doi: 10.1152/ajpgi.00422.2003 291:G297-G306, 2006. First published 6 April 2006; Am J Physiol Gastrointest Liver Physiol Heikinheimo and Theodore C. Simon Joyce K. Divine, Lora J. Staloch, Hanna Haveri, Christopher W. Rowley, Markku in activating the rat liver fatty acid binding protein gene Cooperative interactions among intestinal GATA factors You might find this additional info useful... 13 other HighWire-hosted articles: This article has been cited by http://ajpgi.physiology.org/content/291/2/G297#cited-by including high resolution figures, can be found at: Updated information and services http://ajpgi.physiology.org/content/291/2/G297.full can be found at: AJP - Gastrointestinal and Liver Physiology about Additional material and information http://www.the-aps.org/publications/ajpgi This information is current as of July 26, 2016. website at http://www.the-aps.org/. 20814-3991. Copyright © 2006 the American Physiological Society. ISSN: 0193-1857, ESSN: 1522-1547. Visit our published 12 times a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD involving normal or abnormal function of the gastrointestinal tract, hepatobiliary system, and pancreas. It is publishes original articles pertaining to all aspects of research AJP - Gastrointestinal and Liver Physiology by guest on July 26, 2016 http://ajpgi.physiology.org/ Downloaded from by guest on July 26, 2016 http://ajpgi.physiology.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi: 10.1152/ajpgi.00422.2003291:G297-G306, 2006. First published 6 April 2006;Am J Physiol Gastrointest Liver Physiol
Heikinheimo and Theodore C. SimonJoyce K. Divine, Lora J. Staloch, Hanna Haveri, Christopher W. Rowley, Markkuin activating the rat liver fatty acid binding protein geneCooperative interactions among intestinal GATA factors
You might find this additional info useful...
13 other HighWire-hosted articles: This article has been cited by http://ajpgi.physiology.org/content/291/2/G297#cited-by
including high resolution figures, can be found at: Updated information and serviceshttp://ajpgi.physiology.org/content/291/2/G297.full
can be found at:AJP - Gastrointestinal and Liver Physiology about Additional material and information
http://www.the-aps.org/publications/ajpgi
This information is current as of July 26, 2016.
website at http://www.the-aps.org/. 20814-3991. Copyright © 2006 the American Physiological Society. ISSN: 0193-1857, ESSN: 1522-1547. Visit our published 12 times a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MDinvolving normal or abnormal function of the gastrointestinal tract, hepatobiliary system, and pancreas. It is
publishes original articles pertaining to all aspects of researchAJP - Gastrointestinal and Liver Physiology
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from

Cooperative interactions among intestinal GATA factors in activating the ratliver fatty acid binding protein gene
Joyce K. Divine,1,2 Lora J. Staloch,2 Hanna Haveri,3
Christopher W. Rowley,2 Markku Heikinheimo,3 and Theodore C. Simon2
1Division of Biology and Biomedical Sciences and 2Department of Pediatrics, Washington UniversitySchool of Medicine, St. Louis, Missouri; and 3Children’s Hospital and Program for Developmentaland Reproductive Biology, Biomedicum Helsinki, University of Helsinki, Helsinki, Finland
Submitted 25 September 2003; accepted in final form 21 February 2006
Divine, Joyce K., Lora J. Staloch, Hanna Haveri, ChristopherW. Rowley, Markku Heikinheimo, and Theodore C. Simon. Co-operative interactions among intestinal GATA factors in activating therat liver fatty acid binding protein gene. Am J Physiol GastrointestLiver Physiol 291: G297–G306, 2006. First published April 6, 2006;doi:10.1152/ajpgi.00422.2003.—GATA-4, GATA-5, and GATA-6are endodermal zinc-finger transcription factors that activate numer-ous enterocytic genes. GATA-4 and GATA-6 but not GATA-5 arepresent in adult murine small intestinal enterocytes, and we nowreport the simultaneous presence of all three GATA factors in murinesmall intestinal enterocytes before weaning age. An immunohisto-chemical survey detected enterocytic GATA-4 and GATA-6 at birthand 1 wk of age and GATA-5 at 1 wk but not birth. Interactionsamong GATA factors were explored utilizing a transgene constructedfrom the proximal promoter of the rat liver fatty acid binding proteingene (Fabp1). GATA-4 and GATA-5 but not GATA-6 activate theFabp1 transgene through a cognate binding site at �128. A dose-response assay revealed a maximum in transgene activation by bothfactors, where additional factor did not further increase transgeneactivity. However, at saturated levels of GATA-4, additional trans-gene activation was achieved by adding GATA-5 expression con-struct, and vice versa. Similar cooperativity occurred with GATA-5and GATA-6. Identical interactions were observed with a targettransgene consisting of a single GATA site upstream of a minimalpromoter. Furthermore, GATA-4 and GATA-5 or GATA-5 andGATA-6 bound to each other in solution. These results are consistentwith tethering of one GATA factor to the Fabp1 promoter throughinteraction with a second GATA factor to produce increased targetgene activation. Cooperative target gene activation was specific to anintestinal cell line and may represent a mechanism by which genes areactivated in the small intestinal epithelium during the period beforeweaning.
fabp1; GATA-4; GATA-5; GATA-6; suckling-weaning transition
THE SIX MEMBERS OF THE GATA transcription factor family shareconserved zinc-finger DNA-binding domains that recognizethe same DNA sequence, WGATAR (34). GATA factors bindto DNA as monomers, and only one GATA factor occupies thecognate DNA binding site at a time (21). GATA-1, GATA-2,and GATA-3 are found in hematopoietic cells, whereasGATA-4, GATA-5, and GATA-6 are found in endodermic andmesodermic tissues, including the intestine. Numerous genesexpressed in the intestinal epithelium are activated in culturedintestinal epithelial cell lines by GATA-4 (4, 12, 16, 20, 45),GATA-5 (12, 16, 20, 27, 30), and GATA-6 (1, 16, 18, 20). We
previously performed an immunohistochemical survey forGATA factors in the adult intestine and found that GATA-4and GATA-6 are expressed in adult murine small intestinalenterocytes, with GATA-6 also present in crypt epithelial cells(12). We did not detect GATA-5 in the adult murine intestine(12), although the Gata5 transcript has been detected in theintestine of both late gestation and adult mice (14, 36).
The presence in a single cell of multiple GATA factors thatrecognize the same DNA binding site raises the possibility thatinteractions between GATA factors may occur in transactivat-ing target genes. Although all GATA factors recognize aWGATAR sequence with high affinity, some specificity ingene activation has been observed. GATA-4 is utilized prefer-entially over GATA-6 in the regulation of myosin �-chain and�-chain genes (7). This specificity has been attributed topreferential binding of individual GATA factors to a particularsite, perhaps through interaction with sequences flanking thecore GATA binding sequence (3, 7, 49). Interactions withcofactors or other transcription factors that bind nearby aGATA site may also mediate preferential binding or activity ofindividual GATA factors (4, 10, 12, 13, 30). GATA-4 issubject to phosphorylation (8, 31, 35, 46) or acetylation (26),which can increase DNA binding. Intestinal extracts showedonly GATA-4 but not GATA-6 binding to sequences contain-ing GATA binding sites despite the presence of both factors inthis tissue, suggesting regulation of binding (4, 14). Anotherpossible means of interaction between GATA factors present ina single cell is direct binding between factors. GATA-1 iscapable of a homotypic interaction that results in increasedtarget gene transcription (50) and is required for gene regula-tion in vivo (38). All GATA factors contain two highly con-served zinc fingers that mediate DNA binding. The homotypicGATA-1 interaction is achieved by binding of the moreCOOH-terminal zinc finger with the more NH2-terminal finger,allowing formation of multimeric complexes in the absence ofDNA (9, 33, 50), dependent on six lysine residues (38). Thehighly conserved nature of the zinc-finger domains among allthe GATA factors suggests that heterotypic interactions mayalso occur. Binding of GATA-4 to GATA-6 bound to themyosin heavy-chain promoter has been described (7). In thiscase, only GATA-6 bound the DNA, but GATA-4 was tetheredto the promoter through interaction with GATA-6, allowingGATA-4 to contribute to gene activation. These results suggestthat factor interactions may contribute to target gene regulation
Address for reprint requests and other correspondence: T. C. Simon, Wash-ington Univ. School of Medicine, Dept. of Pediatrics, Campus Box 8208, St.Louis, MO 63110 (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Am J Physiol Gastrointest Liver Physiol 291: G297–G306, 2006.First published April 6, 2006; doi:10.1152/ajpgi.00422.2003.
0193-1857/06 $8.00 Copyright © 2006 the American Physiological Societyhttp://www.ajpgi.org G297
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from

in tissues such as the intestinal epithelium where multipleGATA factors are present. We explored this possibility utiliz-ing the liver fatty acid binding protein gene (Fabp1) as amodel.
