ENVIRONMENTAL BIOTECHNOLOGY Copper resistance of biofilm cells of the plant pathogen Xylella fastidiosa Carolina M. Rodrigues & Marco A. Takita & Helvécio D. Coletta-Filho & Jacqueline C. Olivato & Raquel Caserta & Marcos A. Machado & Alessandra A. de Souza Received: 13 June 2007 / Revised: 26 September 2007 / Accepted: 28 September 2007 / Published online: 9 November 2007 # Springer-Verlag 2007 Abstract Xylella fastidiosa is a phytopathogen that causes diseases in different plant species. The development of disease symptoms is associated to the blockage of the xylem vessels caused by biofilm formation. In this study, we evaluated the sensitivity of biofilm and planktonic cells to copper, one of the most important antimicrobial agents used in agriculture. We measured the exopolysaccharides (EPS) content in biofilm and planktonic cells and used real-time reverse transcription polymerase chain reaction to evaluate the expression of the genes encoding proteins involved in cation/multidrug extru- sion (acrA/B, mexE/czcA, and metI) and others associated with different copper resistance mechanisms (copB, cutA1, cutA2, and cutC) in the X. fastidiosa biofilm formed in two different media. We confirmed that biofilms are less susceptible to copper than planktonic cells. The amount of EPS seems to be directly related to the resistance and it varies according to the media where the cells are grown. The same was observed for gene expression. Nevertheless, some genes seem to have a greater importance in biofilm cells resistance to copper. Our results suggest a synergistic effect between diffusion barriers and other mechanisms associated with bacterial resistance in this phytopathogen. These mechanisms are important for a bacterium that is constantly under stress conditions in the host. Keywords metal . EPS . real-time RT-PCR Introduction Xylella fastidiosa is the causative agent of citrus variegated chlorosis (CVC), a disease present in almost half of the citrus growing areas in Brazil (http://www.fundecitrus.com. br). CVC strongly affects sweet orange production in the North, Northwest, and Central areas of the Sao Paulo State, Brazil, leading to important economic losses. Growers in Brazil have undertaken several control measures including pruning of affected branches and eradication of highly diseased plants to remove the inoculum, spraying of insecticides to reduce the population of transmission vectors (sharpshooters), and production of seedlings in greenhouses covered by plastic and laterally protected by screens. These measures have increased the cost of production that could reach US$286–322 million per year (Fernandes 2003). Because of its importance, X. fastidiosa was the first phytopathogen to have its complete genome sequenced (Simpson et al. 2000). The symptoms of CVC result from water stress caused by a blockage of the xylem vessels (Osiro et al. 2004; De Souza et al. 2004). The vessels are clogged by biofilm cells, and biofilms have many important biological characteristics including increased resistance to a broad range of anti- microbial agents (Mah and O’Toole 2001). These resistance Appl Microbiol Biotechnol (2008) 77:1145–1157 DOI 10.1007/s00253-007-1232-1 DO01232; No of Pages C. M. Rodrigues : M. A. Takita : H. D. Coletta-Filho : J. C. Olivato : R. Caserta : M. A. Machado : A. A. de Souza (*) Centro APTA Citros Sylvio Moreira, Instituto Agronômico de Campinas, Cordeirópolis, Brazil e-mail: [email protected] C. M. Rodrigues Universidade Estadual Paulista (UNESP), Instituto de Biociências de Botucatu, Botucatu, Brazil M. A. Takita Centro de Pesquisa e Desenvolvimento de Recursos Genéticos Vegetais, Instituto Agronômico de Campinas, Campinas, Brazil R. Caserta Universidade Estadual de Campinas (UNICAMP), Campinas, Brazil

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ENVIRONMENTAL BIOTECHNOLOGY
Copper resistance of biofilm cells of the plant pathogenXylella fastidiosa
Carolina M. Rodrigues & Marco A. Takita &
Helvécio D. Coletta-Filho & Jacqueline C. Olivato &
Raquel Caserta & Marcos A. Machado &
Alessandra A. de Souza
Received: 13 June 2007 /Revised: 26 September 2007 /Accepted: 28 September 2007 / Published online: 9 November 2007# Springer-Verlag 2007
Abstract Xylella fastidiosa is a phytopathogen that causesdiseases in different plant species. The development of diseasesymptoms is associated to the blockage of the xylem vesselscaused by biofilm formation. In this study, we evaluated thesensitivity of biofilm and planktonic cells to copper, one of themost important antimicrobial agents used in agriculture. Wemeasured the exopolysaccharides (EPS) content in biofilmand planktonic cells and used real-time reverse transcriptionpolymerase chain reaction to evaluate the expression of thegenes encoding proteins involved in cation/multidrug extru-sion (acrA/B, mexE/czcA, and metI) and others associatedwith different copper resistance mechanisms (copB, cutA1,cutA2, and cutC) in the X. fastidiosa biofilm formed in twodifferent media. We confirmed that biofilms are lesssusceptible to copper than planktonic cells. The amount ofEPS seems to be directly related to the resistance and itvaries according to the media where the cells are grown. Thesame was observed for gene expression. Nevertheless, some
genes seem to have a greater importance in biofilm cellsresistance to copper. Our results suggest a synergistic effectbetween diffusion barriers and other mechanisms associatedwith bacterial resistance in this phytopathogen. Thesemechanisms are important for a bacterium that is constantlyunder stress conditions in the host.
Keywords metal . EPS . real-time RT-PCR
Introduction
Xylella fastidiosa is the causative agent of citrus variegatedchlorosis (CVC), a disease present in almost half of thecitrus growing areas in Brazil (http://www.fundecitrus.com.br). CVC strongly affects sweet orange production in theNorth, Northwest, and Central areas of the Sao Paulo State,Brazil, leading to important economic losses. Growers inBrazil have undertaken several control measures includingpruning of affected branches and eradication of highlydiseased plants to remove the inoculum, spraying ofinsecticides to reduce the population of transmissionvectors (sharpshooters), and production of seedlings ingreenhouses covered by plastic and laterally protected byscreens. These measures have increased the cost ofproduction that could reach US$286–322 million per year(Fernandes 2003). Because of its importance, X. fastidiosawas the first phytopathogen to have its complete genomesequenced (Simpson et al. 2000).
The symptoms of CVC result from water stress caused bya blockage of the xylem vessels (Osiro et al. 2004; De Souzaet al. 2004). The vessels are clogged by biofilm cells, andbiofilms have many important biological characteristicsincluding increased resistance to a broad range of anti-microbial agents (Mah and O’Toole 2001). These resistance
Appl Microbiol Biotechnol (2008) 77:1145–1157DOI 10.1007/s00253-007-1232-1
DO01232; No of Pages
C. M. Rodrigues :M. A. Takita :H. D. Coletta-Filho :J. C. Olivato : R. Caserta :M. A. Machado :A. A. de Souza (*)Centro APTA Citros Sylvio Moreira,Instituto Agronômico de Campinas,Cordeirópolis, Brazile-mail: [email protected]
C. M. RodriguesUniversidade Estadual Paulista (UNESP),Instituto de Biociências de Botucatu,Botucatu, Brazil
M. A. TakitaCentro de Pesquisa e Desenvolvimento de Recursos GenéticosVegetais, Instituto Agronômico de Campinas,Campinas, Brazil
R. CasertaUniversidade Estadual de Campinas (UNICAMP),Campinas, Brazil

mechanisms involve exopolymer matrices, changes in geneexpression, and metabolic alterations that make the microbi-ological population difficult to control (Costerton et al. 1995;Teitzel and Parsek 2003; Surdeau et al. 2006).
It was recently demonstrated that there is an increase ingene expression in the X. fastidiosa biofilm compared toplanktonic cells. Some of the upregulated genes encodecation/multidrug efflux pumps that are associated withdefense mechanisms (De Souza et al. 2004). However, nostudy has been done to evaluate whether X. fastidiosa cellsin biofilm are more resistant or not to antimicrobialcompounds compared to planktonic cells. Moreover, noinformation regarding the association of these genes withresistance is available for this organism.
