Large Molecule Therapeutics Construction and Characterization of Novel, Completely Human Serine Protease Therapeutics Targeting Her2/neu Yu Cao, Khalid A. Mohamedali, John W. Marks, Lawrence H. Cheung, Walter N. Hittelman, and Michael G. Rosenblum Abstract Immunotoxins containing bacterial or plant toxins have shown promise in cancer-targeted therapy, but their long-term clinical use may be hampered by vascular leak syndrome and immunogenicity of the toxin. We incorporated human granzyme B (GrB) as an effector and generated completely human chimeric fusion proteins containing the humanized anti-Her2/neu single-chain antibody 4D5 (designated GrB/4D5). Introduction of a pH-sensitive fusogenic peptide (designated GrB/4D5/26) resulted in comparatively greater specific cytotoxicity although both constructs showed similar affinity to Her2/ neu–positive tumor cells. Compared with GrB/4D5, GrB/4D5/26 showed enhanced and long-lasting cellular uptake and improved delivery of GrB to the cytosol of target cells. Treatment with nanomolar concentrations of GrB/4D5/26 resulted in specific cytotoxicity, induction of apoptosis, and efficient downregulation of PI3K/Akt and Ras/ERK pathways. The endogenous presence of the GrB proteinase inhibitor 9 did not impact the response of cells to the fusion construct. Surprisingly, tumor cells resistant to lapatinib or Herceptin, and cells expressing MDR-1 resistant to chemotherapeutic agents showed no cross-resistance to the GrB-based fusion proteins. Administration (intravenous, tail vein) of GrB/4D5/26 to mice bearing BT474 M1 breast tumors resulted in significant tumor suppression. In addition, tumor tissue excised from GrB/4D5/26–treated mice showed excellent delivery of GrB to tumors and a dramatic induction of apoptosis compared with saline treatment. This study clearly showed that the completely human, functionalized GrB construct can effectively target Her2/neu–expressing cells and displays impressive in vitro and in vivo activity. This construct should be evaluated further for clinical use. Mol Cancer Ther; 12(6); 979–91. Ó2013 AACR. Introduction Bacterial and plant toxin-based immunotoxins have shown remarkable potency and specificity, but a number of obstacles limit their clinical application (1, 2). The toxin component of these fusion proteins can elicit vascular damage leading to loss of vascular integrity (vascular leak syndrome, VLS; refs. 3, 4). Immune responses to the toxins in patients also result in rapid clearance of subsequent courses of therapy (5, 6). Toxin immunogenicity is being addressed by engineering B-cell epitopes on the structure (7, 8), but these molecules may be difficult to humanize completely (9). A new class of immunotoxins have recently been devel- oped containing cytotoxic human proteins (10, 11). Gran- zyme B (GrB) is a well-known serine protease generated by cytotoxic lymphocytes to induce apoptotic cell death in target cells (12, 13). Our group first showed that various fusion constructs targeting tumor cells and tumor endo- thelium and containing GrB have impressive proapopto- tic and cytotoxic activity (14–18). Several other groups have since confirmed these findings using other GrB- containing constructs (19, 20). Because endogenous GrB is present in plasma in both normal and pathologic states, it is unlikely that this molecule would engender an immune response. Dalken and colleagues have described a GrB/FRP5 fusion construct targeting Her2/neu, which displayed selective and rapid tumor cell killing in vitro (21). How- ever, studies have shown that the fusion construct required the presence of the endosome-disrupting agent chloroquine for biologic activity and suggested that an endosomal release process may be necessary for Her2/ neu–targeted agents. Studies by Wang and colleagues suggested that incorporation of a furin-sensitive linker into GrB-based fusion constructs may promote effective Authors' Affiliation: Immunopharmacology and Targeted Therapy Lab- oratory, Department of Experimental Therapeutics, MD Anderson Cancer Center, Houston, Texas Note: Supplementary data for this article are available at Molecular Cancer Therapeutics Online (http://mct.aacrjournals.org/). Current address for Y. Cao: Department of Chemistry, The Scripps Research Institute, La Jolla, CA 92037. Corresponding Author: Michael G. Rosenblum, Department of Experi- mental Therapeutics, MD Anderson Cancer Center, Houston, TX 77054. Phone: 713-792-3554; Fax: 713-794-4261; E-mail: [email protected] doi: 10.1158/1535-7163.MCT-13-0002 Ó2013 American Association for Cancer Research. Molecular Cancer Therapeutics www.aacrjournals.org 979 on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Large Molecule Therapeutics
Construction and Characterization of Novel, CompletelyHuman Serine Protease Therapeutics Targeting Her2/neu
Yu Cao, Khalid A. Mohamedali, John W. Marks, Lawrence H. Cheung, Walter N. Hittelman, andMichael G. Rosenblum
AbstractImmunotoxins containing bacterial or plant toxins have shown promise in cancer-targeted therapy, but
their long-term clinical use may be hampered by vascular leak syndrome and immunogenicity of the
toxin. We incorporated human granzyme B (GrB) as an effector and generated completely human
chimeric fusion proteins containing the humanized anti-Her2/neu single-chain antibody 4D5 (designated
GrB/4D5). Introduction of a pH-sensitive fusogenic peptide (designated GrB/4D5/26) resulted in
comparatively greater specific cytotoxicity although both constructs showed similar affinity to Her2/
neu–positive tumor cells. Compared with GrB/4D5, GrB/4D5/26 showed enhanced and long-lasting
cellular uptake and improved delivery of GrB to the cytosol of target cells. Treatment with nanomolar
concentrations of GrB/4D5/26 resulted in specific cytotoxicity, induction of apoptosis, and efficient
downregulation of PI3K/Akt and Ras/ERK pathways. The endogenous presence of the GrB proteinase
inhibitor 9 did not impact the response of cells to the fusion construct. Surprisingly, tumor cells resistant
to lapatinib or Herceptin, and cells expressing MDR-1 resistant to chemotherapeutic agents showed no
cross-resistance to the GrB-based fusion proteins. Administration (intravenous, tail vein) of GrB/4D5/26
to mice bearing BT474 M1 breast tumors resulted in significant tumor suppression. In addition, tumor
tissue excised from GrB/4D5/26–treated mice showed excellent delivery of GrB to tumors and a dramatic
induction of apoptosis compared with saline treatment. This study clearly showed that the completely
human, functionalized GrB construct can effectively target Her2/neu–expressing cells and displays
impressive in vitro and in vivo activity. This construct should be evaluated further for clinical use.
Mol Cancer Ther; 12(6); 979–91. �2013 AACR.
IntroductionBacterial and plant toxin-based immunotoxins have
shown remarkable potency and specificity, but a numberof obstacles limit their clinical application (1, 2). The toxincomponent of these fusion proteins can elicit vasculardamage leading to loss of vascular integrity (vascular leaksyndrome,VLS; refs. 3, 4). Immune responses to the toxinsin patients also result in rapid clearance of subsequentcourses of therapy (5, 6). Toxin immunogenicity is beingaddressed by engineering B-cell epitopes on the structure
(7, 8), but these molecules may be difficult to humanizecompletely (9).
A new class of immunotoxins have recently been devel-oped containing cytotoxic human proteins (10, 11). Gran-zyme B (GrB) is a well-known serine protease generatedby cytotoxic lymphocytes to induce apoptotic cell death intarget cells (12, 13). Our group first showed that variousfusion constructs targeting tumor cells and tumor endo-thelium and containing GrB have impressive proapopto-tic and cytotoxic activity (14–18). Several other groupshave since confirmed these findings using other GrB-containing constructs (19, 20). Because endogenous GrBis present in plasma in both normal and pathologic states,it is unlikely that this molecule would engender animmune response.
Dalken and colleagues have described a GrB/FRP5fusion construct targeting Her2/neu, which displayedselective and rapid tumor cell killing in vitro (21). How-ever, studies have shown that the fusion constructrequired the presence of the endosome-disrupting agentchloroquine for biologic activity and suggested that anendosomal release process may be necessary for Her2/neu–targeted agents. Studies by Wang and colleaguessuggested that incorporation of a furin-sensitive linkerinto GrB-based fusion constructs may promote effective
Authors' Affiliation: Immunopharmacology and Targeted Therapy Lab-oratory, Department of Experimental Therapeutics, MD Anderson CancerCenter, Houston, Texas
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
Current address for Y. Cao: Department of Chemistry, The ScrippsResearch Institute, La Jolla, CA 92037.
