ORIGINAL ARTICLE Comparison of Three Hypothermic Target Temperatures for the Treatment of Hypoxic Ischemia: mRNA Level Responses of Eight Genes in the Piglet Brain Linus Olson & Stuart Faulkner & Karin Lundströmer & Aron Kerenyi & Dorka Kelen & M. Chandrasekaran & Ulrika Ådén & Lars Olson & Xavier Golay & Hugo Lagercrantz & Nicola J. Robertson & Dagmar Galter Received: 29 July 2012 / Revised: 6 September 2012 / Accepted: 25 September 2012 # Springer Science+Business Media New York 2012 Abstract Hypothermia can reduce neurodevelopmental disabilities in asphyxiated newborn infants. However, the optimal cooling temperature for neuroprotection is not well defined. We studied the effects of transient piglet brain hypoxic ischemia (HI) on transcriptional activity of eight genes and if mRNA level alterations could be counteracted by whole body cooling to 35, 33.5 or 30 °C. BDNF mRNA was globally upregulated by the insult, and none of the cooling temperatures counteracted this change. In contrast, MANF mRNA was downregulated, and these changes were modestly counteracted in different brain regions by hypothermic treatment at 33.5 °C, while 30 °C aggravated the MANF mRNA loss. MAP2 mRNA was markedly down- regulated in all brain regions except striatum, and cooling to 33.5 °C modestly counteract this downregulation in the cortex cerebri. There was a tendency for GFAP mRNA levels in core, but not mantle regions to be downregulated and for these changes to be modestly counteracted by cool- ing to 33.5 or 35 °C. Cooling to 30 °C caused global GFAP mRNA decrease. HSP70 mRNA tended to become upregu- lated by HI and to be more pronounced in cortex and CA1 of hippocampus during cooling to 33.5 °C. We conclude that HI causes alterations of mRNA levels of many genes in superficial and deep piglet brain areas. Some of these changes may be beneficial, others detrimental, and lowering body temperature partly counteracts some, but not all changes. There may be general differences between core and mantle regions, as well as between the different cooling temperatures for protection. Comparing the three studied temperatures, cooling to 33.5 °C, appears to provide the best cooling temperature compromise. Keywords Neonatal asphyxia . Hypoxic ischemic encephalopathy . Therapeutic hypothermia . Neuronal and glial markers . In situ hybridisation . Piglets Introduction Hypoxic ischemia (HI) during childbirth can lead to perma- nent brain dysfunctional states and constitutes a worldwide health problem, not the least in developing countries. While transient mild hypothermia initiated within the first few L. Olson (*) : U. Ådén : H. Lagercrantz Department of Women’ s and Children’ s Health, Astrid Lindgren Children’ s Hospital, Karolinska Institutet, 17176 Stockholm, Sweden e-mail: [email protected] L. Olson : K. Lundströmer : L. Olson : D. Galter Department of Neuroscience, Karolinska Institutet, Stockholm, Sweden L. Olson Division of Energy Processes, Chemical Engineering and Technology, KTH Royal Institute of Technology, Stockholm, Sweden S. Faulkner : A. Kerenyi : D. Kelen : M. Chandrasekaran : N. J. Robertson Institute for Women’ s Health, University College London, London, UK X. Golay Institute of Neurology, University College London, London, UK Transl. Stroke Res. DOI 10.1007/s12975-012-0215-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Comparison of Three Hypothermic Target Temperaturesfor the Treatment of Hypoxic Ischemia: mRNA LevelResponses of Eight Genes in the Piglet Brain
Linus Olson & Stuart Faulkner & Karin Lundströmer &
Aron Kerenyi & Dorka Kelen & M. Chandrasekaran &
Ulrika Ådén & Lars Olson & Xavier Golay &
Hugo Lagercrantz & Nicola J. Robertson & Dagmar Galter
Received: 29 July 2012 /Revised: 6 September 2012 /Accepted: 25 September 2012# Springer Science+Business Media New York 2012
Abstract Hypothermia can reduce neurodevelopmentaldisabilities in asphyxiated newborn infants. However, theoptimal cooling temperature for neuroprotection is not welldefined. We studied the effects of transient piglet brainhypoxic ischemia (HI) on transcriptional activity of eightgenes and if mRNA level alterations could be counteractedby whole body cooling to 35, 33.5 or 30 °C. BDNF mRNAwas globally upregulated by the insult, and none of thecooling temperatures counteracted this change. In contrast,MANF mRNA was downregulated, and these changeswere modestly counteracted in different brain regions by
hypothermic treatment at 33.5 °C, while 30 °C aggravatedthe MANF mRNA loss. MAP2 mRNAwas markedly down-regulated in all brain regions except striatum, and cooling to33.5 °C modestly counteract this downregulation in thecortex cerebri. There was a tendency for GFAP mRNAlevels in core, but not mantle regions to be downregulatedand for these changes to be modestly counteracted by cool-ing to 33.5 or 35 °C. Cooling to 30 °C caused global GFAPmRNA decrease. HSP70 mRNA tended to become upregu-lated by HI and to be more pronounced in cortex and CA1 ofhippocampus during cooling to 33.5 °C. We conclude thatHI causes alterations of mRNA levels of many genes insuperficial and deep piglet brain areas. Some of thesechanges may be beneficial, others detrimental, and loweringbody temperature partly counteracts some, but not allchanges. There may be general differences between coreand mantle regions, as well as between the different coolingtemperatures for protection. Comparing the three studiedtemperatures, cooling to 33.5 °C, appears to provide thebest cooling temperature compromise.
Keywords Neonatal asphyxia . Hypoxic ischemicencephalopathy . Therapeutic hypothermia . Neuronal andglial markers . In situ hybridisation . Piglets
Introduction
Hypoxic ischemia (HI) during childbirth can lead to perma-nent brain dysfunctional states and constitutes a worldwidehealth problem, not the least in developing countries. Whiletransient mild hypothermia initiated within the first few
L. Olson (*) :U. Ådén :H. LagercrantzDepartment of Women’s and Children’s Health,Astrid Lindgren Children’s Hospital, Karolinska Institutet,17176 Stockholm, Swedene-mail: [email protected]
L. Olson :K. Lundströmer : L. Olson :D. GalterDepartment of Neuroscience, Karolinska Institutet,Stockholm, Sweden
L. OlsonDivision of Energy Processes,Chemical Engineering and Technology,KTH Royal Institute of Technology,Stockholm, Sweden
S. Faulkner :A. Kerenyi :D. Kelen :M. Chandrasekaran :N. J. RobertsonInstitute for Women’s Health, University College London,London, UK
X. GolayInstitute of Neurology, University College London,London, UK
Transl. Stroke Res.DOI 10.1007/s12975-012-0215-4

hours after birth can counteract several aspects of such braindamage in asphyxiated newborn infants [1–11], furtherresearch is needed to understand pathogenic mechanismsand how cooling parameters should be optimized to bettercounteract brain damage. Thus, additional information isneeded about how to best induce hypothermia, when itneeds to be initiated and for how long it should bemaintained. Further research is also needed with respectto optimal temperature for neuroprotection, and how coolingaffects different areas of brains with sizes similar to thatof the newborn infant.