Biofilms are capable of adsorbing heavy metal cationsfrom their surroundings and sequestering them in theextracellular matrix, which contributes to heavy metalresistance (Surdeau et al. 2006; Harrison et al. 2006). Thehigher biofilm resistance seems to involve efflux pumps,like the ones belonging to the resistance-nodulation-celldivision (RND) protein family, the ATP-binding cassette(ABC) transporters, the cation translocating P-typeATPases, and others mechanisms (Mah and O’Toole2001; Ramage et al. 2002). The RND proteins worktogether with members of the membrane fusion proteinfamily (MFP). Many RND proteins cooperate with yet athird protein that belongs to the family of outer membranefactors (OMF). These three proteins form an efflux proteincomplex, which may export its substrate from the cyto-plasm directly to the outside. Members of this superfamilyare involved in the transport of a broad range of structurallyunrelated molecules to the exterior of the cells includingmost classes of antibiotics, biocides, cationic antimicrobialpeptides, associated homoserine lactones, quorum-sensingmolecule, and others (Saier et al.1994; Poole 2001, 2002;Yang et al. 2006). The ABC transporters form a mechanismknown as multidrug resistance that transports molecules bycoupling drug/lipid efflux with energy derived from thehydrolysis of ATP (Chang 2003). The P-type ATPases areamong the commonest pumping mechanisms used to exportmetal ions out of cells or into organelles. The pump designis conserved from bacteria to humans and is used fortransporting a wide range of elements (Dameron andHarrison 1998). Some of these systems are widespreadand serve in the basic defense of the cell against heavymetals, but some are highly specialized and occur only infew bacteria (Nies 2003). Other mechanisms seem to beinvolved with metal resistance like the cut genes fromEscherichia coli. These genes are possibly involved in theuptake, intracellular storage, delivery, and efflux of copper.Nevertheless, not much is known about the mechanism bywhich these genes are involved in resistance to this metal(Rouch and Brown 1997).
The resistance to antimicrobial compounds in biofilmcells is known for many different bacteria, but littleinformation is available for plant pathogens. For X.fastidiosa, this information would be extremely relevantconsidering that its main pathogenicity mechanism isbiofilm formation inside the xylem vessels. Moreover, inagriculture, copper-containing compounds are among themain antimicrobial compounds used to limit the spread ofplant pathogenic bacteria and fungi on vegetable and fruitscrops (Voloudakis et al. 2005).
In this study, we investigated the effect of copper onbiofilm and planktonic X. fastidiosa cells grown in definedand undefined media. Under our experimental conditions,biofilms were less susceptible to copper than planktoniccells. We also chose genes previously found to beupregulated in previous studies of either biofilm or virulentcells for gene expression analysis (acrA/B, mexA/czcD,metI) (De Souza et al. 2003, 2004). We also wanted to testthe families known to be specifically related to copperresistance in other organisms like copB and cut genes.Other mechanisms that confer copper resistance in E. colilike the CueO and Cus systems were not found in X.fastidiosa and, therefore, could not be evaluated. Thisresistance seems to be a result of the synergistic effect ofdifferent mechanisms including high amount of exopoly-saccharides (EPS) and gene expression that could lead tothe synthesis of components of different systems involvedin the resistance to antimicrobial compounds.
Materials and methods
Bacterial strains and media
The 9a5c bacterial strain of X. fastidiosa subsp. pauca(Schaad et al. 2004) was used in this study. This strain wasthe same used for genome sequencing and obtained from theInstitut National de La Recherche Agronomique (INRA,Bordeaux, France). Bacterial cells were inoculated into theplant to maintain their pathogenicity state and avoidattenuation due to successive passages in the axenic medium.Petioles and stems were aseptically ground in phosphate-buffered saline (PBS) and the suspension was spread onperiwinkle wilt medium (PW) (Davis et al. 1981). The firstcolonies were observed between 10 and 15 days afterinoculation and such cells were used in the experiments.
MIC determination of planktonic cells
To obtain planktonic cells of X. fastidiosa, severalindividual colonies were transferred to a polypropylenetube containing 3 ml of PW broth. When the A600nm
reached 0.2, corresponding to 108 colony-forming unit
1146 Appl Microbiol Biotechnol (2008) 77:1145–1157

(CFU)/ml, the cells were transferred to a 250-ml flaskcontaining 50 ml of either PW or a defined medium for X.fastidiosa (XDM2) (Lemos et al. 2003). Swimming cellswere weekly transferred for at least ten times. After thispoint, the cells that did not adhere to the glass surfaces wereutilized as the planktonic growth condition.
Planktonic cells in the stationary phase, obtained after10 days of growth, were used in minimal inhibitoryconcentration (MIC) experiments. The population is inhigh density with minimal growth under this condition,providing a better comparison to biofilm cells. Aliquots of1 ml from planktonic stationary phase cells were transfer toNunclon delta SI Multidish 24 wells (Nunc A/S, Roskilde,Denmark). In each well, CuSO4 was inoculated to aconcentration ranging from 0.1 to 5.0 mM. Control withoutCuSO4 was included in all experiments. The MIC wasdetermined after 24 h of growth at 28°C. The MIC wasconsidered as the lowest concentration required for thecomplete inhibition of cell growth after 24 h of incubationin the presence of CuSO4. Complete inhibition wasassumed if a maximum of one colony was present ontriplicate plates inoculated with undiluted culture (Broounet al. 2000).
MIC determination of biofilm cells
Cells in biofilms were obtained by inoculating severalindividual colonies in a polypropylene tube containing 3 mlof PW broth. When the A600nm reached 0.2, correspondingto 108 CFU/ml, the tubes were vortexed and the cells weretransferred to a 250-ml flask containing 50 ml of either PWor XDM2 broth. After 2 to 3 days of growth at 28°C in arotary shaker at 120 rpm, a thin biofilm formation wasobserved attached to the glass at the medium–air interface.Biofilms were also formed on cover glasses at the bottomof Nunclon delta SI Multidish 24 wells. After 15 days,CuSO4 was added to the flasks and the wells in differentconcentrations. For determining the MIC to biofilm cells,we used the MIC for planktonic cells as the initialconcentration in the tests and increased from that. Fourdifferent CuSO4 concentrations ranging from 3.5 to 7 mMwere evaluated. After 24 h of incubation in the presence ofthe CuSO4, the biofilm was scrapped, dislodged, andwashed twice in Milli-Q water. For each CuSO4 concen-tration tested, cells were collected from three differentflasks or cover glasses. The MIC was determined after 24 hof growth at 28°C as described above for the planktoniccells.
Determination of EPS from planktonic and biofilm cells
Planktonic and biofilm cells were grown as described abovein both defined and undefined media. The cells were
collected by centrifugation at 8,000×g for 6 min. Thepellets were weighted and adjusted by the lowest amount ofcells obtained (25 mg). The EPS content was determined bythe phenolsulfuric method (Dubois et al. 1956) usingglucose as standard. The experiment was carried out usingsix repetitions, and the means were compared by the t test(P≤0.05) using the Assistat 7.3 beta software (http://assistat.sites.uol.com.br).
RNA isolation and cDNA synthesis
For the analysis of gene expression in biofilm cells withsubminimum inhibitory concentrations (sub-MICs) andminimum inhibitory CuSO4 concentrations, we followedthe same procedure described for MIC determination. Forthis analysis, cells attached to the glass at the medium–airinterface were used. A fraction of the cells collected after24 h of incubation in the presence of CuSO4 was washed bycentrifugation at 8,000×g for 5 min at 4°C with diethylpyr-ocarbonate-treated water. The pellet was stored at −80°Cuntil RNA extraction.