Corresponding Author: Michael G. Rosenblum, Department of Experi-mental Therapeutics, MD Anderson Cancer Center, Houston, TX 77054.Phone: 713-792-3554; Fax: 713-794-4261; E-mail:[email protected]
doi: 10.1158/1535-7163.MCT-13-0002
�2013 American Association for Cancer Research.
MolecularCancer
Therapeutics
www.aacrjournals.org 979
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

cytoplasmic delivery of an active GrB fragment into targetcells (22). However, the recombinant molecule seemed tobe stable only when generated in situ by protein-expres-sing transfected cells.
We previously examined a series of anti-Her2/neusingle-chain antibodies (scFv) fused to the recombinantgelonin (rGel) toxin, and clearly showed that scFvs withintermediate affinity (Kd 10
�11 mol/L) as opposed to highaffinity (Kd 10
�12 mol/L) were optimal carriers of proteintoxins (23, 24). Therefore, we used an intermediate-affin-ity, humanized anti-Her2/neu scFv-designated 4D5 forthe construction of our GrB-containing fusion constructs.In this study, we provided data on the cytotoxicity ofHer2/neu–targeted GrB fusions against a panel of humantumor cell lines and explored the mechanism of in vitroactivity of these fusion constructs. Finally, we showed thein vivo antitumor efficacy of the functionalized GrB chi-meric protein against a human breast xenograft model.
Materials and MethodsCell lines
The cell lines BT474M1,NCI-N87, Calu3,MDAMB435,and Me180 were all obtained from American Type Cul-ture Collection (ATCC). The human breast cancer celllines MDA MB453 and eB-1 were generously suppliedby Drs. Zhen Fan and Dihua Yu (MD Anderson CancerCenter, Houston, TX). The BT474 M1 Herceptin- andlapatinib-resistant cells were derived fromBT474M1 cellsafter a 12-month selection in the continuous presence of 1mmol/L Herceptin or 1.5 mmol/L lapatinib. BT474 M1MDR-1 cells were generated by the transfection of plas-mid pHaMDR1 to parental BT474M1 cells. TheHEK 293Tcell line was supplied by Dr. Bryant G. Darnay (MDAnderson Cancer Center). All cell lines were maintainedin Dulbecco’s Modified Eagle Medium or RPMI-1640medium supplemented with 10% heat-inactivated FBS,2 mmol/L L-glutamine, and 1 mmol/L antibiotics.
Cell lines were validated by short tandem repeat(STR) DNA fingerprinting using the AmpF‘STR Identi-filer Kit according to the manufacturer’s instructions(Applied Biosystems). The STR profiles were comparedwith known ATCC fingerprints (ATCC.org), to theCell Line Integrated Molecular Authentication database(CLIMA) version 0.1.200808 (http://bioinformatics.istge.it/clima/; Nucleic Acids Research 37:D925-D932PMCID: PMC2686526) and to the MD Anderson finger-print database. The STR profiles matched known DNAfingerprints or were unique.
Plasmid constructionThe GrB/4D5/26, GrB/4D5, GrB/26, and GrB DNA
constructs were generated by an overlapping PCR meth-od. Illustrations of the constructs are shown in Fig. 1A.Wedesigned a universal 218 linker (GSTSGSGKPGSGEG-STKG) incorporated between the individual componentsof GrB, 4D5, or peptide 26. Peptide 26 (AALEALAEA-LEALAEALEALAEAAAA) was generated from the 29-residue amphipathic peptide without the 3 C-terminal
amino acids, which are responsible for dimerization (25).All construct genes were cloned into the mammalian cellexpression vector pSecTag (Life Technologies).
Expression, purification, andactivationofGrB-basedproteins
The GrB-based proteins were expressed in HEK293T host cells and purified by immobilized metalaffinity chromatography as described in SupplementaryMethods.
Determination of Kd by ELISAThe Kd value and specificity of GrB-based protein
samples were evaluated by ELISA on Her2/neu extracel-lular domain (ECD), Her2/neu-positive BT474 M1 cells,and Her2/neu–negative Me180 cells. Rabbit anti-c-mycantibody and horseradish peroxidase–conjugated goatanti-rabbit immunoglobulin G were used as tracers inthis assay, as described previously (24).
GrB activity assaysThe enzymatic activity of the GrB component was
determined in a continuous colorimetric assay usingN-a-t-butoxycarbonyl-L-alanyl-L-alanyl-L-aspartyl-thio-benzylester (BAADT) as a specific substrate (18). Assaysconsisted of commercial human GrB (Enzyme Systems
GrBGrB/4D5/26
26
A.A. M.W. P.I.
GrB/4D5 502 53932.4 9.56
GrB/4D5/26 546 57900.6 9.23
GrBGrB/4D5
Nonreduced Reduced
150
kDa
100
75
50
35
25
A
B
Figure 1. Construction and preparation of GrB-based fusion constructs.A, schematic diagram of GrB fusion constructs containing scFv 4D5 andGrBwithout or with fusogenic peptide 26. B, purifiedGrB-based chimericproteins were analyzed by SDS–PAGE under nonreducing conditions.
Cao et al.
Mol Cancer Ther; 12(6) June 2013 Molecular Cancer Therapeutics980
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

Products) or GrB-based fusion proteins in BAADT at25�C. The change in absorbance at 405 nm was measuredon a Thermomax plate reader. Increases in sample absor-bance were converted to enzymatic rates by using anextinction coefficient of 13,100 cm�1 (mol/L)�1 at 405 nm.The specific activity of GrB-based fusion proteins wascalculated using native GrB as the standard.
Internalization analysisImmunofluorescence-based internalization studies
were conducted using BT474 M1 and Me180 cells. Cellswere treated with 25 nmol/L GrB/4D5/26 for 4 hoursand subjected to immunofluorescent staining with anti-GrB antibody [fluorescein isothiocyanate (FITC)-conju-gated secondary antibody]. Nuclei were counterstainedwith propidium iodide (PI). Visualization of immuno-fluorescence was conducted with a Zeiss LSM510 con-focal laser scanning microscope Zeiss LSM510 (CarlZeiss).
In vitro cytotoxicity assaysLog-phase cells were seeded (�5� 103/well) in 96-well
plates and allowed to attach overnight. Cells were furtherincubated with various concentrations of GrB-basedfusion proteins, GrB, or medium at 37�C for 72 hours.Cell viability was determined using the crystal violetstaining method as described previously (23).
Annexin V/PI stainingThe Annexin V/PI staining assay was used to quanti-
tatively determine the percentage of cells undergoingapoptosis after exposure toGrB/4D5/26. Cellswere seed-ed onto 6-well plates (5 � 105 cells/ well) and incubatedwith 100 nmol/L GrB/4D5/26 at 37�C for 24 or 48 hours.Aliquots of cells were washed with PBS and then incu-bated with Annexin V–FITC antibody. PI solution wasadded at the end of the incubation, and the cells wereanalyzed immediately by flow cytometry.
Cytochrome c release assay and Bax translocationAfter treatment with GrB/4D5 or GrB/4D5/26, cells
were collected and resuspended with 0.5 mL of 1�cytosol extraction buffer mix (BioVision) and thenhomogenized in an ice-cold glass homogenizer. Thehomogenate was centrifuged, and the supernatant wascollected and labeled as the cytosolic fraction. Thepellet was resuspended in 0.1 mL of mitochondrialextraction buffer and saved as the mitochondrialfraction. Aliquots of each cytosolic and mitochondrialfraction were analyzed by Western blotting with anti-bodies recognizing cytochrome c and Bax (Santa CruzBiotechnology).
Assays for caspase activation and apoptosisWestern blot analysis was used to identify activation of
caspases-3 and -9 as well as PARP cleavage. In addition,apoptosis was analyzed using antibodies recognizing Bcl-2 and BID (Santa Cruz Biotechnology).