Studies of thermoregulation in newborns of differentspecies suggest that rectal temperature can be used as anindicator of inner brain temperature [12–15]. However, re-cent studies have stressed the fact that temperature variationdue to type of cooling device can influence outcome [5,16–18]. Moreover, active cooling methods may lead todifferent lowering of the temperature in different braincompartments [19–21], and thus optimal cooling parametersmight differ between brain areas [22].
Stress to the neonatal brain, such as caused by HI, can beassumed to be associated with a very large number of geneactivity alterations, which in turn are typically reflected byaltered levels of mRNA. Some of these changes may occuras active, protective or compensatory responses to thehypoxic ischemic state, while other gene activity changes(typically downregulations of transcriptional activity)may be forced by the lack of energy in the system and/or metabolic abnormalities that follow the HI [23]. Insitu hybridization using radioactive oligomer probesallows direct detection and quantification of mRNAspecies in defined brain areas, down to the level ofsingle neurons, in minute tissue samples (a single 14-μmcryostat section), without the need for reverse transcriptaseor PCR steps.
Here, we address one of the key cooling issues, the roleof cooling temperature, in a well-established piglet modelfor neonatal hypoxic ischemia in which cooling to 33.5 °Cas well as the combination of cooling and xenon has beenshown to have beneficial effects ([24]; Olson et al., manu-script under submission). Anesthetized piglets were sub-jected to HI and kept anesthetized for 48 h withoutcooling, or while cooled to a rectal temperature of 35, 33.5or 30 °C. In a previous study (Olson et al., manuscript undersubmission), in which cooling to a rectal temperature of33.5 °C was used as one protective measure, we studiedthe cellular expression of mRNA encoded for by a selectionof genes with key roles in the brain. Messenger RNAspecies encoded for by the same set of genes was thereforechosen in the present study and included two trophic factors,MANF and BDNF, the immediate early gene HSP70, themicrotubule-associated protein MAP2, the astroglial inter-mediate filament protein GFAP, and the Nogo receptor,
NgR1, a key regulator of brain plasticity. Like in our previ-ous study, we also monitored mRNA encoding the twolactate dehydrogenases LDH-A and LDH-B, important forglucose/pyruvate regulation in the brain [25].
Material and Methods
Hypoxic Ischemia Model and Cooling Protocol
All experimentations were under the UK Home OfficeGuidelines (Animals (Scientific Procedures) Act 1986) andSwedish guidelines. Twenty-eight large, white male pigletsaged <24 h were anesthetised and surgically prepared asdescribed previously [24].
Cerebral Hypoxic Ischemia
A magnetic resonance spectroscopy (MRS) surface coil wassecured on the cranium, and the animal was positioned in a9.4 Tesla magnetic resonance spectrometer (Varian, UK).Transient HI was next induced by inflating the vascularoccluders and reducing fractional inspired oxygen (FiO2)to 12 % (v/v). During HI, cerebral energetics were moni-tored every 2 min by 31P-MRS, and β-nucleotide triphos-phate (NTP, mainly ATP) was measured. When β-NTP hadfallen to 40 % of baseline, FiO2 was adjusted in order tostabilise β-NTP at that level for 12.5 min; after which, theoccluders were deflated and FiO2 normalised. 31P spectrawere acquired for a further 1 h to monitor recovery. The timeintegral of the β-NTP peak area decrement relative to theexchangeable phosphate pool during HI and the first 1 h ofresuscitation was quantified, and acute energy depletion wasdetermined [26].
Experimental Groups
Post-resuscitation piglets were randomised into fourgroups: (1) normothermia (rectal temperature (Trec) 38.5 °Cthroughout), or whole-body cooling to (2) Trec 35 °C, (3)Trec 33.5 °C, or (4) Trec 30 °C during 2–26 h post-insult(all groups n07). Normothermic piglets were maintainedat their target Trec using a warmed water mattress aboveand below the animal; hypothermia piglets were cooled(by cooling the water mattress) to their target Trec over90 min starting 2 h after HI. At 26 h after HI, cooledpiglets were re-warmed to normothermia at 0.5 °C/h.Forty-eight hours post-HI, piglets were euthanized withpentobarbital, and the brain was cardiac perfused withcold 4 % paraformaldehyde, removed along with majororgans and processed for histology and immunohisto-chemistry. Immunohistochemical and MRS data for thisseries are presented elsewhere.
Transl. Stroke Res.

In Situ Hybridization and mRNA Quantification
One hemisphere from each piglet was used for in situhybridization studies, while the other half was used forparallel studies using other histopathological techniques (cf[24]; Olson et al., manuscript under submission). Afterrinsing in sucrose and guided by available literature on pigbrain anatomy [27], different brain areas were dissected,frozen and used to obtain serial 14-μm coronal cryostat(Microm, HM500M) sections. These were thawed ontocoated slides (Super frost, Menzel-Gläser, Braunschweig,Germany) and stored at −20 °C until use. Oligonucleotideprobes specific to the mRNA species of interest (Olson etal., manuscript under submission) were labelled with [33P]dATP (Perkin-Elmer, Massachusetts, USA) by terminaldeoxynucleotidyl transferase (Fermenta, Helsingborg,Sweden), purified and used to hybridize sections over nightat 42 °C. The in situ hybridization protocol has been described[28]; see also Olson et al. (manuscript under submission).Slides were exposed to X-ray film (Biomax, Kodak,Rochester, NY, USA) for 2–5 weeks to obtain autoradio-graphic images of probe distribution in the brain sections.The selected probes corresponded to the following sequen-ces: LDHA, NM_001172363.1, nt:1412–1363; LDHB,NM_001113287.1, nt: 353–505; GFAP, GI: 29335682, nt:89–138; HSP70, NM_001123127, nt: 1676–1627; MAP2,XM_001926244, nt: 265–217; MANF, GI: 52351138, nt:223–272; BDNF, NM_214259, nt: 561–512; NgR, NgR,XM_001927797.1, nt:876–827.
For each X-ray film, a 14C step standard (AmershamAutoradiographic Microscale, Amersham, USA) was in-cluded. Digitized images were analysed using appropriatesoftware (ImageJ), and densities were compared to the 14Cstep standards. Statistical analyses were made using appro-priate software (Prism version 5). We carried out the statis-tical analyses of naïve vs. HI using Student’s t test, while HIwas tested by ANOVA versus the different treatments.Significant differences are shown as p<0.5, p<0.01 andp<0.001.