Total RNAwas isolated from X. fastidiosa cells using theRNeasy RNA extraction kit (Qiagen) and treated with theRNase-Free DNase Set (Qiagen). RNA concentration andits integrity were analyzed in the Agilent 2100 Bioanalyzerusing a RNA Nano Labchips kit. For cDNA synthesis, aconcentration of 1 μg of total RNAwas mixed with 3 μg ofrandom hexamers (Invitrogen) in a final volume of 10 μl.Annealing was accomplished by incubation for 15 min at75°C, followed by the addition of 5 μl SuperScript IIreaction buffer (Invitrogen), 1 μl of 0.1 M of dithiothreitol,1 μl dNTP mix (10 mM dATP, 10 mM dGTP, 10 mMdTTP, 10 mM dCTP), 1 μl of RNaseOut (40 U/μlInvitrogen), and 1 μl of SuperScript II reverse transcriptase(200 U/μl Invitrogen) to the reaction. To rule out thepossibility of amplification from contaminating DNA, PCRusing the RNA as template was done in parallel.
Primer design and validation
All the primers used in this study (Table 1) were designedin the PrimerExpress software version 2.0 (AppliedBiosystems) using DNA sequences obtained in the X.fastidiosa web site (http://aeg.lbi.ic.unicamp.br/xf/).
The specificity of the primers was checked by sequenc-ing the amplicons with the DNA Kit Big Dye TerminatorCycle Sequencing Ready Reaction v 3.0. The sequenceswere obtained in an ABI 3730 automatic sequencer(Applied Biosystems). The nucleotide sequences wereevaluated by aligning them with the original sequencesusing the BlastN tool (Altschul et al. 1997). Moreover,melt-curve analysis was performed from 60°C to 95°C foreach primer at the end of the real-time reverse transcription
Appl Microbiol Biotechnol (2008) 77:1145–1157 1147

polymerase chain reaction (RT-PCR) to confirm theamplification of a unique product for each gene.
To evaluate the amplification efficiency of the differentprimers, standard curves were constructed by plotting the Ct
values against the logarithm of cDNA copies of a set ofstandard solutions. Standard deviations were calculated forreplicate measurements. To estimate the accuracy of thereal-time RT-PCR, we evaluated the correlation coefficients(R2). The slope of the standard curve was used to calculatethe real-time RT-PCR efficiency according to the formulaE=10(−1/slope)−1. A PCR efficiency of 1 indicates thehighest efficiency. Primers that showed efficiency between0.9 and 1 were considered adequate for our experiments.
Gene expression analysis by real-time RT-PCR
Total RNA was extracted from biofilm cells grown in PWmedium containing 3, 4, 5, 6, and 7 mM of CuSO4 and inXDM2 medium with 3, 5, and 7 mM of CuSO4. Thebiofilm without CuSO4 was used as calibrator (control).
Real-time RT-PCR was performed for each of theresistance-associated genes and for the endogenous controlon cDNA templates prepared from total RNA. The petCgene, which encodes an ubiquinol cytochrome C oxidore-ductase, was used as the endogenous control in theexperiment. It was chosen for having similar expressionlevels (P≤0.05) in real-time RT-PCR analyses of the X.fastidiosa growing in biofilm with or without CuSO4 indifferent concentrations. Reactions were prepared accordingto the following setup: 12.5 μl of SYBR Green PCR master
mix (Applied Biosystems), 50 ng of each primer set(Table 1), 1 μl of cDNA, and water to 25 μl. Theamplification condition was 1 cycle at 50°C for 2 min,95°C for 10 min, followed by 40 cycles of 95°C for 1 s, and1 min at 60°C. No template controls were also included todetect any spurious signals arising from the amplification ofany DNA contamination or primer dimers formed duringthe reaction. The products for each primer set were alsosubjected to melt-curve analyses. This procedure wascarried out from 60°C to 95°C to ensure that the resultingfluorescence was originated from a single PCR product anddid not result from a spurious amplification. The real-timeRT-PCR was done using the ABI PRISM 7000 SequenceDetector System (Applied Biosystems). The results wereanalyzed with the ABI PRISM 7000 SDS version 1.1 usingrelative quantification analyses (Applied Biosystems).Detection of the PCR products was measured by monitor-ing the increase in fluorescence caused by the binding ofthe SYBR green dye to double-stranded DNA. A fluores-cence threshold was set automatically to 0.2. The endog-enous control was used to normalize the samples fordifferences in the amounts of cDNA added to each reactionmixture. The results were normalized using the thresholdcycle (Ct) obtained for the endogenous control present inthe same plate. Ct is defined as the first amplification cycleat which fluorescence indicating PCR products is detectableabove the threshold. For normalization, we utilized theequation: ΔCt=Ct (target gene)−Ct (endogenous control).The fold increase of the target gene in the different days ofthe biofilm formation was determined by the equation:
Table 1 Genes evaluated andnucleotide sequences of theprimers used for real-timeRT-PCR
a http://aeg.lbi.ic.unicamp.br/xf/b Endogenous control
ORF numbera Gene name Primer sequence Fragment size(in base pairs)
XF2093 acrA F—5′ TGGCCAACAGCGAGTTACTG 3′ 151R—5′ ACTTCCCTGGTGCTGTGCTT 3′
XF2094 acrB F—5′ TGATTCCGGTCGCCTTACTC 3′ 151R—5′ GTCATGATGCGCTCGACATT 3′
XF2084 mexE F—5′ CGCTTGCCCTAACAGCATGT 3′ 151R—5′ AAGCCAAGGTCGCTTTGAAC 3′
XF2083 czcA F—5′ GAATCGGTACGCCAACAGATG 3′ 151R—5′ TGTTGAGCGCAGCAAAGATG 3′
XF0874 metI F—5′ GAGCAACCACCTGGCAACTC 3′ 151R—5′ TAGGCGACGTCACCCAATC 3′
XF0133 copB F—5′ TCAGCGGTTGATCCTGCAA 3′ 151R—5′ GCGCTCCCAACCCACATAC 3′
XF0619 cutA1 F—5′ TTCTCGCGTATTGGTACAGGAA 3′ 141R—5′ TTCACATGCACAGCGTTGGT 3′
XF0620 dsbB or cutA2 F—5′ TTTGGAAAATCACCCGTCGTA 3′ 151R—5′ ACGGCTCTCACCGCTTTTAG 3′
XF1341 cutC F—5′ GGAGTGTTGCGTGCGTCTT 3′ 151R—5′ ATCCGCACTGACATCAATCG 3′
XF0910 petCa F—5′ TCCAGCCAGGTCAGCAGAAC 3′ 151R—5′ ACCAAAAAAGTCAACAACACTAGGAA 3′
1148 Appl Microbiol Biotechnol (2008) 77:1145–1157
ΔΔCt=ΔCt (sample)−ΔCt (calibrator). The calibrator isthe value obtained for one specific sample. Therefore, thefold increase is always obtained in relation to the specificcalibrator utilized. The values obtained for biofilm withoutCuSO4 were utilized as calibrator. The relative quantifica-tion was obtained by 2−ΔΔCt (Livak and Schmittgen 2001).Statistical analyses of fold change of resistance-associatedgenes transcripts were evaluated by ANOVA, and themeans were compared by the t test (P≤0.05) using theAssistat 7.3 beta software (http://assistat.sites.uol.com.br).