Fabp1 is abundantly expressed in small intestinal entero-cytes beginning at the time of epithelial cytodifferentiation atembryonic day 17 in rodents (39). Enterocytic Fabp1 expres-sion remains high throughout the preweaning period and adult-hood and is primarily regulated at the transcriptional level (2,22). We have utilized a transgene constructed from rat Fabp1nucleotides �596 to �21 to study enterocytic gene regulation(11). A transgene constructed of Fabp1 nucleotides �596 to�21 relative to the start site of transcription is active in allsmall intestinal epithelial cells, including enterocytes (41).There are three consensus GATA sites in the proximal Fabp1promoter, and GATA factors activate the Fabp1 transgene incultured cells through interaction with these sites (11).GATA-4 and GATA-5 activate the Fabp1 transgene in cul-tured cells through preferential interaction of the most proxi-mal of the three GATA binding sites present in the promoter(12). The potential interactions between pairs of endodermalGATA factors in activating the Fabp1 transgene were deter-mined in intestinal Caco-2 cells. We found that GATA-4,GATA-5, and GATA-6 were simultaneously present in thesmall intestinal epithelium of mice before weaning, that coop-erative functional interactions occurred between specific pairsof GATA factors, and that heterotypic and homotypic bindingoccurred among these factors.
MATERIALS AND METHODS
Immunohistochemistry. Approval was obtained from the Institu-tional Animal Care and Use Committee for all experiments involvinganimals. Immunohistochemistry for murine GATA-4, GATA-5, andGATA-6 was performed as previously described (12). The normalsmall intestinal tissue samples were harvested from FVB/N mice onpostnatal days 1, 7, 12, 15, 18, and 21. Briefly, tissues were fixed in4% paraformaldehyde and embedded in paraffin before sectioning.Primary antibodies were polyclonal goat anti-mouse GATA-4 IgG,polyclonal goat anti-mouse GATA-5 IgG, and polyclonal rabbit anti-mouse GATA-6 IgG (all from Santa Cruz Biotechnology), used atrespective dilutions of 1:200, 1:50, and 1:50. Nonimmune IgG wasused as the primary antibody for negative controls. Primary antibodieswere visualized with a commercial avidin-biotin immunoperoxidasesystem (Vectastain Elite ABC Kit, Vector Laboratories) and substrate3,3�-diaminobenzidine (Sigma). Staining for each sample was re-peated at least twice.
Plasmids. The construction of the Fabp1-transgene and GATA-expression plasmids has been described (11), as has construction ofthe plasmid containing a GATA site upstream of a minimal promoter(47). Expression plasmids for GATA:Renilla luciferase (luc) fusionproteins were constructed as follows. The Renilla luciferase codingsequence was amplified from pRL-TK (Promega) with two rounds ofPCR using reverse primer 5�-GATCCGAATTCTTATTGTTCAT-TTTTGAGAACTCGCTCAACGAA-3� with first forward primer5�-CGGCAGCAAGGGCCGCGACAGCAAGAAGGTGCGC-GCCGCCCGCACTTCGAAAGTTTATGATCCAGAACAAAGG-3�to produce an amplimer used as template for amplification with the samereverse primer and second forward primer 5�-AGGGCGAATTCGGTA-CCGCTACGTCTACGCTGCAGGGCGGCGGCGGCAGCAAGG-GCCGCGACAGCAAGAAGGT-3�. This amplimer was cut with EcoRIand inserted into pSG5 (Stratagene) cut with the same enzyme to createpTS453, which contains the Renilla luciferase in frame with an amino-terminal linker sequence GGGGSKGRDSKKVRAAR that contains PstI
and KpnI sites just upstream of the linker coding sequence. pTS453 wasdigested with PstI/KpnI, and amplimers containing the coding sequencefrom GATA-4, GATA-5, or GATA-6 were inserted to create pTS468(GATA-4:luc fusion), pTS469 (GATA-5:luc fusion), or pTS473 (GATA-6:luc fusion). The GATA-4 coding sequence amplimer was producedfrom pTS186 (11) with primers 5�-AATTCGGTACCGCCGCCAC-CATGTACCAAAGCCTGGCCATGGCCGCCAAC-3� and 5�-GCC-GCCCTGCAGCGCGGTGATTATGTCCCCATGACTGTCAGC-3�and digested with PstI and KpnI. The GATA-5 coding sequenceamplimer was produced from pTS218 (12) with primers 5�-AAT-TCGGTACCGCCGCCACCATGTACCAAAGCTTGGCGTTAGC-CCAGAGC-3� and 5�-CCGCCCTGCAGGGCCAAGGCCAGAGCA-CACCAGGTCTCCTG-3� and digested with PstI and KpnI. TheGATA-6 coding sequence amplimer was produced from pTS236 (12)with primers 5�-AATTCGGTACCGCCGCCACCATGTACCA-GACCCTCGCCGCCCTGTCCAGC-3� and 5�-CCGGCCCCTG-CAGGGCCAGGGCCAGAGCACACCAAGAATCCTG-3� and di-gested with SbfI and KpnI. Bacterial expression plasmids for productionof the glutathione-S-transferase (GST)/GATA fusion proteins were pro-duced by introducing the coding sequence for GATA-4 or GATA-5 intopGEX-2T (Amersham). The murine GATA-4 sequence was amplifiedfrom pTS186 (11) with primers incorporating a BamHI site at thefive-prime end (5�-GCACGGATCCTACCAAAGCCTGGCCATG-3�)and an EcoRI site at the three-prime end (5�-GCACGAATTCT-TACGCGGTGATTATGT-3�). The vector and PCR fragment were di-gested with both enzymes and ligated together to create plasmid pTS351.The murine GATA-5 sequence was amplified from pTS218 (11) withprimers incorporating an EcoRI site at the five-prime end (5�-GCAGAATTCCATACCAAAGCTTGGCGTTA-3�) and an EcoRI siteat the three-prime end (5�-CGACGAATTCCTAGGCCAAGGCCA-GAGC-3�). The vector and PCR fragment were digested with bothenzymes and ligated together to create plasmid pTS352. The absence oferrors in the GATA protein coding sequences in both plasmids wasverified by sequencing.
Cell lines and transfection assays. Caco-2 and HepG2 cells werefrom American Type Culture Collection and were maintained asrecommended, and HeLa cells were a kind gift from Alan Schwartz(Washington University). Transient transfections were performed bythe calcium phosphate precipitation method essentially as described inRef. 11. Briefly, the transfection mixtures contained an Fabp1 reporterplasmid, GATA factor expression plasmid(s), and pGL3 controlplasmid (Promega) that was included to normalize for differences intransfection efficiency. The total amount of plasmid DNA (5–9�g/well) was kept constant in a given experiment by addition of pSG5plasmid. Cells were seeded into six-well plates the day before trans-fection so that cell density at the time of transfection was 50%. Assayswere performed 48 h after transfection when the cells just reachedconfluence. Human growth hormone (hGH) transgene reporter wasdetected in the media using a specific radioimmunoassay (NicholsInstitute), whereas luciferase production from pGL3 control wasdetected with a commercial kit (Promega). Values were calculated asthe average of the three replicate wells for each DNA mixture, anderror was calculated as standard deviation. Values are reported as foldactivation over the activity of the native transgene with no addedtranscription factor expression plasmids. All experiments were re-peated at least twice with similar results.
Solution binding assays. Solution binding assays were performedas described with modifications (11). GST-GATA fusion proteinswere produced in BL21 (DE3) Escherichia coli, purified with gluta-thione resin, and dialyzed against a buffer containing 50 mM Tris(pH � 7.5), 150 mM NaCl, 100 �M ZnCl2, 0.3% Nonidet P-40, and1 mM dithiothreitol. 35S-labeled target proteins were synthesized frommammalian expression plasmids pTS186 (GATA-4), pTS218(GATA-5), and pTS236 (GATA-6) using the TNT T7 coupled reticu-locyte lysate kit (Promega). Binding assays were conducted by mixing4 �g of GST or GST-GATA fusion protein with 20 �l of each targetsynthesis reaction in a total volume of 400 �l binding buffer (20 mM
G298 INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from
HEPES, pH � 7.5; 100 mM KCl, 5 mM EDTA, 5 mM EGTA, and0.5% BSA). Binding was allowed to occur for 2 h at 4°C, then 20 �lglutathione Sepharose beads were added, and the reaction continuedovernight. The beads were then washed once in binding buffer andtwice with binding buffer without BSA. Proteins were eluted from thebeads by boiling in SDS sample buffer and separated by SDS-PAGE.Radiolabeled proteins were visualized with a phosphor capture system.
