Identification and characterization of novel carbapenemases Dissertation to obtain the degree Doctor Rerum Naturalium (Dr. rer. nat.) at the Faculty of Biology and Biotechnology International Graduate School of Biosciences Ruhr-University Bochum Department of Medical Microbiology Advisor: Prof. Dr. Sören G. Gatermann Second advisor: Prof. Dr. Franz Narberhaus Niels Ernst Pfennigwerth from Essen Bochum, April 2015 DISSERTATION submitted by

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Identification and characterizationof novel carbapenemases
Dissertation to obtain the degree
Doctor Rerum Naturalium (Dr. rer. nat.)
at the Faculty of Biology and Biotechnology
International Graduate School of Biosciences
Ruhr-University Bochum
Department of Medical Microbiology
Advisor: Prof. Dr. Sören G. GatermannSecond advisor: Prof. Dr. Franz Narberhaus
Niels Ernst Pfennigwerthfrom Essen
Bochum, April 2015
D I S S E R T A T I O N
submitted by

Identifizierung und Charakterisierungneuer Carbapenemasen
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften (Dr. rer. nat)
an der Fakultät für Biologie und Biotechnologie
Internationale Graduiertenschule Biowissenschaften
Ruhr-Universität Bochum
Abteilung für Medizinische Mikrobiologie
Referent: Prof. Dr. Sören G. GatermannKorreferent: Prof. Dr. Franz Narberhaus
Niels Ernst PfennigwerthEssen
Bochum, April 2015
D I S S E R T A T I O N
eingereicht von

Danksagung
Viele haben zu einem erfolgreichen Gelingen dieser Dissertation beigetragen. Einigen möchte ich
besonders danken.
Meinem Doktorvater Herrn Prof. Dr. Sören G. Gatermann danke ich sehr für seine fortwährende
Unterstützung, sein großes Vertrauen in meine Arbeit und die Möglichkeit, in diesem
interessanten Fachbereich zu promovieren.
Herrn Prof. Dr. Franz Narberhaus danke ich sehr für die freundliche Übernahme des Korreferats.
Herrn Dr. Alexander Stang und Herrn Prof. Klaus Überla danke ich für die Möglichkeit, das in
dieser Arbeit gefundene Plasmid in der Abteilung für Virologie zu sequenzieren.
Allen Mitarbeitern der Abteilung für medizinische Mikrobiologie danke ich für das tolle, nette
und freundschaftliche Arbeitsklima und für eine Hilfsbereitschaft, die nie zu enden scheint.
Besonders danke ich hierbei Frau Anja Kaminski für die Hilfe bei den isoelektrischen
Fokussierungen, Frau Anke Albrecht für ihre unverzichtbare Unterstützung bei den
Lokalisationsstudien und Frau Susanne Friedrich für ein immer offenes Ohr bei experimentellen
Problemen.
Danke auch an meine Masterstudent(in)en Lisei Meining, Alexander Hoffmann und Felix Lange
und meine S-Moduler für ihr Mitwirken an Teilen dieser Arbeit.
Ein besonders großer Dank geht an meine KoMaNePf-Mitinsassen Dr. Sandra Neumann,
Dr. Lennart Marlinghaus und Dr. Miriam Korte-Berwanger, ohne euch wären die letzten vier
Jahre um mindestens 90% unlustiger gewesen. Auch für viele fachliche Diskussionen - vielen
Dank!
(Fast) last, but not least: Ein riesiggroßer Dank geht an Herrn Dr. Martin Kaase für seine zu jeder
Zeit freundschaftliche Unterstützung, die zahllosen fruchtbaren fachlichen Diskussionen, das
kritische Korrekturlesen von Postern, Manuskripten und dieser Arbeit und als wandelndes
Lexikon für alle Fragen bezüglich der medizinischen Mikrobiologie. Vielen Dank!
Ein Dank, der so groß ist, dass ich ihn nicht in Worten auszudrücken vermag, gebührt zu guter
Letzt meinen Eltern, meiner Schwester und meiner Frau Freya, die mich zu jeder Zeit
bedingungslos unterstützt, ermutigt und aufgebaut haben. Vielen, vielen Dank!

Contents I
Contents
Contents ................................................................................................................................................ I
List of Figures ................................................................................................................................... IV
List of Tables ...................................................................................................................................... V
Abbreviations .................................................................................................................................. VI
1 Introduction ............................................................................................................................... 1
1.1 β-lactam antibiotics ....................................................................................................................................... 1
1.2 Target structures of β-lactam antibiotics: The bacterial cell wall synthesis ......................... 5
1.3 Mechanisms of antibiotic resistance ...................................................................................................... 8
1.4 β-lactamases .................................................................................................................................................. 10
1.4.1 Class A β-lactamases ......................................................................................................................... 12
1.4.2 Class B β-lactamases ......................................................................................................................... 13
1.4.3 Class C β-lactamases ......................................................................................................................... 14
1.4.4 Class D β-lactamases ........................................................................................................................ 14
1.5 Carbapenemases and their distribution ............................................................................................ 15
1.6 Mobility of β-lactamase genes ................................................................................................................ 16
1.7 Pseudomonas aeruginosa .......................................................................................................................... 19
1.8 Citrobacter freundii ..................................................................................................................................... 19
1.9 Objectives of this work .............................................................................................................................. 20
2 Material and Methods .......................................................................................................... 22
2.1 Material ............................................................................................................................................................ 22
2.1.1 Instruments .......................................................................................................................................... 22
2.1.2 Disposable material .......................................................................................................................... 23
2.1.3 Chemicals .............................................................................................................................................. 24
2.1.4 Antibiotics ............................................................................................................................................. 25
2.1.5 Wafers containing antibiotics ....................................................................................................... 26
2.1.6 Antibiotic gradient test strips ....................................................................................................... 26

Contents II
2.1.7 Kits und standards ............................................................................................................................ 26
2.1.8 Enzymes ................................................................................................................................................. 27
2.1.9 Antibodies ............................................................................................................................................. 27
2.2 Microbial strains, plasmids and oligonuclotides ............................................................................ 28
2.2.1 Microbial strains ................................................................................................................................ 28
2.2.2 Plasmids ................................................................................................................................................. 28
2.2.3 Oligonucleotides ................................................................................................................................. 29
2.3 Methods ........................................................................................................................................................... 37
2.3.1 Microbiological methods ................................................................................................................ 37
2.3.2 Phenotypic methods for antibiotic resistance analysis ..................................................... 39
2.3.3 Molecular biology methods ........................................................................................................... 40
2.3.4 Biochemical methods ....................................................................................................................... 46
2.3.5 In silico methods ................................................................................................................................. 50
3 Results ....................................................................................................................................... 51
3.1 The search for novel carbapenemases ............................................................................................... 51
3.1.1 Identification of IMP-31 in Pseudomonas aeruginosa NRZ-00156 ................................ 51
3.1.2 Identification of OXA-233 in Citrobacter freundii NRZ-02127 ........................................ 55
3.1.3 Identification of KHM-2 in Pseudomonas aeruginosa NRZ-03096 ................................. 58
3.2 Analysis of the genetic environment of blaIMP-31, blaOXA-233 and blaKHM-2 ............................... 61
3.2.1 Genetic environment of blaIMP-31 .................................................................................................. 61
3.2.2 Genetic environment of blaOXA-233 ................................................................................................ 62
3.2.3 Genetic environment of blaKHM-2 .................................................................................................. 63
3.3 Localization of blaIMP-31, blaOXA-233 and blaKHM-2 ................................................................................ 64
3.3.1 Localization of blaIMP-31 .................................................................................................................... 64
3.3.2 Localization of blaOXA-233 .................................................................................................................. 65
3.3.3 Localization of blaKHM-2 .................................................................................................................... 66
3.4 Impact of IMP-31, OXA-233 and KHM-2 on β-lactam resistance ............................................. 67
3.4.1 Impact of IMP-31 on β-lactam resistance ................................................................................ 68
3.4.2 Impact of OXA-233 on β-lactam resistance ............................................................................. 69

Contents III
3.4.3 Impact of KHM-2 on β-lactam resistance ................................................................................. 72
3.4.4 Comparison of IMP-31, OXA-233 and KHM-2 ........................................................................ 73
3.5 Purification of IMP-31, OXA-233 and KHM-2 .................................................................................. 74
3.6 Determination of kinetic parameters .................................................................................................. 77
3.6.1 Determination of kinetic parameters for IMP-31 ................................................................. 77
3.6.2 Determination of kinetic parameters for OXA-233 ............................................................. 79
3.6.3 Determination of kinetic parameters for KHM-2 ................................................................. 81
3.6.4 Comparison of the hydrolytic efficiencies of IMP-31, OXA-233 and KHM-2 ............. 82
3.7 Determination of the isoelectric point of IMP-31, OXA-233 and KHM-2 ............................. 83
3.8 Sequencing and characterization of the blaOXA-233 carrying plasmid pMB3018 ................ 84
4 Discussion ................................................................................................................................ 89
4.1 Identification of IMP-31 ............................................................................................................................ 89
4.2 Identification of OXA-233 ........................................................................................................................ 94
4.3 Identification of KHM-2 ............................................................................................................................ 95
4.4 Catalytic characteristics of IMP-31, OXA-233 and KHM-2 ......................................................... 98
4.4.1 Characteristics of IMP-31 ............................................................................................................... 98
4.4.2 Characteristics of OXA-233 ..........................................................................................................101
4.4.3 Characteristics of KHM-2 ..............................................................................................................106
4.5 Characterization of the blaOXA-233-carrying plasmid pMB3018 ...............................................108
4.6 Comparison of IMP-31, KHM-2 and OXA-233 and concluding remarks .............................109
5 Summary................................................................................................................................. 111
6 Zusammenfassung .............................................................................................................. 113
7 Bibliography .......................................................................................................................... 115
8 Appendix................................................................................................................................. 132
Publications ................................................................................................................................... 137
Curriculum vitae .......................................................................................................................... 139

List of Figures IV
List of Figures
Figure 1.1 Chemical structures of the backbone of β-lactam antibiotics.................................................... 3
Figure 1.2 Chemical structures of imipenem, meropenem, ertapenem and doripenem. .................... 4
Figure 1.3 Chemical structure of peptidoglycan from E. coli. .......................................................................... 6
Figure 1.4 Action of a serine β-lactamase against carbapenems. ............................................................... 12
Figure 1.5 Schematic organization of transporter insertion sequences and transposons. (A) ...... 17
Figure 1.6 Schematic structure of a class 1 integron. ...................................................................................... 19
Figure 3.1 Modified Hodge Test and EDTA-CDT of P. aeruginosa NRZ-00156. .................................... 51
Figure 3.2 Amino acid sequence alignment of IMP-31, IMP-35 and IMP-1. ........................................... 53
Figure 3.3 Phylogenetic analysis of IMP-31. ........................................................................................................ 54
Figure 3.4 Modified Hodge Test of C. freundii NRZ-02127. ........................................................................... 55
Figure 3.5 Amino acid sequence alignment of OXA-233, OXA-17 and OXA-10. .................................... 57
Figure 3.6 Modified Hodge Test and EDTA-CDT of P. aeruginosa NRZ-03096. .................................... 58
Figure 3.7 Amino acid sequence alignment of KHM-2 and KHM-1. ........................................................... 60
Figure 3.8 Genetic environment of blaIMP-31 in P. aeruginosa NRZ-00156. .............................................. 61
Figure 3.9 Genetic environment of blaOXA-233 in C. freundii NRZ-02127. .................................................. 62
Figure 3.10 Genetic environment of blaKHM-2 in P. aeruginosa NRZ-03096. ........................................... 63
Figure 3.11 Localization of blaIMP-31......................................................................................................................... 65
Figure 3.12 Localization of blaOXA-233. ..................................................................................................................... 66
Figure 3.13 Localization of blaKHM-2. ........................................................................................................................ 67
Figure 3.14 Ion exchange (A) and gel filtration (B) chromatograms of the KHM-2 FPLC. ............... 75
Figure 3.15 SDS-PAGE analysis of enzyme preparations of IMP-31, IMP-1, OXA-233, OXA-10,
KHM-2 and KHM-1. ........................................................................................................................................................ 76
Figure 3.16 Hydrolysis assay of IMP-31 for imipenem and Michaelis-Menten plot. .......................... 78
Figure 3.17 CO2-dependent imipenem hydrolysis of OXA-233. ................................................................. 80
Figure 3.18 Isoelectric focussing of OXA-233, OXA-10, IMP-31, IMP-1, KHM-2 and KHM-1. ......... 84
Figure 3.19 Circular map of pMB3018. .................................................................................................................. 85
Figure 3.20 Comparison of pMB3018, pJIE137, p271A, pECS01 and pTR3. .......................................... 87
Figure 4.1 Comparison of the genetic environment of blaIMP-31 and blaIMP-35. ........................................ 92
Figure 4.2 Crystal structure and homology model of the active site of IMP-1 (A) and IMP-31 (B).
..............................................................................................................................................................................................100
Figure 4.3 Crystal structure and homology model of the active sites of OXA-10 (A) and OXA-233
(B). .......................................................................................................................................................................................104
Figure 4.4 Chemical structures of ceftazidime, aztreonam and penicillin G. .......................................105
Figure 4.5 Homology models of KHM-1 (A) and KHM-2 (B). ......................................................................107

List of Tables V
List of Tables
Table 1.1 Classification schemes for β-lactamases according to Bush & Jacoby (2010) and Ambler
(1980). ................................................................................................................................................................................. 11
Table 2.1. Microbial strains used in this study. .................................................................................................. 28
Table 2.2. Plasmids used in this study. .................................................................................................................. 28
Table 2.3: Oligonucleotides used in this study. .................................................................................................. 29
Table 3.1 β-lactam MICs of P. aeruginosa NRZ-00156. ................................................................................... 52
Table 3.2 MLS typing of P. aeruginosa NRZ-00156. .......................................................................................... 55
Table 3.3 β-lactam MICs of C. freundii NRZ-02127. .......................................................................................... 56
Table 3.4 β-lactam MICs of P. aeruginosa NRZ-03096. ................................................................................... 59
Table 3.5 MLS typing of P. aeruginosa NRZ-03096. .......................................................................................... 60
Table 3.6 β-lactam MICs of E. coli TOP10 transformed with the pBK-CMV vector and
IMP-31/IMP-1 expressing E. coli TOP10. .............................................................................................................. 68
Table 3.7 β-lactam MICs of E. coli TOP10 transformed with the pBK-CMV vector and
OXA-233/OXA-10 expressing E. coli TOP10. ....................................................................................................... 70
Table 3.8 β-lactam MICs of the E. coli C600 OXA-233 pMB3018-transconjugant and E. coli C600.
................................................................................................................................................................................................ 71
Table 3.9 β-lactam MICs of E. coli TOP10 transformed with the pBK-CMV vector and
KHM-2/KHM-1 expressing E. coli TOP10. ............................................................................................................ 72
Table 3.10 Relative MIC increases of E. coli TOP10 producing IMP-31, OXA-233 and KHM-2....... 73
Table 3.11 Kinetic parameters of IMP-31. ............................................................................................................ 79
Table 3.12 Kinetic parameters of OXA-233.......................................................................................................... 80
Table 3.13 Kinetic parameters of KHM-2. ............................................................................................................ 81
Table 3.14 Comparison of the hydrolytic efficiencies of IMP-31, OXA-233 and KHM-2. .................. 83

Abbreviations VI
Abbreviations
All abbreviations that are not listed here are either part of the International System of Units
(Système international d’unités, SI) or abbreviations of chemicals that are mentioned in the
Materials and Methods section (Chapter 2).
aa Amino acid
A. dest Aqua destilata (lat.), distilled water
AMP Ampicillin
AmpR Ampicillin resistance
AP Alkaline phosphatase
BLAST Basic Local Alignment Search Tool
bp Base pairs
BSA Bovine serum albumin
CHDL Carbapenem-hydrolyzing class D β-lactamase
CDT Combined-disk test
DNA Deoxyribonucleid acid
ECDC European Centre for disease prevention and control
ESBL Extended-spectrum β-lactamase
ETP Ertapenem
EUCAST European Committee on Antimicrobial Susceptiblity
Testing
FOX Cefoxitin
FPLC Fast protein liquid chromatography
GF Gel filtration
HAI Healthcare-associated infections
IEF Isoelectric focussing
IEX Ion exchange
IMP Imipenem
IR Inverted repeats
kb kilo base pairs
KmR Kanamycin resistance
mAU milli absorbance units
MBL metallo-β-lactamase
Mbp Mega base pairs

Abbreviations VII
MCS Multiple cloning site
MDR Multidrug-resistant
MEM Meropenem
MIC Minimal inhibitory concentration
NCBI National Centre for Biotechnology Information
NRZ National Reference Laboratory for multidrug-resistant
Gram-negative bacteria (“Nationales Referenzzentrum für
Gram-negative Krankenhauserreger”)
OD Optical density
ORF Open reading frame
PAGE Polyacrylamide gel electrophoresis
PCR Polymerase chain reaction
PFGE Pulsed-field gel electrophoresis
pI Isoelectric point
RifR Rifampicin resistance
(r)RNA (ribosomal) Ribonucleic acid
TBE Tris-boric acid-EDTA buffer
v/v volume per volume
w/v weight per volume

Introduction 1
1 Introduction
Antibiotic resistance in clinically relevant bacteria is a major challenge to healthcare systems
worldwide. Especially the ongoing spread and diversification of resistance mechanisms in Gram-
negative pathogens is a worrying development. Gram-negative pathogens, such as
Escherichia coli, Klebsiella pneumoniae, other members of the Enterobacteriaceae,
Pseudomonas aeruginosa and Acinetobacter baumannii can cause severe infections and are a
major threat to critically ill hospitalized patients (Gaynes & Edwards, 2005). Studies of the
European Centre for Disease Prevention and Control (ECDC) estimated that 1.9 to 5.2 million
patients per year in Europe are infected with bacterial pathogens in context of a medical
treatment and that 75 % of these healthcare-associated infections (HAI) result from
hospitalization (Suetens et al., 2013). In an ECDC surveillance study with data from over 250,000
patients affected by HAI in 2011 and 2012, infections with E. coli were the most prevalent with
15.9 %, followed by Staphylococcus aureus infections with 12.3 % (Suetens et al., 2013). While
the main focus of antimicrobial treatment of the last decades was set on S. aureus infections,
especially with the methicillin-resistant S. aureus (MRSA), the most threatening development
nowadays is the increasing number of Gram-negative pathogens that are resistant to antibiotics
(ECDC, 2013; Suetens et al., 2013). Many Gram-negative species are intrinsically resistant to
single antibiotics, but in the last decades, these pathogens have acquired numerous resistance
genes, becoming multidrug-resistant (MDR) or pan-resistant and limiting the treatment options
in many cases dramatically (Falagas & Bliziotis, 2007). In this context, antibiotic resistance has
been listed as one of the greatest threats to human health in the most recent World Economic
Forum Global Risks Reports (World Economic Forum, 2013 & 2014). As there is a lack in
development of novel antibiotics against Gram-negative pathogens due to economical and
organizational reasons (Appelbaum, 2012) and as only few novel antibacterial drugs are
expected to be clinically available in the next years, the situation is predicted to escalate further
(Boucher et al., 2013). In this context, the identification and characterization of resistance
mechanisms in Gram-negative bacteria and the correct treatment of patients infected with these
bacteria in combination with strict hygiene management is the major challenge to antimicrobial
treatment and infection control precautions for the next years.
1.1 β-lactam antibiotics
β-lactam antibiotics are the most important class of antibiotics and were first discovered in 1929
by Sir Alexander Fleming, as he observed the inhibitory effect of a Penicillium notatum mycelium
that contaminated a Staphylococcus colony on an agar plate. Although Fleming was not the first
to observe the antibiosis between fungal and bacteria, he was the first to study one of the

Introduction 2
substances that inhibit bacterial growth and named it penicillin (Kong et al., 2010). In 1940,
penicillin was purified at higher levels and was sucessfully used to treat patients with S. aureus
infections. Penicillin became finally available in the open market in 1946 (Kong et al., 2010).
Several derivatives of penicillin were found or developed in the following decades, constituting
four groups of β-lactams: the penicillins, the cephalosporins, the carbapenems and the
monobactams (Kong et al., 2010; Papp-Wallace et al., 2011).
Penicillins
The penicillins were the first β-lactams in clinical use and were widely used in the beginning of
the antibiotic era. The structure of the molecules is based upon the four-membered β-lactam
ring and an annulated five-membered thiazolidine ring with varying side chains (Figure 1.1).
The thiazolidine ring exhibits sulfur at position C-1. Penicillins are classified into several groups
based upon their origin. The natural penicillins benzylpenicillin (Penicillin G) and
phenoxymethylpenicillin (Penicillin V) were isolated from different variants of
Penicillium chrysogenum and are higly active against sensitive strains of Gram-positive cocci,
therefore sparing most current strains of S. aureus (Mascaretti, 2003). Methicillin on the other
hand is an antistaphylococcal β-lactamase-resistant penicillin and was widely used in therapy
against S. aureus infections but is no longer available nowadays. Other members of this group
are the isoxazolyl-penicillins oxacillin, cloxacillin and dicloxacillin. The aminopenicillins include
ampicillin, bacampicillin and amoxicillin. They have a broader spectrum, including several Gram-
negatives like E. coli or Proteus mirabilis, as they are more capable of penetrating the outer
membrane of these bacteria (Mascaretti, 2003). The last group are the antipseudomonal
pencillins, which are semisynthetic derivates of penicillanic acid. They are categorized into two
subgroups: the carboxypenicillins, including carbenicillin and ticarcillin and the
ureidopenicillins, which include piperacillin and mezlocillin. Notably, piperacillin shows high
activity against P. aeruginosa and Enterobacteriaceae, making it an important treatment option
for infections with these species (Mascaretti, 2003).
Cephalosporins
The first cephalosporin, cephalosporin C, was isolated in 1953 from Cephalosporium acremonium
and the structure was determined in 1961 (Abraham & Newton, 1961). Cephalosporins consist
of the β-lactam ring, an annulated six-membered dihydrothiazine ring and two varying side
chains (Figure 1.1). They are categorized into four to five generations based upon their
characteristics regarding antimicrobial activity, resistance to β-lactamases and membrane
penetrability (Mascaretti, 2003). The first generation includes cephalotin, cefazolin and others

Introduction 3
Figure 1.1 Chemical structures of the backbone of β-lactam antibiotics. All β-lactam antibiotics share the four-membered β-lactam ring. Penicillins and cephalosporins possess a sulfur in the annulated thiazolidine ring while carbapenems exhibit a carbon at this position. In cephalosporins, the thiazolidine ring is six-membered, while it is five-membered in penicillins and carbapenems. that show high antibacterial activity against Gram-positive cocci, but are less effective against
E. coli, P. mirabilis and Klebsiella pneumoniae. The second generation is subgrouped and includes
the true cephalosporins, the cephamycins and the carbacephems. The cephalosporins of this
group exhibit higher activity against Haemophilus influenzae, Neisseria meningitidis,
staphylococci and streptococci than first-generation cephalosporins. An example for this group
is cefuroxime. Cephamycins on the other hand show increased antibacterial action against Gram-
negative bacteria and Bacteroides spp. and possess a –OCH3 group as a third side chain,
increasing their stability to certain β-lactamases and their antibacterial activity. They are less
effecive against staphylococci and streptococci (Mascaretti, 2003). Examples for clinically used
cephamycins are cefoxitin and cefotetan. Loracarbef is the only carbacephem and is not a true
cephalosporin but closely related. The third-generation cephalosporins, or oxyimino-
cephalosporins, exhibit significantly higher activity against Gram-negative bacteria than the first
and second generations. They are more stable to β-lactamases and have a broader spectrum,
including E. coli, Klebsiella spp., P. mirabilis, Citrobacter spp., Serratia marcescens,
Streptococcus pneumoniae, Streptococcus pyogenes and others (Mascaretti, 2003). Clinically
important members of this generation are cefotaxime, ceftriaxone and ceftazidime. The fourth
generation of cephalosporins is characterized by higher antimicrobial activity against some
Enterobacteriaceae, with cefepime and cefpirome being the only members of this generation
(Mascaretti, 2003). Two novel cephalosporins with activity against MRSA are ceftobiprole and
ceftaroline, which are classified as the fifth generation of cephalosporins (Bush et al., 2007;
Saravolatz et al., 2011).
Carbapenems
The first carbapenem, thienamycin, was discovered in 1976 in Streptomyces cattleya and served
as the model compound for all carbapenems. In contrast to many penicillins and cephalosporins,
the antimicrobial activity was shown for a broad range of bacteria, including even Gram-
negative organisms that are intrinsically resistant to many β-lactams, like P. aeruginosa (Tally et

Introduction 4
al., 1978; Weaver et al., 1979; Fainstein et al., 1982). In contrast to penicillins and
cephalosporins, the carbapenems exhibit a carbon for sulfur substitution at position C-1 of the
five-membered annulated ring (Figure 1.1). This carbon atom is responsible for the increased
stability against β-lactamases and the broad-spectrum of this class of β-lactams (Papp-Wallace et
al., 2011). As thienamycin was unstable in aqueous solutions, the search for derivatives was
intensified, leading to the development of imipenem. Imipenem became clinically available in
1985 and demonstrated high target affinity and stability against β-lactamases (Hashizume et al.,
1984; Kong et al., 2010). Imipenem is the N-formimidoyl derivative of thienamycin (Figure 1.2)
and is active against many Gram-positive and Gram-negative species. It has an increased
inhibitory effect on most members of the Enterobacteriaceae and can be used to treat
P. aeruginosa infections when combined with an aminoglycoside (Mascaretti, 2003). As
imipenem is metabolized by the human renal dehydropeptidase-1 (DHP-1), it is combined with
an inhibitor of this enzyme, cilastatin, in therapeutic use (Kropp et al., 1982; Norrby et al., 1983).
Today, three other carbapenems besides imipenem are in clinical use: meropenem, ertapenem
and doripenem. Meropenem possesses a 1-β-methyl group on position C-1 of the carbapenem
backbone (Figure 1.2) and is active against a broad range of Gram-positive and Gram-negative
pathogens with slightly elevated activity against Gram-negatives compared to imipenem. It is
significantly more stable against degradation by DHP-1 (Mascaretti, 2003) due to the 1-β-methyl
group. Ertapenem also possesses a 1-β-methyl group on position C-1 (Figure 1.2) and has high
activity against many Gram-positive and Gram-negative bacteria, but is weak against
Figure 1.2 Chemical structures of imipenem, meropenem, ertapenem and doripenem. The structure is based upon the β-lactam ring and an annulated five-membered thiazolidine ring. In contrast to imipenem, meropenem, ertapenem and doripenem possess a methyl group at position C-1 of the thiazolidine ring, confering stability against the human renal dehydropeptidase DHP-1.

Introduction 5
Acinetobacter spp. and Pseudomonas aeruginosa (Zhanel et al., 2005; Burkhardt et al., 2007).
Doripenem on the other hand shows excellent activity against P. aeruginosa but also reduced
activity against Acinetobacter spp. (Paterson & Depestel, 2009). The structure of doripenem is
very similar to meropenem, with the dimethylcarbamoyl side chain of meropenem replaced with
a sulfamoylaminomethyl group in doripenem (Figure 1.2).
Carbapenems are considered as antibiotics of last resort and should exclusively be used for
therapy of critically ill patients infected with multidrug-resistant bacteria that are still
susceptible to carbapenems (Papp-Wallace et al., 2011).
Monobactams
Monobactams are characterized by their molecular structure, which exhibits a four-membered
β-lactam ring without any annulated secondary ring structure in contrast to the bicyclic
penicillins, cephalosporins and carbapenems (Singh, 2004). The only clinically available member
of this group is aztreonam, a totally synthetic antibiotic. It has specific activity against a wide
range of β-lactamase-producing Gram-negative bacteria, including P. aeruginosa (Mascaretti,
2003). Furthermore, aztreonam shows increased stability to β-lactamases and has a high and
exclusive affinity for the PBP3 transpeptidase of Gram-negative bacteria, also known as FtsI
(Mascaretti, 2003; Kong et al., 2010).
1.2 Target structures of β-lactam antibiotics: The bacterial cell wall synthesis
The mode of action of β-lactam antibiotics is the inhibition of cell wall synthesis in Gram-positive
and Gram-negative bacteria. The cell wall of bacteria is located outside of the cytoplasmic
membrane of almost all bacteria and protects the cell integrity by withstanding the turgor
(Vollmer et al., 2008). The cell shape is also influenced by the cell wall and it is important for the
anchoring of other components of the cell envelope, for example transmembrane proteins
(Dramsi et al., 2008) or teichonic acids (Neuhaus & Baddiley, 2003). While cell walls are found in
nearly every bacterial species that is clinically relevant, they are absent in Mycoplasmas,
Planctomyces, Rickettsia spp. and Chlamydiae (Vollmer et al., 2008). The cell wall is formed by
layers of the polymeric molecule peptidoglycan, which is illustrated in Figure 1.3. Peptidoglycan
is formed by chains of repeating units of the disaccharide N-acetylglucosamine-N-acetylmuramic
acid (GlcNAC-MurNAc) that are cross-linked by short polypeptides, while the saccharides are
linked by β-1→4 bonds (Vollmer et al., 2008; Silhavy et al., 2010). The cross-linking peptide stem
is most often formed by L-Ala-γ-D-Glu-meso-A2pm-D-Ala-D-Ala, where diaminopimelic acid
(A2pm) can be replaced by L-Lys. The terminal D-Ala is present only in the nascent molecule and
is lost in the mature form (Vollmer et al., 2008). The cross-linking occurs between the carboxyl
group of D-Ala and the amino group of the diaminopimelic acid or lysine and the peptide stems
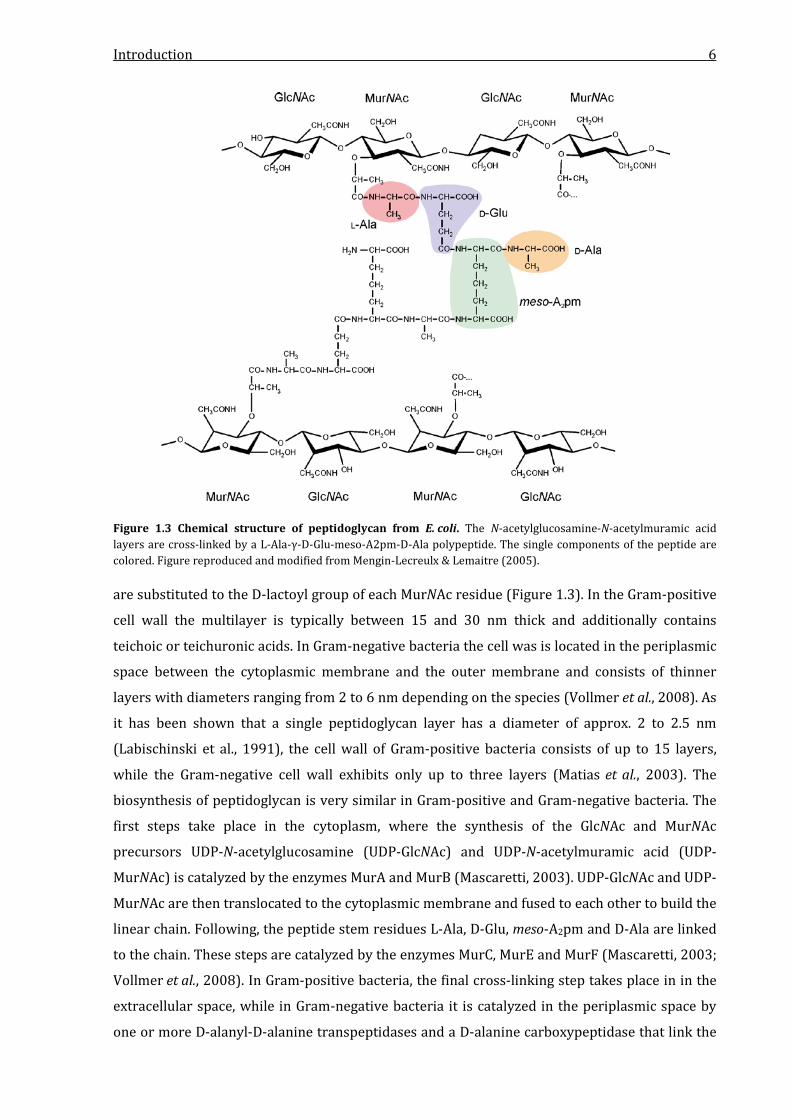
Introduction 6
Figure 1.3 Chemical structure of peptidoglycan from E. coli. The N-acetylglucosamine-N-acetylmuramic acid layers are cross-linked by a L-Ala-γ-D-Glu-meso-A2pm-D-Ala polypeptide. The single components of the peptide are colored. Figure reproduced and modified from Mengin-Lecreulx & Lemaitre (2005). are substituted to the D-lactoyl group of each MurNAc residue (Figure 1.3). In the Gram-positive
cell wall the multilayer is typically between 15 and 30 nm thick and additionally contains
teichoic or teichuronic acids. In Gram-negative bacteria the cell was is located in the periplasmic
space between the cytoplasmic membrane and the outer membrane and consists of thinner
layers with diameters ranging from 2 to 6 nm depending on the species (Vollmer et al., 2008). As
it has been shown that a single peptidoglycan layer has a diameter of approx. 2 to 2.5 nm
(Labischinski et al., 1991), the cell wall of Gram-positive bacteria consists of up to 15 layers,
while the Gram-negative cell wall exhibits only up to three layers (Matias et al., 2003). The
biosynthesis of peptidoglycan is very similar in Gram-positive and Gram-negative bacteria. The
first steps take place in the cytoplasm, where the synthesis of the GlcNAc and MurNAc
precursors UDP-N-acetylglucosamine (UDP-GlcNAc) and UDP-N-acetylmuramic acid (UDP-
MurNAc) is catalyzed by the enzymes MurA and MurB (Mascaretti, 2003). UDP-GlcNAc and UDP-
MurNAc are then translocated to the cytoplasmic membrane and fused to each other to build the
linear chain. Following, the peptide stem residues L-Ala, D-Glu, meso-A2pm and D-Ala are linked
to the chain. These steps are catalyzed by the enzymes MurC, MurE and MurF (Mascaretti, 2003;
Vollmer et al., 2008). In Gram-positive bacteria, the final cross-linking step takes place in in the
extracellular space, while in Gram-negative bacteria it is catalyzed in the periplasmic space by
one or more D-alanyl-D-alanine transpeptidases and a D-alanine carboxypeptidase that link the

Introduction 7
lineal peptidoglycan chain units. The D-alanyl-D-alanine transpeptidases and the D-alanine
carboxypeptidase are also known as penicillin-binding-proteins (PBPs), as they are the primary
target of β-lactam antibiotics (Mascaretti, 2003).
Several PBPs have been described, which significantly differ in β-lactam affinity and vary from
species to species. In E. coli, six PBPs, PBP1 to PBP6, were identified (Spratt & Pardee, 1975) and
numbered descending according to their molecular weight (Mascaretti, 2003). Similar numbers
were found in P. aeruginosa, Enterobacter cloacae, Salmonella typhimurium and S. marcescens
(Georgopapadakou & Liu, 1980; Kong et al., 2010). Gram-positive cocci on the other hand
possess only four PBPs, while some Bacillus species express up to eight (Suginaka et al., 1972).
Especially the low molecular mass PBPs, PBP5, PBP6 and PBP7 were only found in bacilli
(Georgopapadakou & Liu, 1980). PBP1 of E. coli is subdivided into three components, PBP1a,
PBP1b and PBP1c (Spratt & Jobanputra, 1977; Schiffer & Holtje, 1999). PBP1a and PBP1b
function as transglycosylases and transpeptidases, while PBP1c is only a transglycosylase and
the exact function of PCP1c is not known (Sauvage et al., 2008). PBP1-like enzymes catalyze the
peptidoglycan synthesis at the growing zones of the cell wall sides and are effectively inhibited
by pencillin G, most cephalosporins, imipenem and doripenem (Mascaretti, 2003; Breilh et al.,
2013). PBP2 and PBP3 are transpeptidases. While PBP2 catalyzes the initiation of peptidoglycan
insertion at growth sites, PBP3 is needed for formation of the cross-wall at cell division (Spratt,
1975; Mascaretti, 2003; den Blaauwen et al., 2008). PBP2 is one of the main target structures of
all carbapenems, whereas PBP3 strongly binds many cephalosporins, piperacillin, meropenem,
doripenem and aztreonam (Mascaretti, 2003; Breilh et al., 2013). The lower molecular mass
PBPs of E. coli play a role in cell separation, peptidoglycan maturation or recycling (Sauvage et
al., 2008). PBP4 (divided into PBP4a and PBP4b) and PBP7 function as endopeptidases that
cleave cross-bridges between two glycan chains. PBP5 is the major carboxypeptidase that
cleaves the terminal D-Ala-D-Ala bond. This cleavage prevents the transpeptidation of the stem
peptide (Sauvage et al., 2008). The role of PBP6a and PBP6b is not completely understood, but
both enzymes are carboxypeptidases like PBP5 and are assumed to be involved in the control of
peptidoglycan extent and/or peptidoglycan recycling (Sauvage et al., 2008). While PBP4a and
PBP4b have high affinity for penicillin G, ampicillin and imipenem, PBP5 is a major target
structure of cefoxitin and imipenem. PBP7 has high affinity for all carbapenems (Mascaretti,
2003; Breilh et al., 2013).
Inhibition of PBPs by β-lactam antibiotics
The bacterial cell wall is subject to permanent maintenance, controlled degradation and
resynthesis. An inhibition of the essential enzymes involved in this process inevitably leads to
instability of the wall, resulting in lysis and cell death (Mascaretti, 2003). The inactivation of
PBPs by β-lactams and therewith the inhibition of cell wall synthesis is based upon the covalent

Introduction 8
binding and the formation of a stable acyl-ester between the PBP and the antibiotic (Zapun et al.,
2008). β-lactams mimic the D-Ala-D-Ala dipeptide necessary for peptidoglycan crosslinking and
are bound by the PBPs. The active site serine of the PBP attacks the carbonyl group of the β-
lactam ring which leads to the opening of the ring and covalent binding to the enzyme. As this
complex is hydrolyzed with extremely low efficiency, it is equivalent to an inactivation of the
enzyme (Zapun et al., 2008). From crystal structure analysis, several PBP-β-lactam binding
characteristics were analyzed, showing similarities to the PBP4a-α-aminopimelyl-ε-D-alanyl acyl
anzyme and therewith the binding of PBPs to cell wall components (Sauvage et al., 2008). Crystal
structures showed that the active site serine of PBPs is covalently linked to the antibiotic and the
amide group of the β-lactam side chain is inserted between the second motif and the backbone
of the β3 sheet of the PBP. In addition, the thiazolidine ring-associated carboxylate binds to one
or both hydroxyl groups of the PBPs KTGT motif. As a third characteristic, the carbonyl oxygen
of the β-lactam lies in the oxyanion hole of the PBP (Sauvage et al., 2008). As the PBP4 enzymes,
PBP5, the PBP6 enzymes and PBP7 are not essential for growth in E. coli (Denome et al., 1999),
the bacteriolytic effect of β-lactam antibiotics is based upon the inhibition of the PBP1 enzymes,
PBP2 and PBP3 (Mascaretti, 2003; Sauvage et al., 2008).
1.3 Mechanisms of antibiotic resistance
Antibiotic resistance can be caused by a variety of molecular mechanisms. The resistance can be
based upon antibiotic target mutation or modification, prevention of drug penetration, active
efflux of antibiotics, bypass of antibiotic inhibition or enzymatic inactivation of the antibiotic
substance (Blair et al., 2015).
Target mutation
As many antibiotics specifically bind to their targets, a mutation of the target can lead to a
decreased or prevented binding, leading to insusceptibility to the antibiotic. An example for this
mechanism of resistance is the resistance to quinolones in several Gram-negative bacteria and
Staphylococcus aureus. Quinolones inhibit the the bacterial enzymes DNA gyrase and
topoisomerase IV that are responsible for negative supercoil introduction into the DNA (Kim &
Hooper, 2014). Mutations in the gyrA and parC genes lead to changes in the active site of the
enzyme, resulting in decreased inhibition by quinolones and increased resistance (Kim &
Hooper, 2014).
Enzymatic target modification
The resistance to an antibiotic can be based upon target modification. One example is the
methylation of the ribosomal 23S subunit by the chloramphenicol-florfenicol (cfr)
methytransferase. The cfr gene was first described in staphylococci, but is meanwhile found in

Introduction 9
many Gram-positive and Gram-negative pathogens (Shen et al., 2013). The gene encodes for a
methyltransferase which specifically methylates A2503 in the 23S rRNA, confering resistance to
different classes of antibiotics that target the ribosomal 23S rRNA subunit, for example
streptogamins and lincosamides (Long et al., 2006).
Enzymatic bypass
The most well-known example for enzymatic bypass of antibiotic inhibition is the methicillin-
resistant S. aureus (MRSA). This bacterium is resistant to almost all β-lactam antibiotics and
harbours the mecA gene or, more recently, the mecC gene. These genes code for the alternative
transpeptidase PBP2a that is not inhibited by β-lactams except ceftobiprole and ceftaroline
(Hartman & Tomasz, 1984; Lim & Strynadka, 2002; Bush et al., 2007; Garcia-Alvarez et al.,
2011). As the mode of action of β-lactams is the inhibition of bacterial cell wall synthesis, an
alternative transpeptidase can replace the function of the inhibited enzymes, allowing cell
growth.
Reduced permeability
In Gram-negative bacteria, many antibiotics have to enter the periplasm through non-specific
channels, the outer membrane porins (Miller et al., 1972). By mutation or downregulation of the
opr genes and by replacement of porins with more-specific channel proteins, the uptake of
antibiotics into the cell can be reduced, resulting in increased resistance (Balasubramanian et al.,
2011). For example, the mutation or loss of the OprD porin in Gram-negative bacteria can lead to
higher resistance against the carbapenem imipenem (Sanbongi et al., 2009).
Active efflux
An example for active efflux of antibiotics is the resistance to tetracyclines based on the
expression of tet genes. These genes code for membrane transporters that specifically export
tetracyclines and are found in both Gram-positive and Gram-negative pathogens (Kong et al.,
2009). Furthermore, transporters that are able to export a wide range of antibiotics, the
multidrug resistance efflux pumps, have been described. The best characterized MDR efflux
pumps are the resistance nodulation division (RND) family exporters (Blair et al., 2015). RND
transporters are able to confer clinically relevant levels of resistance against an extremely wide
range of antibiotics (Piddock, 2006) and are found mostly in Gram-negative bacteria (Blair et al.,
2015).
Enzymatic modification of antibiotics
The most important mechanism of resistance in Gram-negative bacteria is the enzymatic
degradation or modification of antibiotics. For example, aminoglycoside resistance is mediated

Introduction 10
by production of phosphotransferases (APH), acetyltransferases (AAC) or
nucleotidyltransferases (ANT) which modify the antibiotics, leading to an inactivation
(Abrahams, 1941). ANTs catalyze the transfer of an AMP from an ATP molecule to a hydroxyl
group in the aminoglycoside and thereby inactivate the drug. APHs transfer a phosphate residue
to the aminoglycoside at different positions and are grouped into seven subgroups (Ramirez &
Tolmasky, 2010). However, the most important group is the AAC group of enzymes. These
enzymes catalyze the acetylation of -NH2 groups in the aminoglycoside molecule at different
positions, subgrouping them into the AAC(1), AAC(3), AAC(2´) and AAC(6´) enzymes. (Ramirez &
Tolmasky, 2010).
However, the by far most clinically relevant example of enzymatic inactivation is the hydrolysis
of β-lactam antibiotics by β-lactamases.
1.4 β-lactamases
Resistance to β-lactam antibiotics in Gram-negative bacteria can be based upon four
mechanisms: i) The enzymatic bypass by expression of a β-lactam-resistant alternative
transpeptidase, as it the case for MRSA; ii) the loss of porins which leads to reduced outer
membrane permeability; iii) the mutation of one or more PBPs and iv) the enzymatic
inactivation by β-lactamases (Drawz & Bonomo, 2010).
β-lactamases are bacterial enzymes encoded by bla genes that can specifically bind and
hydrolyse β-lactam antibiotics, leading to the irreversible destruction of the drug. They are the
most common cause of resistance to β-lactams (Livermore, 1995) and in 2015, more than 1,500
unique β-lactamase protein sequences have been assigned (http://www.lahey.org/studies).
These are distinguished by their unique 3-letter name and a number (e.g. NDM-1 for “New Delhi
metallo-β-lactamase 1”). The enzymes can roughly be classified by their substrate spectrum.
Narrow-spectrum β-lactamases are able to hydrolyze penicillins, while extended-spectrum β-
lactamases (ESBLs) are able to hydrolyze penicillins and cephalosporins. Carbapenemases on
the other hand are able to hydrolyze penicillins, carbapenems and mostly cephalosporins and
thus can be the cause for resistance against almost all β-lactam antibiotics (Cantón et al., 2012a).
However, two more detailed classification schemes for β-lactamases exist. The first system is
based on functional characteristics, such as preferred substrates or inhibitor profiles. The aim of
the functional classification is a correlation of enzymes to their phenotype in clinical isolates
(Bush et al., 1995; Bush & Jacoby, 2010). The second scheme was developed by Ambler (1980)
and is based on the amino acid sequences of the enzymes. It classifies β-lactamases into
molecular class A, B, C and D enzymes. This scheme is commonly used in the literature and will
be the one used in this study. Both systems and their characteristics are summarized in Table
1.1. The hydrolysis mechanism can be based upon two enzyme architectures. β-lactamases of
the molecular classes A, C and D possess a serine residue in their active site that is responsible
Introduction 11
for an nucleophilic attack of the hydroxyl group of the serine on the carbonyl group of the β-
lactam ring (Figure 1.4). This results in the formation of a covalent acyl ester. Hydrolysis of the
ester utilizing a catalytic water molecule finally leads to the separation of the complex, leaving
the intact and active enzyme and the inactivated β-lactam (Livermore, 1995; Drawz & Bonomo,
2010). With the formation of a covalenty bound acyl enzyme, the mechanism is similar to the
inhibition of PBPs by β-lactams (Ghuysen, 1991). In contrast to PBPs, where the hydrolysis is of
such a low rate that it effectively leads to an inhibition, the hydrolysis by serine β-lactamases is
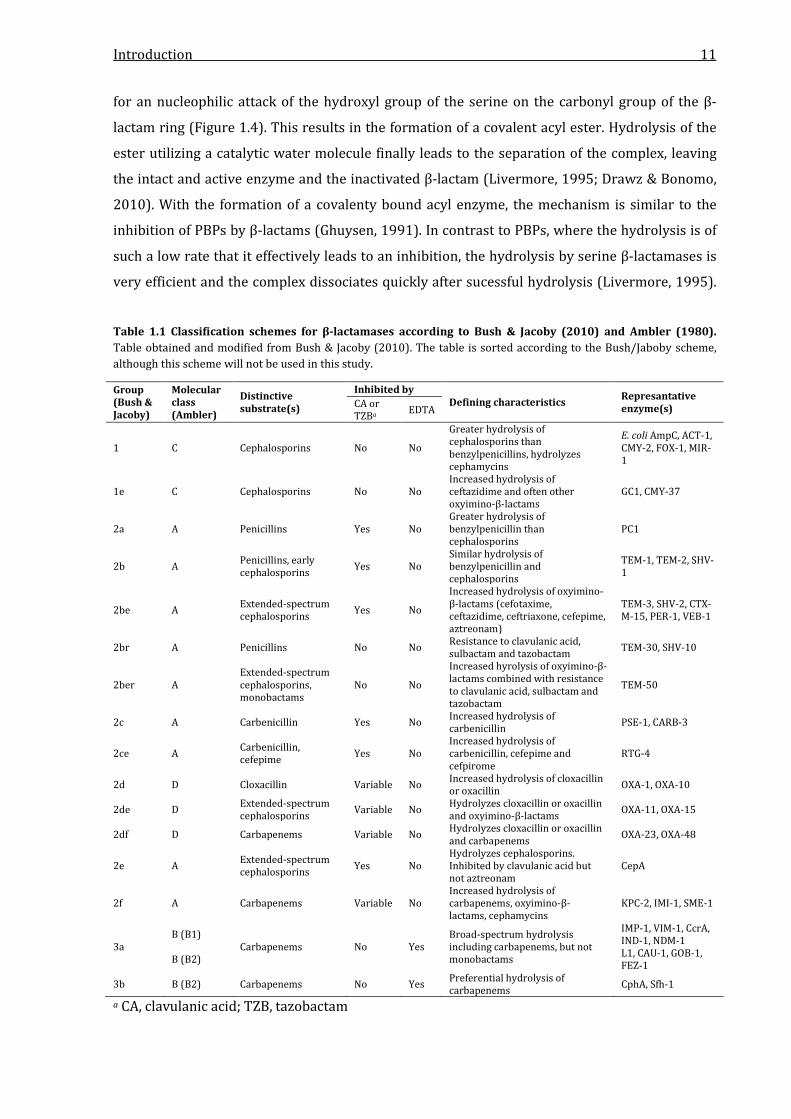
very efficient and the complex dissociates quickly after sucessful hydrolysis (Livermore, 1995).
Table 1.1 Classification schemes for β-lactamases according to Bush & Jacoby (2010) and Ambler (1980). Table obtained and modified from Bush & Jacoby (2010). The table is sorted according to the Bush/Jaboby scheme, although this scheme will not be used in this study.
Group (Bush & Jacoby)
Molecular class (Ambler)
Distinctive substrate(s)
Inhibited by Defining characteristics Represantative
enzyme(s) CA or TZBa EDTA
1 C Cephalosporins No No
Greater hydrolysis of cephalosporins than benzylpenicillins, hydrolyzes cephamycins
E. coli AmpC, ACT-1, CMY-2, FOX-1, MIR-1
1e C Cephalosporins No No Increased hydrolysis of ceftazidime and often other oxyimino-β-lactams
GC1, CMY-37
2a A Penicillins Yes No Greater hydrolysis of benzylpenicillin than cephalosporins
PC1
2b A Penicillins, early cephalosporins Yes No
Similar hydrolysis of benzylpenicillin and cephalosporins
TEM-1, TEM-2, SHV-1
2be A Extended-spectrum cephalosporins Yes No
Increased hydrolysis of oxyimino-β-lactams (cefotaxime, ceftazidime, ceftriaxone, cefepime, aztreonam)
TEM-3, SHV-2, CTX-M-15, PER-1, VEB-1
2br A Penicillins No No Resistance to clavulanic acid, sulbactam and tazobactam TEM-30, SHV-10
2ber A Extended-spectrum cephalosporins, monobactams
No No
Increased hyrolysis of oxyimino-β-lactams combined with resistance to clavulanic acid, sulbactam and tazobactam
TEM-50
2c A Carbenicillin Yes No Increased hydrolysis of carbenicillin PSE-1, CARB-3
2ce A Carbenicillin, cefepime Yes No
Increased hydrolysis of carbenicillin, cefepime and cefpirome
RTG-4
2d D Cloxacillin Variable No Increased hydrolysis of cloxacillin or oxacillin OXA-1, OXA-10
2de D Extended-spectrum cephalosporins Variable No Hydrolyzes cloxacillin or oxacillin
and oxyimino-β-lactams OXA-11, OXA-15
2df D Carbapenems Variable No Hydrolyzes cloxacillin or oxacillin and carbapenems OXA-23, OXA-48
2e A Extended-spectrum cephalosporins Yes No
Hydrolyzes cephalosporins. Inhibited by clavulanic acid but not aztreonam
CepA
2f A Carbapenems Variable No Increased hydrolysis of carbapenems, oxyimino-β-lactams, cephamycins
KPC-2, IMI-1, SME-1
3a B (B1) B (B2)
Carbapenems No Yes Broad-spectrum hydrolysis including carbapenems, but not monobactams
IMP-1, VIM-1, CcrA, IND-1, NDM-1 L1, CAU-1, GOB-1, FEZ-1
3b B (B2) Carbapenems No Yes Preferential hydrolysis of carbapenems CphA, Sfh-1
a CA, clavulanic acid; TZB, tazobactam

Introduction 12
Figure 1.4 Action of a serine β-lactamase against carbapenems. After binding, the β-lactam ring is attacked by the free hydroxyl of the enzymes active site serine residue, yielding a covalent azyl ester. Hydrolysis of the ester with the help of a catalytic water molecule finally leads to the dissociation of the complex and the β-lactam antibiotic is irreversibly inactivated. Figure obtained and modified from Wilson et al. (2010). In contrast to serine-β-lactamases, metallo-β-lactamases (MBL) utilize one or two zinc ions that
coordinate a water molecule which is used for the attack on the β-lactams’ amide bond. In
addition, MBLs do not covalently bind to the β-lactam (Drawz & Bonomo, 2010).
1.4.1 Class A β-lactamases
TEM-1, the first class A serine β-lactamase, was identified in 1965. It was the first plasmid-
mediated β-lactamase described and nowadays, TEM-type enzymes, together with SHV-type β-
lactamases are frequently found in Gram-negative clinical isolates (Drawz & Bonomo, 2010). In
the early 1980s, shortly after the introduction of extended-spectrum cephalosporins cefotaxime
and ceftazidime, the first class A ESBLs were identified that conferred resistance against these
antibiotics (Drawz & Bonomo, 2010). Today, enzymes of the CTX-M type are the most important
class A ESBLs, as the encoding genes are often located on highly transmissible plasmids that
spread into a wide range of Gram-negative pathogens (Bonnet, 2004; Drawz & Bonomo, 2010).
Although these enzymes are able to hydrolyze penicillins, narrow- and extended-spectrum
cephalosporins and aztreonam, they are inhibited by the commercially available β-lactamase
inhibitors sulbactam, tazobactam and clavulanic acid (Drawz & Bonomo, 2010). In contrast, the
class A carbapenemases are able to hydrolyze all β-lactams, including carbapenems and
monobactams, but are still inhibited by the mentioned substances (Bonnet, 2004). The most
important class A carbapenemases are NMC/IMI, SME and KPC-type enzymes and certain GES
variants (Diene & Rolain, 2014). All enzymes of this class share a highly conserved STKF motif at
the amino acid positions 70 to 73 according to the class A β-lactamase standard numbering
scheme with the Ser70 residue beeing the active site serine that covalently binds the β-lactam
ring (Ambler et al., 1991). Although most class A β-lactamase genes are found on plasmids,
several chromosomally located or integron-bourne class A genes (e.g. GES-1) have been
described (Drawz & Bonomo, 2010).