Results
Copper resistance level of planktonic and biofilm cells
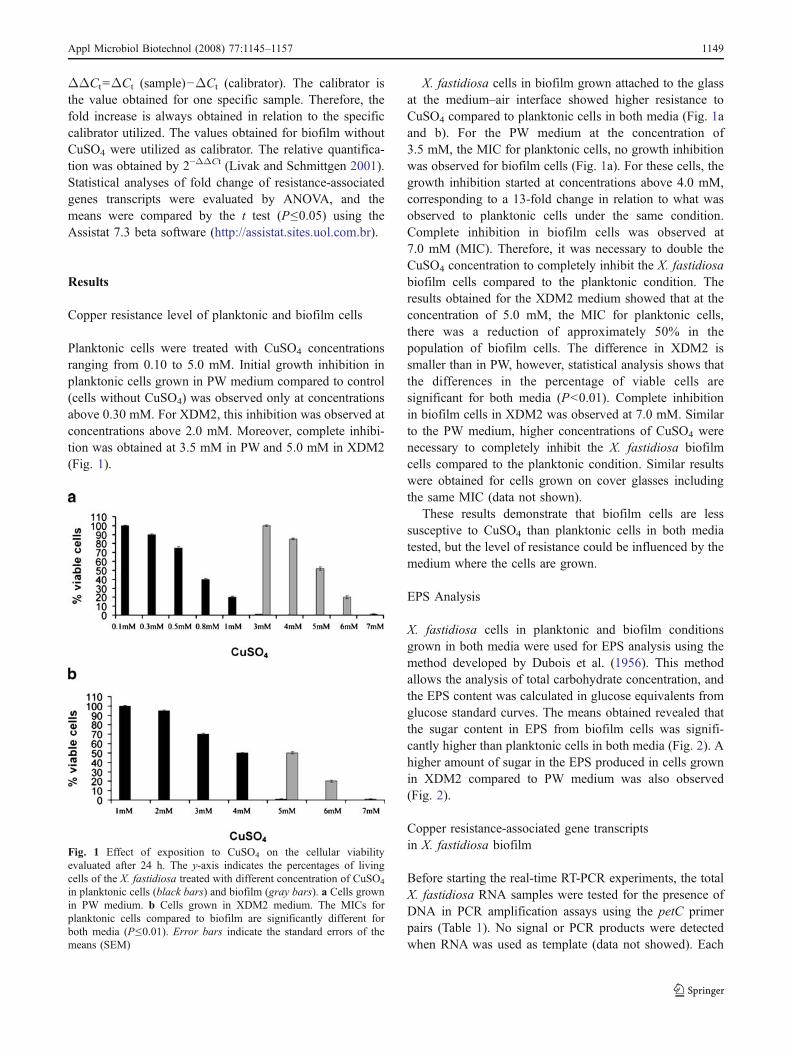
Planktonic cells were treated with CuSO4 concentrationsranging from 0.10 to 5.0 mM. Initial growth inhibition inplanktonic cells grown in PW medium compared to control(cells without CuSO4) was observed only at concentrationsabove 0.30 mM. For XDM2, this inhibition was observed atconcentrations above 2.0 mM. Moreover, complete inhibi-tion was obtained at 3.5 mM in PW and 5.0 mM in XDM2(Fig. 1).
X. fastidiosa cells in biofilm grown attached to the glassat the medium–air interface showed higher resistance toCuSO4 compared to planktonic cells in both media (Fig. 1aand b). For the PW medium at the concentration of3.5 mM, the MIC for planktonic cells, no growth inhibitionwas observed for biofilm cells (Fig. 1a). For these cells, thegrowth inhibition started at concentrations above 4.0 mM,corresponding to a 13-fold change in relation to what wasobserved to planktonic cells under the same condition.Complete inhibition in biofilm cells was observed at7.0 mM (MIC). Therefore, it was necessary to double theCuSO4 concentration to completely inhibit the X. fastidiosabiofilm cells compared to the planktonic condition. Theresults obtained for the XDM2 medium showed that at theconcentration of 5.0 mM, the MIC for planktonic cells,there was a reduction of approximately 50% in thepopulation of biofilm cells. The difference in XDM2 issmaller than in PW, however, statistical analysis shows thatthe differences in the percentage of viable cells aresignificant for both media (P<0.01). Complete inhibitionin biofilm cells in XDM2 was observed at 7.0 mM. Similarto the PW medium, higher concentrations of CuSO4 werenecessary to completely inhibit the X. fastidiosa biofilmcells compared to the planktonic condition. Similar resultswere obtained for cells grown on cover glasses includingthe same MIC (data not shown).
These results demonstrate that biofilm cells are lesssusceptive to CuSO4 than planktonic cells in both mediatested, but the level of resistance could be influenced by themedium where the cells are grown.
EPS Analysis
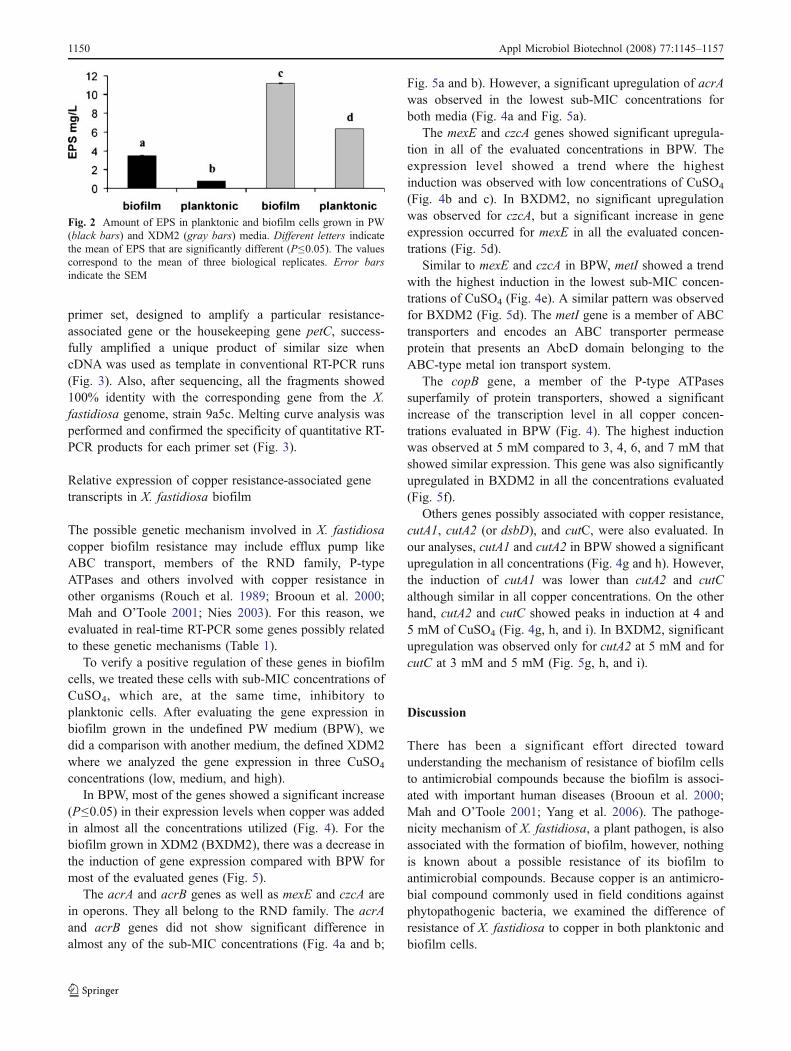
X. fastidiosa cells in planktonic and biofilm conditionsgrown in both media were used for EPS analysis using themethod developed by Dubois et al. (1956). This methodallows the analysis of total carbohydrate concentration, andthe EPS content was calculated in glucose equivalents fromglucose standard curves. The means obtained revealed thatthe sugar content in EPS from biofilm cells was signifi-cantly higher than planktonic cells in both media (Fig. 2). Ahigher amount of sugar in the EPS produced in cells grownin XDM2 compared to PW medium was also observed(Fig. 2).
Copper resistance-associated gene transcriptsin X. fastidiosa biofilm
Before starting the real-time RT-PCR experiments, the totalX. fastidiosa RNA samples were tested for the presence ofDNA in PCR amplification assays using the petC primerpairs (Table 1). No signal or PCR products were detectedwhen RNA was used as template (data not showed). Each
Fig. 1 Effect of exposition to CuSO4 on the cellular viabilityevaluated after 24 h. The y-axis indicates the percentages of livingcells of the X. fastidiosa treated with different concentration of CuSO4
in planktonic cells (black bars) and biofilm (gray bars). a Cells grownin PW medium. b Cells grown in XDM2 medium. The MICs forplanktonic cells compared to biofilm are significantly different forboth media (P≤0.01). Error bars indicate the standard errors of themeans (SEM)
Appl Microbiol Biotechnol (2008) 77:1145–1157 1149
primer set, designed to amplify a particular resistance-associated gene or the housekeeping gene petC, success-fully amplified a unique product of similar size whencDNA was used as template in conventional RT-PCR runs(Fig. 3). Also, after sequencing, all the fragments showed100% identity with the corresponding gene from the X.fastidiosa genome, strain 9a5c. Melting curve analysis wasperformed and confirmed the specificity of quantitative RT-PCR products for each primer set (Fig. 3).