Impact on cell signaling pathwaysAfter treatment, cell lysates were analyzed by Western
blotting with antibodies recognizingHer2/neu and phos-phorylated (p)-mTOR (S2448; Cell Signaling Technology)as well as p-Her2/neu (Tyr877), p-Her2/neu (Tyr 1221/1222), EGF receptor, p-EGF receptor (Thr845), Her3, p-Her3 (Tyr1328), IGF1 receptor, p-IGF1 receptor (Tyr 1165/1166), estrogen receptor (ER), progesterone receptor (PR),Akt, p-Akt, extracellular signal–regulated kinase (ERK),p-ERK (Thr 177/Thr 160), PTEN, proteinase inhibitor 9(PI-9), and b-actin (all from Santa Cruz Biotechonology).Immunoreactive proteins were visualized by enhancedchemiluminescence.
In vivo efficacy studiesWe used Balb/c nude mice to evaluate the in vivo effect
of GrB/4D5/26 against aggressive breast cancer. Eachmouse received a weekly subcutaneous injection of 3mg/kg estradiol cypoinate (26) starting 2 weeks beforethe injection of 1� 107 BT474M1 cells into the right flank.On the third day after cell inoculation, mice were injectedintravenously (tail vein) eitherwith saline orGrB/4D5/26(44 mg/kg) 5 times per week for 2 weeks. Animals weremonitored and tumors were measured (calipers) for anadditional 50 days.
Immunofluorescence analysisTwenty-four hours after the final injection of saline or
GrB/4D5/26, mice were sacrificed and tumor sampleswere frozen immediately in preparation for sectionslides. The sample slides were incubated with eitheranti-GrB antibody (FITC-conjugated secondary anti-body) or a terminal deoxynucleotidyl transferase–medi-ated dUTP nick end labeling (TUNEL) reaction mixture,as well as with an anti-mouse CD31 antibody (phyco-erythrin-conjugated secondary antibody), and were fur-ther subjected to nuclear counterstaining with Hoechst33342. Immunofluorescence observation was conductedunder a Zeiss Axioplan 2 imaging microscope (CarlZeiss).
ResultsConstruction, expression, and purification of GrB-based fusions
The sequence of the humanized anti-Her2/neu scFv4D5 was derived from the published Herceptin light-and heavy-chain variable domain sequences (27). Pre-vious observations suggested that use of fusogenicpeptides facilitates endosomal escape and delivery oflarge molecules into the cytotol (28, 29). We thereforeincorporated the fusogenic peptide 26 (Fig. 1A). GrB-based fusions were generated by fusing GrB to 4D5with (designated GrB/4D5/26) or without (designatedGrB/4D5) the addition of pH-sensitive fusogenic pep-tide 26 (AALEALAEALEALAEALEALAEAAAA) tothe C-terminal of the construct. Furthermore, GrB andGrB/26 were used as controls. All fusion proteins wereexpressed in human embryonic kidney cells (HEK
Novel Functionalized GrB-Targeting Her2/neu
www.aacrjournals.org Mol Cancer Ther; 12(6) June 2013 981
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

293T). Following purification, the final products migrat-ed at the expected molecular weights, with a purity ofmore than 95% (Fig. 1B).
Analysis of binding affinityThe binding affinities (Kd values) of GrB/4D5/26 and
GrB/4D5 were assessed by ELISA using purified Her2/neu ECD, Her2/neu–positive BT474 M1 human breastcancer cells, and Her2/neu–negative Me180 human cer-vical cancer cells. Both fusions specifically bound toHer2/neu ECD and BT474 M1 cells but not to Me180 cells (Fig.2A). The apparent Kd values were determined by calcu-lating the concentration of fusion constructs that pro-duced half-maximal specific binding. GrB/4D5 andGrB/4D5/26 showed apparent Kd values of 0.329 and0.469 nmol/L, respectively, to Her2/neu ECD and 0.383and 0.655 nmol/L, respectively, to BT474 M1 cells. Theseresults are in general agreement with the published Kd
value for native Herceptin to the Her2/neu receptor (0.15nmol/L; ref. 27).
Enzymatic assay of GrB-based fusionsTo assess the biologic activity of the GrB component of
the fusions, we compared the ability of the constructs tocleave the substrate BAADTwith that of native, authenticGrB (Fig. 2B). GrB/4D5 and GrB/4D5/26 had intact GrBenzymatic activity (1.54 � 105 and 1.57 � 105 U/mmoL,respectively). These activities were comparable with thatof the native GrB standard (1.19� 105 U/mmoL). Becausethe pro-GrB fusion constructs contain purification tags onthe N-terminal end of GrB and render the moleculeenzymatically inactive, these proteins were unable tocause hydrolysis of BAADT.
Cellular uptake andGrBdelivery of fusion constructsImmunofluorescence staining was conducted with
BT474M1andMe180 cells. TheGrBmoiety of both fusionswas observed primarily in the cytosol after treatmentwitha fusion protein in BT474 M1 cells but not in Me180 cells(Fig. 2C), showing that both constructs were efficient incell binding and internalization after exposure to Her2/neu–positive cells. The internalization efficiency of the
ELISA of GrB-based fusions on Her2/neu ECD
1.8
1.5
1.2
0.9
0.6
0.3
0.0
GrB/4D5GrB/4D5/26Control
10–3 10–2 10–1 100
Conc. (nmol/L)101 102
Ab
so
rban
ce a
t 405 n
m
1.8
1.5
1.2
0.9
0.6
0.3
0.0
GrB/4D5GrB/4D5/26Control
Pro-GrB/4D5/26
Pro-GrB/4D5
GrB standard
GrB/4D5/26
GrB/4D5
GrB/4D5
GrB/4D5
0 10
0 0.5 1 2 4 8 12 24 36 48 h 0 0.5 1 2 4 8 12 24 36 48 h
0.20
B
A
C
D
0.15
0.10
0.05
0.0020 30 40 50 60 70
Time (min)
GrB/4D5/26
GrB/4D5/26
PBS
BT474 M1
Full-length GrB fusion
Free GrB
β-Actin
Me 180
10–3 10–2 10–1 100
Conc. (nmol/L)101 102
Ab
so
rban
ce a
t 405 n
m
Ab
so
rba
nc
e a
t 4
05
nm
0.8
0.6
0.4
0.2
0.0
GrB/4D5GrB/4D5/26
10–3 10–2 10–1 100
Conc. (nmol/L)101 102
Ab
so
rban
ce a
t 405 n
m
ELISA of GrB-based fusions on Me180 cellsELISA of GrB-based fusions on Her2/neu ECD
Figure 2. Characterization and comparison of GrB-based fusion proteins. A, Kd of the fusion constructs to Her2/neu ECD, Her2/neu–positive BT474M1 cells,and Her2/neu–negative Me180 cells by ELISA. B, enzymatic activity of GrB moiety of fusion proteins compared with native GrB. C, internalizationanalysis of BT474M1 cells andMe180 cells after 4 hours of treatment with 25 nmol/L functionalized GrB fusions. Cells were subjected to immunofluorescentstaining with anti-GrB antibody (FITC-conjugated secondary), with PI nuclear counterstaining. D, Western blot analysis of intracellular behavior of25 nmol/L GrB fusion constructs in BT474 M1 cells.
Cao et al.
Mol Cancer Ther; 12(6) June 2013 Molecular Cancer Therapeutics982
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

fusions was further examined by time-dependent West-ern blot analysis of the GrB signal (full-length GrB fusionþ free GrB; Fig. 2D). Both constructs internalized rapidlyinto BT474 M1 cells within 30 minutes. Compared withGrB/4D5, GrB/4D5/26 displayed enhanced and long-lasting cell internalization. The intracellular delivery ofGrB after endocytosis of GrB/4D5 or GrB/4D5/26 alsowas assessed by time-dependent Western blotting (freeGrB). We observed no GrB delivery by GrB/4D5 up to 48hours of treatment,whereasGrBdelivery byGrB/4D5/26was observed starting at approximately 4 hours of treat-ment and presented a tremendously high level of free GrBup to 48 hours (Fig. 2D).