RESULTS
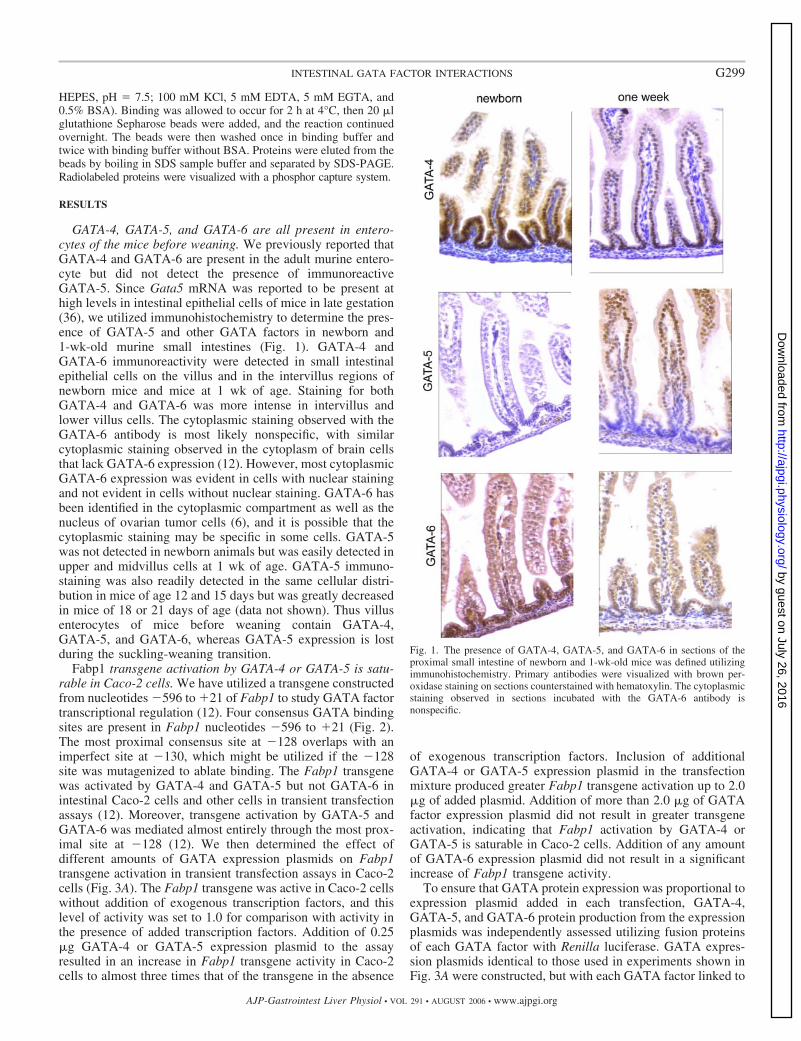
GATA-4, GATA-5, and GATA-6 are all present in entero-cytes of the mice before weaning. We previously reported thatGATA-4 and GATA-6 are present in the adult murine entero-cyte but did not detect the presence of immunoreactiveGATA-5. Since Gata5 mRNA was reported to be present athigh levels in intestinal epithelial cells of mice in late gestation(36), we utilized immunohistochemistry to determine the pres-ence of GATA-5 and other GATA factors in newborn and1-wk-old murine small intestines (Fig. 1). GATA-4 andGATA-6 immunoreactivity were detected in small intestinalepithelial cells on the villus and in the intervillus regions ofnewborn mice and mice at 1 wk of age. Staining for bothGATA-4 and GATA-6 was more intense in intervillus andlower villus cells. The cytoplasmic staining observed with theGATA-6 antibody is most likely nonspecific, with similarcytoplasmic staining observed in the cytoplasm of brain cellsthat lack GATA-6 expression (12). However, most cytoplasmicGATA-6 expression was evident in cells with nuclear stainingand not evident in cells without nuclear staining. GATA-6 hasbeen identified in the cytoplasmic compartment as well as thenucleus of ovarian tumor cells (6), and it is possible that thecytoplasmic staining may be specific in some cells. GATA-5was not detected in newborn animals but was easily detected inupper and midvillus cells at 1 wk of age. GATA-5 immuno-staining was also readily detected in the same cellular distri-bution in mice of age 12 and 15 days but was greatly decreasedin mice of 18 or 21 days of age (data not shown). Thus villusenterocytes of mice before weaning contain GATA-4,GATA-5, and GATA-6, whereas GATA-5 expression is lostduring the suckling-weaning transition.
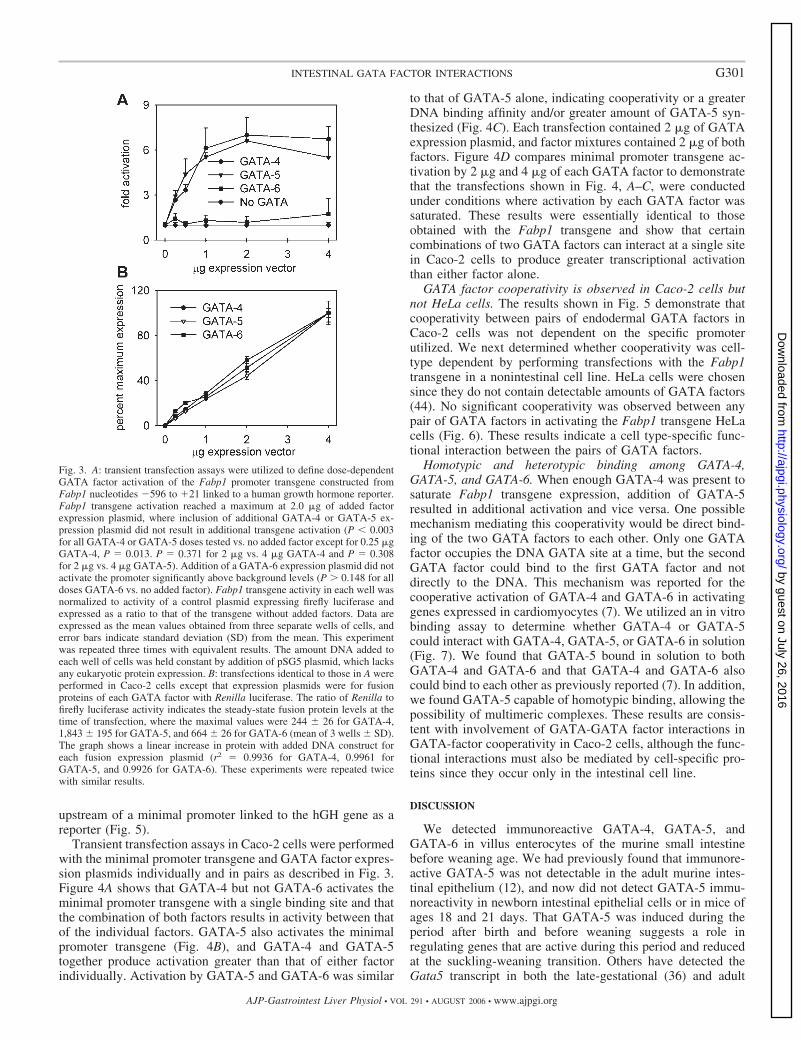
Fabp1 transgene activation by GATA-4 or GATA-5 is satu-rable in Caco-2 cells. We have utilized a transgene constructedfrom nucleotides �596 to �21 of Fabp1 to study GATA factortranscriptional regulation (12). Four consensus GATA bindingsites are present in Fabp1 nucleotides �596 to �21 (Fig. 2).The most proximal consensus site at �128 overlaps with animperfect site at �130, which might be utilized if the �128site was mutagenized to ablate binding. The Fabp1 transgenewas activated by GATA-4 and GATA-5 but not GATA-6 inintestinal Caco-2 cells and other cells in transient transfectionassays (12). Moreover, transgene activation by GATA-5 andGATA-6 was mediated almost entirely through the most prox-imal site at �128 (12). We then determined the effect ofdifferent amounts of GATA expression plasmids on Fabp1transgene activation in transient transfection assays in Caco-2cells (Fig. 3A). The Fabp1 transgene was active in Caco-2 cellswithout addition of exogenous transcription factors, and thislevel of activity was set to 1.0 for comparison with activity inthe presence of added transcription factors. Addition of 0.25�g GATA-4 or GATA-5 expression plasmid to the assayresulted in an increase in Fabp1 transgene activity in Caco-2cells to almost three times that of the transgene in the absence
of exogenous transcription factors. Inclusion of additionalGATA-4 or GATA-5 expression plasmid in the transfectionmixture produced greater Fabp1 transgene activation up to 2.0�g of added plasmid. Addition of more than 2.0 �g of GATAfactor expression plasmid did not result in greater transgeneactivation, indicating that Fabp1 activation by GATA-4 orGATA-5 is saturable in Caco-2 cells. Addition of any amountof GATA-6 expression plasmid did not result in a significantincrease of Fabp1 transgene activity.
To ensure that GATA protein expression was proportional toexpression plasmid added in each transfection, GATA-4,GATA-5, and GATA-6 protein production from the expressionplasmids was independently assessed utilizing fusion proteinsof each GATA factor with Renilla luciferase. GATA expres-sion plasmids identical to those used in experiments shown inFig. 3A were constructed, but with each GATA factor linked to
Fig. 1. The presence of GATA-4, GATA-5, and GATA-6 in sections of theproximal small intestine of newborn and 1-wk-old mice was defined utilizingimmunohistochemistry. Primary antibodies were visualized with brown per-oxidase staining on sections counterstained with hematoxylin. The cytoplasmicstaining observed in sections incubated with the GATA-6 antibody isnonspecific.
