REVIEW Contribution of Callosal Connections to the Interhemispheric Integration of Visuomotor and Cognitive Processes Tilman Schulte & Eva M. Müller-Oehring Received: 6 February 2010 / Accepted: 6 April 2010 / Published online: 17 April 2010 # Springer Science+Business Media, LLC 2010 Abstract In recent years, cognitive neuroscience has been concerned with the role of the corpus callosum and interhemispheric communication for lower-level processes and higher-order cognitive functions. There is empirical evidence that not only callosal disconnection but also subtle degradation of the corpus callosum can influence the transfer of information and integration between the hemi- spheres. The reviewed studies on patients with callosal degradation with and without disconnection indicate a dissociation of callosal functions: while anterior callosal regions were associated with interhemispheric inhibition in situations of semantic (Stroop) and visuospatial (hierarchical letters) competition, posterior callosal areas were associated with interhemispheric facilitation from redundant information at visuomotor and cognitive levels. Together, the reviewed research on selective cognitive functions provides evidence that the corpus callosum contributes to the integration of perception and action within a subcortico-cortical network promoting a unified experience of the way we perceive the visual world and prepare our actions. Keywords Corpus callosum . Crossed-uncrossed difference (CUD) . Redundant targets effect (RTE) . Functional lateralization . Stroop . Hierarchical perception . Attention . Neuroimaging The pioneering work of Norman Geschwind (1965) introduced the idea that deficits in language, learning, emotion and action are manifestations of a ‘disconnection syndrome’ that result not only from cortical lesions but from white matter lesions disrupting the connectivity between sensory and association cortices. Geschwind’ s model included both connections between brain regions and the functional specialization of association cortices (Catani and ffytche 2005). The concept of a distributed neural circuitry has inspired investigations into how brain connectivity enables hemispheric specialization for mental processes, which range from basic perception (Nassi and Callaway 2009) to higher cognitive functions and con- sciousness (Galaburda and Geschwind 1980; Zaidel and Iacoboni 2003). A common theme of these investigations is how perceptions, thoughts and actions emerge from the integration of sensory, cognitive and motor inputs received or processed in separate cerebral hemispheres. Pathways Subserving Intra-and Interhemispheric Communication Cortico-cortical communication within and between hemi- spheres relies on a widespread white matter network that also connects to subcortical gray matter. The white matter tracts can be divided in association pathways that are intrahemispheric fibers connecting cortices of the frontal, parietal, temporal and occipital lobes and commissural pathways that are interhemispheric fibers connecting the cortical areas of both hemispheres (Aralasmak et al. 2006). Geschwind’ s disconnection hypothesis assumed that not cortical damage alone but cortical disconnection by white matter fiber pathway disruption makes substantial contri- T. Schulte (*) : E. M. Müller-Oehring SRI International, Neuroscience Program, 333 Ravenswood Avenue, Menlo Park, CA 94025-3493, USA e-mail: [email protected] E. M. Müller-Oehring Department of Psychiatry & Behavioral Sciences, Stanford University School of Medicine, Stanford, CA 94305, USA Neuropsychol Rev (2010) 20:174–190 DOI 10.1007/s11065-010-9130-1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REVIEW
Contribution of Callosal Connections to the InterhemisphericIntegration of Visuomotor and Cognitive Processes
Tilman Schulte & Eva M. Müller-Oehring
Received: 6 February 2010 /Accepted: 6 April 2010 /Published online: 17 April 2010# Springer Science+Business Media, LLC 2010
Abstract In recent years, cognitive neuroscience has beenconcerned with the role of the corpus callosum andinterhemispheric communication for lower-level processesand higher-order cognitive functions. There is empiricalevidence that not only callosal disconnection but also subtledegradation of the corpus callosum can influence thetransfer of information and integration between the hemi-spheres. The reviewed studies on patients with callosaldegradation with and without disconnection indicate adissociation of callosal functions: while anterior callosalregions were associated with interhemispheric inhibition insituations of semantic (Stroop) and visuospatial (hierarchicalletters) competition, posterior callosal areas were associatedwith interhemispheric facilitation from redundant informationat visuomotor and cognitive levels. Together, the reviewedresearch on selective cognitive functions provides evidencethat the corpus callosum contributes to the integration ofperception and action within a subcortico-cortical networkpromoting a unified experience of the way we perceive thevisual world and prepare our actions.
Keywords Corpus callosum . Crossed-uncrossed difference(CUD) . Redundant targets effect (RTE) . Functionallateralization . Stroop . Hierarchical perception . Attention .
Neuroimaging
The pioneering work of Norman Geschwind (1965)introduced the idea that deficits in language, learning,emotion and action are manifestations of a ‘disconnectionsyndrome’ that result not only from cortical lesions butfrom white matter lesions disrupting the connectivitybetween sensory and association cortices. Geschwind’smodel included both connections between brain regionsand the functional specialization of association cortices(Catani and ffytche 2005). The concept of a distributedneural circuitry has inspired investigations into how brainconnectivity enables hemispheric specialization for mentalprocesses, which range from basic perception (Nassi andCallaway 2009) to higher cognitive functions and con-sciousness (Galaburda and Geschwind 1980; Zaidel andIacoboni 2003). A common theme of these investigations ishow perceptions, thoughts and actions emerge from theintegration of sensory, cognitive and motor inputs receivedor processed in separate cerebral hemispheres.
Pathways Subserving Intra-and InterhemisphericCommunication
Cortico-cortical communication within and between hemi-spheres relies on a widespread white matter network thatalso connects to subcortical gray matter. The white mattertracts can be divided in association pathways that areintrahemispheric fibers connecting cortices of the frontal,parietal, temporal and occipital lobes and commissuralpathways that are interhemispheric fibers connecting thecortical areas of both hemispheres (Aralasmak et al. 2006).Geschwind’s disconnection hypothesis assumed that notcortical damage alone but cortical disconnection by whitematter fiber pathway disruption makes substantial contri-
T. Schulte (*) : E. M. Müller-OehringSRI International, Neuroscience Program,333 Ravenswood Avenue,Menlo Park, CA 94025-3493, USAe-mail: [email protected]
E. M. Müller-OehringDepartment of Psychiatry & Behavioral Sciences,Stanford University School of Medicine,Stanford, CA 94305, USA
Neuropsychol Rev (2010) 20:174–190DOI 10.1007/s11065-010-9130-1

butions to functional deficits in neurological and psychiatricdisorders.
Intrahemispheric Communication Several cortico-corticalassociation and cortico-subcortical projection pathwaysensure communication within each hemisphere. Superiorlongitudinal fasciculus (SLF) fibers, for example, connecttemporo-parieto-occipital regions with the frontal lobe.While left-hemispheric SLF fibers have been related tospeech (Duffau et al. 2002) and language comprehension(Tanabe et al. 1987), right-hemispheric fibers have beenrelated to the spatial-attention network (Doricchi andTomaiuolo 2003). The inferior longitudinal fasciculi con-nect temporal to occipital lobes in each hemisphere andhave been associated with higher-order visual functionssuch as object recognition (left-lateralized; Salvan et al.2004), face recognition, and visually evoked emotions(bilateral or right-lateralized; Bauer 1982; Habib 1986).The inferior fronto-occipital fasciculi connect visual asso-ciation areas with the frontal eye fields, and there is someevidence that they mediate simultaneous perception intra-hemipherically needed for selecting visual information incrowded scenes (right-lateralized or bilateral; Battelli et al.2003; Rizzo and Vecera 2002). The cingulum is a largefiber bundle of the limbic system connecting cingulate andhippocampal gyri with amygdala, nucleus accumbens,thalamus and frontal cortices, areas involved in emotion,reward and executive control. Thus, each hemisphere hasextensive cortico-cortical and cortico-subcortical connec-tions (Mori et al. 2002). The myriad information trans-ferred via these fiber tracts and processed in eachhemisphere is also available for integration between thetwo hemispheres via commissural pathways to form aunified perception.
Interhemispheric Communication The major commissuralpathway is the corpus callosum (CC), a thick band of white
matter fibers consisting of more than 200 million axonsconnecting the right and left cerebral hemispheres of thebrain and playing an important role in interhemispherictransfer of information (Aboitiz et al. 1992; Banich 1995;Banich and Shenker 1994; Innocenti 1986; Schmahmannand Pandya 2006). Although callosal fibers mostly projectto homotopic regions of the hemispheres (e.g., parietal lobeto parietal lobe, etc.) along an anterior-posterior gradient(McCulloch and Garol 1941; Putnam et al. 2008), somecallosal fibers also project to heterotopic regions (e.g.,parietal to frontal lobe, etc.) (Clarke and Zaidel 1994; Cook1984; Rakic and Yakovlev 1968). The CC has nomacroscopic anatomical landmarks that delimit anatomicaland functionally distinct regions (Hofer and Frahm 2006).It has been geometrically divided into three regions(Duara et al. 1991; Sullivan et al. 2002) (1) an anteriorpart (genu+rostrum) that connects frontal and premotorregions of the two hemispheres, (2) a middle part (body)that interconnects motor, somatosensory and parietal areas,and (3) a posterior part of the corpus callosum (splenium)that interconnects temporal and occipital cortices (Fig. 1)(Park et al. 2008; Putnam et al. 2009; Wahl et al. 2007) butthere have been numerous geometric models for differentiatingsections of the CC (e.g., Ryberg et al. 2007; Witelson 1989;see also Chanraud et al., in this issue). Although ourunderstanding of the organization of CC white matterpathways has considerably increased (Poupon et al. 2001;Putnam et al. 2009) in the advent of recent development ofdiffusion tensor imaging (DTI) techniques (Baird and Warach1998; Le Bihan et al. 2001; Le Bihan 2003; Moseley et al.1990; Pfefferbaum et al. 2003; Sullivan and Pfefferbaum2003; Wahl et al. 2007; Westerhausen et al. 2009) relativelylittle is still known about the relation between callosal regionsand interhemispheric integration of sensory and cognitiveinformation (Bloom and Hynd 2005).
In humans, primary visual, auditory and motor cortexfunctions are highly lateralized with the left cerebral
Sagittal Axial Coronal
Fig. 1 Sagittal, axial andcoronal section of the humanbrain (The SRI24 multichannelatlas of normal adult humanbrain structure; Rohlfing et al.2010) illustrating the anterior(genu), middle (body) andposterior (splenium) sectors ofcorpus callosum (CC). Otherillustrated interhemisphericconnections are the anteriorcommissure (AC) and thesuperior colliculi (SC)
Neuropsychol Rev (2010) 20:174–190 175

hemisphere processing right-sided sensory input and motoroutput and the right cerebral hemisphere processing left-sided sensory input and motor output. Input from bothhemispheres needs to be integrated and selected for furtherhigher-order cognitive processing and for a unified motorresponse (Stephan et al. 2003). From an evolutionarystandpoint the CC is assumed to play an integral role inthe development of higher cognitive processes and hemi-spheric specialization (Doron and Gazzaniga 2008). Duringhuman evolution, brain and CC size increased considerably.Some studies suggest that the communication speedbetween the two hemispheres is faster with increased brainsize (Caminiti et al. 2009; Innocenti 1995) and that largercallosal area indicate faster interhemispheric communica-tion (Jancke and Steinmetz 1994; Paul et al. 2007), whileothers suggest a progressive slowing of interhemisphericcommunication in larger brains (Ringo et al. 1994).Interestingly, recent animal studies have suggested thatmore lateralized brains are better to act directly on manysources of information at the same time (Magat and Brown2009). For example, chimpanzees that use one hand tofish termites out of their mounds are more efficientthan chimpanzees that are ambidextrous (Marchant andMcGrew 1996).
Although the CC is the largest interhemispheric com-missure, other commissural pathways include the anterior,hippocampal, posterior, habenuar, and tectal commissuresand also the two sides of the brainstem reticular formationhave extensive connections (Aralasmak et al. 2006). Thesuperior colliculus and the tectal commissure are thought toplay a role in visual transfer and to maintain interhemispherictransfer of basic visual information in patients with callosalagenesis, callosotomy and commissurotomy (Tardif andClarke 2002). Nevertheless, studies in acallosal and split-brain patients state quite clearly that absence or loss of CCintegrity contributes to impairment in sensory and cognitiveintegration (Fabri et al. 2001; Yamauchi et al. 1997). In fact,many years of research have demonstrated a relationshipbetween callosal connectivity and prolonged interhemispherictransfer time in split-brain patients and in acallosal patients(Corballis 1998; Iacoboni et al. 2000; Marzi et al. 1991;Mooshagian et al. 2009; Paul et al. 2007; Reuter-Lorenzet al. 1995; Roser and Corballis 2002). Less radicaldisruption of callosal fiber integrity such as seen in patientswith multiple sclerosis (Warlop et al. 2008), schizophrenia(Schrift et al. 1986; Woodruff et al. 1997), alcoholism(Hutner and Oscar-Berman 1996; Schulte et al. 2004,2005, 2008), autism (Keary et al. 2009), and even innormal aging (Jeeves and Moes 1996; Schulte et al. 2005)has been also associated with prolonged interhemispherictransfer time (IHTT).
Callosal Function in Interhemispheric Communication
Different callosal functions have been proposed for inter-hemispheric communication. For example, the hemisphericinsulation view states that the CC acts as a shield to reduceinterference and to prevent potentially harmful intrusionsfrom the opposite hemisphere (Kinsbourne and Hicks 1978;Liederman and Meehan 1986), whereas the callosalinhibition view assumes that stimulus-driven activity inone hemisphere suppresses activity in the other, thereby, forinstance, causing a shift in attention to the contralateral sideof the more activated hemisphere (Chiarello and Maxfield1996). Callosal reciprocal inhibition may further equilibrateactivation and prevent attention from being skewed to thecontralateral side when one hemisphere is disproportion-ately active (Kinsbourne 2003). Interhemispheric coopera-tion between the hemispheres has been assumed to be moreadvantageous than within hemispheric processing in situa-tions of high processing demand (Banich and Belger 1990).Interhemispheric sharing of information and the need forinhibiton or isolation from each other are not mutuallyexclusive and may depend on the kind of informationprocessed. It is, for example, well established that the twohemispheres have different functions, with a left-hemisphericsuperiority in processing speech and language and a right-hemispheric superiority for certain visuospatial functions.Thus, when one hemisphere is superior to the other inprocessing specific information, intrahemispheric processingmay be more advantageous, and callosal isolation orinhibition would increase processing efficiency and perfor-mance. By contrast, when both hemispheres are qualified toprocess the same information, sharing of hemisphericresources may be more advantageous and require callosalinterhemispheric cooperation (Bloom and Hynd 2005).
Interhemispheric Communication for Visuomotorand Cognitive Functions
First, we will provide an overview of the role of the CC inlower-level processes and interhemispheric visuomotorintegration as documented in recent behavioral and neuro-imaging studies on the crossed-uncrossed difference, anestimation of interhemispheric transfer time (Poffenberger1912), and on redundancy gain, a measure of visuomotorintegration (Miller 1991; Mordkoff and Yantis 1991, 1993).Secondly, we will explore the contribution of the CC tohigher-order cognitive functions, such as attention andexecutive control, and to integrating the left hemisphere,specialized for processing local details and language, andright hemisphere, specialized for processing holistic, global
176 Neuropsychol Rev (2010) 20:174–190