Relative expression of copper resistance-associated genetranscripts in X. fastidiosa biofilm
The possible genetic mechanism involved in X. fastidiosacopper biofilm resistance may include efflux pump likeABC transport, members of the RND family, P-typeATPases and others involved with copper resistance inother organisms (Rouch et al. 1989; Brooun et al. 2000;Mah and O’Toole 2001; Nies 2003). For this reason, weevaluated in real-time RT-PCR some genes possibly relatedto these genetic mechanisms (Table 1).
To verify a positive regulation of these genes in biofilmcells, we treated these cells with sub-MIC concentrations ofCuSO4, which are, at the same time, inhibitory toplanktonic cells. After evaluating the gene expression inbiofilm grown in the undefined PW medium (BPW), wedid a comparison with another medium, the defined XDM2where we analyzed the gene expression in three CuSO4
concentrations (low, medium, and high).In BPW, most of the genes showed a significant increase
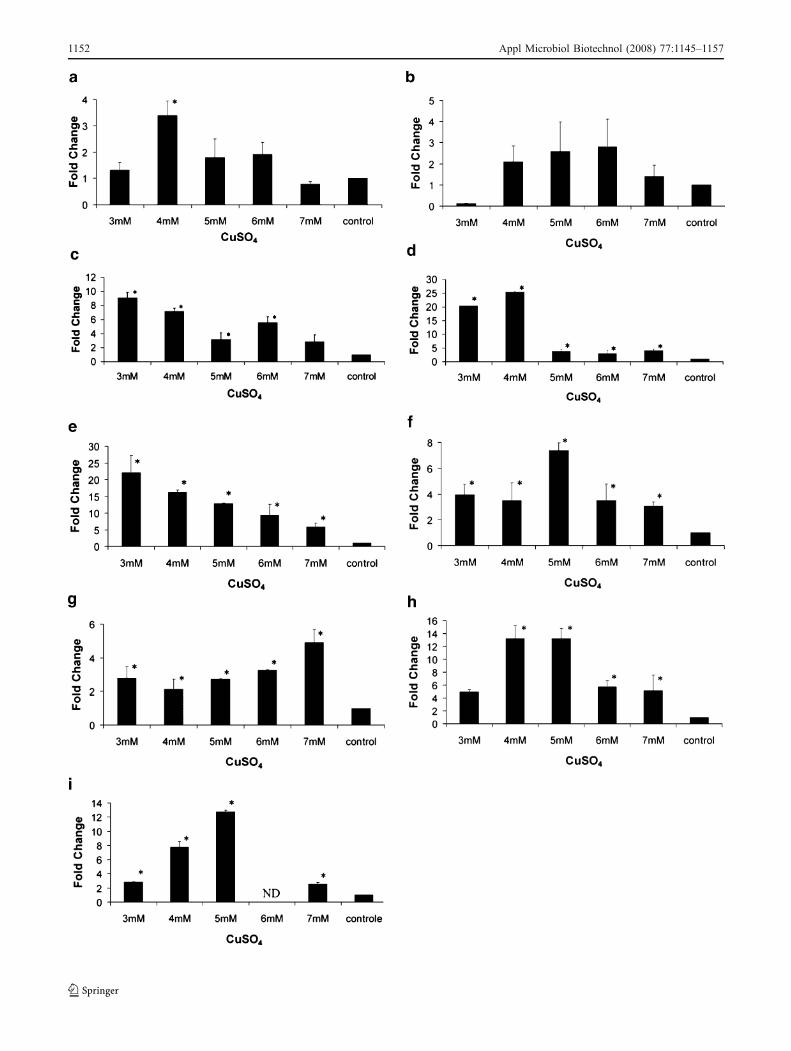
(P≤0.05) in their expression levels when copper was addedin almost all the concentrations utilized (Fig. 4). For thebiofilm grown in XDM2 (BXDM2), there was a decrease inthe induction of gene expression compared with BPW formost of the evaluated genes (Fig. 5).
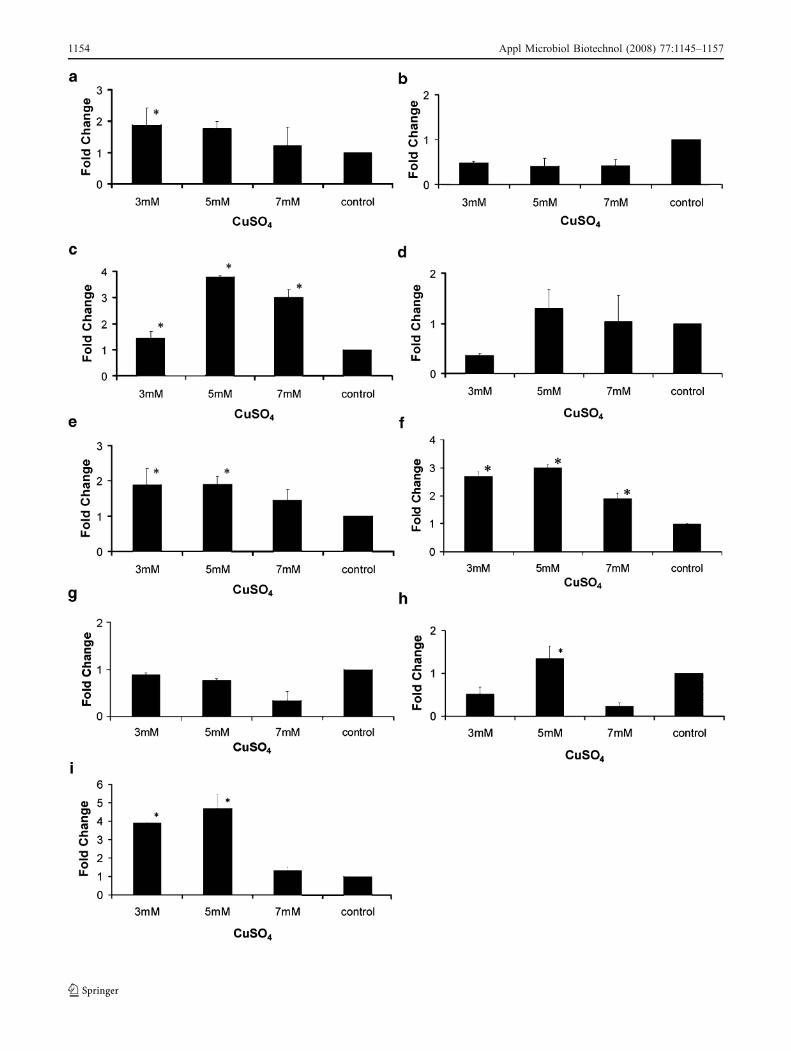
The acrA and acrB genes as well as mexE and czcA arein operons. They all belong to the RND family. The acrAand acrB genes did not show significant difference inalmost any of the sub-MIC concentrations (Fig. 4a and b;
Fig. 5a and b). However, a significant upregulation of acrAwas observed in the lowest sub-MIC concentrations forboth media (Fig. 4a and Fig. 5a).
The mexE and czcA genes showed significant upregula-tion in all of the evaluated concentrations in BPW. Theexpression level showed a trend where the highestinduction was observed with low concentrations of CuSO4
(Fig. 4b and c). In BXDM2, no significant upregulationwas observed for czcA, but a significant increase in geneexpression occurred for mexE in all the evaluated concen-trations (Fig. 5d).