Introduction 13
1.4.2 Class B β-lactamases
Class B β-lactamases, or metallo-β-lactamases (MBLs), differ substantially from the other
classes. Instead of an active site serine the hydrolysis mechanism uses one or two zinc ions that
are coordinated in the active site of the enzyme (Gupta, 2008a). By coordination of a water
molecule by the zinc ions and the use of the -OH group of the water the enzyme performs the
hydrolytic attack on the amide bond of the β-lactam substrate, resulting in an opening of the ring
(Drawz & Bonomo, 2010). Because of their unique hydrolysis mechanism, MBLs are not
inhibited by clinically available inhibitors like sulbactam, clavulanic acid or tazobactam. In vitro,
MBLs can be inhibited by EDTA, which chelates the zinc ions that are necessary for hydrolysis,
making them unavailable to the β-lactamase (Drawz & Bonomo, 2010). In contrast to the class A,
C and D enzymes that belong to the acyltransferases of the SxxK superfamily, MBLs belong to
their own superfamily, also including enzymes with non-β-lactamase functions (Cornaglia et al.,
2011).
The substrate spectrum of MBLs differs between the numerous enzyme variants. For example,
the CphA metallo-β-lactamase of Aeromonas hydrophila has a rather narrow substrate spectrum
while extended range enzymes like the VIM- or IMP-type MBLs are able to hydrolyze all β-
lactams, including carbapenems, but sparing monobactams (Cornaglia et al., 2011). MBLs are
subcategorized into three subclasses. The B1 subclass enzymes require at least one zinc ion in
their active site to be fully active. The most clinically relevant members of this subclass are the
VIM, IMP and NDM enzymes (Nordmann & Poirel, 2014). The B2 enzymes, for example CphA,
require only a single zinc ion and are even inhibidted by a second one, while the B3 MBLs
essentially require two zinc ions, for example the L1 MBL from Stenotrophomonas maltophilia
(Cornaglia et al., 2011). L1 and other dicationic enzymes coordinate the β-lactam by the
carboxylate and carbonyl groups. After binding, the carbonyl is polarized by one of the zinc ions
and attacked by the -OH group of a water molecule. This leads to an anionic state of the nitrogen
in the β-lactam, which is than protonated, leaving the opened β-lactam ring. The source of this
proton is still unknown. For B2 enzymes it is proposed that the water molecule is not
coordinated by the single zinc ion, but by the enzyme residues His118 or Asp120 and that the
zinc ion is responsible for coordination of the β-lactam nitrogen (Drawz & Bonomo, 2010). The
zinc binding ligands are highly conserved between the members of each subclass. Among the
most clinical relevant subclass B1 enzymes, the first zinc ion is bound by the amino acid residues
His116, His118 and His196, while the second one binds to the residues Asp120, Cys221 and
His263, following the Class B β-lactamases standard numbering scheme (Garau et al., 2004).
MBL encoding genes can be chromosomally located (e.g. L1 from S. maltophilia) or plasmid-
bourne like blaVIM or blaNDM and are often found within integron structures (Cornaglia et al.,
2011).

Introduction 14
1.4.3 Class C β-lactamases
The class C β-lactamases, or AmpC enzymes, are serine-β-lactamases. In 1940, the E. coli AmpC
was the first enzyme reported to inactivate penicillin (Abraham & Chain, 1940). The most AmpC
enconding genes are located on the bacterial chromosome, but plasmid-bourne AmpC enzymes
are becoming more prevalent (Drawz & Bonomo, 2010). AmpC genes can be found in many
Enterobacteriaceae like Enterobacter spp., Citrobacter freundii or E. coli and in P. aeruginosa or
A. baumannii, while Klebsiella spp., Salmonella spp. and Proteus spp. normally do not harbour
chromosomal AmpC encoding genes (Jacoby, 2009). In most cases, the expression level of blaAmpC
genes is rather low, but in some species can be induced by exposure to certain β-lactams,
especially cefoxitin and imipenem (Bennett & Chopra, 1993; Babic et al., 2006). The induction
mechanism is based on the conformational change of the transcriptional regulator AmpR that is
induced by binding of cell wall fragments that are formed under β-lactam treatment. This has an
important clinical impact, as strains susceptible to β-lactams can become resistant during
therapy (Jacoby, 2009; Drawz & Bonomo, 2010). In addition, AmpCs are sometimes
overexpressed in clinical isolates, resulting from mutations in the ampD or ampC genes that lead
to hyperinducibility or to constitutive expression (Jacoby, 2009). Although carbapenems are
hydrolyzed with only weak activity, an AmpC overexpression combined with a porin loss and
efflux systems can lead to increased carbapenem resistance in clinical P. aeruginosa isolates
(Jacoby, 2009). Examples for AmpC enzymes are CMY-2, ACT-1, DHA-1 and the E. coli AmpC
(Bush & Jacoby, 2010).
1.4.4 Class D β-lactamases
With currently over 450 variants assigned, class D serine β-lactamases are one of the largest
group of β-lactam hydrolyzing enzymes. They are also known as OXA-type enzymes, named after
their initial characteristic: the ability to hydrolyze oxacillin with higher efficiencies than class A
β-lactamases (Drawz & Bonomo, 2010). They display very low levels of homology to Class A and
C β-lactamases (Massova & Mobashery, 1998) and are a very heterogenous group of enzymes
that is found in a wide variety of Gram-negative bacteria with clinical importance. They were
mostly identified in P. aeruginosa, E. coli, P. mirabilis and A. baumannii isolates (Leonard et al.,
2013). OXA-type β-lactamase genes are characterized as highly mobile, as most of them have
been found on plasmids, in transposons or within mobile integrons (Poirel et al., 2010). Contrary
to mobile blaOXA genes, it was found that every A. baumannii strain intrinsically harbours the
chromosomally encoded OXA-51 β-lactamase (Evans & Amyes, 2014).
While many OXA-type enzymes are described as narrow-spectrum β-lactamases or ESBLs (e.g.
OXA-2, OXA-10 and OXA-20), the class also harbours carbapenemases that are known as
carbapenem-hydrolyzing class D β-lactamases (CHDLs) with OXA-48 beeing the most prominent
and clinically relevant one (Poirel et al., 2010; Leonard et al., 2013). OXA β-lactamases can

Introduction 15
significantly differ from each other with homologies of only 30 % and the enzymes are
subgrouped, for example into the OXA-2, OXA-10 and OXA-23-like enzymes (Evans & Amyes,
2014). Despite their great difference, OXA enzymes share several highly conserved regions, with
one of them beeing the region around the serine amino acid residue at position 70, relative to
the class D β-lactamase numbering system (De Luca et al., 2011). This residue is part of the
STFK motif (positions 70 to 73) and is the active site serine that covalently binds the β-lactam
substrate. The two other highly conserved regions are the YGN motif at the positions 144 to 146
and the KTG motif at the positions 216 to 218. These motifs are found in almost all OXA enzymes
(Poirel et al., 2010).
1.5 Carbapenemases and their distribution
As previously described, carbapenemases are found in the molecular classes A, B and D.
Although these enzymes differ in their hydrolytic efficiency against various β-lactam substrates,
they are often conferring high level resistance to carbapenems in clinical Gram-negative isolates
(Queenan & Bush, 2007). The carbapenemases of the Ambler class A are the IMI/NMC, SME, KPC
and GES-type enzymes (Diene & Rolain, 2014). GES-1 has been described as an ESBL, but novel
variants of this enzyme like GES-2 or GES-5 have been found that exhibited significant
carbapenem hydrolysis (Nordmann et al., 2012). SME, IMI and NMC enzymes are usually
chromosomally encoded, whereas GES and KPC enzymes are plasmid-encoded (Diene & Rolain,
2014). The currently clinically most relevant class A carbapenemase is KPC-2, which was
originally identified in a K. pneumoniae isolate in the U.S. in 1996 but is nowadays found in many
Gram-negative species and has spread globally within a few years (Nordmann & Poirel, 2014).
All class B metallo-β-lactamases are classified as carbapenemases. While MBLs are intrinsic for
many environmental and opportunistic bacterial species, several acquired mobile MBLs have
been identified since the early 1990s (Walsh et al., 2005). They were mostly found in clinical
P. aeruginosa strains or in Enterobacteriaceae (Nordmann et al., 2012). The most common MBLs
belong to the IMP, VIM and NDM type, but also other types have been described that are found
less frequent, for example GIM, KHM, FIM and SIM (Queenan & Bush, 2007; Sekiguchi et al.,
2008; Pollini et al., 2013; Diene & Rolain, 2014; Nordmann & Poirel, 2014). MBL genes can be
located on conjugable plasmids or mobile transposons and are distributed worldwide with
several regional accumulations (Diene & Rolain, 2014). In many cases, MBL genes are found
within integron structures or as part of larger transposons (Walsh et al., 2005; Cornaglia et al.,
2011). Currently, VIM-2 is the most reported MBL wordwide and is mostly found in southern
Europe (Greece, Spain and Italy) and in South Korea and Taiwan (Nordmann & Poirel, 2014).
NDM enzymes on the other hand are mostly found on the Indian subcontinent (India, Pakistan
and Sri Lanka) but have also rapidly spread worldwide since their first description in 2009 and
NDM-1 is currently one of the most clinically relevant carbapenemases (Yong et al., 2009;

Introduction 16
Nordmann & Poirel, 2014). The third important group of MBLs are the IMP-type enzymes.
IMP-type MBLs were the first acquired MBLs to be identified in 1991 and have spread into many
Gram-negative species with clinical importance since then (Cornaglia et al., 2011). So far, 50 IMP
variants have been assigned (http://www.lahey.org/studies) and these enzymes have spread
worldwide, mostly in P. aeruginosa and A. baumannii strains (Nordmann & Poirel, 2014).
Class D carbapenemases, or CHDLs, can be plasmid- or chromosomally encoded (Diene & Rolain,
2014). The clinically most relevant OXA carbapenemase is the plasmid-encoded OXA-48, which
has been primarily found in Enterobacteriaceae. It was first described in K. pneumoniae in 2003
and has spread widely since. OXA-48 is mainly found in Turkey and most other countries of the
Mediterranean area, but is also frequently found in nearly all European countries and Northern
Africa (Diene & Rolain, 2014; Nordmann & Poirel, 2014). Other important OXA-type
carbapenemases are the OXA-23-, OXA-24- and OXA-58-like enzymes which are found
worldwide and mainly in A. baumannii isolates (Walsh, 2010). They can be chromosome- or
plasmid-encoded (Evans & Amyes, 2014).
1.6 Mobility of β-lactamase genes
β-lactamase genes or resistance genes in general can be transferred between bacteria with
various mechanisms. The two clinically most important mechanisms that mediate this horizontal
gene transfer are conjugative transposable elements and conjugable plasmids (Diene & Rolain,
2014). Conjugable transposable elements are genetic structures that encode all functions
necessary for their own intercellular transfer and are subgrouped into conjugable transposons
(Tn) and insertion sequences (IS) (Siguier et al., 2014).
Insertion sequences
Insertion sequences (IS) are relatively small DNA structures (0.7 to 2.5 kb) that carry one or two
open reading frames (ORFs) that encode for transposases. Transposases are multifunctional
enzymes that catalyze the excision and the transfer of DNA sequences (Siguier et al., 2014). IS
are bordered by short terminal inverted repeat sequences that function as recognition sites for
the transposase (Darmon & Leach, 2014). ISs can jump into the chromsome as well as into
plasmids (Siguier et al., 2014). Although a classical IS does not harbour additional genes, many IS
families are more complex and can carry passenger genes that encode for regulatory proteins,
methyltransferases or antibiotic resistance (Figure 1.5). They are known as transporter ISs
(Siguier et al., 2014). IS elements have frequently been reported as carriers for β-lactamase
genes. For example, the blaOXA-48 gene is almost always flanked by one or two copies of the
insertion sequence IS1999 (Evans & Amyes, 2014).

Introduction 17
Figure 1.5 Schematic organization of transporter insertion sequences and transposons. (A) Organization of a typical transporter IS. The IS is flanked by two short inverted repeat regions (IRL and IRR) that encompass one or two transposase encoding genes and one or more passenger genes. When the IS is inserted, a short sequence of the target DNA is often duplicated, resulting in direct repeats (DR) that encompass the IS. (B) Organization of a typical transposon. The transposon is flanked by larger inverted repeat regions and carries multiple genes that are responsible for transposition. It can also carry additional accessory genes that can be resistance genes or other genes conferring a phenotypical advantage to the host cell. Figure obtained and modified from Darmon & Leach (2014).
Transposons
Transposons are large DNA structures with sizes ranging from 2.5 to 60 kb (Darmon & Leach,
2014) and encode for site-specific DNA recombinases that function as integrases, resolvases and
invertases (Burrus et al., 2002). These enzymes catalyze the integration and excision of DNA, the
resolution of co-integrates and the inversion of DNA fragments (Darmon & Leach, 2014). Like
ISs, transposons can be integrated into the chromosome or into plasmids (Darmon & Leach,
2014). Transposons usually possess long terminal inverted repeats and often harbour accessory
genes that confer an phenotypic advantage to their host, for example antibiotic or heavy metal
resistance genes (Darmon & Leach, 2014). The structure of a typical transposon is illustrated in
Figure 1.5. Complex conjugative transposons are called composite transposons, that possess ISs
at both ends and can excise themselves for conjugation to another cell (Darmon & Leach, 2014).
A large number of β-lactamase gene carrying transposons have been described, for example
Tn4401 that carries the blaKPC-2 gene (Cuzon et al., 2010). Tn2006 in A. baumannii is carrying the
blaOXA-23 gene and is almost allways a composite transposon that is bracketed on both sides by
the insertion sequence ISAbaI (Diene & Rolain, 2014). ISAba1 has also been reported as a carrier
for blaOXA-51-like, blaOXA-58-like and blaOXA-235-like genes (Evans & Amyes, 2014).
Plasmids
Self-transmissible conjugative plasmids are large DNA molecules that encode the proteins
involved in their own transfer from a donor cell to a recipient cell via conjugation. They exist
separately from the bacterial chromsome and are replicated independently from it, although the
replication infrastructure is mainly provided by the host cell (Bennett, 2008). The size of
plasmids ranges between a few thousand to hundreds of thousands of base pairs and in most
cases, they are circular molecules, although linear plasmids exist, for example in

Introduction 18
Streptomyces spp. or Borellia burgdorferi (Snyder & Champness, 2007). All conjugable plasmids
exhibit two important regions, the oriV and oriT regions. The oriV (V for vegetative) region is the
origin of replication and is the main determinant for the plasmid host range and the copy
number regulation, although conjugative plasmids are mostly single copy molecules (Snyder &
Champness, 2007). Another important function of the oriV is determination of the
incompatibility type, which is a regulative mechanism that determines the stable coexistance of
two or more plasmids in one cell. If two plasmids cannot coexist stably in the cell, they share the
same incompatibility (Inc) type (Snyder & Champness, 2007). The oriT (T for transfer) is the
origin of the rolling-circle replication during conjugation (Snyder & Champness, 2007). The
genes necessary for transfer are the tra genes, which occur in various combinations and are
correlated to the plasmids Inc-type (Snyder & Champness, 2007). Usually, plasmids carry genes
that confer a growth advantage for the host cell. These can be resistance determinants and since
the first detection of antibiotic resistance, plasmids have been the major distributives of
antimicrobial resistance genes (Bennett, 2008). Many important carbapenemase genes are
plasmid-mediated, for example OXA-48, NDM-1, KPC-2, and VIM-1 and in many cases, the genes
are part of integrons (Smith Moland et al., 2003; Poirel et al., 2004b; Loli et al., 2006; Johnson &
Woodford, 2013).
Integrons
Integrons are genetic structures that efficiently capture and express genes. They are often part
of larger insertion sequences or transposons and thus can be mobilized (Mazel, 2006). The
structure of integrons is characterized by several core features. The first feature is the intI gene
encoding for an integrase, which catalyzes the recombination between incoming gene cassettes
and the second core feature, the attI site. This site is an integron-associated recombination site.
The third core feature is the expression of captured genes by one or two integron-associated
promoters (Gillings, 2014). Novel genes are acquired by insertion of circular gene cassettes,
which usually consist of a single ORF and the attC element (Hall et al., 1991). The gene is
inserted by site-specific recombination between the attI and attC sites and this process is
catalyzed by the integrase (Gillings, 2014). While integrons were classified into five groups at
first, it is nowadays known that hundreds of different integron classes exist, based on their
respective intI sequences (Boucher et al., 2007). The most clinically relevant classes of integrons
are the classes 1, 2 and 3, which are all linked to insertion sequences and transposons,
conferring a mobility of the integrated gene cassettes. The most frequently found integrons that
are associated with antibiotic resistance genes are the class 1 integrons (Mazel, 2006). The
structure of a typical class 1 integron is shown in Figure 1.6. Class 1 integrons consist of two
highly conserved regions, the 5´CS region, which includes the intI1 gene and the attI site and the

Introduction 19
Figure 1.6 Schematic structure of a class 1 integron. The conserved integron regions consist of the integrase-encoding intI1 gene and the qacEΔ1/sul1 open reading frame. Resistance gene cassettes (gene + attC site) can be acquired and inserted at the attI site and are expressed under the control of the promoters Pc and P2. 3´CS region, including the partially deleted gene qacEΔ1 and the sul1 gene that confer resistance
against quarternary ammonium compounds and sulfonamides (Mazel, 2006). They possess
three promoter structures that are Pint, Pc and P2 (Collis & Hall, 1995). Pint is the promoter of the
intI1 gene, while Pc and P2 control the expression of the integrated genes cassettes and can occur
in several variations, resulting in different expression levels (Papagiannitsis et al., 2009). Class 1
integrons have been described as carriers of blaIMP, blaVIM, blaOXA and aac-type genes and play an
important role for the dissemination of antibiotic resistance (Walsh et al., 2005; Voulgari et al.,
2013).
1.7 Pseudomonas aeruginosa
P. aeruginosa is a Gram-negative opportunistic pathogen that can cause a wide range of severe
nosocomial infections. It normally inhabits the soil and surface in aqueous environments and
exhibits several intrinsic antibiotic resistance determinants, such as low permeability,
expression of efflux systems and an inducible AmpC β-lactamase (Gellatly & Hancock, 2013).
P. aeruginosa is one of the most common pathogens causing respiratory infections in
hospitalized patients. In almost all cases infections occur only in patients with poor health status
and as most clinical P. aeruginosa strains carry multiple resistance genes in addition to their
intrinsic resistance mechanisms isolates are often multidrug resistant. Consequently, morbidity
and mortality based on P. aeruginosa infections are rather high (Gellatly & Hancock, 2013).
Apart from pneumonia, the bacterium is also capable of infecting the urinary tract and soft tissue
(e.g. after burns) and can cause bacteremia, keratitis and other infections (Gellatly & Hancock,
2013). P. aeruginosa strains have frequently been described as carriers of carbapenemases, with
the majority beeing MBLs of the IMP, VIM, NDM and GIM families (Diene & Rolain, 2014).
1.8 Citrobacter freundii
Like P. aeruginosa, C. freundii is a Gram-negative opportunistic pathogen and can cause severe
nosocomial infections in neonates or immunocompromised adults or older children (Doran,
1999). As a member of the Enterobacteriaceae, C. freundii belongs to the resident commensal
flora of the human gastrointestinal tract, although it is assumed that Citrobacter species have a
wide environmental distribution (Janda & Abbot, 2006). It can cause infections of the central

Introduction 20
nervous system, bacteremia and urinary tract infections. Rarely, C. freundii strains are described
in the context of wound infections, respiratory tract infections and gastroenteritis cases (Janda &
Abbot, 2006). Expression of β-lactamases is common in Citrobacter species and C. freundii
always harbours a chromosomally-encoded ampC gene. It has also been described as a carrier
for several plasmid-encoded ESBLs like CTX-M-3 or carbapenemases of the KPC, OXA-48 and
IMP type (Janda & Abbot, 2006; Diene & Rolain, 2014).
1.9 Objectives of this work
At the German National Reference Laboratory for multidrug-resistant Gram-negative bacteria at
the Ruhr-University Bochum, Gram-negative clinical isolates with increased carbapenem
resistance are analyzed for the molecular basis of resistance. The resistance is analyzed with
phenotypic and genetic methods, including several antibiotic disk-based tests and a PCR
screening on the most common carbapenemase genes, but also on genes that are less frequent.
In some clinical isolates, the cause of resistance can only be identified phenotypically and there
is a chance that these isolates harbour novel resistance genes or variants of existing ones. As it
has been shown that already one single amino acid substitution can significantly change the
hydrolysis characteristics of a β-lactamase, the identification and characterization of these
enzymes is important for both clinical diagnostics and antimicrobial therapy.
In this study, three clinical isolates were analyzed on the molecular basis of carbapenem
resistance: P. aeruginosa NRZ-00156, C. freundii NRZ-02127 and P. aeruginosa NRZ-03096.
P. aeruginosa NRZ-00156 was isolated in 2008 from an ingunial swab from a patient
hospitalized in Western Germany and showed high carbapenem resistance and a clear
carbapenemase phenotype. This phenotype was inhibited by EDTA, indicating the potential
production of a metallo-β-lactamase. However, all diagnostic PCRs for MBL genes were negative
and it was suspected that this isolate harboured a novel MBL gene.
C. freundii NRZ-02127 was isolated in 2011 from tracheal aspirate from a patient hospitalized in
Southern Germany and showed elevated carbapenem resistance but was susceptible to
oxyimino-cephalosporins. It also showed a carbapenemase phenotype that was inhibited by
clavulanic acid. It was suspected that this isolate harboured a class D β-lactamase, although the
resistance to carbapenems and the inhibition by clavulanic acid did not match to any described
OXA-type enzyme and diagnostic PCRs for blaOXA-48-like genes were negative.
P. aeruginosa NRZ-03096 was isolated in 2012 from an anal swab from a patient hospitalized in
Northern Germany and also showed high carbapenem resistance and an MBL phenotype, as the
resistance was inhibited by EDTA. Like in P. aeruginosa NRZ-00156, diagnostic PCRs covering all
common MBL genes were negative and it was suspected that this isolate produced a novel
carbapenemase.

Introduction 21
The main objectives that were adressed in this work are: i) the search for novel
carbapenemases; ii) the characterization of the genetic environment of the carbapenemase
genes and their localization; iii) the phenotypic characterization of the enzymes and their impact
on resistance in vivo and iv) the biochemical characterization of the novel enzymes.
The identification of novel carbapenemases was adressed by PCR and shotgun cloning
approaches and phenotypic characterization by resistance analyses in isogenic E. coli strains.
The characterization of the genetic environment was adressed by sequencing and in silico DNA
sequence analysis techniques, while the localization of the genes was analyzed by Southern
blotting experiments and 454-sequencing of plasmids.
The biochemical characterization was adressed by overexpression and purification of the native
unmodified novel enzymes and by obtaining the kinetic parameters Km and kcat for the most
important β-lactam substrates with in vitro hydrolysis assays.
These experiments were performed to make a statement on the capabilities of novel
carbapenemases, their ongoing diversification, their ability to spread and their potential
significance for future carbapenem resistance developments in Gram-negative bacteria of
clinical importance.

Material and Methods 22
2 Material and Methods
All instruments or materials that were used in this study but not listed here were standard lab
equipment. All solutions and lab materials for bacterial cell cultures and molecular techniques
were autoclaved or filtered sterile before use. All buffers and media were prepared in A. dest
except noted otherwise.
2.1 Material
2.1.1 Instruments
Cell disruptors:
PowerLyzer 24 MO BIO
Sonifier W-250 D Branson
Chromatography instruments:
ÄKTA Pure 25 L FPLC system GE Healthcare
HiPrep 16/60 S-200 HR GF column GE Healthcare
HiPrep 26/10 desalting column GE Healthcare
HiTrap SP HP 5 ml IEX column GE Healthcare
Centrifuges & rotors:
Centrifuge 3K12, rotor #11223 Sigma-Aldrich
Biofuge Pico, rotor #3325 Thermo Scientific-Heraeus
Megafuge 1.0R, rotor #3360 Heraeus
Fresco 21, rotor #75003424 Thermo Scientific-Heraeus
J2-HS, rotors JA-14 & JA-20 Beckmann
Electrophoresis instruments:
CHEF DR III PFGE chamber Bio-Rad
DNA gel electrophoresis chamber Renner
Multiphor II IEF chamber GE Healthcare
SDS -PAGE chamber P8DS Thermo Scientific / Owl Scientific
GelDoc XR+ gel documentation system Bio-Rad
HyperCassette exposure cassette Amersham
Incubators:
Incubator BB 16 Heraeus Instruments
Incubator shaker 3033 GFL
innova 4330 New Brunswick Scientific
PureLab Classic water destiller Elga

Material and Methods 23
Next-generation sequencing instruments:
DNA nebulizer Roche
DynaMag magnetic particle concentrator Invitrogen
GS Junior 454 sequencing system Roche
GS Junior Bead Counter Roche
GS Junior Titanium PicoTiterPlater Roche
PCR cyclers:
Labcycler basic Sensoquest
Labcycler gradient Sensoquest
Mastercycler personal Eppendorf
Mastercycler epgradient Eppendorf
pH-Meter 761 Calimatic Knick
Photometers and accessories:
BioPhotometer Eppendorf
BioSpectrometer kinetic Eppendorf
Eppendorf µCuvette G1.0 Eppendorf
Power supply:
Gene Power Supply 200/400 Pharmacia
Power Pac 300 Power Supply BioRad
Qubit 2.0 fluorometer Life Technologies
Thermomixers:
ThermoMixer C Eppendorf
TS1 Thermoshaker Biometra
Vacu-Blot Southern blotting chamber Biometra
VITEK Densichek densitometer bioMérieux
2.1.2 Disposable material
CleanGel IEF GE Healthcare
Sterile cotton tips Heinz Herenz Hamburg
CryoPure cryotubes Sarstedt
Disposable PFGE Plug Mold Bio-Rad
Filtropur S 0.2 sterile filter Sarstedt
Fitropur S 0.45 sterile filter Sarstedt
Gel blot paper Schleicher & Schüll
Glass beads 0.1 mM Scientific Industries
HyperFilm MP autoradiography film GE Healthcare
MH2 Mueller-Hinton agar plates bioMérieux

Material and Methods 24
Nylon membrane, positively charged Roche
UVette cuvettes Eppendorf
UV-transparent cuvettes Sarstedt
2.1.3 Chemicals
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid
(HEPES) AppliChem
β-Mercaptoethanol AppliChem
Acetic acid p.a. VWR
Acrylamide (30%) Rotiphorese Gel 30 Roth
AEBSF-hydrochloride AppliChem
Ammonium persulfate (APS) Roth
Bovine serum albumine (BSA) Fraktion V AppliChem
Brilliant Blue R-250 Roth
Bromophenole blue Merck
Calcium chloride J.T. Baker
Clavulanic acid SmithKline-Beecham
Chloro-5-substituted adamantyl-1,2-
dioxetane phosphate (CSPD) Roche
Defibrinated sheep blood Thermo Scientific
Dimethyl sulfoxide (DMSO) Biomol
DNA-Polymerasepuffer GE Healthcare
Deoxyribose nucleoside triphosphates (dNTPs) Applied Biosystems,
Thermo Scientific
Dipotassium phosphate Merck
Disodium phosphate J.T. Baker
Ethanol p.a. Sigma-Aldrich
Ethylenediaminetetraacetic acid (EDTA) Merck
Ethidiumbromide AppliChem
Formamide J.T. Baker
Glycerol J.T. Baker
Hydrogen chloride 36-38% J.T. Baker
InCert agarose Lonza
Isopropyl alcohol J.T. Baker
Magnesium chloride J.T. Baker
Magnesium sulfate J.T. Baker
Maleic acid Merck

Material and Methods 25
Manganese(II) chloride Merck
Methanol p.a. J.T. Baker
Monopotassium phosphate Riedel-de Haën
Monosodium phosphate J.T. Baker
Nitrocefin Calbiochem
Petroleum Roth
Polyoxyethylene (20) cetyl ether (Brij 58) Sigma-Aldrich
Roti-Nanoquant Roth
Rubidium chloride AppliChem
Seakem Gold Agarose Lonza
Sodium dodecyl sulfate AppliChem, Sigma
Sodium hydroxide J.T. Baker
Sodium chloride J.T. Baker
Sodium citrate Merck
Sodium lauroyl sarcosinate Sigma-Aldrich
StarPure agarose StarLab
Sucrose AppliChem
Tetramethylethylenediamine (TEMED) Merck
Tris(hydroxymethyl)aminomethane-HCl (Tris-HCl) AppliChem
Tris(hydroxymethyl)aminomethane (Trizma base) Sigma-Aldrich
Tween20 AppliChem
Zinc sulfate AppliChem
2.1.4 Antibiotics
Penicillin G Molekula
Ampicillin Molekula, AppliChem
Oxacillin Molekula
Piperacillin Molekula
Cefotaxime Molekula, Aventis
Ceftazidime Molekula
Cefoxitin Molekula, Infektiopharm
Imipenem Molekula
Ertapenem Molekula, MSD
Meropenem Molekula, Hexal
Aztreonam Molekula
Kanamycin AppliChem
Chloramphenicol AppliChem

Material and Methods 26
Sodium azide Riedel-de Haën
Rifampicin AppliChem
Streptomycin AppliChem
2.1.5 Wafers containing antibiotics
Susceptibility disks Thermo Scientific Oxoid
Ampicillin 10 µg, Ampicillin/Sulbactam 10/10 µg, Piperacillin 30 µg, Piperacillin/Tazobactam
36 µg, Amoxicillin 10 µg, Amoxicillin/Clavulanic acid 30 µg, Cefotaxim 5 µg, Cefoxitin 30 µg,
Imipenem 10 µg, Meropenem 10 µg, Doripenem 10 µg, Ertapenem 10 µg
2.1.6 Antibiotic gradient test strips
Etest bioMérieux
Ampicillin, Ampicillin/Sulbactam, Piperacillin, Piperacillin/Tazobactam, Amoxicillin,
Amoxicillin/Clavulanic acid, Temocillin, Cephalothine, Cefuroxime, Cefoxitin, Cefotaxime,
Ceftriaxon, Cefepime, Ceftazidime, Imipenem, Meropenem, Doripenem, Ertapenem, Aztreonam,
Gentamicin, Tobramicin, Amikacin, Doxycyclin, Tetracyclin, Minocyclin, Tigecyclin,
Ciprofloxacin, Levofloxacin, Colistin, Nitrofurantoin, Chloramphenicol, Fosfomycin,
Trimethoprim/Sulfamethoxazole, MBL Etest Imipenem, MBL Etest Meropenem
2.1.7 Kits und standards
Kits:
Expand Long Range PCR System Roche
FastStart High Fidelity PCR System Roche
GS Junior Titanium emPCR Kit (Lib-L) Roche
GS Junior Maintenance Wash Kit Roche
GS Junior Titanium PicoTiterPlate Kit Roche
GS Junior Titanium Sequencing Kit Roche
GS Rapid Library Kit Roche
Nucleospin Tissue Kit Macherey-Nagel
Nucleospin Plasmid Kit Macherey-Nagel
Nucleospin Gel and PCR cleanup Kit Macherey-Nagel
NucleoBond PC 100 Kit Macherey-Nagel
PCR DIG Probe Synthesis Kit Roche
Qubit dsDNA HS Assay Kit Life Technologies
Qubit Protein Assay Kit Life Technologies

Material and Methods 27
Universal GenomeWalker 2.0 Kit Clontech
Standards:
CHEF DNA Size Marker #170-3605 Bio-Rad
CHEF DNA Size Marker #170-3667 Bio-Rad
GeneRuler DNA Ladder Mix Thermo Scientific
Lambda-Ladder PFG marker New England Biolabs
Low range PFG marker New England Biolabs
PageRuler Plus Prestained Thermo Scientific
2.1.8 Enzymes
FastDigest DNA restriction endonucleases Thermo Scientific
I-Ceu I restriction endonuclease New England Biolabs
Lysozyme AppliChem
Nuclease S1 restriction endonuclease Thermo Scientific
Pfu proof-reading DNA Polymerase Thermo Scientific
Proteinase K Boehringer Mannheim
Pwo SuperYield DNA Polymerase Roche
RNase A AppliChem
T4 DNA Ligase Thermo Scientific, Roche
Taq DNA Polymerase Roche, Peqlab
All enzyme reactions were performed within the appropriate buffer supplied by the
manufacturer.
2.1.9 Antibodies
Anti-Digoxigenin-AP, Fab fragments Roche

Material and Methods 28
2.2 Microbial strains, plasmids and oligonuclotides
2.2.1 Microbial strains
All microbial strains used in this study are listed in Table 2.1.
Table 2.1. Microbial strains used in this study.
Strain Relevant characteristics Reference / Source
E. coli TOP10 Cloning host, lacI- Invitrogen
E. coli J53 NaN3R Clowes & Rowley (1954)
E. coli C600 + RifR RifR RUB
E. coli ATCC 25922 Control strain for various resistance tests LCG Standards
P. aeruginosa NRZ-00156 Clinical isolate RUB
C. freundii NRZ-02127 Clinical isolate RUB
P. aeruginosa NRZ-03096 Clinical isolate RUB
2.2.2 Plasmids
All plasmids used or constructed in this study are listed in Table 2.2.
Table 2.2. Plasmids used in this study.
Plasmid Relevant characteristics Reference / Source pBK-CMV Cloning & expression vector, KmR Invitrogen
pMB3002 pBK-CMV derivative carrying a gDNA fragment from C. freundii NRZ-02127
Meining (2012)
pMB3006 pBK-CMV derivative carrying the blaOXA-233 gene
Meining (2012)
pMB3007 pBK-CMV derivative carrying the blaIMP-31 gene
This study
pMB3010 pBK-CMV derivative carrying the blaIMP-1 gene
This study
pMB3011 pBK-CMV derivative carrying a gDNA fragment from P. aeruginosa NRZ-00156 including blaIMP-31
This study
pMB3013 pBK-CMV derivative carrying a gDNA fragment from P. aeruginosa NRZ-03096 including blaKHM-2
Hoffmann (2013)
pMB3014 pBK-CMV derivative carrying the blaKHM-2 gene
Hoffmann (2013)
pMB3018 Wildtype plasmid from C. freundii NRZ-02127
This study
pMB3026 pBK-CMV derivative carrying the blaOXA-10 gene
Lange (2014)
pEX-A2-KHM-1 pEX-A2 derivative carrying the synthesized blaKHM-1 gene
This study
pMB3037 pBK-CMV derivative carrying the blaKHM-1 gene
This study
Material and Methods 29
2.2.3 Oligonucleotides
All Oligonucleotides used in this study are listed in Table 2.3.
Table 2.3: Oligonucleotides used in this study. Recognition sites for restriction endonucleases are underlined.
Oligonucleotide Sequence (5´to 3´) Purpose Reference
IMP-X_seq_fw ACG CAG CAG GGC AGT CGC C
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
IMP-X_seq_rev GTT GGG GCA GTC CCG CTT GG
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
IMP-X_seq_up AAT ACT GCC TTT GAT TTT AT
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
IMP-X_seq_dwn GAA AAC TCA TTT AGT GGC GT
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
IMP-X_seq_fw ACG CAG CAG GGC AGT CGC C
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
IMP-X_seq_rev GTT GGG GCA GTC CCG CTT GG
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
IMP-X_seq_up AAT ACT GCC TTT GAT TTT AT
Sequencing of blaIMP-31 from gDNA of P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP1 GAG CTT CTT AAA AAG AAC GGT AAT GCG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP2 GCC GAA AAC TCA TTT AGT GGC GTT AGC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP3 CTC AGC CCC TTA GCT CTG CGT TAG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP2-5 CTT GGG AGC AGG CTG TTA AGG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP4 GGC AAC CAG AAT ATC AGT GGT G
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP5 GTA CCT CGC TGT TGG CCA GGT CGA AAC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP5-1 CCC GTA TTT CAA CAA ATC GCC AG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP6 CAA AGT ACA GCA TCG TGA CCA AC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP7 CCG AGC AAC TTG CGA GCG ATC CG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study