forms and spatial attention functions (Aziz-Zadeh et al.2006; Gazzaniga 2000; Gazzaniga 2005; Hugdahl andDavidson 2003).
The Role of the CC for the Integration of Lower-LevelFunctions
Interhemispheric Transfer Time and the Corpus CallosumApproaches describing and analyzing interhemisphericintegration of information in humans include simplereaction time (RT) tasks in which targets are presented inthe same (uncrossed) or opposite (crossed) visual field inrelation to the responding hand (Poffenberger 1912; Zaideland Iacoboni 2003). The difference between crossedand uncrossed reaction times relative to the respondinghand (Crossed-Uncrossed Difference, CUD; crossed: lefthemifield/right hand and right hemifield/left hand;uncrossed: left hemifield/left hand, right hemifield/righthand) provides an index of interhemispheric transfer time(IHTT), because the uncrossed response can be processedwithin the same hemisphere, whereas crossed responsesrequire transfer of visuomotor information between hemi-spheres via the CC (Corballis 1998, 2002; Iacoboni andZaidel 1995, 2004; Mooshagian et al. 2008; Pollmann andZaidel 1998; Tettamanti et al. 2002).
In normal subjects the CUD is approximately 4–6 ms(Marzi et al. 1991), and shorter in younger than older adults(Schulte et al. 2004; Reuter-Lorenz and Stanczak 2000).Hanajima et al. (2001) found that muscle responses totranscranial stimulation over the left-hand motor cortex canbe facilitated when transcranial stimulation has beenapplied 4–5 ms earlier over the right-hand motor cortex,about the same time as the CUD value estimated frombehavioral studies in normal subjects. In acallosal andcommissurotomized patients the CUD is prolonged (∼30–70 ms) because of longer reaction times to lateralizedstimuli in the crossed compared to the uncrossed conditionowing to the indirect route required for interhemispherictransfer (e.g., Berlucchi et al. 1971; Berlucchi et al. 1995;Clarke and Zaidel 1989; Forster and Corballis 1998, 2000).
There is an ongoing debate whether the CUD is basedsolely on the anatomy of callosal fibers or whether theCUD may also be mediated by attentional components(Mooshagian et al. 2008). Support for an anatomicalinterpretation of the CUD comes from DTI studies. DTIyields estimates of fractional anisotropy (FA) that indexeswhite matter structural integrity and measures the orienta-tional displacement and distribution of water molecules invivo across tissue components (Pierpaoli and Basser 1996).FA is typically higher in fibers with a homogeneous orlinear structure than in tissue with an inhomogenousstructure, such as areas with pathology (Lansberg et al.
2001; Neumann-Haefelin et al. 2000) or crossing fibers(Pfefferbaum and Sullivan 2003; Virta et al. 1999). DTIstudies have revealed correlations between subtle variationsin regional white matter callosal microstructure andbehavioral measures of IHTT and cognitive ability (Muetzelet al. 2008; Schulte et al. 2005; Sullivan et al. 2010). Inadolescence (9–23 years), for example, age-relatedincreases in callosal FA were associated with betterbimanual coordination using an alternating finger-tappingtask (Muetzel et al. 2008), and in adults (20–78 years), age-related white matter degradation correlated with poorermotor and cognitive performance (Sullivan et al. 2010). Innormal aging, Schulte et al. (2005) found that low FA incallosal genu and splenium correlated with longer IHTTmeasured with the CUD (Poffenberger paradigm, 1912).By contrast, Westerhausen et al. (2006) did not find acorrelation between callosal diffusion parameters and theCUD (2.5 ms) but did find negative correlations betweenmean diffusion and IHTT, derived from the P100 event-related potential. Methodological variables such as stimulusparameters (eccentricity and luminance, fixation control,forehead-chin rest) (Schiefer et al. 2001), calculation ofthe CUD using median or mean reaction times (RT)(Braun et al. 1995), and a reduced variance by choosing ahomogenous sample of young male healthy collegestudents (Westerhausen et al. 2006) may have contributedto the relative insensitivity of behavioral CUD measures(Bashore 1981).
There is also evidence that attention influences CUDmeasures. In a functional magnetic resonance imaging(fMRI) study, Weber et al. (2005) tested healthy right-handed volunteers with a modified Poffenberger paradigmin which the attentional focus was implicitly directedtoward the contra-or ipsilateral hemifield relative to theresponding hand. They found that crossed relative touncrossed trials (CUD) activated anterior callosal area, butwhen attentionally manipulated, activated superior colliculiand right superior parietal cortex, a region typicallyassociated with attentional shifts. This is consistent withfindings of Iacoboni and Zaidel (2004) suggesting thatshifts in spatial attention (Corbetta et al. 1995) can criticallyinfluence sensory-motor integration (Creem-Regehr 2009).Activations of premotor and parietal cortex regionsobserved with fMRI also map to fibers linking throughthe mid-body and anterior regions of the CC (Iacoboni andZaidel 2004; Schulte et al. 2005). Recently, Mooshagianand colleagues examined the influence of spatial attentionalshifts on CUD in patients with agenesis of the CC andcomplete commissurotomy (2009) and in healthy subjects(2008) by alternating between natural (arms uncrossed) andunnatural arm positions (arms crossed). The CUD wassmaller with arms crossed (unnatural arms position) thanuncrossed (natural arms position) in patients as well as
Neuropsychol Rev (2010) 20:174–190 177

normal controls indicating that the relative arm positionis sufficient to modulate the CUD even in the absence ofthe CC.
To summarize, the CUD indexes the time required forinformation to cross the CC (Berlucchi et al. 1995; Fendrichet al. 2004), but there is also evidence that the CUD canbe modulated by spatial attention (Braun et al. 1995;Mooshagian et al. 2008). Other possible factors thatinfluence the CUD are age (Jeeves and Moes 1996; Schulteet al. 2004; Reuter-Lorenz and Stanczak 2000) and sexdifferences in callosal structure and size (Luders et al. 2003).In a recent event related potential (ERP) experiment, Moeset al. (2007) revealed shorter and more symmetrical IHTTfor women than men. It appears that IHTT betweenhemispheres is faster from right-to-left than from left-to-right (Barnett and Corballis 2005; Iwabuchi and Kirk 2009).The reduced asymmetry in women is mainly due to shorterleft-to-right IHTT in women than men. Different assump-tions have been made to explain this asymmetry in IHTTsuch as a smaller average size in the right occipital loberesulting in fewer callosal fibers projecting from left-to-rightposterior areas (Saron and Davidson 1989), faster axonalconduction in the right hemisphere relative to the left(Barnett and Corballis 2005), or the degree of hemisphericspecialization as suggested by a reversal of asymmetrictransfer times for verbal and non-verbal stimuli (Nowickaet al. 1996; Rugg and Beaumont 1978). Alternatively,differences in the distribution of gray and white matter inthe two cerebral hemispheres have been proposed to reflectneuronal organization promoting inter-or intrahemisphericprocessing with more gray matter relative to white matter inthe left hemisphere than in the right, particularly in thefrontal regions, emphasizing parallel intrahemispheric pro-cessing rather than interhemispheric transfer (Gur et al.1980). Current volumetric magnetic resonance imaging(Pujol et al. 2002) and DTI (Hagmann et al. 2003; Krauset al. 2007) protocols have reported left–right asymmetries ofthe superior longitudinal tract, the upper cerebellar peduncleand in frontal and temporal white matter supporting theassumption that anatomical differences between both hemi-spheres and the proportion of gray and white matter mayunderlie functional asymmetry.
Parallel Processing, Redundancy Gain and the CorpusCallosum To explore our visual environment, we integratevisual information from each visual hemifield as it reachesthe contralateral cerebral hemisphere. Because of theanatomical arrangement of the human visual system thatprojects visual inputs from each visual hemifield to thecontralateral visual cortex, the independent contributionsfrom each hemisphere and the role of the corpus callosumfor the integration of parallel visuomotor processes can bereadily assessed by comparing paired with single targets
presented to one or both visual hemifields. Typically,responses are faster to stimulus pairs than single stimuli, aphenomenon called the redundant targets effect (RTE), orsummation effect (Miller 1986; Mordkoff and Yantis 1991;Roser and Corballis 2003). The RTE is usually tested withbrief light flashes presented for less than 150ms to prevent eyemovements that would shift the visual field (Reuter-Lorenzet al. 1995; Miniussi et al. 1998; Iacoboni et al. 2000).
Researchers have proposed two explanations for redun-dancy gains from paired stimulation, the ‘horse race’ model(Raab 1962) and the ‘co-activation’ model (Miller 1982). Iftwo stimuli, one within each hemisphere, are processedindependently or in parallel, the faster processed stimulus‘wins the race’ (Miller and Ulrich 2003; Raab 1962).Statistically, the probability of eliciting a fast response istwice as high when two stimuli are presented simultaneouslythan when only one is presented. The probability or ‘race’model assumes that each stimulus is transmitted alongseparate channels and the response is triggered as soon as adecision is made by either one of the two channels (Mordkoffand Yantis 1991). An alternative to the ‘race’ model isrequired to explain the situation when gain from doublestimulus presentations exceeds probability (Raab 1962),i.e., an enhanced redundancy gain. Here, co-activation fromotherwise independent processing channels may occur toincrease signal strength of redundant targets and produce aspeeded response (Miller 1982).
Millers’ use of the term co-activation was theoretical andnot founded on measured brain activity. Recent studieshave associated enhanced redundancy gain (co-activationmodel) with neural correlates and implied a possible role ofthe CC (Bucur et al. 2005; Miniussi et al. 1998; Savazziand Marzi 2002; Schulte et al. 2006a; Turatto et al. 2004),in contrast to the horse race model that assumes twoindependent processing channels and does not require thecorpus callosum. For example, studies on split brainpatients and on normal subjects testing redundancy gainfor paired targets within the same visual hemifield, i.e.,intrahemispheric summation, and in both hemifields, i.e.,interhemispheric summation, report larger redundancy gainsfor paired bilateral than unilateral stimulation (Iacoboni et al.2000; Marks and Hellige 2003; Ouimet et al. 2009;Pollmann and Zaidel 1998; Reuter-Lorenz et al. 1995;Schulte et al. 2006a) indicating that interhemisphericcommunication may mediate neural co-activation. However,split-brain and acallosal patients typically show abnormallyenhanced bilateral RTEs compared to healthy adults, whichimplies that other than callosal commissures can mediateinterhemispheric processing advantages (Aglioti et al. 1996;Corballis 1998; Corballis et al. 2003; Iacoboni et al. 2000;Reuter-Lorenz et al. 1995; Roser and Corballis 2002, 2003).It has been argued that healthy individual may not benefit asmuch from a bilateral presentation as do the split-brain
178 Neuropsychol Rev (2010) 20:174–190