Similar to mexE and czcA in BPW, metI showed a trendwith the highest induction in the lowest sub-MIC concen-trations of CuSO4 (Fig. 4e). A similar pattern was observedfor BXDM2 (Fig. 5d). The metI gene is a member of ABCtransporters and encodes an ABC transporter permeaseprotein that presents an AbcD domain belonging to theABC-type metal ion transport system.
The copB gene, a member of the P-type ATPasessuperfamily of protein transporters, showed a significantincrease of the transcription level in all copper concen-trations evaluated in BPW (Fig. 4). The highest inductionwas observed at 5 mM compared to 3, 4, 6, and 7 mM thatshowed similar expression. This gene was also significantlyupregulated in BXDM2 in all the concentrations evaluated(Fig. 5f).
Others genes possibly associated with copper resistance,cutA1, cutA2 (or dsbD), and cutC, were also evaluated. Inour analyses, cutA1 and cutA2 in BPW showed a significantupregulation in all concentrations (Fig. 4g and h). However,the induction of cutA1 was lower than cutA2 and cutCalthough similar in all copper concentrations. On the otherhand, cutA2 and cutC showed peaks in induction at 4 and5 mM of CuSO4 (Fig. 4g, h, and i). In BXDM2, significantupregulation was observed only for cutA2 at 5 mM and forcutC at 3 mM and 5 mM (Fig. 5g, h, and i).
Discussion
There has been a significant effort directed towardunderstanding the mechanism of resistance of biofilm cellsto antimicrobial compounds because the biofilm is associ-ated with important human diseases (Brooun et al. 2000;Mah and O’Toole 2001; Yang et al. 2006). The pathoge-nicity mechanism of X. fastidiosa, a plant pathogen, is alsoassociated with the formation of biofilm, however, nothingis known about a possible resistance of its biofilm toantimicrobial compounds. Because copper is an antimicro-bial compound commonly used in field conditions againstphytopathogenic bacteria, we examined the difference ofresistance of X. fastidiosa to copper in both planktonic andbiofilm cells.
Fig. 2 Amount of EPS in planktonic and biofilm cells grown in PW(black bars) and XDM2 (gray bars) media. Different letters indicatethe mean of EPS that are significantly different (P≤0.05). The valuescorrespond to the mean of three biological replicates. Error barsindicate the SEM
1150 Appl Microbiol Biotechnol (2008) 77:1145–1157
In our experimental conditions, planktonic cells grownin PW and XDM2 were more susceptible to copper thanthose grown in biofilm. However, the susceptibility ofplanktonic cells in XDM2 was lower than in PW. PW is anundefined medium that has different undefined componentsin its composition and has been used for growing all X.fastidiosa species (Davis et al. 1981). XDM2 is a defined
medium whose formulation was based in the X. fastidiosagenome annotation, and it is composed of glucose,vitamins, and amino acids, some of them are abundant inthe xylem sap.
It is possible that the lower susceptibility of planktoniccells in XDM2 compared with PW is due to the amount ofEPS produced when the cells are grown in XDM2. EPS can
Fig. 3 Validation of the primers utilized in real-time quantitativereverse transcription-polymerase chain reaction. We evaluated thegenes a acrA. b acrB. c czcA. d mexE. e metI. f copB. g cutA1.h cutA2. i cutC. j petC (endogenous control). The graphs illustrate themelting peak profiles for each gene. The amplicons were run inagarose gel (1.0%) electrophoresis. Lane L 1 Kb-plus DNA ladder
(Invitrogen); lanes 1, 2, 3, 4, and 5 amplification of cDNA frombiofilm cells incubated during 24 h with 3, 4, 5, 6, or 7 mM of CuSO4,respectively. Rn is the normalized reporter signal, which represents thefluorescence signal of the reporter dye divided by the fluorescencesignal of the passive reference dye
Appl Microbiol Biotechnol (2008) 77:1145–1157 1151
1152 Appl Microbiol Biotechnol (2008) 77:1145–1157

adsorb metal cations and, therefore, reduce the amount ofthese cations in the surrounding environment (Harrisonet al. 2006). Different components of the XDM2 mediumcould be responsible for the complexion of more cations incomparison to the components of PW. This could alsoaffect the different responses observed for the differentmedia.
Our results indicate that X. fastidiosa planktonic cellswere more susceptible to copper than cells in biofilm.These results were similar to what was found in the copperresistance of Pseudomonas aeruginosa biofilm where theminimal concentration necessary to completely kill thepopulation was 6 mM (Teitzel and Parsek 2003), while forX. fastidiosa, we observed a concentration of 7 mM. X.fastidiosa biofilm showed a much smaller increase inresistance to this metal in relation to planktonic cells,twofold, compared to P. aeruginosa where a >60-folddifference was observed. On the other hand, these authorsalso observed a small increase in resistance (twofold)when they evaluated its resistance to another metal, lead.In addition, planktonic cells when in high density showhigher resistance to antimicrobial compounds (Spoeringand Lewis 2001). No work has ever compared the X.fastidiosa biofilm and planktonic cells in relation toresistance to antimicrobial compounds. Because we useda high-density cell population in our experiments, wecould infer that it might have a higher resistance to thesecompounds, which could explain the small difference inresistance of biofilm and planktonic cells in both mediatested.
It has been shown that biofilms themselves are notsimply a diffusion barrier to antimicrobial compounds, butrather that bacteria within these microbial communitiesemploy distinct mechanisms to resist the action of antimi-crobial agents (Mah et al. 2003; Mukherjee et al. 2003).These mechanisms include cation/multidrug efflux of thecells (Nies 2003; Poole 2001, 2002). The genetics andbiochemistry of heavy metal resistance mechanisms havebeen carefully studied in free-swimming organisms butthere are not many studies concerning the participation of
these systems in biofilm cells (Teitzel and Parsek 2003). InX. fastidiosa, we investigated genes related to three effluxsystems: ABC transport, RND protein family, P-typeATPases, and other genes that could be related to copperresistance, homologs of cutA1, cutA2, and cutC from E. coli(Rouch and Brown 1997). Some of these genes werechosen for real-time RT-PCR analyses not only for theirpossible roles in copper resistance but also due to theexpression in X. fastidiosa biofilm cells obtained in PWmedium (De Souza et al. 2003, 2004).
For the RND efflux protein complex, we evaluated theexpression of four genes, acrA, acrB, mexE, and czcA.This complex transports a broad range of structurallyunrelated molecules to the exterior of the cells (Poole2004). It has recently been verified that TolC, the outermembrane protein component of the RND family ofMDR, is functional and necessary for pathogenicity and,probably, in planta survival of X. fastidiosa (Reddy et al.2007). Phenolic compounds also induce the expression ofan acrB homologue in Erwinia chrysanthemi. Therefore,successful phytopathogen can co-opt plant-signalingchemicals to induce enhanced tolerance to toxic plantantimicrobial chemicals (Ravirala et al. 2007). In X.fastidiosa, the MFP-encoding genes, acrA and mexE,show the conserved protein domain of acriflavin resistanceprotein A (AcrA). On the other hand, acrB and czcA showthe conserved protein domain of acriflavin resistanceprotein B (AcrB), an integral membrane protein. AcrBcooperates with AcrA and an outer membrane channelTolC to extrude the target molecule. Our work shows thatthe presence of copper leads to a strong upregulation inBPW mainly in mexE/czcA. For BXDM2, only mexE wassignificantly induced by copper, however, a similar trendwas observed for czcA.
The other gene analyzed was metI, which encodes aprotein belonging to the ATP-binding cassette (ABC)transporters family and shows an AbcD domain. The ABCtransporters related with antimicrobial compound effluxhave been identified in several organisms (Pattery et al.1999). However, the role of ABC transporters in biofilmresistance to antimicrobial compounds was only recentlyinvestigated in Candida albicans where biofilm cellsshowed upregulation of ABC transporters in presence ofthe antifungal fluconazole (Ramage et al. 2002). In X.fastidiosa, metI was upregulated in all evaluated concen-trations of copper, however, a higher induction wasobserved in the lowest sub-MIC concentrations.