G299INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from

Renilla luciferase with a flexible 17-amino-acid linker. TheGATA-4 and GATA-5 but not GATA-6 fusion proteins acti-vated the Fabp1 transgene but with less activity than the nativefactors (22.9% for GATA-4 and 80.8% for GATA-5 at the 1�g level). Steady-state fusion protein levels increased propor-tionally with added dosage of expression plasmid as deter-mined by Renilla luciferase activity (Fig. 3B), suggesting thatthe plateaus of transgene activation and lack of GATA-6activity are not the result of limiting protein synthesis. Thetotal amount of plasmid DNA added to each well of cells ineach transfection experiment was kept constant by addition ofpSG5 plasmid, which lacks any eukaryotic protein expression.
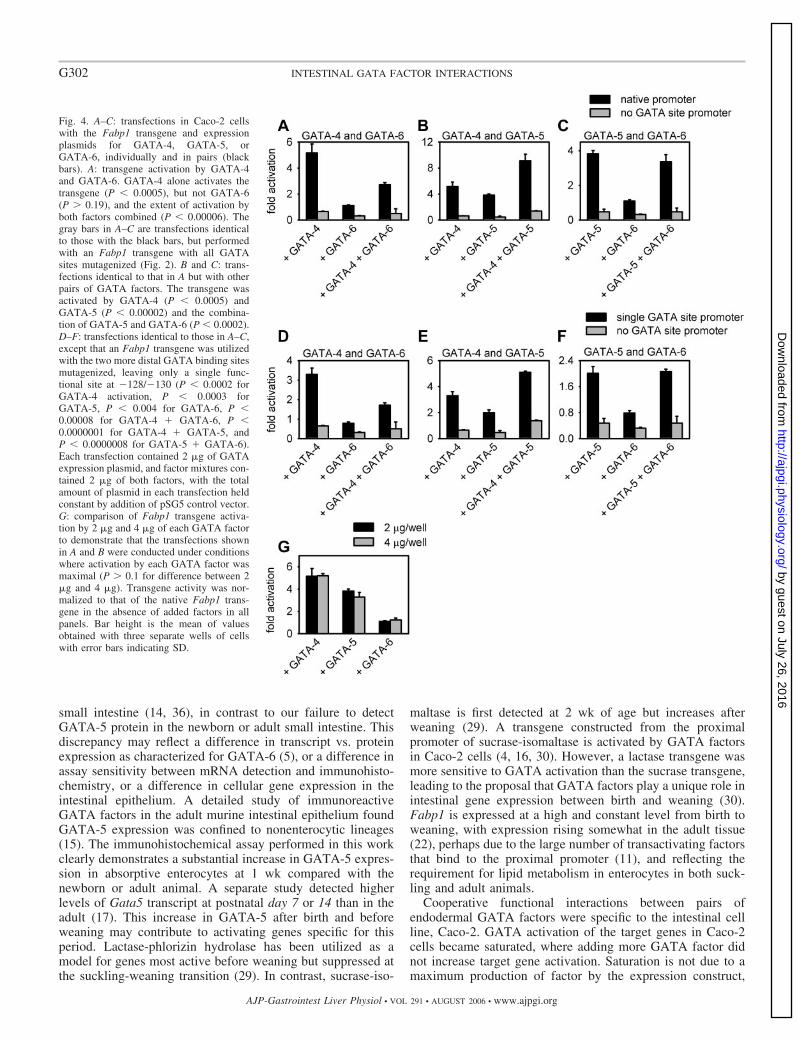
GATA factors exhibit cooperative activation of the Fabp1transgene in Caco-2 cells. GATA-4 and GATA-5 but notGATA-6 activate the Fabp1 transgene in Caco-2 cells. SinceGATA-4, GATA-5, and GATA-6 are present in overlappingcell populations in the intestinal epithelium, we explored po-tential interactions among GATA factors in Fabp1 transgeneactivation in Caco-2 cells (Fig. 4). Fabp1 transgene activationby pairs of GATA factors was compared with activation byindividual factors. Since only one GATA factor can bind toeach GATA site (21), activation by a pair of GATA factorstogether would be predicted to be between the activations ofthe two factors individually. This effect was observed withGATA-4 and GATA-6 (Fig. 3A), where Fabp1 transgeneactivation by the two factors in combination lies between thevalues obtained with each factor individually. However, adifferent result was obtained with the combination of GATA-4and GATA-5 or GATA-5 and GATA-6. These combinations ofendodermal GATA factors resulted in a significantly greatertransgene activation than predicted by competitive interaction,although the interaction of GATA-5 and GATA-6 could be dueto the greater amount of GATA-5 present or a greater bindingaffinity of GATA-5 than GATA-6. These factor interactionswere mediated through the three Fabp1 GATA binding sites,since a transgene with all these sites mutagenized was notactivated by any individual GATA factor or combination ofGATA factors (gray bars).
The cooperative activation observed with pairs of GATAfactors could be due to GATA factor binding at more than one
GATA site in the Fabp1 promoter, although we found thatindividual GATA factors activated the Fabp1 transgene onlythrough the most proximal site (12). This possibility was testedwith a transgene constructed from an Fabp1 promoter se-quence with the GATA sites at �228 and �556 inactivated bymutagenesis (Fig. 2). Figure 4, D–F, shows transfections per-formed with this transgene but otherwise identical to thoseshown in Fig. 4, A–C. Despite the presence of only a singlefunctional GATA site, cooperative Fabp1 transgene activationwas observed with GATA-4 and GATA-5 or GATA-5 andGATA-6, although interaction between GATA-5 and GATA-6might be due to increased quantity or greater DNA bindingaffinity of GATA-5 than GATA-6. The gray bars in Fig. 4,D–F, are transfections with an Fabp1 transgene with all GATAsites mutagenized, confirming that activation by individual orpaired GATA factors is dependent on the intact GATA bindingsite. The transfections shown in Fig. 4, A–F, were conductedwith saturating amounts of GATA factor expression plasmid,as verified by the comparison with additional wells of cells thatreceived higher amounts of expression plasmid during thesame experiment (Fig. 4G).
GATA factors exhibit cooperative activation of the syntheticminimal promoter transgene in Caco-2 cells. Cooperativeactivation of the Fabp1 transgene by GATA-4 and GATA-5 orGATA-5 and GATA-6 might be due to interactions with otherfactors present endogenously in Caco-2 cells and that bind tothe Fabp1 promoter. For example, we have shown thatGATA-4, GATA-5, and GATA-6 functionally and physicallyinteract with HNF-1� to activate the Fabp1 transgene, and thatFabp1 transgene activation by GATA family factors is reducedin Caco-2 cells when an HNF-1 binding site at �95 is mu-tagenized (12). We found that activation of the Fabp1 trans-gene with a mutagenized HNF-1 binding site by both GATA-4and GATA-5 was decreased compared with the native trans-gene, but the same relative degree of cooperation was observedbetween the two factors (data not shown). To determine po-tential effects of transcription factors besides HNF-1 familyfactors, we assessed cooperativity between GATA factorsutilizing a transgene (47) consisting of a single GATA site
Fig. 2. Rat liver fatty acid binding protein gene (Fabp1) nucleotides �596 to �21 are diagrammed with the location and sequence of the four GATA bindingsites indicated. The sequence of each GATA binding site is compared with the GATA factor matrix in the TRANSFAC database (48), with vertical linesindicating a matching nucleotide. The promoter is numbered relative to the start site of transcription (�1) with each GATA binding site designated by the mostproximal nucleotide of the consensus binding site as indicated. The �128 and �130 GATA binding overlap, allowing GATA factors to bind to one or the othersite but not both sites simultaneously. Mutations introduced to abrogate factor binding are underlined, with the substituted bases shown above the sequence.
G300 INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from
upstream of a minimal promoter linked to the hGH gene as areporter (Fig. 5).
Transient transfection assays in Caco-2 cells were performedwith the minimal promoter transgene and GATA factor expres-sion plasmids individually and in pairs as described in Fig. 3.Figure 4A shows that GATA-4 but not GATA-6 activates theminimal promoter transgene with a single binding site and thatthe combination of both factors results in activity between thatof the individual factors. GATA-5 also activates the minimalpromoter transgene (Fig. 4B), and GATA-4 and GATA-5together produce activation greater than that of either factorindividually. Activation by GATA-5 and GATA-6 was similar
to that of GATA-5 alone, indicating cooperativity or a greaterDNA binding affinity and/or greater amount of GATA-5 syn-thesized (Fig. 4C). Each transfection contained 2 �g of GATAexpression plasmid, and factor mixtures contained 2 �g of bothfactors. Figure 4D compares minimal promoter transgene ac-tivation by 2 �g and 4 �g of each GATA factor to demonstratethat the transfections shown in Fig. 4, A–C, were conductedunder conditions where activation by each GATA factor wassaturated. These results were essentially identical to thoseobtained with the Fabp1 transgene and show that certaincombinations of two GATA factors can interact at a single sitein Caco-2 cells to produce greater transcriptional activationthan either factor alone.