individuals because an intact corpus callosum assures fasttransfer of information between hemispheres even when onlyone hemisphere is stimulated directly by visual input, i.e.,when a single lateralized stimulus is presented.
But how can enhanced RTEs in the split-brain dependupon convergence of information from separated hemi-spheres when at the same time prolonged CUDs in split-brain patients reflect slow subcortical transmission? Toexplicate this paradox in the split-brain, Reuter-Lorenz etal. (1995) proposed in their “and-or model” that the RTE isbased on response competition between the cerebral hemi-spheres that depends on callosal connectivity. According tothis model, co-activation occurs at a response selectionstage (see also Miller 2004; Roser and Corballis 2003), andacts as an ‘and’ gate that requires input from bothhemispheres, while prolonged CUDs in split-brain patientsare due to the use of relatively inefficient ipsilateralpathways in making crossed responses. Corballis et al.(2002) assumed that summation is due to cortical projectionto a subcortical arousal system that is normally inhibitedby the corpus callosum, while in the split-brain patient,interhemispheric inhibition is released, resulting in aparadoxically enhanced redundant targets effect.
Even though the CC is the largest commissure at thecortical level, there are other pathways connecting thehemispheres: the anterior and posterior commissures, andsubcortical projections through the hippocampal, habenular,and intercollicular brain systems (Bayard et al. 2004).Activation of the superior colliculi, part of the retino-tectalpathway within the visual brain system, has been associatedwith the initiation of fast responses (e.g., eye movements:Klier et al. 2003; arm-reaching movements: Dean et al.1989; and even shifts in attention that do not involve anyovert movements: Ignashchenkova et al. 2004). Iacoboni etal. (2000) proposed that the superior colliculi and theposterior callosal area connecting visual extrastriate areasare the key structures for interhemispheric neural co-activation explaining visuomotor integration betweenhemispheres: Activity in the extrastriate cortex followingvisual input feeds into the superior colliculi, which thenfeeds back to extrastriate cortices. In split-brain patientswith long interhemispheric transfer times (IHTT) asmeasured with the CUD, feedback to the superior colliculiwould be asynchronous and sum up over time to be largerthan in normal subjects with short IHTT and synchronoussuperior colliculus input. The larger superior colliculusactivation would feed a stronger signal back to theextrastriate cortices, which in turn send stronger activationto the premotor cortex, speeding the response. Consistentwith his model, Iacoboni et al. (2000) found enhancedRTEs in split-brain patients with long CUDs (> 15 ms) butnot in patients with short CUDs (< 15 ms). Furtherevidence for an involvement of subcortical structures, such
as the superior colliculus, or cortico-subcortical interactions(Roser and Corballis 2003) comes from split-brain studies(Corballis 1998; Savazzi and Marzi 2004) demonstratingreduced RTEs with equiluminant stimuli that are invisibleto the superior colliculi and thus restrict processing tocortical pathways of the retino-geniculate parvocellularsystem (Livingstone and Hubel 1987). However, despiteconverging evidence for a perceptual explanation for theRTE (Corballis et al. 2003; Iacoboni et al. 2000; Miniussiet al. 1998; Murray et al. 2001; Schulte et al. 2006a),others argue that there could also be a premotor explana-tion (Diederich and Colonius 1987; Giray and Ulrich1993; Iacoboni and Zaidel 2003; Ouimet et al. 2009).
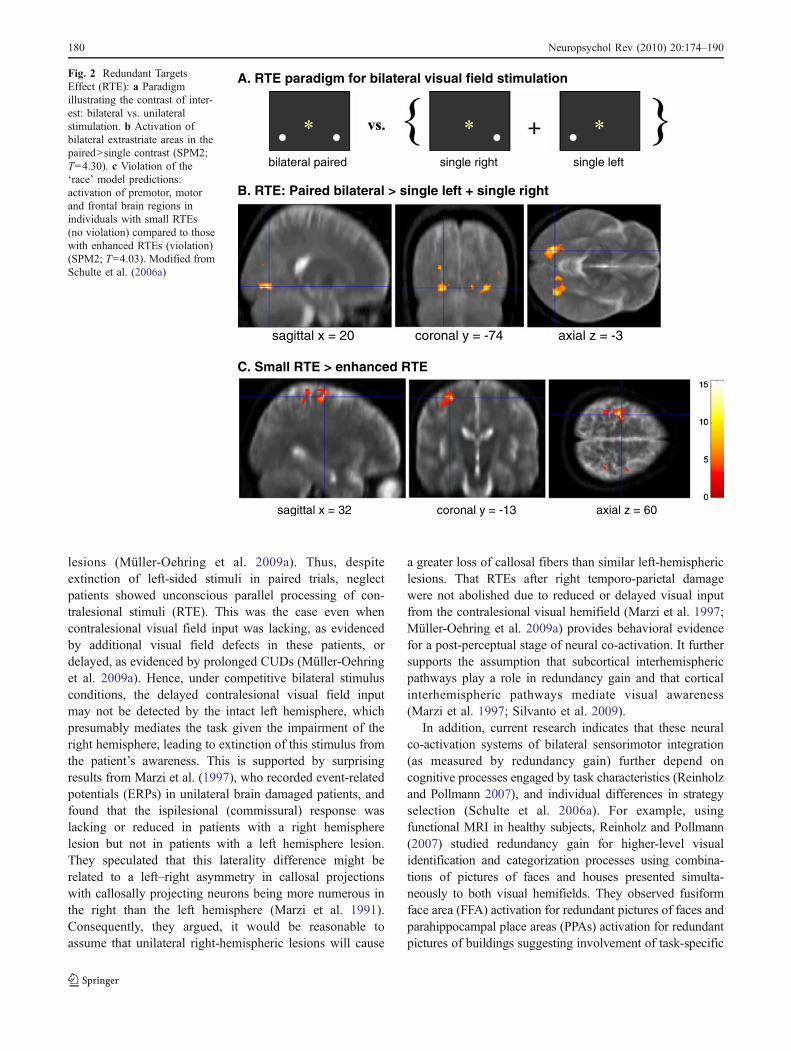
In healthy subjects, considerable inter-subject variabilityhas been observed: approximately half showing enhancedRTEs that violate the ‘race’ model while half do not(Corballis 2002; Schulte et al. 2006a). This normalvariability in RTE suggests involvement of different neuralloci (extrastriate, premotor or frontal cortical areas) depend-ing on strategy selection, task demands, and stimuluscharacteristics. In an fMRI study comparing normalindividuals with small and enhanced RTEs, we foundevidence for both perceptual and premotor/decisional locifor the RTE. Whereas all individuals showed bilateralactivation of extrastriate cortices to paired in contrast tosummed single stimulus conditions, individuals with smallRTEs activated frontal and premotor areas more than thosewith large RTEs (Schulte et al. 2006a) (Fig. 2).
Our analysis was between individuals, whereas that ofMooshagian et al. (2008) compared activation between fast‘co-activation’ trials and slower ‘not co-activation’ trialswithin the same healthy subjects. They found righttemporo-parietal junction activation for ‘co-activation’trials, specifically in the angular gyrus. This finding is ofparticular interest because the temporo-parietal junction hasbeen implicated in spatial attention and, when damaged, inthe occurrence of visuo-spatial neglect (Lux et al. 2006;Mort et al. 2003; Müller-Oehring et al. 2007, 2009a;Shirani et al. 2009). A phenomenon closely associated withvisuo-spatial neglect is ‘extinction to paired simultaneousstimulation,’ when patients see single visual stimuli, but failto report contralesional stimuli when an ipsilesionalstimulus is presented at the same time. In this case, thecontralesional stimulus is extinguished from awareness(Meister et al. 2006; Schendel and Robertson 2002).
In patients with unilateral left extinction after righttemporo-parietal damage, enhanced redundancy gains havebeen observed even when one stimulus in a pair waspresented to the left hemifield and therefore was notperceived (Marzi et al. 1996; Müller-Oehring et al. 2009a,b, 2010). We recently found that visual extinction of left-sided stimuli, but not RTE, was correlated with prolongedCUDs in neglect patients with right temporo-parietal
Neuropsychol Rev (2010) 20:174–190 179
lesions (Müller-Oehring et al. 2009a). Thus, despiteextinction of left-sided stimuli in paired trials, neglectpatients showed unconscious parallel processing of con-tralesional stimuli (RTE). This was the case even whencontralesional visual field input was lacking, as evidencedby additional visual field defects in these patients, ordelayed, as evidenced by prolonged CUDs (Müller-Oehringet al. 2009a). Hence, under competitive bilateral stimulusconditions, the delayed contralesional visual field inputmay not be detected by the intact left hemisphere, whichpresumably mediates the task given the impairment of theright hemisphere, leading to extinction of this stimulus fromthe patient’s awareness. This is supported by surprisingresults from Marzi et al. (1997), who recorded event-relatedpotentials (ERPs) in unilateral brain damaged patients, andfound that the ispilesional (commissural) response waslacking or reduced in patients with a right hemispherelesion but not in patients with a left hemisphere lesion.They speculated that this laterality difference might berelated to a left–right asymmetry in callosal projectionswith callosally projecting neurons being more numerous inthe right than the left hemisphere (Marzi et al. 1991).Consequently, they argued, it would be reasonable toassume that unilateral right-hemispheric lesions will cause
a greater loss of callosal fibers than similar left-hemisphericlesions. That RTEs after right temporo-parietal damagewere not abolished due to reduced or delayed visual inputfrom the contralesional visual hemifield (Marzi et al. 1997;Müller-Oehring et al. 2009a) provides behavioral evidencefor a post-perceptual stage of neural co-activation. It furthersupports the assumption that subcortical interhemisphericpathways play a role in redundancy gain and that corticalinterhemispheric pathways mediate visual awareness(Marzi et al. 1997; Silvanto et al. 2009).
In addition, current research indicates that these neuralco-activation systems of bilateral sensorimotor integration(as measured by redundancy gain) further depend oncognitive processes engaged by task characteristics (Reinholzand Pollmann 2007), and individual differences in strategyselection (Schulte et al. 2006a). For example, usingfunctional MRI in healthy subjects, Reinholz and Pollmann(2007) studied redundancy gain for higher-level visualidentification and categorization processes using combina-tions of pictures of faces and houses presented simulta-neously to both visual hemifields. They observed fusiformface area (FFA) activation for redundant pictures of faces andparahippocampal place areas (PPAs) activation for redundantpictures of buildings suggesting involvement of task-specific
A. RTE paradigm for bilateral visual field stimulation
bilateral paired single right single left
B. RTE: Paired bilateral > single left + single right
sagittal x = 20 coronal y = -74 axial z = -3
C. Small RTE > enhanced RTE
sagittal x = 32 coronal y = -13 axial z = 60
Fig. 2 Redundant TargetsEffect (RTE): a Paradigmillustrating the contrast of inter-est: bilateral vs. unilateralstimulation. b Activation ofbilateral extrastriate areas in thepaired>single contrast (SPM2;T=4.30). c Violation of the‘race’ model predictions:activation of premotor, motorand frontal brain regions inindividuals with small RTEs(no violation) compared to thosewith enhanced RTEs (violation)(SPM2; T=4.03). Modified fromSchulte et al. (2006a)
180 Neuropsychol Rev (2010) 20:174–190