Material and Methods 30
Oligonucleotide Sequence (5´to 3´) Purpose Reference
GW-IMP-31-GSP7-1 GAG CTC TGG TTG AGT TGC TGT TC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP8 GCC GTG TAC ATG GTT CAA ACA CGC C
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP8-1 GCG AGC GAT CCG ATG CTA CGA GAA AG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP9 CAG ATG GTC CAG CCG TGT ACA TG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP9-1 GGT GAA TGC GGG AAA CGT TAA GTG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP10 GAG GCT TTT GAG GAC GCT GAG AAC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP11 GCC CAT ATG GCA CGA TCG TTT CG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31GSP11-1 GCC GCA GAC GCC TCA TAT GTA TAT C
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31GSP11-2 CTG CTC AAG AAA TTC TAC AAG AGC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31GSP12 GCC GTG TAC ATG GTT CAA ACA CG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-GSP13 CAT TGA CGC GGT ATT TGG ACC AG
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31GSP14 CTG GAA ATG TAT CTC AAC CAG C
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31GSP15 CAA GTG AGG GCA TCA TTG GTG GC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-AP3 GAT CTG CGC CAC CTG ATC AAC ACT G
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-AP3-1 GCC TTG CGC ACC TTT ACG AGG ATC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-AP4 CGA ATT GTT AGA CCG CGC TTA GAA G
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-AP5 GAT ACT TCG TCG AGG GCG ACT GTC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
GW-IMP-31-AP6 GTA AAG CTG CTC TCC CTC GGT TC
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
Material and Methods 31
Oligonucleotide Sequence (5´to 3´) Purpose Reference
GW-IMP-31-AP7 CTC GAT GGA AGG GTT AGG CAT C
Sequencing of the blaIMP-31 genetic context from P. aeruginosa NRZ-00156
This study
M13_uni CGA CGT TGT AAA ACG ACG GCC AGT
Sequencing of pBK-CMV based plasmids
New England Biolabs
M13_rev CAG GAA ACA GCT ATG AC Sequencing of pBK-CMV based plasmids
New England Biolabs
OXA-X_seq.fw GGC CGC CCC TCA TGT CAA AC
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_fw1 TAG AAT GGC TCT CCC TTT TC
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_fw.2 ATG TCC TCT GGT AAA CGG GT
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_fw3 CTC ACT GCT TCT GCG CTG TT
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_fw4 ATC TTC AAA GTC CGG CAT CG
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_fw5 TTT ACG AAG TTT CTC ACC GCC
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_rev.1 ACG GCT TCG GCA GAG AAC TC
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_rev.2 GCC ACT CAT AGA GCA TCG CA
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_rev.3 GGG TCA AGG ATC TGG ATT TC
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_rev.4 GTC GGC TTC TGA CGT TCA GT
Sequencing of pMB3002 Meining (2012)
pMB3002_seq_rev5 CCA TTA ATG TTC CGC AAA TC
Sequencing of pMB3002 Meining (2012)
OXA233_Int_rev ACG GCT TCG GCA GAG AAC TC
Sequencing of pMB3002 Meining (2012)
OXA233_Int_fw AGG CGG CAC CTG AAT ATC TAG T
Sequencing of pMB3002 Meining (2012)
Oxa233_Int2_fw2 TGT TCA ATG ATC CCG AGG TC
Sequencing of pMB3002 This study
Oxa233_Int2_rev2 TCA TAG AGC ATC GCA AGG TC
Sequencing of pMB3002 This study
Oxa233_Int1_fw2 CAT TGC AAT GCT GAA TGG AG
Sequencing of pMB3002 This study
Oxa233_Int1_fw3 CGA GGT CAC CAA GAT CCA AA
Sequencing of pMB3002 This study
Oxa233_Int1_rev3 TTC GTG CCT TCA TCC GTT TC
Sequencing of pMB3002 This study
pMB3013_seq_fw1 CGA CAG GTG CCG GCA CAC GCG ATG
Sequencing of pMB3013 Hoffmann (2013)
pMB3013_seq_rv1 CAG AAC AAA CTA ATG AAT TGC
Sequencing of pMB3013 Hoffmann (2013)
pMB3013_seq_fw2 GAT GTG CGC AAC GCA GAA C
Sequencing of pMB3013 Hoffmann (2013)
pMB3013_seq_rv2 CCT GTC TTT GAC AAG CAG ACC
Sequencing of pMB3013 Hoffmann (2013)
pMB3013_seq_rv3 CAC GGC GTC TTG AGC TGA TAC
Sequencing of pMB3013 Hoffmann (2013)
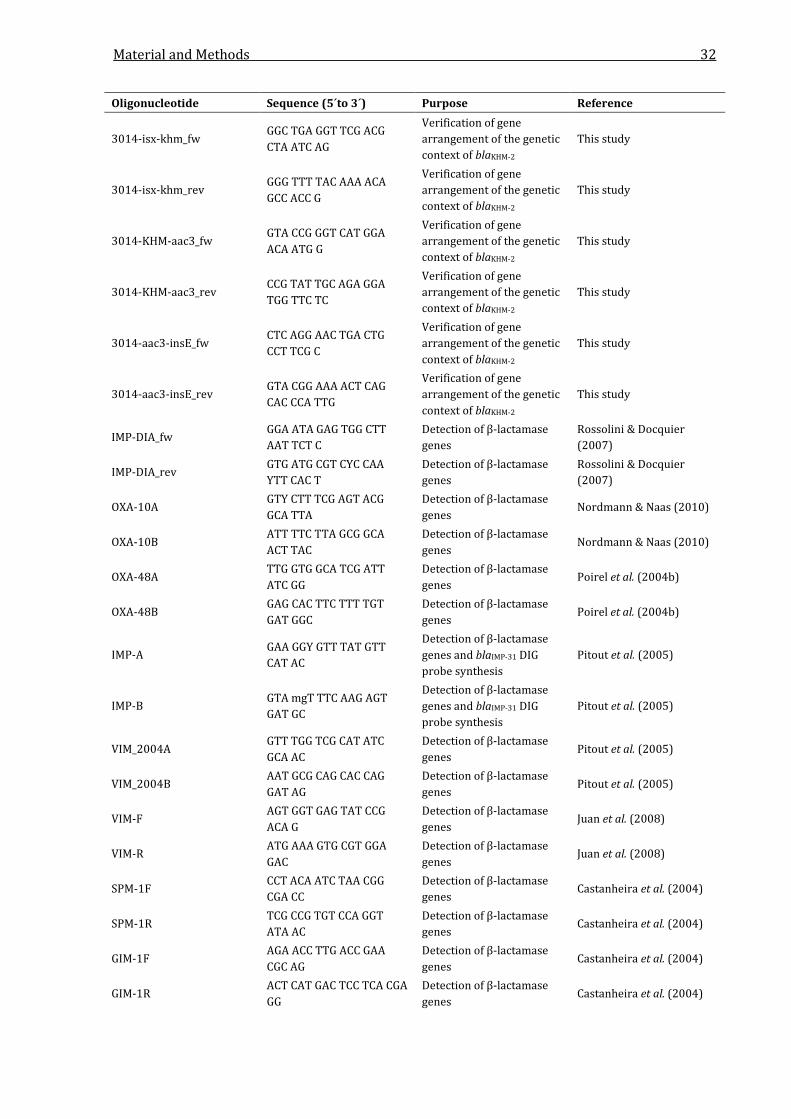
Material and Methods 32
Oligonucleotide Sequence (5´to 3´) Purpose Reference
3014-isx-khm_fw GGC TGA GGT TCG ACG CTA ATC AG
Verification of gene arrangement of the genetic context of blaKHM-2
This study
3014-isx-khm_rev GGG TTT TAC AAA ACA GCC ACC G
Verification of gene arrangement of the genetic context of blaKHM-2
This study
3014-KHM-aac3_fw GTA CCG GGT CAT GGA ACA ATG G
Verification of gene arrangement of the genetic context of blaKHM-2
This study
3014-KHM-aac3_rev CCG TAT TGC AGA GGA TGG TTC TC
Verification of gene arrangement of the genetic context of blaKHM-2
This study
3014-aac3-insE_fw CTC AGG AAC TGA CTG CCT TCG C
Verification of gene arrangement of the genetic context of blaKHM-2
This study
3014-aac3-insE_rev GTA CGG AAA ACT CAG CAC CCA TTG
Verification of gene arrangement of the genetic context of blaKHM-2
This study
IMP-DIA_fw GGA ATA GAG TGG CTT AAT TCT C
Detection of β-lactamase genes
Rossolini & Docquier (2007)
IMP-DIA_rev GTG ATG CGT CYC CAA YTT CAC T
Detection of β-lactamase genes
Rossolini & Docquier (2007)
OXA-10A GTY CTT TCG AGT ACG GCA TTA
Detection of β-lactamase genes
Nordmann & Naas (2010)
OXA-10B ATT TTC TTA GCG GCA ACT TAC
Detection of β-lactamase genes
Nordmann & Naas (2010)
OXA-48A TTG GTG GCA TCG ATT ATC GG
Detection of β-lactamase genes
Poirel et al. (2004b)
OXA-48B GAG CAC TTC TTT TGT GAT GGC
Detection of β-lactamase genes
Poirel et al. (2004b)
IMP-A GAA GGY GTT TAT GTT CAT AC
Detection of β-lactamase genes and blaIMP-31 DIG probe synthesis
Pitout et al. (2005)
IMP-B GTA mgT TTC AAG AGT GAT GC
Detection of β-lactamase genes and blaIMP-31 DIG probe synthesis
Pitout et al. (2005)
VIM_2004A GTT TGG TCG CAT ATC GCA AC
Detection of β-lactamase genes
Pitout et al. (2005)
VIM_2004B AAT GCG CAG CAC CAG GAT AG
Detection of β-lactamase genes
Pitout et al. (2005)
VIM-F AGT GGT GAG TAT CCG ACA G
Detection of β-lactamase genes
Juan et al. (2008)
VIM-R ATG AAA GTG CGT GGA GAC
Detection of β-lactamase genes
Juan et al. (2008)
SPM-1F CCT ACA ATC TAA CGG CGA CC
Detection of β-lactamase genes
Castanheira et al. (2004)
SPM-1R TCG CCG TGT CCA GGT ATA AC
Detection of β-lactamase genes
Castanheira et al. (2004)
GIM-1F AGA ACC TTG ACC GAA CGC AG
Detection of β-lactamase genes
Castanheira et al. (2004)
GIM-1R ACT CAT GAC TCC TCA CGA GG
Detection of β-lactamase genes
Castanheira et al. (2004)

Material and Methods 33
Oligonucleotide Sequence (5´to 3´) Purpose Reference
SIM1-F TAC AAG GGA TTC GGC ATC G
Detection of β-lactamase genes
Lee et al. (2005)
SIM1-R TAA TGG CCT GTT CCC ATG TG
Detection of β-lactamase genes
Lee et al. (2005)
NDM-1_a_fw CAA TAT TAT GCA CCC GGT CG
Detection of β-lactamase genes
Kaase (unpublished)
NDM-1_a_rev CCT TGC TGT CCT TGA TCA GG
Detection of β-lactamase genes
Kaase (unpublished)
DIM-1_a_fw GTA CCT GAG CTA AGA ATC GAG
Detection of β-lactamase genes
Kaase (unpublished)
DIM-1_a_rev_neu CGG CTG GAT TGA TTT GTT AGA G
Detection of β-lactamase genes
Kaase (unpublished)
AIM-1_a_fw GAA ACG TCG CTT CAC CCT G
Detection of β-lactamase genes
Kaase (unpublished)
AIM-1_a_rev ACC AGG ATG TCG CAG TCG AG
Detection of β-lactamase genes
Kaase (unpublished)
KHM-1_a_fw GCT CTT GTT ATA TCG TTT GGT C
Detection of β-lactamase genes
Kaase (unpublished)
KHM-1_a_rev CAT TGT TGC ATT GCT ATA ACG G
Detection of β-lactamase genes
Kaase (unpublished)
NDM-1_b_fw CCT CAA CTG GAT CAA GCA GG
Detection of β-lactamase genes
Kaase (unpublished)
NDM-1_b_rev GAC AAC GCA TTG GCA TAA GTC
Detection of β-lactamase genes
Kaase (unpublished)
FIM-1_F GAA GCA CAT GGA AAA CTG GG
Detection of β-lactamase genes
Pollini et al. (2013)
FIM-1_R GAT GGG CGA ATG AGA CAG C
Detection of β-lactamase genes
Pollini et al. (2013)
IMI-A ATA GCC ATC CTT GTT TAG CTC
Detection of β-lactamase genes
Aubron et al. (2005)
IMI-B TCT GCG ATT ACT TTA TCC TC
Detection of β-lactamase genes
Aubron et al. (2005)
GES-C GTT TTG CAA TGT GCT CAA CG
Detection of β-lactamase genes
Weldhagen & Prinsloo (2004)
GES-D TGC CAT AGC AAT AGG CGT AG
Detection of β-lactamase genes
Weldhagen & Prinsloo (2004)
KPC_5 TGT CAC TGT ATC GCC GTC Detection of β-lactamase genes
Yigit et al. (2001)
KPC_10 CTC AGT GCT CTA CAG AAA ACC
Detection of β-lactamase genes
Yigit et al. (2001)
KPC_fw AAC AAG GAA TAT CGT TGA TG
Detection of β-lactamase genes
Pasteran et al. (2008)
KPC_rev AGA TGA TTT TCA GAG CCT TA
Detection of β-lactamase genes
Pasteran et al. (2008)
KPC_a_fw CTG TAT CGC CGT CTA GTT CTG
Detection of β-lactamase genes
Kaase (unpublished)
KPC_a_rev GTC GTG TTT CCC TTT AGC CA
Detection of β-lactamase genes
Kaase (unpublished)
KPC_b_fw AAT ATC TGA CAA CAG GCA TGA CGG
Detection of β-lactamase genes
Kaase (unpublished)
KPC_b_rev GTT GAC GCC CAA TCC CTC GA
Detection of β-lactamase genes
Kaase (unpublished)

Material and Methods 34
Oligonucleotide Sequence (5´to 3´) Purpose Reference
KPC_c_fw CCG CCG CCA ATT TGT TGC TG
Detection of β-lactamase genes
Kaase (unpublished)
KPC_c_rev TTA CTG CCC GTT GAC GCC CA
Detection of β-lactamase genes
Kaase (unpublished)
OXA-10_end.fw AAA TCC ATT CCC ACC AAA ATC A
Sequencing of integrons or genes correlated with integrons
This study
aadA6_start_rev ATG TCT AAC TTT GTT TTA GGG CGA C
Sequencing of integrons or genes correlated with integrons
This study
aadA6_end_fw GAG CGG AAT GTA GTG CTT ACC TT
Sequencing of integrons or genes correlated with integrons
This study
integron_5CS GGC ATC CAA GCA GCA AG Sequencing of integrons or genes correlated with integrons
Levesque et al. (1995)
integron_3CS AAG CAG ACT TGA CCT GA Sequencing of integrons or genes correlated with integrons
Levesque et al. (1995)
INT-F CTC TCA CTA GTG AGG GGC
Sequencing of integrons or genes correlated with integrons
Juan et al. (2008)
INT-R ATG AAA ACC GCC ACT GCG
Sequencing of integrons or genes correlated with integrons
Juan et al. (2008)
integron_5CS GGC ATC CAA GCA GCA AG Sequencing of integrons or genes correlated with integrons
Falcone et al. (2009)
integron_3CS CTC TCA AGA TTT TAA TGC GGA TG
Sequencing of integrons or genes correlated with integrons
Falcone et al. (2009)
qacE_delta1 GCC AAC TAT TGC GAT AAC
Sequencing of integrons or genes correlated with integrons
Schneider et al. (2008)
qacE-F GAA AGG CTG GCT TTT TCT TG
Sequencing of integrons or genes correlated with integrons
Juan et al. (2008)
qacE-R ATT ATG ACG ACG CCG AGT C
Sequencing of integrons or genes correlated with integrons
Juan et al. (2008)
int1_fw GCC TGT TCG GTT CGT AAG CT
Sequencing of integrons or genes correlated with integrons
Toleman et al. (2005)
QacR_rev CGG ATG TTG CGA TTA CTT CG
Sequencing of integrons or genes correlated with integrons
Toleman et al. (2005)
aadA6_fw GGA GCA GCA ACG ATG TTA CG
Sequencing of integrons or genes correlated with integrons
This study
aadA6_rev TTG CTG CGC TGT ACC AAA TG
Sequencing of integrons or genes correlated with integrons
This study

Material and Methods 35
Oligonucleotide Sequence (5´to 3´) Purpose Reference
sul1_class1_rev CCG ACT TCA GCT TTT GAA GGT TC
Sequencing of integrons or genes correlated with integrons
This study
qacE_class1_rev CAA TTA TGA GCC CCA TAC CTA CAA AG
Sequencing of integrons or genes correlated with integrons
This study
acsA-F_ampl_curran ACC TGG TGT ACG CCT CGC TGA C
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
acsA-R_ampl_curran GAC ATA GAT GCC CTG CCC CTT GAT
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
acsA-F_seq_curran GCC ACA CCT ACA TCG TCT AT
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
acsA-R_seq_curran AGG TTG CCG AGG TTG TCC AC
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
aroE-F_ampl_curran TGG GGC TAT GAC TGG AAA CC
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
aroE-R_ampl_curran TAA CCC GGT TTT GTG ATT CCT ACA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
aroE-F_seq_curran ATG TCA CCG TGC CGT TCA AG
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
aroE-R_seq_curran TGA AGG CAG TCG GTT CCT TG
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
guaA-F_ampl_curran CGG CCT CGA CGT GTG GAT GA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
guaA-R_ampl_curran GAA CGC CTG GCT GGT CTT GTG GTA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
guaA-F_seq_curran AGG TCG GTT CCT CCA AGG TC
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
guaA-R_seq_curran GAC GTT GTG GTG CGA CTT GA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
mutL-F_ampl_curran CCA GAT CGC CGC CGG TGA GGT G
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
mutL-R_ampl_curran CAG GGT GCC ATA GAG GAA GTC
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
mutL-F_seq_curran AGA AGA CCG AGT TCG ACC AT
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
mutL-R_seq_curran GGT GCC ATA GAG GAA GTC AT
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
nuoD-F_ampl_curran ACC GCC ACC CGT ACT G MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
nuoD-R_ampl_curran TCT CGC CCA TCT TGA CCA MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
nuoD-F_seq_curran ACG GCG AGA ACG AGG ACT AC
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
nuoD-R_seq_curran TGG CGG TCG GTG AAG GTG AA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
ppsA-F_ampl_curran GGT CGC TCG GTC AAG GTA GTG G
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
ppsA-R_ampl_curran GGG TTC TCT TCT TCC GGC TCG TAG
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
ppsA-F_seq_curran GGT GAC GAC GGC AAG CTG TA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
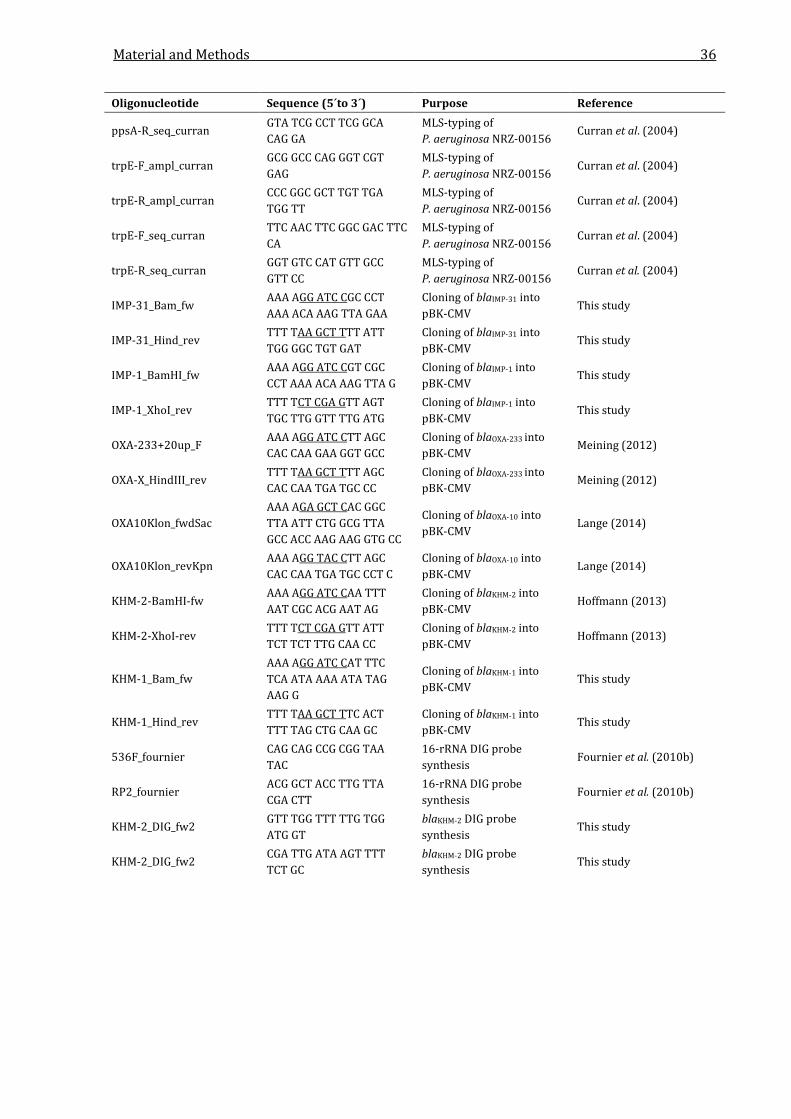
Material and Methods 36
Oligonucleotide Sequence (5´to 3´) Purpose Reference
ppsA-R_seq_curran GTA TCG CCT TCG GCA CAG GA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
trpE-F_ampl_curran GCG GCC CAG GGT CGT GAG
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
trpE-R_ampl_curran CCC GGC GCT TGT TGA TGG TT
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
trpE-F_seq_curran TTC AAC TTC GGC GAC TTC CA
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
trpE-R_seq_curran GGT GTC CAT GTT GCC GTT CC
MLS-typing of P. aeruginosa NRZ-00156
Curran et al. (2004)
IMP-31_Bam_fw AAA AGG ATC CGC CCT AAA ACA AAG TTA GAA
Cloning of blaIMP-31 into pBK-CMV
This study
IMP-31_Hind_rev TTT TAA GCT TTT ATT TGG GGC TGT GAT
Cloning of blaIMP-31 into pBK-CMV
This study
IMP-1_BamHI_fw AAA AGG ATC CGT CGC CCT AAA ACA AAG TTA G
Cloning of blaIMP-1 into pBK-CMV
This study
IMP-1_XhoI_rev TTT TCT CGA GTT AGT TGC TTG GTT TTG ATG
Cloning of blaIMP-1 into pBK-CMV
This study
OXA-233+20up_F AAA AGG ATC CTT AGC CAC CAA GAA GGT GCC
Cloning of blaOXA-233 into pBK-CMV
Meining (2012)
OXA-X_HindIII_rev TTT TAA GCT TTT AGC CAC CAA TGA TGC CC
Cloning of blaOXA-233 into pBK-CMV
Meining (2012)
OXA10Klon_fwdSac AAA AGA GCT CAC GGC TTA ATT CTG GCG TTA GCC ACC AAG AAG GTG CC
Cloning of blaOXA-10 into pBK-CMV
Lange (2014)
OXA10Klon_revKpn AAA AGG TAC CTT AGC CAC CAA TGA TGC CCT C
Cloning of blaOXA-10 into pBK-CMV
Lange (2014)
KHM-2-BamHI-fw AAA AGG ATC CAA TTT AAT CGC ACG AAT AG
Cloning of blaKHM-2 into pBK-CMV
Hoffmann (2013)
KHM-2-XhoI-rev TTT TCT CGA GTT ATT TCT TCT TTG CAA CC
Cloning of blaKHM-2 into pBK-CMV
Hoffmann (2013)
KHM-1_Bam_fw AAA AGG ATC CAT TTC TCA ATA AAA ATA TAG AAG G
Cloning of blaKHM-1 into pBK-CMV
This study
KHM-1_Hind_rev TTT TAA GCT TTC ACT TTT TAG CTG CAA GC
Cloning of blaKHM-1 into pBK-CMV
This study
536F_fournier CAG CAG CCG CGG TAA TAC
16-rRNA DIG probe synthesis
Fournier et al. (2010b)
RP2_fournier ACG GCT ACC TTG TTA CGA CTT
16-rRNA DIG probe synthesis
Fournier et al. (2010b)
KHM-2_DIG_fw2 GTT TGG TTT TTG TGG ATG GT
blaKHM-2 DIG probe synthesis
This study
KHM-2_DIG_fw2 CGA TTG ATA AGT TTT TCT GC
blaKHM-2 DIG probe synthesis
This study

Material and Methods 37
2.3 Methods
2.3.1 Microbiological methods
2.3.1.1 Growth and storage of bacterial cells
Bacterial cells were grown at 37 °C on LB, MacConkey, Mueller-Hinton and Columbia blood agar
plates or in liquid LB medium supplemented with appropriate antibiotics, if necessary. Agar
plates containing bacterial colonies were stored at 4 °C. For longterm storage, 800 µl of 88 %
glycerol were added to 800 µl of an overnight culture and the mixture was frozen in liquid
nitrogen. The cultures were stored at -80 °C.
LB medium 1 % (w/v) NaCl
(AppliChem) 1 % (w/v) Tryptone
0.5 % (w/v) Yeast extract
LB agar 1.5 % (w/v) Agar
(AppliChem) in LB medium
MacConkey agar 1.7 % (w/v) Peptone from gelatin
(Merck) 0.15 % (w/v) Peptone from casein
0.15 % (w/v) Peptone from meat
0.5 % (w/v) NaCl
1 % (w/v) Lactose
0.15 % (w/v) Bile salt mixture
0.003 % (w/v) Neutralred
0.0001 % (w/v) Crystalviolett
1.35 % Agar
Columbia blood agar 2.3 % (w/v) Peptone
(Thermo Scientific Oxoid) 0.1 % (w/v) Starch
1 % (w/v) NaCl
1 % (w/v) Agar
4 % (v/v) Defibrinated sheep blood

Material and Methods 38
2.3.1.2 Determination of bacterial growth in fluid cultures
Bacterial growth was determined by measuring the optical density (OD) of a culture at a
wavelength of 600 nm. A OD600 of 1 corresponds to 1-5 x 108 cells per ml. Turbidity of bacterial
suspensions in 0.8 % NaCl was optically measured using a VITEK Densichek (bioMérieux) in
relation to McFarland standards.
2.3.1.3 Preparation of chemically competent E. coli cells
100 ml of LB medium were supplemented with 2 ml Mg2+ solution and inoculated with 2 ml of an
overnight E. coli culture. The cells were grown to an OD600 of 0.5 at 37 °C. The cells were
harvested by centrifugation (4,000 × g, 4 °C, 10 min) and the pellet was resuspended in 50 ml
TMF buffer, previously cooled to 4 °C. After 1 h incubation on ice the cells were harvested again
(3,000 × g, 4 °C, 10 min) and the pellet was resuspended in 10 ml of cold TMF buffer and
supplemented with 3 ml of 88 % glycerol. The cells were aliquoted in reaction tubes to 250 µl
each and stored at -80 °C.
TMF buffer 40 mM MnCl2
50 mM RbCl
Mg2+ solution 500 mM MgCl2
500 mM MgSO4
2.3.1.4 Transformation of competent E. coli cells
A 250 µl aliquot of competent E. coli cells was thawed on ice. After addition of 1-3 µl of purified
plasmid DNA or 20 µl of a ligation mixture the cells were incubated on ice for 30 min.
Transformation was performed by heat shocking at 42 °C for 2 min. After heat shock 700 µl of LB
medium were added, followed by 1 h incubation at 37 °C at 300 rpm in an incubation shaker.
The cells were harvested by centrifugation (16,000 × g, 1 min), the supernatant was discarded
except for approx. 100 µl and the cell pellet was resuspended in the remaining supernatant. The
cells were plated on LB agar plates containing appropriate antibiotics and were incubated
overnight at 37 °C.
2.3.1.5 Conjugation experiments
For conjugation experiments, bacterial isolates and a recipient strain were mixed and inoculated
as a spot of around 1 cm onto Columbia blood agar plates. As recipient strains either E. coli C600
(rifampicin resistant) or E. coli J53 (sodium azide resistant) were used. After 18 h of incubation
transconjugants were selected on LB agar containing 100 mg/l ampicillin and selective
antibiotic (100 mg/l rifampicin or 200 mg/l sodium azide).

Material and Methods 39
2.3.2 Phenotypic methods for antibiotic resistance analysis
2.3.2.1 Disk diffusion antibiotic susceptibility test
Resistance to antibiotics was analyzed using paper disks containing single antibiotics or
combinations of antibiotics and β-lactamase inhibitors (Thermo Scientific Oxoid). Bacteria from
agar plates or liquid cultures were suspended in 0.8 % NaCl until a McFarland of 0.5-0.8 was
reached. MH2 agar plates (bioMérieux) were evenly inoculated with bacteria using a sterile
cotton tip. Antibiotic disks were placed on the plate with at least 1 cm distance from each other
or the agar edge. The plates were incubated overnight at 37 °C and resistance to antibiotics was
evaluated by measuring the inhibition zone diameters. The results were interpreted following
the guidelines of the European Committee on Antimicrobial Susceptibility Testing (EUCAST).
2.3.2.2 EDTA-Combined disk test (EDTA-CDT)
For phenotypic analysis of MBL production, the analyzed strain was plated on MH2 agar plates
as described in 2.3.2.1. Antibiotik disks containing carbapenems were placed on the plate in
duplicates and 10 µl of 0.5 M EDTA were pipetted on the disk. The plates were incubated
overnight at 37 °C and the inhibition zone diameters were measured. An increased inhibition
zone of the carbapenem/EDTA disk compared to the one without EDTA indicated an MBL
production.
2.3.2.3 Modified Hodge Test
To analyze if a bacterial strain was producing a carbapenemase, MH2 agar plates were
inoculated with E. coli ATCC 25922 as described in 2.3.2.1. Disks containing imipenem,
meropenem and ertapenem were placed on the inoculated agar and colony material from the
test strain was streaked in a straight line from the edge of one disk to another disk. The plates
were incubated overnight at 37 °C and the plate was examinend for clover leaf-like indentation
of E.coli ATCC 25922 growth along of the the test strain streak within the disk diffusion zone. A
growth of E.coli ATCC 25922 into the zone was interpreted as positive and no growth as negative.
2.3.2.4 Determination of minimal inhibitory concentration (MIC)
Determination of MICs was performed using the Etest gradient test strips (bioMérieux). MH2
agar plates were inoculated with bacteria as described in 2.3.2.1. The gradient strips were
placed onto the agar plate with sterile tweezers. The plates were incubated overnight at 37 °C.
The MIC was determined by reading the scale of the gradient strip at the position were bacterial
growth reached the strip. MIC values of clinical isolates were interpreted following the EUCAST
guidelines (http://www.eucast.org/clinical_breakpoints/; 15 March 2015, date last accessed)
(EUCAST, 2015)

Material and Methods 40
2.3.3 Molecular biology methods
2.3.3.1 Preparation of genomic DNA from Gram-negative bacteria
Genomic DNA from Gram-negative bacteria was extracted using the NucleoSpin Tissue kit
(Macherey-Nagel) following the manufacturer’s instructions.
2.3.3.2 Preparation of plasmid DNA from E. coli
Plasmid DNA from E. coli was extracted using the NucleoSpin Plasmid kit (Macherey-Nagel)
following the manufacturer’s instructions. For plasmids larger than 10 kb, the NucleoBond® PC
100 kit was used following the manufacturer’s instructions.
2.3.3.3 Purification of PCR-Products and DNA fragments from agarose gels
PCR products and DNA fragments from agarose gels were purified using the NucleoSpin Gel and
PCR Clean-up kit (Macherey-Nagel) following the manufacturer’s instructions.
2.3.3.4 Quick preparation of plasmid DNA from E. coli cells
For quick extraction of plasmid DNA from E. coli, the procedure described by Holmes & Quigley
(1981) was used with several modifications. Cells were grown in 4 ml of LB media overnight and
harvested by centrifugation (16,000 × g, 1 min). The pellet was resuspended in 50 µl boiling-
prep buffer and incubated for 3 min at room temperature, followed by incubation at 95 °C for
1 min. The samples were cooled on ice for 5 min and centrifuged (16,000 × g, 4 °C, 10 min). The
supernatant contained the plasmid DNA.
Boiling-prep buffer 10 mM Tris/HCL, pH 8.0
(Storage at -20 °C) 1 mM EDTA
15 % (w/v) Sucrose
2 mg/ml Lysozym
0.2 mg/ml RNAseA
0.1 mg/ml BSA
2.3.3.5 Agarose gel electrophoresis of DNA fragments
To determine the size of DNA fragments and for preparative purifaction of PCR products, DNA
fragments were separated by agarose gel electrophoresis using gels containing 0.8 % (w/v)
agarose in TBE buffer. The samples were mixed with loading buffer and applied to a gel.
Separation of DNA fragments was performed at a constant voltage of 50-150 V. After
electrophoresis, the gel was incubated in 0.05 % ethidium bromide for 10-30 min. After a
washing step in A. dest, the DNA bands were visualized under UV illumination using the Gel Doc

Material and Methods 41
XR+ system (Bio-Rad). The GeneRulerTM DNA Ladder Mix (Thermo Scientific) served as a size
marker.
TBE-buffer 89 mM Tris
89 mM Boronic acid
2 mM EDTA
pH 7.8
Loading buffer 0.5 % (w/v) Bromphenol blue
43 % (v/v) Glycerol
100 mM EDTA
2.3.3.6 Digestion of bacterial DNA with restriction endonucleases
Restriction of bacterial DNA was performed using FastDigest® restriction endonucleases and the
FastDigest buffer system (Thermo Scientific). The reactions contained 1-3 µl of appropriate
endonucleases and up to 16 µl of purified DNA and were carried out at 37 °C for 2-30 min in a
total volume of 20 µl. Restriction of vector DNA for ligation purposes additionally contained 1 µl
of FastAP alkaline phosphatase to prevent religation by dephosphorylation of the restricted
vector ends. For of ligation reactions, the enzymes were inactivated at 65-80 °C for 5-10 min
after restriction.
2.3.3.7 Ligation of DNA fragments
Ligation of DNA fragments with T4 DNA ligase was performed by mixing 1 µl of restricted and
dephosphorylated target vector DNA with 0.5-6 µl of the desired restricted DNA insert. The DNA
was incubated with 1 µl of T4 DNA ligase in 1 x T4 DNA ligase buffer (Thermo Scientific) in a
total volume of 20 µl for 1 h or overnight at room temperature.
2.3.3.8 Polymerase chain reaction (PCR) for amplification of DNA fragments
Amplifikation of DNA fragments up to 2 kb was performed as described by Saiki et al. (1988)
using Taq, Pfu and Pwo DNA polymerases in their appropriate buffer. For amplification of larger
DNA fragments (up to 8 kb), the FastStart High Fidelity kit (Roche) or the Expand Long Range
PCR System (Roche) were used following the manufacturer’s instructions and the supplied
buffers.
2.3.3.9 Genome walking
To amplify and sequence larger parts of unknown genomic DNA starting from a known gene
sequence, the Universal GenomeWalker 2.0 kit (Clontech) was used following the

Material and Methods 42
manufacturer’s instructions. Total DNA of bacteria was digested with various restriction
endonucleases to obtain uncloned DNA libraries. The DNA fragments were ligated with adaptor
nucleotides with known sequences. The fragment-adaptor nucleotides were amplified by PCR
with an adaptor-specific and a gene-specific oligonuncleotide and the products were sequenced
(2.3.3.12).
2.3.3.10 DNA synthesis
Synthesis of bacterial DNA was performed by Eurofins Genomics and provided subcloned into
the pEX-A2 vector.
2.3.3.11 Determination of DNA and protein concentration in aqueous solutions
DNA concentrations and quality were determined photometrically using the Eppendorf
BioSpectrometer with the Eppendorf µCuvette G1.0 by measuring the absorbance of a sample at
260 and 280 nm. Protein concentrations were either determined using the modified Bradford
assay Roti-Nanoquant (Roth) following the manufacturer’s instructions or using the Qubit 2.0
fluorometer with the Qubit Protein Assay Kit (Life Technologies).
2.3.3.12 Sequencing of plasmid and genomic DNA
Sequencing of bacterial DNA was performed in publication quality with Sanger-sequencing by
third-party companies (Seqlab, GATC) following the companies DNA sample requirements.
2.3.3.13 Sequencing of plasmid DNA by 454-pyrosequencing
Large wildtype plasmids were sequenced using the 454 next generation sequencing technique
and the GS Junior system (Roche) at the Department of Virology (RUB) following the
manufacturer’s manuals. 300 ng of purified plasmid DNA were fragmented by nebulization and
damaged ends were repaired by T4 Ligase and Taq Polymerase. An adaptor molecule was
ligated with the DNA fragments and the fragments were bound to magnetic beads. Small
fragments were removed by washing and pelleting steps using the magnetic beads within a
magnetic tube holder. The library quality was analyzed by gel electrophoresis after separating
the DNA fragments from the magnetic beads by addition of TE buffer. The library was quantified
fluorometrically and the library sample concentration was calculated on the basis of a standard
curve. The DNA library was bound to capture beads and amplified by random amplified PCR in a
96-well plate. After amplification, the library was recovered from the plate and washed with
ethanol and isopropanol for removal of PCR reagents. The library was transferred to Enrichment
beads by several washing steps and the sequencing oligonucleotides were annealed. The library
was adjusted to a total amount of approx. 500,000 enriched beads, determined using the GS
Junior Bead Counter. 454 sequencing was performed by loading the bead-bound DNA library on

Material and Methods 43
a GS Junior Titanium Pico TiterPlate, preloaded with the enzymes required for the sequencing
reaction. The sequence of the DNA fragments was determined by single nucleotide addition. If a
nucleotide is ligated to the single stranded DNA, the released pyrophosphate is converted into
ATP by an ATP sulfurylase and used by a luciferase to convert luciferin to oxyluciferin which
leads to light emission. The light emission is detected by a bioluminescence camera and the DNA
is sequenced. After sequencing, the DNA sequence fragments were assembled using the GS
Junior Sequencer software.
2.3.3.14 Shotgun-cloning approach for the identification of bacterial resistance genes
In order to determine the molecular mechanism of increased β-lactam resistance of Gram-
negative clinical isolates, genomic DNA of the isolates was digested with restriction
endonucleases whose specific recognition sites statistically occur with high frequency in Gram-
negative bacterial DNA. Namely, BamHI (GGATCC), HindIII (AAGCTT), EcoRI (GAATCC), XhoI
(CTCGAG), MboI and Sau3AI (both GATC) were used. The aim was to receive relatively small
DNA fragments with sizes from 0.5-6 kb. For MboI, digestion was performed for only 2-5 min, as
the MboI recognition site (GATC) is very frequent in bacterial DNA and a longer incubation
would lead to DNA fragments too small for efficient cloning. After restriction, the DNA fragments
were ligated (2.3.3.7) with the pBK-CMV vector which was previously digested with the same
restriction enzyme (for MboI: BamHI) and the ligation preparation was transformed into E. coli
TOP10 (2.3.1.4). The cells were plated on LB agar plates containing 50-100 mg/l ampicillin and
50 mg/l kanamycin to select for cells that received complete genes that confer higher levels of β-
lactam resistance. The plates were incubated overnight at 37 °C. In case of bacterial growth,
plasmid DNA was extracted from the cells (2.3.3.2) and the insert of the pBK-CMV vector was
sequenced using appropriate oligonucleotides (Table 2.3 & 2.3.3.12).
2.3.3.15 Multilocus sequence typing (MLST) and clonal complex analysis
The sequence type of P. aeruginosa strains was determined following the instructions of Curran
et al. (2004). Fragments of seven housekeeping genes (acsA, aroE, guaA, mutL, nuoD, ppsA and
trpE) were amplified by PCR (2.3.3.8) and sequenced (2.3.3.12). The sequence type of the isolate
was identified with the sequence definition tool of the P. aeruginosa MLST web site
(http://pubmlst.org/paeruginosa/). eBURST analysis was performed to determine the clonal
complex to which the isolate belongs using the eBURST software (Feil et al., 2004).
2.3.3.16 Pulsed field gel electrophoresis (PFGE)
PFGE was used to separate genomic and plasmid DNA by size with lengths of up to 3 Mbp. The
DNA samples were preparated by inoculating 10 ml of TN buffer with colony material from agar
plates to a McFarland of 0.3-0.8. After a centrifugation step (4,000 × g, 4 °C, 15 min) the

Material and Methods 44
supernatant was discarded and the pellet was resuspended in 500 µl EC buffer. The solution was
warmed to 45 °C and RNase A and lysozyme were added to a final concentration of 20 µg/l and
2 mg/ml, respectively. 500 µl pre-warmed InCert agarose were added and mixed with the
bacterial suspension by inverting the tube 2-3 times. Immediately after mixing, the suspension
was transfered into the slots of a Disposable Plug Mold (Bio-Rad) with 100 µl each. The gel plugs
were incubated for 10 min at 4 °C until the plugs were solid. The plugs were incubated in a
thermoshaker for 1 h at 37 °C and 300 rpm in 1 ml EC lysis buffer. Afterwards, the lysis buffer
was removed and the plugs were incubated 18-24 h at 50 °C and 300 rpm with proteinase K
with a final concentration of 1 mg/ml. After the incubation the buffer was removed and the plugs
were washed two times in TE buffer with 1 mM AEBSF for 2 h at 37 °C and 300 rpm, followed by
two washing steps with TE buffer for 1 h. After washing, the plugs were stored in ES buffer
overnight. As the next step the plugs were incubated two times for 15 min in TN buffer, followed
by incubation in the specific buffer for the restriction enzyme supplied by the manufacturer. For
plasmid PFGE, the DNA was restricted with Nuclease S1, which linearizes plasmid DNA. For
chromosomal localization studies, the DNA was restricted with I-Ceu I, which recognizes a highly
conserved region in rrn genes (Liu et al., 1993). After restriction the plugs were stored in ES
buffer at 4 °C. PFGE was performed by inserting the gel plugs into the slots of a 1 % SeaKem
agarose gel and the gel was run with switch times from 1-200 s at 6 V/cm with an angle of 120 °
for 12-36 h in TBE buffer (Bio-Rad). The DNA bands were visualized under UV illumination and
the gel was subjected to Southern blotting, if applicable.
TN buffer 1 M NaCl
10 mM Tris/HCl, pH 7.6
pH 7.6
EC buffer 1 M NaCl
6 mM Tris/HCl, pH 7.6
100 mM EDTA
0.5 % (w/v) Brij 58
0.2 % (w/v) Sodium deoxycholate
0.5 % (v/v) Sodium lauroyl sarcosinate
pH 7.6
ES buffer 0.5 M EDTA
1 % (v/v) Sodium lauroyl sarcosinate

Material and Methods 45
TE buffer 10 mM Tris/HCl, pH 7.5
1 mM EDTA
2.3.3.17 Southern Blotting and hybridization with gene specific probes
For Southern Blotting experiments, DNA fragments were separated by PFGE (2.3.3.16) and the
gel was placed onto a positively charged nylon membrane, previously wetted with SSCx2 buffer.
The gels were covered with 0.25 M HCl for depurination and a vacuum was applied for 20 min.
Afterwards the HCl was removed and the gel was covered with 1 M NaOH. Transfer of DNA to
the membrane was achieved by vacuum application for 90 min. The membrane was washed two
times in SSCx2 buffer and dried for 30 min at 120 °C. Gene specific digoxigenin-labeled probes
were produced by using the PCR DIG Probe Synthesis kit (Roche) with gene-specific
oligonucleotides following the manufacturer’s instructions. The membrane was incubated for
30 min at 42 °C in hybridization buffer. 10 µl of the DIG PCR product were added to 40 ml of
hybridization buffer and the membrane was incubated with the probe overnight at 42 °C with
gentle shaking. After probe incubation the blot was washed two times in washing buffer 1 at
room temperature for 5 min, followed by two washing steps in washing buffer 2 at 65 °C for
15 min. Afterwards the membrane was incubated in DIG washing buffer for 2 min and in DIG
blocking buffer for 30 min with gentle shaking. After blocking, the blot was incubated with 4 µl
Anti-DIG-AP in 40 ml DIG blocking buffer for 30 min. The membrane was washed two times with
DIG washing buffer for 15 min and transfered into DIG substrate buffer. The phosphatase
reaction was started by plating 4 µl of CSPD in 0.4 ml DIG substrate buffer evenly over the
membrane. Chemiluminescence signals were detected with HyperFilmTM autoradiography films
(GE Healthcare).
SSC×20 buffer 3 M NaCl
0.3 M Sodium citrate
pH 7.0
DIG maleate buffer 0.1 M Maleic acid
0.15 M NaCl
Blocking stock 10 % (w/v) Blocking Reagent (Roche)
in DIG Maleate buffer

Material and Methods 46
Hybridization buffer 50 % (v/v) Formamide
2 % (v/v) Blocking stock
0.1 % (v/v) Sodium lauroyl sarcosinate
0.02 % (v/v) SDS
in SSC×5
Washing buffer 1 0.1 % (v/v) SDS
in SSC×2
Washing buffer 2 0.1 % (v/v) SDS
in SSC×0.5
DIG washing buffer 0.3 % (v/v) Tween20
in DIG maleate buffer
DIG blocking buffer 1 % (v/v) Blocking stock
in DIG maleate buffer
DIG substrate buffer 0.1 M Tris, pH 9.5
0.1 M NaCl
0.05 M MgCl2
2.3.4 Biochemical methods
2.3.4.1 Purification of β-lactamases by fast protein liquid chromatography (FPLC)
E. coli TOP10 cells expressing a β-lactamase were grown in 4 l LB medium at 37 °C for 18 h and
harvested by centrifugation (3,600 × g, 4 °C, 30 min). Cell pellets were resuspended in 50 ml
buffer H or 0.1 M sodium phosphate buffer, depending on the expressed β-lactamase. Metallo-β-
lactamase-producing cells were resuspended in buffer H while Ambler class D β-lactamase
producing cells were resuspended in sodium phosphate buffer. The cells were disrupted by
sonication at 4 °C. The lysates were cleared by centrifugation (48.400 × g, 4 °C, 30 min), filtered
through a Filtropur S 0.2 µm filter (Sarstedt) and desalted with a HiPrep 26/10 desalting column
(GE Healthcare) using the Äkta Pure automated FPLC system (GE Healthcare). The column was
previously equilibrated with buffer H or 0.1 M phosphate buffer. After desalting, the protein-
containing fractions were loaded onto a 5 ml HiTrap SP HP ion exchange column (GE
Healthcare) at a flow rate of 2 ml/min, previously equilibrated with buffer H or 0.1 M phosphate
buffer. The column was washed with 10 ml of buffer H and bound proteins were eluted using a
linear NaCl gradient (0 to 1 M). Fractions containing high amounts of protein were loaded onto a

Material and Methods 47
HiPrep 16/60 S-200 HR gel filtration column (GE Healthcare) equilibrated with buffer H or 0.1 M
phosphate buffer, both supplemented with 0.15 M NaCl at a flow rate of 0.8 ml/min. β-lactamase
containing fractions were identified by incubating 5 µl of eluate with 5 µl of a 1 mM solution of
the chromogenic cephalosporin nitrocefin. A color change from yellow to red indicated the
presence of a β-lactamase in the sample. Fractions that contained high amounts of β-lactamase
activity were pooled, immediately frozen in liquid nitrogen and stored at -80 °C.
Buffer H 50 mM HEPES
50 µM ZnSO4
pH 7.5
Sodium phosphate buffer 0.1 M Sodium phosphate
pH 5.9
2.3.4.2 SDS polyacrylamide gel electrophoresis
Separation of proteins based on their molecular weight was performed as described by Laemmli
(1970). Protein samples were mixed with SDS sample buffer and were incubated at 100 °C for
10 min. 15-20 µl of the sample were loaded onto a 4 % acrylamide stacking gel and seperated by
gel electrophoresis in an 11 % acrylamide running gel. Electrophoresis was performed at a
voltage of 20-150 V for 1-2 h in SDS running buffer. The gels were stained with Coomassie
Brilliant blue after completion of electrophoresis. The PageRuler Plus Prestained standard
(Thermo Scientific) served as a size marker.
SDS sample buffer 10 % (w/v) SDS
5 % (v/v) β-Mercaptoethanol
0.5 % (w/v) Bromphenol blue
250 mM Tris/HCl, pH 6.8
50 % (v/v) Glycerol
4 % stacking gel 0.5 M Tris/HCl pH 6.8
4 % (v/v) Acrylamide/Bisacrylamide
(37.5 : 1)
0.4 % (w/v) SDS
0.06 % (v/v) TEMED
0.1 % (w/v) APS

Material and Methods 48
11 % running gel 0.5 M Tris/HCl, pH 8.8
11 % (v/v) Acrylamide/Bisacrylamide
(37.5 : 1)
0.1 % (w/v) SDS
0.04 % (v/v) TEMED
0.1 % (w/v) APS
SDS running buffer 25 mM Tris/HCl, pH 8.3
250 mM Glycine
0.1 % (w/v) SDS
2.3.4.3 Isoelectric focussing
For determination of the isoelectric point of β-lactamases cell-free extracts or purified β-
lactamases were subjected to isoelectric focussing. Cell free extracts were prepared from 10 ml
of an overnight culture. The cells were disrupted by vigorous shaking with glass beads using the
PowerLyzer 24. The lysate was cleared by centrifugation (16,000 × g, 5 min). For isoelectric
focusing 2 µl of the supernatant or 2 µl of purified enzyme solution were applied onto a CleanGel
IEF gel (GE Healthcare) together with reference standards. The gel was cooled to 7 °C during the
procedure and was run for 30 min at 500 V, 20 mA and 5 W, for 90 min at 1700 V, 20 mA and
25 W and for 30 min at 2000 V, 20 mA and 30 W. β-lactamases were visualized by adding 2 ml of
1 mM Nitrocefin spread evenly over the gel.
2.3.4.4 Determination of kinetic parameters of purified β-lactamases
Hydrolysis of β-lactam antibiotics was monitored by measuring the absorbance changes
resulting from the opening of the β-lactam ring. All measurements were performed at 25 °C in
buffer H or buffer P in a total volume of 500 µl. The following wavelenghts were used for β-
lactam hydrolysis monitoring with an Eppendorf BioSpectrometer (Eppendorf):
235 nm Penicillin G, Ampicillin, Piperacillin
260 nm Oxacillin, Cefoxitine, Ceftazidime, Cefotaxime
300 nm Imipenem, Meropenem, Ertapenem
320 nm Aztreonam
The substrate concentrations ranged from 0.5 to 1,600 µM, depending on hydrolytic efficiency.
Hydrolysis was started by enzyme addition and the reaction was monitored over 10 min with

Material and Methods 49
enzyme concentrations ranging from 0.01 to 0.2 µM. For each substrate, the molar extinction
coefficient (ε) was determined using the Lambert-Beer law.
𝐴 = 𝜖 ∙ 𝑐 ∙ ℓ
A: absorption
c: concentration
ℓ: path length
The initial rate slopes were calculated from the the reactions linear phase with linear regression
using the GraphPad Prism 6 software. The initial velocity of the reaction was determined with a
modified Lambert-Beer law and the following formula:
0V =𝛥𝐴
𝜀 ∙ ℓ ∙ ∆𝑡
V0: initial reaction velocity
ΔA/Δt: absorbtion change per time
ε: molar extinction coefficient of the substrate
ℓ: path length
The Vmax and Km kinetic parameters were determined with nonlinear regression using the
Michaelis-Menten equitation with the GraphPad Prism 6 software. The turnover number kcat was
determined by dividing Vmax by the enzyme concentration.
Buffer H 50 mM HEPES
50 µM ZnSO4
pH 7.5
Buffer P 0.1 M Sodium phosphate
50 mM NaHCO3
pH 7.0

Material and Methods 50
2.3.5 In silico methods
2.3.5.1 In silico DNA and amino acid sequence analysis
Computational DNA sequence analysis was performed using various bioinformatic tools. In silico
restriction, ligation, cloning and comparison of DNA and protein sequences were performed
using the Clone Manager 5 software (Sci-Ed). Bacterial promotor structures were analyzed using
the online web tools PromoterHunter (Klucar et al., 2010), BPROM (Solovyev & Salamov, 2011)
and SCOPE (Carlson et al., 2007). Integron promoters were analyzed following the classifications
of Papagiannitsis et al. (2009). Annotation of DNA sequences was performed using RAST (Aziz et
al., 2008; Overbeek et al., 2014) and by manual annotation using BLAST
(http://blast.ncbi.nlm.nih.gov) and the NCBI nucleotide database
(http://www.ncbi.nlm.nih.gov/). Graphical alignment of large DNA sequences was performed
using Mauve (Darling et al., 2004; Darling et al., 2010). Prediction of N-terminal signal peptide
sequences was done using the SignalP server (Petersen et al., 2011). Circular views of plasmids
were constructed using DNAPlotter (Carver et al., 2009). Inc-typing of plasmid sequences was
performed using the web-based PlasmidFinder 1.2 software (Carattoli et al., 2014).
2.3.5.2 Phylogenetic analysis of β-lactamases
Amino acid sequences were aligned using the ClustalW2 algorithm in MEGA6 software (Tamura
et al., 2013). Phylogenetic trees based on alignments were constructed using the neighbour-
joining method with 1000 times bootstrapping and the Dayhoff model in MEGA6. Cluster
analysis was performed using the Ctree software (Archer & Robertson, 2007).
2.3.5.3 Tertiary structure modelling
Protein structure modelling based on amino acid homology was done using the SWISS-MODEL
webserver (Arnold et al., 2006; Guex et al., 2009; Kiefer et al., 2009; Biasini et al., 2014). Protein
models were visualized using PyMol (http://www.pymol.org/pymol).
2.3.5.4 SDS-PAGE analysis
Relative quantification of bands detected in SDS gels was performed using the GelDoc XR+
software (Bio-Rad).