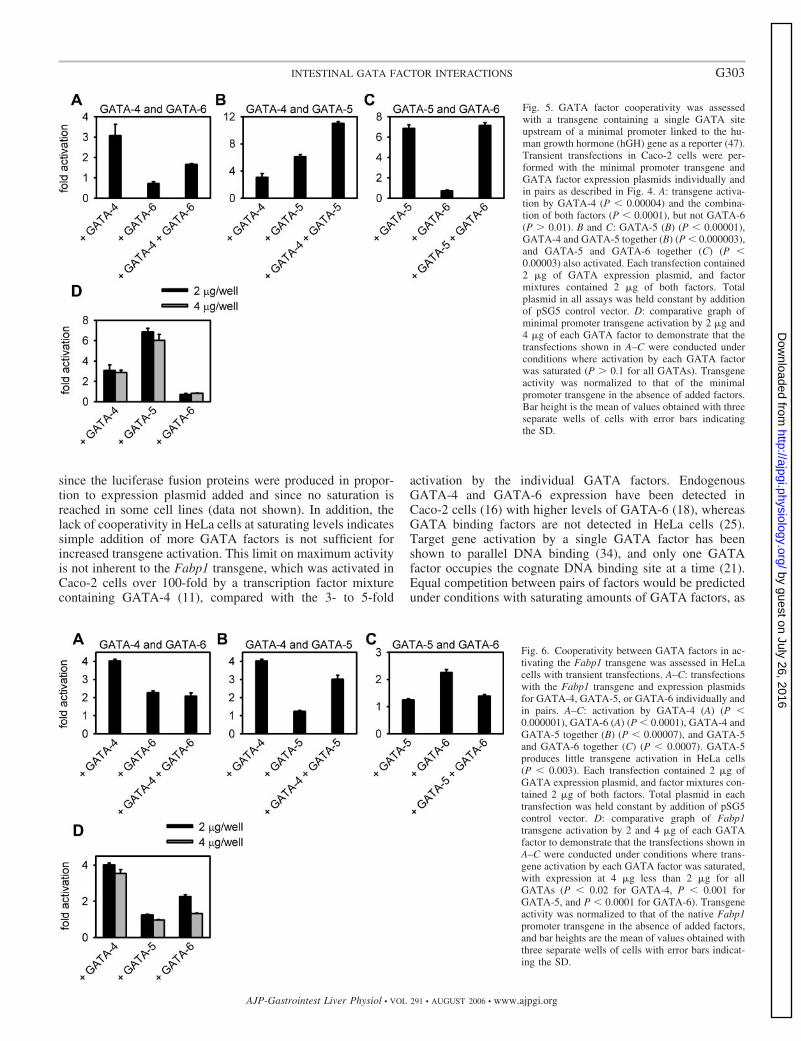
GATA factor cooperativity is observed in Caco-2 cells butnot HeLa cells. The results shown in Fig. 5 demonstrate thatcooperativity between pairs of endodermal GATA factors inCaco-2 cells was not dependent on the specific promoterutilized. We next determined whether cooperativity was cell-type dependent by performing transfections with the Fabp1transgene in a nonintestinal cell line. HeLa cells were chosensince they do not contain detectable amounts of GATA factors(44). No significant cooperativity was observed between anypair of GATA factors in activating the Fabp1 transgene HeLacells (Fig. 6). These results indicate a cell type-specific func-tional interaction between the pairs of GATA factors.
Homotypic and heterotypic binding among GATA-4,GATA-5, and GATA-6. When enough GATA-4 was present tosaturate Fabp1 transgene expression, addition of GATA-5resulted in additional activation and vice versa. One possiblemechanism mediating this cooperativity would be direct bind-ing of the two GATA factors to each other. Only one GATAfactor occupies the DNA GATA site at a time, but the secondGATA factor could bind to the first GATA factor and notdirectly to the DNA. This mechanism was reported for thecooperative activation of GATA-4 and GATA-6 in activatinggenes expressed in cardiomyocytes (7). We utilized an in vitrobinding assay to determine whether GATA-4 or GATA-5could interact with GATA-4, GATA-5, or GATA-6 in solution(Fig. 7). We found that GATA-5 bound in solution to bothGATA-4 and GATA-6 and that GATA-4 and GATA-6 alsocould bind to each other as previously reported (7). In addition,we found GATA-5 capable of homotypic binding, allowing thepossibility of multimeric complexes. These results are consis-tent with involvement of GATA-GATA factor interactions inGATA-factor cooperativity in Caco-2 cells, although the func-tional interactions must also be mediated by cell-specific pro-teins since they occur only in the intestinal cell line.
DISCUSSION
We detected immunoreactive GATA-4, GATA-5, andGATA-6 in villus enterocytes of the murine small intestinebefore weaning age. We had previously found that immunore-active GATA-5 was not detectable in the adult murine intes-tinal epithelium (12), and now did not detect GATA-5 immu-noreactivity in newborn intestinal epithelial cells or in mice ofages 18 and 21 days. That GATA-5 was induced during theperiod after birth and before weaning suggests a role inregulating genes that are active during this period and reducedat the suckling-weaning transition. Others have detected theGata5 transcript in both the late-gestational (36) and adult
Fig. 3. A: transient transfection assays were utilized to define dose-dependentGATA factor activation of the Fabp1 promoter transgene constructed fromFabp1 nucleotides �596 to �21 linked to a human growth hormone reporter.Fabp1 transgene activation reached a maximum at 2.0 �g of added factorexpression plasmid, where inclusion of additional GATA-4 or GATA-5 ex-pression plasmid did not result in additional transgene activation (P � 0.003for all GATA-4 or GATA-5 doses tested vs. no added factor except for 0.25 �gGATA-4, P � 0.013. P � 0.371 for 2 �g vs. 4 �g GATA-4 and P � 0.308for 2 �g vs. 4 �g GATA-5). Addition of a GATA-6 expression plasmid did notactivate the promoter significantly above background levels (P 0.148 for alldoses GATA-6 vs. no added factor). Fabp1 transgene activity in each well wasnormalized to activity of a control plasmid expressing firefly luciferase andexpressed as a ratio to that of the transgene without added factors. Data areexpressed as the mean values obtained from three separate wells of cells, anderror bars indicate standard deviation (SD) from the mean. This experimentwas repeated three times with equivalent results. The amount DNA added toeach well of cells was held constant by addition of pSG5 plasmid, which lacksany eukaryotic protein expression. B: transfections identical to those in A wereperformed in Caco-2 cells except that expression plasmids were for fusionproteins of each GATA factor with Renilla luciferase. The ratio of Renilla tofirefly luciferase activity indicates the steady-state fusion protein levels at thetime of transfection, where the maximal values were 244 26 for GATA-4,1,843 195 for GATA-5, and 664 26 for GATA-6 (mean of 3 wells SD).The graph shows a linear increase in protein with added DNA construct foreach fusion expression plasmid (r2 � 0.9936 for GATA-4, 0.9961 forGATA-5, and 0.9926 for GATA-6). These experiments were repeated twicewith similar results.
G301INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from
small intestine (14, 36), in contrast to our failure to detectGATA-5 protein in the newborn or adult small intestine. Thisdiscrepancy may reflect a difference in transcript vs. proteinexpression as characterized for GATA-6 (5), or a difference inassay sensitivity between mRNA detection and immunohisto-chemistry, or a difference in cellular gene expression in theintestinal epithelium. A detailed study of immunoreactiveGATA factors in the adult murine intestinal epithelium foundGATA-5 expression was confined to nonenterocytic lineages(15). The immunohistochemical assay performed in this workclearly demonstrates a substantial increase in GATA-5 expres-sion in absorptive enterocytes at 1 wk compared with thenewborn or adult animal. A separate study detected higherlevels of Gata5 transcript at postnatal day 7 or 14 than in theadult (17). This increase in GATA-5 after birth and beforeweaning may contribute to activating genes specific for thisperiod. Lactase-phlorizin hydrolase has been utilized as amodel for genes most active before weaning but suppressed atthe suckling-weaning transition (29). In contrast, sucrase-iso-
maltase is first detected at 2 wk of age but increases afterweaning (29). A transgene constructed from the proximalpromoter of sucrase-isomaltase is activated by GATA factorsin Caco-2 cells (4, 16, 30). However, a lactase transgene wasmore sensitive to GATA activation than the sucrase transgene,leading to the proposal that GATA factors play a unique role inintestinal gene expression between birth and weaning (30).Fabp1 is expressed at a high and constant level from birth toweaning, with expression rising somewhat in the adult tissue(22), perhaps due to the large number of transactivating factorsthat bind to the proximal promoter (11), and reflecting therequirement for lipid metabolism in enterocytes in both suck-ling and adult animals.