higher-order visual object-selective areas in visuomotorintegration of redundant information.
Despite the ongoing debate over the cortical locus ofco-activation, results from split-brain research clearlydemonstrate that interhemispheric integration of simplepercepts can occur without the presence of the CC throughsubcortical transmission of information that is projected tocortical sites in both hemispheres. In the split-brain,enhanced RTEs have been interpreted as an absence ofcallosal inhibition (Corballis et al. 2002; Zaidel andIacoboni 2003). Yet, in healthy individuals, a recent studyshowed greater RTEs for equiluminant than high-contraststimuli implying cortical rather than subcortical integrationmechanisms (Schulte et al. 2004), in contrast to split-brainpatients, who typically show reduced RTEs with equilu-minant stimuli (Corballis 1998; Roser and Corballis 2002).Furthermore, callosal degradation without disconnection,e.g., in chronic alcoholics (Lim and Helpern 2002; Kubickiet al. 2003; Pfefferbaum and Sullivan 2003) has been foundto correlate with reduced RTE with equiluminant stimuli(Schulte et al. 2005). These results imply a cooperative roleof the CC and that such cooperation is reduced by subtledegradation of callosal integrity. In addition, data in healthysubjects with an intact CC showed bilateral extrastriateactivation for RTEs (Schulte et al. 2006a) (Fig. 2) andtemporo-parietal activations for enhanced RTEs (Mooshagianet al. 2008) and support an interhemispheric cooperationmodel for neural summation. Subtle degradation ofcallosal structure also occurs in healthy aging (Bartzokiset al. 2004; Bucur et al. 2005; Madden et al. 2009;Sullivan et al. 2001) and can result in disturbed bilateralintegration of visual (Müller-Oehring et al. 2007; Schulteet al. 2004, 2005), cognitive (Kennedy and Raz 2009;Zahr et al. 2009), and motor functions (Bartzokis et al.2008; Sullivan et al. 2006, 2010).
In summary, the role of the corpus callosum for theintegration of lower-level visuomotor functions is contro-versial. While split-brain research indicates that the CC actsin an inhibitory fashion within a subcortico-corticalnetwork (Corballis et al. 2002; Roser and Corballis 2003),recent research on callosal degradation without disconnec-tion suggests that the corpus callosum acts cooperatively.The review of the few studies on interhemisphericcommunication emanating from the right temporo-parietaljunction further suggests a special role of the corpuscallosum in conscious perception (Marzi et al. 1991,1997; Müller-Oehring et al. 2009a). Activation of the righttemporo-parietal junction appears to mediate co-activationfor visuomotor integration in conjunction with homologoustemporo-parietal regions in the contralateral hemisphere(Mooshagian et al. 2008) probably associated with bilateralattentional mechanisms normally equilibrated by reciprocal
callosal inhibition (Kinsbourne 1993) supporting consciousperception (Reuter-Lorenz et al. 1995).
The Role of the CC for the Integration of HigherCognitive Functions
Hemispheric specialization for higher-order cognition suchas the left-hemispheric dominance for language and right-hemispheric dominance for visuospatial attention (Damasioand Damasio 1993; Gazzaniga 2005) has been assumed tobe a consequence of interhemispheric connectivity, i.e., thelikelihood of intrahemispheric processing and hemisphericspecialization of brain functions increases with increasedtime constraints of callosal conduction delay (Hopkins andRilling 2000). Although the functions of the CC are moreassociated with transferring sensory information and visuo-motor integration (Banich 1998) recent studies highlightthe pivotal role of CC for higher-order cognitive functionssuch as identification of complex stimuli (Banich et al.2000; Schulte et al. 2006b; Skiba et al. 2000). Studyingneuropsychiatric diseases, such as alcoholism, that aremarked by subtle white matter neuropathology, particularlyof the CC (Harper and Kril 1988, 1990; Harper andMatsumoto 2005; Pfefferbaum and Sullivan 2002, 2005;Pfefferbaum et al. 2006; Tarnowska-Dziduszko et al. 1995),provides a window of opportunity to investigate the contri-bution of CC degeneration to the functional integration ofinformation across the hemispheres to perform complex tasks(Hiatt and Newman 2007).
It has been assumed that both hemispheres interactwith each other in a dynamic push-pull fashion to equalizethe direction of visuospatial attention (Kinsbourne 1977,Kinsbourne and Bruce 1987; Reuter-Lorenz et al. 1990;Corbetta et al. 2005). In this activation-orienting model it ispostulated that visuospatial attention is biased in thedirection contralateral to the more activated hemisphere(Schulte et al. 2001). Based on the assumption that dynamicinteractions between the hemispheres via the CC affectattentional processing (Banich 1998), we recently testedwhether the integrity of the CC in alcoholics predicts thefunctioning of neural systems known to underlie higher-order component processes of selective attention andconflict processing. To identify the role of hemisphericlateralization and interhemispheric transfer within thefronto-parietal attention system, we developed a lateralizedStroop paradigm (Schulte et al. 2006b). For nearly 75 years,the Stroop color-word effect has provided a rich paradigmfor parsing and manipulating processes of attention andconflict in the context of stimuli naturally compellingbecause of their semantic content (Stroop 1935; MacLeod1991). Processing words at their semantic level is involun-
Neuropsychol Rev (2010) 20:174–190 181

tary and automatic and naturally overrides perceptual tags,such as the color of the ink of a written word that has nointrinsic value (Carter et al. 2000; MacDonald et al. 2000;Schulte et al. 2009). The Stroop effect can be conceptuallyconsidered as a left hemisphere lateralized task with greaterinterference arising from the semantic content of the word(MacLeod 1991). Because of its preferentially processingof verbal information (e.g., language, speech, and writing),the left hemisphere should be more affected than the rightby semantic competition (Luo et al. 1999). This lateralityeffect was demonstrated in a study (Weekes and Zaidel1996), which used separate color words and color patchesand found greater Stroop interference when color wordswere presented to the left than right hemisphere indepen-dent of the location of the color patch (i.e., within the samehemifield or in the opposite hemifield).
In our lateralized Stroop match-to-sample task, weanalyzed hemispheric preference for Stroop word-colorinformation by comparing reaction times to right visualfield trials (preferred left cerebral hemisphere for wordprocessing) with reaction times to left visual field trials(non-preferred right hemisphere). We found that controlsubjects and patients with chronic alcoholism differed inperformance as a function of lateralized stimulus presenta-tions and attentional processes when color cues matched ordid not match the word’s color (Schulte et al. 2006b): Formatching colors, both groups showed a right visual fieldadvantage; for nonmatching colors, however, controlsshowed a left visual field advantage, whereas alcoholicsshowed no visual field advantage. We speculated thatcontrols were more successful than alcoholics in disengag-ing their attention from the invalidly cued color to correctlyprocess the color of the Stroop stimulus in nonmatch colortrials. Subsequent disengagement and shift of attentionaway from the invalidly cued color to the correct target colorof the Stroop stimulus is associated with right-hemispherefunctions (Blumstein and Cooper 1974; Christman 2001;Hartje et al. 1985; Posner 1980) and may be responsible forthe left visual field advantage in nonmatch trials observed inhealthy controls. This left visual field advantage in nonmatchtrials was correlated with a larger callosal splenium area incontrols but not alcoholics indicating that informationpresented to the non-preferred hemisphere is transmittedvia the splenium to the hemisphere that is specialized forefficient processing. This processing route for visual hemi-field information was disrupted in alcoholics, possibly as aresult of callosal thinning.
Furthermore, our visual world can be decomposed intocomponent features and then integrated into more complexstimuli, objects and scenes—a concept originated from inves-tigations of the visual cortex (Felleman et al. 1997; Pandya andSanides 1973). Thus, whole-part perception is based on the
extraction of higher-order or global features (such as thesilhouette of a face, or street scene) and details on the locallevel (e.g., facial details like eyes and nose, or details in a streetscene like cars and traffic lights). A widely used paradigm tostudy whole-part perception is a target detection task that useshierarchical letters, where a large (global) letter is made ofsmaller (local) letters, modeling the hierarchical structure ofvisual world scenes (Fink et al. 1997; Navon 1977).
Current models of local-global perception assume thatlocal and global processes proceed in parallel, with a lefthemisphere advantage in processing local features and aright hemisphere advantage in processing the global“gestalt”, as indicated by brain lesion studies (Delis et al.1986, 1988; Lamb et al. 1989; Robertson et al. 1988, 1991;Yamaguchi et al. 2000) and neuroimaging studies in healthysubjects (Evans et al. 2000; Fink et al. 1996, 1999; Han etal. 2002; Heinze et al. 1998; Weber et al. 2000; Yamaguchiet al. 2000). Thus, the study of local-global perception maycontribute to our understanding of the role of the CC forhigher-order visual integration.
Although RT studies using lateralized hemifield presen-tations have not always shown local-global hemisphericdifferences (Blanca et al. 1994; Evert and Kmen 2003;Hübner 1997; Yovel et al. 2001), electrophysiologicalstudies using central stimulus presentations do support thishemispheric processing bias for left-local and right-globalinformation (Heinze and Münte 1993; Heinze et al. 1998;Malinowski et al. 2002; Yamaguchi et al. 2000). Together,these studies suggest that, in principle, each hemisphere canprocess local and global information; however, hemisphericprocessing advantages emerge when local and globalinformation is processed simultaneously in both hemi-spheres. The competition hypothesis (Han et al. 2002;Volberg and Hübner 2004) proposes that with centralstimulation both hemispheres have simultaneous access tothe same visual information and the specialized hemisphereassigns more resources to a given local or global targetlevel bringing hemispheric differences for local and globalfeature processing to light (Fig. 3). With lateralizedpresentations, the hemisphere contralateral to the stimulatedhemifield receives the visual information prior to theipsilateral hemisphere that receives information after inter-hemispheric transfer through the CC. The resulting timedifference in processing between hemispheres may elimi-nate interhemispheric competition, and also hemisphericdifferences associated with local and global feature pro-cessing. Results from an ERP study (Han et al. 2002) thatfound hemispheric differences only for centrally presentedhierarchical stimuli and not for lateralized stimulus pre-sentations, are consistent with the competition hypothesis.
Local-global paradigms have yielded two measures: (a)the difference in response times to local and global targets,
182 Neuropsychol Rev (2010) 20:174–190

or precedence effect, which indexes parallel local andglobal processes assuming independent processing channelsfor local and global features, and (b) the difference inresponse times to incongruent (e.g., a global F made up oflocal Ts) and congruent stimuli (e.g., a global T made up oflocal Ts), or interference effect, which indexes local-globalinhibition in situations of competition. Several studiesindicate a global-over-local precedence effect, i.e., fasterresponses to global than local features, and global interfer-ence, i.e., slower responses when the global feature wasincongruent than when it was congruent with the localfeature (Navon 1977). However, local precedence and localinterference have also been reported (Müller-Oehring et al.2007). Which level—local or global—is processed fasterdepends on stimulus size (Kinchla and Wolfe 1979), thesize ratio between global and local forms (Kimchi andPalmer 1982), the stimulus’ location in the visual field(central or peripheral) (Kimchi 1992; Lamb and Robertson1987, 1988), and attentional selection (Broadbent 1977;Han et al. 2001; Pomerantz 1983; Robertson et al. 1993;Weissman et al. 2005; Yamaguchi et al. 2000).
Up to date, only a few studies have investigated the roleof the CC in local-global precedence and interference. In1993, Robertson et al. found that split-brain patients did notshow global interference despite normal global precedence.However, Weekes et al. (1997) repeated the experiment intwo of the same split-brain patients and found bothpreserved global precedence and global interference. Thus,in contrast to Robertson et al. (1993), their findings implythat the CC is not necessary for eliciting global interference.Rather, because each hemisphere can process local andglobal information, interference effects occur within one
hemisphere in the absence of the CC. The different resultsfrom these two studies may be due to slight differences instimulus characteristics and presentation: Robertson et al.(1993) used stimulus eccentricities of 2.7° while Weekes etal. (1997) used stimulus eccentricities of 1° for each visualhemifield. Given that there is a 1–2° stripe overlap at theretinal vertical meridian between the two visual hemifields,the smaller stimulus eccentricity and enhanced stimulus-background contrast used by Weekes et al. (1997) may havein fact sent information to the left and right visual corticesand contributed to intrahemispheric local-global competi-tion. Thus neither study provides clear evidence for callosalinvolvement in local-global interference.
Our own recent studies in normal aging (Müller-Oehringet al. 2007), alcoholism (Müller-Oehring et al. 2009b), andHIV infection (Müller-Oehring et al. 2010), all conditionswith callosal thinning or impaired integrity, provide newevidence for CC involvement in local-global processing. Inhealthy aging, for example, we found that larger localprecedence effects and greater local interference correlatedwith smaller callosal genu area (Müller-Oehring et al.2007). In addition to precedence and interference, we alsotested the relation between the CC and local-globalfacilitation. Local-global facilitation effects have beenfound previously for attentional cueing (Lamb and Yund2000; Robertson et al. 1993), and repetitive priming (Hanet al. 2003; Schatz and Erlandson 2003). We found, that inhealthy subjects posterior callosal splenium area wasassociated with local-global facilitation from repetitionpriming independent of age (Müller-Oehring et al. 2007).Converging evidence for a differentiated role of the CC inlocal-global integration comes from two recent DTI studieson chronic alcoholism (Müller-Oehring et al. 2009b) andHIV-infection (Müller-Oehring et al. 2010) indicating thatlocal-global interference is mediated by anterior callosalgenu integrity and local-global facilitation by posteriorcallosal splenium integrity.
Together, these findings suggest that the CC mediateslateralized higher-order cognition such as whole-partperceptual integration. Specifically, anterior callosal integ-rity appears to mediate local-global interference (callosalinhibition), whereas posterior callosal integrity seems tomediate local-global facilitation (callosal cooperation).Thus, component processes of visuospatial perception andattention are attributable, at least in part, to the integrity ofcallosal pathways, relevant for the integration of lateralizedbrain functions.
Conclusion
Geschwind (1965) formulated the idea that many neuro-logical disorders can be understood as ‘disconnections
Fig. 3 Model of whole-partperception: Parallel intrahemi-spheric and transcallosal inter-hemispheric processing of localand global features. With centralstimulation, the specializedhemisphere assigns moreresources to a given local orglobal target level promotinghemispheric differences withglobal features (e.g., big letter F)preferentially processed in theright cerebral hemisphere andlocal features (e.g., smallletters T) in the left hemisphere(competition hypothesis)
Neuropsychol Rev (2010) 20:174–190 183