Results 51
3 Results
The ongoing spread and diversification of carbapenemases in Gram-negative pathogens is a
major clinical problem. At the German National Reference Laboratory for multidrug-resistant
Gram-negative bacteria, three strains attracted attention as the molecular basis of carbapenem
resistance was could not be determined in the routine diagnostic process. It was suspected that
these isolates harbour novel carbapenemases.
3.1 The search for novel carbapenemases
3.1.1 Identification of IMP-31 in Pseudomonas aeruginosa NRZ-00156
P. aeruginosa NRZ-00156 was found to be highly resistant to carbapenems in routine diagnostics
and showed a metallo-β-lactamase phenotype. As all PCRs for MBL-type genes were negative, it
was suspected that the isolate harboured a novel MBL.
The isolate was analyzed phenotypically by a modified Hodge Test and an EDTA-CDT to ensure
the production of a metallo-β-lactamase. The results of these tests are shown in Figure 3.1. The
modified Hodge Test indicated the production and secretion of a carbapenemase, as the
indicator strain was able to grow along the streak of P. aeruginosa NRZ-00156. The EDTA-CDT
was clearly positive with an increase in the inhibition zone diameter of 10 mm for
imipenem/EDTA and 6 mm for meropenem/EDTA, while the control showed an increase of only
Figure 3.1 Modified Hodge Test and EDTA-CDT of P. aeruginosa NRZ-00156. (A) Modified Hodge Test. The indicator strain E. coli ATCC 25922 was plated on an MH2 agar plate and P. aeruginosa NRZ-00156 was streaked between disks containing imipenem (IPM), meropenem (MEM) and ertapenem (ETP). An invaginating growth of the indicator strain along the test strain streak is highlighted by white arrows. (B) EDTA-CDT. P. aeruginosa NRZ-00156 was plated on an MH2 agar plate. Carbapenem disks were placed in duplicate and EDTA was added to one of the disks. A blank disk with EDTA served as a control.

Results 52
4 mm. Consequently, the test indicated the production of an MBL by the isolate. For more
detailed resistance analysis, the minimal inhibitory concentrations (MIC) for various β-lactams
were determined. The results of the MIC analysis are shown in Table 3.1. According to EUCAST
criteria, the isolate was resistant to piperacillin, piperacillin/tazobactam, cefepime and
ceftazidime. Regarding the β-lactams not covered by the EUCAST criteria as not commonly used
for therapy against P. aeruginosa due to intrinsic resistance, the isolate showed MICs that often
exceeded the detection range. Susceptibility was detected only for the monobactam aztreonam.
With MICs higher than 32 mg/l, the isolate was resistant to the carbapenems imipenem,
meropenem and doripenem which was in accordance with the production of a potent
carbapenemase. To ensure that the isolate did not harbour a known carbapenemase gene that
was accidentally not detected in routine diagnostics, a PCR screening on VIM, IMP, NDM, KHM,
SPM, GIM, SIM, DIM, AIM and FIM-type carbapenemase genes was performed. Surprisingly, a
Table 3.1 β-lactam MICs of P. aeruginosa NRZ-00156. Shown are the MICs detected by Etest strips and their interpretation according to EUCAST criteria (R, resistant; I, intermediate, S, susceptible).
Antibiotic P. aeruginosa NRZ-00156
Interpretation according to EUCAST criteria
Ampicillin >256a -b
Ampicillin-sulbactam >256 -
Piperacillin 96 R
Piperacillin-tazobactam 64 R
Amoxicillin >256 -
Amoxicillin-clavulanate 48 -
Temocillin >1024 -
Cephalotin >256 -
Cefuroxime >256 -
Cefoxitin >256 -
Cefotaxime >256 -
Ceftriaxone >256 -
Cefepime >256 R
Ceftazidime >256 R
Imipenem >32 R
Meropenem >32 R
Doripenem >32 R
Ertapenem >32 -
Aztreonam 8 S
aThe MIC was higher than detectable by Etest strips, which usually have a concentration range of up to 256 mg/l. bNo clinical MIC EUCAST-breakpoint data are available for these antibiotics for P. aeruginosa.

Results 53
PCR with the oligonucleotides IMP-A and IMP-B (Table 2.3) yielded an amplificate for IMP-type
genes with a size of approximately 600 bp (data not shown). As the oligonucleotides IMP-A and
IMP-B are degenerate and bind to a large number of blaIMP-type genes, the amplificate was
sequenced and showed distinct differences to all other known blaIMP sequences that were
publicly available. To sequence the whole open reading frame (ORF) of the potentially new blaIMP
variant, a combination of oligonucleotides for conserved class 1 integron regions and blaOXA-10-like
genes was used for PCRs, as it is known that blaIMP genes are often associated with blaOXA genes
within class 1 integron structures. PCRs with the oligonucleotide combinations 5’CS/IMP-B and
IMP-A/OXA-10B (Table 2.3) yielded amplificates that covered the whole blaIMP ORF and a few
hundred base pairs of the flanking genetic environment. The ORF had a size of 738 bp and coded
for a 245 amino acid protein. On nucleotide level, the sequence showed a homology of 86 % to
blaIMP-8 and blaIMP-24. With regard to blaIMP-1, it showed a homology of only 83 %. On amino acid
level, the novel IMP variant showed a homology of 84.1 % to IMP-8 and 83.7 % to IMP-2, IMP-19,
IMP-20 and IMP-24. With only 80.0 % homology, IMP-31 was the most divergent IMP-type
enzyme relative to the reference enzyme IMP-1. The nucleotide and protein sequences were
submitted to the international β-lactamase numbering institution (K. Bush & G. Jacoby, Lahey
Clinic Medical Centre, Burlington, U.S.; http://www.lahey.org/studies) and the enzyme was
assigned as IMP-31. The nucleotide sequence of blaIMP-31 was submitted to the NCBI database
(accession number KF148593.1). Shortly after, the amino acid sequence of another novel
IMP-type enzyme, IMP-35, was published by another working group and showed a homology of
96.7 % to IMP-31, making IMP-35 the current next nearest relative. An alignment of the amino
acid sequences of IMP-31, IMP-35 and IMP-1 is shown in Figure 3.2. Consisting of 245 amino
Figure 3.2 Amino acid sequence alignment of IMP-31, IMP-35 and IMP-1. The highly conserved zinc binding ligands of IMP-type enzymes are marked with asterisks.

Results 54
acid residues, IMP-31 lacked one C-terminal amino acid compared to IMP-35 and IMP-1. In
contrast to all other known IMP variants, where the C-terminus is mostly formed by a KKXSXPSX
motif, the C-terminal amino acid sequence of IMP-31 was KNHHSPK and therewith showed a
significant difference. IMP-31 showed 54 amino acid substitutions compared to IMP-38,
resulting in an identity of only 78.2 % which is highest diversity of all known IMP variants
compared to each other (data not shown). IMP-31 showed no alterations of the highly conserved
zinc binding ligands of subclass B1 MBLs.
To correlate IMP-31 with all other IMP-type enzymes with publicly available amino acid
sequences, a phylogenetic tree was constructed and this tree is shown in Figure 3.3. Cluster
analysis using the CTree software clustered the enzymes into thirteen groups, the IMP-1
(including IMP-3, IMP-4, IMP-6, IMP-10, IMP-25, IMP-26, IMP-30, IMP-34, IMP-38, IMP-40 and
IMP-42), IMP-2 (including IMP-8, IMP-19, IMP-20 and IMP-24), IMP-5 (including IMP-7, IMP-15,
IMP-28 and IMP-43), IMP-9 (including IMP-45), IMP-11 (including IMP-21, IMP-41 and IMP-44),
IMP-12 (including only IMP-12), IMP-13 (including IMP-33 and IMP-37), IMP-14 (including
Figure 3.3 Phylogenetic analysis of IMP-31. The tree was constructed based on aligned amino acid sequences of all 42 IMP-type MBLs with publicly available sequences after removal of their N-terminal signal peptides. Construction was performed using the neighbour-joining method with 1000 times bootstrapping and the Dayhoff model. Replicate tree percentages during bootstrapping are shown next to the branches. Clusters were analyzed using the C-tree algorithm and are indicated by parenthezised numbers. Scale: 0.02 substitutions per site.

Results 55
IMP-32 and IMP-48), IMP-16 (including IMP-22), IMP-18 (including only IMP-18), IMP-27
(including only IMP-27), IMP-29 (including only IMP-29) and IMP-31 (including IMP-35) groups.
The analysis showed that IMP-31 and IMP-35 formed a cluster that showed the highest diversity
to any other IMP-type enzyme cluster.
To acquire more information on the isolate P. aeruginosa NRZ-00156 and to be able to classify
the isolate in an epidemiological context, the MLS type of the isolate was determined by
amplification and sequencing of seven P. aeruginosa housekeeping genes. The sequence types
and the corresponding MLS type were determined using the sequence definition tool of the
P. aeruginosa MLST web site (http://pubmlst.org/paeruginosa/). The results are summarized in
Table 3.2. Analysis of the allele types showed that P. aeruginosa NRZ-00156 expressed an allelic
profile consistent with ST235, which belongs to the clonal complex CC235.
Table 3.2 MLS typing of P. aeruginosa NRZ-00156. Listed are the seven P. aeruginosa MLST housekeeping genes and the corresponding allele types of P. aeruginosa NRZ-00156.
Gene acsA aroE guaA mutL nuoD ppsA trpE allele type 38 11 3 13 1 2 4
3.1.2 Identification of OXA-233 in Citrobacter freundii NRZ-02127
C. freundii NRZ-02127 attracted attention in routine diagnostics, as the isolate showed
susceptibility to oxyimino-cephalosporins but elevated resistance to carbapenems, which was
inhibited by clavulanic acid. As PCRs for the most common class A and D β-lactamase genes were
negative, it was suspected that the isolate harboured a novel β-lactamase.
To ensure the production of a carbapenemase by C. freundii NRZ-02127, the isolate was analyzed
by a modified Hodge Test, which is shown in Figure 3.4. The growth of the indicator strain along
the streak of C. freundii NRZ-02127 indicated a carbapenemase secretion by the isolate. To
Figure 3.4 Modified Hodge Test of C. freundii NRZ-02127. The indicator strain E. coli ATCC 25922 was plated on an MH2 agar plate and C. freundii NRZ-02127 was streaked between disks containing imipenem (IPM), meropenem (MEM) and ertapenem (ETP). An invaginating growth of the indicator strain along the test strain streak is highlighted by white arrows.

Results 56
further characterize the resistance phenotype of the isolate, the MICs for various β-lactams were
determined. The results are summarized in Table 3.3. The isolate was resistant to penicillins and
showed a noticable inhibition by clavulanic acid, but not by sulbactam or tazobactam. It was
resistant to cefuroxime, but showed very low MICs for oxyimino-cephalosporins and was
interpreted as susceptible to cefepime and intermediate to ceftazidime according to the EUCAST
criteria. Carbapenem MICs were interpreted as intermediate for imipenem and meropenem and
resistant for doripenem and ertapenem. To identify the molecular basis of this resistance
phenotype, shotgun cloning experiments were performed. The experiments yielded a β-lactam
resistant clone that showed the same resistance profile as C. freundii NRZ-02127 with increased
resistance to carbapenems but not to oxyimino- cephalosporins. Sequencing of the insert of the
recombinant plasmid pMB3002 revealed an 801-bp ORF coding for a protein consisting of
Table 3.3 β-lactam MICs of C. freundii NRZ-02127. Shown are the MICs detected by Etest strips and their interpretation according to EUCAST criteria (R, resistant; I, intermediate, S, susceptible).
Antibiotic C. freundii NRZ-02127
Interpretation according
to EUCAST criteria
Ampicillin >256a R
Ampicillin-sulbactam >256 R
Piperacillin >256 R
Piperacillin-tazobactam >256 R
Amoxicillin >256 R
Amoxicillin-clavulanate 64 R
Temocillin 64 -b
Cephalotin >256 -
Cefuroxime 24 R
Cefoxitin >256 -
Cefotaxime 0.75 S
Ceftriaxone 0.75 -
Cefepime 0.38 S
Ceftazidime 1.5 I
Imipenem 3 I
Meropenem 6 I
Doripenem 3 R
Ertapenem >32 R
Aztreonam 0.5 S
aThe MIC was higher than detectable by Etest strips, which usually have a concentration range of up to 256 mg/l. bNo clinical MIC EUCAST-breakpoint data are available for these antibiotics for Enterobacteriaceae according.

Results 57
266 amino acids. A BLAST homology search revealed that the sequence was 99.8 % identical to
the blaOXA-17 gene and 99.75 % to the blaOXA-19 gene. Sequence analysis showed that the gene
exhibited a single nucleotide substitution compared to the blaOXA-17 gene at position 349 from
guanine to tymine. Compared to the blaOXA-10 gene, the sequence showed an additional
substitution at position 218 from alanine to guanine. The sequence of the novel blaOXA gene was
submitted to the international β-lactamase numbering institution and the encoded enzyme was
assigned as OXA-233. The nucleotide sequence of blaOXA-233 was submitted to the NCBI database
(accession number KJ657570.1). OXA-233 was compared with the two next nearest relatives
OXA-17 and OXA-10 and an alignment of the amino acid sequences is shown in Figure 3.5. The
amino acid sequences of OXA-233 and OXA-17 differed in a valine to phenylalanine substitution
at the highly conserved position 117. Compared to OXA-10, OXA-233 exhibited an additional
point mutation at position 73 from asparagine to serine, while this mutation is also found in
OXA-17. This resulted in identities of 99.2 % to OXA-10 and 99.6 % to OXA-17. The highly
conserved STFK-motif at positions 67 to 70 which includes the active site serine was not altered
in OXA-233. No MLS typing scheme existed for C. freundii at that time, and the MLS type of the
isolate could not be determined. As class D β-lactamases are a very heterogenous group of
enzymes and as OXA-233 was closely related solely to enzymes of the OXA-10 subgroup, no
further phylogenetic analysis was performed.
Figure 3.5 Amino acid sequence alignment of OXA-233, OXA-17 and OXA-10. Highly conserved regions of class D β-lactamases are highlighted. The active site serine residue of class D β-lactamase is marked with an asterisk.

Results 58
3.1.3 Identification of KHM-2 in Pseudomonas aeruginosa NRZ-03096
Like P. aeruginosa NRZ-00156, P. aeruginosa NRZ-03096 attracted attention in routine
diagnostics as the isolate exhibited high carbapenem resistance that was inhibited by EDTA,
indicating an MBL production. As all diagnostic PCRs were negative, the isolate was suspected to
harbour a novel MBL.
The isolate was analyzed phenotypically for metallo-β-lactamase production by a modified
Hodge Test and an EDTA-CDT. The results are shown in Figure 3.6 and the isolate indicated a
carbapenemase secretion in the Hodge Test. MBL production was indicated by increased
inhibition zone diameters of 13 mm for imipenem/EDTA (10 to 23 mm) and 19 mm (6 to
25 mm) for meropenem/EDTA, while the control showed an inhibition zone diameter of 19 mm.
Determination of the MICs for β-lactams and interpretation according the EUCAST criteria
showed that the isolate was susceptible to piperacillin and piperacillin/tazobactam and resistant
to cefepime and ceftazidime with MICs of 64 and >256 mg/l, respectively (Table 3.4). The isolate
was susceptible to imipenem with an MIC of only 3 mg/l, while it was intermediate for
meropenem and resistant to doripenem with MICs of 6 and 8 mg/l, respectively. The MIC for
ertapenem was >32 mg/l. Regarding the antibiotics with no EUCAST breakpoints, the isolate
showed high MICs for ampicillin, amoxicillin and most cephalosporins with an MIC of >256 mg/l.
Although the carbapenem MICs were not as high as for the IMP-31 containing isolate
P. aeruginosa NRZ-00156, the observed values indicated the presence of a carbapenem
resistance mechanism.
To exclude that the isolate harboured a known carbapenemase gene that was not detected in
Figure 3.6 Modified Hodge Test and EDTA-CDT of P. aeruginosa NRZ-03096. (A) Modified Hodge Test. The indicator strain E. coli ATCC 25922 was plated on an MH2 agar plate and P. aeruginosa NRZ-03096 was streaked between disks containing imipenem (IPM), meropenem (MEM) and ertapenem (ETP). A growth of the indicator strain along the test strain streak is highlighted by white arrows. (B) EDTA-CDT. P. aeruginosa NRZ-03096 was plated on an MH2 agar plate. Carbapenem disks were placed in duplicate and EDTA was added to one of the disks. A blank disk with EDTA served as a control.

Results 59
Table 3.4 β-lactam MICs of P. aeruginosa NRZ-03096. Shown are the MICs detected by Etest strips and their interpretation according to EUCAST criteria (R, resistant; I, intermediate, S, susceptible).
Antibiotic
P. aeruginosa
NRZ-03096
Interpretation according
to EUCAST criteria
Ampicillin >256a -b
Ampicillin-sulbactam >256 -
Piperacillin 4 S
Piperacillin-tazobactam 2 S
Amoxicillin >256 -
Amoxicillin-clavulanate >256 -
Temocillin >1024 -
Cephalotin >256 -
Cefuroxime >256 -
Cefoxitin >256 -
Cefotaxime >256 -
Ceftriaxone >256 -
Cefepime 64 R
Ceftazidime >256 R
Imipenem 3 S
Meropenem 6 I
Doripenem 8 R
Ertapenem >32 -
Aztreonam 1.5 I
aThe MIC was higher than detectable by Etest strips, which usually have a concentration range of up to 256 mg/l. bNo clinical MIC EUCAST-breakpoint data are available for these antibiotics for Enterobacteriaceae.
routine diagnostics, a PCR screening on VIM, IMP, NDM, KHM, SPM, GIM, SIM, DIM, AIM and FIM-
typ MBL genes was performed, but all PCRs were negative. To identify the putative novel
carbapenemase gene, a shotgun cloning approach was taken. As experiments using genomic
DNA that was digested with HindIII, EcoRI, XhoI and BamHI did not yield any recombinant
clones, MboI was used for restriction. The partially digested genomic DNA was then ligated with
the BamHI-digested pBK-CMV vector, as the MboI and BamHI overhangs (GATC) are compatible.
Finally, the MboI experiments yielded a clone with increased resistance for carbapenems and the
4575-bp insert of the contained recombinant plasmid pMB3013 was sequenced using
oligonucleotides listed in Table 2.3. It harboured a 726-bp ORF that coded for a 241 amino acid
protein. On both nucleotide and protein level the sequences showed a homology of only 74.3 %

Results 60
Figure 3.7 Amino acid sequence alignment of KHM-2 and KHM-1. The residues known as subclass B1 zinc binding residues of are marked with asterisks. to blaKHM-1, coding for the metallo-β-lactamase KHM-1. The sequence of the novel blaKHM gene
was submitted to the international β-lactamase numbering institution and the encoded enzyme
was assigned as KHM-2. An alignment of the amino acid sequences of KHM-2 and KHM-1 is
shown in Figure 3.7. Compared to KHM-1, KHM-2 showed no alterations in the highly conserved
zinc binding residues, but exhibited a threonine to aspartic acid substitution at position 100,
which is part of the conserved HXHXD zinc binding motif. With only 74.3 % homology, KHM-2
showed one of the greatest distances to the next nearest relative within the Ambler subclass B1.
As KHM-2 and KHM-1 were the only members of the KHM group, no further phylogenetic
analysis was performed. Compared zo other subclass B1 enzymes, KHM-2 showed similarities of
only 54 % to IMP-1, 29 % to VIM-2 and 29 % to NDM-1.
To acquire more information on the isolate P. aeruginosa NRZ-03096 and to be able to classify
the isolate in an epidemiological context, the MLS type of the isolate was determined by
amplification and sequencing of seven P. aeruginosa housekeeping genes. Like for
P. aeruginosa NRZ-00156, the sequence types and the corresponding MLS type were determined
using the sequence definition tool of the P. aeruginosa MLST web site
(http://pubmlst.org/paeruginosa/). The allele types are summarized in Table 3.5 and showed
that P. aeruginosa NRZ-03096 expressed an unknown allelic profile. This was based on a point
mutation and two insertions in the 3´ region of the aroE gene. The closest match for the aroE
sequence type was type 5, resulting in MLST 395 beeing the nearest relative to the sequence
type expressed by P. aeruginosa NRZ-03096.
Table 3.5 MLS typing of P. aeruginosa NRZ-03096. Listed are the seven P. aeruginosa MLST housekeeping genes and the corresponding allele types (ST) of P. aeruginosa NRZ-03096. Parts of the results were obtained by Hoffmann (2013).
Gene acsA aroE guaA mutL nuoD ppsA trpE allele type 6 closest match: 5 1 1 1 12 1

Results 61
3.2 Analysis of the genetic environment of blaIMP-31, blaOXA-233 and blaKHM-2
The genetic environment of β-lactamase genes can have a significant influence on the gene
expression level and the ability of the gene to be horizontally transferred to other bacteria. To
identify genetic structures like integrons or transposable elements, the genetic context of the
three novel carbapenemase genes blaIMP-31, blaOXA-233 and blaKHM-2 was analyzed by cloning and
sequencing techniques.
3.2.1 Genetic environment of blaIMP-31
To further explore the genetic environment of the blaIMP-31 gene, a shotgun cloning approach was
performed. Shotgun cloning experiments with MboI finally yielded one single E. coli TOP10 clone
that showed increased resistance for carbapenems. The insert of the contained pBK-CMV
derivative plasmid pMB3011 was sequenced and the 2767-bp insert covered the whole blaIMP-31
ORF. In addition, it covered the neighboring blaOXA-10-like gene which was identified as blaOXA-35.
However, the shotgun cloning approach failed to provide significant additional information on
the surrounding regions, as apart from the two genes mentioned, the insert only covered 330 bp
of the 3´region of an intI1 gene upstream of the blaIMP-31 gene and 193 bp of a sequence with high
similarities to an aminoglycoside-acetyltransferase encoding gene downstream of the blaOXA-35
gene. To further analyze the genetic environment of blaIMP-31, a genome walking approach was
chosen. Using the Universal GenomeWalker 2.0 kit (Clontech), a DNA fragment with a size of
approximately 6 kb was amplified and sequenced using oligonucleotides listed in Table 2.3. By
combination of the sequences obtained from PCRs and genome walking it was possible to
assemble 4.8 kb of the genomic environment of the blaIMP-31 gene. A schematic of the genetic
environment of blaIMP-31 is shown in Figure 3.8. Sequence analysis showed that the gene was part
of a disrupted class 1 integron as the first gene cassette directly after the attI site. Further
downstream, gene cassettes containing blaOXA-35, aac(6’)-Ib, aac(3)-Ic and aphA15 genes were
identified. Downstream of the aphA15 gene cassette, the integron was disrupted by a
transposon-like structure, consisting of a tniC gene, which encodes for a site-specific
Figure 3.8 Genetic environment of blaIMP-31 in P. aeruginosa NRZ-00156. Conserved integron structures are shown in grey. The sequences of the integron promoters PcH2 and the inactive P2 are shown below and framed grey. The -35 and -10 boxes are marked with bold letters.

Results 62
recombinase and 525 bp of the tnpA gene, coding for the transposase A protein. Consequently,
the obtained sequences did not cover the full putative transposon and the missing 3´CS region of
the class 1 integron. The sequence was analyzed for direct and inverted repeats that could serve
as integration sites for the putative transposon, but as the transposon was not fully covered by
the obtained sequence, no repeat regions could be identified that could be associated to the
putative transposon. However, a potential repeat region with a high GC content and sequences of
multiple identical bases was identified downstream of the aphA15 gene. Analysis of the integron
promoter region revealed that the integron gene cassettes were expressed under the control of
the hybrid PcH2 promoter, consisting of the perfect -35 box TTGACA and the -10 box TAAGCT,
separated by a 17-bp spacer. The P2 promoter exhibited a 14-bp spacer region between the -35
box TTGTTA and the -10 box TACAGT and was missing the insertion of three guanine bases
which optimize the spacing in active P2 variants, resulting in a probably weak or inactive P2
promoter. 3.2.2 Genetic environment of blaOXA-233
The pBK-CMV derivative pMB3002 was obtained from shotgun cloning experiments and
harboured an insert with a size of 9102 bp. The insert was fully sequenced using
oligonucleotides listed in Table 2.3 and sequence analysis revealed that the blaOXA-233 gene was
part of a class 1 integron as the second gene cassette. Upstream, an aac(6´)-Ib gene was
identified, coding for an aminoglycoside-acetyltransferase. Downstream, the blaOXA-233 gene was
followed by the conserved 3´CS region of the integron, consisting of the genes qacEΔ1 and sul1. A
schematic of the genetic environment of blaOXA-233 is shown in Figure 3.9. In silico promoter
analysis revealed that the integron cassettes were under the control of a strong Pc promoter,
combined with an inactive P2 promoter. The Pc promoter exhibited the perfect -35 box TTGACA
and the -10 box TAAACT, resulting in a strong promoter. Like in the blaIMP-31 carrying integron,
the P2 promoter was inactive with a TTGTTA -35 box and a TACAGT -10 box that were
separated by only 14 spacing base pairs. In the sequence covered by the insert of pMB3002, no
Figure 3.9 Genetic environment of blaOXA-233 in C. freundii NRZ-02127. Conserved integron structures are shown in grey. The sequences of the integron promoters PcH2 and the inactive P2 are shown below and highlighted grey. The -35 and -10 boxes are marked with bold letters. Parts of this figure are based on results obtained by Meining (2012).

Results 63
transposon or IS structures were identified. Furthermore, no direct or inverted repeats flanking
the integron could be found.
3.2.3 Genetic environment of blaKHM-2
To acquire further information on the genetic environment of blaKHM-2, the 4575-bp insert of the
recombinant plasmid pMB3013 was fully sequenced using oligonucleotides listed in Table 2.3. A
schematic of the genetic environment is shown in Figure 3.10. Upstream of the blaKHM-2 gene, a
930-bp part of an ORF was identified. It showed 74 % identity to a gene coding for a putative
transposase of the ISXo2 family. Downstream of the blaKHM-2 gene, an ORF coding for a 262 amino
acid protein was identified. A BLAST homology search yielded a single hit that had an identity of
68 % to the putative gene. This sequence was annotated as an aac(3´) gene in the nucleotide
database of the National Centre for Biotechnology Information (NCBI) and was found in a
Gloeobacter violaceus whole genome sequence and consequently annotated to code for an
aminoglycoside-acetyltransferase. Downstream of the putative aac gene, the gene for a putative
insE family transposase was identified. As the region contained two putative transposase genes,
the sequence was analyzed for direct and inverted repeats. Upstream of the blaKHM-2 gene, a
palindromic sequence was identified (CCAATCATATTAATTGGATTGG) that could serve as an
insertion site for either the Isxo2 or InsE transposase, but no equivalent repeat was found in the
rest of the sequence covered by the pMB3013 insert. The rest of the sequence did not contain
any noticable repeat or inverted repeat regions.
In silico promoter analysis of the genetic environment revealed that the promoter of the blaKHM-2
gene was located 52 bp upstream of the ATG triplet and exhibited the -35 box TCGACA and the -
10 box AAATTA with a 17-bp spacing sequence. The sequence covered by the insert of pMB3013
did not contain any integron-like structures associated with the blaKHM-2 gene.
Figure 3.10 Genetic environment of blaKHM-2 in P. aeruginosa NRZ-03096. Putative transposon structure genes are shown as grey arrows. The sequences of the promoter structures upstream of the blaKHM-2 gene are shown below. The promoter of the blaKHM-2 gene is framed grey and the -35 and -10 boxes are marked with bold letters. The ATG triplet of the blaKHM-2 gene is marked with black framed bold letters. Parts of this figure are based on results obtained by Hoffmann (2013).

Results 64
As the shotgun cloning approach for KHM-2 was performed using MboI as restriction enzyme for
genomic DNA digestion and as digestion of DNA by MboI results in relatively small DNA
fragments, it was possible that the sequence of the insert was assembled by ligation of multiple
gDNA fragments from different regions of the DNA and did not represent the actual organization
in P. aeruginosa NRZ-03096. To verify that the arrangement shown in Figure 3.10 reflects the
actual arrangement in the isolate, PCRs that covered the 3`and 5`ends of neighboring ORFs
(isxO2/blaKHM-2; blaKHM-2/aac(3´)-like; aac(3´)-like/insE) were designed and performed with total
DNA from P. aeruginosa NRZ-03096 using oligonucleotides listed in Table 2.3. All used
combinations of oligonucleotides yielded PCR products of the expected size (data not shown)
and this was taken as verification, that the sequence arrangement reflected the actual state in
the isolate.
3.3 Localization of blaIMP-31, blaOXA-233 and blaKHM-2
Resistance genes can be chromosome- or plasmid-encoded. Plasmid-encoded genes can be
mobilized by conjugation of the plasmid, while chromosome-encoded genes can be mobilized by
transconjugable transposons, which is a less effective mechanism of gene distribution than
conjugative plasmids. In this context, it was analyzed if the novel carbapenemase genes
identified in this thesis were plasmid-encoded or if they were part of the chromosome of the
respective isolate.
3.3.1 Localization of blaIMP-31
To identify the localization of the IMP-31 encoding gene, total DNA from P. aeruginosa
NRZ-00156 was digested with nuclease S1 and I-CeuI and separated by PFGE. Nuclease S1 cuts
circular DNA molecules exactly once, leading to linearization of plasmids and chromosomes.
I-CeuI recognizes and digests a 26-bp sequence in bacterial rrn genes which code for the 23S
ribosomal subunit. As P. aeruginosa usually harbours four copies of the 23S rDNA that are
located exclusively on the chromosome, a digestion with I-CeuI yields four genomic DNA
fragments. As the 16S rDNA is also chromosome-located and neighbored to the 23S rDNA, each
of the fragments should contain a single copy of an intact 16S rDNA.
PFGE analysis after nuclease S1-digestion showed no detectable linearized plasmid bands and
indicated that the isolate did not harbour any plasmid that could be the carrier for blaIMP-31 (data
not shown). Consequently, it was suggested that the gene was chromosome-located. A digestion
with I-CeuI, followed by PFGE yielded four fragments with sizes of approximately 900 kb,
1,000 kb, 1,300 kb and 2,200 kb. The results are shown in Figure 3.11. Southern blotting and
hybridization with digoxigenin (DIG)-labeled DNA probes specific for blaIMP-31 and the 16S rDNA

Results 65
Figure 3.11 Localization of blaIMP-31. Total DNA of P. aeruginosa NRZ-00156 was digested with I-CeuI, separated by PFGE and subjected to Southern Blotting. Hybridization was performed with blaIMP-31 and 16S rDNA gene-specific probes. The figure shows the ethidiumbromide-stained PFGE-lanes of the size Marker (M), the total DNA (L) and the corresponding hybridized blots (IMP and 16S). Signals were detected using an Anti-DIG-AP-coupled antibody, the alkaline phosphatase substrate CSPD and autoradiography films. The blaIMP-31 signal and the corresponding 16S signal are indicated with an arrow.
was performed and with the 16S probe, four signals at the exact same size as in the EtBr-stained
gel were detected, corresponding to the four chromosome fragments. Hybridization with a
blaIMP-31 specific probe yielded a weak, but detectable signal at the size of the 2,200 kb fragment.
As the signals detected for blaIMP-31 and the 16S rDNA matched the 2,200 kb fragment detected in
PFGE, it was indicated that the blaIMP-31 gene was chromosome-located in P. aeruginosa
NRZ-00156.
3.3.2 Localization of blaOXA-233
As it is known that OXA-type carbapenemases are often plasmid-encoded, transconjugation
experiments were performed with the OXA-233 carrying isolate C. freundii NRZ-02127. Finally,
the experiments yielded a β-lactam resistant E. coli C600 clone which exhibited the same
resistance profile as the clinical isolate with increased resistance to carbapenems but
susceptibility to oxyimino-cephalosporins (Table 3.8) and was PCR-positive for blaOXA-233 (data
not shown). Both the clinical isolate and the transconjugant were subsequently analyzed by
nuclease S1 digestion and PFGE, followed by Southern blotting and hybridization with a
blaOXA-233 specific probe. The results of these experiments are shown in Figure 3.12. In PFGE
analysis, the isolate C. freundii NRZ-02127 showed three plasmid bands that had a size of
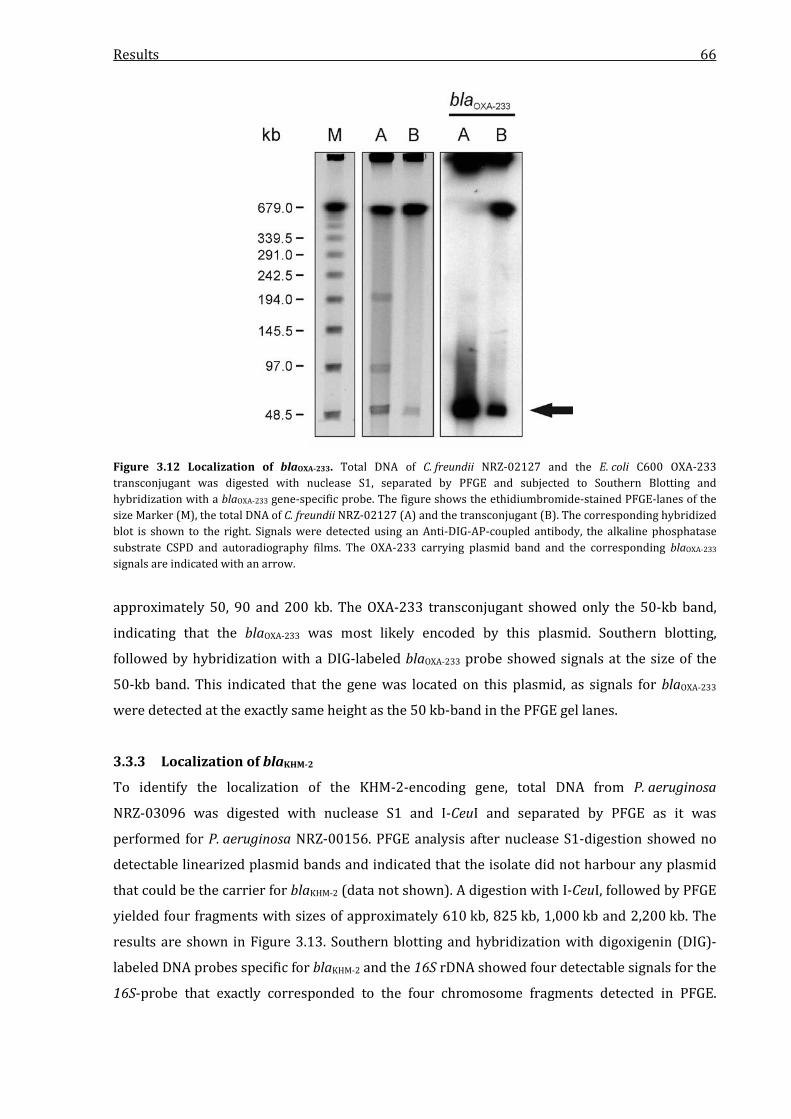
Results 66
Figure 3.12 Localization of blaOXA-233. Total DNA of C. freundii NRZ-02127 and the E. coli C600 OXA-233 transconjugant was digested with nuclease S1, separated by PFGE and subjected to Southern Blotting and hybridization with a blaOXA-233 gene-specific probe. The figure shows the ethidiumbromide-stained PFGE-lanes of the size Marker (M), the total DNA of C. freundii NRZ-02127 (A) and the transconjugant (B). The corresponding hybridized blot is shown to the right. Signals were detected using an Anti-DIG-AP-coupled antibody, the alkaline phosphatase substrate CSPD and autoradiography films. The OXA-233 carrying plasmid band and the corresponding blaOXA-233 signals are indicated with an arrow.
approximately 50, 90 and 200 kb. The OXA-233 transconjugant showed only the 50-kb band,
indicating that the blaOXA-233 was most likely encoded by this plasmid. Southern blotting,
followed by hybridization with a DIG-labeled blaOXA-233 probe showed signals at the size of the
50-kb band. This indicated that the gene was located on this plasmid, as signals for blaOXA-233
were detected at the exactly same height as the 50 kb-band in the PFGE gel lanes.
3.3.3 Localization of blaKHM-2
To identify the localization of the KHM-2-encoding gene, total DNA from P. aeruginosa
NRZ-03096 was digested with nuclease S1 and I-CeuI and separated by PFGE as it was
performed for P. aeruginosa NRZ-00156. PFGE analysis after nuclease S1-digestion showed no
detectable linearized plasmid bands and indicated that the isolate did not harbour any plasmid
that could be the carrier for blaKHM-2 (data not shown). A digestion with I-CeuI, followed by PFGE
yielded four fragments with sizes of approximately 610 kb, 825 kb, 1,000 kb and 2,200 kb. The
results are shown in Figure 3.13. Southern blotting and hybridization with digoxigenin (DIG)-
labeled DNA probes specific for blaKHM-2 and the 16S rDNA showed four detectable signals for the
16S-probe that exactly corresponded to the four chromosome fragments detected in PFGE.

Results 67
Figure 3.13 Localization of blaKHM-2. Total DNA of P. aeruginosa NRZ-03096 was digested with I-CeuI, separated by PFGE and subjected to Southern Blotting. Hybridization was performed with blaKHM-2 and 16S rDNA gene-specific probes. The figure shows the ethidiumbromide-stained PFGE-lanes of the size Marker (M), the total DNA (L) and the corresponding hybridized blots (KHM and 16S). Signals were detected using an Anti-DIG-AP-coupled antibody, the alkaline phosphatase substrate CSPD and autoradiography films. The blaKHM-2 signal and the corresponding 16S signal are indicated with an arrow.
Hybridization with a blaKHM-2 specific probe yielded detectable signal at the size of the 2,200 kb
fragment. As the signals detected for blaIMP-31 and the 16S rDNA matched the 2,200 kb fragment
detected in PFGE, it was indicated that the blaKHM-2 gene was chromosome-located in
P. aeruginosa NRZ-03096.
3.4 Impact of IMP-31, OXA-233 and KHM-2 on β-lactam resistance
To analyze the effect of expression of IMP-31, OXA-233 and KHM-2 on the resistance against
β-lactam antibiotics, the encoding genes were cloned into the pBK-CMV vector and the resulting
plasmids were transformed into E. coli TOP10. E. coli TOP10 is a K12 determinant that is lacking
a functional LacI protein due to a point mutation in the lacI gene. This results in a constitutive
expression of the lac operon and other genes that are under the control of a lac promoter. As the
expression of genes which are cloned into the MCS of the pBK-CMV vector is controlled by such a
promoter, these genes are constitutively expressed in E. coli TOP10. By determination of the
minimal inhibitory concentration (MIC) for various β-lactam antibiotics for E. coli TOP10
expressing the genes identified in this study, the influence of the production of IMP-31, OXA-233
and KHM-2 on β-lactam resistance was analyzed in relation to E. coli TOP10 carrying the empty
pBK-CMV vector and not producing a β-lactamase. Contrary to the MIC data for the clinical
isolates, the data were not interpreted according to the EUCAST criteria as these criteria are not
applicable to laboratory E. coli K12 determinant strains.

