Combined SP-A/bleomycin effect on cytokines by THP-1 cells; Impact of surfactant lipids on this effect Weixiong Huang 1 , Guirong Wang 1 , David S. Phelps 2 , Hamid Al-Mondhiry 3 , and Joanna Floros 1,2 Departments of 1 Cellular and Molecular Physiology, 2 Pediatrics, and 3 Medicine, Penn State College of Medicine, Hershey, PA 17033, USA Running Title: SP-A/bleomycin and Cytokines Corresponding author Joanna Floros, Ph.D. Departments of Cellular and Molecular Physiology, H166 Penn State College of Medicine 500 University Drive Hershey, PA 17033, USA Telephone: (717) 531-6972 Fax: (717) 531-7667 E-mail: [email protected] Copyright 2002 by the American Physiological Society. AJP-Lung Articles in PresS. Published on February 22, 2002 as DOI 10.1152/ajplung.00434.2001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Combined SP-A/bleomycin effect on cytokines by THP-1
cells; Impact of surfactant lipids on this effect
Weixiong Huang1, Guirong Wang1, David S. Phelps2, Hamid Al-Mondhiry3,
and Joanna Floros 1,2
Departments of 1Cellular and Molecular Physiology, 2Pediatrics, and 3Medicine,
Penn State College of Medicine, Hershey, PA 17033, USA
Running Title: SP-A/bleomycin and Cytokines
Corresponding author
Joanna Floros, Ph.D.
Departments of Cellular and Molecular Physiology, H166
Penn State College of Medicine
500 University Drive
Hershey, PA 17033, USA
Telephone: (717) 531-6972
Fax: (717) 531-7667
E-mail: [email protected]
Copyright 2002 by the American Physiological Society.
AJP-Lung Articles in PresS. Published on February 22, 2002 as DOI 10.1152/ajplung.00434.2001

2
Abstract
Surfactant protein A (SP-A) plays a role in host defense and inflammation in the lung. In the
present study we investigated the hypothesis that SP-A is involved in bleomycin-induced
pulmonary fibrosis. We studied the effects of human SP-A on bleomycin-induced cytokine
production and mRNA expression in THP-1 macrophage-like cells and obtained the following
results. 1) Bleomycin-treated THP-1 cells increased TNF-α, IL-8, and IL-1β production in dose
and time-dependent patterns as we have observed with SP-A. TNF-α levels were unaffected by
treatment with cytosine arabinoside. 2) The combined bleomycin/SP-A effect on cytokine
production is additive by ribonuclease protection assay and synergistic by ELISA. 3) Although
the bleomycin effect on cytokine production was not significantly affected by the presence of
surfactant lipid, the additive and synergistic effect of SP-A/bleomycin on cytokine production
was significantly reduced. We speculate that the elevated cytokine levels resulting from the
bleomycin/SP-A synergism are responsible for bleomycin-induced pulmonary fibrosis and that
surfactant lipids can help ameliorate pulmonary complications observed during bleomycin
chemotherapy.
Keywords: chemotherapeutic agent, ELISA, synergistic effect, Ribonuclease Protection Assay
(RPA)

3
Introduction
Bleomycin is a group of glycopeptides isolated from Streptomyces verticillus. Although it is
an effective antineoplastic agent, bleomycin-induced pulmonary fibrosis sometimes becomes
fatal and limits the usefulness of the drug (34; 39). The histological features of pulmonary
fibrosis in human and animal studies include inflammatory cell recruitment, fibroblast
proliferation and collagen synthesis (5). A number of studies concerning the pathogenesis of
pulmonary fibrosis have focused on the role of inflammatory cells, especially alveolar
macrophage, in the fibrotic process. Bleomycin induces inflammatory cells from human and
animal lung to secrete multifunctional cytokines, such as TNF-α, IL-1β, IL-8, and TGF-β (8; 9;
13; 33; 48). In a clinical study, TNF-α has been shown to be significantly increased after
bleomycin infusion (35).
The mechanism of bleomycin-induced cytokine production is not well understood. The
cytotoxic effect of bleomycin is believed to be related to DNA damage that is characterized by
the appearance of DNA damage-inducible proteins (25) and apoptosis (29). There is also
increased activity of NF-kB, which may result from the increase of reactive oxygen species by
bleomycin (26). NF-kB is a transcriptional factor that regulates the expression of many cytokine
genes (50). Among these, TGF-β is considered to be an important cytokine related to fibroblast
proliferation and collagen synthesis (13; 48) and TNF-α is considered to be a central mediator in
bleomycin-induced pulmonary fibrosis (27; 28; 49). TNF-α receptor knockout mice have been
shown to be protected from lung injury after exposure to bleomycin (27; 28).