The cation translocating P-type ATPases are amongstthe more common pumping mechanisms used to exportmetal ions out of cells (Dameron and Harrison 1998). InX. fastidiosa, we evaluated copB that encodes a P-typeATPase. In our experiment, the expression of this genewas increased at 5 mM of CuSO4. The expression of this
�Fig. 4 Relative quantitation of resistance-associated genes by real-time quantitative polymerase chain reaction. cDNA were preparedfrom biofilm cells grown in the undefined PW medium (BPW) andincubated during 24 h with 3, 4, 5, 6, 7 mM, or without (control)CuSO4. The samples were used for quantitation in the ABI PRISM7000 Sequence Detector System (Applied Biosystems). The measureswere normalized using the threshold cycles (Cts) obtained for theamplifications of the endogenous control run in the same plate. Thevalues represent the fold increase in gene expression compared withvalues obtained for cDNA from biofilm without CuSO4 (calibrator).The results are averages of three biological repetitions. a acrA. b acrB.c mexE. d czcA. e metI. f copB. g cutA1. h cutA2. i cutC. Asteriskindicates significant difference (P≤0.05) between the mean valuescompared to the control. Error bars indicate the SEM
Appl Microbiol Biotechnol (2008) 77:1145–1157 1153
1154 Appl Microbiol Biotechnol (2008) 77:1145–1157

gene in X. fastidiosa biofilm is only observed in thepresence of copper, suggesting that, as in other organisms,this gene is involved with copper homeostasis and couldbe upregulated in biofilm cells. The increase in geneexpression is similar to Enterococcus hirae where copBhas a function of expelling excess copper (Ordermatt et al.1994).
In E. coli, other genes possibly involved in the uptake,intracellular storage, delivery, and efflux of copper arecutABCDEF. The association of these genes in copperhomeostasis is made due to the results obtained in apreliminary characterization of copper-sensitive mutants.However, few of these genes have been directly linked tocopper metabolism, transport, or regulation and most of thecut genes are probably only indirectly involved in copperhomeostasis (Rouch et al. 1989). The cutA locus consists oftwo operons, one containing a single open reading frame(ORF) encoding a cytoplasmic protein of 13 kDa (CutA1)and the other consisting of two genes encoding a 50-kDadisulfide isomerase (CutA2 or DsbD) and a 24-kDa(CutA3) inner membrane protein. Among the cut genessimilar to E. coli only cutA1, cutA2, and cutC were found inthe X. fastidiosa genome. These genes showed differentbehaviors in the presence of copper. Induction of cutA1 waslower than the other cut genes in BPW and no significantchange was observed in BXDM2. On the other hand, cutA2and cutC showed upregulation in both biofilm conditions.Other mechanisms that confer copper resistance in E. colilike the CueO and Cus systems were not found in X.fastidiosa.
The lower level of induction in gene expressionobserved in BXDM2 compared to PW could be explainedby the presence of a higher amount of EPS as mentionedbefore. Adsorption of cations by EPS could reduce theamount of copper that penetrates the cells and, therefore,less pumping activity would be necessary to eliminate thiscompound to the outside of the cell. Another possibilitywould be that some of these genes are already highlyexpressed in biofilm cells when grown in XDM2 without
copper. Therefore, the increase of gene expression inresponse to the addition of copper to the medium wouldnot be significant as observed in the experiments.
One of the characteristics associated with microbialbiofilms is their increased resistance to antimicrobialcompounds. However, little is currently known about thephenotypic and genotypic changes that occur during thetransition from planktonic to biofilm mode of growth,especially in phytopathogenic bacteria. These biofilmswere intrinsically resistant to copper independent of themedia where they were formed. Nonetheless, the level ofgene expression could vary according to the environmentwhere the biofilm is formed. We have shown that theresistance of the X. fastidiosa biofilm seems to beassociated with an array of different mechanisms leadingto resistance of the cells. These mechanisms could involveEPS production, extrusion of the metal through differentpumps, and a probable action of cut (copper homeostasisprotein) genes. The gene expression analyses indicate thatresistance to copper in X. fastidiosa is a complexphenomenon that cannot be explained by just onemechanism; it is instead multifactorial and may involvedifferent molecular mechanisms of resistance.
Another interesting observation was that at a concen-tration of 7 mM, which was the inhibitory concentrationfor cell growth in culture, we were able to purify goodquality RNA and verified the expression of the genesassociated with copper resistance in all the evaluatedrepetitions. This result suggests that copper can penetrate,but fail to kill the cells. Some hypothesis to explain thisfinding could be speculated, the cells could be in aresistant physiological state or a bacterial programmed celldeath is taking place, a recently described mechanism inbacteria related with survival in stress environment(Engelberg-Kulka et al. 2006). For this reason, there is adrastic reduction in the population and because of thefastidious character of the X. fastidiosa growth, the cellsthat remain alive are not sufficient for allowing growth inthe culture medium. On the other hand, in the host plant,after the action of the antimicrobial compound, the livingcells could recover and recolonize the environment. Thiscould represent an important strategy for the survival ofthe population and also make it very difficult forcontrolling when biofilm is formed. These hypothesesneed to be further investigated.
Acknowledgments This work was supported by research grantsfrom Fundação de Amparo à Pesquisa do Estado de São Paulo(FAPESP: 04/14576-2) and Conselho Nacional de DesenvolvimentoCientífico e Tecnológico (CNPq: 094167-2005-2). M.A.T., H.D.C-F.,M.A.M., and A.A.S. are recipients of a research fellowship fromCNPq.
�Fig. 5 Relative quantitation of resistance-associated genes by real-time quantitative polymerase chain reaction. cDNA were preparedfrom biofilm cells grown in the defined XDM2 medium (BXDM2)and incubated during 24 h with 3, 5, 7 mM, or without (control)CuSO4. The samples were used for the quantitation in the ABI PRISM7000 Sequence Detector System (Applied Biosystems). The measureswere normalized using the threshold cycles (Cts) obtained for theamplifications of the endogenous control run in the same plate. Thevalues represent the fold increase in gene expression compared withvalues obtained for cDNA from biofilm without CuSO4 (calibrator).The results are averages of three biological repetitions. a acrA. b acrB.c mexE. d czcA. e metI. f copB. g cutA1. h cutA2. i cutC. Asteriskindicates significant difference (P≤0.05) between the mean valuescompared to the control. Error bars indicate the SEM
Appl Microbiol Biotechnol (2008) 77:1145–1157 1155

References
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic AcidsRes 25:3389–3402
Brooun A, Liu S, Lewis K (2000) A dose–response study of antibioticresistance in Pseudomonas aeruginosa biofilms. AntimicrobAgents Chemother 44:640–646