Results 68
3.4.1 Impact of IMP-31 on β-lactam resistance
To study the impact of production of IMP-31 on β-lactam resistance, the encoding gene was
cloned into the pBK-CMV vector, yielding the recombinant plasmid pMB3007. The plasmid was
then transformed into E. coli TOP10. The gene coding for the IMP reference enzyme IMP-1 was
also cloned into the pBK-CMV vector (yielding pMB3010) and transformed into E. coli TOP10 to
serve as a reference. E. coli TOP10 transformed with the pBK-CMV vector was used as a control.
The MICs obtained from these experiments are summarized in Table 3.6. Compared to the
control strain, IMP-31 producing E. coli TOP10 showed increased resistance to all tested β-
lactams. The MIC for ampicillin was increased over 10-fold from 1.5 mg/l to 16 mg/l and 12-fold
for ampicillin/sulbactam (1.0 to 12 mg/l). Piperacillin and piperacillin/tazobactam MICs were
only slightly increased from 1.0 mg/l to 3 mg/l and 0.75 mg/l to 3 mg/l, which corresponds to a
3-fold and 4-fold increase. Production of IMP-31 further resulted in a 128-fold increase in the
MIC for amoxicillin. The MIC for amoxicillin/clavulanate however was only increased 11-fold. Table 3.6 β-lactam MICs of E. coli TOP10 transformed with the pBK-CMV vector and IMP-31/IMP-1 expressing E. coli TOP10. MIC increases relative to the control strain (E. coli TOP10/pBK-CMV) are shown in parentheses.
MIC (mg/l)
Antibiotic E. coli TOP10/pBK-CMV
E. coli TOP10/pMB3007
IMP-31
E. coli TOP10/pMB3010
IMP-1
Ampicillin 1.5 16 (11×) 48 (32×)
Ampicillin-sulbactam 1.0 12 (12×) 48 (48×)
Piperacillin 1.0 3 (3×) 3 (3×)
Piperacillin-tazobactam 0.75 3 (4×) 3 (4×)
Amoxicillin 2 256 (128×) >256 (>128×)
Amoxicillin-clavulanate 1.5 16 (11×) 32 (21×)
Temocillin 8 16 (2×) 32 (4×)
Cephalotin 3 64 (21×) >256 (>85×)
Cefuroxime 2 64 (32×) >256 (>128×)
Cefoxitin 4 >256 (>64×) >256 (>64×)
Cefotaxime 0.032 3 (94×) 16 (500×)
Ceftriaxone 0.047 3 (64×) 32 (681×)
Cefepime 0.023 0.75 (32×) 3 (130×)
Ceftazidime 0.25 32 (128×) >256 (>1024×)
Imipenem 0.19 0.38 (2×) 1.5 (8×)
Meropenem 0.016 0.19 (12×) 0.75 (47×)
Doripenem 0.016 0.19 (12×) 0.5 (31×)
Ertapenem 0.006 0.19 (32×) 1.0 (167×)
Aztreonam 0.094 0.094 (1×) 0.094 (1×)

Results 69
Temocillin MICs were only 2-fold increased from 8 mg/l to 16 mg/l. The MIC increases for first
and second generation cephalosporins ranged from over 64-fold for cefoxitin (4 to >256 mg/l)
to 21-fold for cephalotin (3 to 64 mg/l). Third generation cephalosporin MICs were increased
94-fold for cefotaxime, 64-fold for ceftriaxone, 32-fold for cefepime and 128-fold for ceftazidime.
The MIC for imipenem was increased from 0.19 mg/l to 0.38 mg/l, which was only a 2-fold
increase, while the meropenem and doripenem MICs were both increased 12-fold from
0.016 mg/l to 0.19 mg/l. With a 32-fold increase, ertapenem showed the highest carbapenem
MIC elevation with values of 0.006 and 0.19 mg/l for the control strain and the IMP-31
producing strain, respectively. The expression of IMP-31 had no effect on the MIC for aztreonam.
Compared to the IMP-1 producing strain, the MICs of the IMP-31 producing E. coli TOP10 were
generally lower with only the MICs for piperacillin and piperacillin/tazobactam beeing 3 mg/l
for both strains. Production of IMP-1 led to MICs of 48 mg/l for ampicillin and
ampicillin/sulbactam, while the MIC for amoxicillin was similar to the IMP-31 strain. The
greatest differences between IMP-1 and IMP-31 were seen for the oxyimino-cephalosporins
cefotaxime, ceftriaxone and ceftazidime with MICs of 16 mg/l, 32 mg/l and >256 mg/l,
corresponding to a 500-fold, 681-fold and >1024-fold increase relative to the control strain.
Carbapenem MICs were elevated 8-fold for imipenem, 47-fold for meropenem, 31-fold for
doripenem and 167-fold for ertapenem, showing significantly higher MICs than IMP-31
producing E. coli TOP10. Like for IMP-31, the expression of IMP-1 had no effect on the MIC for
aztreonam. In general, the production of IMP-31 led to increased MICs for almost all tested β-
lactams, although production of the reference enzyme IMP-1 resulted in even higher MICs.
3.4.2 Impact of OXA-233 on β-lactam resistance
To analyze the impact of OXA-233 production on β-lactam resistance, the blaOXA-233 was cloned
into the pBK-CMV vector, yielding the recombinant plasmid pMB3006. To serve as a reference,
the blaOXA-10 gene was cloned the same way (yielding pMB3026). Both strains were analyzed in
MIC studies and the results are shown in Table 3.7. E. coli TOP10 cells transformed with the
pBK-CMV vector served as a control. The β-lactam MICs were also determined for the E. coli
C600 OXA-233 transconjugant that carried the plasmid pMB3018 from the clinical isolate. E. coli
C600 without any plasmid served as a control and the results are shown in Table 3.8.
MIC determination showed that the OXA-233 producing strain exhibited elevated β-lactam
resistance against most tested antibiotics. The pMB3006 harbouring strain showed an MIC of
>256 mg/l for ampicillin and amoxicillin, resulting in a more than 170-fold increase compared to
the control strain. For piperacillin and amoxicillin, MICs of 16 mg/l and >256 mg/l were
detected. β-lactam-inhibitor combinations led to significantly decreased MICs, indicating an
inhibition of OXA-233 by sulbactam, tazobactam and clavulanic acid. Regarding cephalosporins,
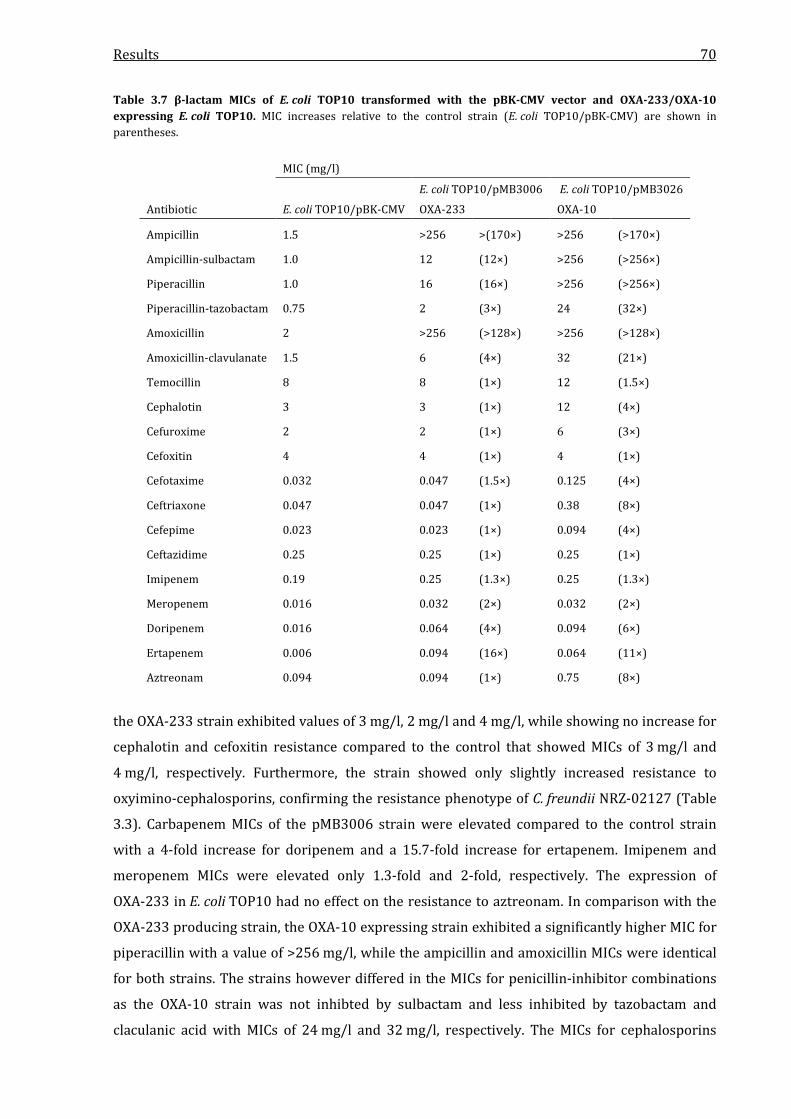
Results 70
Table 3.7 β-lactam MICs of E. coli TOP10 transformed with the pBK-CMV vector and OXA-233/OXA-10 expressing E. coli TOP10. MIC increases relative to the control strain (E. coli TOP10/pBK-CMV) are shown in parentheses.
MIC (mg/l)
Antibiotic E. coli TOP10/pBK-CMV
E. coli TOP10/pMB3006
OXA-233
E. coli TOP10/pMB3026
OXA-10
Ampicillin 1.5 >256 >(170×) >256 (>170×)
Ampicillin-sulbactam 1.0 12 (12×) >256 (>256×)
Piperacillin 1.0 16 (16×) >256 (>256×)
Piperacillin-tazobactam 0.75 2 (3×) 24 (32×)
Amoxicillin 2 >256 (>128×) >256 (>128×)
Amoxicillin-clavulanate 1.5 6 (4×) 32 (21×)
Temocillin 8 8 (1×) 12 (1.5×)
Cephalotin 3 3 (1×) 12 (4×)
Cefuroxime 2 2 (1×) 6 (3×)
Cefoxitin 4 4 (1×) 4 (1×)
Cefotaxime 0.032 0.047 (1.5×) 0.125 (4×)
Ceftriaxone 0.047 0.047 (1×) 0.38 (8×)
Cefepime 0.023 0.023 (1×) 0.094 (4×)
Ceftazidime 0.25 0.25 (1×) 0.25 (1×)
Imipenem 0.19 0.25 (1.3×) 0.25 (1.3×)
Meropenem 0.016 0.032 (2×) 0.032 (2×)
Doripenem 0.016 0.064 (4×) 0.094 (6×)
Ertapenem 0.006 0.094 (16×) 0.064 (11×)
Aztreonam 0.094 0.094 (1×) 0.75 (8×)
the OXA-233 strain exhibited values of 3 mg/l, 2 mg/l and 4 mg/l, while showing no increase for
cephalotin and cefoxitin resistance compared to the control that showed MICs of 3 mg/l and
4 mg/l, respectively. Furthermore, the strain showed only slightly increased resistance to
oxyimino-cephalosporins, confirming the resistance phenotype of C. freundii NRZ-02127 (Table
3.3). Carbapenem MICs of the pMB3006 strain were elevated compared to the control strain
with a 4-fold increase for doripenem and a 15.7-fold increase for ertapenem. Imipenem and
meropenem MICs were elevated only 1.3-fold and 2-fold, respectively. The expression of
OXA-233 in E. coli TOP10 had no effect on the resistance to aztreonam. In comparison with the
OXA-233 producing strain, the OXA-10 expressing strain exhibited a significantly higher MIC for
piperacillin with a value of >256 mg/l, while the ampicillin and amoxicillin MICs were identical
for both strains. The strains however differed in the MICs for penicillin-inhibitor combinations
as the OXA-10 strain was not inhibted by sulbactam and less inhibited by tazobactam and
claculanic acid with MICs of 24 mg/l and 32 mg/l, respectively. The MICs for cephalosporins

Results 71
were higher compared to the OXA-233 strain, although the increases were relatively low with
values of 0.125 mg/l for cefotaxime or 0.094 mg/l for cefepime. Surprisingly, the OXA-10 strain
showed nearly the same MICs for carbapenems as the OXA-233, which showed a 1.3-fold
increase for imipenem, a 2-fold increase for meropenem, a 4-fold increase for doripenem and a
16-fold increase for ertapenem. In contrast to OXA-233, the production of OXA-10 led to an MIC
increase for aztreonam from 0.094 mg/l to 0.75 mg/l. β-lactam MICs were also determined for
the OXA-233 transconjugant. It showed the same resistance profile as the clinical isolate and the
pMB3006 strain with high level resistance to penicillins that was inhibited by sulbactam,
tazobactam and clavulanic acid and susceptibility to oxyimino-cephalosporins. Like in the other
OXA-233 strain, carbapenem MICs were distinctly elevated except for imipenem, where the MIC
was not increased compared to the E. coli C600 control strain.
Table 3.8 β-lactam MICs of the E. coli C600 OXA-233 pMB3018-transconjugant and E. coli C600. MIC increases relative to the control strain (E. coli C600) are shown in parentheses.
MIC (mg/l)
Antibiotic E. coli C600 E. coli C600/pMB3018
Ampicillin 1.5 >256 (>170×)
Ampicillin-sulbactam 1.5 24 (16×)
Piperacillin 0.75 >256 (>340×)
Piperacillin-tazobactam 0.75 8 (11×)
Amoxicillin 3 >256 (>85×)
Amoxicillin-clavulanate 3 8 (3×)
Temocillin 3 12 (4×)
Cephalotin 3 8 (3×)
Cefuroxime 2 3 (2×)
Cefoxitin 2 4 (2×)
Cefotaxime 0.032 0.064 (2×)
Ceftriaxone 0.047 0.064 (1.4×)
Cefepime 0.016 0.023 (1.4×)
Ceftazidime 0.125 0.19 (1.5×)
Imipenem 0.19 0.19 (1×)
Meropenem 0.012 0.094 (8×)
Doripenem 0.023 0.19 (8×)
Ertapenem 0.004 0.19 (48×)
Aztreonam 0.047 0.064 (1.4×)
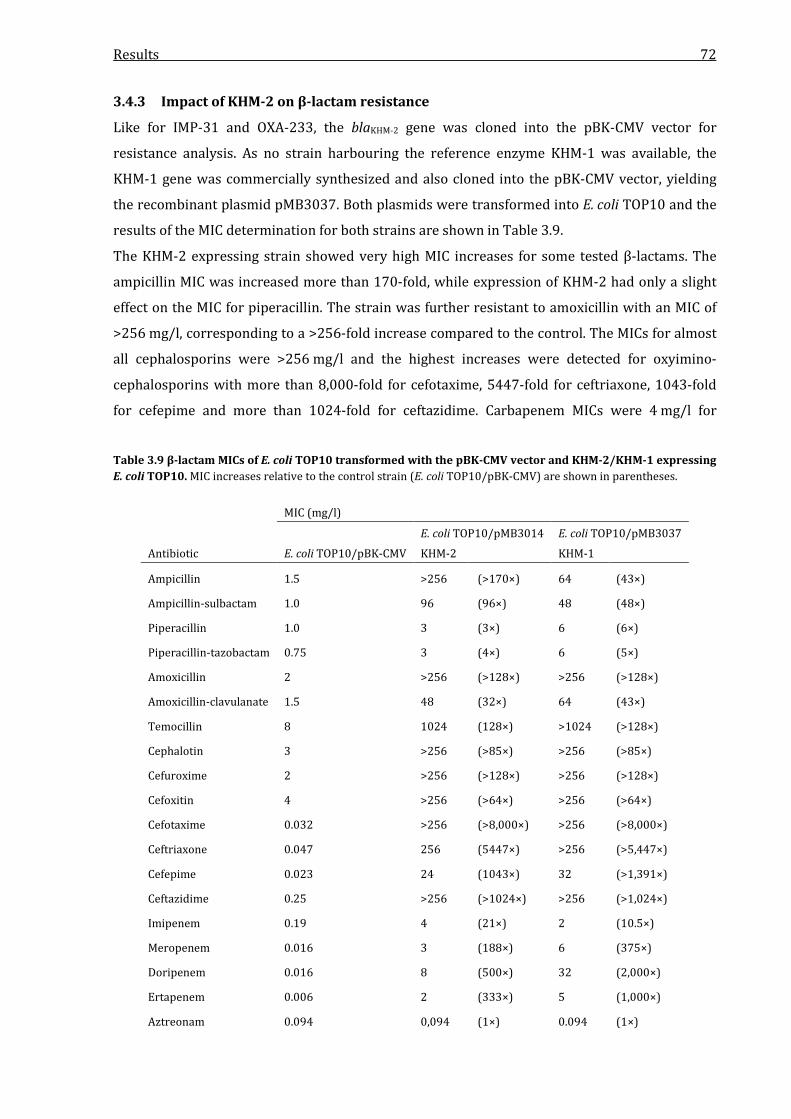
Results 72
3.4.3 Impact of KHM-2 on β-lactam resistance
Like for IMP-31 and OXA-233, the blaKHM-2 gene was cloned into the pBK-CMV vector for
resistance analysis. As no strain harbouring the reference enzyme KHM-1 was available, the
KHM-1 gene was commercially synthesized and also cloned into the pBK-CMV vector, yielding
the recombinant plasmid pMB3037. Both plasmids were transformed into E. coli TOP10 and the
results of the MIC determination for both strains are shown in Table 3.9.
The KHM-2 expressing strain showed very high MIC increases for some tested β-lactams. The
ampicillin MIC was increased more than 170-fold, while expression of KHM-2 had only a slight
effect on the MIC for piperacillin. The strain was further resistant to amoxicillin with an MIC of
>256 mg/l, corresponding to a >256-fold increase compared to the control. The MICs for almost
all cephalosporins were >256 mg/l and the highest increases were detected for oxyimino-
cephalosporins with more than 8,000-fold for cefotaxime, 5447-fold for ceftriaxone, 1043-fold
for cefepime and more than 1024-fold for ceftazidime. Carbapenem MICs were 4 mg/l for
Table 3.9 β-lactam MICs of E. coli TOP10 transformed with the pBK-CMV vector and KHM-2/KHM-1 expressing E. coli TOP10. MIC increases relative to the control strain (E. coli TOP10/pBK-CMV) are shown in parentheses.
MIC (mg/l)
Antibiotic E. coli TOP10/pBK-CMV
E. coli TOP10/pMB3014
KHM-2
E. coli TOP10/pMB3037
KHM-1
Ampicillin 1.5 >256 (>170×) 64 (43×)
Ampicillin-sulbactam 1.0 96 (96×) 48 (48×)
Piperacillin 1.0 3 (3×) 6 (6×)
Piperacillin-tazobactam 0.75 3 (4×) 6 (5×)
Amoxicillin 2 >256 (>128×) >256 (>128×)
Amoxicillin-clavulanate 1.5 48 (32×) 64 (43×)
Temocillin 8 1024 (128×) >1024 (>128×)
Cephalotin 3 >256 (>85×) >256 (>85×)
Cefuroxime 2 >256 (>128×) >256 (>128×)
Cefoxitin 4 >256 (>64×) >256 (>64×)
Cefotaxime 0.032 >256 (>8,000×) >256 (>8,000×)
Ceftriaxone 0.047 256 (5447×) >256 (>5,447×)
Cefepime 0.023 24 (1043×) 32 (>1,391×)
Ceftazidime 0.25 >256 (>1024×) >256 (>1,024×)
Imipenem 0.19 4 (21×) 2 (10.5×)
Meropenem 0.016 3 (188×) 6 (375×)
Doripenem 0.016 8 (500×) 32 (2,000×)
Ertapenem 0.006 2 (333×) 5 (1,000×)
Aztreonam 0.094 0,094 (1×) 0.094 (1×)

Results 73
imipenem, 3 mg/l for meropenem, 8 mg/l for doripenem and 2 mg/l for ertapenem,
corresponding to a 21-fold, 188-fold, 500-fold and 333-fold increase compared to the control
strain, respectively. The strain expressing the reference enzyme KHM-1 showed lower MIC for
ampicillin and ampicillin/sulbactam, while the MIC increases for penicillin, amoxicillin and
cephalosporins were mostly the same or equal. Regarding carbapenems, production of KHM-1
led to MICs of 2 mg/l for imipenem, 6 mg/l for meropenem, 32 mg/l for doripenem and 5 mg/l
for ertapenem. In comparison to the KHM-2 strain, resistance to imipenem was decreased, but
significantly higher for doripenem with a four-times higher MIC. Like for IMP-31 and IMP-1, the
expression of KHM-2 and KHM-1 showed no effect on the MIC for aztreonam.
3.4.4 Comparison of IMP-31, OXA-233 and KHM-2
A comparison of the relative MIC increases conferred by production of the three novel
carbapenemases identified in this study is shown in Table 3.10. Compared to each other, the
Table 3.10 Relative MIC increases of E. coli TOP10 producing IMP-31, OXA-233 and KHM-2. Shown are the MIC increases in relation to the control strain E. coli TOP10/pBK-CMV. The data are taken from Table 3.6, Table 3.7 and Table 3.8.
MIC increases relative to the control strain E. coli TOP10/pBK-CMV
Antibiotic
E. coli TOP10/pMB3007
IMP-31
E. coli TOP10/pMB3006
OXA-233
E. coli TOP10/pMB3014
KHM-2
Ampicillin 11× >170× >170×
Ampicillin-sulbactam 12× 12× 96×
Piperacillin 3× 16× 3×
Piperacillin-tazobactam 4× 3× 4×
Amoxicillin 128× >128× >128×
Amoxicillin-clavulanate 11× 4× 32×
Temocillin 2× 1× 128×
Cephalotin 21× 1× >85×
Cefuroxime 32× 1× >128×
Cefoxitin >64× 1× >64×
Cefotaxime 94× 1.5× >8,000×
Ceftriaxone 64× 1× 5447×
Cefepime 32× 1× 1043×
Ceftazidime 128× 1× >1024×
Imipenem 2× 1.3× 21×
Meropenem 12× 2× 188×
Doripenem 12× 4× 500×
Ertapenem 32× 16× 333×
Aztreonam 1× >170× 1×

Results 74
enzymes showed distinct differences in their impact on β-lactam resistance. Regarding
penicillins, production of both OXA-233 and KHM-2 led to significantly higher MICs for
ampicillin than production of IMP-31. Both MBL expressing strains showed only slightly
elevated MICs for piperacillin and piperacillin/tazobactam. The resistance of the OXA-233 strain
towards penicillins was clearly affected by all inhibitors, while the MBL strains were only
affected by clavulanic acid. The highest cephalosporin MIC increases were detected for the
KHM-2 strain, while the IMP-31 strain was also resistant but with lower total values. In contrast,
the production of OXA-233 had nearly no effect on most cephalosporin MICs. The weakest MIC
increases for carbapenems were also detected for the OXA-233 strain, as the IMP-31 strain
showed increases in resistance of 12 to 32-fold. The KHM-2 strain however showed the highest
detected carbapenem MIC increases with up to 500-fold increased resistance. In conclusion,
production of KHM-2 led to significantly higher β-lactam MICs than production of IMP-31, while
the OXA-233 strain overall showed the lowest MIC increases. Despite their differences,
production of all three enzymes led to increased MICs for carbapenems.
3.5 Purification of IMP-31, OXA-233 and KHM-2
In order to characterize the three novel carbapenemases IMP-31, OXA-233 and KHM-2
biochemically by determining the kinetic parameters Km and kcat in in vitro hydrolysis assays, the
enzymes and their respective reference enzymes (IMP-1, OXA-10 and KHM-2) had to be purified
at a high level. As overexpression experiments with His-tagged β-lactamases did yield very low
amounts of purified protein in other studies (Meining, 2012; Hoffmann, 2013; Lange, 2014), the
decision was made to purify the unmodified, native enzymes from larger culture volumes. To
acquire a satisfying amount of purified protein, cell extracts from a four-liter E. coli TOP10
culture that harboured one of the respective plasmids pMB3007 (IMP-31), pMB3010 (IMP-1),
pMB3006 (OXA-233), pMB3026 (OXA-10), pMB3014 (KHM-2) or pMB3037 (KHM-1) were
subjected to two chromatography steps. The first chromatography step for purification was an
ion exchange step, separating the respective expressed β-lactamase from other proteins with a
different isoelectric point (pI). As the resulting enzyme preperations still contained
contaminating proteins that had the same or similar pI, a gel filtration chromatography was
performed as the second purification step. The ion exchange fractions subjected to gel filtration
were chosen on the basis of nitrocefin hydrolysis. Nitrocefin is a chromogenic cephalosporin that
changes its color from yellow to red when hydrolyzed, enabling an easy detection of β-lactamase
activity in protein fractions. Typical chromatograms from the ion exchange and gel filtration
steps of the purification of KHM-2 are shown in Figure 3.14. Exemplary chromatograms for the
other five purified enzymes are shown in the appendix section of this study. In the ion exchange
chromatography of cell extracts from the KHM-2 producing E. coli TOP10 strain, the extract was

Results 75
applied to the column and high amounts of protein with up to 3,000 milli absorbance units
(mAU) were detected during the process. After washing, elution was started by increasing the
NaCl concentration with a linear gradient, resulting in changes of the surface charges and elution
of the bound protein. The proteins eluted in a single peak that corresponded to an absorbance
signal of 2,000 mAU. The corresponding fractions were analyzed for nitrocefin hydrolysis and
the ones containing the highest β-lactamase activity were pooled and subjected to gel filtration
chromatography. In this second purification step the proteins were seperated by size and a
Figure 3.14 Ion exchange (A) and gel filtration (B) chromatograms of the KHM-2 FPLC. Shown are the chromogram curves monitored during the chromatography process. The curves show the amount of detected milli absorbance units (mAU) relative to the volume passed through the columns. Red lines indicate the collected fractions. The ion exchange step for KHM-2 was performed at a pH of 7.5.

Results 76
single peak was detected after a volume of 67 ml with an absorbance signal of 683 mAU. The
corresponding fractions containing high amount of protein were again analyzed for nitrocefin
hydrolysis. Fractions showing high activity were pooled and subjected to SDS-PAGE analysis.
The results of these analyses are shown in Figure 3.15 for all six enzymes purified in this study.
SDS-PAGE analysis showed that all six enzymes were successfully purified with high purity
grades and the relative quantity was determined using the GelDox Xr+ software (Bio-Rad).
IMP-31 was detected as a band corresponding to a molecular weight of approximately 25 kDa. A
second band was detected at a lower weight, but had a clearly lower intensity and relative
quantification showed that the IMP-31 enzyme preparation was 85 % pure. IMP-1 was purified
near homogenity and was detected as a single band at a weight of approximately 25 kDa with a
Figure 3.15 SDS-PAGE analysis of enzyme preparations of IMP-31, IMP-1, OXA-233, OXA-10, KHM-2 and KHM-1. IMP-31 (A), IMP-1 (B), OXA-233 (C), OXA-10 (D), KHM-2 (E) and KHM-1 (F) were purified by ion exchange and gel filtration chromatography and 10 µl of the preparations were subjected to SDS-PAGE. Shown are images of the Coomassie-stained polyacrylamide gels after electrophoresis. The molecular weights of the size marker bands are stated in kDa.

Results 77
purity of 96 %. As the calculated molecular weight for IMP-31 and IMP-1 was 25.116 and
25.113 kDa, respectively, SDS-PAGE analysis showed that the purified proteins had the correct
weight. OXA-233 had a calculated weight of 27.571 kDa and was detected as a distinct band at a
weight of approximately 25 kDa, while the gel also exhibited a few other significantly smaller
bands of contaminating proteins. The calculated purity of the enzyme was 90 %. OXA-10 was
also purified near homogenity with a purity of 99 % and was shown as a single distinct band at a
weight of approximately 25 kDa, while the calculated weight was 27.550 kDa. SDS-PAGE analysis
of the KHM-2 enzyme preparation exhibited a clear band at a weight of 25 kDa. The preparation
was slightly contaminated by other proteins that showed weak bands and one protein with a
distinct band at a weight corresponding to 55 kDa. However, the calculated purity of the enzyme
preparation was 86 %. Like IMP-1 and OXA-10, KHM-1 was purified near homogenity with a
purity of 99 % and was detected as a single distinct band in SDS-PAGE at 25 kDa.
To acquire a sufficient amount of protein for in vitro hydrolysis assays, all six enzymes were
purified in triplicate. Determination of the protein concentration of the enzyme preparations
showed that more than 1 mg of purified enzyme was obtained for each preparation of IMP-31,
IMP-1, OXA-233, OXA-10, KHM-2 and KHM-1.
3.6 Determination of kinetic parameters
As β-lactams absorb light at ultraviolet wavelengths, it is possible to monitor the enzymatic
hydrolysis of these antibiotics spectrophotometrically and to perform Michaelis-Menten kinetics
with the obtained data. To analyze the biochemical characteristics of the novel carbapenemases
IMP-31, OXA-233 and KHM-2, the purified enzymes and their respective reference enzymes
were subjected to determination of the kinetic parameters Km and kcat. Km is the Michaelis
constant and an inverse indicator of the affinity of the substrate to the enzyme, while kcat is the
turnover number and specifies the amount of substrate molecules that are converted to product
per second. The quotient kcat/Km finally is an indicator for the hydrolytic efficiency of the
enzyme. By monitoring the absorbance changes in in vitro hydrolysis assays, these kinetic
parameters were determined for all six enzymes and various β-lactam antibiotics using
nonlinear regression. An example of an absorbance curve and a Michaelis-Menten plot is shown
in Figure 3.16.
3.6.1 Determination of kinetic parameters for IMP-31
The kinetic data obtained for IMP-31 and IMP-1 are summarized in Table 3.11 and showed that
IMP-31 and IMP-1 significantly differed in their hydrolytic activity. Regarding penicillins, IMP-31
showed kcat values of 81 s-1 for penicillin G, 6.6 s-1 for ampicillin and 0.6 s-1 for piperacillin, while
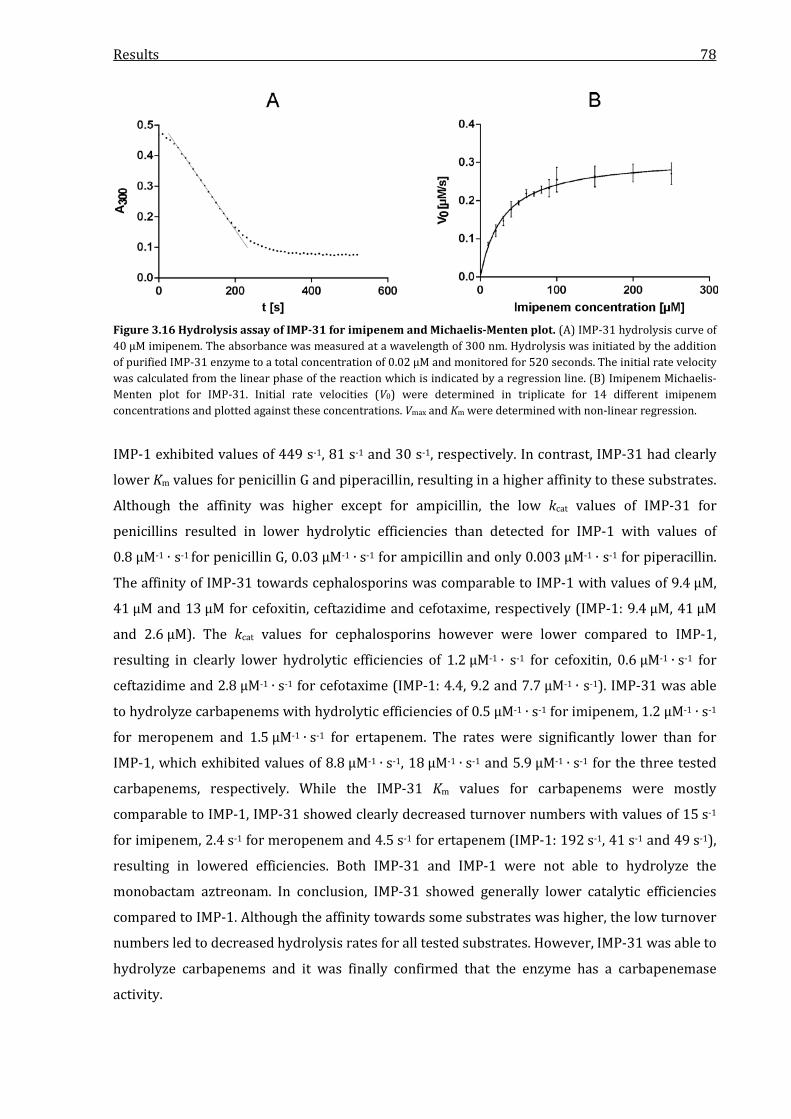
Results 78
Figure 3.16 Hydrolysis assay of IMP-31 for imipenem and Michaelis-Menten plot. (A) IMP-31 hydrolysis curve of 40 µM imipenem. The absorbance was measured at a wavelength of 300 nm. Hydrolysis was initiated by the addition of purified IMP-31 enzyme to a total concentration of 0.02 µM and monitored for 520 seconds. The initial rate velocity was calculated from the linear phase of the reaction which is indicated by a regression line. (B) Imipenem Michaelis-Menten plot for IMP-31. Initial rate velocities (V0) were determined in triplicate for 14 different imipenem concentrations and plotted against these concentrations. Vmax and Km were determined with non-linear regression.
IMP-1 exhibited values of 449 s-1, 81 s-1 and 30 s-1, respectively. In contrast, IMP-31 had clearly
lower Km values for penicillin G and piperacillin, resulting in a higher affinity to these substrates.
Although the affinity was higher except for ampicillin, the low kcat values of IMP-31 for
penicillins resulted in lower hydrolytic efficiencies than detected for IMP-1 with values of
0.8 µM-1 ∙ s-1 for penicillin G, 0.03 µM-1 ∙ s-1 for ampicillin and only 0.003 µM-1 ∙ s-1 for piperacillin.
The affinity of IMP-31 towards cephalosporins was comparable to IMP-1 with values of 9.4 µM,
41 µM and 13 µM for cefoxitin, ceftazidime and cefotaxime, respectively (IMP-1: 9.4 µM, 41 µM
and 2.6 µM). The kcat values for cephalosporins however were lower compared to IMP-1,
resulting in clearly lower hydrolytic efficiencies of 1.2 µM-1 ∙ s-1 for cefoxitin, 0.6 µM-1 ∙ s-1 for
ceftazidime and 2.8 µM-1 ∙ s-1 for cefotaxime (IMP-1: 4.4, 9.2 and 7.7 µM-1 ∙ s-1). IMP-31 was able
to hydrolyze carbapenems with hydrolytic efficiencies of 0.5 µM-1 ∙ s-1 for imipenem, 1.2 µM-1 ∙ s-1
for meropenem and 1.5 µM-1 ∙ s-1 for ertapenem. The rates were significantly lower than for
IMP-1, which exhibited values of 8.8 µM-1 ∙ s-1, 18 µM-1 ∙ s-1 and 5.9 µM-1 ∙ s-1 for the three tested
carbapenems, respectively. While the IMP-31 Km values for carbapenems were mostly
comparable to IMP-1, IMP-31 showed clearly decreased turnover numbers with values of 15 s-1
for imipenem, 2.4 s-1 for meropenem and 4.5 s-1 for ertapenem (IMP-1: 192 s-1, 41 s-1 and 49 s-1),
resulting in lowered efficiencies. Both IMP-31 and IMP-1 were not able to hydrolyze the
monobactam aztreonam. In conclusion, IMP-31 showed generally lower catalytic efficiencies
compared to IMP-1. Although the affinity towards some substrates was higher, the low turnover
numbers led to decreased hydrolysis rates for all tested substrates. However, IMP-31 was able to
hydrolyze carbapenems and it was finally confirmed that the enzyme has a carbapenemase
activity.
Results 79
Table 3.11 Kinetic parameters of IMP-31. The parameters were determined using nonlinear regression and the Michaelis-Menten equitation. Parameters for IMP-1 were determined to serve as a control and reference. Km values are shown in µM and kcat values in s-1. The experiments were performed at 25 °C.
IMP-31 IMP-1
Substrate kcat (s-1)a Km (µM)a kcat/Km (µM-1 ∙ s-1) kcat (s-1) Km (µM)
kcat/Km (µM-1 ∙ s-1)
Penicillin G 81 ± 4.1 108 ± 31 0.8 449 ± 34 256 ± 30 1.8 Ampicillin 6.6 ± 0.5 255 ± 23 0.03 81 ± 10 192 ± 51 0.4 Piperacillin 0.6 ± 0.09 185 ± 27 0.003 30 ± 2.3 684 ± 106 0.04 Cefoxitin 11 ± 0.8 9.4 ± 1.6 1.2 41 ± 0.3 9.4 ± 1.2 4.4 Ceftazidime 24 ± 1.2 41 ± 8.1 0.6 377 ± 0.9 41 ± 4.8 9.2 Cefotaxime 36 ± 4.1 13 ± 2.3 2.8 20 ± 0.4 2.6 ± 0.2 7.7 Imipenem 15 ± 1.5 30 ± 3.1 0.5 192 ± 26 22 ± 4.4 8.8 Meropenem 2.4 ± 1.0 2.0 ± 0.7 1.2 41 ± 7.2 2.3 ± 0.7 18 Ertapenem 4.5 ± 0.5 3.0 ± 0.4 1.5 49 ± 4.3 8.3 ± 2.1 5.9 Aztreonam NHb NH - NH NH -
a kcat and Km values represent the means of three independent experiments with three different enzyme preparations ± standard deviations. b NH, no hydrolysis was detected with a substrate concentration of up to 1 mM and an enzyme concentration of up to 200 nM.
3.6.2 Determination of kinetic parameters for OXA-233
As it has been shown that the active site serine of class D β-lactamases is carboxylated in vivo,
the determination of kinetic parameters for OXA-233 and OXA-10 was performed with sodium
bicarbonate as a CO2 source in the buffer used for hydrolysis assays. The kinetic data obtained
from these experiments are shown in Table 3.12. OXA-233 exhibited clearly lower turnover
numbers for penicillins than OXA-10, with kcat values of 39 s-1 for penicillin G, 343 s-1 and 117 s-1
for oxacillin, for which the kinetic parameters were determined instead of piperacillin due to the
presence of an OXA-type enzyme. As OXA-10 exhibited kcat values of 144 s-1 for penicillin G,
690 s-1 for ampicillin and 357 s-1 for oxacillin and as OXA-233 showed higher Km values, the
novel enzyme exhibited a lower hydrolytic efficiency for these substrates. Regarding
cephalosporins, the hydrolysis rates of OXA-233 for cefoxitin and ceftazidime were extremely
low. Although hydrolysis was detectable, the rate was too low to determine the kinetic
parameters, as even with extremely high enzyme concentrations of up to 200 nM, the initial rate
was not determinable from the monitored absorbance curves. OXA-10 on the other hand was
able to hydrolyze these cephalosporins with a low, but determinable rate of 0.003 µM-1 ∙ s-1. For
cefotaxime, OXA-233 showed a very low hydrolytic efficiency with a Km/kcat ratio of only
0.003 µM-1 ∙ s-1, while OXA-10 exhibited an over 10-fold increased value of 0.035 µM-1 ∙ s-1 for this
substrate. OXA-233 was able to hydrolyze carbapenems with hydrolysis rates of 0.075, 0.2 and
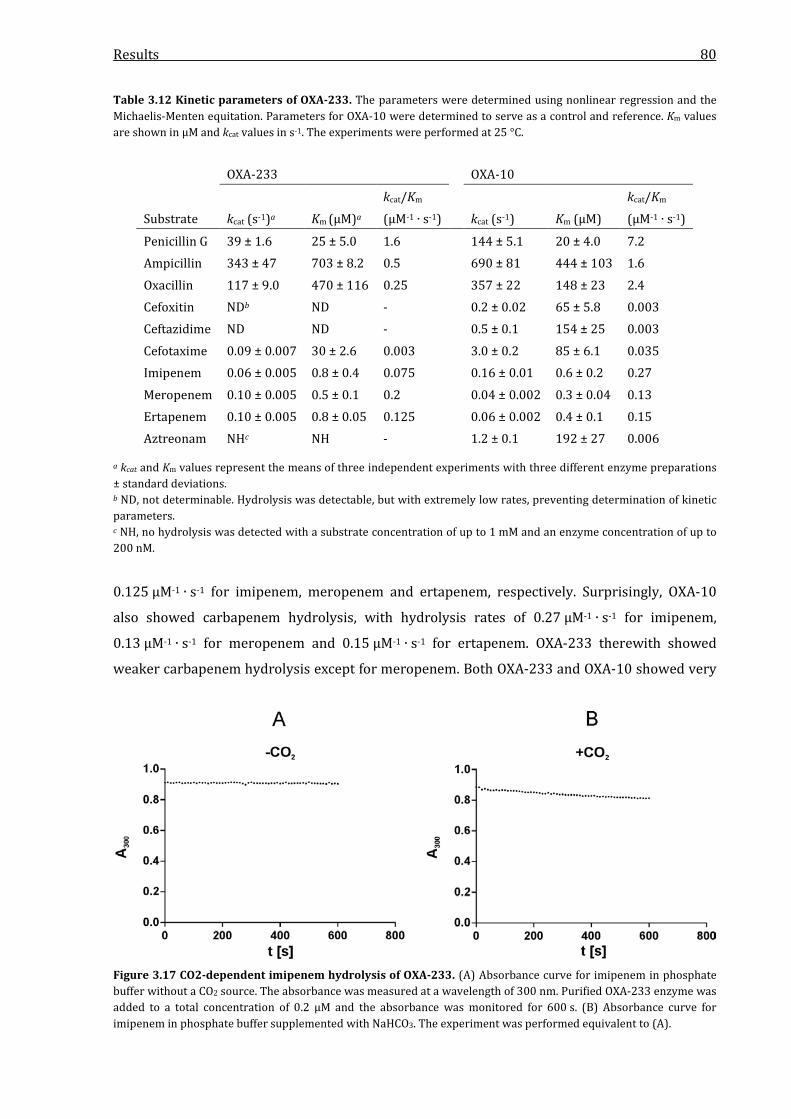
Results 80
Table 3.12 Kinetic parameters of OXA-233. The parameters were determined using nonlinear regression and the Michaelis-Menten equitation. Parameters for OXA-10 were determined to serve as a control and reference. Km values are shown in µM and kcat values in s-1. The experiments were performed at 25 °C.
OXA-233 OXA-10
Substrate kcat (s-1)a Km (µM)a kcat/Km (µM-1 ∙ s-1) kcat (s-1) Km (µM)
kcat/Km (µM-1 ∙ s-1)
Penicillin G 39 ± 1.6 25 ± 5.0 1.6 144 ± 5.1 20 ± 4.0 7.2 Ampicillin 343 ± 47 703 ± 8.2 0.5 690 ± 81 444 ± 103 1.6 Oxacillin 117 ± 9.0 470 ± 116 0.25 357 ± 22 148 ± 23 2.4 Cefoxitin NDb ND - 0.2 ± 0.02 65 ± 5.8 0.003 Ceftazidime ND ND - 0.5 ± 0.1 154 ± 25 0.003 Cefotaxime 0.09 ± 0.007 30 ± 2.6 0.003 3.0 ± 0.2 85 ± 6.1 0.035 Imipenem 0.06 ± 0.005 0.8 ± 0.4 0.075 0.16 ± 0.01 0.6 ± 0.2 0.27 Meropenem 0.10 ± 0.005 0.5 ± 0.1 0.2 0.04 ± 0.002 0.3 ± 0.04 0.13 Ertapenem 0.10 ± 0.005 0.8 ± 0.05 0.125 0.06 ± 0.002 0.4 ± 0.1 0.15 Aztreonam NHc NH - 1.2 ± 0.1 192 ± 27 0.006
a kcat and Km values represent the means of three independent experiments with three different enzyme preparations ± standard deviations. b ND, not determinable. Hydrolysis was detectable, but with extremely low rates, preventing determination of kinetic parameters. c NH, no hydrolysis was detected with a substrate concentration of up to 1 mM and an enzyme concentration of up to 200 nM.
0.125 µM-1 ∙ s-1 for imipenem, meropenem and ertapenem, respectively. Surprisingly, OXA-10
also showed carbapenem hydrolysis, with hydrolysis rates of 0.27 µM-1 ∙ s-1 for imipenem,
0.13 µM-1 ∙ s-1 for meropenem and 0.15 µM-1 ∙ s-1 for ertapenem. OXA-233 therewith showed
weaker carbapenem hydrolysis except for meropenem. Both OXA-233 and OXA-10 showed very
Figure 3.17 CO2-dependent imipenem hydrolysis of OXA-233. (A) Absorbance curve for imipenem in phosphate buffer without a CO2 source. The absorbance was measured at a wavelength of 300 nm. Purified OXA-233 enzyme was added to a total concentration of 0.2 µM and the absorbance was monitored for 600 s. (B) Absorbance curve for imipenem in phosphate buffer supplemented with NaHCO3. The experiment was performed equivalent to (A).
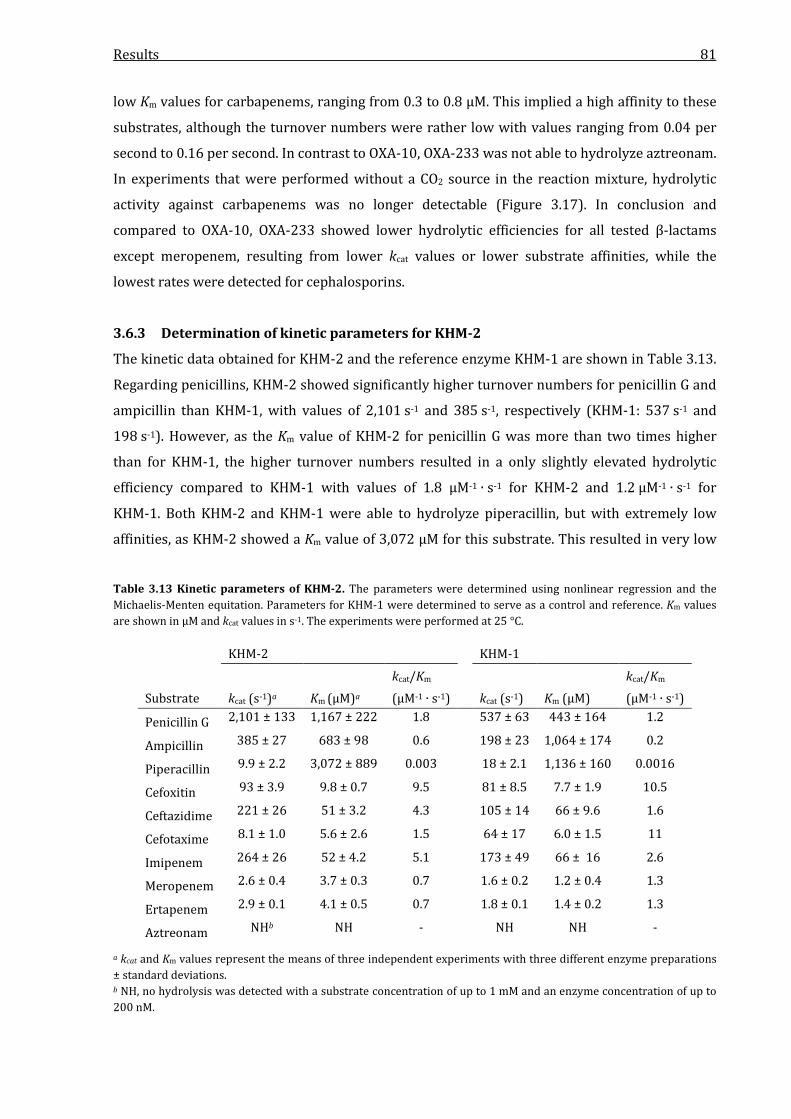
Results 81
low Km values for carbapenems, ranging from 0.3 to 0.8 µM. This implied a high affinity to these
substrates, although the turnover numbers were rather low with values ranging from 0.04 per
second to 0.16 per second. In contrast to OXA-10, OXA-233 was not able to hydrolyze aztreonam.
In experiments that were performed without a CO2 source in the reaction mixture, hydrolytic
activity against carbapenems was no longer detectable (Figure 3.17). In conclusion and
compared to OXA-10, OXA-233 showed lower hydrolytic efficiencies for all tested β-lactams
except meropenem, resulting from lower kcat values or lower substrate affinities, while the
lowest rates were detected for cephalosporins.
3.6.3 Determination of kinetic parameters for KHM-2
The kinetic data obtained for KHM-2 and the reference enzyme KHM-1 are shown in Table 3.13.
Regarding penicillins, KHM-2 showed significantly higher turnover numbers for penicillin G and
ampicillin than KHM-1, with values of 2,101 s-1 and 385 s-1, respectively (KHM-1: 537 s-1 and
198 s-1). However, as the Km value of KHM-2 for penicillin G was more than two times higher
than for KHM-1, the higher turnover numbers resulted in a only slightly elevated hydrolytic
efficiency compared to KHM-1 with values of 1.8 µM-1 ∙ s-1 for KHM-2 and 1.2 µM-1 ∙ s-1 for
KHM-1. Both KHM-2 and KHM-1 were able to hydrolyze piperacillin, but with extremely low
affinities, as KHM-2 showed a Km value of 3,072 µM for this substrate. This resulted in very low
Table 3.13 Kinetic parameters of KHM-2. The parameters were determined using nonlinear regression and the Michaelis-Menten equitation. Parameters for KHM-1 were determined to serve as a control and reference. Km values are shown in µM and kcat values in s-1. The experiments were performed at 25 °C.
KHM-2 KHM-1
Substrate kcat (s-1)a Km (µM)a kcat/Km (µM-1 ∙ s-1) kcat (s-1) Km (µM)
kcat/Km (µM-1 ∙ s-1)
Penicillin G 2,101 ± 133 1,167 ± 222 1.8 537 ± 63 443 ± 164 1.2
Ampicillin 385 ± 27 683 ± 98 0.6 198 ± 23 1,064 ± 174 0.2
Piperacillin 9.9 ± 2.2 3,072 ± 889 0.003 18 ± 2.1 1,136 ± 160 0.0016
Cefoxitin 93 ± 3.9 9.8 ± 0.7 9.5 81 ± 8.5 7.7 ± 1.9 10.5
Ceftazidime 221 ± 26 51 ± 3.2 4.3 105 ± 14 66 ± 9.6 1.6
Cefotaxime 8.1 ± 1.0 5.6 ± 2.6 1.5 64 ± 17 6.0 ± 1.5 11
Imipenem 264 ± 26 52 ± 4.2 5.1 173 ± 49 66 ± 16 2.6
Meropenem 2.6 ± 0.4 3.7 ± 0.3 0.7 1.6 ± 0.2 1.2 ± 0.4 1.3
Ertapenem 2.9 ± 0.1 4.1 ± 0.5 0.7 1.8 ± 0.1 1.4 ± 0.2 1.3
Aztreonam NHb NH - NH NH -
a kcat and Km values represent the means of three independent experiments with three different enzyme preparations ± standard deviations. b NH, no hydrolysis was detected with a substrate concentration of up to 1 mM and an enzyme concentration of up to 200 nM.