In vitro cytotoxic effects of GrB-based fusionsGrB-based fusionswere then tested against a number of
tumor cell lines. After 72 hours exposure, GrB/4D5/26showed specific cytotoxicity to Her2/neu–positive cells,with IC50 values of less than 100 nmol/L (Table 1), andGrB/4D5 showed cytotoxic effects at somewhat higherdoses (>200 nmol/L). In addition, GrB/26 showed min-imal cytotoxicity at doses more than 600 nmol/L, but nosignificant activity of GrB itself was observed at doses upto 1.5 mmol/L. When Her2/neu–positive MDA MB453cells were pretreated with Herceptin (5 mmol/L) for 6hours and then treatedwithGrB/4D5/26 for 72 hours, thecytotoxicity ofGrB/4D5/26was reduced (Supplementary
Fig. S1), thereby showing a requirement for antigen bind-ing of the GrB/4D5/26 construct.
We further investigated the expression levels of theendogenous PI-9 in different tumor cells (SupplementaryFig. S2 and Table 1). These studies failed to find anassociation between the response of cells to the cytotox-icity of theGrB constructs and the endogenous expressionof PI-9. This may suggest that factors other than PI-9 mayaccount for the observed differences inGrB/4D5/26 cyto-toxicity to Her2/neu–expressing target cells.
Cytotoxic effects of GrB/4D5/26 against cellsresistant to Herceptin or lapatinib
Acquired resistance to Herceptin or lapatinib can bemediated by concomitant upregulation of Her2/neudownstreamsignalingpathwaysor activationof signalingthrough the ER pathway (30). In this study, we developeda model of Herceptin- and lapatinib-resistant variants ofBT474 M1 cells. Parental BT474 M1 cells were readilysensitive to both Herceptin (IC50, 52.5 nmol/L) and lapa-tinib (IC50, 34.7 nmol/L; Table 2). Herceptin-resistant cellsshowed resistance to Herceptin [IC50, 10.1 mmol/L; foldresistance (FR), 192] but remained sensitive to lapatinib(IC50, 32.4 nmol/L). Lapatinib-resistant cells showedresistance to high micromolar concentrations of bothHerceptin (IC50, 74.1 mmol/L; FR, 1411) and lapatinib(IC50, 8.2 mmol/L; FR, 237). As shown in Table 2, cells
Table 1. Comparative IC50 values of GrB-based fusion constructs against various types of tumor cell lines
IC50 (nmol/L)
Cell line TypeHer2/neulevel
PI-9level GrB/4D5/26 GrB/4D5 GrB/26 GrB
BT474 M1 Breast þþþþ þ 29.3 253.3 905.5 >1,500.0Calu3 Lung þþþþ þþþþþ 40.5 242.4 863.0 >1,500.0NCI-N87 Gastric þþþþ þ 90.4 629.0 1,106.0 >1,500.0MDA MB453 Breast þþþ þ 56.8 436.0 694.2 >1,500.0eB-1 Breast þþ — 93.1 551.3 1,134.5 >1,500.0MDA MB435 Breast þ — >500.0 >750.0 1,031 >1,500.0Me180 Cervical þ þ >500.0 >750.0 >1,500.0 >1,500.0
NOTE: þ, indicates the Her2/neu expression level in different cancer cells.
Table 2. Cytotoxic effects of Her2/neu–targeted therapeutic agents on IC50 values in BT474 M1 cells andresistant variants
IC50 (nmol/L) with (FR)a
Agent BT474 M1
BT474 M1Herceptin-resistant
BT474 M1lapatinib-resistant
BT474 M1 þEGFb
BT474 M1 þNRG-1c
BT474 M1 þb-estradiold
Herceptin 52.5 (1) 10,100.5 (192) 74,100.0 (1,411) 26,305.0 (501) 23,033.0 (439) 74.1 (1)Lapatinib 34.7 (1) 32.4 (1) 8,225.0 (237) 543.0 (16) 547.1 (16) 33.9 (1)GrB/4D5/26 32.9 (1) 26.8 (1) 66.1 (2) 21.7 (1) 18.1 (1) 31.3 (1)
aFR represents IC50 of agent on BT474 M1–resistant variants/that on BT474 M1 parental cells.b,c,dCells were pretreated with b20 ng/mL EGF, c50 ng/mL NRG-1, or d10 ng/mL b-estradiol for 72 hours before drug treatment.
Novel Functionalized GrB-Targeting Her2/neu
www.aacrjournals.org Mol Cancer Ther; 12(6) June 2013 983
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

resistant toHerceptin showed equivalent sensitivity to theGrB/4D5/26 construct (IC50, �30 nmol/L for both Her-ceptin-resistant and parental BT474 M1 cells, respective-ly). For lapatinib-resistant cells, the IC50 was marginallyincreased (2-fold) compared with parental cells (66.1 vs.32.9 nmol/L).
We also showed that addition of EGF or neuregulin-1(NRG-1) growth factor, but not b-estradiol, to BT474M1 parental cells can circumvent the cellular cytotoxicresponses to Herceptin and lapatinib. Seventy-two hoursof pretreatment of BT474 M1 cells with 20 ng/mL EGF or50 ng/mLNRG-1 resulted in a 400- to 500-fold increase inresistance toHerceptin and a 16-fold increase in resistanceto lapatinib (Table 2). However, treatment of these resis-tant cells resulted in no cross-resistance to GrB/4D5/26fusions compared with parental BT474 M1 cells.
A significant observation was that incubation of cellswith GrB/4D5/26 in the presence of chloroquine didnot improve cytotoxicity toward these cells (Supple-mentary Fig. S3). This finding showed that the fuso-
genic peptide 26 efficiently releases GrB fusion pro-teins from intracellular vesicles, thereby allowingaccess to cytosolic GrB substrates and induction ofapoptosis.
Mechanistic studies of GrB/4D5/26 cytotoxicityWe conducted a panel of experiments to assess the
potential of GrB-based fusions to initiate the proteolyticcascade culminating in apoptosis of BT474 M1 parental,Herceptin-, and lapatinib-resistant cells.
Annexin V/PI staining. Cells were incubated for 24 or48 hours with 100 nmol/L GrB/4D5/26 and apoptosiswas detected via Annexin V/PI staining. GrB/4D5/26induced apoptosis in BT474 M1 parental, Herceptin- andlapatinib-resistant cells, as indicatedby the reducedviablepopulation combined with greater populations of earlyapoptosis (Fig. 3A). No apoptosis was induced by 100nmol/L GrB/4D5 in any of these cells (SupplementaryFig. S4).Her2/neu–negativeMe180 cellswere not affectedby either construct.
Figure 3. Effects of GrB-based fusions on apoptotic pathways of BT474 M1 parental, Herceptin-, and lapatinib-resistant cells. A, detection of apoptosis ofGrB/4D5/26 by Annexin V/PI staining assay. Me180 cells served as a Her2/neu–negative control group. B, Western blot analysis of cleavage andactivationof caspases-3 and -9 aswell asPARPbyGrB-based fusion constructs. C,Western blot analysis of apoptosis kinetics andspecificity ofGrB/4D5/26.Cells were treated with GrB/4D5/26 for up to 24 hours with or without 100 mmol/L zVAD-fmk for 24 hours in parental or Herceptin-resistant cells and for up to48 hours in lapatinib-resistant cells.
Cao et al.
Mol Cancer Ther; 12(6) June 2013 Molecular Cancer Therapeutics984
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002
Activation of caspases. Caspase activation in BT474M1 cellswas detected byWestern blot analysis. Treatmentwith GrB/4D5/26 resulted to the cleavage of caspase-3,�9, andPARP in all cells, but no activation occurredwhencellswere treatedwithGrB/4D5 (Fig. 3B). ComparedwithBT474 M1 parental and Herceptin-resistant cells, the acti-vations of caspase-9, -3, and PARP were delayed in lapa-tinib-resistant cells, which coincided with the observeddecreased cytotoxic effects.We further assessed the kinetics of PARP cleavage
induced by GrB/4D5/26 on BT474 M1 parental, Her-ceptin- and lapatinib-resistant cells, and found thatcleavage occurred after 2 hours of drug exposure forparental and Herceptin-resistant cells but at 24 hours forlapatinib-resistant cells (Fig. 3C). In addition, in thepresence of the pan-caspase inhibitor zVAD-fmk, PARP
cleavage of GrB/4D5/26 was partially inhibited in allcells. This finding is in agreement with a mechanismrelying on GrB activity for caspase-3 cleavage followedby PARP cleavage.