Results
Examination of cellular mRNA expression patterns in aseparate control piglet subjected to 48-h anaesthesia onlysuggests that the mRNA species analysed are present at thesame levels and patterns as in naïve brains (data not shown).As expected, the effects of HI on mRNA levels of theselected set of genes in different brain areas agree largelywith the findings in another set of piglets (Olson et al.,manuscript under submission). Likewise, the effects ofcooling to 33.5 °C are largely similar to the observationsin our other data set, showing that our experimental system
generates reproducible results and setting the stage forcomparing this cooling temperature to milder (35 °C) andmore severe (30 °C) cooling. Examples of mRNA amountsin cortex cerebri of naive piglets, and piglets subjected tohypoxic ischemia without or with 48 h cooling to 33.5 °Care shown in Fig. 1 for Map2, HSP70 and MANF. Figures 2and 3 present the quantification of the mRNA levels for thesix genes.
BDNF mRNA
BDNF mRNA was low in all areas of the naive newbornpiglet brain. Quantitative data on the effects of hypoxicischemia and the three different cooling temperatures areshown in Fig. 2. HI increased BDNF mRNA levels in cortexand in other investigated areas. We did not find any markedeffects of any of the cooling temperatures on these BDNFmRNA increases, although the mean levels were marginallylower when cooling to 33.5 °C compared to 35 or 30 °C.
MANF mRNA
MANF mRNA levels were generally high, and the gene wasexpressed both in neurons and non-neural cells (Fig. 2;signals in white matter). HI decreased MANF mRNA levelsin all examined areas. Cooling to 33.5 °C appeared tomodestly counteract the effects of HI in all areas. Whilelevels were not significantly improved by cooling to 33.5 °Cin any individual area, the consistency of this pattern hasonly a ≈3 % likelihood ((1/2)5) to occur. In cortex, striatumand thalamus, decreasing the piglet rectal temperature to30 °C caused lower levels of MANF mRNA than in any ofthe other four tested conditions (naive, hypoxic ischemia orhypoxic ischemia + cooling to 33.5 or 35 °C). Cooling to35 °C did not alter MANF mRNA levels. It should be notedthat none of the individual effects of cooling reachedsignificance.
HSP70 mRNA
HSP70 mRNA levels were low in the naive piglet brain andincreased in most areas in response to HI (Fig. 2). A ten-dency for cooling to cause a further increase of HSP70mRNA levels was noted in the cortex and hippocampusand somewhat more pronounced when cooling to 33.5 °C,as compared to 35 or 30 °C. However, although all threecooling temperatures lead to higher mean HSP70 mRNAlevels in both cortex and hippocampus compared to normo-temperature (chance of being random observation: (1/2)6≈1,5 %), none of these observations was individually signif-icant. Notably, mean levels of HSP70 mRNA in striatumwere slightly lower in animals cooled to 33.5 °C in both ourprevious and in the present study.
Transl. Stroke Res.
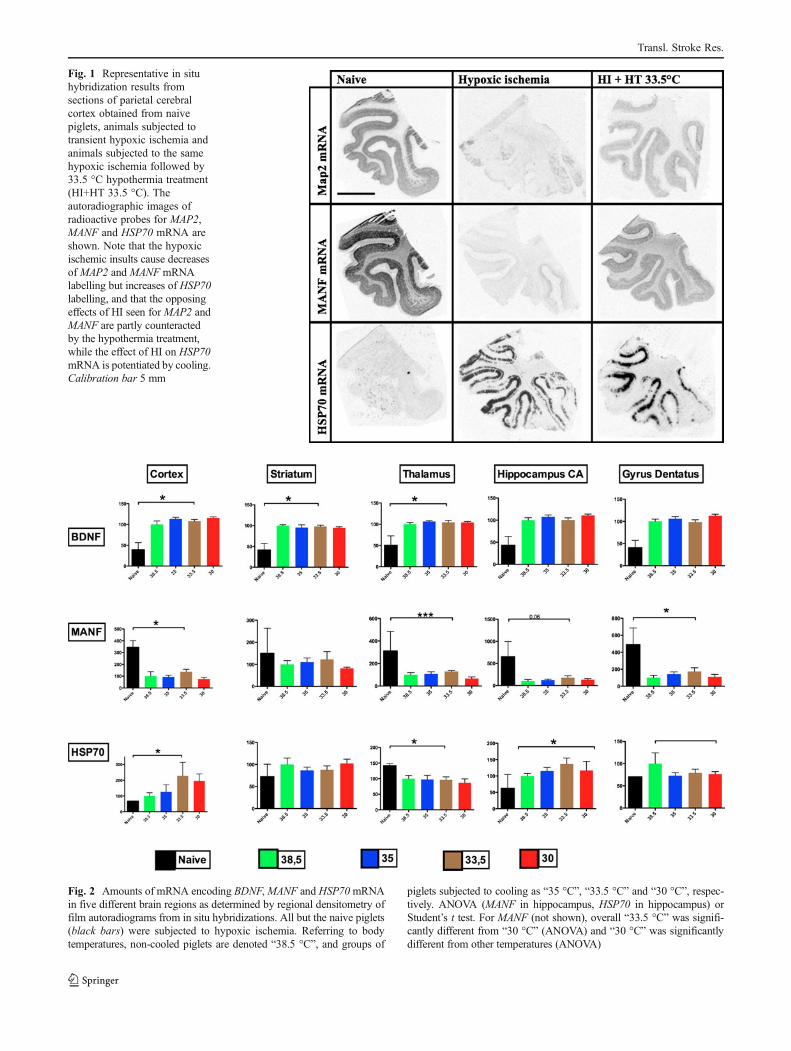
Fig. 1 Representative in situhybridization results fromsections of parietal cerebralcortex obtained from naivepiglets, animals subjected totransient hypoxic ischemia andanimals subjected to the samehypoxic ischemia followed by33.5 °C hypothermia treatment(HI+HT 33.5 °C). Theautoradiographic images ofradioactive probes for MAP2,MANF and HSP70 mRNA areshown. Note that the hypoxicischemic insults cause decreasesof MAP2 and MANF mRNAlabelling but increases of HSP70labelling, and that the opposingeffects of HI seen for MAP2 andMANF are partly counteractedby the hypothermia treatment,while the effect of HI on HSP70mRNA is potentiated by cooling.Calibration bar 5 mm
Fig. 2 Amounts of mRNA encoding BDNF,MANF and HSP70mRNAin five different brain regions as determined by regional densitometry offilm autoradiograms from in situ hybridizations. All but the naive piglets(black bars) were subjected to hypoxic ischemia. Referring to bodytemperatures, non-cooled piglets are denoted “38.5 °C”, and groups of
piglets subjected to cooling as “35 °C”, “33.5 °C” and “30 °C”, respec-tively. ANOVA (MANF in hippocampus, HSP70 in hippocampus) orStudent’s t test. For MANF (not shown), overall “33.5 °C” was signifi-cantly different from “30 °C” (ANOVA) and “30 °C” was significantlydifferent from other temperatures (ANOVA)
Transl. Stroke Res.