Cooperative functional interactions between pairs ofendodermal GATA factors were specific to the intestinal cellline, Caco-2. GATA activation of the target genes in Caco-2cells became saturated, where adding more GATA factor didnot increase target gene activation. Saturation is not due to amaximum production of factor by the expression construct,
Fig. 4. A–C: transfections in Caco-2 cellswith the Fabp1 transgene and expressionplasmids for GATA-4, GATA-5, orGATA-6, individually and in pairs (blackbars). A: transgene activation by GATA-4and GATA-6. GATA-4 alone activates thetransgene (P � 0.0005), but not GATA-6(P 0.19), and the extent of activation byboth factors combined (P � 0.00006). Thegray bars in A–C are transfections identicalto those with the black bars, but performedwith an Fabp1 transgene with all GATAsites mutagenized (Fig. 2). B and C: trans-fections identical to that in A but with otherpairs of GATA factors. The transgene wasactivated by GATA-4 (P � 0.0005) andGATA-5 (P � 0.00002) and the combina-tion of GATA-5 and GATA-6 (P � 0.0002).D–F: transfections identical to those in A–C,except that an Fabp1 transgene was utilizedwith the two more distal GATA binding sitesmutagenized, leaving only a single func-tional site at �128/�130 (P � 0.0002 forGATA-4 activation, P � 0.0003 forGATA-5, P � 0.004 for GATA-6, P �0.00008 for GATA-4 � GATA-6, P �0.0000001 for GATA-4 � GATA-5, andP � 0.0000008 for GATA-5 � GATA-6).Each transfection contained 2 �g of GATAexpression plasmid, and factor mixtures con-tained 2 �g of both factors, with the totalamount of plasmid in each transfection heldconstant by addition of pSG5 control vector.G: comparison of Fabp1 transgene activa-tion by 2 �g and 4 �g of each GATA factorto demonstrate that the transfections shownin A and B were conducted under conditionswhere activation by each GATA factor wasmaximal (P 0.1 for difference between 2�g and 4 �g). Transgene activity was nor-malized to that of the native Fabp1 trans-gene in the absence of added factors in allpanels. Bar height is the mean of valuesobtained with three separate wells of cellswith error bars indicating SD.
G302 INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from
since the luciferase fusion proteins were produced in propor-tion to expression plasmid added and since no saturation isreached in some cell lines (data not shown). In addition, thelack of cooperativity in HeLa cells at saturating levels indicatessimple addition of more GATA factors is not sufficient forincreased transgene activation. This limit on maximum activityis not inherent to the Fabp1 transgene, which was activated inCaco-2 cells over 100-fold by a transcription factor mixturecontaining GATA-4 (11), compared with the 3- to 5-fold
activation by the individual GATA factors. EndogenousGATA-4 and GATA-6 expression have been detected inCaco-2 cells (16) with higher levels of GATA-6 (18), whereasGATA binding factors are not detected in HeLa cells (25).Target gene activation by a single GATA factor has beenshown to parallel DNA binding (34), and only one GATAfactor occupies the cognate DNA binding site at a time (21).Equal competition between pairs of factors would be predictedunder conditions with saturating amounts of GATA factors, as
Fig. 5. GATA factor cooperativity was assessedwith a transgene containing a single GATA siteupstream of a minimal promoter linked to the hu-man growth hormone (hGH) gene as a reporter (47).Transient transfections in Caco-2 cells were per-formed with the minimal promoter transgene andGATA factor expression plasmids individually andin pairs as described in Fig. 4. A: transgene activa-tion by GATA-4 (P � 0.00004) and the combina-tion of both factors (P � 0.0001), but not GATA-6(P 0.01). B and C: GATA-5 (B) (P � 0.00001),GATA-4 and GATA-5 together (B) (P � 0.000003),and GATA-5 and GATA-6 together (C) (P �0.00003) also activated. Each transfection contained2 �g of GATA expression plasmid, and factormixtures contained 2 �g of both factors. Totalplasmid in all assays was held constant by additionof pSG5 control vector. D: comparative graph ofminimal promoter transgene activation by 2 �g and4 �g of each GATA factor to demonstrate that thetransfections shown in A–C were conducted underconditions where activation by each GATA factorwas saturated (P 0.1 for all GATAs). Transgeneactivity was normalized to that of the minimalpromoter transgene in the absence of added factors.Bar height is the mean of values obtained with threeseparate wells of cells with error bars indicatingthe SD.
Fig. 6. Cooperativity between GATA factors in ac-tivating the Fabp1 transgene was assessed in HeLacells with transient transfections. A–C: transfectionswith the Fabp1 transgene and expression plasmidsfor GATA-4, GATA-5, or GATA-6 individually andin pairs. A–C: activation by GATA-4 (A) (P �0.000001), GATA-6 (A) (P � 0.0001), GATA-4 andGATA-5 together (B) (P � 0.00007), and GATA-5and GATA-6 together (C) (P � 0.0007). GATA-5produces little transgene activation in HeLa cells(P � 0.003). Each transfection contained 2 �g ofGATA expression plasmid, and factor mixtures con-tained 2 �g of both factors. Total plasmid in eachtransfection was held constant by addition of pSG5control vector. D: comparative graph of Fabp1transgene activation by 2 and 4 �g of each GATAfactor to demonstrate that the transfections shown inA–C were conducted under conditions where trans-gene activation by each GATA factor was saturated,with expression at 4 �g less than 2 �g for allGATAs (P � 0.02 for GATA-4, P � 0.001 forGATA-5, and P � 0.0001 for GATA-6). Transgeneactivity was normalized to that of the native Fabp1promoter transgene in the absence of added factors,and bar heights are the mean of values obtained withthree separate wells of cells with error bars indicat-ing the SD.
G303INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from

we observed with GATA-4 and GATA-6. However, GATA-4and GATA-5 or GATA-5 and GATA-6 displayed a muchlarger degree of activation when combined than predicted bycompetition for a single site under saturating conditions. Analternative interpretation of the observed activation byGATA-5 and GATA-6 together is that increased synthesisand/or DNA binding of GATA-5 ameliorates any competitiveaction of GATA-6.
One mechanism for cooperativity between pairs of factors isheterotypic binding, where one factor tethers the other to theDNA, thus allowing both to contribute to gene activation. Thistype of interaction has been observed in cardiac cells forGATA-4 and GATA-6 (7). Homotypic interactions have alsobeen reported for GATA-1 (9, 50). Large complexes ofGATA-1 have been detected in the nucleus of GATA-1-expressing cells, and these same complexes containedGATA-2 or GATA-3 when these factors were also present(14). GATA-1 homotypic binding occurs through interactionsat the zinc fingers (33), and the zinc-finger motif has beenfound to have a general role in protein-protein interactions aswell as the DNA-binding function originally described in Ref.32. GATA-1 homotypic binding is dependent on lysine resi-dues conserved among GATA-1, GATA-2, and GATA-3, butthese residues are not present in GATA-4, GATA-5, orGATA-6 (38). We found that GATA-5 but not GATA-4 wascapable of homotypic binding, and heterotypic binding couldoccur between GATA-4 and GATA-5 or GATA-5 andGATA-6, as well as the previously described interaction be-tween GATA-4 and GATA-6. These results are consistent withextensive interactions between GATA factors independent ofDNA binding.
The existence of a maximum limit for GATA factor activa-tion suggests that additional proteins besides the GATA factorsare involved in target gene activation and that these accessoryproteins are limiting. The observed cooperativity also is con-sistent with a mechanism where each GATA factor actsthrough an independent accessory factor to activate the targetgene. Additional evidence for cell-specific factors critical forGATA factor transactivation of target genes is provided by theobservation that GATA family Fabp1 transgene activationdiffers between cell lines. GATA-4 or GATA-5 but notGATA-6 activates the Fabp1 transgene in Caco-2 cells,whereas GATA-4 or GATA-6 but not GATA-5 activates theFabp1 transgene in HeLa cells. Cell type-specific activation ofa lactase-phlorizin hydrolase transgene by GATA-5 has beenalso been observed (30). The cell type-specific cooperativityalso provides evidence for the presence of proteins mediatingGATA target gene activation. Endogenous GATA factors may
also influence transgene activation in the cell lines. HeLa cellshave no detectable GATA binding activity (30), whereasCaco-2 cells contain GATA-4, GATA-5, and GATA-6, withGATA-6 the predominant form (18). Despite the ubiquitousnature of in vitro binding between the GATA factors, weobserved cooperative functional interactions only betweenGATA-4 and GATA-5 or GATA-5 and GATA-6 in Caco-2cells. Competitive interactions were observed betweenGATA-4 and GATA-6 in Caco-2 cells and all pairs tested inHeLa cells. A cooperative interaction between GATA-4 andGATA-6 has been reported in cardiac cells (7). These resultssuggest that a functional interaction between the factors isdependent on cell-specific factors that may stabilize the inter-action in each cell type.