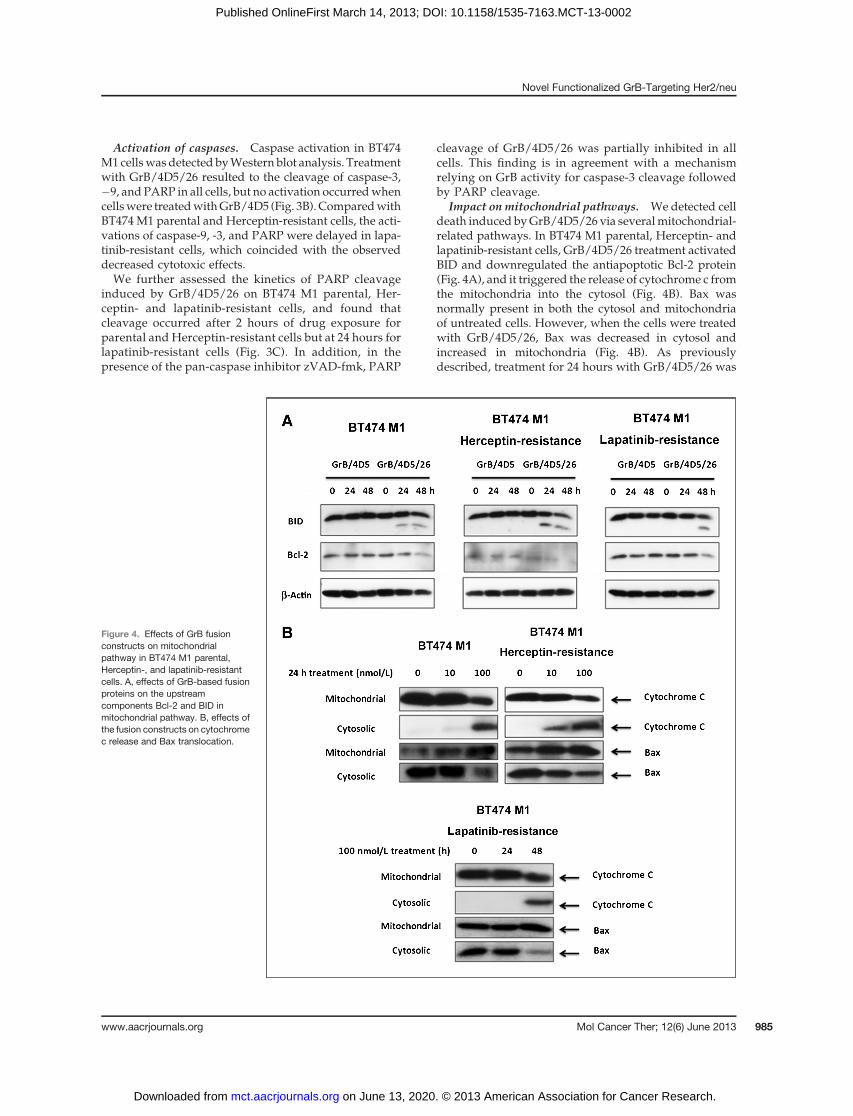
Impact on mitochondrial pathways. We detected celldeath induced byGrB/4D5/26 via severalmitochondrial-related pathways. In BT474 M1 parental, Herceptin- andlapatinib-resistant cells, GrB/4D5/26 treatment activatedBID and downregulated the antiapoptotic Bcl-2 protein(Fig. 4A), and it triggered the release of cytochrome c fromthe mitochondria into the cytosol (Fig. 4B). Bax wasnormally present in both the cytosol and mitochondriaof untreated cells. However, when the cells were treatedwith GrB/4D5/26, Bax was decreased in cytosol andincreased in mitochondria (Fig. 4B). As previouslydescribed, treatment for 24 hours with GrB/4D5/26 was
Figure 4. Effects of GrB fusionconstructs on mitochondrialpathway in BT474 M1 parental,Herceptin-, and lapatinib-resistantcells. A, effects of GrB-based fusionproteins on the upstreamcomponents Bcl-2 and BID inmitochondrial pathway. B, effects ofthe fusion constructs on cytochromec release and Bax translocation.
Novel Functionalized GrB-Targeting Her2/neu
www.aacrjournals.org Mol Cancer Ther; 12(6) June 2013 985
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

shown to activate the mitochondrial pathway in bothBT474 M1 parental and Herceptin-resistant cells, but thisactivation was delayed in lapatinib-resistant cells.
Effects of GrB fusions on Her- and ER-associatedsignaling pathways
We next examined the mechanistic effects of the con-structs on Her- and ER-related signaling events in BT474M1 parental cells and the resistant variants. As shown inSupplementary Fig. S5, cells resistant to Herceptin hadenhanced Her family receptor activity but reduced levelsof PRandPI-9. In contrast, in lapatinib-resistant cells therewas total downregulation of Her family receptor activitybut higher levels of ER, PR, and PI-9.
Cells treatedwithGrB/4D5orGrB/4D5/26 showed theeffects on these signaling pathways, corresponding to thecytotoxic results we observed (Fig. 5). Treatment withGrB/4D5/26 remarkedly inhibited phosphorylation ofHer2/neu and its downstream molecules Akt, mTOR,and ERK, which are critical events in Her2/neu signalingcascade. In contrast, GrB/4D5 showed a comparativelyreduced effect on these pathways.We observed a reducedER level among all cells. Evidence from other researchers
has shown that upregulation of the ER pathway in ER-and Her2/neu–positive cell lines with lapatinib createsan escape/survival pathway (30, 31), but GrB/4D5/26seem to be able to inactivate all the signaling pathwaysin these cells. We also observed the delaying signalingeffects of GrB/4D5/26 on lapatinib-resistant cells com-pared with parental or Herceptin-resistant cells, whichwas in agreement with the apoptotic cell death resultsobserved for the lapatinib-resistant cells. Notably, therewas an increased mRNA and protein level of PI-9 in thisresistant line but not in the parental or Herceptin-resis-tant cells (Supplementary Figs. S5 and S6). Takentogether, these results suggest that activation of the ERpathway upregulates the expression of PI-9 and thisresults in a slight inhibition of GrB/4D5/26 activity anda delay in apoptotic cell death compared with parentalcells.
Our investigation suggests that the GrB/4D5/26 fusionis more cytotoxic than GrB/4D5 construct to Her2/neu–positive cells, even those that have acquired resis-tance to the traditional Her2/neu therapeutic agents Her-ceptin and lapatinib. The cytotoxicity results coincidewith the observed effects on signal transduction and
Figure 5. Western blot analyses ofthe effects of GrB/4D5 and GrB/4D5/26 in BT474 M1 parental,Herceptin-, and lapatinib-resistantcells on Her2/neu and ER signalingpathways. Cells were treated with100 nmol/L functionalized GrBconstructs for 24 or 48 hours, andtotal cell lysates were quantifiedand further measured by Westernblot analysis of pHer2/neu, pAkt,pmTOR, pERK, ER, PR, and PI-9level.
Cao et al.
Mol Cancer Ther; 12(6) June 2013 Molecular Cancer Therapeutics986
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

monitoring these pathways may be useful as a monitor ofdrug efficacy.
Effects of GrB/4D5/26 on the MDR-1–expressingcellsMultidrug resistance (MDR) is a phenomenon, which
results from various reasons. The most-characterizedcause of MDR is the overexpression of a 170-kDa mem-brane glycoprotein known as P-glycoprotein (Pgp). Toverify the effects of GrB-based fusions on the Her2/neu–positive cells with MDR-1 expression, we generat-ed the BT474 M1 MDR-1 cells by the transfection ofplasmid pHaMDR1 to parental BT474 M1 cells. Asshown in Table 3, compared with parental cells, BT474M1 MDR-1 showed 209-fold resistance to Taxol and 89-fold resistance to vinblastin. However, we could notobserve the cross-resistance of MDR-1 cells to GrB/4D5and GrB/4D5/26 constructs. Therefore, GrB-basedfusion constructs show a wide range cytotoxicity totarget cells even those with acquired resistance to che-motherapeutic agents.