GFAP mRNA
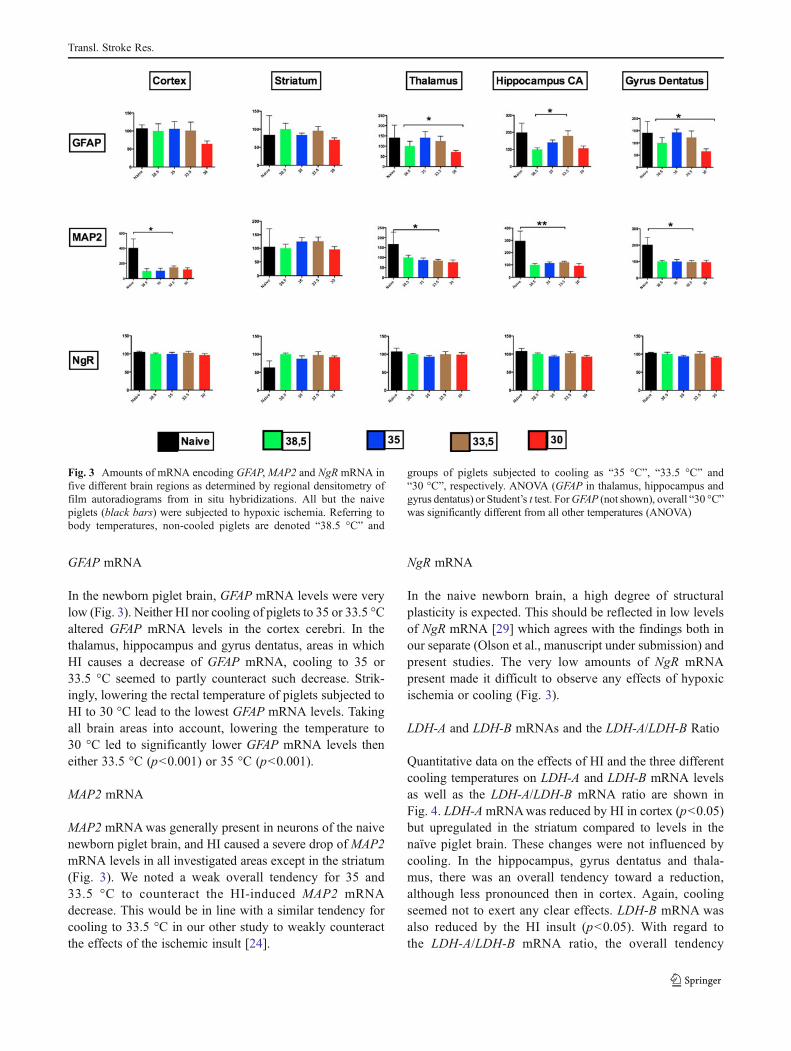
In the newborn piglet brain, GFAP mRNA levels were verylow (Fig. 3). Neither HI nor cooling of piglets to 35 or 33.5 °Caltered GFAP mRNA levels in the cortex cerebri. In thethalamus, hippocampus and gyrus dentatus, areas in whichHI causes a decrease of GFAP mRNA, cooling to 35 or33.5 °C seemed to partly counteract such decrease. Strik-ingly, lowering the rectal temperature of piglets subjected toHI to 30 °C lead to the lowest GFAP mRNA levels. Takingall brain areas into account, lowering the temperature to30 °C led to significantly lower GFAP mRNA levels theneither 33.5 °C (p<0.001) or 35 °C (p<0.001).
MAP2 mRNA
MAP2 mRNAwas generally present in neurons of the naivenewborn piglet brain, and HI caused a severe drop of MAP2mRNA levels in all investigated areas except in the striatum(Fig. 3). We noted a weak overall tendency for 35 and33.5 °C to counteract the HI-induced MAP2 mRNAdecrease. This would be in line with a similar tendency forcooling to 33.5 °C in our other study to weakly counteractthe effects of the ischemic insult [24].
NgR mRNA
In the naive newborn brain, a high degree of structuralplasticity is expected. This should be reflected in low levelsof NgR mRNA [29] which agrees with the findings both inour separate (Olson et al., manuscript under submission) andpresent studies. The very low amounts of NgR mRNApresent made it difficult to observe any effects of hypoxicischemia or cooling (Fig. 3).
LDH-A and LDH-B mRNAs and the LDH-A/LDH-B Ratio
Quantitative data on the effects of HI and the three differentcooling temperatures on LDH-A and LDH-B mRNA levelsas well as the LDH-A/LDH-B mRNA ratio are shown inFig. 4. LDH-A mRNAwas reduced by HI in cortex (p<0.05)but upregulated in the striatum compared to levels in thenaïve piglet brain. These changes were not influenced bycooling. In the hippocampus, gyrus dentatus and thala-mus, there was an overall tendency toward a reduction,although less pronounced then in cortex. Again, coolingseemed not to exert any clear effects. LDH-B mRNA wasalso reduced by the HI insult (p<0.05). With regard tothe LDH-A/LDH-B mRNA ratio, the overall tendency
Fig. 3 Amounts of mRNA encoding GFAP,MAP2 and NgR mRNA infive different brain regions as determined by regional densitometry offilm autoradiograms from in situ hybridizations. All but the naivepiglets (black bars) were subjected to hypoxic ischemia. Referring tobody temperatures, non-cooled piglets are denoted “38.5 °C” and
groups of piglets subjected to cooling as “35 °C”, “33.5 °C” and“30 °C”, respectively. ANOVA (GFAP in thalamus, hippocampus andgyrus dentatus) or Student’s t test. ForGFAP (not shown), overall “30 °C”was significantly different from all other temperatures (ANOVA)
Transl. Stroke Res.