syndromes’ in which white matter damage degradesnetwork functions leading to specific cognitive disabilitiessuch as visuospatial neglect (Doricchi et al. 2008).Numerous studies have since contributed to our currentunderstanding of interhemispheric sharing of informationfrom early visual input to complex decision-making. Manyof these studies have focused on split-brain patients whosecerebral commissures had been surgically disconnected toimprove intractable epilepsy (for reviews see Gazzaniga2000, 2005; Lassonde and Ouimet 2010). The recentadvance of neuroimaging techniques such as DTI andfMRI have provided the opportunity to study CC functionin situations of callosal degradation without disconnection.These studies have documented a central role of the CC forinterhemispheric visuomotor integration and for higher-order cognitive functions, and have shown that even subtledegradation of the CC in neurologically impaired patientscan be related to deficits in the transfer of informationbetween the hemispheres. For visuomotor integration, split-brain research suggests that the CC exerts an inhibitoryfunction on the hemispheres and that absence of callosalinhibition results in enhanced bilateral processing advan-tages. Recent DTI and fMRI studies that tested bihemi-spheric visuomotor integration in the connected brain,however, provide evidence for callosal cooperation. Whetherthe specific role of the CC for visuomotor integration isinhibitory or cooperative remains unclear. At this point, itappears plausible to assume that callosal inhibition andcooperation are not mutually exclusive but depend oncomplex interactions within a subcortico-cortical networkthat probably equilibrate hemispheric activation according totask demands.
A further aim of this review was to focus on the role ofthe CC for higher-order lateralized cognitive functions.Together, the reviewed research provides clear indicationthat the CC contributes to the integration of perception andaction, promoting a unified experience of the way weperceive the visual world and prepare our actions. Ofparticular interest is emerging evidence from neuroimagingthat the CC employs a differentiated role with callosal areastransmitting different types of information depending on thecortical destination of connecting fibers. Studies on patientswith callosal degradation indicated that anterior callosalfibers linking frontal and premotor areas of the twohemispheres were associated with inhibitory functions insituations of semantic competition (Stroop) and local-globalinterference, whereas posterior callosal areas connectingtemporo-parietal and occipital cortical regions were relatedwith facilitation from redundant targets and local-globalfeatures. Thus, to achieve an interhemispheric balancebetween component brain functions, the CC appears toexert both functional inhibition and excitation (Bloom andHynd 2005).
Future directions should include new imaging techniquessuch as fiber tractography in combination with functionalneuroimaging and electrophysiological methods to explorethe specific role of regional callosal connectivity for brainfunctions in the healthy brain and callosal degradation.
Acknowledgments We thank Margaret J. Rosenbloom for com-ments on the manuscript. Preparation of this article was supported byNational Institutes of Health research grants: AA018022, AA010723,AA005965, AA012388, AA017168, AA017432
Disclosures The authors declare that no conflicts of interest areassociated with the preparation of this article.
References
Aboitiz, F., Scheibel, A. B., Fisher, R. S., & Zaidel, E. (1992).Individual differences in brain asymmetries and fiber composition inthe human corpus callosum. Brain Research, 598, 154–161.
Aglioti, S., Smania, N., Manfredi, M., & Berlucchi, G. (1996).Disownership of left hand and objects related to it in a patientwith right brain damage. Neuroreport, 20, 293–296.
Aralasmak, A., Ulmer, J. L., Kocak, M., Salvan, C. V., Hillis, A. E., &Yousem, D. M. (2006). Association, commissural, and projectionpathways and their functional deficit reported in literature.Journal of Computer Assisted Tomography, 30, 695–715.
Aziz-Zadeh, L., Koski, L., Zaidel, E., Mazziotta, J., & Iacoboni, M.(2006). Lateralization of the human mirror neuron system.Journal of Neuroscience, 26, 2964–2970.
Baird, A. E., & Warach, S. (1998). Magnetic resonance imaging ofacute stroke. Journal of Cerebral Blood Flow & Metabolism, 18,583–609.
Banich, M. T. (1995). Interhemispheric processing: Theoretical andempirical considerations. In R. Davidson & K. Hugdahl (Eds.),Brain asymmetry (pp. 427–450). Cambridge: MIT.
Banich, M. T. (1998). The missing link: the role of interhemisphericinteraction in attentional processing. Brain and Cognition, 36,128–157.
Banich, M. T., & Belger, A. (1990). Interhemispheric interaction:how do the hemispheres divide and conquer a task? Cortex, 26,77–94.
Banich, M. T., & Shenker, J. (1994). Investigations of interhemisphericprocessing: methodological considerations. Neuropsychology, 8,263–277.
Banich, M. T., Milham, M. P., Atchley, R., Cohen, N. J., Webb, A.,Wszalek, T., et al. (2000). fMRI studies of Stroop tasks revealunique roles of anterior and posterior brain systems in attentionalselection. Journal of Cognitive Neuroscience, 12, 988–1000.
Barnett, K. J., & Corballis, M. C. (2005). Speeded right-to-leftinformation transfer: the result of speeded transmission in right-hemisphere axons? Neuroscience Letters, 380, 88–92.
Bartzokis, G., Sultzer, D., Lu, P. H., Nuechterlein, K. H., Mintz, J., &Cummings, J. L. (2004). Heterogeneous age-related breakdownof white matter structural integrity: implications for cortical“disconnection” in aging and Alzheimer’s disease. Neurobiologyof Aging, 25, 843–851.
Bartzokis, G., Lu, P. H., Tingus, K., Mendez, M. F., Richard, A.,Peters, D. G., et al. (2008). Lifespan trajectory of myelin integrityand maximum motor speed. Neurobiology of Aging. In press.
Bashore, T. R. (1981). Vocal and manual reaction time estimates ofinterhemispheric transmission time. Psychology Bulletin, 89,352–368.
184 Neuropsychol Rev (2010) 20:174–190

Battelli, L., Cavanagh, P., Martini, P., & Barton, J. J. (2003). Bilateraldeficits of transient visual attention in right parietal patients.Brain, 126, 2164–2174.
Bauer, R. M. (1982). Visual hypoemotionality as a symptom ofvisual-limbic disconnection in man. Archives of Neurology, 39,702–708.
Bayard, S., Gosselin, N., Robert, M., & Lassonde, M. (2004).Inter-and intra-hemispheric processing of visual event-relatedpotentials in the absence of the corpus callosum. Journal ofCognitive Neuroscience, 16, 401–414.
Berlucchi, G., Heron, W., Hyman, R., Rizzolatti, G., & Umilta, C.(1971). Simple reaction times of ipsilateral and contralateral handto lateralized visual stimuli. Brain, 94, 419–430.
Berlucchi, G., Aglioti, S., Marzi, C. A., & Tassinari, G. (1995).Corpus callosum and simple visuomotor integration. Neuro-psychologia, 33, 923–936.
Blanca, M. J., Zalabardo, C., Garcia-Criado, F., & Siles, R.(1994). Hemispheric differences in global and local process-ing dependent on exposure duration. Neuropsychologia, 32,1343–1351.
Bloom, J. S., & Hynd, G. W. (2005). The role of the corpus callosum ininterhemispheric transfer of information: excitation or inhibition?Neuropsychology Review, 15, 59–71.
Blumstein, S., & Cooper, W. E. (1974). Hemispheric processing ofintonation contours. Cortex, 10, 146–158.
Braun, C. M., Daigneault, S., Dufresne, A., Miljours, S., & Collin, I.(1995). Does so-called interhemispheric transfer time depend onattention? American Journal of Psychology, 108, 527–546.
Broadbent, D. E. (1977). The hidden preattentive process. AmericanPsychologist, 32, 109–118.
Bucur, B., Madden, D. J., & Allen, P. A. (2005). Age-relateddifferences in the processing of redundant visual dimensions.Psychology and Aging, 20, 435–446.
Caminiti, R., Ghaziri, H., Galuske, R., Hof, P. R., & Innocenti, G. M.(2009). Evolution amplified processing with temporally dis-persed slow neuronal connectivity in primates. Proceedings ofthe National Academy of Sciences of the United States ofAmerica, 106, 19551–19556.
Carter, C. S., MacDonald, A. M., Botvinick, M., Ross, L. L., Stenger,V. A., Noll, D., et al. (2000). Parsing executive processes:strategic vs. evaluative functions of the anterior cingulate cortex.Proceedings of the National Academy of Sciences of the UnitedStates of America, 97, 1944–1948.
Catani, M., & ffytche, D. H. (2005). The rises and falls ofdisconnection syndromes. Brain, 128, 2224–2239.
Chiarello, C., & Maxfield, L. (1996). Varieties of interhemisphericinhibition, or how to keep a good hemisphere down. Brain andCognition, 30, 81–108.
Christman, S. D. (2001). Individual differences in stroop and local-global processing: a possible role of interhemispheric interaction.Brain and Cognition, 45, 97–118.
Clarke, J. M., & Zaidel, E. (1989). Simple reaction times to lateralizedlight flashes. Varieties of interhemispheric communication routes.Brain, 112, 849–870.
Clarke, J. M., & Zaidel, E. (1994). Anatomical-behavioral relationships:corpus callosum morphometry and hemispheric specialization.Behavioural Brain Research, 64, 185–202.
Cook, N. D. (1984). Callosal inhibition: the key to the brain code.Behavioral Science, 29, 98–110.
Corballis, M. C. (1998). Interhemispheric neural summation in theabsence of the corpus callosum. Brain, 121, 1795–1807.
Corballis, M. C. (2002). Hemispheric interactions in simple reactiontime. Neuropsychologia, 40, 423–434.
Corballis, M. C., Hamm, J. P., Barnett, K. J., & Corballis, P. M.(2002). Paradoxical interhemispheric summation in the splitbrain. Journal of Cognitive Neuroscience, 14, 1151–1157.
Corballis, M. C., Corballis, P. M., & Fabri, M. (2003). Redundancygain in simple reaction time following partial and completecallosotomy. Neuropsychologia, 42, 71–81.
Corbetta, M., Shulman, G. L., Miezin, F. M., & Petersen, S. E. (1995).Superior parietal cortex activation during spatial attention shiftsand visual feature conjunction. Science, 270, 802–805.
Corbetta, M., Tansy, A. P., Stanley, C. M., Astafiev, S. V., Snyder, A. Z.,& Shulman, G. L. (2005). A functional MRI study of preparatorysignals for spatial location and objects. Neuropsychologia, 43,2041–2056.
Creem-Regehr, S. H. (2009). Sensory-motor and cognitive functionsof the human posterior parietal cortex involved in manualactions. Neurobiology of Learning and Memory, 91, 166–171.
Damasio, A. R., & Damasio, H. (1993). Brain and language, In“Mind and Brain”, a Scientific American Book. New York:Freeman.
Doricchi, F., Thiebaut de Schotten, M., Tomaiuolo, F., & Bartolomeo,P. (2008). White matter (dis)connections and gray matter (dys)functions in visual neglect: gaining insights into the brainnetworks of spatial awareness. Cortex, 44, 983–995.
Dean, P., Redgrave, P., & Westby, G. W. (1989). Event or emergency?Two response systems in the mammalian superior colliculus.Trends in Neurosciences, 12, 137–147.
Delis, D. C., Robertson, L. C., & Efron, R. (1986). Hemisphericspecialization of memory for visual hierarchical stimuli. Neuro-psychologia, 24, 205–214.
Delis, D. C., Kiefner, M., & Fridlund, A. J. (1988). Visuospatialdysfunction following unilateral brain damage: dissociations inhierarchical and hemispatial analysis. Journal of Clinical andExperimental Neuropsychology, 10, 421–431.
Diederich, A., & Colonius, H. (1987). Intersensory facilitation in themotor component? A reaction time analysis. PsychologicalResearch, 49, 23–29.
Doricchi, F., & Tomaiuolo, F. (2003). The anatomy of neglect withouthemianopia: a key role for parietal-frontal disconnection? Neuro-report, 14, 2239–2243.
Doron, K. W., & Gazzaniga, M. S. (2008). Neuroimaging techniquesoffer new perspectives on callosal transfer and interhemisphericcommunication. Cortex, 44, 1023–1029.
Duffau, H., Capelle, L., Sichez, N., Denvil, D., Lopes, M., Sichez, J.P., et al. (2002). Intraoperative mapping of the subcorticallanguage pathways using direct stimulations. An anatomo-functional study. Brain, 125, 199–214.
Duara, R., Kushch, A., Gross-Glenn, K., Barker, W. W., Jallad, B.,Pascal, S., et al. (1991). Neuroanatomic differences betweendyslexic and normal readers on magnetic resonance imagingscans. Archives of Neurology, 48, 410–416.
Evans, M. A., Shedden, J. M., Hevenor, S. J., & Hahn, M. C. (2000).The effect of variability of unattended information on global andlocal processing: evidence for lateralization at early stages ofprocessing. Neuropsychologia, 38, 225–239.
Evert, D. L., & Kmen, M. (2003). Hemispheric asymmetries for globaland local processing as a function of stimulus exposure duration.Brain and Cognition, 51, 115–142.
Fabri, M., Polonara, G., Del Pesce, M., Quattrini, A., Salvolini, U., &Manzoni, T. (2001). Posterior corpus callosum and interhemi-spheric transfer of somatosensory information: an fMRI andneuropsychological study of a partially callosotomized patient.Journal of Cognitive Neuroscience, 13, 1071–1079.
Felleman, D. J., Burkhalter, A., & Van Essen, D. C. (1997). Corticalconnections of areas V3 and VP of macaque monkey extrastriatevisual cortex. The Journal of Comparative Neurology, 379, 21–47.
Fendrich, R., Hutsler, J. J., & Gazzaniga, M. S. (2004). Visual andtactile interhemispheric transfer compared with the method ofPoffenberger. Experimental Brain Research, 158, 67–74.
Neuropsychol Rev (2010) 20:174–190 185