Chang G (2003) Multidrug resistance ABC transporters. FEBS Lett27:102–105
Costerton JW, Lewandowski Z, Caldwell DE, Korber D, Lappin-ScottHM (1995) Microbial biofilms. Annu Rev Microbiol 49:711–745
Dameron CT, Harrison MD (1998) Mechanisms for protection againstcopper toxicity. Am J Clin Nutr 67:1091S–1097S
Davis MJ, French WJ, Schaad NW (1981) Axenic culture of thebacteria associated with phony disease of peach and plum scald.Curr Microbiol 5:311–316
De Souza AA, Takita MA, Coletta-Filho HD, Caldana C, GoldmanGH, Yanai GM, Muto NH, Costa de Oliveira R, Nunes LR,Machado MA (2003) Analysis of gene expression in two growthstates of Xylella fastidiosa and its relationship with pathogenicity.Mol Plant Microb Interact 16:867–875
De Souza AA, Takita MA, Coletta-Filho HD, Caldana C, Yanai GM,Muto NH, Costa De Oliveira R, Nunes LR, Machado MA (2004)Gene expression profile of the plant pathogen Xylella fastidiosaduring biofilm formation in vitro. FEMS Microbiol Lett237:341–353
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956)Colorimetric method for determination of sugars and relatedsubstances. Anal Chem 28:350–354
Engelberg-Kulka H, Amitai S, Kolodkin-Gal I, Hazan R (2006)Bacterial programmed cell death and multicellular behavior inbacteria. PLoS Genet 2:e135
Fernandes WB Jr (2003) Analysis of the world processed orangeindustry. Ph.D. thesis. University of Florida, Gainesville, FL,p 108
Harrison JJ, Rabiei M, Turner RJ, Badry EA, Sproule KM, Ceri H(2006) Metal resistance in Candida biofilmes. FEMS MicrobiolEcol 3:479–491
Lemos EG, Alves LM, Campanharo JC (2003) Genomics-baseddesign of defined growth media for the plant pathogen Xylellafastidiosa. FEMS Microbiol Lett 219:39–45
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expressiondata using real-time quantitative PCR and the 22DDCT method.Methods 25:402–408
Mah TF, Pitts B, Pellock B, Walker GC, Stewart PS, O’Toole GA(2003) A genetic basis for Pseudomonas aeruginosa biofilmantibiotic resistance. Nature 426:306–310
Mah TFC, O’Toole GA (2001) Mechanisms of biofilm resistance toantimicrobial agents. Trends Microbiol 9:34–39
Mukherjee PK, Chandra J, Kuhn DM, Ghannoum MA (2003)Mechanism of fluconazole resistance in Candida albicansbiofilms: phase-specific role of efflux pumps and membranesterols. Infect Immun 71:4333–4340
Nies DH (2003) Efflux-mediated heavy metal resistance inprokaryotes. FEMS Microbiol Rev 27:313–339
Ordermatt A, Krapf R, Solioz M (1994) Induction of the putativecopper ATPases, CopA and CopB, of Enterococcus hirae by Ag+and Cu2, and Ag+ extrusion by CopB. Biochem Biophys ResCommun 202:44–48
Osiro D, Colnago LA, Otoboni AMMB, Lemos EGM, Souza AA,Coletta-Filho HD, Machado MA (2004) A kinetic model forXylella fastidiosa adhesion, biofilm formation, and virulence.FEMS Microbiol Lett 236:313–318
Pattery T, Hernalsteens JP, De Greve H (1999) Identification andmolecular characterization of a novel Salmonella enteritidispathogenicity islet encoding an ABC transporter. Mol Microbiol33:791–805
Poole K (2001) Multidrug efflux pumps and antimicrobial resistancein Pseudomonas aeruginosa and related organisms. J MolMicrobiol Biotechnol 3:255–264
Poole K (2002) Mechanisms of bacterial biocide and antibioticresistance. J Appl Microbiol 92:55S–64S
Poole K (2004) Efflux-mediated multiresistance in gram-negativebacteria. Clin Microbiol Infect 10:12–26
Ramage G, Bachmann S, Patterson TF, Wickes BL, Lopez-Ribot JL(2002) Investigation of multidrug efflux pumps in relation tofluconazole resistance in Candida albicans biofilms. J Antimi-crob Chemother 49:973–980
Ravirala RS, Barabot RD, Wheeler DM, Reverchon S, Tatum O,Malouf J, Liu H, Pritchard L, Hedley PE, Birch PRJ, Toth IK,Payton P, San Francisco MJD (2007) Efflux pump geneexpression in Erwinia chrysanthemi is induced by exposure tophenolic acids. Mol Plant Microbe Interact 20:313–320
Reddy JD, Reddy SL, Hopkins DL, Gabriel DW (2007) TolC isrequired for pathogenicity of Xylella fastidiosa in Vitis viniferagrapevines. Mol Plant Microbe Interact 20:403–410
Rouch D, Camakaris J, Lee BTO (1989) Copper transport in E. coli.In: Hamer DH, Winge DR (eds) Metal ion homeostasis:molecular biology and chemistry. Liss, New York, pp 469–477
Rouch DA, Brown NL (1997) Copper-inducible transcriptionalregulation at two promoters in the Escherichia coli copperresistance determinant pco. Microbiology 143:1191–1202
Saier Jr. MH, Tam, R, Reizer, A, Reizer, J (1994) Two novelfamilies of bacterial membrane proteins concerned withnodulation, cell division and transport. J Mol MicrobiolBiotechnol 11:841–847
Schaad NW, Postnikova E, Lact G, Fatmi M, Chang CJ (2004)Xylella fastidiosa subspecies: X. fastidiosa subsp piercei, subsp.nov., X. fastidiosa subsp. multiplex subsp. nov., and X.fastidiosa subsp. pauca subsp. nov. Syst Appl Microbiol27:290–300
Simpson AJG, Reinach FC, Arruda P, Abreu FA, Acencio M,Alvarenga R, Alves LMC, Araya JE, Baia GS, Baptista CS,Barros MH, Bonaccorsi ED, Bordin S, Bové JM, Briones MRS,Bueno MRP, Camargo AA, Camargo LEA, Carraro DM, CarrerH, Colauto NB, Colombo C, Costa FF, Costa MCR, Costa-NetoCM, Coutinho LL, Cristofani M, Dias-Neto E, Docena C, El-Dorry H, Facincani AP, Ferreira AJS, Ferreira VCA, Ferro JA,Fraga JS, França SC, Franco MC, Frohme M, Furlan LR, GarnierM, Goldman GH, Goldman MHS, Gomes SL, Gruber A, Ho PL,Hoheisel JD, Junqueira ML, Kemper EL, Kitajima JP, KriegerJE, Kuramae EE, Laigret F, Lambais MR, Leite LCC, LemosEGM, Lemos MVF, Lopes SA, Lopes CR, Machado JA,Machado MA, Madeira AMBN, Madeira HMF, Marino CL,Marques MV, Martins EAL, Martins EMF, Matsukuma AY,Menck CFM, Miracca EC, Miyaki CY, Monteiro-Vitorello CB,Moon DH, Nagai MA, Nascimento ALTO, Netto LES, Nhani Jr,A, Nobrega FG, Nunes LR, Oliveira MA, De Oliveira MC, DeOliveira RC, Palmieri DA, Paris A, Peixoto BR, Pereira GAG,Pereira Jr HA, Pesquero JB, Quaggio RB, Roberto PG,Rodrigues V, Rosa AJ De M, De Rosa Jr, VE, De Sá RG,
1156 Appl Microbiol Biotechnol (2008) 77:1145–1157

Santelli RV, Sawasaki HE, Da Silva ACR, Da Silva AM, DaSilva FR, Silva Jr, WA, Da Silveira JF, Silvestri MLZ,Siqueira WJ, De Souza AA, De Souza AP, Terenzi MF, TruffiD, Tsai SM, Tsuhako MH, Vallada H, Van Sluys MA,Verjovski-Almeida S, Vettore AL, Zago MA, Zatz M,Meidanis J, Setubal JC (2000) The genome sequence of theplant pathogen Xylella fastidiosa. The Xylella fastidiosa consor-tium of the Organization for Nucleotide Sequencing andAnalysis. Nature 40:151–159
Spoering AL, Lewis K (2001) Biofilms and planktonic cells ofPseudomonas aeruginosa have similar resistance to killing byantimicrobials. J Bacteriol 183:6746–6751
Surdeau N, Laurent-Maquin D, Bouthors S, Gelle MP (2006)Sensitivity of bacterial biofilms and planktonic cells to a newantimicrobial agent, Oxsil 320N. J Hosp Infect 62:487–493
Teitzel GM, Parsek MR (2003) Heavy metal resistance of biofilmand planktonic Pseudomonas aeruginosa. Appl EnvironMicrobiol 69:2313–2320
Voloudakis AE, Reignier TM, Cooksey DA (2005) Regulation ofResistance to copper in Xanthomonas axonopodis pv. vesicatoria.Appl Environ Microbiol 71:782–789
Yang S, Lopez CR, Zechiedrich EL (2006) Quorum sensing andmultidrug transporters in Escherichia coli. Proc Natl Acad SciU S A 103:2386–2391
Appl Microbiol Biotechnol (2008) 77:1145–1157 1157
Related Documents