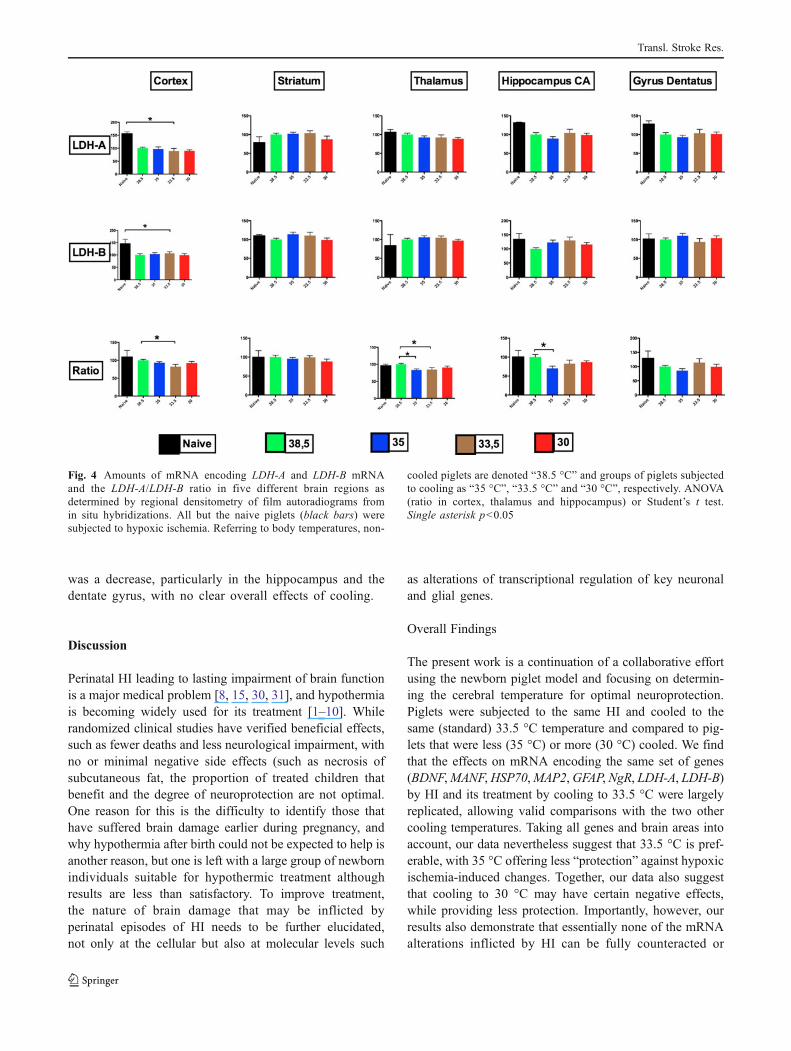
was a decrease, particularly in the hippocampus and thedentate gyrus, with no clear overall effects of cooling.
Discussion
Perinatal HI leading to lasting impairment of brain functionis a major medical problem [8, 15, 30, 31], and hypothermiais becoming widely used for its treatment [1–10]. Whilerandomized clinical studies have verified beneficial effects,such as fewer deaths and less neurological impairment, withno or minimal negative side effects (such as necrosis ofsubcutaneous fat, the proportion of treated children thatbenefit and the degree of neuroprotection are not optimal.One reason for this is the difficulty to identify those thathave suffered brain damage earlier during pregnancy, andwhy hypothermia after birth could not be expected to help isanother reason, but one is left with a large group of newbornindividuals suitable for hypothermic treatment althoughresults are less than satisfactory. To improve treatment,the nature of brain damage that may be inflicted byperinatal episodes of HI needs to be further elucidated,not only at the cellular but also at molecular levels such
as alterations of transcriptional regulation of key neuronaland glial genes.
Overall Findings
The present work is a continuation of a collaborative effortusing the newborn piglet model and focusing on determin-ing the cerebral temperature for optimal neuroprotection.Piglets were subjected to the same HI and cooled to thesame (standard) 33.5 °C temperature and compared to pig-lets that were less (35 °C) or more (30 °C) cooled. We findthat the effects on mRNA encoding the same set of genes(BDNF,MANF,HSP70,MAP2,GFAP, NgR, LDH-A, LDH-B)by HI and its treatment by cooling to 33.5 °C were largelyreplicated, allowing valid comparisons with the two othercooling temperatures. Taking all genes and brain areas intoaccount, our data nevertheless suggest that 33.5 °C is pref-erable, with 35 °C offering less “protection” against hypoxicischemia-induced changes. Together, our data also suggestthat cooling to 30 °C may have certain negative effects,while providing less protection. Importantly, however, ourresults also demonstrate that essentially none of the mRNAalterations inflicted by HI can be fully counteracted or
Fig. 4 Amounts of mRNA encoding LDH-A and LDH-B mRNAand the LDH-A/LDH-B ratio in five different brain regions asdetermined by regional densitometry of film autoradiograms fromin situ hybridizations. All but the naive piglets (black bars) weresubjected to hypoxic ischemia. Referring to body temperatures, non-
cooled piglets are denoted “38.5 °C” and groups of piglets subjectedto cooling as “35 °C”, “33.5 °C” and “30 °C”, respectively. ANOVA(ratio in cortex, thalamus and hippocampus) or Student’s t test.Single asterisk p<0.05
Transl. Stroke Res.

compensated for by cooling at any of the three chosencooling temperatures.
Cooling Protocols
A large part of the standard protocol used today to studyhypothermic therapy in clinical and large animal settings isbased on the work by Gunn and collaborators on sheep [32].The temperature was set to 33–34 °C and the cooling time forhumans to 72 h, based partly on rodent work [33]. Hypother-mia was to be initiated no later than 6 h after birth, as describedby Thoresen et al. [23], to avoid secondary energy failure.
In addition to the cooling temperature, factors influencingthe therapeutic effect of hypothermia include the intervalbetween insult and the induction of hypothermia [34, 35] toobtain maximum benefit [35, 36] and the length of cooling.Short cooling, such as 1 h, has been reported to be ineffec-tive in piglets with regard to both behaviour and brainpathology, even if initiated soon (30 min) after a hypoxicischemic event [37]. The present study was carried out onbrain hemispheres from piglets subjected to HI and random-ized to different hypothermia treatments groups. The selec-tion and treatment before and during the experiment of the48 h of hypothermia follow the protocol documented indetail elsewhere [24] and in which the interval to treatmentwas chosen according to available knowledge. We chose thesame set of eight protein-encoding genes that we have usedin a separate study (Olson et al., manuscript under submis-sion) and that mediate early reactions to neuronal stress(HSP70), neurotrophic proteins (BDNF andMANF), dendriticand somatic cytoskeletal integrity (MAP2), astroglial reactiv-ity (GFAP), regulation of synaptic plasticity (NgR), as well aslactate/pyruvate ratio in the brain (LDH-A and LDH-B).