Antitumor activity of GrB/4D5/26 fusions inxenograft modelsWe evaluated the ability of the GrB/4D5/26 fusion
construct to inhibit the growth of established BT474 M1tumor xenografts in nude mice after systemic adminis-tration. Tumors were subcutaneously inoculated withBT474 M1 cells on day 0, and treatment was initiated onday 3. Treatment consisted of an intravenous injection fora total of 10 days with saline or 44 mg/kg GrB/4D5/26.Compared with saline, GrB/4D5/26 greatly slowedtumor progression over 50 days of observation (Fig.6A). There were no obvious toxic effects of GrB/4D5/26 on mice at this dose, suggesting that the maximum-tolerated dose at this schedule had not been reached.Finally, we determined the localization of GrB/4D5/26
after administration to mice bearing BT474 M1 tumors.Immunofluorescence staining confirmed that GrB/4D5/26 localized quickly and specifically in tumor tissue (Fig.6B). This observation further suggested that GrB/4D5/26can effectively target tumor cells overexpressing Her2/
neu in vivo and can show significant tumor growth-sup-pressive effects in the absence of observable toxicity.Staining of tumor tissue nuclei with TUNEL (Fig. 6C)clearly showed that the tumor tissues displayed apoptoticnuclei in the GrB/4D5/26 treatment group. In addition,the intratumoral distribution of GrB/4D5/26 seemed toconcentrate primarily in areas with extensive apoptoticresponse (compare Grb/4D5/26 distribution in Fig. 6B,with TUNEL staining in Fig. 6C).
DiscussionAntibody-based therapeutic agents are one of the fast-
est growing areas in the cancer therapeutic field. Two ofthe most promising strategies to enhance the antitumoractivity of antibodies are antibody–drug conjugates(ADC) and immunotoxins. There are now a number ofclinically effectiveADCs showing remarkable activity (32,33) and many of these constructs were driven by theimpressive success of the trastuzumab–DM1 (T–DM1)conjugate. On the other hand, there are limitations withADCs, such as facile aggregation, off-target toxicity, andpotential resistance from MDR-positive cells (34, 35).
The use of immunotoxins has always been a promisingalternative strategy for cancer-targeted therapy, but thepotential for antigenicity precluding prolonged treatmentcycles (36, 37) and the unresolved issue of VLS havelimited interest in the overall development of this area.Currently, a number of groups have focused on thedevelopment of either deimmunized or fully human toxintype molecules as payloads (1, 10, 38, 39).
In this study, we constructed novel human anti-Her2/neu immunotoxins containing human GrB as anapoptosis-inducing effector. GrB seems to be an idealpayload for targeted therapeutic applications in partbecause this serine protease exerts a multimodal andwell-known mechanism of cytotoxic action (40, 41). Ofinterest, this study found that inhibitors of caspaseactivation had little impact on the overall cytotoxicityof the construct, attesting to the presence of multiple,redundant, proapoptotic pathways activated by thismolecule and suggesting that emergence of resistanceto this class of agents may be difficult from a biologicperspective.
In a nominal cytotoxic process, GrB penetrates directlyinto target cells through the action of perforin-mediatedtransmembrane pores. This process bypasses the lyso-somal compartment allowingGrB accessibility directly tocytosolic substrates (42). Internalization of GrB throughantibody-mediated events provides tumor cell specific-ity but in the case of Her2/neu, internalization likelyproceeds through the lysosomal compartment. For ouroptimal construct, we included a 26-residue, fusogenicpeptide. At neutral pH, this peptide has a randomconfiguration, but under acidic lysosomal conditions,this peptide assumes an amphipathic helix, thereby dis-rupting the lysosomal membrane allowing improveddelivery of the fusion construct into the cytosol (25).Studies on target cells show that the incorporation of
Table 3. Cytotoxicity of chemical agents andGrB-based fusions on MDR-1–expressing cells
IC50 (nmol/L)
BT474M1
BT474 M1MDR-1 FRa
Taxol 5.2 1,047.3 209Vinblastin 1.3 105.1 89GrB/4D5 311.8 318.9 1GrB/4D5/26 34.1 35.5 1
aFR represents IC50 of agent on BT474M1MDR-1 cells/thaton BT474 M1 parental cells.
Novel Functionalized GrB-Targeting Her2/neu
www.aacrjournals.org Mol Cancer Ther; 12(6) June 2013 987
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

the fusogenic peptide dramatically augments intracellu-lar delivery and biologic activity of the GrB-containingconstruct.
Previous studies determined that bacterially expressedGrB fusion proteins targeting cell surface antigens otherthan Her2/neu have excellent cytotoxic activity (17, 43).The folding of the protein and the absence of carbohy-drates on the final product could impact their biologicactivity compared with proteins produced inmammaliancells. We expressed the GrB/4D5 fusion in bacterial cellsand the purified protein showed similar cytotoxicity tothat produced in mammalian cells (data not shown). Thissuggests that intracellular routing of chimeric GrB mole-cules depends on the type of target receptor and was notaffected by the presence of carbohydrates on the chimericmolecule.
Dalken and colleagues described the construction andbiologic activity of Her2/neu–targeted fusion constructGrB/FRP5 (21). This agent was shown to be specificallycytotoxic to target cells with IC50 values in the subna-nomolar range but the cytotoxic activity was dependenton the addition of the lysomotropic agent chloroquine.In the absence of chloroquine, the cytotoxicity of theagent was reduced 10- to 300-fold, thus suggesting thatthe construct may have been primarily sequestered intothe lysozomal compartment and not available to acti-vate apoptotic cascade mechanisms. The incorporationof the fusogenic, pH-sensitive peptide 26 in our con-struct seemed to circumvent the need for a lysomotropicagent to augment the activity of GrB fusion and itprovided a greater concentration of target protein inthe cell. The use of this peptide did not seem to impact
1,500
1,250
1,000
750
500
250
0
Saline
GrB/4D5/26
Saline
GrB/4D5/26
Saline
GrB/4D5/26
0 10 20 30 40 50
Time (d)
OverlayGrBCD31Hoechst 33342B
A
C OverlayTUNELCD31Hoechst 33342
Tu
mo
r vo
lum
e (
mm
3)
Figure 6. Tumor apoptotic activityof GrB/4D5/26 in BT474 M1 tumorxenografts. A,micewith BT474M1flank tumors were intravenouslyinjected with saline or 44 mg/kgGrB/4D5/26 at the indicated times(arrows). Mean tumor volume wascalculated as W � L � H asmeasured with digital calipers. B,immunofluorescence staining oftumor samples after intravenousinjection of saline andGrB/4D5/26.Twenty-four hours after injection,animal was sacrificed and frozentumor sections were prepared anddetected by anti-GrB antibody(green) and anti-mouse CD31antibody (red). Hoechst 33342(blue) was used for DNA staining.C, apoptosis detection in tumortissue by TUNEL assay.
Cao et al.
Mol Cancer Ther; 12(6) June 2013 Molecular Cancer Therapeutics988
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

the enzymatic activity of the GrB component nor did itinfluence the binding activity of the 4D5 to Her2/neureceptor. Finally, the presence of the 26 component didnot seem to augment the nonspecific toxicity of theconstruct against antigen-negative cells in vitro nor didit increase the apparent toxicity of the construct duringintravenous administration in our xenograft studies.The antitumor efficacy studies showed that admin-
istration of GrB/4D5/26 to mice bearing well-devel-oped BT474 M1 tumors was effective at total doses of 44mg/kg. This dose translates to a total dose of approx-imately 140 mg/m2. In comparison, studies of the ADCT–DM1 conjugate in the BT474 tumor model used dosesof 3 to 15 mg/kg (�10–50 mg/m2; ref. 44), which islower than the doses for the GrB construct. However,our study showed that there were no deaths or weightloss during the treatment schedule, suggesting thesafety and tolerability of GrB-based agents. Althoughwe did not observe complete regression of tumor xeno-grafts, alternative schedules or higher doses need to beexamined.The Her2/neu–targeted therapeutic agents Herceptin
and lapatinib have significantly improved outcomes incancer treatment, but their use is limited by resistanceand tolerability issues (45, 46). Evaluating the cytotoxic-ity of functionalized GrB fusions to Herceptin- or lapa-tinib-resistant cells represents an important step. Ourresults suggested that GrB/4D5/26 inhibits the prolifer-ation and survival of resistant cells as a result of caspase-dependent and -independent apoptotic effects. In addi-tion, our investigation into cellular signaling indicatedthat GrB/4D5/26 could efficiently downregulate thephosphorylation of Her2/neu and ER family members,resulting in inhibition of both PI3K/Akt and Ras/ERKpathways.The development of MDR mechanisms affecting
groups of therapeutic agents has been shown to be acentral problem resulting in reduced response in cancertreatment (47). The emergence of MDR phenotypes couldalso be a serious problem for the application of ADCs(48, 49). Studies by Kovtun and colleagues (50) reportedthat ADCs using polyethylene glycol (PEG)-basedhydrophilic linkers showed higher retention in MDR-1–expressing cells than similar conjugates made with thenonpolar linker succinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate (SMCC), which is found inT–DM1. Therefore, the emergence of MDR may providecross-resistance to T–DM1, due to the efflux of free drugupon intracellular release from the antibody. In con-trast, the current studies show that expression of MDRdoes not provide cross-resistance to GrB-based fusionconstructs and this seems to be a significant advantageover the conventional ADC approach.The only intracellular inhibitor of human GrB is the
nucleocytoplasmic serpin, PI-9. PI-9 has been found tobe endogenously expressed in lymphocytes, dendriticcells, and mast cells for self-protection against GrB-mediated apoptosis (40, 41). This may suggest that the
endogenous PI-9 level in cancer cells could inhibit theGrB activity of our target molecules. However, ourstudies did not show any relationship between PI-9levels and cell sensitivity to GrB/4D5/26 in Her2/neu–positive cells.