Results 82
hydrolytic efficiencies of 0.003 µM-1 ∙ s-1 and 0.0016 µM-1 ∙ s-1 for KHM-2 and KHM-1,
respectively. Both enzymes were able to hydrolyze cefoxitin and ceftazidime with similar kcat and
Km values, resulting in similar efficiencies. For cefotaxim, KHM-2 however showed a significantly
decreased activity, as the turnover number was eight times lower than for KHM-1 with a value of
8.1 s-1, while KHM-1 exhibited a value of 64 s-1. This resulted in a hydrolytic efficiency of only
1.5 µM-1 ∙ s-1, while KHM-1 was able to hydrolyze this substrate with a rate of 11 µM-1 ∙ s-1.
Regarding carbapenems, KHM-2 and KHM-1 clearly differed in their ability to hydrolyze
imipenem. KHM-2 exhibited a kcat value of 264 s-1 and a Km value of 52 µM, while KHM-1 showed
a turnover number of only 173 s-1 and a similar Km value of 66 µM, resulting in a two times
higher efficiency of KHM-2. Meropenem and ertapenem on the other hand were hydrolyzed with
similar rates of 0.7 µM-1 ∙ s-1 for KHM-2 and 1.3 µM-1 ∙ s-1 for KHM-1, as the kcat and Km values
were also relatively similar. Like the other metallo-β-lactamases characterized in this study,
both KHM-2 and KHM-1 were not able to hydrolyze the monobactam aztreonam. In conclusion,
KHM-2 showed a more efficient hydrolysis of penicillins than KHM-1, while cephalosporin
hydrolysis was similar except for cefotaxime. Both enzymes hydrolyzed carbapenems with
KHM-2 showing a two times higher hydrolytic efficiency against imipenem.
3.6.4 Comparison of the hydrolytic efficiencies of IMP-31, OXA-233 and KHM-2
The kinetic parameters of the three novel carbapenemases identified and characterized in this
study are listed comparatively in Table 3.14. Compared to each other, KHM-2 showed the
highest penicillin hydrolysis rates. While the rate for penicillin G was comparable to OXA-233
with values of 1.8 µM-1 ∙ s-1 and 1.6 µM-1 ∙ s-1, respectively, IMP-31 hydrolyzed this substrate with
a rate of 0.8 µM-1 ∙ s-1 and furthermore showed a significantly lower rate for ampicillin
(0.03 µM-1 ∙ s-1) than OXA-233 and KHM-1 (0.5 and 0.6 µM-1 ∙ s-1). Regarding cephalosporins,
KHM-2 showed the highest rates for cefoxitin and ceftazidime, while IMP-31 exhibited clearly
lower rates. OXA-233 was able to hydrolyze these antibiotics, but with extremely low hydrolyis
rates, preventing a determination of the kcat and Km parameters. For cefotaxime, hydrolysis rates
were determinable for all three enzymes and IMP-31 showed the highest rate with 2.8 µM-1 ∙ s-1,
while the rate for OXA-233 was again very low with 0.003 µM-1 ∙ s-1. The hydrolytic activity of
IMP-31, OXA-233 and KHM-2 against carbapenem antibiotics showed that OXA-233 was a rather
weak carbapenemase with rates of only 0.075 µM-1 ∙ s-1 for imipenem, 0.2 µM-1 ∙ s-1 for
meropenem and 0.125 µM-1 ∙ s-1 for ertapenem. Although IMP-31 also showed a relatively low
hydrolysis rate for imipenem (0.5 µM-1 ∙ s-1), the enzyme exhibited the highest detected rates for
meropenem and ertapenem with values of 1.2 µM-1 ∙ s-1 and 1.5 µM-1 ∙ s-1, respectively. Imipenem
on the other hand was most efficiently hydrolyzed by KHM-2 with a rate of 5.1 µM-1 ∙ s-1, while
the enzyme showed rates of 0.7 µM-1 ∙ s-1 for both meropenem and ertapenem. In conclusion, the
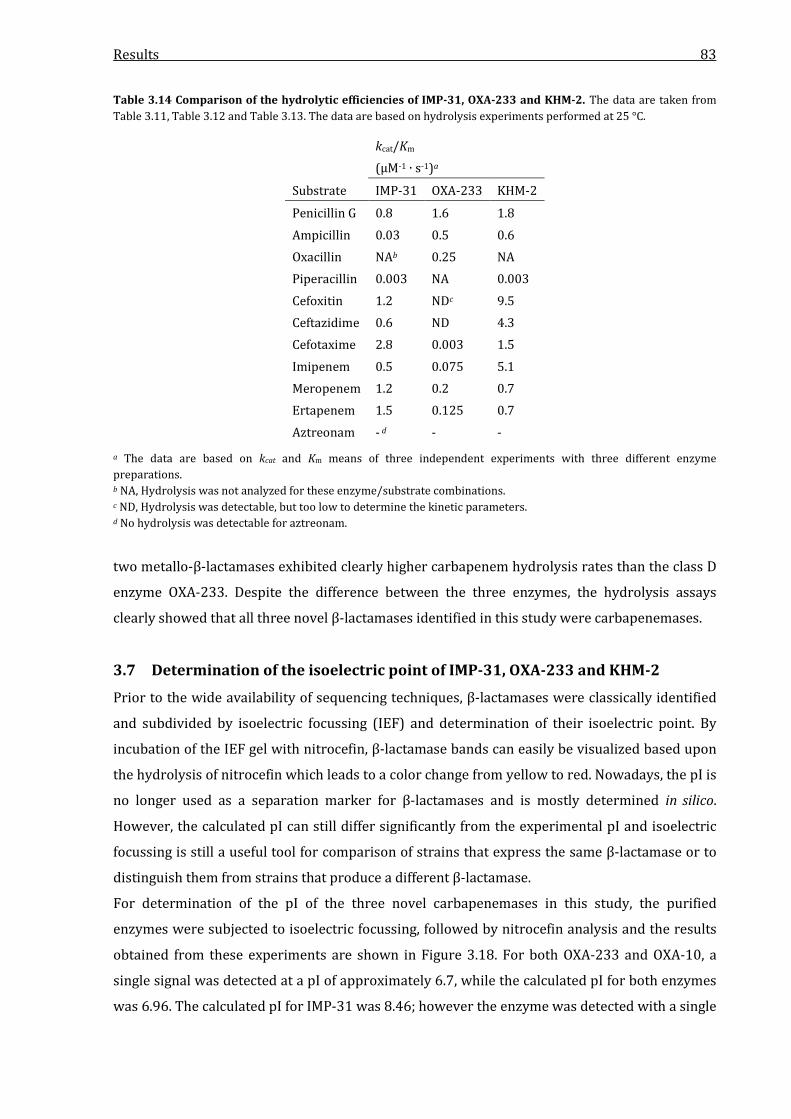
Results 83
Table 3.14 Comparison of the hydrolytic efficiencies of IMP-31, OXA-233 and KHM-2. The data are taken from Table 3.11, Table 3.12 and Table 3.13. The data are based on hydrolysis experiments performed at 25 °C.
kcat/Km (µM-1 ∙ s-1)a
Substrate IMP-31 OXA-233 KHM-2
Penicillin G 0.8 1.6 1.8 Ampicillin 0.03 0.5 0.6 Oxacillin NAb 0.25 NA Piperacillin 0.003 NA 0.003 Cefoxitin 1.2 NDc 9.5 Ceftazidime 0.6 ND 4.3 Cefotaxime 2.8 0.003 1.5 Imipenem 0.5 0.075 5.1 Meropenem 1.2 0.2 0.7 Ertapenem 1.5 0.125 0.7 Aztreonam - d - -
a The data are based on kcat and Km means of three independent experiments with three different enzyme preparations. b NA, Hydrolysis was not analyzed for these enzyme/substrate combinations. c ND, Hydrolysis was detectable, but too low to determine the kinetic parameters. d No hydrolysis was detectable for aztreonam.
two metallo-β-lactamases exhibited clearly higher carbapenem hydrolysis rates than the class D
enzyme OXA-233. Despite the difference between the three enzymes, the hydrolysis assays
clearly showed that all three novel β-lactamases identified in this study were carbapenemases.
3.7 Determination of the isoelectric point of IMP-31, OXA-233 and KHM-2
Prior to the wide availability of sequencing techniques, β-lactamases were classically identified
and subdivided by isoelectric focussing (IEF) and determination of their isoelectric point. By
incubation of the IEF gel with nitrocefin, β-lactamase bands can easily be visualized based upon
the hydrolysis of nitrocefin which leads to a color change from yellow to red. Nowadays, the pI is
no longer used as a separation marker for β-lactamases and is mostly determined in silico.
However, the calculated pI can still differ significantly from the experimental pI and isoelectric
focussing is still a useful tool for comparison of strains that express the same β-lactamase or to
distinguish them from strains that produce a different β-lactamase.
For determination of the pI of the three novel carbapenemases in this study, the purified
enzymes were subjected to isoelectric focussing, followed by nitrocefin analysis and the results
obtained from these experiments are shown in Figure 3.18. For both OXA-233 and OXA-10, a
single signal was detected at a pI of approximately 6.7, while the calculated pI for both enzymes
was 6.96. The calculated pI for IMP-31 was 8.46; however the enzyme was detected with a single
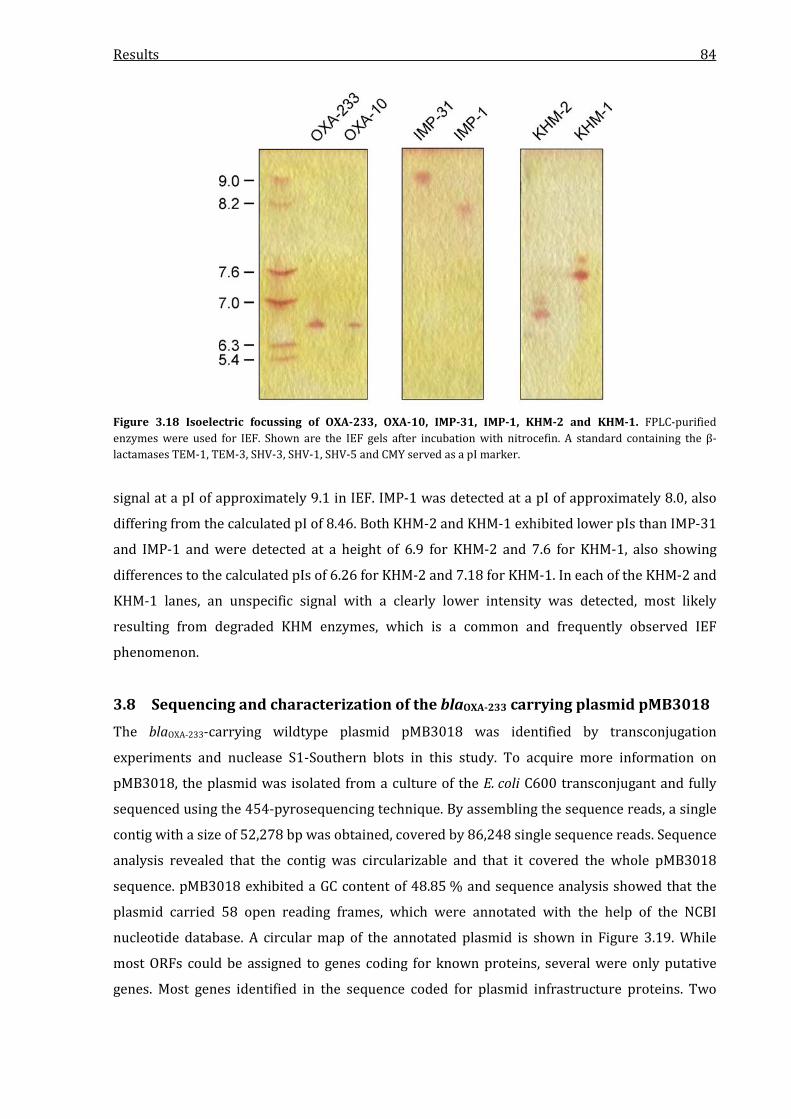
Results 84
Figure 3.18 Isoelectric focussing of OXA-233, OXA-10, IMP-31, IMP-1, KHM-2 and KHM-1. FPLC-purified enzymes were used for IEF. Shown are the IEF gels after incubation with nitrocefin. A standard containing the β-lactamases TEM-1, TEM-3, SHV-3, SHV-1, SHV-5 and CMY served as a pI marker.
signal at a pI of approximately 9.1 in IEF. IMP-1 was detected at a pI of approximately 8.0, also
differing from the calculated pI of 8.46. Both KHM-2 and KHM-1 exhibited lower pIs than IMP-31
and IMP-1 and were detected at a height of 6.9 for KHM-2 and 7.6 for KHM-1, also showing
differences to the calculated pIs of 6.26 for KHM-2 and 7.18 for KHM-1. In each of the KHM-2 and
KHM-1 lanes, an unspecific signal with a clearly lower intensity was detected, most likely
resulting from degraded KHM enzymes, which is a common and frequently observed IEF
phenomenon.
3.8 Sequencing and characterization of the blaOXA-233 carrying plasmid pMB3018
The blaOXA-233-carrying wildtype plasmid pMB3018 was identified by transconjugation
experiments and nuclease S1-Southern blots in this study. To acquire more information on
pMB3018, the plasmid was isolated from a culture of the E. coli C600 transconjugant and fully
sequenced using the 454-pyrosequencing technique. By assembling the sequence reads, a single
contig with a size of 52,278 bp was obtained, covered by 86,248 single sequence reads. Sequence
analysis revealed that the contig was circularizable and that it covered the whole pMB3018
sequence. pMB3018 exhibited a GC content of 48.85 % and sequence analysis showed that the
plasmid carried 58 open reading frames, which were annotated with the help of the NCBI
nucleotide database. A circular map of the annotated plasmid is shown in Figure 3.19. While
most ORFs could be assigned to genes coding for known proteins, several were only putative
genes. Most genes identified in the sequence coded for plasmid infrastructure proteins. Two

Results 85
Figure 3.19 Circular map of pMB3018. The outer circle displays the size in bp, the inner circle represents the GC content plotted against the average of the complete sequence with pale green indicating a GC content higher and purple indicating a GC content lower than the average of the total sequence. Genes are color-coded, depending on functional annotations: blue, conjugative transfer; orange, plasmid replication and maintenance; red, antimicrobial resistance; green, gene integration or transposition; and grey, putative functions or hypothetical proteins.
large transfer operons were identified, containing the tra genes A, B, C, D, E, F, G, H, I, J, K, L, M
and O, the endonuclease encoding gene nuc and the oriT region. The second locus consisted of
the tra genes K, J, I and the partially deleted fipA gene. Other genes carried by pMB3018 that play
a role in plasmid stability, antirestriction mechanisms, host range determination or regulation of
conjugation were stbA, B and C, ardR, B and K and kikA. The oriV region was identified at the
positions 17020 to 18035, containing a repA replicase gene. Regarding genes conferring
antibiotic resistance, the aac(6’), qacEΔ and sul genes carried by the blaOXA-233 integron were the
only ones identified on pMB3018. Beside two genes that code for putative phage integrases, two
transposase genes, IS6100 and ISSen4, were identified. A total of 17 ORFs identified in the

Results 86
sequence were not annotable as a BLAST search yielded only hits for hypothetical genes without
a predicted function.
As regions with significant differences in the GC content are often indicators for the insertion of
mobile elements, the sequence was analyzed for these regions. The results are visualized in
Figure 3.19 as the GC content plot. Two regions were identified that significantly differed from
the average. The first region consisted of the two ORFs with unknown function neighbored to
the ISSen4 transposase gene at positions 8253 to 11067. This region exhibited a GC content of
only 30.6 %, while the average of the plasmid was 48.85 % as noted before. The second region
consisted of the blaOXA-233 carrying integron and the neighboring IS6100 transposase gene with a
GC content of 56.6 %. The blaOXA-233 gene however had a GC content of only 42 % and if not taken
into consideration, the GC content of the region was 59 % and therewith more than 10 % higher
than the average of pMB3018. The sequence was furthermore analyzed for repeat regions as
these often serve as markers of a transposon or insertion sequence. Flanking the IS6100 gene,
two inverted repeat regions with a length of 123 bp each were identified, indicating the presence
of a transposon that consisted of only the transposase gene. Regarding the blaOXA-233-carrying
integron, a 13-bp inverted repeat region was identified upstream of the intI gene, with the
counterpart located downstream of the IS6100 transposase gene. No other repeat regions were
identified in the sequence which were correlatable to a potential insertion sequence or
transposon, especially not to the ISSen4 transposase gene. Replicon type analysis revealed that
the plasmid had no known replicon type, but was related to the IncN type. A BLAST homology
search revealed four plasmids with homologies to pMB3018, however they showed significant
differences. The closest relative was the NDM-1 carrying plasmid pJIE137 (accession number
NG_037697.1) with a coverage of 71 % and an identity of 95 %. The next relatives with high
homology scores were pECS01 (accession number KJ413946.1) and pTR3 (accession number
JQ349086.2), both showing a coverage of 62 % and an identity of 97 %. p271A (accession
number JF785549.1) finally was the fourth relative with acoverage of 52 % and an identity of
97 %. A schematic comparison of pMB3018 and the four other plasmids is shown in Figure 3.20.
Using the progressiveMauve algorithm, ten regions with high homologies were identified in
pMB3018 that were also found in one or more of the related plasmids. The regions found in all
five plasmids were the two tra regions, including their neighbored genes kikA, ΔfipA and the
stbACB genes and the oriV region including the repA and ardK genes. The region containing the
genes ardR, ardB, ccgC and mpr were also found in pJIE137, pECS01 and pTR3. In p271A, only a
part of this region including the mpr gene was identified. The 6-kb region containing the ISSen4
transposase gene that was part of p271A, pECS01 and pTR3 was shortened in pMB3018,
although the ISSen4 gene was still present. The pMB3018 region from 8252 to 16368 bp which
contained the two phage integrase genes and the three neighbored putative genes was not found
in any of the four related plasmids.

Results 87
Figu
re 3
.20
Com
pari
son
of p
MB3
018,
pJI
E137
, p27
1A, p
ECS0
1 an
d pT
R3. T
he a
lignm
ent w
as g
ener
ated
usi
ng th
e pr
ogre
ssiv
eMau
ve a
lgor
ithm
. Reg
ions
with
hig
h ho
mol
ogie
s ar
e co
lore
d an
d si
mila
ritie
s ar
e in
dica
ted
with
a h
omol
ogy
plot
. The
ann
otat
ed g
enes
cor
resp
ondi
ng to
the
iden
tifie
d re
gion
s of p
MB3
018
are
show
n ab
ove.

Results 88
Among the four plasmids related to pMB3018, only pJIE137 exhibited regions with similarities
to the resistance gene region of pMB3018. Analysis showed that pJIE137 also carried a class 1
integron and the regions that showed high homologies to pMB3018 were the ones coding for the
conserved integron genes intI1, qacEΔ1 and sul1. In addition, pJIE137 showed a homology to the
IS6100 transposase gene identified in pMB3018. In conclusion, the differences found in this
analysis showed the distinct differences of pMB3018 in comparison to other known IncN-related
plasmids.

Discussion 89
4 Discussion
The ongoing spread and diversification of carbapenemases in Gram-negative pathogens is one of
the most urgent problems for antimicrobial therapy of healthcare-associated infections. As these
β-lactamases can significantly differ in their ability to hydrolyze different β-lactam antibiotics,
the identification and characterization of these enzymes is crucial for clinical diagnostics and
correct antimicrobial treatment. In this study, three carbapenem-resistant Gram-negative
clinical isolates from patients hospitalized in Germany were analyzed for the presence of a novel
carbapenemase.
4.1 Identification of IMP-31
The isolate P. aeruginosa NRZ-00156 showed a carbapenemase phenotype, as the modified
Hodge Test indicated secretion of a carbapenemase into the medium, although this has been
described as rather based on leakage than secretion (Livermore, 1995). As carbapenem-
resistant clinical P. aeruginosa isolates are often carriers of metallo-β-lactamases (Walsh, 2010;
Diene & Rolain, 2014), the isolate was analyzed by an EDTA-CDT. The inhibition of carbapenem
resistance by EDTA clearly indicated the presence of an MBL, as these enzymes require one or
two zinc ions to perform the nucleophilic attack on the β-lactam ring. It has to be noted, that
EDTA itself also has an inhibitory effect on cell growth, as seen for the blank disk with EDTA.
This is also based on the chelating characteristics of EDTA, making free metal ions unavailable
for the bacteria (Root et al., 1988). However, the inhibitory effect of EDTA is considerably
weaker than growth inhibition by carbapenem antibiotics.
β-lactam MICs of the isolate were determined and clearly showed increased resistance to
carbapenems. Although carbapenem resistance can be based upon other mechanisms, such as
loss of the OprD porin, an MIC of >32 mg/l for all tested carbapenems further indicated the
presence of a carbapenemase, as mutation or loss of porin very rarely leads to carbapenem MICs
higher than 32 mg/l (Livermore, 2001). In contrast to MICs for other penicillins and
cephalosporins, the isolate did not show elevated MICs for piperacillin. This has been shown for
many metallo-β-lactamases, although it has not been described as a typical characteristic (Laraki
et al., 1999; Franceschini et al., 2000; Poirel et al., 2000; Cornaglia et al., 2011; Yong et al., 2012).
Consequently, the lower piperacillin MIC was only a hint for a potential MBL presence. The
susceptibility towards aztreonam finally was another indicator for an MBL production, as these
enzymes are not able to hydrolyze this antibiotic (Cornaglia et al., 2011).
The identification of the blaIMP-31 gene in the isolate by PCR and sequencing was surprising, as
the same PCR for blaIMP genes was performed prior to this study in the routine diagnostic

Discussion 90
process. It remains unclear why the gene was not detected in clinical diagnostics, but the most
likely explanation would be an insufficient quality of the DNA used for PCR analysis.
IMP-type (IMP for “active on imipenem”) carbapenemases show a continuous worldwide spread
among almost all Gram-negative pathogens, but are mostly found in P. aeruginosa and
A. baumannii isolates (Zhao & Hu, 2011). The first IMP-type enzyme was identified in 1988 in a
P. aeruginosa strain from Japan (Watanabe et al., 1991) and since then, blaIMP genes have been
increasingly detected wordwide (Zhao & Hu, 2011). In Europe, IMP-type carbapenemases have
been reported from Austria, Italy, the Czech Republic, France, the UK, Slovakia and Germany
(Riccio et al., 2000; Tysall et al., 2002; Neuwirth et al., 2007; Ohlasova et al., 2007; Duljasz et al.,
2009; Nemec et al., 2010; Pournaras et al., 2013). To date, 50 unique IMP-type enzymes have
been assigned (http://www.lahey.org/studies/), showing a high diversity of their amino acid
sequences with up to 22 % differences. At the time of the discovery of IMP-31, the enzyme
showed a very high diversity towards all other known IMP-type enzymes with only 84.1 %
homology to the next nearest relative IMP-8. The greatest distance was found in comparison to
IMP-38 to which IMP-31 showed 54 single amino acid substitutions, resulting in a homology of
only 78.2 %, which is currently the highest diversity between any IMP-type enzymes. In
addition, IMP-31 was the most distant enzyme compared to the reference enzyme IMP-1 with a
homology of only 80.0 %. However, the description of IMP-35 shortly later revealed a closer
relative with a homology of 96.7 %. Unlike IMP-35 and most other IMP variants that consist of
246 amino acid residues, IMP-31 was a 245 amino acid protein, shortened by one C-terminal
residue. In general, the C-terminus of IMP-31 was significantly different from those of other
IMP-type enzymes. For most IMP-type enzymes the C-terminus is formed by a KKPSXPSN motif,
with the first two lysine residues being conserved in all known IMP variants. For IMP-31 the C-
terminal sequence was KNHHSPK, making the C-terminus of the enzyme the most divergent
compared to all other IMP-type metallo-β-lactamases. Although relatively highly conserved, the
function of the C-terminus of IMP-type enzymes is still unknown, so it can only be speculated
about the influence of the altered C-terminus. For serine-β-lactamases, it is thought that they
originate from penicillin-binding-proteins and they share several structure similarities (Kelly et
al., 1986). As the C-terminus of PBPs is involved in interaction with the membranes (Harris et al.,
2002), the C-terminus of serine-β-lactamase could have the same function. However, MBLs
represent a completely different class of enzymes, as they do not belong to the SxxK
acetyltransferases superfamiliy like serine-β-lactamases and are part of their own superfamily
of metallo-enzymes (Cornaglia et al., 2011). Consequently, nothing equivalent is known for the
C-terminus of MBLs. As the signal peptide for the periplasmatic localization of β-lactamases is
located at the N-terminus (Pradel et al., 2009), it is furthermore unlikely that the altered C-
terminus of IMP-1 has a direct influence on secretion of the enzyme.

Discussion 91
The phylogenetic analysis of IMP-31 and the clustering of all IMP-type enzymes into thirteen
groups underlined the growing evolutionary complexity of the IMP family. The phylogenetic tree
showed IMP-31 to be a member of a new phylogenetic cluster that consists only of IMP-31 and
IMP-35 and is the most distant cluster of the IMP family. With a greater evolutionary distance to
a putative common ancestor even greater than IMP-35, IMP-31 further illustrated the immense
diversity of IMP-type carbapenemases and IMP-31 in particular. As IMP-35 was found in a
P. aeruginosa isolate in the Dutch-German border region and as the IMP-31 carrying strain was
referred to the National Reference Laboratory for Gram-negative pathogens from a clinical
diagnostics lab from North-Rhine-Westphalia, it can be assumed that the IMP-31 cluster has
established itself in Western Germany. Furthermore, the isolate P. aeruginosa NRZ-00156
expressed the allelic profile ST235, which has been frequently reported as a carrier of different
carbapenemases, e.g. IMP-1, IMP-6, VIM-2, GES-6 or PER-1 (Libisch et al., 2008; Seok et al., 2011;
Sardelic et al., 2012; Botelho et al., 2015; Shimizu et al., 2015). ST235 is an international
P. aeruginosa high-risk clone which belongs to the clonal complex CC235 (Maatallah et al., 2011).
CC235 has been frequently reported in context with the production of several metallo-β-
lactamases and ESBLs like IMP-7, VIM-4 or GES-1 in European countries (Nemec et al., 2010;
Samuelsen et al., 2010; Larché et al., 2012). The identification of IMP-31 in such a sequence type
is a strong indicator for a potential spread of this strain and the IMP-31 MBL in healthcare
settings.
With a GC content of ~39 %, it is very clear that all IMP variants are introduced into the the
species P. aeruginosa, which has a GC content of ~66 %, but the source of these genes is still
unknown. It can be assumed that IMP genes originate from intrinsic genes of one or several
closely related environmental bacterial species, although these progenitors are hitherto
unknown, unlike the progenitors of e.g. CTX-M, OXA-48 or OXA-23 (Poirel et al., 2004a; Poirel et
al., 2008; Cantón et al., 2012b). Due to the high diversity of IMP-31 and IMP-35 compared to
other variants it can be speculated, that they rather represent a de novo mobilization from the
unknown environmental source than an evolutionary diversification from other IMP-type
enzymes within P. aeruginosa. This would further imply the danger of the continuing
introduction of novel resistance genes into bacterial species of medical importance.
Most metallo-β-lactamase genes and all blaIMP genes in particular were found within integron
structures (Zhao & Hu, 2011). This was also the case for blaIMP-31. The gene showed a genetic
environment very similar to blaIMP-35, which also was the first gene cassette in a class 1 integron,
followed by blaOXA-35 and aac(6’)-Ib genes (Pournaras et al., 2013). However, the sequence
downstream of the aac(6’)-Ib gene showed major differences to the blaIMP-35-carrying integron
with two additional resistance genes, aac(3)-Ic and aphA15, which were cassettes of the
integron. A comparison of the genetic environments is shown in Figure 4.1. The blaIMP-31 integron

Discussion 92
Figure 4.1 Comparison of the genetic environment of blaIMP-31 and blaIMP-35. Grey boxes indicate similar sequences. The figure for the blaIMP-35 genetic environment was obtained and modified from Pournaras et al. (2013).
was furthermore disrupted by the transposon-associated genes tniC and tnpA, encoding a
recombinase of the invertase/resolvase family (Radstrom et al., 1994) and the transposase A.
The disruption of a class 1 integron by the tniC gene has been described for a blaVIM-2-carrying
putative transposon that is related to the worldwide dispersed Tn5090/Tn402 transposon,
which is proposed to be the progenitor of the common class 1 type of integrons (Radstrom et al.,
1994; Toleman et al., 2007; Moyo et al., 2015). Although Tn5090/Tn402 consists of the
additional genes orf6, tniB and tniA downstream of the tniC gene, which all code for
transposition enzymes, the presence of tniC is sufficient for transposition (Toleman et al., 2007).
This strongly suggests a possible transposon-mediated mobilization of blaIMP-31, as it has also
been shown for a Tn5090/Tn402-associated blaVIM-1-carrying integron (Tato et al., 2010). As the
sequence obtained by PCRs and genome walking did not cover the whole putative transposon
structure, for example the inverted repeat region that would be located upstream of the intI
gene, it was however not definitely possible to determine the functionality of the putative
transposon. In addition, it is not clear if the transposon-like structure only disrupted the
integron or if the conserved 3’CS end has been completely deleted and replaced by the
transposon elements, as it has been shown for a VIM-2 carrying class 1 integron (Samuelsen et
al., 2009). In contrast, the blaIMP-35-carrying integron was not disrupted by such elements. The
differing gene arrangement of the genetic environments of the two MBL genes however is
difficult to explain. Integron gene cassettes are always integrated next to the attI site and the
gene arrangement correlates with the chronological order of integration. This means that the
aphA15 and aac(3)-Ic gene cassettes of the blaIMP-31-carrying integron were the first gene
cassettes that were integrated. Subsequently, the aac(6’)-Ib, blaOXA-35 and blaIMP-31 genes must
have been integrated in this order and the exact same genetic events must have been occurred in
another integron with the only difference beeing the integration of a very closely related blaIMP
gene. In conclusion, this implies that it is rather unlikely that IMP-31 and IMP-35 originated
directly from each other or that one of the encoding integron-bourne genes simply mutated
while integrated. It is more likely that both genes represent two separate integration events;

Discussion 93
however the exact same order of the two following genes is still remarkable. In addition, the
blaIMP-35-harbouring isolate expressed another sequence type, ST622, although this sequence
type is very closely related to ST235 (Pournaras et al., 2013). This further implied that the two
genes did not originate directly from each other and possibly represent two separate de novo
mobilizations from a still unknown source of IMP-type carbapenemase genes.
Expression of the blaIMP-31 gene cassette and the following cassettes of the disrupted integron
was controlled by a typical integron promoter structure. Integron promoters are part of the
integrase gene and the attI site and several types of promoters have been identified and
classified on the basis of their impact on gene cassette expression. The promoter found in the
blaIMP-31-carrying integron was the Pc hybrid 2 promoter (PcH2), consisting of the strong -35
region TTGACA and the weaker -10 box TAAGCT (Papagiannitsis et al., 2009). This combination
has been identified as a weaker promoter than the perfect strong integron promoter, which
exhibits the -10 box TAAACT instead (Collis & Hall, 1995). It has been shown that expression of
genes controlled by the PcH2 variant is reduced almost 4-fold in comparison to the strong
variant (Papagiannitsis et al., 2009). The P2 promoter was identified directly adjacent to the intI
gene and was missing the insertion of three guanine bases. In the active variant of P2, this
insertion leads to an optimization of the spacing between the -35 and -10 boxes to 17 bp. A lack
of this insertion leads to an inactivation of the promoter (Collis & Hall, 1995). An active P2
promoter has been described as a compensator for a weak or hybrid Pc promoter
(Papagiannitsis et al., 2009), but this was absent in the present integron. Consequently, it can be
assumed that the expression of the IMP-31-encoding gene was not at the highest possible level.
As the isolate showed high carbapenem resistance that was exclusively reversible by inhibition
with EDTA (and thus showed the significant contribution of IMP-31 towards resistance) an
expression sufficient to confer high level resistance was indicated.
Contrary to many other acquired MBL genes that are plasmid-encoded (Walsh et al., 2005),
Southern blot experiments showed that the blaIMP-31 gene was located on the bacterial
chromosome. A number of IMP variants were described as chromosome-encoded, e.g. IMP-24
(Lee et al., 2008), IMP-28 (Perez-Llarena et al., 2012) or IMP-33 (Deshpande et al., 2013). On the
other hand, several other enzymes like IMP-29 (Jeannot et al., 2012) or IMP-34 (Shigemoto et al.,
2013) have been described as plamid-encoded. As integrons can not mobilize themselves to
move to another organism, it must be assumed that the blaIMP-31 gene was mobilized from an
unknown source into the P. aeruginosa isolate NRZ-00156. Consequently, a conjugative plasmid
or conjugative transposon must have been present for mobilization of blaIMP-31 and latter could
still be the case. As gene cassettes in integrons furthermore can easily be excised and integrated
into another integron or transposon structure, it is very likely that the blaIMP-31 gene is
mobilizable into other organisms.

Discussion 94
4.2 Identification of OXA-233
The isolate C. freundii NRZ-02127 also exhibited a carbapenemase phenotype with a positive
modified Hodge Test. However, the resistance spectrum that was detected in MIC analysis did
not match any known carbapenemase profile, as the high resistance towards penicillins, the
inhibition by clavulanic acid and the susceptibility to cephalosporins did neither fit to class A
carbapenemases nor to MBLs or OXA-48. A sparing of oxyimino-cephalosporins is untypical for
class A carbapenemases, although they are inhibited by clavulanic acid (Nordmann et al., 2012).
MBLs also do usually not spare cephalosporins and are unsusceptible to clavulanic acid
(Cornaglia et al., 2011). OXA-48 finally shows lowered hydrolysis of cephalosporins, but is also
not inhibited by clavulanic acid (Poirel et al., 2004b). Therefore, the discovery of the OXA-10
related enzyme OXA-233 was surprising, as OXA-10-like enzymes were classified as narrow- or
extended-spectrum enzymes without carbapenemase activity (Poirel et al., 2010; Evans &
Amyes, 2014). Enzymes of the OXA-10 group were detected first in the U.S. (Korfhagen et al.,
1975) and are found worldwide today (Poirel et al., 2010). In Europe, OXA-10 related enzymes
have been reported in P. aeruginosa isolates from Turkey (Hall et al., 1993; Danel et al., 1995;
Danel et al., 1998; Danel et al., 1999), Germany (Pournaras et al., 2013), Croatia (Sardelic et al.,
2012), Bulgaria (Vatcheva-Dobrevska et al., 2013), Portugal (Moura et al., 2012), France (Aubert
et al., 2001; Poirel et al., 2001; Fournier et al., 2010a; Hocquet et al., 2011) and Denmark
(Hansen et al., 2014).
OXA-233 was very closely related to OXA-35 and OXA-10 with one and two amino acid
substitutions, respectively. At position 117, OXA-233 possessed a phenylalanine residue, while
all other OXA variants exhibit a valine, isoleucine or rarely, leucine (Leonard et al., 2013). As this
mutation was the only one that divided OXA-233 and OXA-17 and as both OXA-10 and OXA-17
and most other OXA-10 related enzymes were classified as only narrow- or extended-spectrum
enzymes (Poirel et al., 2010), it was assumed that the V177F substitution was responsible for
the carbapenemase activity.
blaOXA-233 was identified to be part of a conserved and intact class 1 integron, as it has been
shown for numerous other OXA-type genes, for example blaOXA-28 (Poirel et al., 2001), blaOXA-35
(Aubert et al., 2001), blaOXA-142 (Liu et al., 2014) or blaOXA-320 (Cicek et al., 2014). The OXA-233
encoding gene was the second gene cassette of the integron. As the expression of gene cassettes
within an integron continously decreases with an increasing distance from the promoter
(Gillings, 2014), a lower expression than for the adjacent aac(6´)-Ib gene can be assumed.
However, as the in silico promoter analysis of the integron revealed that a strong Pc promoter
was present, the effect should be minimized as the strong variant of the Pc promoter shows an
expression level almost four times higher than the PcH2 variant identified in the blaIMP-31-
carrying integron, as previously described. As the clinical isolate furthermore exhibited elevated
MICs for penicillins and carbapenems and as OXA-233 seemed to be the only β-lactamase

Discussion 95
present in the isolate, the influence of the position in the integron on expression can be assumed
as rather low. Like for the blaIMP-31-carrying integron, the P2 promoter was found in its inactive
variant, missing the space-optimizing insertion of three guanines between the -35 and -10 boxes
and therewith it can be assumed that P2 had no influence on the expression of blaOXA-233. No
transposon structures were identified in the genetic environment of the blaOXA-233 gene and
analysis of the full sequence of the plasmid pMB3018 showed that the gene was not obviously
part of a transposon, which will be discussed later.
The localization of blaOXA-233 was identified by Southern blot analysis with a blaOXA-233 specific
probe, which showed that the blaOXA-233 was located on this transconjugable plasmid (pMB3018)
with a size of approximately 50 kb. Nuclease S1 digestion revealed two other plasmid bands of
C. freundii NRZ-02127. These bands are likely to represent two other plasmids with a greater
size than pMB3018, but it is also possible that they resulted from circularized and supercoiled
forms of pMB3018 due to an incomplete digestion by nuclease S1. As the hybridization however
showed no signals for blaOXA-233 corresponding to these bands and as the OXA-233
transconjugant did not exhibit these additional bands, an incomplete digestion is very unlikely.
The signals that were detected at a size of approximately 680 kb represented unspecific
hybridizations with the intense bands observed in PFGE at the same size. This is a common
phenomenon in PFGE analyses.
Almost all blaOXA group genes have been found on plasmids, with the exception of the more
recently found intrinsic OXA variants in some Acinetobacter species, for example OXA-213,
OXA-235 or OXA-309 (Evans & Amyes, 2014). The most prominent and clinically relevant
plasmid-encoded CHDL is OXA-48, which can be harboured by the plasmid pOXA-48a and close
relatives which have shown an almost worldwide spread (Poirel et al., 2012a). Among the
OXA-10 group, many enzymes have been described as plasmid-encoded, for example OXA-7 in
E. coli, OXA-10 in P. aeruginosa and OXA-17 also in P. aeruginosa (Philippon et al., 1983;
Medeiros et al., 1985; Danel et al., 1999). The plasmid localization of blaOXA-233 in combination
with the presence of a class 1 integron implies a high ability to spread into other organisms. As
pMB3018 is at least conjugable from C. freundii into E. coli, it is very likely that it is also
transferable into other Enterobacteriaceae species of clinical importance.
4.3 Identification of KHM-2
Like P. aeruginosa NRZ-00156, the isolate P. aeruginosa NRZ-03096 showed a carbapenemase
phenotype in the modified Hodge Test. This was further underlined by the performance of the
EDTA-CDT which indicated the presence of an MBL. However, the inhibition zone increase for
the blank control disk was much greater than for the IMP-31 producing strain with 12 mm for
P. aeruginosa NRZ-00156 and 19 mm for P. aeruginosa NRZ-03096. This phenomenon is
frequently observed in clinical diagnostics and is based upon the differing susceptibilities of the

Discussion 96
different tested isolates to EDTA (Pitout et al., 2005; Galani et al., 2008). Like the IMP-31 isolate,
P. aeruginosa NRZ-03096 showed low MICs towards piperacillin, further indicating an MBL
production, as simultaneously MICs for other penicillins, cephalosporins and carbapenems were
elevated. Carbapenem MICs however were only slightly elevated and the isolate was susceptible
to imipenem and only indermediate resistant to meropenem according to the EUCAST criteria,
which would have been unusal in case of an MBL production.
Shotgun cloning experiments however revealed that P. aeruginosa NRZ-03096 harboured a
novel metallo-β-lactamase which had a homology of only 74.3 % to the KHM-1 enzyme (KHM for
“Kyorin Health Science MBL1”), which has been first described in 2008 and was found only once
in a clinical C. freundii isolate from Japan (Sekiguchi et al., 2008). As the homology to the next
relative was very low compared to other β-lactamases of the same type, it was considered that
the novel enzyme could represent a novel enzyme type, but as the threshold for the definition of
a new type is proposed at <73 % homology (George Jacoby, personal communication), the
enzyme was designated as KHM-2. The mutations of KHM-2 in comparison to KHM-1 were
spread widely over the whole enzyme, but the highly conserved zinc-binding motifs of subclass
B1 enzymes were not altered. As stated in the results section, this applied only to the histidine
residues directly involved in zinc-binding, as the sequence exhibited a threonine to aspartic acid
substitution at position 100, which is part of the conserved HXHXD zinc binding motif. A
potential influence on the zinc binding efficiency will be discussed later.
Regarding the genetic environment of the blaKHM-2 gene, it was surprising that no integron
structures were found, as most other MBL genes were described as integron-bourne (Cornaglia
et al., 2011). The identification of two putative transposase genes adjacent to the blaKHM-2 gene
however, strongly suggests a mobility of the gene, although no repeat regions could be idenified
as associated with the putative transposon structure. The insE transposase gene has been
described as part of a large insertion sequence flanked by two IS903 elements, which are
members of the IS5 family (Sekizuka et al., 2011). In IS903, insE was furthermore identified as
associated with the chaperon-enconding genes groEL and groES upstream of the 5´ end of insE.
An association of insE with these genes was also identified in a blaNDM-1-carrying transposon
from A. baumannii, while insE was also not associated with repeat regions in this context (Pfeifer
et al., 2011), as it was detected for blaKHM-2. As the sequence of the genetic environment of
blaKHM-2 obtained by shotgun cloning did not provide information on the genes located further
downstream of the insE gene, a possible association with the chaperon-encoding genes groEL
and groES could not be analyzed. Regarding the gene coding for the putative transposase of the
ISXo2 family, it was not possible to identify any putative repeat region adjacent to the ORF,
especially as the obtained sequence from shotgun cloning did not cover the whole ORF. ISXo2
has been described as an insertion sequence found in Xanthomonas oryzae pv. oryzae
(Rajeshwari & Sonti, 2000) but has not yet been described as beeing associated with any type of

Discussion 97
antibiotic resistance. However, a BLAST homology search for the isxo2-like sequence yielded
another hit with 74 % similarity that belonged to the whole genome sequence of the
P. aeruginosa ST111 outbreak strain PA38182 (Genbank HG530068.1). Several putative β-
lactamase and MBL genes are annotated in this sequence; however, no further information on
this sequence is currently available. Regarding the aac(3´)-like gene adjacent to the blaKHM-2 gene,
the BLAST search yielded only a single hit and it remained unclear, why the repective ORF was
annotated as an aac-type gene in the whole genome sequence of the Gloeobacter violaceus PCC
7421 strain (Genbank accession number BA000045.2). As the sequence of the putative gene
showed no homologies to any other aac-like sequences in the NCBI database, it must be assumed
that the annotation in the NCBI database is incorrect. Consequently, the function of the ORF
downstream of the blaKHM-2 gene could not be surely identified.
A comparison of the genetic environments of blaKHM-2 and blaKHM-1 was difficult, as only little is
known about the genetic environment of the KHM-1 encoding gene in the C. freundii isolate from
Japan. In the vicinity of blaKHM-1 a 360-bp ORF that encodes the hypothetical protein VP1798 of
Vibrio parahaemolyticus has been described (Sekiguchi et al., 2008). This ORF was not identified
in the genetic environment of blaKHM-2, so a more comprehensive comparison of the genetic
contexts of both genes was not possible.
According to the literature, the blaKHM-1 gene was found only once in the C. freundii isolate from
Japan (Sekiguchi et al., 2008) and no further cases where the gene was found in clinical isolate
have been described since 2008. Consequently, an efficient spread of the blaKHM-1-carrying
plasmid can be excluded. P. aeruginosa NRZ-03096 was referred to the NRZ from a German
diagnostics lab and it is remarkable, that the only two members of a group of MBL enzymes are
found in two species of a different order and with such a huge geographical distance. As KHM-2
and KHM-1 furthermore show distinct differences to each other and with respect to the large
geographical distance between the two isolates, it can be speculated that KHM-2 did not
originate from the Japanese KHM-1 and rather represents a de novo mobilization from an
unknown environmental source of blaKHM-type genes. With a GC content of ~43 % it is
furthermore implicated that KHM-type genes were mobilized into C. freundii and P. aeruginosa,
which usually have higher GC contents of 51 % and 66 %, respectively. While the blaKHM-1 gene
was found on a plasmid which was conjugable from C. freundii into E. coli W1895 (Sekiguchi et
al., 2008), the analysis of the gene localization in this study showed that the blaKHM-2 gene was
chromosomally-encoded in P. aeruginosa NRZ-03096. Consequently, the blaKHM-2 gene must have
been mobilized into P. aeruginosa by a plasmid or a transposon-mediated mechanism. As two
putative transposases were found adjacent to the gene, one of these could be responsible for the
integration in the chromosome. Furthermore, the chromosomally localization of the gene was
another indicator that the gene was mobilized into P. aeruginosa from another species, as no
KHM-like genes have been described in this species so far. However, it was remarkable that the

Discussion 98
isolate expressed a sequence type similar to ST395. ST395 has only rarely been described in the
context of antibiotic resistance in clinical isolates. The only published cases were from France
and Hungary, but ST395 was not reported as a carrier of carbapenemases in these cases (Libisch
et al., 2009; Cholley et al., 2011; Slekovec et al., 2012; Valot et al., 2014). Consequently, a rapid
spread of this KHM-2-carrying P. aeruginosa sequence type in healthcare settings remains
questionable, as other sequence types like ST235 are significantly more prevalent worldwide.
4.4 Catalytic characteristics of IMP-31, OXA-233 and KHM-2
The catalytic properties of β-lactamases are essential for their ability to confer resistance against
various β-lactam antibiotics and the knowledge of kinetic properties for these enzymes is crucial
for correct antibiotic therapy. The characterization of the impact of amino acid mutations on
enzyme functionality is furthermore important for the understanding of the structural
properties of these enzymes of high clinical importance.
4.4.1 Characteristics of IMP-31
Interestingly, E. coli TOP10 cells heterologously producing IMP-31 showed relatively low MIC
increases compared to the IMP-1 strain. Although MICs for all tested carbapenems and most
other β-lactams were distinctly increased in relation to the control strain with up to more than
128-fold increased resistance, the total values were rather low. The IMP-1 strain showed
significantly higher MICs for all tested β-lactams except piperacillin, for which the MICs were
only slightly increased for both IMP strains. For example, the MIC for ceftazidime was 32 mg/l
for the IMP-31 strain, representing one of the highest increases in relation to the control strain.
For the IMP-1 strain, the MIC for ceftazidime was higher than 256 mg/l and thereby higher than
detectable by Etest strips, indicating a significant difference in the catalytic properties between
the two enzymes. It was shown for other IMP-type enzymes, that heterologous expression of the
enzymes in E. coli led to MICs only slightly higher than for the control strains. For example, a
strain expressing IMP-13 was shown to exhibit MICs of 0.125 mg/l for meropenem and
ertapenem, while the IMP-1 strain showed values of 2 mg/l for both antibiotics in the respective
study. The control strain without a β-lactamase showed values of 0.015 mg/l, resulting in an 8-
fold and 133-fold increase for IMP-13 and IMP-1, respectively (Santella et al., 2011). For a strain
expressing the IMP-18 variant, MICs of 1 mg/l, 0.06 mg/l and 0.12 mg/l were detected for
imipenem, meropenem and ertapenem, representing 17-fold, 4-fold and 8-fold increases in
relation to the control strain. An IMP-1 expressing strain on the other hand showed a 33-fold
increase in the MIC for imipenem and 133-fold MIC increases for meropenem and ertapenem in
the respective study (Borgianni et al., 2011). A comparison with the MICs mediated by the next
nearest relative of IMP-31, IMP-35, was not possible as no MIC data were available for this
enzyme (Pournaras et al., 2013). The second nearest relative IMP-8 however was described to

Discussion 99
mediate MIC increases of 15-fold for imipenem and 8-fold for meropenem, respectively (Yan et
al., 2001). Compared to MIC increases mediated by the production of the third next relatives
IMP-2 and IMP-19 it was noticable that expression of these enzymes resulted in significantly
higher increases for imipenem than an expression of IMP-31, while the increases were similar
for meropenem (Riccio et al., 2000; Neuwirth et al., 2007). With regard to the MIC increases
reported from other studies, the increases mediated by production of IMP-31 remained low in
comparison to the influence of an IMP-1 expression, but were nontheless higher than it was
detected for several other IMP-type enzymes.
The kinetic parameters that were determined for IMP-31 were in good agreement with most of
the MIC data. IMP-31 showed a generally lower hydrolytic activity against β-lactam antibiotics
than the reference enzyme IMP-1. Although IMP-31 was able to hydrolyze all tested penicillins,
cephalosporins and carbapenems, the catalytic efficiencies were rather low, the highest value
was 2.8 µM-1 ∙ s-1. In contrast, the highest hydrolysis rate for IMP-1 was 18 µM-1 ∙ s-1. The inability
of IMP-31 (and IMP-1) to hydrolyze the monobactam aztreonam was expected, as this is a key
characteristic of MBL enzymes (Cornaglia et al., 2011). As no kinetic data were available for
IMP-35 (Pournaras et al., 2013), a direct comparison of the kinetic parameters of IMP-31 and
IMP-35 was not possible. Also no kinetic data were available for the next nearest relative IMP-8
(Yan et al., 2001). Compared to IMP-2 and IMP-19, which were the next relatives with available
kinetic data, IMP-31 showed lower hydrolytic efficiencies for imipenem and meropenem than
IMP-2 (Riccio et al., 2000). In comparison to IMP-19, IMP-31 showed higher rates for both
antibiotics (Neuwirth et al., 2007). This was contrasting the MIC comparisons, where IMP-19
was shown to mediate a higher relative increase than IMP-31. Regarding penicillins and
cephalosporins, IMP-31 showed higher catalytic efficiencies for ampicillin and ceftazidime than
IMP-2. Compared to IMP-19, IMP-31 showed lower hydrolysis rates for penicllin G, but higher
efficiencies against cefoxitin and ceftazidime. Although the catalytic efficiency of IMP-31 were
lower than for the reference enzyme IMP-1, the high MICs of the clinical isolate P. aeruginosa
NRZ-00156 indicated that even a weaker carbapenemase is apparently sufficient to confer high
levels of carbapenem resistance in clinical isolates, as it has also been shown for IMP-8 , IMP-13
and IMP-18 (Yan et al., 2001; Toleman et al., 2003a; Hanson et al., 2006). Most clinical
P. aeruginosa strains possess additional resistance mechanisms like porins loss, efflux pumps or
additional β-lactamases which act in concert to confer detectable carbapenem resistance
(Livermore, 2001). This has also been shown in this study, as the isolate P. aeruginosa
NRZ-00156 carried the blaOXA-35 gene as a second β-lactamase gene. It can be furthermore
assumed that the expression level of genes under the control of a class 1 integron promoter is
much higher in a wildtype strain than in an E. coli K12-derived laboratory strain such as TOP10,
where both IMP-31 and IMP-1 were expressed under the control of a lac promoter, which has
been described as relatively weak (Deuschle et al., 1986).