Lactate
LDH-A and LDH-B generates the M and H subunits form-ing the five different tetrameric LDH isoforms which drivepyruvate–lactate inter-conversions [25], and lactate levelsincrease rapidly in the brain if circulation is compromised.Recently, it was demonstrated that brain lactate levels alsoincrease during aging due to progressive mitochondrial dys-function [25]. Using MRS, brain lactate/Cr and lactate/NAApeak area ratios were monitored [24] in the piglets subjectedto our separate study [38] showing increased levels in thecompromised brain and normalization by cooling and xenontreatment. Similar MRS studies for the present experimentsuggest similar results and indicate that 33.5 °C rectal tem-perature is needed to obtain protection with this measure incore regions of the brain, while cooling to 35 °C is asufficient cooling to protect mantle regions [39]. In line withour previous findings, LDH-A mRNAwas decreased by thehypoxic ischemic event, most marked in the mantle region
(cortex), while we saw no clear influence on this effect bycooling. There was also a modest decrease in the LDH-A/LDH-B mRNA ratio, possibly beneficial in terms of subse-quent formation of H-rich tetrameric enzyme increasing theformation of pyruvate from the excess lactate. Given theknown fast lactate responses, we assume that the transcrip-tional changes also occurred early after the hypoxic ische-mic challenge and that the mRNA alterations measured atsacrifice might be smaller than those measured earlier dur-ing the experiment.
BDNF mRNA
Messenger RNA encoding BDNF [40] is expressed by neu-rons of the adult pig and rodent brain, as shown by in situhybridization [41], and rapidly upregulated by neuronalactivity [29, 42–44]. The global increase of BDNF RNAin response to HI is in agreement with its known rapidresponses to neuronal activation and disturbances. The factthat this is presumably adequate, protective measure was notaffected by any of the cooling temperatures, suggests BDNFupregulation is an important first line response, much like thatof immediate early genes such asHSP70, which is also uphelddespite cooling. A significant dampening effect of cooling to33.5 °C noted in the hippocampus in our previous study(Olson et al., manuscript under submission) was now reducedto a weak tendency, possibly due to somewhat lower sensitiv-ity of the present study or temporal differences in terms oflength of cooling periods and time since HI at sacrifice.Previous work has demonstrated that the delivery of BDNFto the brain of neonatal rats subjected to HI counteractsbehavioural impairments [45, 46], findings that are in linewith the current observations that endogenous BNDF mRNAincrease in response to HI.
MANF mRNA
Lindholm et al. [47] recently characterized MANF as aprotein with neurotrophic effects which is widely expressedin the brain and elsewhere. We found that MANF mRNA isconstitutively present at much higher levels in the newbornpiglet than BDNF mRNA. However, unlike BDNF mRNA,MANF mRNA levels are downregulated uniformly andwithout exception by hypoxic ischemia. The different tem-peratures of hypothermia did not influence this response.The dramatically different responses of the BDNF andMANF genes to a HI challenge demonstrate that alterationsof mRNA levels can occur in both directions. Hence,increases as well as decreases can be active, compensatorychanges as well as forced by the ischemic condition. TheMANF result also exemplifies the fact that not all mRNAinsult-induced alterations, that remain manifest at the timeof sacrifice, can be normalized by cooling.
Transl. Stroke Res.

HSP70 mRNA
HSP70 is upregulated by cellular stress, including ischemia[48], and rescues partially denatured proteins. In agreementwith these findings as well as with our own previous results(Olson et al., manuscript under submission), HSP70 mRNAlevels were clearly increased by HI in the present study.Also, in agreement with the previous results, HSP70 mRNAlevels tended to be further upregulated by cooling to 33.5 °C.While individual observations did not reach significance,the slightly lower levels of HSP70 mRNA in challengedpiglets cooled to 33.5 °C was also in line with the previousfindings (Olson et al., manuscript under submission).Because HSP70 mRNA levels are higher when cooling to30 °C than cooling to 35 or 33.5 °C in the striatum, one mayspeculate that in the core area striatum, a lower rectal tem-perature was needed to see a pattern more similar to that ofthe mantle area of the cortex cerebri. Our piglet data arecompatible with the recent findings with respect to theeffects of different hypothermia temperatures on membersof the HSP70 family in rats [49].
GFAP mRNA
When astrocytes are activated by stress such as brain injury,GFAP levels increase markedly, signifying transformationinto reactive astrocytes. As expected, GFAP mRNA levelswere low in the newborn piglet brain. HI and cooling to 33.5or 35 °C seemed not to cause marked effects. However,cooling to 30 °C was associated with low GFAP mRNAlevels and, thus, seemed to be too low a temperature for thepresent cooling needs.
MAP2 mRNA
Messenger RNA encoded by MAP2, a gene specificallyexpressed by neurons [50], was decreased by HI in all fiveexamined areas and markedly so in parietal cortex, hippo-campal CA1 and the dentate gyrus. This is in agreementwith our findings in another set of piglets (Olson et al.,manuscript under submission). The severe loss of MAP2mRNA, a condition that is present after careful re-warming2–3 days after the insult, suggests this neuropathologicalfeature cannot be effectively counteracted by any of thecooling temperatures.
NgR mRNA
In line with high neuronal plasticity during development,levels of NgR mRNA were generally low. This made smalleffects of HI and treatments difficult to assess. In the adult(as noted in rats and mice), NgR levels are higher in the cortexand hippocampus, and activity-induced downregulation [51],
presumably allowing synaptic reorganisation to carry novellasting memories [29], is therefore easy to observe.
Concluding Remarks
Improvement of current cooling protocols is needed if alarger proportion of individuals is to be rescued from per-manent brain pathologies. In our previous (Olson et al.,manuscript under submission) and the present studies, wesuggest that analysis of transcriptional activity of selectedgenes in defined brain areas may help obtain a more com-plete picture of brain pathology. We present principal differ-ences between several classes of genes, such as increasedversus decreased mRNA levels in response to the insult, andthe fact that the responses to hypothermic treatment arerarely complete normalization and often absent altogether.The overall result from the present study of different coolingtemperatures is that 30 °C is too cold, and that perhaps 35 °Cis too warm. Thus, lowering the body temperature to 33.5 °Cappears somewhat better, although it rarely rectifies HI-induced alterations. A recent study of a number of physio-logical parameters, blood analyses and cardiac monitoring inthe same set of piglets also concludes that cooling to 30 °C ismetabolically detrimental and also associated with episodesof cardiac arrest and loss of piglets [52].
For a selected cooling temperature, further work is neededto monitor the time course of alterations of transcriptionalactivities. This may help in designing the optimal time toinitiate cooling and for how long it should be maintained.The current protocol states that cooling should be initiatedwithin 6 h. Because some of the transcriptional changes can beassumed to be beneficial, particularly the earliest changes ofimmediate early genes, there is reason to further investigate ifsuch changes can resist cooling, not to start cooling too early.
We conclude that studies of the transcriptional activity ofkey brain genes, as revealed by in situ hybridization, offeradditional information about abnormal events in specificregions of the newborn brain subjected to HI. This approachmay help in understanding the many early changes of genefunction that occur and how treatments, such as hypothermia,may counter such gene activity changes.