Fink, G. R., Halligan, P. W., Marshall, J. C., Frith, C. D., Frackowiak,R. S. J., & Dolan, R. J. (1996). Where in the brain does visualattention select the forest and the trees? Nature, 382, 626–628.
Fink, G. R., Halligan, P. W., Marshall, J. C., Frith, C. D., Frackowiak,R. S., & Dolan, R. J. (1997). Neural mechanisms involved in theprocessing of global and local aspects of hierarchically organizedvisual stimuli. Brain, 120, 1779–1791.
Fink, G. R., Marshall, J. C., Halligan, P. W., & Dolan, R. J. (1999).Hemispheric asymmetries in global/local processing are modu-lated by perceptual salience. Neuropsychologia, 37, 31–40.
Forster, B., & Corballis, M. C. (1998). Interhemispheric transmissiontimes in the presence and absence of the forebrain commissures:effects of luminance and equiluminance. Neuropsychologia, 36,925–934.
Forster, B., & Corballis, M. C. (2000). Interhemispheric transfer ofcolour and shape information in the presence and absence of thecorpus callosum. Neuropsychologia, 38, 32–45.
Galaburda, A. M., & Geschwind, N. (1980). The human languageareas and cerebral asymmetries. Revue Medicale de la SuisseRomande, 100, 119–128.
Gazzaniga, M. S. (2000). Cerebral specialization and interhemisphericcommunication: does the corpus callosum enable the humancondition? Brain, 123, 1293–1326.
Gazzaniga, M. S. (2005). Forty-five years of split-brain research andstill going strong. Nature Reviews Neuroscience, 6, 653–659.
Geschwind, N. (1965). Disconnexion syndromes in animals and man.I. Brain, 88, 237–294.
Giray, M., & Ulrich, R. (1993). Motor coactivation revealed byresponse force in divided and focused attention. Journal ofExperimental Psychology: Human Perception and Performance,19, 1278–1291.
Gur, R. C., Packer, I. K., Hungerbuhler, J. P., Reivich, M., Obrist, W.D., Amarnek, W. S., et al. (1980). Differences in the distributionof gray and white matter in human cerebral hemispheres. Science,207, 1226–1228.
Habib, M. (1986). Visual hypoemotionality and prosopagnosiaassociated with right temporal lobe isolation. Neuropsychologia,24, 577–582.
Hagmann, P., Thiran, J. P., Jonasson, L., Vandergheynst, P., Clarke, S.,Maeder, P., et al. (2003). DTI mapping of human brainconnectivity: statistical fibre tracking and virtual dissection.Neuroimage, 19, 545–554.
Han, S., He, X., Yund, E. W., & Woods, D. L. (2001). Attentionalselection in the processing of hierarchical patterns: an ERP study.Biological Psychology, 5, 31–48.
Han, S., Weaver, J. A., Murray, S. O., Kang, X., Yund, E. W., &Woods, D. L. (2002). Hemispheric asymmetry in global/localprocessing: effects of stimulus position and spatial frequency.Neuroimage, 17, 1290–1299.
Han, S., Yund, E. W., & Woods, D. L. (2003). An ERP study of theglobal precedence effect: the role of spatial frequency. ClinicalNeurophysiology, 114, 1850–1865.
Hanajima, R., Ugawa, Y., Machii, K., Mochizuki, H., Terao, Y.,Enomoto, H., et al. (2001). Interhemispheric facilitation of thehand motor area in humans. Journal of Physiology, 531, 849–859.
Harper, C. G., & Kril, J. J. (1988). Corpus callosal thickness inalcoholics. British Journal of Addiction, 83, 577–580.
Harper, C. G., & Kril, J. J. (1990). Neuropathology of alcoholism.Alcohol and Alcoholism, 25, 207–216.
Harper, C., & Matsumoto, I. (2005). Ethanol and brain damage.Current Opinion in Pharmacology, 5, 73–78.
Hartje, W., Willmes, K., & Weniger, D. (1985). Is there parallel andindependent hemispheric processing of intonational and phoneticcomponents of dichotic speech stimuli? Brain and Language, 24,83–99.
Heinze, H. J., & Münte, T. F. (1993). Electrophysiological correlatesof hierarchical stimulus processing: dissociation between onsetand later stages of global and local target processing. Neuro-psychologia, 31, 841–852.
Heinze, H. J., Hinrichs, H., Scholz, M., Burchert, W., & Mangun, G. R.(1998). Neural mechanisms of global and local processing. Acombined PET and ERP study. Journal of Cognitive Neuroscience,10, 485–498.
Hiatt, K. D., & Newman, J. P. (2007). Behavioral evidence ofprolonged interhemispheric transfer time among psychopathicoffenders. Neuropsychology, 21, 313–318.
Hofer, S., & Frahm, J. (2006). Topography of the human corpuscallosum revisited-comprehensive fiber tractography using diffusiontensor magnetic resonance imaging. Neuroimage, 32, 989–994.
Hopkins, W. D., & Rilling, J. K. (2000). A comparative MRI studyof the relationship between neuroanatomical asymmetry andinterhemispheric connectivity in primates: implication for theevolution of functional asymmetries. Behavioral Neuroscience,114, 739–748.
Hübner, R. (1997). The effect of spatial frequency on globalprecedence and hemispheric differences. Perception & Psycho-physics, 59, 187–201.
Hugdahl, K., & Davidson, R. J. (2003). The asymmetrical brain(pp. 259–302). Cambridge: MIT.
Hutner, N., & Oscar-Berman, M. (1996). Visual laterality patterns forthe perception of emotional words in alcoholic and agingindividuals. Journal of Studies on Alcohol, 57, 144–154.
Iacoboni, M., & Zaidel, E. (1995). Channels of the corpus callosum.Evidence from simple reaction times to lateralized flashes in thenormal and the split brain. Brain, 118, 779–788.
Iacoboni, M., & Zaidel, E. (2003). Interhemispheric visuo-motorintegration in humans: the effect of redundant targets. EuropeanJournal of Neuroscience, 17, 1981–1986.
Iacoboni, M., & Zaidel, E. (2004). Interhemispheric visuo-motorintegration in humans: the role of the superior parietal cortex.Neuropsychologia, 42, 419–425.
Iacoboni, M., Ptito, A., Weekes, N. Y., & Zaidel, E. (2000). Parallelvisuomotor processing in the split brain: cortico-subcorticalinteractions. Brain, 123, 759–769.
Ignashchenkova, A., Dicke, P. W., Haarmeier, T., & Their, P. (2004).Neuron-specific contribution of the superior colliculus to overtand covert shifts of attention. Nature Neuroscience, 7, 56–64.
Innocenti, G. M. (1986). Postnatal development of corticocorticalconnections. Italian Journal of Neurological Sciences, Suppl, 5,25–28.
Innocenti, G. M. (1995). Exuberant development of connections, andits possible permissive role in cortical evolution. Trends inNeurosciences, 18, 397–402.
Iwabuchi, S. J., & Kirk, I. J. (2009). Atypical interhemisphericcommunication in left-handed individuals. Neuroreport, 20, 166–169.
Jancke, L., & Steinmetz, H. (1994). Interhemispheric transfer time andcorpus callosum size. Neuroreport, 5, 2385–2388.
Jeeves, M. A., & Moes, P. (1996). Interhemispheric transfer timedifferences related to aging and gender. Neuropsychologia, 34,627–636.
Keary, C. J., Minshew, N. J., Bansal, R., Goradia, D., Fedorov, S.,Keshavan, M. S., et al. (2009). Corpus callosum volume andneurocognition in autism. Journal of Autism and DevelopmentalDisorders, 39, 834–841.
Kennedy, K. M., & Raz, N. (2009). Aging white matter and cognition:differential effects of regional variations in diffusion propertieson memory, executive functions, and speed. Neuropsychologia,47, 916–927.
Kimchi, R. (1992). Primacy of wholistic processing and global/localparadigm: a critical review. Psychological Bulletin, 112, 24–38.
186 Neuropsychol Rev (2010) 20:174–190

Kimchi, R., & Palmer, S. E. (1982). Form and texture in hierarchicallyconstructed patterns. Journal of Experimental Psychology:Human Perception and Performance, 8, 521–535.
Kinchla, R. A., & Wolfe, J. M. (1979). The order of visual processing:“Top–down,” “bottom–up”, or “middle–out”. Perception &Psychophysics, 25, 225–231.
Kinsbourne, M. (1977). Hemi-neglect and hemisphere rivalry. Advancesin Neurology, 18, 41–49.
Kinsbourne, M. (2003). The corpus callosum equilibrates hemisphericactivation. In E. Zaidel & M. Iacoboni (Eds.), The parallel brain:The cognitive neuroscience of the corpus callosum (pp. 271–281). Cambridge: MIT.
Kinsbourne, M., & Hicks, R. B. (1978). Functional cerebral space: Amodel for overflow, transfer and interference effects in humanperformance: A tutorial review. In J. Requin (Ed.), Attention andperformance VII. Hillsdale: Erlbaum.
Kinsbourne, M. (1993). Orientational bias model of unilateral neglect:Evidence from attentional gradients within hemispace. In I. H.Robertson & J. C. Marshall (Eds.), Unilateral neglect: Clinicaland experimental studies (pp. 63–86). Hillsdale, NJ: Erlbaum.
Kinsbourne, M., & Bruce, R. (1987). Shift in visual laterality withinblocks of trials. ACTA Psychologica (Amsterdam), 66, 139–155.
Klier, E. M., Wang, H., & Crawford, J. D. (2003). Three-dimensionaleye-head coordination is implemented downstream from thesuperior colliculus. Journal of Neurophysiology, 89, 2839–2853.
Kraus, M. F., Susmaras, T., Caughlin, B. P., Walker, C. J., Sweeney, J.A., & Little, D. M. (2007). White matter integrity and cognitionin chronic traumatic brain injury: a diffusion tensor imagingstudy. Brain, 130, 2508–2519.
Kubicki, M., Westin, C. F., Nestor, P. G., Wible, C. G., Frumin, M.,Maier, S. E., et al. (2003). Cingulate fasciculus integritydisruption in schizophrenia: a magnetic resonance diffusiontensor imaging study. Biological Psychiatry, 54, 1171–1180.
Lamb, M. R., & Robertson, L. C. (1987). Do response time advantageand interference reflect the order of processing of global-and local-level information? Perception & Psychophysics, 46, 254–258.
Lamb, M. R., & Robertson, L. C. (1988). The processing ofhierarchical stimuli: effects of retinal locus, locational uncertainty,and stimulus identity. Perception & Psychophysics, 44, 172–181.
Lamb, M. R., & Yund, E. W. (2000). The role of spatial frequency incued shifts of attention between global and local forms.Perception & Psychophysics, 62, 753–761.
Lamb, M. R., Robertson, L. C., & Knight, R. T. (1989). Attention andinterference in the processing of global and local information:effects of unilateral temporal–parietal junction lesions. Neuro-psychologia, 27, 471–483.
Lansberg, M. G., Thijs, V. N., O’Brien, M. W., Ali, J. O., deCrespigny, A. J., Tong, D. C., et al. (2001). Evolution of apparentdiffusion coefficient, diffusion-weighted, and T2-weighted signalintensity of acute stroke. American Journal of Neuroradiology,22, 637–644.
Lassonde, M., & Ouimet, C. (2010). The split-brain. Wiley Interdisci-plinary Reviews: Cognitive Science, 1, 191–202.
Le Bihan, D. (2003). Looking into the functional architecture ofthe brain with diffusion MRI. Nature Reviews Neuroscience, 4,469–480.
Le Bihan, D., Mangin, J. F., Poupon, C., Clark, C. A., Pappata, S., Molko,N., et al. (2001). Diffusion tensor imaging: concepts and applica-tions. Journal of Magnetic Resonance Imaging, 13, 534–546.
Liederman, J., & Meehan, P. (1986). When is between-hemispheredivision of labor advantageous? Neuropsychologia, 24, 863–874.
Lim, K. O., & Helpern, J. A. (2002). Neuropsychiatric applications ofDTI—a review. NMR in Biomedicine, 15, 587–593.
Livingstone, M. S., & Hubel, D. H. (1987). Psychophysical evidencefor separate channels for the perception of form, color,movement, and depth. Journal of Neuroscience, 7, 3416–3468.
Luders, E., Rex, D. E., Narr, K. L., Woods, R. P., Jancke, L.,Thompson, P. M., et al. (2003). Relationships between sulcalasymmetries and corpus callosum size: gender and handednesseffects. Cerebral Cortex, 13, 1084–1093.
Luo, Y. J., Hu, S., Weng, X. C., & Wei, J. H. (1999). Effects ofsemantic discrimination of Chinese words on N400 componentof event-related potentials. Perceptual and Motor Skills, 89, 185–193.
Lux, S., Thimm, M., Marshall, J. C., & Fink, G. R. (2006). Directedand divided attention during hierarchical processing in patientswith visuo-spatial neglect and matched healthy volunteers.Neuropsychologia, 44, 436–444.
MacDonald, A. W., 3rd, Cohen, J. D., Stenger, V. A., & Carter, C. S.(2000). Dissociating the role of the dorsolateral prefrontal andanterior cingulate cortex in cognitive control. Science, 288,1835–1838.
MacLeod, C. M. (1991). Half a century of research on the Stroopeffect: an integrative review. Psychological Bulletin, 109, 163–203.
Madden, D. J., Spaniol, J., Costello, M. C., Bucur, B., White, L. E.,Cabeza, R., et al. (2009). Cerebral white matter integrity mediatesadult age differences in cognitive performance. Journal ofCognitive Neuroscience, 21, 289–302.
Magat, M., & Brown, C. (2009). Laterality enhances cognition inAustralian parrots. Proceedings of the Royal Society. Biologicalsciences, 276, 4155–4162.
Malinowski, P., Hubner, R., Keil, A., & Gruber, T. (2002). Theinfluence of response competition on cerebral asymmetries forprocessing hierarchical stimuli revealed by ERP recordings.Experimental Brain Research, 144, 136–139.
Marchant, L. F., & McGrew, W. C. (1996). Laterality of limb functionin wild chimpanzees of Gombe National Park: comprehensivestudy of spontaneous activities. Journal of Human Evolution, 30,427–443.
Marks, N. L., & Hellige, J. B. (2003). Interhemispheric interaction inbilateral redundancy gain: effects of stimulus format. Neuropsy-chology, 17, 578–593.
Marzi, C. A., Bisiacchi, P., & Nicoletti, R. (1991). Is interhemispherictransfer of visuomotor information asymmetric? Evidence from ameta-analysis. Neuropsychologia, 29, 1163–1177.
Marzi, C. A., Smania, N., Martini, M. C., Gambina, G., Tomelleri, G.,Palamara, A., et al. (1996). Implicit redundant-targets effect invisual extinction. Neuropsychologia, 34, 9–22.
Marzi, C. A., Fanini, A., Girelli, M., Ipata, A. E., Miniussi, C.,Smania, N., et al. (1997). Is extinction following parietal damagean interhemispheric disconnection phenomenon? In P. Their & H.O. Karnath (Eds.), Parietal lobe contribution to orientation in 3Dspace. Heidelberg: Springer.
McCulloch, W. S., & Garol, H. W. (1941). Cortical origin anddistribution of corpus callosum and anterior commissure in themonkey (Macaca mulatta). Journal of Neurophysiology, 4, 555–563.
Meister, I. G., Wienemann, M., Buelte, D., Grunewald, C., Sparing, R.,Dambeck, N., et al. (2006). Hemiextinction induced by transcranialmagnetic stimulation over the right temporo-parietal junction.Neuroscience, 142, 119–123.
Miller, J. (1982). Divided attention: evidence for coactivation withredundant signals. Cognitive Psychology, 14, 247–279.
Miller, J. (1986). Timecourse of coactivation in bimodal dividedattention. Perception & Psychophysics, 40, 331–343.
Miller, J. (1991). Channel interaction and the redundant-targets effectin bimodal divided attention. Journal of Experimental Psychology:Human Perception and Performance, 17, 160–169.
Miller, J. O. (2004). Exaggerated redundancy gain in the split brain: Ahemispheric coactivation account. Cognitive Psychology, 49,118–154.
Neuropsychol Rev (2010) 20:174–190 187