Discussion 100
It has been shown for many other MBLs and IMP-type enzymes in particular, that relatively
small changes in the amino acid sequence can lead to significantly altered catalytic properties.
For example, IMP-10 differs by only one single amino acid substitution from IMP-1 but shows
almost no hydrolysis of penicillin G and ampicillin (Iyobe et al., 2002). As IMP-31 and IMP-1
differ from each other by 50 amino acid substitutions, it was very likely that this large number of
mutations had an influence on the catalytic behaviour or the tertiary structure of the enzyme.
For more detailed analysis, the tertiary structure of IMP-31 was modelled using the IMP-2
crystal structure as the template, as IMP-2 shows a higher homology to IMP-31 than IMP-1. A
comparison of the IMP-31 protein model and the crystal structure of IMP-1 is shown in Figure
4.2. Although the model of IMP-31 showed slight changes in the distances between the second
group of zinc binding ligands and the respective zinc ion compared to IMP-1, these changes were
rather low with the highest difference shown with an increase from 2.3 to 2.7 Å for His197. The
distance between the two coordinated zinc ions was also only slightly changed from 3.5 to 3.6 Å
but as it has been proposed that the distance between the zincs is important for the coordination
of the catalytic water molecule in IMP-1 (Yamaguchi et al., 2005), this could have an influence on
the hydrolytic properties. However, the quality score of the model was only 0.85 and therefore
the possibility of a deviation of the model from the true structure of IMP-31 remained. As the
mutations of IMP-31 are spread widely over the whole enzyme, it could also be possible that the
altered characteristics result from structural changes not directly related to the active site. It is
thought that substrate binding and hydrolysis of MBLs is influenced by a flexible loop near the
active site which is formed by a tryptophan or phenylalanine residue (Palzkill, 2013). It has been
proposed that this loop supports the tight binding of substrates and the stabilization of the
Figure 4.2 Crystal structure and homology model of the active site of IMP-1 (A) and IMP-31 (B). The highly conserved zinc binding ligands of the active site are colored in purple and the tryptophan of the flexible loop at position 81 is colored in blue. Distances between the zinc binding ligands and the two zinc ions are indicated by dashed lines and denoted in Å. The IMP-1 crystal structure was taken from PDB accession number 4UAM.2. The IMP-31 homology model was constructed using the SWISS-Model server using the crystal structure of IMP-2 (PDB accession number 4UBQ.1) as a template. The figures were rendered using PyMOL.

Discussion 101
active site ligands. In IMP-type enzymes, this residue is a tryptophan located at position 46 and
although this residue was not mutated in IMP-31, five surrounding residues showed alterations
compared to IMP-1 (Figure 3.2). This could possibly have an influence on the function of the
flexible loop and thereby influence the catalytic efficiency, as it has been shown for IMP-1 that
mutations of this loop result in significantly altered turnover numbers (Moali et al., 2003). In the
IMP-31 model, this loop was slightly altered and the conserved tryptophan was rotated, which
could possibly affect the ability of the loop to stabilize the active site. However, further structure
analysis including crystallization is needed to surely identify the reason for the catalytic
behaviour of IMP-31, as these hypotheses are based on homology modelling.
In conclusion, IMP-31 was shown to have a distinct carbapenemase activity that is very likely to
confer high levels of β-lactam resistance in Gram-negative clinical isolates.
4.4.2 Characteristics of OXA-233
OXA-233 showed distinct differences in the ability to confer β-lactam resistance in comparison
to OXA-10. While E. coli TOP10 expressing OXA-10 showed high MICs for penicillins and
penicillin/inhibitor combinations, the OXA-233 strain clearly showed an inhibition by
sulbactam, tazobactam and clavulanic acid, which is very atypical for a class D enzyme,
especially for OXA-10-like enzymes. OXA-type enzymes that were described as inhibited by
clavulanic acid are OXA-12 (Rasmussen et al., 1994; Walsh et al., 1995), OXA-18 (Philippon et al.,
1997), OXA-20 (Naas et al., 1998), OXA-45 (Toleman et al., 2003b), OXA-53 (Mulvey et al., 2004)
and OXA-63 (Meziane-Cherif et al., 2008), but these do not belong to the OXA-10 group. No
OXA-10-like enzymes have so far been described as inhibited by this compound. As expected, the
OXA-233 expressing strain showed significantly lower MICs for piperacillin-tazobactam and
amoxicillin-clavulanate than for the single antibiotics. Clavulanic acid, sulbactam and
tazobactam are clinically used β-lactam inhibitors that share structural similarity with penicillin.
While clavulanic acid was isolated from Streptomyces clavuligerus and is clinically used in the
salt form clavulanate, sulbactam and tazobactam are synthetic penicillinate sulfones (Reading &
Cole, 1977; English et al., 1978; Fisher et al., 1980). The mechanism of action of these substances
is similar to the hydrolysis of β-lactam antibiotics; however, the hydrolysis rate is extremely low,
leading to an inhibition of the enzyme similar to the inhibition of PBPs by β-lactams (Drawz &
Bonomo, 2010). The inhibition effectiveness is known to depend on the inhibitor/enzyme
combination, for example TEM-1 needs 160 clavulanate molecules for inactivation, while SHV-1
requires only 60 (Drawz & Bonomo, 2010). Consequently, it can be hypothesized that the two
amino acid substitions of OXA-233 relative to OXA-10 lead to a reduced amount of molecules
that is necessary for inhibition. In addition, one or both of the substitutions seem to mediate the
susceptibility of OXA-233 towards sulbactam, which was not detectable for OXA-10.

Discussion 102
Regarding the MICs for cephalosporins, the inability of OXA-233 to confer increased resistance
towards cephalotin, cefuroxime, cefoxitin, cefotaxime, ceftriaxone, cefepime and ceftazidime was
remarkable, as the next nearest relative OXA-17, which differs in only one amino acid
substitution to OXA-233, has been described as an extended-spectrum enzyme with high activity
on cephalosporins (Danel et al., 1999). This indicated that the apparent changes in the substrate
spectrum were more likely based on the V117F substitution than on N73S. Consistent with the
literature, the OXA-10 strain showed only slightly increased cephalosporin resistance and no
increase for ceftazidime. As expected, expression of OXA-233 led to increased MICs for
carbapenems, although the increase was very low. It has been shown for several “weak” OXA
enzymes, that even if they are not capable of high β-lactam hydrolysis rates, they can confer
clinically relevant resistance in Gram-negative wildtype strains (Antunes et al., 2014). In
addition, it can be assumed that expression of OXA-233 and OXA-10 in E. coli TOP10 was at a
relatively low level, as the expression was not induced and controlled by a relatively weak lac
promoter. Furthermore, the higher MIC increases for the OXA-233 transconjugant, where the
blaOXA-233 gene was under control of the strong integron promoter, demonstrated the effect of an
assumable higher expression level.
As the next nearest relatives OXA-10 and OXA-17 were described as unable to hydrolyze
carbapenems, it was initially assumed that the mutation at position 117 from valine to
phenylalanine was responsible for the carbapenemase activity of OXA-233. But surprisingly,
production of OXA-10 also led to increased MICs for carbapenems. This was remarkable, as
enzymes of the OXA-10 group had always been described as narrow- or extended-spectrum
enzymes with no activity on carbapenems (Poirel et al., 2010; Leonard et al., 2013; Evans &
Amyes, 2014). However, in 2014 it was shown that probably all class D enzymes (including
OXA-10) are in fact carbapenemases and that many previous characterizations of OXA-type
enzymes were inaccurate (Antunes et al., 2014). This was explained by the fact that OXA
enzymes possess a highly conserved lysine residue at position 70 (K70), which is an important
part of the active site and plays a crucial role in β-lactam hydrolysis. K70 is suggested to activate
the attacking groups in both acylation and deacylation reactions during the opening of the β-
lactam ring and therewith thought to influence the ability of the β-lactamase to quickly bind
another substrate molecule (Schneider et al., 2009). This lysine residue has been shown to be
carboxylated in vivo (Golemi et al., 2001) and it has further been shown that the carboxylation is
required for full enzyme activity (Schneider et al., 2009). Antunes et al. (2014) stated that almost
all biochemical characterizations of OXA-type enzymes were performed without the
supplementation of a CO2 source to the reaction mixture and that this could lead to
decarboxylation of K70, resulting in reduced enzyme activity. They further showed that the
narrow-spectrum enzymes OXA-2 and OXA-10 in fact were able to hydrolyze carbapenems with
low efficiency when a CO2 source was present. With this background, biochemical

Discussion 103
characterization of OXA-233 and OXA-10 was performed with addition of a CO2 source to the
reaction mixture in this study. Determination of kinetic parameters clearly showed that both
OXA-233 and OXA-10 were able to hydrolyze carbapenems in vitro and that the carbapenemase
activity was CO2-dependent, confirming the findings of Antunes and colleagues (2014).
Hydrolytic efficiencies observed for OXA-233 were rather low compared to OXA-10, showing
higher values for all tested substrates except for meropenem, which was the only substrate for
which OXA-233 showed a higher activity. The greatest difference was detected for
cephalosporins as the hydrolysis rates for cefoxitin and ceftazidime of OXA-233 were too low to
be determined with the experimental setup used in this study. However, cephalosporin
hydrolysis rates of OXA-10 were also extremely low. In general, the catalytic data of both
OXA-233 and OXA-10 reflected the results obtained from the MIC analyses for both OXA-233 and
OXA-10. Interestingly, OXA-233 showed a significantly lower affinity towards ampicillin and
oxacillin and in contrast to OXA-10 was not able to hydrolyze aztreonam. This was also
surprising as all other OXA-10-like enzymes have been described as beeing capable of a
moderate aztreonam hydrolysis (Poirel et al., 2010).
It has been mentioned before, that only two amino acid substitutions (N73S and V117F)
distinguish OXA-10 and OXA-233. Consequently, at least one of these mutations must be the
reason for the differences in catalytic behaviour. In all OXA enzymes except OXA-233, the highly
conserved position 117 is always occupied by valine, isoleucine or rarely, leucine and is an
important hydrophobic active site residue as a part of the “omega loop” (Schneider et al., 2009;
Poirel et al., 2010; Leonard et al., 2013). It has been shown that mutation of V117 to aspartic acid
leads to a decarboxylation of the important active site lysine K70, resulting in a loss of enzyme
activity (Schneider et al., 2009). In OXA-233, position 117 is occupied by phenylalanine. The
aromatic side chain of phenylalanine is more hydrophobic than the ones of valine, isoleucine or
leucine, which could enhance the tertiary structure stability of the active site or the whole
enzyme. It has been shown for a large variety of proteins, that increased hydrophobicity
positively influences stability of the tertiary structure (Kellis et al., 1988; Pace et al., 2011).
Consequently, it can be speculated that an increased hydrophobicity of the active site of
OXA-233 could lead to a decreased flexibility of the site and thereby to lower hydrolysis rates.
However, this would not explain several differences in the catalytic behaviours of OXA-233 and
OXA-10 as the hydrolytic efficiency of OXA-233 was not overall decreased and it was slightly
elevated towards meropenem and similar for ertapenem. In addition, a high hydrophobicity of
the active site is beneficial for efficient carboxylation of K70 (Leonard et al., 2013) and
consequently, replacement of V117 with an even more hydrophobic residue should rather result
in increased enzyme activity. In site-directed mutagenesis experiments that were performed
with OXA-1 by Buchman and colleagues (2012), it was shown that a substitution of V117 to

Discussion 104
Figure 4.3 Crystal structure and homology model of the active sites of OXA-10 (A) and OXA-233 (B). The highly conserved residues of the active site are colored in purple. Position 117 is colored in red. The OXA-10 structure was taken from PDB accession number 2WGW.1.B. The OXA-233 homology model was constructed using the SWISS-Model server. The figures were rendered using PyMOL.
phenylalanine leads to significantly lower MICs for ampicillin when expressed in E. coli.
However, lowered MICs for ampicillin were not detected for OXA-233 in this study, but as
OXA-233 and OXA-1 only show a homology of 26 %, this could be based on other structural
differences. To gain more detailed information on the potential structure of OXA-233 and the
influence of the V117F mutation, a homology model was constructed, based on the crystal
structure of OXA-10 (PDB accession number 2WGW.1.B). A comparison of the structure models
of the active site of OXA-233 and OXA-10 is shown in Figure 4.3. The homology model of
OXA-233 showed that the large aromatic side chain of phenylalanine extends into the space of
the active site. This would definitely lead to alterations in the distances between the important
active site residues (S67, K70, S115, W154, L155, K205 and G207) and F117, possibly affecting
carboxylation of K70 and the catalytic behaviour. It has to be noted that modelling of the
carboxylation of K70 was not possible and a comparison with the crystal structure image clearly
indicates that in case of a carboxylation, the side ring of F117 and the carboxy group of K70
could collide, further implicating a possible conformational change of the active site in OXA-233.
For this reason, no atom distances were calculated as they would be too inaccurate. Another
explanation for the catalytic characteristics of OXA-233 could be a sterical hindrance of the
binding of some β-lactams, as the measured Km values were mostly higher than detected for
OXA-10. Especially oxyimino-cephalosporin hydrolysis could be influenced, as their R2 side
chains are larger than the ones of penicillins and carbapenems (Figure 4.4). This could also be an
explanation for the inability of OXA-233 to hydrolyze aztreonam, which also possesses a
relatively large R2 group. However, this would not explain the extremely low hydrolysis of

Discussion 105
Figure 4.4 Chemical structures of ceftazidime, aztreonam and penicillin G. Ceftazidime is shown as a representative for oxyimino-cephalosporins. The R2 side chains of oxyimino-cephalosporins and aztreonam is significantly larger than the R2 chain of penicillin G and carbapenems (Figure 1.2).
cefoxitin, which possesses a R2 side chain comparable to penicillin G. To verify the hypothesis
that the V177F substitution is responsible for the altered hydrolytic properties of OXA-233,
further structure analysis, including crystallization and substrate binding modelling, is needed.
In conclusion, it was clearly shown that OXA-233 has a CO2-dependent carbapenemase activity
and that this activity leads to increased resistance against many β-lactam antibiotics. As
OXA-233 showed extremely weak hydrolysis of cephalosporins, these antibiotics could still
represent a therapy option against OXA-233-producing Gram-negative pathogens. It has been
proposed that oxyimino-cephalosporins are a potential therapy option for OXA-48-producing
Enterobacteriaceae without an ESBL association, as like OXA-233, OXA-48-like enzymes (except
OXA-163) show weak hydrolysis of these antibiotics (Poirel et al., 2012b). However, only few
clinical data are available to support the use of these antibiotics with only one published study
showing a successful treatment of a neonate infected by an OXA-48-producing K. pneumoniae
strain in France with a combination of cefotaxim and amikacin (Levast et al., 2011).
Furthermore, a sucessful treatment of a patient also infected with an OXA-48-producing
K. pneumoniae strain using a combination of ceftazidime and avibactam has been reported from
Spain (Mora-Rillo et al., unpublished data). Another study performed with a peritonitis model in
mice also showed that treatment with ceftazidime was an efficient therapy against OXA-48-
producing K. pneumoniae (Mimoz et al., 2012). Regarding these data, it can be suggested that
oxyimino-cephalosporins remain an option also for OXA-233 producing Enterobacteriaceae that
do not possess other cephalosporin resistance mechanisms. This is furthermore supported by
the MIC data obtained for C. freundii NRZ-02127, which was shown to be susceptible or
intermediate to cefotaxime, cefepime and ceftazidime, indicating potential therapy options
against producers of this novel class D carbapenemase. In addition, the observed inhibition by
clavulanic acid in the MIC studies might be clinically relevant, but has to be further analyzed in
future experiments.

Discussion 106
4.4.3 Characteristics of KHM-2
An expression of KHM-2 in E. coli TOP10 resulted in significantly higher MICs for all tested β-
lactams except piperacillin and aztreonam. Compared to the only known relative, KHM-1, the
resistance spectrum showed higher MIC increases for ampicillin and amoxicillin, indicating a
more efficient hydrolysis of these substrates. Cephalosporin MICs were on a comparable level
for both strains. Regarding carbapenems, MICs were also elevated for both strains, with the
KHM-2 strain showing a 2-fold higher increase for imipenem, while the KHM-1 strain showed
higher increases for meropenem, doripenem and ertapenem. The MICs which were detected for
both strains were significantly higher than compared to those of the control strain with
increases of up to 8,000-fold for cefotaxime. The kinetic analysis of KHM-2 and KHM-1 reflected
the resistance spectrum observed in the MIC studies for most tested substrates with penicillin G,
cefoxitin and ceftazidime beeing well hydrolyzed with efficiencies of up to 10.5 µM-1 ∙ s-1.
Interestingly, the hydrolysis rate of KHM-2 for imipenem was higher than the rate for
ceftazidime, while it was distinctly lower for the reference enzyme KHM-1. Consistent with the
MIC data, KHM-1 had higher rates for meropenem and ertapenem. The greatest difference in
catalytic efficiency was detected for cefotaxime, which was also 10-fold more efficiently
hydrolyzed by KHM-1. Like all other MBLs characterized in this study, KHM-2 was not able to
hydrolyze aztreonam.
As no information is available in the literature regarding the structure or specific amino acids of
KHM-1 and as this enzyme furthermore shows a homology of only 74 % to KHM-2, it was
difficult to form a hypothesis for the influence of the mutations of KHM-2 on β-lactam hydrolysis.
As the zinc binding ligands of KHM-2 and KHM-1 are identical to other subclass B1 MBls (Garau
et al., 2004) and were not altered in KHM-2, a weaker zinc binding, possibly influencing the
hydrolytic efficiency, is rather unlikely. However, the substitution at position 100 from
threonine to aspartic acid could have an influence on the zinc coordination. The first zinc
binding site of subclass B1 MBLs is formed by two histidines at the consensus positions 116, 118
and 196, while the second zinc binding site is formed by an aspartic acid residue at postion 120,
a cysteine at position 221 and a histidine at position 263 according to the MBL standard
numbering scheme (Garau et al., 2004). In the KHM-1 and KHM-2 sequences, these residues
correspond to the positions 97, 99 and 159 for the first binding site and 101, 178 and 217 for the
second binding site. For KHM-2, the T100D subsitution leads to two neighbored aspartic acid
residues, which might compete as zinc-binding ligands. This could result in an alteration of the
distances between the two zinc ions or the three ligands of the second zinc binding site. As these
residues play a crucial role for correct coordination of the zinc ions for the nucleophilic attack on
the β-lactam ring (Palzkill, 2013), a distance alteration could possibly affect the hydrolytic
efficiency. To gather more information on the putative influences of the mutations and to
substantiate these hypotheses, the structures of KHM-2 and KHM-1 were modelled based on

Discussion 107
homologies with the crystal structure of IMP-1, which was the next nearest relative to KHM-type
MBLs, but with an identity of only 58.33 % to KHM-1 and 61.36 % to KHM-2. The homology
models are shown in Figure 4.5. Modelling showed that the zinc binding site of KHM-2 could in
fact be influenced by the T100D substitution, as the zinc binding sites of both aspartic acid
residues are oriented in the same direction, which could lead to disturbances in zinc
coordination. This could lead to altered hydrolytic characteristics, as it has been shown for
IMP-1 that mutations of Asp120 (MBL standard numbering scheme; Asp101 in KHM-type
enzymes) can significantly influence the hydrolytic efficiency due to alterations in the distance
between the two coordinated zinc ions (Yamaguchi et al., 2005). Regarding the conserved
tryptophan of the flexible loop, the models showed that the mutations of the surrounding
residues in KHM-2 probably lead to a conformational change of the loop, resulting in an
increased distance of the tryptophan to the active site. As it has been shown for IMP-1,
mutations of this residue can affect the kcat values for various substrates (Moali et al., 2003).
Consequently, the increased distance to the active site in KHM-2 could influence the hydrolytic
characteristics of KHM-2. However, as the model qualities were rather low due to the low
homology to the template crystal structure, these conclusions remain hypothetical and have to
be confirmed by crystal structure analysis of both KHM-2 and KHM-1. As the differences in
substrate hydrolysis between KHM-2 and KHM-1 were furthermore rather diverse with some
rates beeing higher and some lower for KHM-2, these can not be fully explained on the basis of
Figure 4.5 Homology models of KHM-1 (A) and KHM-2 (B). The highly conserved zinc binding ligands of the active site are colored in purple for the first ligand group and in blue for the second ligand group. The T100D substitution in KHM-2 is colored in cyan and the conserved tryptophans of the flexible loop of MBLs are colored in orange. Both models were constructed using the SWISS-Model server. The crystal structure of IMP-1 (PDB accession number 1ddk.1) which was the next nearest related structure (58.33 % homology to KHM-1 and 61.36 % to KHM-2) was used as a template. The figures were rendered using PyMOL.

Discussion 108
the models as the mutations mentioned here mostly led to overall lowered or increased activity
in IMP-1 (Moali et al., 2003; Yamaguchi et al., 2005).
In conclusion, it was shown that KHM-2 is a novel metallo-β-lactamase with a high
carbapenemase activity, showing distinct differences to the next nearest relative KHM-1 in
catalytic behaviour. It can be hypothesized that these altered characteristics are based on
several mutations of amino acid residues in the vicinity of the highly conserved residues of the
zinc binding motifs and the flexible loop of subclass B1 MBLs.
4.5 Characterization of the blaOXA-233-carrying plasmid pMB3018
The plasmid pMB3018 was related to the four other IncN-like plasmids pJIE137, p271A, pECS01
and pTR3. The plasmid backbone of all five plasmids showed high homologies and it has been
suggested that this variant of the IncN-type be classified as a novel subgroup named IncN2
(Poirel et al., 2011). The main characteristic of the other IncN2-type plasmids is the presence of
a complete tra locus with the tra genes K, J and I beeing separated from the main locus. These
genes code for subunits of the sex pilus or for proteins with various functions necessary for
conjugational transfer, e.g. plasmid stability proteins or components of the relaxosome (Zatyka
& Thomas, 1998). In pJIE137 the traKJI-locus is located downstream of the main tra-locus,
separated by the ΔfipA gene (Partridge et al., 2012), which was also found in pMB3018. In p271A
and pTR3 the traKJI-locus is located more distant to the main locus on the 3´-extremity of the
oriT (Poirel et al., 2011; Chen et al., 2012), showing one of the main differences regarding the
plasmid backbone to pMB3018. A region that was found in all five related plasmids was the
region containing the stbA, stbB and stbC genes next to the traKJI-locus, which are predicted to
code for plasmid stability proteins. While p271A, pTR3 and pECS01 were described as carrying
the blaNDM-1 gene, bracketed by the two insertion sequences ISEc33 and ISSen4, pJIE137 carries a
blaCTX-M-16 gene and is missing the insertion sequences (Partridge et al., 2012). In pMB3018
however, ISSen4 was present, but not associated with any resistance gene and without its
counterpart ISEc33. It can be hypothesized that the blaNDM-1-carrying insertion sequence was
disrupted by other mobile genetic elements in pMB3018, as several ORFs coding for phage
integrases or hypothetical transposases were located next to ISSen4. This was supported by the
absence of repeat sequences that could be correlated to ISSen4, as the second repeat could have
been deleted during an integration of the phage integrase genes. Another option could be an
incomplete excision of the NDM-1 IS, resulting in the remaining ISSen4. pJIE137 possesses a 5.2-
kb region that corresponds to the CUP (conserved upstream repeat)-controlled regulon of the
IncN plasmid R46 (Delver & Belogurov, 1997). This region consists of the ardR, ardB and ardK
genes, coding for antirestriction proteins, a gene coding for a single-strand DNA binding protein
(ssb) and the repA gene. While Ard proteins provide protection from the restriction enzymes of
the recipient during conjugation (Wilkins, 2002), the single-strand binding protein is predicted

Discussion 109
to have a protective function in conjugation which is not known in detail (Delver & Belogurov,
1997). In contrast to pJIE137, the plasmids p271A, pTR3 and pECS01 contain a partially deleted
CUP region, missing the ardR and ardB genes. In pMB3018, all genes of the CUP region of
pJIE137 were identified but were disrupted by the large putative transposon or IS structure that
contained the two putative phage integrase genes and ISSen4. As this region showed a
significantly lower GC content compaired to the rest of the plasmid, it can be assumed that it
represents the result of an integration event of mobilized DNA from an unknown source. This
contradicts the hypothesis of the disrupted NDM-1 IS and rather indicates that the ISSen4 in
pMB3018 represents an independent insertion event, as the NDM-1 IS in p271A, pTR3 and
pECS01 is not neighbored to the CUP region (Poirel et al., 2011; Chen et al., 2012; Netikul et al.,
2014). A unique feature of pMB3018 was the blaOXA-233-carrying class 1 integron which was
neither found in pJIE137 nor in the other three related plasmids. Although pJIE137 carried a
class 1 integron, it was located at a different region than in pMB3018 and did not contain any β-
lactamase genes (Partridge et al., 2012). The blaOXA-233-carrying integron was neighbored by
IS6100, but as IS6100 was bracketed by two 123-bp inverted repeats it possibly represented a
complete insertion sequence element. However, the identification of a 13-bp inverted repeat
bracketing the integron and IS6100 was consistent with a perfect transposon structure and it is
as well possible that the blaOXA-233 integron was mobilized into pMB3018 by this structure.
In conclusion and regarding the differences to pJIE137 and the p271A-like plasmids, it is more
likely that pMB3018 originated from pJIE137 or a common ancestor, as the two plasmids shared
the same organization of the two tra loci and the CUP region, which however was disrupted by a
large putative transposon in pMB3018. The blaOXA-233 gene was a unique feature of pMB3018 and
no related integron structure was found in any other IncN2 plasmid deposited in the NCBI
database, demonstrating the immense diversity of β-lactamase gene carrying genetic structures.
4.6 Comparison of IMP-31, KHM-2 and OXA-233 and concluding remarks
Compared to each other, the three novel carbapenemase described in this study showed
significantly different substrate profiles. The metallo-β-lactamases IMP-31 and KHM-2 showed
an efficient hydrolysis of penicillins, cephalosporins and carbapenems as it has been described
as a common characteristic of MBL enzymes and represents the most clinically relevant feature
of this group of β-lactamases (Walsh et al., 2005; Gupta, 2008b; Cornaglia et al., 2011). In most
cases, the presence of an MBL in a clinical Gram-negative pathogen is equivalent to the almost
complete exclusion of β-lactam antibiotics for therapy. As MBL genes are very often
accompanied by additional resistance mechanisms against various classes of antibiotics, MBL-
producing isolates can easily become pan-resistant (Maltezou, 2009). This is further aggravated
by the fact that no MBL inhibitors are available for clinical use (Drawz & Bonomo, 2010).
Although it has been shown for many metallo-β-lactamase that these enzymes are not able to

Discussion 110
hydrolyze piperacillin with high efficiencies, this was never stated as a common characteristic of
MBLs (Walsh et al., 2005; Gupta, 2008b; Maltezou, 2009; Cornaglia et al., 2011). As all four MBLs
analyzed in this study showed very low hydrolysis rates for penicillin and as the KHM-2-
harbouring isolate P. aeruginosa NRZ-03096 was susceptible to piperacillin according to
EUCAST criteria, this antibiotic might still represent a possible treatment option, even in the
presence of an MBL. However, this could only be an option in the case of the absence of
additional piperacillin resistance mechanisms such as porin loss, exporter pumps or expression
of another β-lactamase. Regarding all other tested β-lactams, both IMP-31 and KHM-2 showed a
distinct effect on the resistance of E. coli, but it was remarkable that the KHM-producing strains
showed significantly higher MIC increases than the IMP and OXA expressing strains. This either
indicated a very efficient hydrolysis of β-lactam antibiotics by KHM-2 and KHM-1 or a
significantly higher expression in E. coli TOP10 than the IMP- and OXA-expressing strains. But as
the kinetic parameters of both KHM-type enzymes were not very different from the IMP and
OXA-type enzymes characterized in this study, this indicated a significantly increased expression
in E. coli TOP10. However, the specific reason for this assumed higher expression remains
unclear. In conclusion, both IMP-31 and KHM-2 had a significant impact on β-lactam resistance
and it must be assumed that both enzymes are able to confer clinically relevant resistance levels
for β-lactams and carbapenems in particular in Gram-negative species of clinical importance. In
contrast to the broad spectrum of IMP-31 and KHM-2, the class D enzyme OXA-233 showed very
low hydrolysis of cephalosporins but was able to hydrolyze carbapenems if supplied with a CO2
source. As sufficient CO2 sources are probably abundant at infection sites, the experimental
conditions used in this study likely reflect the situation in vivo and it can be assumed that the
confirmation of a carbapenemase activity of OXA-10-like enzymes might be clinically relevant.
In conclusion, it was shown for all three discovered novel β-lactamases that they possess a
distinct carbapenemase activity and can confer increased β-lactam resistance in E. coli, including
resistance to carbapenems. As the OXA-233-encoding gene was located on a transconjugable
plasmid and as the blaIMP-31 and blaKHM-2 genes were very likely located in transposon structures
on the chromosome, it can be furthermore assumed that all three enzymes are able to spread
into other organisms and might play an important role in the future for multidrug-resistance in
Gram-negative pathogens.

Summary 111
5 Summary
The increasing number of carbapenemase-producing Gram-negative pathogens responsible for
healthcare-associated infections is a major clinical problem. Consequently, the identification and
characterization of novel carbapenemase genes and their encoded enzymes is crucial for both
clinical diagnostics and antimicrobial therapy. In this thesis, three carbapenem-resistant Gram-
negative clinical isolates of the species Pseudomonas aeruginosa and Citrobacter freundii were
analyzed on the presence of a novel carbapenemase and three novel enzymes were successfully
discovered by PCR techniques and shotgun cloning approaches: IMP-31, OXA-233 and KHM-2.
While IMP-31 and KHM-2 were metallo-β-lactamases of the molecular β-lactamase class B,
OXA-233 was an OXA-10 related class D enzyme. By characterization of the genetic environment
of the three novel β-lactamase genes and analysis of the gene localization it was shown that
blaIMP-31 and blaOXA-233 were part of a class 1 integron, while blaKHM-2 was not part of such a
genetic structure. With pulsed-field gel electrophoresis experiments and Southern blot
hybridizations, it was shown that the IMP-31 and KHM-2 encoding genes were located on the
bacterial chromosome of the clinical isolates, implying that the two MBL genes were integrated
into the respective chromosome by a transposon-mediated mechanism. On the other hand, the
blaOXA-233 gene was identified on the conjugable plasmid pMB3018 with a size of 52 kb which was
fully sequenced by 454-pyrosequencing and was shown to be a member of the IncN2
incompatibility group. As the genes identified in this study showed a significantly different GC
content compared to the species they were found in, it must be assumed that they were
mobilized into these species from a still unknown source. The impact of a production of IMP-31,
OXA-233 and KHM-2 on β-lactam resistance was analyzed by determination of the minimal
inhibitory concentration (MIC) for various β-lactam antibiotics for Escherichia coli strains
expressing the three novel enzymes and their respective reference enzymes. The analysis
showed that production of all three enzymes leads to significantly increased resistance against
most β-lactams and carbapenems in particular. By determination of the kinetic parameters kcat
and Km, which reflect the catalytic efficiency of an enzyme, FPLC-purified IMP-31, OXA-233,
KHM-2 and their respective reference enzymes were characterized biochemically by in vitro
hydrolysis assays. For each enzyme, the kinetic parameters were determined for ten different β-
lactam substrates using non-linear regression and the analyses showed that all three enzymes
are distinct carbapenemases which are likely to confer clinically relevant carbapenem resistance
levels in Gram-negative pathogens. IMP-31 and KHM-2 showed hydrolysis of almost every tested
β-lactam, while OXA-233 was lacking a high hydrolytic efficiency for cephalosporins, indicating a
possible treatment option. Carbapenem hydrolysis of OXA-233 was CO2 dependent and
confirmed the recent finding, that probably all class D enzymes are able to hydrolyze

Summary 112
carbapenems when supplied with a CO2 source. The identification and characterization of
IMP-31, OXA-233 and KHM-2 in this thesis underlines the ongoing spread and diversification of
carbapenemases in Gram-negative species of clinical importance.

Zusammenfassung 113
6 Zusammenfassung
Die zunehmende Zahl von Carbapenemase-produzierenden Gram-negativen Krankheitserregern
ist ein immenses klinisches Problem. Daher ist die Identifizierung und Charakterisierung neuer
Varianten dieser Enzyme von großer Bedeutung für die klinische Diagnostik und die korrekte
Antibiotikatherapie. In dieser Arbeit wurden drei klinische Isolate der Spezies Pseudomonas
aeruginosa und Citrobacter freundii mittels diverser PCR-Techniken und Shotgun-
Klonierungsexperimenten auf das Vorhandensein von neuen Carbapenemasen hin untersucht.
Hierbei konnten drei neue Enzyme identifiziert werden: IMP-31, OXA-233 und KHM-2. Während
IMP-31 und KHM-2 Metallo-β-Laktamasen der molekularen Klasse B waren, stellte OXA-233 ein
OXA-10-ähnliches Mitglied der Klasse D β-Laktamasen dar. Durch die Charakterisierung der
genetischen Umgebung der drei neuen β-Laktamasegene konnte gezeigt werden, dass sowohl
das blaIMP-31- als auch das blaKHM-2-Gen Bestandteil eines Klasse 1 Integrons waren, während das
blaOXA-233-Gen nicht in einer derartigen genetischen Struktur vorlag. Mittels
Pulsfeldgelektrophoreseanalysen, gefolgt von Southern Blot-Hybridisierungen, konnte gezeigt
werden, dass die IMP-31- und KHM-2-kodierenden Gene waren auf dem Chromosom des
jeweiligen Isolats lokalisiert waren, was auf eine Transposon-vermittelte Integration schließen
ließ. Das OXA-233-kodierende Gen hingegen wurde auf dem konjugierbaren Plasmid pMB3018
lokalisiert, welches eine Größe von 52 kb aufwies. Dieses Plasmid wurde im Rahmen dieser
Arbeit mittels 454-Pyrosequencing komplett sequenziert und die Sequenzanalyse ergab, dass
dieses Plasmid zum IncN2-Inkompatibilitätstyps gehört. Da die in dieser Studie identifizierten
Gene in ihrem GC-Gehalt deutlich von dem der Spezies, in denen sie identifiziert wurden,
abwichen, muss davon ausgegangen werden, dass diese Gene von einer bislang unbekannten
Quelle in diese Spezies mobilisiert wurden. Der Einfluss von IMP-31, OXA-233 und KHM-2 auf
die Resistenz gegenüber β-Laktamantibiotika wurde mittels Bestimmung der minimalen
Hemmkonzentration (MHK) für Escherichia coli-Stämme, die die entsprechenden Enzyme
exprimierten, untersucht. Die Analyse ergab, dass alle drei Enzyme eine deutlich erhöhte
Resistenz gegen β-Laktame und damit auch gegen Carbapeneme vermittelten. Durch die
Bestimmung der kinetischen Parameter kcat und Km, welche ein Maß für die katalytische Effizienz
eines Enzyms darstellen, wurden die drei neuen Carbapenemasen und ihre jeweiligen
Referenzenzyme mittels FPLC aufgereinigt und in in vitro Hydrolyseuntersuchungen
biochemisch charakterisiert. Die Bestimmung der kinetischen Parameter zeigte, dass die drei
Enzyme eine deutliche Carbapenemaseaktivität aufwiesen und es damit höchst wahrscheinlich
ist, dass diese Enzyme eine hohe und damit klinisch relevante Carbapenemresistenz in Gram-
negativen Erregern vermitteln können. Für jedes der sechs Enzyme wurden die kinetischen
Parameter für zehn verschiedene Substrate mittels nichtlinearer Regression bestimmt und es

Zusammenfassung 114
zeigte sich, dass IMP-31 und KHM-2 in der Lage waren, nahezu alle Substrate mit hoher Effizienz
zu hydrolysieren, während OXA-233 keine hohe hydrolytische Aktivität gegenüber
Cephalosporinen besaß. Dies könnte auf eine mögliche Therapieoption hindeuten. Des Weiteren
war die Carbapenemhydrolyse von OXA-233 CO2-abhängig, was die erst vor Kurzem formulierte
Annahme, dass möglicherweise alle Klasse D β-Laktamasen eine CO2-abhängige
Carbapenemasefunktion besitzen, untermauerte. Die Identifizierung und Charakterisierung von
IMP-31, OXA-233 und KHM-2 im Rahmen dieser Arbeit ist ein weiteres Indiz für die
fortwährende Verbreitung und Diversifikation von Carbapenemasen in Gram-negativen Spezies
mit klinischer Relevanz.

Bibliography 115
7 Bibliography
Abraham EP, Chain E (1940) An enzyme from bacteria able to destroy penicillin. Nature, 146: 837. Abraham EP, Newton GG (1961) The structure of cephalesporin C. Biochem J, 79: 377-393. Abrahams A (1941) Chronic Miliary Tuberculosis. Proc R Soc Med, 35(1): 12. Ambler RP (1980) The structure of β-lactamases. Philos Trans R Soc Lond B Biol Sci, 289(1036): 321-331. Ambler RP, Coulson AF, Frere JM, Ghuysen JM, Joris B, Forsman M, Levesque RC, Tiraby G, Waley SG (1991) A standard numbering scheme for the class A β-lactamases. Biochem J, 276 ( Pt 1): 269-270. Antunes NT, Lamoureaux TL, Toth M, Stewart NK, Frase H, Vakulenko SB (2014) Class D β-lactamases: are they all carbapenemases? Antimicrob Agents Chemother, 58(4): 2119-2125. Appelbaum PC (2012) 2012 and beyond: potential for the start of a second pre-antibiotic era? J Antimicrob Chemother, 67(9): 2062-2068. Archer J, Robertson DL (2007) CTree: comparison of clusters between phylogenetic trees made easy. Bioinformatics, 23(21): 2952-2953. Arnold K, Bordoli L, Kopp J, Schwede T (2006) The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics, 22(2): 195-201. Aubert D, Poirel L, Ali AB, Goldstein FW, Nordmann P (2001) OXA-35 is an OXA-10-related β-lactamase from Pseudomonas aeruginosa. J Antimicrob Chemother, 48(5): 717-721. Aubron C, Poirel L, Ash RJ, Nordmann P (2005) Carbapenemase-producing Enterobacteriaceae, U.S. rivers. Emerg Infect Dis, 11(2): 260-264. Aziz RK et al. (2008) The RAST Server: rapid annotations using subsystems technology. BMC Genomics, 9: 75. Babic M, Hujer AM, Bonomo RA (2006) What's new in antibiotic resistance? Focus on β-lactamases. Drug Resist Updat, 9(3): 142-156. Balasubramanian D, Kong KF, Jayawardena SR, Leal SM, Sautter RT, Mathee K (2011) Co-regulation of {beta}-lactam resistance, alginate production and quorum sensing in Pseudomonas aeruginosa. J Med Microbiol, 60(Pt 2): 147-156. Bennett PM (2008) Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic resistance genes in bacteria. Br J Pharmacol, 153 Suppl 1: S347-357. Bennett PM, Chopra I (1993) Molecular basis of β-lactamase induction in bacteria. Antimicrob Agents Chemother, 37(2): 153-158. Biasini M et al. (2014) SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res, 42(Web Server issue): W252-258.

Bibliography 116
Blair JM, Webber MA, Baylay AJ, Ogbolu DO, Piddock LJ (2015) Molecular mechanisms of antibiotic resistance. Nat Rev Microbiol, 13(1): 42-51. Bonnet R (2004) Growing group of extended-spectrum β-lactamases: the CTX-M enzymes. Antimicrob Agents Chemother, 48(1): 1-14. Borgianni L, Prandi S, Salden L, Santella G, Hanson ND, Rossolini GM, Docquier JD (2011) Genetic context and biochemical characterization of the IMP-18 metallo-β-lactamase identified in a Pseudomonas aeruginosa isolate from the United States. Antimicrob Agents Chemother, 55(1): 140-145. Botelho J, Grosso F, Sousa C, Peixe L (2015) Characterization of a new genetic environment associated with GES-6 carbapenemase from a Pseudomonas aeruginosa isolate belonging to the high-risk clone ST235. J Antimicrob Chemother, 70(2): 615-617. Boucher HW, Talbot GH, Benjamin DK, Jr., Bradley J, Guidos RJ, Jones RN, Murray BE, Bonomo RA, Gilbert D (2013) 10 x '20 Progress--development of new drugs active against gram-negative bacilli: an update from the Infectious Diseases Society of America. Clin Infect Dis, 56(12): 1685-1694. Boucher Y, Labbate M, Koenig JE, Stokes HW (2007) Integrons: mobilizable platforms that promote genetic diversity in bacteria. Trends Microbiol, 15(7): 301-309. Breilh D, Texier-Maugein J, Allaouchiche B, Saux MC, Boselli E (2013) Carbapenems. J Chemother, 25(1): 1-17. Buchman JS, Schneider KD, Lloyd AR, Pavlish SL, Leonard DA (2012) Site-saturation mutagenesis of position V117 in OXA-1 β-lactamase: effect of side chain polarity on enzyme carboxylation and substrate turnover. Biochemistry, 51(14): 3143-3150. Burkhardt O, Derendorf H, Welte T (2007) Ertapenem: the new carbapenem 5 years after first FDA licensing for clinical practice. Expert Opin Pharmacother, 8(2): 237-256. Burrus V, Pavlovic G, Decaris B, Guedon G (2002) Conjugative transposons: the tip of the iceberg. Mol Microbiol, 46(3): 601-610. Bush K, Heep M, Macielag MJ, Noel GJ (2007) Anti-MRSA β-lactams in development, with a focus on ceftobiprole: the first anti-MRSA β-lactam to demonstrate clinical efficacy. Expert Opin Investig Drugs, 16(4): 419-429. Bush K, Jacoby GA (2010) Updated functional classification of β-lactamases. Antimicrob Agents Chemother, 54(3): 969-976. Bush K, Jacoby GA, Medeiros AA (1995) A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother, 39(6): 1211-1233. Cantón R et al. (2012a) Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin Microbiol Infect, 18(5): 413-431. Cantón R, Gonzalez-Alba JM, Galan JC (2012b) CTX-M Enzymes: Origin and Diffusion. Front Microbiol, 3: 110.

Bibliography 117
Carattoli A, Zankari E, Garcia-Fernandez A, Voldby Larsen M, Lund O, Villa L, Moller Aarestrup F, Hasman H (2014) In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother, 58(7): 3895-3903. Carlson JM, Chakravarty A, DeZiel CE, Gross RH (2007) SCOPE: a web server for practical de novo motif discovery. Nucleic Acids Res, 35(Web Server issue): W259-264. Carver T, Thomson N, Bleasby A, Berriman M, Parkhill J (2009) DNAPlotter: circular and linear interactive genome visualization. Bioinformatics, 25(1): 119-120. Castanheira M, Toleman MA, Jones RN, Schmidt FJ, Walsh TR (2004) Molecular characterization of a β-lactamase gene, blaGIM-1, encoding a new subclass of metallo-β-lactamase. Antimicrob Agents Chemother, 48(12): 4654-4661. Chen YT, Lin AC, Siu LK, Koh TH (2012) Sequence of closely related plasmids encoding blaNDM-1 in two unrelated Klebsiella pneumoniae isolates in Singapore. PLoS One, 7(11): e48737. Cholley P, Thouverez M, Hocquet D, van der Mee-Marquet N, Talon D, Bertrand X (2011) Most multidrug-resistant Pseudomonas aeruginosa isolates from hospitals in eastern France belong to a few clonal types. J Clin Microbiol, 49(7): 2578-2583. Cicek AC, Duzgun AO, Saral A, Sandalli C (2014) Determination of a novel integron-located variant (blaOXA-320 ) of Class D β-lactamase in Proteus mirabilis. J Basic Microbiol, 54(10): 1030-1035. Clowes RC, Rowley D (1954) Some observations on linkage effects in genetic recombination in Escherichia coli K-12. J Gen Microbiol, 11(2): 250-260. Collis CM, Hall RM (1995) Expression of antibiotic resistance genes in the integrated cassettes of integrons. Antimicrob Agents Chemother, 39(1): 155-162. Cornaglia G, Giamarellou H, Rossolini GM (2011) Metallo-β-lactamases: a last frontier for β-lactams? Lancet Infect Dis, 11(5): 381-393. Curran B, Jonas D, Grundmann H, Pitt T, Dowson CG (2004) Development of a multilocus sequence typing scheme for the opportunistic pathogen Pseudomonas aeruginosa. J Clin Microbiol, 42(12): 5644-5649. Cuzon G et al. (2010) Worldwide diversity of Klebsiella pneumoniae that produce β-lactamase blaKPC-2 gene. Emerg Infect Dis, 16(9): 1349-1356. Danel F, Hall LM, Duke B, Gur D, Livermore DM (1999) OXA-17, a further extended-spectrum variant of OXA-10 β-lactamase, isolated from Pseudomonas aeruginosa. Antimicrob Agents Chemother, 43(6): 1362-1366. Danel F, Hall LM, Gur D, Livermore DM (1995) OXA-14, another extended-spectrum variant of OXA-10 (PSE-2) β-lactamase from Pseudomonas aeruginosa. Antimicrob Agents Chemother, 39(8): 1881-1884. Danel F, Hall LM, Gur D, Livermore DM (1998) OXA-16, a further extended-spectrum variant of OXA-10 β-lactamase, from two Pseudomonas aeruginosa isolates. Antimicrob Agents Chemother, 42(12): 3117-3122. Darling AC, Mau B, Blattner FR, Perna NT (2004) Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res, 14(7): 1394-1403.

Bibliography 118
Darling AE, Mau B, Perna NT (2010) progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS One, 5(6): e11147. Darmon E, Leach DR (2014) Bacterial genome instability. Microbiol Mol Biol Rev, 78(1): 1-39. De Luca F, Benvenuti M, Carboni F, Pozzi C, Rossolini GM, Mangani S, Docquier JD (2011) Evolution to carbapenem-hydrolyzing activity in noncarbapenemase class D β-lactamase OXA-10 by rational protein design. Proc Natl Acad Sci U S A, 108(45): 18424-18429. Delver EP, Belogurov AA (1997) Organization of the leading region of IncN plasmid pKM101 (R46): a regulation controlled by CUP sequence elements. J Mol Biol, 271(1): 13-30. den Blaauwen T, de Pedro MA, Nguyen-Disteche M, Ayala JA (2008) Morphogenesis of rod-shaped sacculi. FEMS Microbiol Rev, 32(2): 321-344. Denome SA, Elf PK, Henderson TA, Nelson DE, Young KD (1999) Escherichia coli mutants lacking all possible combinations of eight penicillin binding proteins: viability, characteristics, and implications for peptidoglycan synthesis. J Bacteriol, 181(13): 3981-3993. Deshpande LM, Davies TA, Blandino G, Nicoletti G, Jones RN, Castanheira M (2013) IMP-33, a new IMP variant detected in Pseudomonas aeruginosa from Sicily. Antimicrob Agents Chemother, 57(12): 6401-6403. Deuschle U, Kammerer W, Gentz R, Bujard H (1986) Promoters of Escherichia coli: a hierarchy of in vivo strength indicates alternate structures. EMBO J, 5(11): 2987-2994. Diene SM, Rolain JM (2014) Carbapenemase genes and genetic platforms in Gram-negative bacilli: Enterobacteriaceae, Pseudomonas and Acinetobacter species. Clin Microbiol Infect, 20(9): 831-838. Doran TI (1999) The role of Citrobacter in clinical disease of children: review. Clin Infect Dis, 28(2): 384-394. Dramsi S, Magnet S, Davison S, Arthur M (2008) Covalent attachment of proteins to peptidoglycan. FEMS Microbiol Rev, 32(2): 307-320. Drawz SM, Bonomo RA (2010) Three decades of β-lactamase inhibitors. Clin Microbiol Rev, 23(1): 160-201. Duljasz W, Gniadkowski M, Sitter S, Wojna A, Jebelean C (2009) First organisms with acquired metallo-β-lactamases (IMP-13, IMP-22, and VIM-2) reported in Austria. Antimicrob Agents Chemother, 53(5): 2221-2222. European Centre for Disease Prevention and Control (2013). Antimicrobial resistance surveillance in Europe. http://www.ecdc.europa.eu/en/publications/Publications/antimicrobial-resistance-surveillance-europe-2013.pdf. English AR, Retsema JA, Girard AE, Lynch JE, Barth WE (1978) CP-45,899, a β-lactamase inhibitor that extends the antibacterial spectrum of β-lactams: initial bacteriological characterization. Antimicrob Agents Chemother, 14(3): 414-419.

