., Berelson, W.M., Klikhammer, G.P., Johnson, K.S.,Coale, K.H., Anderson, R.F., Kumar, N., Burdige, D.J., Hammond,D.E., Brumsack, H.J., McCorkle, D.C., Rushdi, A., 1998.Geochemistry of barium in marine sediments: implications for itsuse as a paleoproxy. Geochimica et Cosmochimica Acta 62,3453–3473.
Météo France, 1995. Atlas climatique de la Nouvelle-Calédonie.Météo-France, Direction interrégionale de Nouvelle-Calédonie etde Wallis et de Futuna, Nouméa. 104 pp.
Montaggioni, L.F., 1988. Holocene reef growth history in mid-platehigh volcanic islands. Proceedings Sixth International Coral ReefSymposium, Townsville, Australia, 3, 455–460.
Montaggioni, L.F., 2000. Postglacial reef growth. Comptes rendus del'Académie des Sciences, Paris, Earth and Planetary Sciences 331,319–330.
Montaggioni, L.F., 2005. History of Indo-Pacific coral reef systemssince the last glaciation: development patterns and controllingfactors. Earth-Science Reviews 71, 1–75.
Montaggioni, L.F., Macintyre, I.G., 1991. Reefs as recorders ofenvironmental changes. Coral Reefs 10 (2), 53–54.
Montaggioni, L.F., Cuet, P., Naïm, O., 1993. Effects of nutrient excesson a modern fringing reef (Réunion island). Geological implica-tions. Proceedings Colloquium on Global Aspects of Coral Reefs,Miami, 397–403.
Moore, W.S., 1997. High fluxes of radium and barium from the mouthof the Ganges–Brahmaputra River during low river dischargesuggest a large ground water source. Earth and Planetary ScienceLetters 150, 141–150.
Müller, A., Gagan, M.K., Lough, J., 2001. Effect of early marinediagenesis on coral reconstructions of surface-ocean 13C/12C andcarbonate saturation state. Global Biogeochemical Cycles 18,GB1033. doi:10.1029/2003GB002112.
Müller, A., Gagan, M.K., Lough, J.M., 2004. Effect of early marinediagenesis on coral reconstructions of surface-ocean 13C/12C andcarbonate saturation state. Global Biogeochemical Cycles 18,GB103. doi:10.1029/2003GB002112.
Neumann, A.C., Macintyre, I.G., 1985. Reef response to sea level rise:keep-up, catch-up or give-up. Proceedings Fifth InternationalCoral Reef Congress, Tahiti 3, 105–110.
Paris, J.P., 1981. Les ressources minérales de la Nouvelle-Calédonie.Mémoire du Bureau de Recherches Géologiques et Minières,Orléans–La Source 113, 219–224.
Pinazo, C., Bujan, S., Douillet, P., Fichez, R., Grenz, C., Maurin,A., 2004. Impact of wind and freshwater inputs on phyto-plankton biomass in the coral reef lagoon of New Caledoniaduring the summer cyclonic period: a coupled tri-dimensionalbiogeochemical modelling approach. Coral Reefs 23 (2),281–296.
Pingitore Jr., N.E., Rangel, Y., Kwarteng, A., 1989. Barium variationin Acropora palmata andMontastrea annularis. Coral Reefs 8 (1),31–36.
Prakash Babu, C., Brumsack, H.-J., Schnetger, B., Böttchert, M.E.,2002. Barium as a productivity proxy in continental marginsediments: a study from the eastern Arabian Sea. Marine Geology184, 189–206.
Quinn, T.M., Sampson, D.E., 2002. A multiproxy approach toreconstructing sea surface conditions using coral skeletongeochemistry. Paleoceanography 17, 1062. doi:10.1029/2000PA000528.

455L.F. Montaggioni et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 237 (2006) 436–455
Quinn, T.M., Tudhope, A.W., 2000. Science and technology ofsubmerged coral drilling: a workshop report. InternationalWorkshop, NSF-JOI/USSAC, Florida, Sept 2000, pp. 1–82.
Reuer, M.K., Boyle, E.A., Cole, J.C., 2003. A mid-twentieth centuryreduction in tropical upwelling inferred from coralline traceelement proxies. Earth and Planetary Science Letters 210,437–452.