WeexaminedGrB sensitivity against lapatinib-resistantcells and found these cells showed a slight (2-fold)increase in the GrB/4D5/26 IC50. This coincided with anupregulation of PI-9 leading to a delay in apoptosis. Thisupregulation may be the indirect result of ER pathwaychanges induced by lapatinib resistance. Therefore, in thecell lines that are both ER- and Her2–positive, for whichupregulation of the ER pathway may occur as an escapepathway, the endogenous GrB inhibitor PI-9 could beupregulated to inhibit GrB activity.
In conclusion, we showed that a novel Her2/neu–targeted functionalized GrB fusion constructs using thepH-sensitive fusogenic peptide 26 as an endosomolyticdomain efficiently promotes the release of GrB into thecytoplasm, resulting in apoptotic cell death in Her2/neu–positive cancer cells. This fusogenic peptide couldbe useful for studying GrB-induced apoptosis withoutthe requirement of perforin or chloroquine. In addition,our studies show that tumor cells highly resistant toeither lapatinib or Herceptin and the cells with MDR-1expression resistant to chemotherapeutic agents werenot cross-resistant to the GrB-based fusion protein.Although the induction of PI-9 expression in lapati-nib-resistant cells delayed the apoptotic cytotoxicity ofGrB/4D5/26, this agent had an IC50 value that was only2-fold higher than parental cells, despite the fact thatresistant cells were more than 200-fold resistant tolapatinib.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: Y. Cao, K.A. Mohamedali, L.H. Cheung, M.G.RosenblumDevelopment of methodology: Y. Cao, K.A. Mohamedali, L.H. CheungAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): Y. Cao, J.W. Marks, L.H. Cheung, W.N.HittelmanAnalysis and interpretation of data (e.g., statistical analysis, biostatis-tics, computational analysis): Y. Cao, K.A. Mohamedali, L.H. Cheung,W.N. HittelmanWriting, review, and/or revision of the manuscript: Y. Cao, K.A. Moha-medali, L.H. Cheung, W.N. HittelmanAdministrative, technical, or material support (i.e., reporting or organiz-ing data, constructing databases): Y. Cao, L.H. Cheung, M.G. RosenblumStudy supervision: L.H. Cheung, M.G. Rosenblum
Grant SupportThis research was supported, in part, by the Clayton Foundation for
Research (M.G. Rosenblum).The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received January 3, 2013; revisedMarch 6, 2013; acceptedMarch 8, 2013;published OnlineFirst March 14, 2013.
Novel Functionalized GrB-Targeting Her2/neu
www.aacrjournals.org Mol Cancer Ther; 12(6) June 2013 989
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

References1. De LC, D'Alessio G. From immunotoxins to immunoRNases. Curr
Pharm Biotechnol 2008;9:210–4.2. Frankel AE. Reducing the immune response to immunotoxin. Clin
Cancer Res 2004;10:13–5.3. SmallshawJE,Ghetie V, Rizo J, Fulmer JR, Trahan LL,GhetieMA, et al.
Genetic engineering of an immunotoxin to eliminate pulmonary vas-cular leak in mice. Nat Biotechnol 2003;21:387–91.
4. Posey JA, Khazaeli MB, Bookman MA, Nowrouzi A, Grizzle WE,Thornton J, et al. A phase I trial of the single-chain immunotoxinSGN-10 (BR96 sFv-PE40) in patients with advanced solid tumors.Clin Cancer Res 2002;8:3092–9.
5. Hall PD, Virella G, Willoughby T, Atchley DH, Kreitman RJ, Frankel AE.Antibody response to DT-GM, a novel fusion toxin consisting of atruncated diphtheria toxin (DT) linked to human granulocyte-macro-phage colony stimulating factor (GM), during a phase I trial of patientswith relapsed or refractory acute myeloid leukemia. Clin Immunol2001;100:191–7.
6. Hertler AA, Spitler LE, Frankel AE. Humoral immune response to a ricinA chain immunotoxin in patients with metastatic melanoma. CancerDrug Deliv 1987;4:245–53.
7. OhS, Todhunter DA, Panoskaltsis-Mortari A, BuchsbaumDJ, Toma S,Vallera DA. A deimmunized bispecific ligand-directed toxin that showsan impressive anti-pancreatic cancer effect in a systemic nude mouseorthotopic model. Pancreas 2012;41:789–96.
8. Onda M, Beers R, Xiang L, Nagata S, Wang QC, Pastan I. An immu-notoxin with greatly reduced immunogenicity by identification andremoval of B cell epitopes. Proc Natl Acad Sci U S A 2008;105:11311–6.
9. MathewM, Verma RS. Humanized immunotoxins: a new generation ofimmunotoxins for targeted cancer therapy. Cancer Sci 2009;100:1359–65.
10. Kurschus FC, Jenne DE. Delivery and therapeutic potential of humangranzyme B. Immunol Rev 2010;235:159–71.
11. De LC, D'Alessio G. Human anti-ErbB2 immunoagents—immunoR-Nases and compact antibodies. FEBS J 2009;276:1527–35.
12. Lobe CG, Havele C, Bleackley RC. Cloning of two genes that arespecifically expressed in activated cytotoxic T lymphocytes. Proc NatlAcad Sci U S A 1986;83:1448–52.
13. Trapani JA, Klein JL, White PC, Dupont B. Molecular cloning of aninducible serine esterase gene from human cytotoxic lymphocytes.Proc Natl Acad Sci U S A 1988;85:6924–8.
14. Kanatani I, Lin X, Yuan X, Manorek G, Shang X, Cheung LH, et al.Targeting granzyme B to tumor cells using a yoked human chorionicgonadotropin. Cancer Chemother Pharmacol 2011;68:979–90.
15. Rosenblum MG, Barth S. Development of novel, highly cytotoxicfusion constructs containing granzyme B: unique mechanisms andfunctions. Curr Pharm Des 2009;15:2676–92.
16. Liu Y, Zhang W, Niu T, Cheung LH, Munshi A, Meyn RE Jr, et al.Targeted apoptosis activation with GrB/scFvMEL modulates melano-ma growth, metastatic spread, chemosensitivity, and radiosensitivity.Neoplasia 2006;8:125–35.
17. Liu Y, Cheung LH, Thorpe P, RosenblumMG.Mechanistic studies of anovel human fusion toxin composed of vascular endothelial growthfactor (VEGF)121 and the serine protease granzyme B: directed apo-ptotic events in vascular endothelial cells. Mol Cancer Ther 2003;2:949–59.
18. Liu Y, Cheung LH,HittelmanWN, RosenblumMG. Targeted delivery ofhuman pro-apoptotic enzymes to tumor cells: in vitro studies describ-ing a novel class of recombinant highly cytotoxic agents. Mol CancerTher 2003;2:1341–50.