Bibliography 119
European Committee on Antimicrobial Susceptibility Testing (2015). Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 5.0, 2015. http://www.eucast.org/clinical_breakpoints/ (15 March 2015, date last accessed). Evans BA, Amyes SG (2014) OXA β-lactamases. Clin Microbiol Rev, 27(2): 241-263. Fainstein V, LeBlanc B, Weaver S, Bodey GP (1982) A comparative in vitro study of thienamycin. Infection, 10(1): 50-52. Falagas ME, Bliziotis IA (2007) Pandrug-resistant Gram-negative bacteria: the dawn of the post-antibiotic era? Int J Antimicrob Agents, 29(6): 630-636. Falcone M, Mezzatesta ML, Perilli M, Forcella C, Giordano A, Cafiso V, Amicosante G, Stefani S, Venditti M (2009) Infections with VIM-1 metallo-β-lactamase-producing Enterobacter cloacae and their correlation with clinical outcome. J Clin Microbiol, 47(11): 3514-3519. Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG (2004) eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol, 186(5): 1518-1530. Fisher J, Belasco JG, Charnas RL, Khosla S, Knowles JR (1980) Β-lactamase inactivation by mechanism-based reagents. Philos Trans R Soc Lond B Biol Sci, 289(1036): 309-319. Fournier D, Hocquet D, Dehecq B, Cholley P, Plesiat P (2010a) Detection of a new extended-spectrum oxacillinase in Pseudomonas aeruginosa. J Antimicrob Chemother, 65(2): 364-365. Fournier PE et al. (2010b) Comprehensive diagnostic strategy for blood culture-negative endocarditis: a prospective study of 819 new cases. Clin Infect Dis, 51(2): 131-140. Franceschini N, Caravelli B, Docquier JD, Galleni M, Frere JM, Amicosante G, Rossolini GM (2000) Purification and biochemical characterization of the VIM-1 metallo-β-lactamase. Antimicrob Agents Chemother, 44(11): 3003-3007. Galani I, Rekatsina PD, Hatzaki D, Plachouras D, Souli M, Giamarellou H (2008) Evaluation of different laboratory tests for the detection of metallo-β-lactamase production in Enterobacteriaceae. J Antimicrob Chemother, 61(3): 548-553. Garau G, Garcia-Saez I, Bebrone C, Anne C, Mercuri P, Galleni M, Frere JM, Dideberg O (2004) Update of the standard numbering scheme for class B β-lactamases. Antimicrob Agents Chemother, 48(7): 2347-2349. Garcia-Alvarez L et al. (2011) Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: a descriptive study. Lancet Infect Dis, 11(8): 595-603. Gaynes R, Edwards JR (2005) Overview of nosocomial infections caused by Gram-negative bacilli. Clin Infect Dis, 41(6): 848-854. Gellatly SL, Hancock RE (2013) Pseudomonas aeruginosa: new insights into pathogenesis and host defenses. Pathog Dis, 67(3): 159-173. Georgopapadakou NH, Liu FY (1980) Penicillin-binding proteins in bacteria. Antimicrob Agents Chemother, 18(1): 148-157.

Bibliography 120
Ghuysen JM (1991) Serine β-lactamases and penicillin-binding proteins. Annu Rev Microbiol, 45: 37-67. Gillings MR (2014) Integrons: past, present, and future. Microbiol Mol Biol Rev, 78(2): 257-277. Golemi D, Maveyraud L, Vakulenko S, Samama JP, Mobashery S (2001) Critical involvement of a carbamylated lysine in catalytic function of class D β-lactamases. Proc Natl Acad Sci U S A, 98(25): 14280-14285. Guex N, Peitsch MC, Schwede T (2009) Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: a historical perspective. Electrophoresis, 30 Suppl 1: S162-173. Gupta V (2008a) Metallo beta lactamases in Pseudomonas aeruginosa and Acinetobacter species. Expert Opin Investig Drugs, 17(2): 131-143. Gupta V (2008b) Metallo β-lactamases in Pseudomonas aeruginosa and Acinetobacter species. Expert Opin Investig Drugs, 17(2): 131-143. Hall LM, Livermore DM, Gur D, Akova M, Akalin HE (1993) OXA-11, an extended-spectrum variant of OXA-10 (PSE-2) β-lactamase from Pseudomonas aeruginosa. Antimicrob Agents Chemother, 37(8): 1637-1644. Hall RM, Brookes DE, Stokes HW (1991) Site-specific insertion of genes into integrons: role of the 59-base element and determination of the recombination cross-over point. Mol Microbiol, 5(8): 1941-1959. Hansen F et al. (2014) Characterization of carbapenem nonsusceptible Pseudomonas aeruginosa in Denmark: a nationwide, prospective study. Microb Drug Resist, 20(1): 22-29. Hanson ND, Hossain A, Buck L, Moland ES, Thomson KS (2006) First occurrence of a Pseudomonas aeruginosa isolate in the United States producing an IMP metallo-β-lactamase, IMP-18. Antimicrob Agents Chemother, 50(6): 2272-2273. Harris F, Brandenburg K, Seydel U, Phoenix D (2002) Investigations into the mechanisms used by the C-terminal anchors of Escherichia coli penicillin-binding proteins 4, 5, 6 and 6b for membrane interaction. Eur J Biochem, 269(23): 5821-5829. Hartman BJ, Tomasz A (1984) Low-affinity penicillin-binding protein associated with β-lactam resistance in Staphylococcus aureus. J Bacteriol, 158(2): 513-516. Hashizume T, Ishino F, Nakagawa J, Tamaki S, Matsuhashi M (1984) Studies on the mechanism of action of imipenem (N-formimidoylthienamycin) in vitro: binding to the penicillin-binding proteins (PBPs) in Escherichia coli and Pseudomonas aeruginosa, and inhibition of enzyme activities due to the PBPs in E. coli. J Antibiot (Tokyo), 37(4): 394-400. Hocquet D, Colomb M, Dehecq B, Belmonte O, Courvalin P, Plesiat P, Meziane-Cherif D (2011) Ceftazidime-hydrolysing β-lactamase OXA-145 with impaired hydrolysis of penicillins in Pseudomonas aeruginosa. J Antimicrob Chemother, 66(8): 1745-1750. Hoffmann A (2013) Identification and characterization of the novel β-lactamase KHM-2. Master thesis, Ruhr-University Bochum. Holmes DS, Quigley M (1981) A rapid boiling method for the preparation of bacterial plasmids. Anal Biochem, 114(1): 193-197.

Bibliography 121
Iyobe S, Kusadokoro H, Takahashi A, Yomoda S, Okubo T, Nakamura A, O'Hara K (2002) Detection of a variant metallo-β-lactamase, IMP-10, from two unrelated strains of Pseudomonas aeruginosa and an Alcaligenes xylosoxidans strain. Antimicrob Agents Chemother, 46(6): 2014-2016. Jacoby GA (2009) AmpC β-lactamases. Clin Microbiol Rev, 22(1): 161-182, Table of Contents. Janda JM, Abbot SL (2006). The Enterobacteria, Second Edition. ASM Press, American Society for Microbiology. Jeannot K, Poirel L, Robert-Nicoud M, Cholley P, Nordmann P, Plesiat P (2012) IMP-29, a novel IMP-type metallo-β-lactamase in Pseudomonas aeruginosa. Antimicrob Agents Chemother, 56(4): 2187-2190. Johnson AP, Woodford N (2013) Global spread of antibiotic resistance: the example of New Delhi metallo-β-lactamase (NDM)-mediated carbapenem resistance. J Med Microbiol, 62(Pt 4): 499-513. Juan C, Beceiro A, Gutierrez O, Alberti S, Garau M, Perez JL, Bou G, Oliver A (2008) Characterization of the new metallo-β-lactamase VIM-13 and its integron-borne gene from a Pseudomonas aeruginosa clinical isolate in Spain. Antimicrob Agents Chemother, 52(10): 3589-3596. Kellis JT, Jr., Nyberg K, Sali D, Fersht AR (1988) Contribution of hydrophobic interactions to protein stability. Nature, 333(6175): 784-786. Kelly JA et al. (1986) On the origin of bacterial resistance to penicillin: comparison of a β-lactamase and a penicillin target. Science, 231(4744): 1429-1431. Kiefer F, Arnold K, Kunzli M, Bordoli L, Schwede T (2009) The SWISS-MODEL Repository and associated resources. Nucleic Acids Res, 37(Database issue): D387-392. Kim ES, Hooper DC (2014) Clinical importance and epidemiology of quinolone resistance. Infect Chemother, 46(4): 226-238. Klucar L, Stano M, Hajduk M (2010) phiSITE: database of gene regulation in bacteriophages. Nucleic Acids Res, 38(Database issue): D366-370. Kong KF, Schneper L, Mathee K (2010) Β-lactam antibiotics: from antibiosis to resistance and bacteriology. APMIS, 118(1): 1-36. Kong V, Beckert L, Awunor-Renner C (2009) A case of beta lactam-induced visual hallucination. N Z Med J, 122(1298): 76-77. Korfhagen TR, Loper JC, Ferrel JA (1975) Pseudomonas aeruginosa R factors determining gentamicin plus carbenicillin resistance from patients with urinary tract colonizations. Antimicrob Agents Chemother, 7(1): 64-68. Kropp H, Sundelof JG, Hajdu R, Kahan FM (1982) Metabolism of thienamycin and related carbapenem antibiotics by the renal dipeptidase, dehydropeptidase. Antimicrob Agents Chemother, 22(1): 62-70.

Bibliography 122
Labischinski H, Goodell EW, Goodell A, Hochberg ML (1991) Direct proof of a "more-than-single-layered" peptidoglycan architecture of Escherichia coli W7: a neutron small-angle scattering study. J Bacteriol, 173(2): 751-756. Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227(5259): 680-685. Lange F (2014) Cloning and characterization of a new variant of the OXA-10 β-lactamase exhibiting carbapenemase-activity from Klebsiella pneumoniae. Master thesis, Ruhr-University Bochum. Laraki N, Franceschini N, Rossolini GM, Santucci P, Meunier C, de Pauw E, Amicosante G, Frere JM, Galleni M (1999) Biochemical characterization of the Pseudomonas aeruginosa 101/1477 metallo-β-lactamase IMP-1 produced by Escherichia coli. Antimicrob Agents Chemother, 43(4): 902-906. Larché J et al. (2012) Rapid identification of international multidrug-resistant Pseudomonas aeruginosa clones by multiple-locus variable number of tandem repeats analysis and investigation of their susceptibility to lytic bacteriophages. Antimicrob Agents Chemother, 56(12): 6175-6180. Lee K, Yum JH, Yong D, Lee HM, Kim HD, Docquier JD, Rossolini GM, Chong Y (2005) Novel acquired metallo-β-lactamase gene, blaSIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob Agents Chemother, 49(11): 4485-4491. Lee MF, Peng CF, Hsu HJ, Chen YH (2008) Molecular characterisation of the metallo-β-lactamase genes in imipenem-resistant Gram-negative bacteria from a university hospital in southern Taiwan. Int J Antimicrob Agents, 32(6): 475-480. Leonard DA, Bonomo RA, Powers RA (2013) Class D β-lactamases: a reappraisal after five decades. Acc Chem Res, 46(11): 2407-2415. Levast M, Poirel L, Carrer A, Deiber M, Decroisette E, Mallaval FO, Lecomte C, Nordmann P (2011) Transfer of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae from Turkey to France. J Antimicrob Chemother, 66(4): 944-945. Levesque C, Piche L, Larose C, Roy PH (1995) PCR mapping of integrons reveals several novel combinations of resistance genes. Antimicrob Agents Chemother, 39(1): 185-191. Libisch B, Balogh B, Fuzi M (2009) Identification of two multidrug-resistant Pseudomonas aeruginosa clonal lineages with a countrywide distribution in Hungary. Curr Microbiol, 58(2): 111-116. Libisch B et al. (2008) Identification of PER-1 extended-spectrum β-lactamase producing Pseudomonas aeruginosa clinical isolates of the international clonal complex CC11 from Hungary and Serbia. FEMS Immunol Med Microbiol, 54(3): 330-338. Lim D, Strynadka NC (2002) Structural basis for the beta lactam resistance of PBP2a from methicillin-resistant Staphylococcus aureus. Nat Struct Biol, 9(11): 870-876. Liu CP, Chen TL, Wang NY, Chow SF, Lin JC, Yan TR (2014) Detection of Pseudomonas aeruginosa isolates carrying the blaOXA-142 extended-spectrum β-lactamase gene in Taiwan. J Microbiol Immunol Infect.

Bibliography 123
Liu SL, Hessel A, Sanderson KE (1993) Genomic mapping with I-Ceu I, an intron-encoded endonuclease specific for genes for ribosomal RNA, in Salmonella spp., Escherichia coli, and other bacteria. Proc Natl Acad Sci U S A, 90(14): 6874-6878. Livermore DM (1995) β-Lactamases in laboratory and clinical resistance. Clin Microbiol Rev, 8(4): 557-584. Livermore DM (2001) Of Pseudomonas, porins, pumps and carbapenems. J Antimicrob Chemother, 47(3): 247-250. Loli A, Tzouvelekis LS, Tzelepi E, Carattoli A, Vatopoulos AC, Tassios PT, Miriagou V (2006) Sources of diversity of carbapenem resistance levels in Klebsiella pneumoniae carrying blaVIM-1. J Antimicrob Chemother, 58(3): 669-672. Long KS, Poehlsgaard J, Kehrenberg C, Schwarz S, Vester B (2006) The Cfr rRNA methyltransferase confers resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics. Antimicrob Agents Chemother, 50(7): 2500-2505. Maatallah M et al. (2011) Population structure of Pseudomonas aeruginosa from five Mediterranean countries: evidence for frequent recombination and epidemic occurrence of CC235. PLoS One, 6(10): e25617. Maltezou HC (2009) Metallo-β-lactamases in Gram-negative bacteria: introducing the era of pan-resistance? Int J Antimicrob Agents, 33(5): 405 e401-407. Mascaretti OA (2003). Bacteria versus Antibacterial Agents - An integrated approach. ASM Press, American Society for Microbiology. Massova I, Mobashery S (1998) Kinship and diversification of bacterial penicillin-binding proteins and β-lactamases. Antimicrob Agents Chemother, 42(1): 1-17. Matias VR, Al-Amoudi A, Dubochet J, Beveridge TJ (2003) Cryo-transmission electron microscopy of frozen-hydrated sections of Escherichia coli and Pseudomonas aeruginosa. J Bacteriol, 185(20): 6112-6118. Mazel D (2006) Integrons: agents of bacterial evolution. Nat Rev Microbiol, 4(8): 608-620. Medeiros AA, Cohenford M, Jacoby GA (1985) Five novel plasmid-determined β-lactamases. Antimicrob Agents Chemother, 27(5): 715-719. Meining L (2012) Biochemical characterization of the novel β-lactamase OXA-233 and analysis of the genetic context of the blaOXA gene. Master thesis, University of Duisburg-Essen. The study was performed at the Ruhr-University Bochum. Mengin-Lecreulx D, Lemaitre B (2005) Structure and metabolism of peptidoglycan and molecular requirements allowing its detection by the Drosophila innate immune system. J Endotoxin Res, 11(2): 105-111. Meziane-Cherif D, Lambert T, Dupechez M, Courvalin P, Galimand M (2008) Genetic and biochemical characterization of OXA-63, a new class D β-lactamase from Brachyspira pilosicoli BM4442. Antimicrob Agents Chemother, 52(4): 1264-1268.

Bibliography 124
Miller AK, Celozzi E, Kong Y, Pelak BA, Kropp H, Stapley EO, Hendlin D (1972) Cephamycins, a new family of β-lactam antibiotics. IV. In vivo studies. Antimicrob Agents Chemother, 2(4): 287-290. Mimoz O, Gregoire N, Poirel L, Marliat M, Couet W, Nordmann P (2012) Broad-spectrum β-lactam antibiotics for treating experimental peritonitis in mice due to Klebsiella pneumoniae producing the carbapenemase OXA-48. Antimicrob Agents Chemother, 56(5): 2759-2760. Moali C, Anne C, Lamotte-Brasseur J, Groslambert S, Devreese B, Van Beeumen J, Galleni M, Frere JM (2003) Analysis of the importance of the metallo-β-lactamase active site loop in substrate binding and catalysis. Chem Biol, 10(4): 319-329. Mora-Rillo M, Loeches-Yagüe B, Muñoz-Velez M, Gomez-Gil R, Paño-Pardo JR, Arribas JR, García-Rodriguez J (2014) Potential Utility of Ceftazidime-Avibactam for OXA-48-producing Enterobacteriaceae: Preliminary in vitro and in vivo data. Interscience Conference of Antimicrobial Agents and Chemotherapy (ICAAC), Abstract F-955. Moura A, Pereira C, Henriques I, Correia A (2012) Novel gene cassettes and integrons in antibiotic-resistant bacteria isolated from urban wastewaters. Res Microbiol, 163(2): 92-100. Moyo S, Haldorsen B, Aboud S, Blomberg B, Maselle SY, Sundsfjord A, Langeland N, Samuelsen O (2015) Identification of VIM-2-producing Pseudomonas aeruginosa from Tanzania is associated with sequence types 244 and 640 and the location of blaVIM-2 in a TniC integron. Antimicrob Agents Chemother, 59(1): 682-685. Mulvey MR, Boyd DA, Baker L, Mykytczuk O, Reis EM, Asensi MD, Rodrigues DP, Ng LK (2004) Characterization of a Salmonella enterica serovar Agona strain harbouring a class 1 integron containing novel OXA-type β-lactamase (blaOXA-53) and 6'-N-aminoglycoside acetyltransferase genes [aac(6')-I30]. J Antimicrob Chemother, 54(2): 354-359. Naas T, Sougakoff W, Casetta A, Nordmann P (1998) Molecular characterization of OXA-20, a novel class D β-lactamase, and its integron from Pseudomonas aeruginosa. Antimicrob Agents Chemother, 42(8): 2074-2083. Nemec A, Krizova L, Maixnerova M, Musilek M (2010) Multidrug-resistant epidemic clones among bloodstream isolates of Pseudomonas aeruginosa in the Czech Republic. Res Microbiol, 161(3): 234-242. Netikul T, Sidjabat HE, Paterson DL, Kamolvit W, Tantisiriwat W, Steen JA, Kiratisin P (2014) Characterization of an IncN2-type blaNDM-1-carrying plasmid in Escherichia coli ST131 and Klebsiella pneumoniae ST11 and ST15 isolates in Thailand. J Antimicrob Chemother, 69(11): 3161-3163. Neuhaus FC, Baddiley J (2003) A continuum of anionic charge: structures and functions of D-alanyl-teichoic acids in gram-positive bacteria. Microbiol Mol Biol Rev, 67(4): 686-723. Neuwirth C, Siebor E, Robin F, Bonnet R (2007) First occurrence of an IMP metallo-β-lactamase in Aeromonas caviae: IMP-19 in an isolate from France. Antimicrob Agents Chemother, 51(12): 4486-4488. Nordmann P, Dortet L, Poirel L (2012) Carbapenem resistance in Enterobacteriaceae: here is the storm! Trends Mol Med, 18(5): 263-272. Nordmann P, Naas T (2010). Chapter 14. β-Lactams and Pseudomonas aeruginosa. Antibiogramm. EKSA Publishing, ASM Press.

Bibliography 125
Nordmann P, Poirel L (2014) The difficult-to-control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clin Microbiol Infect, 20(9): 821-830. Norrby SR et al. (1983) Urinary recovery of N-formimidoyl thienamycin (MK0787) as affected by coadministration of N-formimidoyl thienamycin dehydropeptidase inhibitors. Antimicrob Agents Chemother, 23(2): 300-307. Ohlasova D, Kmet V, Niks M (2007) First report of the carbapenem-resistant Pseudomonas aeruginosa producing IMP-7 metallo-β-lactamase in Slovakia. Int J Antimicrob Agents, 30(4): 370-371. Overbeek R et al. (2014) The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res, 42(Database issue): D206-214. Pace CN et al. (2011) Contribution of hydrophobic interactions to protein stability. J Mol Biol, 408(3): 514-528. Palzkill T (2013) Metallo-β-lactamase structure and function. Ann N Y Acad Sci, 1277: 91-104. Papagiannitsis CC, Tzouvelekis LS, Miriagou V (2009) Relative strengths of the class 1 integron promoter hybrid 2 and the combinations of strong and hybrid 1 with an active p2 promoter. Antimicrob Agents Chemother, 53(1): 277-280. Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA (2011) Carbapenems: past, present, and future. Antimicrob Agents Chemother, 55(11): 4943-4960. Partridge SR, Paulsen IT, Iredell JR (2012) pJIE137 carrying blaCTX-M-62 is closely related to p271A carrying blaNDM-1. Antimicrob Agents Chemother, 56(4): 2166-2168. Pasteran FG, Otaegui L, Guerriero L, Radice G, Maggiora R, Rapoport M, Faccone D, Di Martino A, Galas M (2008) Klebsiella pneumoniae Carbapenemase-2, Buenos Aires, Argentina. Emerg Infect Dis, 14(7): 1178-1180. Paterson DL, Depestel DD (2009) Doripenem. Clin Infect Dis, 49(2): 291-298. Perez-Llarena FJ et al. (2012) Characterization of a novel IMP-28 metallo-β-lactamase from a Spanish Klebsiella oxytoca clinical isolate. Antimicrob Agents Chemother, 56(8): 4540-4543. Petersen TN, Brunak S, von Heijne G, Nielsen H (2011) SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Methods, 8(10): 785-786. Pfeifer Y, Wilharm G, Zander E, Wichelhaus TA, Gottig S, Hunfeld KP, Seifert H, Witte W, Higgins PG (2011) Molecular characterization of blaNDM-1 in an Acinetobacter baumannii strain isolated in Germany in 2007. J Antimicrob Chemother, 66(9): 1998-2001. Philippon AM, Paul GC, Jacoby GA (1983) Properties of PSE-2 β-lactamase and genetic basis for its production in Pseudomonas aeruginosa. Antimicrob Agents Chemother, 24(3): 362-369. Philippon LN, Naas T, Bouthors AT, Barakett V, Nordmann P (1997) OXA-18, a class D clavulanic acid-inhibited extended-spectrum β-lactamase from Pseudomonas aeruginosa. Antimicrob Agents Chemother, 41(10): 2188-2195. Piddock LJ (2006) Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin Microbiol Rev, 19(2): 382-402.

Bibliography 126
Pitout JD, Gregson DB, Poirel L, McClure JA, Le P, Church DL (2005) Detection of Pseudomonas aeruginosa producing metallo-β-lactamases in a large centralized laboratory. J Clin Microbiol, 43(7): 3129-3135. Poirel L, Bonnin RA, Nordmann P (2011) Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob Agents Chemother, 55(9): 4224-4229. Poirel L, Bonnin RA, Nordmann P (2012a) Genetic features of the widespread plasmid coding for the carbapenemase OXA-48. Antimicrob Agents Chemother, 56(1): 559-562. Poirel L, Figueiredo S, Cattoir V, Carattoli A, Nordmann P (2008) Acinetobacter radioresistens as a silent source of carbapenem resistance for Acinetobacter spp. Antimicrob Agents Chemother, 52(4): 1252-1256. Poirel L, Girlich D, Naas T, Nordmann P (2001) OXA-28, an extended-spectrum variant of OXA-10 β-lactamase from Pseudomonas aeruginosa and its plasmid- and integron-located gene. Antimicrob Agents Chemother, 45(2): 447-453. Poirel L, Heritier C, Nordmann P (2004a) Chromosome-encoded ambler class D β-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob Agents Chemother, 48(1): 348-351. Poirel L, Heritier C, Tolun V, Nordmann P (2004b) Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob Agents Chemother, 48(1): 15-22. Poirel L, Naas T, Nicolas D, Collet L, Bellais S, Cavallo JD, Nordmann P (2000) Characterization of VIM-2, a carbapenem-hydrolyzing metallo-β-lactamase and its plasmid- and integron-borne gene from a Pseudomonas aeruginosa clinical isolate in France. Antimicrob Agents Chemother, 44(4): 891-897. Poirel L, Naas T, Nordmann P (2010) Diversity, epidemiology, and genetics of class D β-lactamases. Antimicrob Agents Chemother, 54(1): 24-38. Poirel L, Potron A, Nordmann P (2012b) OXA-48-like carbapenemases: the phantom menace. J Antimicrob Chemother, 67(7): 1597-1606. Pollini S, Maradei S, Pecile P, Olivo G, Luzzaro F, Docquier JD, Rossolini GM (2013) FIM-1, a new acquired metallo-β-lactamase from a Pseudomonas aeruginosa clinical isolate from Italy. Antimicrob Agents Chemother, 57(1): 410-416. Pournaras S, Kock R, Mossialos D, Mellmann A, Sakellaris V, Stathopoulos C, Friedrich AW, Tsakris A (2013) Detection of a phylogenetically distinct IMP-type metallo-β-lactamase, IMP-35, in a CC235 Pseudomonas aeruginosa from the Dutch-German border region (Euregio). J Antimicrob Chemother, 68(6): 1271-1276. Pradel N, Delmas J, Wu LF, Santini CL, Bonnet R (2009) Sec- and Tat-dependent translocation of β-lactamases across the Escherichia coli inner membrane. Antimicrob Agents Chemother, 53(1): 242-248. Queenan AM, Bush K (2007) Carbapenemases: the versatile β-lactamases. Clin Microbiol Rev, 20(3): 440-458, table of contents.

Bibliography 127
Radstrom P, Skold O, Swedberg G, Flensburg J, Roy PH, Sundstrom L (1994) Transposon Tn5090 of plasmid R751, which carries an integron, is related to Tn7, Mu, and the retroelements. J Bacteriol, 176(11): 3257-3268. Rajeshwari R, Sonti RV (2000) Stationary-phase variation due to transposition of novel insertion elements in Xanthomonas oryzae pv. oryzae. J Bacteriol, 182(17): 4797-4802. Ramirez MS, Tolmasky ME (2010) Aminoglycoside modifying enzymes. Drug Resist Updat, 13(6): 151-171. Rasmussen BA, Keeney D, Yang Y, Bush K (1994) Cloning and expression of a cloxacillin-hydrolyzing enzyme and a cephalosporinase from Aeromonas sobria AER 14M in Escherichia coli: requirement for an E. coli chromosomal mutation for efficient expression of the class D enzyme. Antimicrob Agents Chemother, 38(9): 2078-2085. Reading C, Cole M (1977) Clavulanic acid: a β-lactamase-inhiting β-lactam from Streptomyces clavuligerus. Antimicrob Agents Chemother, 11(5): 852-857. Riccio ML, Franceschini N, Boschi L, Caravelli B, Cornaglia G, Fontana R, Amicosante G, Rossolini GM (2000) Characterization of the metallo-β-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of blaIMP allelic variants carried by gene cassettes of different phylogeny. Antimicrob Agents Chemother, 44(5): 1229-1235. Root JL, McIntyre OR, Jacobs NJ, Daghlian CP (1988) Inhibitory effect of disodium EDTA upon the growth of Staphylococcus epidermidis in vitro: relation to infection prophylaxis of Hickman catheters. Antimicrob Agents Chemother, 32(11): 1627-1631. Rossolini GM, Docquier JD (2007). Chapter 9. Class B β-Lactamases. Enzyme-Mediated Resistance to Antibiotics, ASM Press. Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT, Mullis KB, Erlich HA (1988) Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science, 239(4839): 487-491. Samuelsen O, Buaro L, Toleman MA, Giske CG, Hermansen NO, Walsh TR, Sundsfjord A (2009) The first metallo-β-lactamase identified in norway is associated with a TniC-like transposon in a Pseudomonas aeruginosa isolate of sequence type 233 imported from Ghana. Antimicrob Agents Chemother, 53(1): 331-332. Samuelsen O et al. (2010) Molecular epidemiology of metallo-β-lactamase-producing Pseudomonas aeruginosa isolates from Norway and Sweden shows import of international clones and local clonal expansion. Antimicrob Agents Chemother, 54(1): 346-352. Sanbongi Y, Shimizu A, Suzuki T, Nagaso H, Ida T, Maebashi K, Gotoh N (2009) Classification of OprD sequence and correlation with antimicrobial activity of carbapenem agents in Pseudomonas aeruginosa clinical isolates collected in Japan. Microbiol Immunol, 53(7): 361-367. Santella G, Docquier JD, Gutkind G, Rossolini GM, Radice M (2011) Purification and biochemical characterization of IMP-13 metallo-β-lactamase. Antimicrob Agents Chemother, 55(1): 399-401. Saravolatz LD, Stein GE, Johnson LB (2011) Ceftaroline: a novel cephalosporin with activity against methicillin-resistant Staphylococcus aureus. Clin Infect Dis, 52(9): 1156-1163.

Bibliography 128
Sardelic S, Bedenic B, Colinon-Dupuich C, Orhanovic S, Bosnjak Z, Plecko V, Cournoyer B, Rossolini GM (2012) Infrequent finding of metallo-β-lactamase VIM-2 in carbapenem-resistant Pseudomonas aeruginosa strains from Croatia. Antimicrob Agents Chemother, 56(5): 2746-2749. Sauvage E, Kerff F, Terrak M, Ayala JA, Charlier P (2008) The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis. FEMS Microbiol Rev, 32(2): 234-258. Schiffer G, Holtje JV (1999) Cloning and characterization of PBP 1C, a third member of the multimodular class A penicillin-binding proteins of Escherichia coli. J Biol Chem, 274(45): 32031-32039. Schneider I, Keuleyan E, Rasshofer R, Markovska R, Queenan AM, Bauernfeind A (2008) VIM-15 and VIM-16, two new VIM-2-like metallo-β-lactamases in Pseudomonas aeruginosa isolates from Bulgaria and Germany. Antimicrob Agents Chemother, 52(8): 2977-2979. Schneider KD, Bethel CR, Distler AM, Hujer AM, Bonomo RA, Leonard DA (2009) Mutation of the active site carboxy-lysine (K70) of OXA-1 β-lactamase results in a deacylation-deficient enzyme. Biochemistry, 48(26): 6136-6145. Sekiguchi J, Morita K, Kitao T, Watanabe N, Okazaki M, Miyoshi-Akiyama T, Kanamori M, Kirikae T (2008) KHM-1, a novel plasmid-mediated metallo-β-lactamase from a Citrobacter freundii clinical isolate. Antimicrob Agents Chemother, 52(11): 4194-4197. Sekizuka T, Matsui M, Yamane K, Takeuchi F, Ohnishi M, Hishinuma A, Arakawa Y, Kuroda M (2011) Complete sequencing of the blaNDM-1-positive IncA/C plasmid from Escherichia coli ST38 isolate suggests a possible origin from plant pathogens. PLoS One, 6(9): e25334. Seok Y, Bae IK, Jeong SH, Kim SH, Lee H, Lee K (2011) Dissemination of IMP-6 metallo-β-lactamase-producing Pseudomonas aeruginosa sequence type 235 in Korea. J Antimicrob Chemother, 66(12): 2791-2796. Shen J, Wang Y, Schwarz S (2013) Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J Antimicrob Chemother, 68(8): 1697-1706. Shigemoto N et al. (2013) A novel metallo-β-lactamase, IMP-34, in Klebsiella isolates with decreased resistance to imipenem. Diagn Microbiol Infect Dis, 76(1): 119-121. Shimizu W et al. (2015) Persistence and epidemic propagation of a Pseudomonas aeruginosa clone ST235 harboring an IS26-composite transposon carrying the blaIMP-1-integron in Hiroshima, 2005-2012. Antimicrob Agents Chemother. Siguier P, Gourbeyre E, Chandler M (2014) Bacterial insertion sequences: their genomic impact and diversity. FEMS Microbiol Rev, 38(5): 865-891. Silhavy TJ, Kahne D, Walker S (2010) The bacterial cell envelope. Cold Spring Harb Perspect Biol, 2(5): a000414. Singh GS (2004) Β-lactams in the new millennium. Part-I: monobactams and carbapenems. Mini Rev Med Chem, 4(1): 69-92. Slekovec C, Plantin J, Cholley P, Thouverez M, Talon D, Bertrand X, Hocquet D (2012) Tracking down antibiotic-resistant Pseudomonas aeruginosa isolates in a wastewater network. PLoS One, 7(12): e49300.

Bibliography 129
Smith Moland E, Hanson ND, Herrera VL, Black JA, Lockhart TJ, Hossain A, Johnson JA, Goering RV, Thomson KS (2003) Plasmid-mediated, carbapenem-hydrolysing β-lactamase, KPC-2, in Klebsiella pneumoniae isolates. J Antimicrob Chemother, 51(3): 711-714. Snyder L, Champness W (2007). Molecular genetics of bacteria, Third Edition. ASM Press, American Society for Microbiology. Solovyev V, Salamov A (2011). Chapter 4. Automatic Annotation of Microbial Genomes and Metagenomic Sequences. Metagenomics and its Applications in Agriculture, Biomedicine and Environmental Studies. Nova Science Publishers. Spratt BG (1975) Distinct penicillin binding proteins involved in the division, elongation, and shape of Escherichia coli K12. Proc Natl Acad Sci U S A, 72(8): 2999-3003. Spratt BG, Jobanputra V (1977) Mutants of Escherichia coli which lack a component of penicillin-binding protein 1 are viable. FEBS Lett, 79(2): 374-378. Spratt BG, Pardee AB (1975) Penicillin-binding proteins and cell shape in Escherichia coli. Nature, 254(5500): 516-517. Suetens C, Hopkins S, Kolman J, Diaz Högberg L (2013) Point prevalence survey of healthcare associated infections and antimicrobial use in European acute care hospitals 2011-2012, European Centre for Disease Prevention and Control. http://www.ecdc.europa.eu/en/publications/Publications/healthcare-associated-infections-antimicrobial-use-PPS.pdf. Suginaka H, Blumberg PM, Strominger JL (1972) Multiple penicillin-binding components in Bacillus subtilis, Bacillus cereus, Staphylococcus aureus, and Escherichia coli. J Biol Chem, 247(17): 5279-5288. Tally FP, Jacobus NV, Gorbach SL (1978) In vitro activity of thienamycin. Antimicrob Agents Chemother, 14(3): 436-438. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol, 30(12): 2725-2729. Tato M, Coque TM, Baquero F, Canton R (2010) Dispersal of carbapenemase blaVIM-1 gene associated with different Tn402 variants, mercury transposons, and conjugative plasmids in Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob Agents Chemother, 54(1): 320-327. Toleman MA, Biedenbach D, Bennett D, Jones RN, Walsh TR (2003a) Genetic characterization of a novel metallo-β-lactamase gene, blaIMP-13, harboured by a novel Tn5051-type transposon disseminating carbapenemase genes in Europe: report from the SENTRY worldwide antimicrobial surveillance programme. J Antimicrob Chemother, 52(4): 583-590. Toleman MA, Biedenbach D, Bennett DM, Jones RN, Walsh TR (2005) Italian metallo-β-lactamases: a national problem? Report from the SENTRY Antimicrobial Surveillance Programme. J Antimicrob Chemother, 55(1): 61-70. Toleman MA, Rolston K, Jones RN, Walsh TR (2003b) Molecular and biochemical characterization of OXA-45, an extended-spectrum class 2d' β-lactamase in Pseudomonas aeruginosa. Antimicrob Agents Chemother, 47(9): 2859-2863.

Bibliography 130
Toleman MA, Vinodh H, Sekar U, Kamat V, Walsh TR (2007) blaVIM-2-harboring integrons isolated in India, Russia, and the United States arise from an ancestral class 1 integron predating the formation of the 3' conserved sequence. Antimicrob Agents Chemother, 51(7): 2636-2638. Tysall L, Stockdale MW, Chadwick PR, Palepou MF, Towner KJ, Livermore DM, Woodford N (2002) IMP-1 carbapenemase detected in an Acinetobacter clinical isolate from the UK. J Antimicrob Chemother, 49(1): 217-218. Valot B, Rohmer L, Jacobs MA, Miller SI, Bertrand X, Hocquet D (2014) Comparative Genomic Analysis of Two Multidrug-Resistant Clinical Isolates of ST395 Epidemic Strain of Pseudomonas aeruginosa Obtained 12 Years Apart. Genome Announc, 2(3). Vatcheva-Dobrevska R, Mulet X, Ivanov I, Zamorano L, Dobreva E, Velinov T, Kantardjiev T, Oliver A (2013) Molecular epidemiology and multidrug resistance mechanisms of Pseudomonas aeruginosa isolates from Bulgarian hospitals. Microb Drug Resist, 19(5): 355-361. Vollmer W, Blanot D, de Pedro MA (2008) Peptidoglycan structure and architecture. FEMS Microbiol Rev, 32(2): 149-167. Voulgari E, Poulou A, Koumaki V, Tsakris A (2013) Carbapenemase-producing Enterobacteriaceae: now that the storm is finally here, how will timely detection help us fight back? Future Microbiol, 8(1): 27-39. Walsh TR (2010) Emerging carbapenemases: a global perspective. Int J Antimicrob Agents, 36 Suppl 3: S8-14. Walsh TR, Hall L, MacGowan AP, Bennett PM (1995) Sequence analysis of two chromosomally mediated inducible β-lactamases from Aeromonas sobria, strain 163a, one a class D penicillinase, the other an AmpC cephalosporinase. J Antimicrob Chemother, 36(1): 41-52. Walsh TR, Toleman MA, Poirel L, Nordmann P (2005) Metallo-β-lactamases: the quiet before the storm? Clin Microbiol Rev, 18(2): 306-325. Watanabe M, Iyobe S, Inoue M, Mitsuhashi S (1991) Transferable imipenem resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother, 35(1): 147-151. Weaver SS, Bodey GP, LeBlanc BM (1979) Thienamycin: new β-lactam antibiotic with potent broad-spectrum activity. Antimicrob Agents Chemother, 15(4): 518-521. Weldhagen GF, Prinsloo A (2004) Molecular detection of GES-2 extended spectrum β-lactamase producing Pseudomonas aeruginosa in Pretoria, South Africa. Int J Antimicrob Agents, 24(1): 35-38. Wilkins BM (2002) Plasmid promiscuity: meeting the challenge of DNA immigration control. Environ Microbiol, 4(9): 495-500. Wilson BA, Salvers AA, Whitt DD, Winkler ME (2010). Bacterial Pathogenesis - A Molecular Approach, Third Edition. ASM Press, American Society for Microbiology. World Economic Forum (2013). Global Risks 2013 - Eighth Edition. http://www.weforum.org/reports/global-risks-2013-eighth-edition. World Economic Forum (2014). Global Risks Report 2014. http://www.weforum.org/reports/global-risks-2014-report.

Bibliography 131
Yamaguchi Y et al. (2005) Probing the role of Asp-120(81) of metallo-β-lactamase (IMP-1) by site-directed mutagenesis, kinetic studies, and X-ray crystallography. J Biol Chem, 280(21): 20824-20832. Yan JJ, Ko WC, Wu JJ (2001) Identification of a plasmid encoding SHV-12, TEM-1, and a variant of IMP-2 metallo-β-lactamase, IMP-8, from a clinical isolate of Klebsiella pneumoniae. Antimicrob Agents Chemother, 45(8): 2368-2371. Yigit H, Queenan AM, Anderson GJ, Domenech-Sanchez A, Biddle JW, Steward CD, Alberti S, Bush K, Tenover FC (2001) Novel carbapenem-hydrolyzing β-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents Chemother, 45(4): 1151-1161. Yong D, Toleman MA, Bell J, Ritchie B, Pratt R, Ryley H, Walsh TR (2012) Genetic and biochemical characterization of an acquired subgroup B3 metallo-β-lactamase gene, blaAIM-1, and its unique genetic context in Pseudomonas aeruginosa from Australia. Antimicrob Agents Chemother, 56(12): 6154-6159. Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, Lee K, Walsh TR (2009) Characterization of a new metallo-β-lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother, 53(12): 5046-5054. Zapun A, Contreras-Martel C, Vernet T (2008) Penicillin-binding proteins and β-lactam resistance. FEMS Microbiol Rev, 32(2): 361-385. Zatyka M, Thomas CM (1998) Control of genes for conjugative transfer of plasmids and other mobile elements. FEMS Microbiol Rev, 21: 291-319. Zhanel GG, Johanson C, Embil JM, Noreddin A, Gin A, Vercaigne L, Hoban DJ (2005) Ertapenem: review of a new carbapenem. Expert Rev Anti Infect Ther, 3(1): 23-39. Zhao WH, Hu ZQ (2011) IMP-type metallo-β-lactamases in Gram-negative bacilli: distribution, phylogeny, and association with integrons. Crit Rev Microbiol, 37(3): 214-226.
Appendix 132
8 Appendix
Appendix 1 Ion exchange (A) and gel filtration (B) chromatograms of the IMP-31 FPLC. Shown are the chromogram curves monitored during the chromatography process. The curves show the amount of detected milli absorbance units (mAU) relative to the volume passed through the columns. Red lines indicate the collected fractions. The ion exchange step for IMP-31 was performed at a pH of 7.5.

Appendix 133
Appendix 2 Ion exchange (A) and gel filtration (B) chromatograms of the IMP-1 FPLC. Shown are the chromogram curves monitored during the chromatography process. The curves show the amount of detected milli absorbance units (mAU) relative to the volume passed through the columns. Red lines indicate the collected fractions. The ion exchange step for IMP-1 was performed at a pH of 7.5.
Appendix 134
Appendix 3 Ion exchange (A) and gel filtration (B) chromatograms of the OXA-233 FPLC. Shown are the chromogram curves monitored during the chromatography process. The curves show the amount of detected milli absorbance units (mAU) relative to the volume passed through the columns. Red lines indicate the collected fractions. The ion exchange step for OXA-233 was performed at a pH of 6.0.

Appendix 135
Appendix 4 Ion exchange (A) and gel filtration (B) chromatograms of the OXA-10 FPLC. Shown are the chromogram curves monitored during the chromatography process. The curves show the amount of detected milli absorbance units (mAU) relative to the volume passed through the columns. Red lines indicate the collected fractions. The ion exchange step for OXA-10 was performed at a pH of 4.9.

Appendix 136
Appendix 5 Ion exchange (A) and gel filtration (B) chromatograms of the KHM-1 FPLC. Shown are the chromogram curves monitored during the chromatography process. The curves show the amount of detected milli absorbance units (mAU) relative to the volume passed through the columns. Red lines indicate the collected fractions. The ion exchange step for KHM-1 was performed at a pH of 7.5.

Publications 137
Publications
Research articles
Pfennigwerth N, Geis G, Gatermann SG, Kaase M (2015) Description of IMP-31, a novel
metallo-β-lactamase found in an ST235 Pseudomonas aeruginosa strain in Western Germany.
J Antimicrob Chemother. [Published online ahead print, pii: dkv079]
Conference posters
Pfennigwerth N, Gatermann SG, Korte M, Neumann S, Marlinghaus L, Kaase M (2011)
Description of a novel IMP carbapenemase, IMP-31, in two Pseudomonas aeruginosa isolates
from Germany. Poster ERP08.
63. Jahrestagung der Deutschen Gesellschaft für Hygiene und Mikrobiologie (DGHM), Essen,
Germany
Pfennigwerth N, Gatermann SG, Kaase M (2012) Description of IMP-31, a novel metallo-β-
lactamase very divergent from other known IMP carbapenemases. Poster P1237.
22nd European Congress for Clinical Microbiology and Infections Diseases (ECCMID), London, UK
Pfennigwerth N, Meining L, Lang R, Gatermann SG, Kaase M (2012) Description of OXA-233,
a novel class D β-lactamase with activity against carbapenems. Poster PRP06.
64. Jahrestagung der Deutschen Gesellschaft für Hygiene und Mikrobiologie (DGHM), Hamburg,
Germany
Pfennigwerth N, Meining L, Lang R, Gatermann SG, Kaase M (2013) Description of OXA-233,
a novel class D carbapenemase inhibited by clavulanic acid. Poster P1278.
23th European Congress for Clinical Microbiology and Infections Diseases (ECCMID), Berlin,
Germany
Pfennigwerth N, Gatermann SG, Kaase M (2013) Phylogenetic analysis of the IMP-type
carbapenemase IMP-31 from Pseudomonas aeruginosa. Poster PRP05.
65. Jahrestagung der Deutschen Gesellschaft für Hygiene und Mikrobiologie (DGHM), Rostock,
Germany

Publications 138
Pfennigwerth N, Hoffmann A, Belmar-Campos C, Gatermann SG, Kaase M (2014)
Description of KHM-2, a novel metallo-β-lactamase found in a clinical Pseudomonas aeruginosa
isolate from Germany. Poster P1120.
24th European Congress for Clinical Microbiology and Infections Diseases (ECCMID), Barcelona,
Spain
Pfennigwerth N, Hoffmann A, Belmar-Campos C, Gatermann SG, Kaase M (2014)
Description of KHM-2, a novel metallo-β-lactamase found in a clinical Pseudomonas aeruginosa
isolate from Germany. Poster PRP42.
66. Jahrestagung der Deutschen Gesellschaft für Hygiene und Mikrobiologie (DGHM), Dresden,
Germany

Curriculum vitae 139
Curriculum vitae
Name Niels Ernst Pfennigwerth
Date of birth 26.05.1985
Place of birth Essen
Nationality German
Family status married
Education
since 04/2011 PhD studies at the Department of Medical Microbiology
Ruhr-University Bochum, Prof. Dr. Sören G. Gatermann
Title: Identification and characterization of novel carbapenemases
10/2008-04/2011 Master studies in Biology
Ruhr-University Bochum
Master thesis Department of Microbial Biology
Ruhr-University Bochum, Prof. Dr. Franz Narberhaus
Title: Analysis of the stability and localisation of the KDO
transferase KdtA from Escherichia coli
10/2005-09/2008 Bachelor studies in Biology
Ruhr-University Bochum
Bachelor thesis Department of General and Molecular Botany
Ruhr-University Bochum, Prof. Dr. Ulrich Kück
Title: Synthesis of Raa4 from Chlamydomonas reinhardtii in E. coli for in vitro
binding studies
08/1991-05/2004 Gymnasium Essen-Überruhr
Graduation: Diploma from German secondary school qualifying for university
admission or matriculation
Other activities
01/2008-02/2008 Internship at the microbiological lab
ZLM GmbH, Essen

Erklärung 140
Erklärung
Hiermit erkläre ich, dass ich die Arbeit selbstständig verfasst und bei keiner anderen Fakultät
eingereicht und dass ich keine anderen als die angegebenen Hilfsmittel verwendet habe. Es
handelt sich bei der heute von mir eingereichten Dissertation um sechs in Wort und Bild völlig
übereinstimmende Exemplare.
Weiterhin erkläre ich, dass digitale Abbildungen nur die originalen Daten enthalten und in
keinem Fall inhaltsverändernde Bildbearbeitung vorgenommen wurde.
Bochum, den
______________________________________
Niels Ernst Pfennigwerth
Related Documents