Ribaud-Laurenti, A., Hamelin, B., Montaggioni, L.F., Cardinal, D.,2001. Diagenesis and its impact on Sr/Ca ratio in HoloceneAcropora corals. International Journal of Earth Sciences 90,438–451.
Rimbu, N., Lohmann, G., Lorenz, S.J., Kim, J.H., Schneider, R.R.,2004. Holocene climate variability as derived from alkenone seasurface temperature and coupled ocean–atmosphere modelexperiments. Climate Dynamics 23, 215–227.
Shen, G.T., Sanford, C.L., 1990. Trace-element indicators of climatevariability in reef-building corals. In: Glynn, P.W. (Ed.), GlobalEcological Consequences of the 1982–83 El Niño–SouthernOscillation. Elsevier, pp. 255–283.
Shen, G.T., Cole, J.E., Lea, D.W., Linn, N.J., McConnaughey, T.A.,Fairbanks, R.G., 1992. Surface ocean variability at Galapagosfrom 1936–1982: calibration of geochemical tracers in corals.Paleoceanography 7 (5), 404–407.
Sinclair, D.J., 2005. Non-river flood barium signals in the skeletons ofcorals from coastal Queensland, Australia. Earth and PlanetaryScience Letters 237, 354–369.
Sinclair, D.J., McCulloch, M.T., 2004. Corals record low mobilebarium concentrations in the Burdekin River during the 1974flood: evidence for limited supply to rivers? Palaeogeography,Palaeoclimatology, Palaeoecology 214 (1–2), 155–174.
Smith, S.V., Buddemeier, R.W., 1992. Global change and coral reefecosystems. Annual Reviews of Ecological Systems 23, 89–118.
Smith, S.V., Buddemeier, R.W., Redalje, R.J., Houck, J.E., 1979.Strontium–calcium thermometry in coral skeletons. Science 204,404–407.
Spiro, B.F., 1971. Diagenesis of some scleractinian corals from theGulf of Elat, Israël. Danmarks Geologie Undergögelse Skr. 21,1–10.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen, K.A., Kromer, B., McCornick, G., Van der Plicht, J., Spurk, M.,1998. INTCAL 98 radiocarbon age calibration, 24,000–0 cal B.P.Radiocarbon 40, 1041–1083.
Swart, P.K., Grottoli, A.G., 2003. Proxy indicators of climate in coralskeletons: a perspective. Coral Reefs 22 (4), 313–315.
Tudhope, A.T., Lea, D.W., Shimmield, G.B., Chilcott, C.P., Head, S.,1996. Monsoon climate and Arabian Sea coastal upwellingrecorded in massive corals from Southern Oman. Palaios 11 (4),347–361.
Tudhope, A.W., Lea, D.W., Shimmield, G.B., Chilcott, C.P.,Scoffin, T.P., Fallick, A.E., Jebb, M., 1997. Climatic recordsfrom massive Porites corals in Papua New-Guinea: a compar-ison of skeletal Ba/Ca, skeletal δ18O and coral rainfall.Proceedings Eighth International Coral Reef Symposium,Panama, 2, 1719–1724.
Wei, G., Sun, M., Li, X., Nie, B., 2000. Mg/Ca, Sr/Ca and U/Ca ratiosof Porites coral from Sanya Bay, Hainan Island, South China Seaand their relationships to sea surface temperature. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology 162, 59–74.
Yokoyama, Y., Esat, T.M., 2004. Long term variations of uraniumisotopes and radiocarbon in the surface water recorded in corals.In: Shiyomi, M., Kawahata, H., Koizumi, H., Tsuda, A. (Eds.),Global Environmental Change in the Ocean and on Land.Terrapub, Tokyo, pp. 279–309.
Yu, K.-F., Zhao, J.-X., Liu, T.-S., Wei, G.-J., Wang, P.-X., Collerson,K.D., 2004. High-frequency winter cooling and reef coralmortality during the Holocene climatic optimum. Earth andPlanetary Science Letters 224, 143–155.
Related Documents