19. Weidle UH, Georges G, Brinkmann U. Fully human targeted cytotoxicfusion proteins: new anticancer agents on the horizon. Cancer Geno-mics Proteomics 2012;9:119–33.
20. Stahnke B, Thepen T, Stocker M, Rosinke R, Jost E, Fischer R, et al.Granzyme B-H22(scFv), a human immunotoxin targeting CD64 inacute myeloid leukemia of monocytic subtypes. Mol Cancer Ther2008;7:2924–32.
21. Dalken B, Giesubel U, Knauer SK, Wels WS. Targeted induction ofapoptosis by chimeric granzyme B fusion proteins carrying antibody
and growth factor domains for cell recognition. Cell Death Differ2006;13:576–85.
22. Wang T, Zhao J, Ren JL, Zhang L, Wen WH, Zhang R, et al. Recom-binant immunoproapoptotic proteins with furin site can translocateand kill HER2-positive cancer cells. Cancer Res 2007;67:11830–9.
23. Cao Y, Marks JD, Marks JW, Cheung LH, Kim S, Rosenblum MG.Construction and characterization of novel, recombinant immunotox-ins targeting the Her2/neu oncogene product: in vitro and in vivostudies. Cancer Res 2009;69:8987–95.
24. Cao Y, Marks JD, Huang Q, Rudnick SI, Xiong C, Hittelman WN, et al.Single-chain antibody-based immunotoxins targeting Her2/neu:design optimization and impact of affinity on antitumor efficacy andoff-target toxicity. Mol Cancer Ther 2012;11:143–53.
25. Turk MJ, Reddy JA, Chmielewski JA, Low PS. Characterization of anovel pH-sensitive peptide that enhances drug release from folate-targeted liposomes at endosomal pHs. Biochim Biophys Acta 2002;1559:56–68.
26. Gully CP, Zhang F, Chen J, Yeung JA, Velazquez-Torres G, Wang E,et al. Antineoplastic effects of an Aurora B kinase inhibitor in breastcancer. Mol Cancer 2010;9:42.
27. Carter P, Presta L, Gorman CM, Ridgway JB, Henner D, Wong WL,et al. Humanization of an anti-p185HER2 antibody for human cancertherapy. Proc Natl Acad Sci U S A 1992;89:4285–9.
28. PlankC,OberhauserB,MechtlerK, KochC,WagnerE. The influenceofendosome-disruptive peptides on gene transfer using synthetic virus-like gene transfer systems. J Biol Chem 1994;269:12918–24.
29. Bongartz JP, Aubertin AM, Milhaud PG, Lebleu B. Improved biologicalactivity of antisense oligonucleotides conjugated to a fusogenic pep-tide. Nucleic Acids Res 1994;22:4681–8.
30. WangYC,Morrison G, Gillihan R, Guo J,Ward RM, Fu X, et al. Differentmechanisms for resistance to trastuzumab versus lapatinib in HER2-positive breast cancers–role of estrogen receptor and HER2 reacti-vation. Breast Cancer Res 2011;13:R121.
31. Liu L, Greger J, Shi H, Liu Y, Greshock J, Annan R, et al. Novelmechanismof lapatinib resistance inHER2-positive breast tumor cells:activation of AXL. Cancer Res 2009;69:6871–8.
32. Casi G, Neri D. Antibody–drug conjugates: basic concepts, examplesand future perspectives. J Control Release 2012;161:422–8.
33. Alley SC, Okeley NM, Senter PD. Antibody–drug conjugates: targeteddrug delivery for cancer. Curr Opin Chem Biol 2010;14:529–37.
34. King HD, Dubowchik GM, Mastalerz H, Willner D, Hofstead SJ,Firestone RA, et al. Monoclonal antibody conjugates of doxorubi-cin prepared with branched peptide linkers: inhibition of aggrega-tion by methoxytriethyleneglycol chains. J Med Chem 2002;45:4336–43.
35. Tai W, Mahato R, Cheng K. The role of HER2 in cancer therapy andtargeted drug delivery. J Control Release 2010;146:264–75.
36. Nagata S, Pastan I. Removal of B cell epitopes as a practical approachfor reducing the immunogenicity of foreign protein-based therapeu-tics. Adv Drug Deliv Rev 2009;61:977–85.
37. Madhumathi J, Verma RS. Therapeutic targets and recent advances inprotein immunotoxins. Curr Opin Microbiol 2012;15:300–9.
38. Liu W, Onda M, Lee B, Kreitman RJ, Hassan R, Xiang L, et al.Recombinant immunotoxin engineered for low immunogenicity andantigenicity by identifying and silencing human B-cell epitopes. ProcNatl Acad Sci U S A 2012;109:11782–7.
39. Onda M, Beers R, Xiang L, Lee B, Weldon JE, Kreitman RJ, et al.Recombinant immunotoxin against B-cell malignancieswith no immu-nogenicity in mice by removal of B-cell epitopes. Proc Natl Acad SciU S A 2011;108:5742–7.
40. Trapani JA, Sutton VR. Granzyme B: pro-apoptotic, antiviral andantitumor functions. Curr Opin Immunol 2003;15:533–43.
41. Chowdhury D, Lieberman J. Death by a thousand cuts: granzymepathways of programmed cell death. Annu Rev Immunol 2008;26:389–420.
42. Motyka B, Korbutt G, Pinkoski MJ, Heibein JA, Caputo A, Hobman M,et al. Mannose 6-phosphate/insulin-like growth factor II receptor is adeath receptor for granzyme B during cytotoxic T cell-induced apo-ptosis. Cell 2000;103:491–500.
Cao et al.
Mol Cancer Ther; 12(6) June 2013 Molecular Cancer Therapeutics990
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

43. Kurschus FC, Bruno R, Fellows E, Falk CS, Jenne DE. Membranereceptors are not required to deliver granzyme B during killer cellattack. Blood 2005;105:2049–58.
44. Lewis Phillips GD, Li G, Dugger DL, Crocker LM, Parsons KL, Mai E,et al. Targeting HER2-positive breast cancer with trastuzumab-DM1, an antibody–cytotoxic drug conjugate. Cancer Res 2008;68:9280–90.
45. Garrett JT, Arteaga CL. Resistance to HER2-directed antibodies andtyrosine kinase inhibitors: mechanisms and clinical implications. Can-cer Biol Ther 2011;11:793–800.
46. Bedard PL, de AE, Cardoso F. Beyond trastuzumab: overcomingresistance to targeted HER-2 therapy in breast cancer. Curr CancerDrug Targets 2009;9:148–62.
47. Hilgeroth A, Hemmer M, Coburger C. The impact of the induction ofmultidrug resistance transporters in therapies by used drugs: recentstudies. Mini Rev Med Chem 2012;12:1127–34.
48. Hurvitz SA, Kakkar R. The potential for trastuzumab emtansine inhuman epidermal growth factor receptor 2 positive metastatic breastcancer: latest evidence and ongoing studies. Ther Adv Med Oncol2012;4:235–45.
49. Murphy CG, Morris PG. Recent advances in novel targeted thera-pies for HER2-positive breast cancer. Anticancer Drugs 2012;23:765–76.
50. Kovtun YV, Audette CA, Mayo MF, Jones GE, Doherty H, Maloney EK,et al. Antibody-maytansinoid conjugates designed to bypass multi-drug resistance. Cancer Res 2010;70:2528–37.
Novel Functionalized GrB-Targeting Her2/neu
www.aacrjournals.org Mol Cancer Ther; 12(6) June 2013 991
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002

2013;12:979-991. Published OnlineFirst March 14, 2013.Mol Cancer Ther Yu Cao, Khalid A. Mohamedali, John W. Marks, et al. Serine Protease Therapeutics Targeting Her2/neuConstruction and Characterization of Novel, Completely Human
Updated version
10.1158/1535-7163.MCT-13-0002doi:
Access the most recent version of this article at:
Cited articles
http://mct.aacrjournals.org/content/12/6/979.full#ref-list-1
This article cites 50 articles, 20 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/12/6/979.full#related-urls
This article has been cited by 3 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/12/6/979To request permission to re-use all or part of this article, use this link
on June 13, 2020. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst March 14, 2013; DOI: 10.1158/1535-7163.MCT-13-0002
Related Documents