Acknowledgments We thank Eva Lindqvist and Karin Pernold for theexcellent technical support. This study is supported by the MärthaLundqvist stiftelse (LO), The Swedish Research Council (HL, LO,DG), Stiftelsen Barncentrum (LO), The British MRC (NR), KarolinskaInstitutet, The Karolinska Distinguished Professor Award (LO), andSwedish Brain Power (LO, DG). The preparation of piglets until sacrificeand administration of hypothermia was undertaken at UCLH/UCL, withfunding from the United Kingdom Department of Health’s NIHRBiomedical Research Centres (NR). In situ hybridization for lactatedehydrogenases is supported by the National Institute on Aging, NIH(AG04418), and in situ hybridization for plasticity genes (BDNF andNgR) is supported by the National Institute on Drug Abuse, NIH (LO).
Transl. Stroke Res.

References
1. Azzopardi D, Brocklehurst P, Edwards D, Halliday H, Levene M,Thoresen M, et al. The TOBY Study. Whole body hypothermia forthe treatment of perinatal asphyxial encephalopathy: a randomisedcontrolled trial. BMC Pediatr. 2008;8:17.
2. Azzopardi DV, Strohm B, Edwards AD, Dyet L, Halliday HL,Juszczak E, et al. Moderate hypothermia to treat perinatal asphyxialencephalopathy. N Engl J Med. 2009;361:1349–58.
3. Shankaran S, Laptook AR, Ehrenkranz RA, Tyson JE, McDonaldSA, Donovan EF, et al. Whole-body hypothermia for neonates withhypoxic-ischemic encephalopathy. N Engl J Med. 2005;353:1574–84.
4. Edwards AD, Brocklehurst P, Gunn AJ, Halliday H, Juszczak E,Levene M, et al. Neurological outcomes at 18 months of age aftermoderate hypothermia for perinatal hypoxic ischaemic encepha-lopathy: synthesis and meta-analysis of trial data. BMJ. 2010;340:c363.
5. Jacobs S, Hunt R, Tarnow-Mordi W, Inder T, Davis P. Cooling fornewborns with hypoxic ischaemic encephalopathy. CochraneDatabase Syst Rev. 2007(4):CD003311.
6. Jacobs SE, Morley CJ, Inder TE, Stewart MJ, Smith KR,McNamara PJ, et al. Whole-body hypothermia for term and near-term newborns with hypoxic-ischemic encephalopathy: a random-ized controlled trial. Arch Pediatr Adolesc Med. 2011;165:692–700.
7. Johnston MV, Fatemi A, Wilson MA, Northington F. Treatmentadvances in neonatal neuroprotection and neurointensive care.Lancet Neurol. 2011;10:372–82.
8. Kurinczuk JJ, White-Koning M, Badawi N. Epidemiology ofneonatal encephalopathy and hypoxic-ischaemic encephalopathy.Early Hum Dev. 2010;86:329–38.
9. Perlman M, Shah PS. Hypoxic-ischemic encephalopathy:challenges in outcome and prediction. J Pediatr. 2011;158:e51–4.
10. Roka A, Azzopardi D. Therapeutic hypothermia for neonatal hyp-oxic ischaemic encephalopathy. Early Hum Dev. 2010;86:361–7.
11. Hobbs C, Thoresen M, Tucker A, Aquilina K, Chakkarapani E,Dingley J. Xenon and hypothermia combine additively, offeringlong-term functional and histopathologic neuroprotection afterneonatal hypoxia/ischemia. Stroke. 2008;39:1307–13.
12. Burnard ED, Cross KW. Rectal temperature in the newborn afterbirth asphyxia. Br Med J. 1958;2:1197–9.
13. Clarke L, Heasman L, Firth K, Symonds ME. Influence of route ofdelivery and ambient temperature on thermoregulation in newbornlambs. Am J Physiol. 1997;272:R1931–9.
14. Okken AKJ. Thermoregulation of sick and low birth weightneonates. LŸbeck: Springer; 1995.
15. Pierrat V, Haouari N, Liska A, Thomas D, Subtil D, Truffert P.Prevalence, causes, and outcome at 2 years of age of newbornencephalopathy: population based study. Arch Dis Child. 2005;90:F257–61.
16. Christensson K, Siles C, Cabrera T, Belaustequi A, de la Fuente P,Lagercrantz H, et al. Lower body temperatures in infants deliveredby caesarean section than in vaginally delivered infants. ActaPaediatr. 1993;82:128–31.
17. Iwata S, Iwata O, Olson L, Kapetanakis A, Kato T, Evans S, et al.Therapeutic hypothermia can be induced and maintained usingeither commercial water bottles or a “phase changing material”mattress in a newborn piglet model. Arch Dis Child. 2009;94:387–91.
18. AgnewDM, Koehler RC, Guerguerian AM, Shaffner DH, TraystmanRJ, Martin LJ, et al. Hypothermia for 24 hours after asphyxic cardiacarrest in piglets provides striatal neuroprotection that is sustained10 days after rewarming. Pediatr Res. 2003;54:253–62.
19. Gluckman PD, Wyatt JS, Azzopardi D, Ballard R, Edwards AD,Ferriero DM, et al. Selective head cooling with mild systemic
hypothermia after neonatal encephalopathy: multicentre rando-mised trial. Lancet. 2005;365:663–70.
20. Iwata S, Iwata O, Thornton JS, Shanmugalingam S, Bainbridge A,Peebles D, et al. Superficial brain is cooler in small piglets:neonatal hypothermia implications. Ann Neurol. 2006;60:578–85.
21. Liu X, Chakkarapani E, Hoque N, Thoresen M. Environmentalcooling of the newborn pig brain during whole-body cooling. ActaPaediatr. 2011;100:29–35.
22. Iwata O, Thornton JS, Sellwood MW, Iwata S, Sakata Y, NooneMA, et al. Depth of delayed cooling alters neuroprotection patternafter hypoxia-ischemia. Ann Neurol. 2005;58:75–87.
23. Thoresen M, Penrice J, Lorek A, Cady E, Wylezinska M, KirkbrideV. Mild hypothermia following severe transient hypoxia-ischaemiaameliorates delayed cerebral energy failure in the newborn piglet.Pediatr Res. 1995;37:667–70.
24. Faulkner S, Bainbridge A, Kato T, Chandrasekaran M, KapetanakisAB, Hristova M, et al. Xenon augmented hypothermia reduces earlylactate/N-acetylaspartate and cell death in perinatal asphyxia. AnnNeurol. 2011;70:133–50.