Miller, J., & Ulrich, R. (2003). Simple reaction time and statisticalfacilitation: a parallel grains model. Cognitive Psychology, 46,101–151.
Miniussi, C., Girelli, M., & Marzi, C. A. (1998). Neural site of theredundant target effect electrophysiological evidence. Journal ofCognitive Neuroscience, 10, 216–230.
Moes, P. E., Brown, W. S., & Minnema, M. T. (2007). Individualdifferences in interhemispheric transfer time (IHTT) as measuredby event related potentials. Neuropsychologia, 45, 2626–2630.
Mooshagian, E., Kaplan, J., Zaidel, E., & Iacoboni, M. (2008). Fastvisuomotor processing of redundant targets: the role of the righttemporo-parietal junction. PLoS One, 3, e2348.
Mooshagian, E., Iacoboni, M., & Zaidel, E. (2009). Spatial attentionand interhemispheric visuomotor integration in the absence of thecorpus callosum. Neuropsychologia, 47, 933–937.
Mordkoff, J. T., & Yantis, S. (1991). An interactive race model ofdivided attention. Journal of Experimental Psychology: HumanPerception and Performance, 17, 520–538.
Mordkoff, J. T., & Yantis, S. (1993). Dividing attention between colorand shape: evidence of coactivation. Perception & Psychophysics,53, 357–366.
Mori, S., Kaufmann, W. E., Davatzikos, C., Stieltjes, B., Amodei, L.,Fredericksen, K., et al. (2002). Imaging cortical association tractsin the human brain using diffusion-tensor-based axonal tracking.Magnetic Resonance in Medicine, 47, 215–223.
Mort, D. J., Malhotra, P., Mannan, S. K., Rorden, C., Pambakian, A.,Kennard, C., et al. (2003). The anatomy of visual neglect. Brain,126, 1986–1997.
Moseley, M. E., Cohen, Y., Kucharczyk, J., Mintorovitch, J., Asgari,H. S., Wendland, M. F., et al. (1990). Diffusion-weighted MRimaging of anisotropic water diffusion in cat central nervoussystem. Radiology, 176, 439–445.
Muetzel, R. L., Collins, P. F., Mueller, B. A. M., Schissel, A., Lim, K. O.,& Luciana, M. (2008). The development of corpus callosummicrostructure and associations with bimanual task performance inhealthy adolescents. Neuroimage, 39, 1918–1925.
Müller-Oehring, E. M., Schulte, T., Raassi, C., Pfefferbaum, A., &Sullivan, E. V. (2007). Local-global interference is modulated byage, sex and anterior corpus callosum size. Brain Research, 1142,189–205.
Müller-Oehring, E. M., Schulte, T., Kasten, E., Poggel, D. A.,Müller, I., Wüstenberg, T., et al. (2009a). Parallel interhemi-spheric processing in hemineglect: relation to visual field defects.Neuropsychologia, 47, 2397–2408.
Müller-Oehring, E. M., Schulte, T., Fama, R., Pfefferbaum, A., &Sullivan, E. V. (2009b). Global-local interference is related tocallosal compromise in alcoholism: a behavior-DTI associationstudy. Alcoholism: Clinical and Experimental Research, 33, 477–489.
Müller-Oehring, E. M., Schulte, T., Rosenbloom, M. J., Pfefferbaum,A., & Sullivan, E. V. (2010). Callosal degradation in HIV-1infection predicts hierarchical perception: A DTI study. Neuro-psychologia, 48, 1133–1143.
Murray, M. M., Foxe, J. J., Higgins, B. A., Javitt, D. C., & Schroeder,C. E. (2001). Visuo-spatial neural response interactions in earlycortical processing during a simple reaction time task: a high-density electrical mapping study. Neuropsychologia, 39, 828–844.
Nassi, J. J., & Callaway, E. M. (2009). Parallel processing strategies ofthe primate visual system. Nature Reviews Neuroscience, 10,360–372.
Navon, D. (1977). Forest before trees: the precedence of globalfeatures in visual perception. Cognitive Psychology, 9, 353–383.
Neumann-Haefelin, T., Moseley, M. E., & Albers, G. W. (2000). Newmagnetic resonance imaging methods for cerebrovascular disease:emerging clinical applications. Annals of Neurology, 47, 559–570.
Nowicka, A., Grabowska, A., & Fersten, E. (1996). Interhemispherictransmission of information and functional asymmetry of thehuman brain. Neuropsychologia, 34, 147–151.
Ouimet, C., Jolicoeur, P., Miller, J., Ptito, A., Paggi, A., Foschi, N.,et al. (2009). Sensory and motor involvement in the enhancedredundant target effect: a study comparing anterior-and totallysplit-brain individuals. Neuropsychologia, 47, 684–692.
Pandya, D. N., & Sanides, F. (1973). Architectonic parcellation of thetemporal operculum in rhesus monkey and its projection pattern.Zeitschrift fur Anatomie und Entwicklungsgeschichte, 139, 127–161.
Park, H. J., Kim, J. J., Lee, S. K., Seok, J. H., Chun, J., Kim, D. I., etal. (2008). Corpus callosal connection mapping using corticalgray matter parcellation and DT-MRI. Human Brain Mapping,29, 503–516.
Paul, L. K., Brown, W. S., Adolphs, R., Tyszka, J. M., Richards, L. J.,Mukherjee, P., et al. (2007). Agenesis of the corpus callosum:genetic, developmental and functional aspects of connectivity.Nature Reviews Neuroscience, 8, 287–299.
Pfefferbaum, A., & Sullivan, E. V. (2002). Microstructural but notmacrostructural disruption of white matter in women withchronic alcoholism. Neuroimage, 15, 708–718.
Pfefferbaum, A., & Sullivan, E. V. (2003). Increased brain whitematter diffusivity in normal adult aging: relationship to anisotropyand partial voluming. Magnetic Resonance in Medicine, 49, 953–961.
Pfefferbaum, A., & Sullivan, E. V. (2005). Disruption of brain whitematter microstructure by excessive intracellular and extracellularfluid in alcoholism: evidence from diffusion tensor imaging.Neuropsychopharmacology, 30, 423–432.
Pfefferbaum, A., Adalsteinsson, E., & Sullivan, E. V. (2003).Replicability of diffusion tensor imaging measurements offractional anisotropy and trace in brain. Journal of MagneticResonance Imaging, 18, 427–433.
Pfefferbaum, A., Adalsteinsson, E., & Sullivan, E. V. (2006).Dysmorphology and microstructural degradation of the corpuscallosum: interaction of age and alcoholism. Neurobiology ofAging, 27, 994–1009.
Pierpaoli, C., & Basser, P. J. (1996). Toward a quantitative assessmentof diffusion anisotropy. Magnetic Resonance in Medicine, 36,893–906.
Poffenberger, A. T. (1912). Reaction time to retinal stimulation withspecial reference to time lost in conduction through nerves center.Archiv fur die gesamte Psychologie, 23, 1–173.
Pollmann, S., & Zaidel, E. (1998). The role of the corpus callosum invisual orienting: importance of interhemispheric visual transfer.Neuropsychologia, 36, 763–774.
Pomerantz, J. R. (1983). Global and local precedence: selectiveattention in form and motion perception. Journal of ExperimentalPsychology: General, 112, 516–540.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal ofExperimental Psychology, 32, 3–25.
Poupon, C., Mangin, J., Clark, C. A., Frouin, V., Régis, J., Le Bihan,D., et al. (2001). Towards inference of human brain connectivityfrom MR diffusion tensor data. Medical Image Analysis, 5, 1–15.
Pujol, J., López-Sala, A., Deus, J., Cardoner, N., Sebastián-Gallés, N.,Conesa, G., et al. (2002). The lateral asymmetry of the humanbrain studied by volumetric magnetic resonance imaging. Neuro-image, 17, 670–679.
Putnam, M. C., Wig, G. S., Grafton, S. T., Kelley, W. M., &Gazzaniga, M. S. (2008). Structural organization of the corpuscallosum predicts the extent and impact of cortical activity in thenondominant hemisphere. Journal of Neuroscience, 28, 2912–2918.
Putnam, M. C., Steven, M. S., Doron, K. W., Riggall, A. C., &Gazzaniga, M. S. (2009). Cortical projection topography of the
188 Neuropsychol Rev (2010) 20:174–190