Many proteins are known to bind to the GATA factors, andinteracting proteins may contribute to cell specificity in het-erotypic binding or family member target gene activation. Fastoxidative-glycolytic (FOG) family factors were identified aswidely expressed regulatory cofactors that bound to GATAfactors, and FOG-2 is critical for GATA-4 function in vivo(42). Cell specificity of GATA-factor function/factor-factorinteraction may also be influenced by the binding of GATAfactors to other cell-specific transcription factors. HNF-1� is atranscription factor found in intestine but not heart, andGATA-4, GATA-5, and GATA-6 all interact functionally andphysically with HNF-1� to activate the Fabp1 transgene (11).HNF-1� is found in Caco-2 cells (11) but not HeLa cells (30)and may contribute to the cell specificity of the observedGATA interactions. We observed the GATA interactions witha Fabp1 promoter transgene lacking an HNF-1 binding site andwith a synthetic promoter with no HNF-1 binding site, butinteractions independent of HNF-1� DNA binding may occur.Direct binding of GATA-4, GATA-5, or GATA-6 to othertranscription factors found in the intestinal epithelium has alsobeen reported (27, 30, 40, 44, 46). Cell-specific protein mod-ification may also contribute to the regulation of interactionsbetween GATA factors. GATA-4 is phosphorylated, and thephosphorylated form has increased stability (7, 28) and in-creased binding to C/EBP�, another transcription factor foundin the intestinal epithelium (43).
Heterotypic binding of the GATA factors could in principletether any GATA factor present in the cell to a competentGATA binding site in any gene through interaction with adifferent GATA factor. Many cells contain at least two of thenonhematopoietic GATA factors: GATA-4, GATA-5, orGATA-6. The intestinal epithelium contains all three of thesefactors in unique spatiotemporal patterns. GATA-4 is presentin differentiated enterocytes of the newborn, suckling, and
Fig. 7. 35S-GATA-4, 35S-GATA-5, or 35S-GATA-6 wereincubated with either glutathione-S-transferase (GST) or afusion protein consisting of GST linked to GATA-4 or toGATA-5. Each solution binding experiment is representedby three lanes, indicated by the 35S-labeled GATA factorabove the lanes. The third lane of the set contains an aliquotof the 35S-labeled GATA factor run directly on the gel as acontrol for synthesis and molecular size marker (labeled“input only”). The first lane of each set is a binding assayconducted with GST as bait and is compared with thesecond lane of each assay where the bait is the GST-GATAfusion protein.
G304 INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from

adult mouse small intestine (12, 44). GATA-6 is present incrypt (proliferating) epithelial cells as well as differentiated(villus) enterocytes in the adult and neonate (12). GATA-5 ispresent in differentiated villus enterocytes of the small intestinethrough 7–15 days of age, diminishing during the suckling-weaning transition, and absent in the adult small intestinalepithelium (12). The existence of three factors with partiallyoverlapping expression patterns in the intestinal epitheliumsuggests separate functions for each GATA factor and/or use ofmultiple factors to achieve expression patterns for a singlefunction that are more complex than possible with a singlefactor. It is interesting to note that the appearance of GATA-5in the intestine before weaning allows cooperative interactionsin enterocytes with both GATA-4 and GATA-6, which arepresent throughout the entire postnatal developmental periodand thus available for interaction. These interactions may becritical for regulating genes expressed uniquely during thisperiod. The possibility of heterotypic interactions betweenthese factors provides an additional rationale for the presenceof multiple factors in a single cell, where target gene activationmay be increased through a single GATA site by multipleGATA factors. There is evidence that different GATA factorscan activate the same genes in the same cells. Intestinal targetgenes are activated by more than one GATA factor in celltransfection assays, including Fabp1 (16, 20, 37). Ectopicexpression of GATA-4, GATA-5, or GATA-6 in Xenopuslaevis oocytes activates the same cardiac-specific genes (23).cGATA-4, cGATA-5, and cGATA-6 are all expressed duringchicken cardiac development, and repression of all three genesresults in a profound defect in cardiogenesis but not loss of anytwo GATA factors (24). However, GATA factor activation oftarget genes is not entirely interchangeable, since Gata6 ex-pression cannot fully rescue the phenotype in endodermaldifferentiation of Gata4-deficient embryonic stem cells (19).All in all, GATA-4, GATA-5, and GATA-6 may have similarindividual as well as cooperative roles in the maturing intesti-nal epithelium.
ACKNOWLEDGMENTS
We are grateful to David Wilson for provision of the synthetic GATA sitetransgene and helpful discussions and to Josh and Lilia Rissman for technicalassistance.
GRANTS
This work was supported in part by National Institute of Diabetes andDigestive and Kidney Diseases Grant DK-56361 (to T. C. Simon), researchGrant 1-FY99–606 from the March of Dimes Birth Defects Foundation (toT. C. Simon), the Washington University Diabetes Research Training CenterGrant DK-20579 (to T. C. Simon), the Washington University DigestiveDiseases Research Core Center Grant DK-52574 (to T. C. Simon), the FinnishPediatric Foundation (to H. Haveri and M. Heikinheimo), and the SigridJuselius Foundation (to M. Heikinheimo).
REFERENCES
1. Al-azzeh ED, Fegert P, Blin N, and Gott P. Transcription factorGATA-6 activates expression of gastroprotective trefoil genes TFF1 andTFF2. Biochim Biophys Acta 1490: 324–332, 2000.
2. Bass NM, Manning JA, Ockner RK, Gordon JI, Seetharam S, andAlpers DH. Regulation of the biosynthesis of two distinct fatty acid-binding proteins in rat liver and intestine. Influences of sex difference andof clofibrate. J Biol Chem 260: 1432–1436, 1985.
3. Boudreau F, Rings EHHM, Swain GP, Sinclair AM, Suh ER, SilbergDG, Scheuermann RH, and Traber PG. A novel colonic repressor
element regulates intestinal gene expression by interacting with Cux/CDP.Mol Cell Biol 22: 5467–5478, 2002.
4. Boudreau F, Rings EHHM, van Wering HM, Kim RK, Swain GP,Krasinski SD, Moffett J, Grand RJ, Suh ER, and Traber PG. Hepa-tocyte nuclear factor-1�, GATA-4, and caudal related homeodomainprotein Cdx2 interact functionally to modulate intestinal gene transcrip-tion. Implication for the developmental regulation of the sucrase-isomalt-ase gene. J Biol Chem 277: 31909–31917, 2002.
5. Brewer A, Nemer G, Gove C, Rawlins F, Nemer M, Patient R, andPizzey J. Widespread expression of an extended peptide sequence ofGATA-6 during murine embryogenesis and non-equivalence of RNA andprotein expression domains. Gene Expr Patterns 2: 123–131, 2002.
6. Capo-chichi CD, Roland IH, Vanderveer L, Bao R, Yamagata T,Hirai H, Cohen C, Hamilton TC, Godwin AK, and Xu XX. Anomalousexpression of epithelial differentiation-determining GATA factors in ovar-ian tumorigenesis. Cancer Res 63: 4967–4977, 2003.
7. Charron F, Paradis P, Bronchain O, Nemer G, and Nemer M.Cooperative interaction between GATA-4 and GATA-6 regulates myo-cardial gene expression. Mol Cell Biol 19: 4355–4365, 1999.
8. Charron F, Tsimiklis G, Arcand M, Robitaille L, Liang Q, MolkentinJD, Meloche S, and Nemer M. Tissue-specific GATA factors are tran-scriptional effectors of the small GTPase RhoA. Genes Dev 15: 2702–2719, 2001.
9. Crossley M, Merika M, and Orkin SH. Self-association of the erythroidtranscription factor GATA-1 mediated by its zinc finger domains. Mol CellBiol 15: 2448–2456, 1995.
10. Di Lisi R, Millino C, Calabria E, Altruda F, Schiaffino S, and AusoniS. Combinatorial cis-acting elements control tissue-specific activation ofthe cardiac troponin I gene in vitro and in vivo. J Biol Chem 273:25371–25380, 1998.
11. Divine JK, McCaul SP, and Simon TC. HNF-1� and endodermaltranscription factors cooperatively activate Fabpl: MODY3 mutationsabrogate cooperativity. Am J Physiol Gastrointest Liver Physiol 285:G62–G72, 2003.
12. Divine JK, Staloch LJ, Haveri H, Jacobsen CM, Wilson DB, Heikin-heimo M, and Simon TC. GATA-4, GATA-5, and GATA-6 activate therat liver fatty acid binding protein gene in concert with HNF-1�. Am JPhysiol Gastrointest Liver Physiol 287: G1086–G1099, 2004.
13. Durocher D and Nemer M. Combinatorial interactions regulating cardiactranscription. Dev Genet 22: 250–262, 1998.
14. Dusing MR, Florence EA, and Wiginton DA. High-level activation bya duodenum-specific enhancer requires functional GATA binding sites.Am J Physiol Gastrointest Liver Physiol 284: G1053–G1065, 2003.
15. Dusing MR and Wiginton DA. Epithelial lineages of the small intestinehave unique patterns of GATA expression. J Mol Histol 36: 15–24, 2005.
16. Fang R, Olds LC, Santiago NA, and Sibley E. GATA family transcrip-tion factors activate lactase gene promoter in intestinal Caco-2 cells. Am JPhysiol Gastrointest Liver Physiol 280: G58–G67, 2001.