25. Ross JM, Oberg J, Brene S, Coppotelli G, Terzioglu M, Pernold K,et al. High brain lactate is a hallmark of aging and caused by a shiftin the lactate dehydrogenase A/B ratio. Proc Natl Acad Sci USA.2010;107:20087–92.
26. Lorek A, Takei Y, Cady EB, Wyatt JS, Penrice J, Edwards AD, etal. Delayed (“secondary”) cerebral energy failure after acutehypoxia-ischemia in the newborn piglet: continuous 48-hourstudies by phosphorus magnetic resonance spectroscopy. PediatrRes. 1994;36:699–706.
27. Felix B, Leger ME, Albe-Fessard D, Marcilloux JC, Rampin O,Laplace JP. Stereotaxic atlas of the pig brain. Brain Res Bull.1999;49:1–137.
28. Dagerlind A, Friberg K, Bean AJ, Hokfelt T. Sensitive mRNAdetection using unfixed tissue: combined radioactive and non-radioactive in situ hybridization histochemistry. Histochemistry.1992;98:39–49.
29. Karlen A, Karlsson TE, Mattsson A, Lundstromer K, Codeluppi S,Pham TM, et al. Nogo receptor 1 regulates formation of lastingmemories. Proc Natl Acad Sci USA. 2009;106:20476–81.
30. Marlow N, Budge H. Prevalence, causes, and outcome at 2 yearsof age of newborn encephalopathy. Arch Dis Child. 2005;90:F193–4.
31. Schulzke SM, Rao S, Patole SK. A systematic review ofcooling for neuroprotection in neonates with hypoxic ischemicencephalopathy—are we there yet? BMC Pediatr. 2007;7:30.
32. Gunn AJ, Gunn TR, de Haan HH, Williams CE, Gluckman PD.Dramatic neuronal rescue with prolonged selective head coolingafter ischemia in fetal lambs. J Clin Invest. 1997;99:248–56.
33. Sirimanne ES, Blumberg RM, Bossano D, Gunning M, EdwardsAD, Gluckman PD, et al. The effect of prolonged modification ofcerebral temperature on outcome after hypoxic-ischemic braininjury in the infant rat. Pediatr Res. 1996;39:591–7.
34. Taylor DL, Mehmet H, Cady EB, Edwards AD. Improvedneuroprotection with hypothermia delayed by 6 hours followingcerebral hypoxia-ischemia in the 14-day-old rat. Pediatr Res.2002;51:13–9.
35. Colbourne F, Corbett D. Delayed postischemic hypothermia: a sixmonth survival study using behavioral and histological assessmentsof neuroprotection. J Neurosci. 1995;15:7250–60.
36. Wagner BP, Nedelcu J, Martin E. Delayed postischemic hypothermiaimproves long-term behavioral outcome after cerebral hypoxia-ischemia in neonatal rats. Pediatr Res. 2002;51:354–60.
37. Laptook AR, Corbett RJ, Burns DK, Sterett R. A limited intervalof delayed modest hypothermia for ischemic brain resuscitation isnot beneficial in neonatal swine. Pediatr Res. 1999;46:383–9.
38. Olson L. On neonatal asphyxia: clinical and animal studiesincluding development of a simple, safe method for therapeutic
Transl. Stroke Res.

hypothermia with global applicability. Thesis. Stockholm: Dep ofWomen’s and Children’s Health, Karolinska Institutet; 2011.
39. Faulkner S, Bainbridge A, Kelen D, Thayyil S, Chandrasekaran M,Kato T, et al. Whole body cooling to 35, 33 and 30 °C in a pigletmodel of hypoxic ischemic brain injury: which is the optimaltemperature? In: Society for Neuroscience, San Diego, CA, USA,2010. p. 1.
40. Lewin GR, Barde YA. Physiology of the neurotrophins. Annu RevNeurosci. 1996;19:289–317.
41. Wetmore C, Ernfors P, Persson H, Olson L. Localization of brain-derived neurotrophic factor mRNA to neurons in the brain by insitu hybridization. Exp Neurol. 1990;109:141–52.
42. Isackson PJ, Huntsman MM, Murray KD, Gall CM. BDNF mRNAexpression is increased in adult rat forebrain after limbic seizures:temporal patterns of induction distinct from NGF. Neuron.1991;6:937–48.
43. Schmidt-Kastner R, Humpel C, Wetmore C, Olson L. Cellularhybridization for BDNF, trkB, and NGF mRNAs and BDNF-immunoreactivity in rat forebrain after pilocarpine-induced statusepilepticus. Exp Brain Res. 1996;107:331–47.
44. Wetmore C, Olson L, Bean AJ. Regulation of brain-derived neuro-trophic factor (BDNF) expression and release from hippocampalneurons is mediated by non-NMDA type glutamate receptors. JNeurosci. 1994;14:1688–700.
45. Han BH, Holtzman DM. BDNF protects the neonatal brain fromhypoxic-ischemic injury in vivo via the ERK pathway. J Neurosci.2000;20(15):5775–81.
46. Almli CR, Levy TJ, Han BH, Shah AR, Gidday JM, HoltzmanDM, et al. BDNF protects against spatial memory deficitsfollowing neonatal hypoxia-ischemia. Exp Neurol. 2000;166(1):99–114.
47. Lindholm P, Peranen J, Andressoo JO, Kalkkinen N, Kokaia Z,Lindvall O, et al. MANF is widely expressed in mammalian tissuesand differently regulated after ischemic and epileptic insults inrodent brain. Mol Cell Neurosci. 2008;39(3):356–71.
48. Giffard RG, Han RQ, Emery JF, Duan M, Pittet JF. Regulation ofapoptotic and inflammatory cell signaling in cerebral ischemia:the complex roles of heat shock protein 70. Anesthesiology.2008;109:339–48.
49. Xiao-Yan X, Yi-Xin X. Effects of graded hypothermia on hypoxic-ischemic brain damage in the neonatal rat. Chin Med Sci J.2011;26:49–53.
50. Lingwood BE, Healy GN, Sullivan SM, Pow DV, Colditz PB.MAP2 provides reliable early assessment of neural injury in thenewborn piglet model of birth asphyxia. J Neurosci Methods.2008;171:140–6.
51. Josephson A, Trifunovski A, Scheele C, Widenfalk J, WahlestedtC, Brene S, et al. Activity-induced and developmental down-regulation of the Nogo receptor. Cell Tissue Res. 2003;311:333–42.
52. Kerenyi A, Kelen D, Faulkner SD, Bainbridge A, ChandrasekaranM, Cady EB, et al. Systemic effects of whole-body cooling to 35 °C,33.5 °C, and 30 °C in a piglet model of perinatal asphyxia: implica-tions for therapeutic hypothermia. Pediatr Res. 2012;71(5):573–82.
Transl. Stroke Res.
Related Documents