human splenium: Hemispheric asymmetry and individual differ-ences. Journal of Cognitive Neuroscience. In press.
Raab, D. H. (1962). Statistical facilitation of simple reaction times.Transactions of the New York Academy of Sciences, 24, 574–590.
Rakic, P., & Yakovlev, P. I. (1968). Development of the corpuscallosum and cavum septi in man. Journal of ComparativeNeurology, 132, 45–72.
Reinholz, J., & Pollmann, S. (2007). Neural basis of redundancyeffects in visual object categorization. Neuroscience Letters, 412,123–128.
Reuter-Lorenz, P. A., & Stanczak, L. (2000). Differential effects ofaging on the functions of the corpus callosum. DevelopmentalNeuropsychology, 18, 113–137.
Reuter-Lorenz, P. A., Kinsbourne, M., & Moscovitch, M. (1990).Hemispheric control of spatial attention. Brain and Cognition,12, 240–266.
Reuter-Lorenz, P. A., Nozawa, G., Gazzaniga, M. S., & Hughes, H. C.(1995). Fate of neglected targets: a chronometric analysis ofredundant target effects in the bisected brain. Journal ofExperimental Psychology. Human Perception and Performance,21, 211–230.
Ringo, J. L., Doty, R. W., Demeter, S., & Simard, P. Y. (1994). Time isof the essence: a conjecture that hemispheric specialization arisesfrom interhemispheric conduction delay. Cerebral Cortex, 4,331–343.
Rizzo, M., & Vecera, S. P. (2002). Psychoanatomical substrates ofBálint’s syndrome. Journal of Neurology, Neurosurgery, andPsychiatry, 72, 162–178.
Robertson, L. C., Lamb, M. R., & Knight, R. T. (1988). Effects oflesions of temporal–parietal junction on perceptual and atten-tional processing in humans. Journal of Neuroscience, 8, 3757–3769.
Robertson, L. C., Lamb, M. R., & Knight, R. T. (1991). Normalglobal-local analysis in patients with dorsolateral frontal lobelesions. Neuropsychologia, 29, 959–967.
Robertson, L. C., Lamb, M. R., & Zaidel, E. (1993). Callosal transferand hemisphere laterality in response to hierarchical patterns:evidence from normal and commissurotomized subjects. Neuro-psychology, 7, 325–342.
Rohlfing, T., Zahr, N. M., Sullivan, E. V., & Pfefferbaum, A. (2010).The SRI24 multichannel atlas of normal adult human brainstructure. Human Brain Mapping. In press.
Roser, M., & Corballis, M. C. (2002). Interhemispheric neuralsummation in the split brain with symmetrical and asymmetricaldisplays. Neuropsychologia, 40, 1300–1312.
Roser, M., & Corballis, M. C. (2003). Interhemispheric neuralsummation in the split brain: effects of stimulus colour and task.Neuropsychologia, 41, 830–846.
Rugg,M. D., &Beaumont, J. G. (1978). Interhemispheric asymmetries inthe visual evoked response: effects of stimulus lateralisation andtask. Biological Psychology, 6, 283–292.
Ryberg, C., Rostrup, E., Stegmann, M. B., Barkhof, F., Scheltens, P.,van Straaten, E. C., et al. (2007). Clinical significance of corpuscallosum atrophy in a mixed elderly population. Neurobiology ofAging, 28, 955–963.
Salvan, C. V., Ulmer, J. L., DeYoe, E. A., Wascher, T., Mathews, V. P.,Lewis, J. W., et al. (2004). Visual object agnosia and pure wordalexia: correlation of functional magnetic resonance imaging andlesion localization. Journal of Computer Assisted Tomography,28, 63–67.
Saron, C. D., & Davidson, R. J. (1989). Visual evoked potentialmeasures of interhemispheric transfer time in humans. BehavioralNeuroscience, 103, 1115–1138.
Savazzi, S., & Marzi, C. A. (2002). Speeding up reaction time withinvisible stimuli. Current Biology, 12, 403–407.
Savazzi, S., & Marzi, C. A. (2004). The superior colliculus subservesinterhemispheric neural summation in both normals and patientswith a total section or agenesis of the corpus callosum. Neuro-psychologia, 42, 1608–1618.
Schatz, J., & Erlandson, F. (2003). Level-repetition effects inhierarchical stimulus processing: timing and location of corticalactivity. International Journal of Psychophysiology, 47, 255–269.
Schendel, K. L., & Robertson, L. C. (2002). Using reaction time toassess patients with unilateral neglect and extinction. Journal ofClinical and Experimental Neuropsychology, 24, 941–950.
Schiefer, U., Strasburger, H., Becker, S. T., Vonthein, R., Schiller, J.,Dietrich, T. J., et al. (2001). Reaction time in automated kineticperimetry: effects of stimulus luminance, eccentricity, andmovement direction. Vision Research, 41, 2157–2164.
Schmahmann, J. D., & Pandya, D. N. (2006). Corpus callosum. In J.D. Schmahmann & D. N. Pandya (Eds.), Fiber pathways of thebrain (pp. 485–496). New York: Oxford.
Schrift, M. J., Bandla, H., Shah, P., & Taylor, M. A. (1986).Interhemispheric transfer in major psychoses. Journal of Nervousand Mental Disease, 174, 203–207.
Schulte, T., Müller-Oehring, E. M., Strasburger, H., Warzel, H., &Sabel, B. A. (2001). Acute effects of alcohol on divided andcovert attention in men. Psychopharmacology, 154, 61–69.
Schulte, T., Pfefferbaum, A., & Sullivan, E. V. (2004). Parallelinterhemispheric processing in aging and alcoholism: relation tocorpus callosum size. Neuropsychologia, 42, 257–271.
Schulte, T., Sullivan, E. V., Muller-Oehring, E. M., Adalsteinsson, E.,& Pfefferbaum, A. (2005). Corpus callosal microstructuralintegrity influences interhemispheric processing: a diffusiontensor imaging study. Cerebral Cortex, 15, 1384–1392.
Schulte, T., Chen, S. H. A., Müller-Oehring, E. M., Adalsteinsson, E.,Pfefferbaum, A., & Sullivan, E. V. (2006a). fMRI Evidence forfrontal modulation of the extrastriate—dependent redundanttargets effect. Neuroimage, 30, 973–982.
Schulte, T., Müller-Oehring, E. M., Pfefferbaum, A., & Sullivan, E. V.(2006b). Callosal involvement in a lateralized Stroop task inalcoholic and healthy subjects. Neuropsychology, 20, 727–736.
Schulte, T., Müller-Oehring, E. M., Pfefferbaum, A., & Sullivan, E. V.(2008). Callosal compromise differentially affects conflict pro-cessing and attentional allocation in alcoholism, HIV-infection,and their comorbidity. Brain Imaging and Behavior, 2, 27–38.
Schulte, T., Müller-Oehring, E. M., Vinco, S., Hoeft, F., Pfefferbaum, A.,& Sullivan, E. V. (2009). Double dissociation between action-drivenand perception-driven conflict resolution invoking anterior versusposterior brain systems. Neuroimage, 48, 381–390.
Silvanto, J., Walsh, V., & Cowey, A. (2009). Abnormal functionalconnectivity between ipsilesional V5/MT+and contralesionalstriate cortex (V1) in blindsight. Experimental Brain Research,193, 645–650.
Shirani, P., Thorn, J., Davis, C., Heidler-Gary, J., Newhart, M.,Gottesman, R. F., et al. (2009). Severity of hypoperfusion indistinct brain regions predicts severity of hemispatial neglect indifferent reference frames. Stroke, 40, 3563–3566.
Skiba, M., Diekamp, B., Prior, H., & Güntürkün, O. (2000).Lateralized interhemispheric transfer of color cues: evidence fordynamic coding principles of visual lateralization in pigeons.Brain and Language, 73, 254–273.
Stephan, K. E., Marshall, J. C., Friston, K. J., Rowe, J. B., Ritzl, A.,Zilles, K., et al. (2003). Lateralized cognitive processes andlateralized task control in the human brain. Science, 301, 384–386.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions.Journal of Experimental Psychology, 12, 643–662.
Sullivan, E. V., & Pfefferbaum, A. (2003). Diffusion tensor imaging innormal aging and neuropsychiatric disorders. European Journalof Radiology, 45, 244–255.
Neuropsychol Rev (2010) 20:174–190 189

Sullivan, E. V., Adalsteinsson, E., Hedehus, M., Ju, C., Moseley, M.,Lim, K. O., et al. (2001). Equivalent disruption of regional whitematter microstructure in ageing healthy men and women.Neuroreport, 12, 99–104.
Sullivan, E. V., Pfefferbaum, A., Adalsteinsson, E., Swan, G. E., &Carmelli, D. (2002). Differential rates of regional brain change incallosal and ventricular size: a 4-year longitudinal MRI study ofelderly men. Cerebral Cortex, 12, 438–445.
Sullivan, E. V., Adalsteinsson, E., & Pfefferbaum, A. (2006).Selective age-related degradation of anterior callosal fiberbundles quantified in vivo with fiber tracking. CerebralCortex, 16, 1030–1039.
Sullivan, E. V., Rohlfing, T., & Pfefferbaum, A. (2010). Quantitativefiber tracking of lateral and interhemispheric white mattersystems in normal aging: relations to timed performance.Neurobiology of Aging, 31, 464–481.
Tanabe, H., Sawada, T., Inoue, N., Ogawa, M., Kuriyama, Y., &Shiraishi, J. (1987). Conduction aphasia and arcuate fasciculus.Acta Neurologica Scandinavica, 76, 422–427.
Tardif, E., & Clarke, S. (2002). Commissural connections of humansuperior colliculus. Neuroscience, 111, 363–372.
Tarnowska-Dziduszko, E., Bertrand, E., & Szpak, G. M. (1995).Morphological changes in the corpus callosum in chronicalcoholism. Folia Neuropathologica, 33, 25–29.
Tettamanti, M., Paulesu, E., Scifo, P., Maravita, A., Fazio, F., Perani, D.,et al. (2002). Interhemispheric transmission of visuomotor informa-tion in humans: fMRI evidence. Journal of Neurophysiology, 88,1051–1058.
Turatto, M., Mazza, V., Savazzi, S., & Marzi, C. A. (2004). The roleof the magnocellular and parvocellular systems in the redundanttarget effect. Experimental Brain Research, 158, 141–150.
Virta, A., Barnett, A., & Pierpaoli, C. (1999). Visualizing andcharacterizing white matter fiber structure and architecture inthe human pyramidal tract using diffusion tensor MRI. MagneticResonance Imaging, 17, 1121–1133.
Volberg, G., & Hübner, R. (2004). On the role of response conflictsand stimulus position for hemispheric differences in global/localprocessing: an ERP study. Neuropsychologia, 42, 1805–1813.
Wahl, M., Lauterbach-Soon, B., Hattingen, E., Jung, P., Singer, O.,Volz, S., et al. (2007). Human motor corpus callosum: topogra-phy, somatotopy, and link between microstructure and function.Journal of Neuroscience, 27, 12132–12138.
Warlop, N. P., Achten, E., Debruyne, J., & Vingerhoets, G. (2008).Diffusion weighted callosal integrity reflects interhemisphericcommunication efficiency in multiple sclerosis. Neuropsychologia,46, 2258–2264.
Weber, B., Schwarz, U., Kneifel, S., Treyer, V., & Buck, A. (2000).Hierarchical visual processing is dependent on the oculomotorsystem. NeuroReport, 11, 241–247.
Weber, B., Treyer, V., Oberholzer, N., Jaermann, T., Boesiger, P.,Brugger, P., et al. (2005). Attention and interhemispheric transfer:a behavioral and fMRI study. Journal of Cognitive Neuroscience,17, 113–123.
Weekes, N. Y., & Zaidel, E. (1996). The effects of proceduralvariations on lateralized Stroop effects. Brain and Cognition, 31,308–330.
Weekes, N. Y., Carusi, D., & Zaidel, E. (1997). Interhemisphericrelations in hierarchical perception: a second look. Neuropsycho-logia, 35, 37–44.
Weissman, D. H., Gopalakrishnan, A., Hazlett, C. J., & Woldorff, M.G. (2005). Dorsal anterior cingulate cortex resolves conflict fromdistracting stimuli by boosting attention toward relevant events.Cerebral Cortex, 15, 229–237.
Westerhausen, R., Kreuder, F., Woerner, W., Huster, R. J., Smit, C.M., Schweiger, E., et al. (2006). Interhemispheric transfer timeand structural properties of the corpus callosum. NeuroscienceLetters, 409, 140–145.
Westerhausen, R., Grüner, R., Specht, K., & Hugdahl, K. (2009).Functional relevance of interindividual differences in temporallobe callosal pathways: a DTI tractography study. CerebralCortex, 19, 1322–1329.
Witelson, S. F. (1989). Hand and sex differences in the isthmus andgenu of the human corpus callosum. A postmortem morpholog-ical study. Brain, 112, 799–835.
Woodruff, P. W., Phillips, M. L., Rushe, T., Wright, I. C., Murray,R. M., & David, A. S. (1997). Corpus callosum size and inter-hemispheric function in schizophrenia. Schizophrenia Re-search, 23, 189–196.
Yamaguchi, S., Yamagata, S., & Kobayashi, S. (2000). Cerebralasymmetry of the “top-down” allocation of attention to globaland local features. Journal of Neuroscience, 20, RC72.
Yamauchi, H., Fukuyama, H., Nagahama, Y., Katsumi, Y., Dong, Y.,Konishi, J., et al. (1997). Atrophy of the corpus callosum,cognitive impairment, and cortical hypometabolism in progres-sive supranuclear palsy. Annals of Neurology, 41, 606–614.
Yovel, G., Levy, J., & Yovel, I. (2001). Hemispheric asymmetries forglobal and local visual perception: effects of stimulus and taskfactors. Journal of Experimental Psychology: Human Perceptionand Performance, 27, 1369–1385.
Zahr, N. M., Rohlfing, T., Pfefferbaum, A., & Sullivan, E. V.(2009). Problem solving, working memory, and motor corre-lates of association and commissural fiber bundles in normalaging: a quantitative fiber tracking study. Neuroimage, 44,1050–1062.
Zaidel, E., & Iacoboni, M. (2003). Poffenberger’s simple reaction timeparadigm for measuring interhemispheric transfer time. In E.Zaidel & M. Iacoboni (Eds.), The parallel brain (pp. 1–8).Cambridge: MIT.
190 Neuropsychol Rev (2010) 20:174–190
Related Documents