17. Fang R, Olds LC, and Sibley E. Spatio-temporal patterns of intestine-specific transcription factor expression during postnatal mouse gut devel-opment. Gene Expr Patterns 6: 426–432, 2006.
18. Fitzgerald K, Bazar L, and Avigan MI. GATA-6 stimulates a cellline-specific activation element in the human lactase promoter. Am JPhysiol Gastrointest Liver Physiol 274: G314–G324, 1998.
19. Fujikura J, Yamato E, Yonemura S, Hosoda K, Masui S, Nakao K,Miyazaki Ji J, and Niwa H. Differentiation of embryonic stem cells isinduced by GATA factors. Genes Dev 16: 784–789, 2002.
20. Gao X, Sedgwick T, Shi YB, and Evans T. Distinct functions areimplicated for the GATA-4, -5, and -6 transcription factors in the regula-tion of intestine epithelial cell differentiation. Mol Cell Biol 18: 2901–2911, 1998.
21. Ghirlando R and Trainor CD. GATA-1 bends DNA in a site-indepen-dent fashion. J Biol Chem 275: 28152–28156, 2000.
22. Gordon JI, Elshourbagy N, Lowe JB, Liao WS, Alpers DH, andTaylor JM. Tissue specific expression and developmental regulation oftwo genes coding for rat fatty acid binding proteins. J Biol Chem 260:1995–1998, 1985.
23. Jiang Y and Evans T. The Xenopus GATA-4/5/6 genes are associatedwith cardiac specification and can regulate cardiac-specific transcriptionduring embryogenesis. Dev Biol 174: 258–270, 1996.
24. Jiang Y, Tarzami S, Burch JB, and Evans T. Common role for each ofthe cGATA-4/5/6 genes in the regulation of cardiac morphogenesis. DevGenet 22: 263–277, 1998.
G305INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from

25. Kanda T, Niot I, Foucaud L, Fujii H, Bernard A, Ono T, and BesnardP. Effect of bile on the intestinal bile-acid binding protein (I-BABP)expression. In vitro and in vivo studies. FEBS Lett 384: 131–134, 1996.
26. Kawamura T, Ono K, Morimoto T, Wada H, Hirai M, Hidaka K,Morisaki T, Heike T, Nakahata T, Kita T, and Hasegawa K. Acety-lation of GATA-4 is involved in the differentiation of embryonic stemcells into cardiac myocytes. J Biol Chem 280: 19682–19688, 2005.
27. Kiela PR, LeSueur J, Collins JF, and Ghishan FK. Transcriptionalregulation of the rat NHE3 gene. Functional interactions between GATA-5and Sp family transcription factors. J Biol Chem 278: 5659–5668, 2003.
28. Kitta K, Day RM, Kim Y, Torregroza I, Evans T, and Suzuki YJ.Hepatocyte growth factor induces GATA-4 phosphorylation and cellsurvival in cardiac muscle cells. J Biol Chem 278: 4705–4712, 2003.
29. Krasinski SD, Estrada G, Yeh KY, Yeh M, Traber PG, Rings EH,Buller HA, Verhave M, Montgomery RK, and Grand RJ. Transcrip-tional regulation of intestinal hydrolase biosynthesis during postnataldevelopment in rats. Am J Physiol Gastrointest Liver Physiol 267: G584–G594, 1994.
30. Krasinski SD, Van Wering HM, Tannemaat MR, and Grand RJ.Differential activation of intestinal gene promoters: functional interactionsbetween GATA-5 and HNF-1�. Am J Physiol Gastrointest Liver Physiol281: G69–G84, 2001.
31. Liang Q, Wiese RJ, Bueno OF, Dai YS, Markham BE, and MolkentinJD. The transcription factor GATA4 is activated by extracellular signal-regulated kinase 1- and 2-mediated phosphorylation of serine 105 incardiomyocytes. Mol Cell Biol 21: 7460–7469, 2001.
32. Mackay JP and Crossley M. Zinc fingers are sticking together. TrendsBiochem Sci 23: 1–4, 1998.
33. Mackay JP, Kowalski K, Fox AH, Czolij R, King GF, and Crossley M.Involvement of the N-finger in the self-association of GATA-1. J BiolChem 273: 30560–30567, 1998.
34. Merika M and Orkin SH. DNA-binding specificity of GATA familytranscription factors. Mol Cell Biol 13: 3999–4010, 1993.
35. Morimoto T, Hasegawa K, Kaburagi S, Kakita T, Wada H, YanazumeT, and Sasayama S. Phosphorylation of GATA-4 is involved in �1-adrenergic agonist-responsive transcription of the endothelin-1 gene incardiac myocytes. J Biol Chem 275: 13721–13726, 2000.
36. Morrisey EE, Ip HS, Tang Z, Lu MM, and Parmacek MS. GATA-5:a transcriptional activator expressed in a novel temporally and spatially-restricted pattern during embryonic development. Dev Biol 183: 21–36,1997.
37. Nemer G, Qureshi ST, Malo D, and Nemer M. Functional analysis andchromosomal mapping of Gata5, a gene encoding a zinc finger DNA-binding protein. Mamm Genome 10: 993–999, 1999.
38. Nishikawa K, Kobayashi M, Masumi A, Lyons SE, Weinstein BM, LiuPP, and Yamamoto M. Self-association of Gata1 enhances transcrip-
tional activity in vivo in zebra fish embryos. Mol Cell Biol 23: 8295–8305,2003.
39. Rubin DC, Ong DE, and Gordon JI. Cellular differentiation in theemerging fetal rat small intestinal epithelium: mosaic patterns of geneexpression. Proc Natl Acad Sci USA 86: 1278–1282, 1989.
40. Sepulveda JL, Vlahopoulos S, Iyer D, Belaguli N, and Schwartz RJ.Combinatorial expression of GATA-4, Nkx2–5, and serum response factordirects early cardiac gene activity. J Biol Chem 277: 25775–25782, 2002.
41. Sweetser DA, Birkenmeier EH, Hoppe PC, McKeel DW, and GordonJI. Mechanisms underlying generation of gradients in gene expressionwithin the intestine: an analysis using transgenic mice containing fatty acidbinding protein-human growth hormone fusion genes. Genes Dev 2:1318–1332, 1988.
42. Tevosian SG, Albrecht KH, Crispino JD, Fujiwara Y, Eicher EM, andOrkin SH. Gonadal differentiation, sex determination and normal Sryexpression in mice require direct interaction between transcription partnersGATA4 and FOG2. Development 129: 4627–4634, 2002.
43. Tremblay JJ, Hamel F, and Viger RS. Protein kinase A-dependentcooperation between GATA and CCAAT/enhancer-binding protein tran-scription factors regulates steroidogenic acute regulatory protein promoteractivity. Endocrinology 143: 3935–3945, 2002.
44. van Wering H, Huibregtse I, van der Zwan S, de Bie M, Dowling L,Grand R, and Krasinski S. Physical interaction between GATA-5 andHNF-1� results in synergistic activation of the human lactase-phlorizinhydrolase promoter. J Biol Chem 277: 27659–27667, 2002.
45. Van Wering HM, Bosse T, Musters A, De Jong E, De Jong N, HogenEsch CE, Boudreau F, Swain GP, Dowling LN, Montgomery RK,Grand RJ, and Krasinski SD. Complex regulation of the lactase-phlorizin hydrolase promoter by GATA-4. Am J Physiol GastrointestLiver Physiol 287: G899–G909, 2004.
46. Wang J, Paradis P, Aries A, Komati H, Lefebvre C, Wang H, andNemer M. Convergence of protein kinase C and JAK-STAT signaling ontranscription factor GATA-4. Mol Cell Biol 25: 9829–9844, 2005.
47. Wilson DB, Dorfman DM, and Orkin SH. A nonerythroid GATA-binding protein is required for function of the human preproendothelin-1promoter in endothelial cells. Mol Cell Biol 10: 4854–4862, 1990.
48. Wingender E, Chen X, Fricke E, Geffers R, Hehl R, Liebich I, KrullM, Matys V, Michael H, Ohnhauser R, MP, Schacherer F, Thiele S,and Urbach S. The TRANSFAC system on gene expression regulation.Nuc Acids Res 29: 281–283, 2001.
49. Yamagata T, Nishida J, Sakai R, Tanaka T, Honda H, Hirano N,Mano H, Yazaki Y, and Hirai H. Of the GATA-binding proteins, onlyGATA-4 selectively regulates the human interleukin-5 gene promoter ininterleukin-5-producing cells which express multiple GATA-binding pro-teins. Mol Cell Biol 15: 3830–3839, 1995.
50. Yang HY and Evans T. Homotypic interactions of chicken GATA-1 canmediate transcriptional activation. Mol Cell Biol 15: 1353–1363, 1995.
G306 INTESTINAL GATA FACTOR INTERACTIONS
AJP-Gastrointest Liver Physiol • VOL 291 • AUGUST 2006 • www.ajpgi.org
by guest on July 26, 2016http://ajpgi.physiology.org/
Dow
nloaded from
Related Documents