4
Pulmonary surfactant is essential for normal lung function. Surfactant protein A (SP-A), in
addition to surfactant-related function (10), plays a role in local host defense and regulation of
inflammatory processes (3; 6). SP-A is a collagenous C-type lectin or collectin (24) and its
carbohydrate recognition domain (CRD) is involved in binding SP-A to pathogens and
promoting phagocytosis of these pathogens by the macrophages (42; 43). In the macrophage-
like THP-1 cell line, human SP-A stimulates production of TNF-α, IL-1β, IL-8, and IL-6 in a
dose- and a time-dependent manner (19; 36; 45). Similar effects are seen in other cells of
monocytic origin from both rats and humans (17; 19). SP-A-enhanced TNF-α production
appears to involve NF-κB activation (14). SP-A also enhances immune cell proliferation (18)
and increases expression of some cell surface proteins (16). In addition, SP-A knockout mice
show an increased susceptibility to infection (21). A recent in vivo study suggests a role for SP-
A in neutrophil recruitment into the lungs of preterm lambs (15). There have also been reports
with other systems in which an anti-inflammatory role has been attributed to SP-A (4).
Surfactant lipids (surfactant TA) can modulate adherence and superoxide production of
neutrophils (37). Surfactant lipids inhibit several SP-A regulated immune cell functions,
including stimulation of macrophages (41). Surfactant lipids and SP-A may be counterregulatory
and changes in the relative amounts of surfactant lipids to SP-A may be important in determining
the immune status of the lung. Although most of SP-A in the normal alveolar space is thought to
be lipid-associated, “lipid-free” SP-A could increase if the balance between SP-A and surfactant
lipid was altered under certain conditions (31). There is evidence that bleomycin-induced lung
injury in animal models is accompanied by qualitative and quantitative changes of surfactant

5
lipids (30; 38). Increased SP-A content in rat has been reported after intratracheal treatment of
bleomycin (32; 44).
We hypothesized that “lipid-free” SP-A, the result of an imbalance of SP-A and surfactant
lipids following bleomycin treatment, enhances the effects of bleomycin on proinflammatory
cytokine production and may be partly responsible for bleomycin-induced pulmonary fibrosis.
In the present study, we examined the effects of SP-A on bleomycin-induced cytokine
production and mRNA expression in THP-1 cells.
Materials and Methods
Cell culture
The THP-1 cell line was obtained from the American Type Culture Collection (Manassas,
VA). Cells were grown in RPMI 1640 medium (Sigma, St. Louis, MO) with 0.05 mM 2-
mercaptoethanol containing 10% fetal calf serum (FCS; Summit Biotechnology, Ft. Collins, CO)
at 37oC in an atmosphere of 5% CO2. The cells were split periodically and used at passages 8-15
in the various experiments. After differentiation with 10-8 M Vitamin D3 for 72h, cells were
pelleted and washed with cold PBS. The cell pellet was then resuspended in complete RPMI
1640 medium with 10% FCS at a density of 2 × 106 cells/ml in 24-well culture plates, and
exposed to bleomycin and SP-A. Cell viability was determined by trypan blue exclusion. Under
the conditions employed in this study, neither bleomycin nor SP-A appeared to have any effect
on the viability of THP-1 cells. Incubations were terminated by pelleting the cells. Supernatants
and /or cell pellet were stored at –80oC until assayed.

6
Bleomycin and native human SP-A
Bleomycin (Blenoxane; Bristol-Myers Squibb Co, Princeton, NJ) solutions were prepared
immediately before use with endotoxin-free saline (American Pharmaceutical Partners, Inc. Los
Angeles, CA). Lipopolysaccharide (LPS) was not detected in the stock solution of bleomycin at
a bleomycin concentration of 5 U/ml (1U=1mg) using the method described below.
SP-A was purified from bronchoalveolar lavage of alveolar proteinosis patients with 1-
butanol extraction (12). After extraction of whole surfactant with butanol, the pellet was
completely dried with a flux of nitrogen gas and then homogenized twice in a freshly prepared
buffer (20 mM n-Octyl β-D-Glucopyranoside, 10mM HEPES, 150 mM NaCl, pH 7.4). After
pelleting, the insoluble protein was dissolved in 5 mM Tris-HCl, pH 7.5 and dialyzed for 48h
against the same buffer. The dialyzed solution was centrifuged (210,000 × g, 4oC, 30 min) and
the supernatant containing SP-A was collected and stored at –80oC. The purified protein was
examined by two-dimensional gel electrophoresis followed by western blotting and silver
staining, and was found to be greater than 98% pure. Protein concentration was determined with
the micro bicinchoninic acid method (Pierce, Rockford, IL) with RNase A as a standard. SP-A
was stored at -80oC. Endotoxin content was determined with the QCL-1000 Limulus amebocyte
lysate assay (Biowhittaker; Walkersville, MD). This test indicated that SP-A used in this study
contained <0.1 pg LPS/10µg SP-A.
Stimulation of THP-1 cells with SP-A and bleomycin
After differentiation with 10-8 M vitamin D3 for 72h, THP-1 cells were pelleted and washed
as described above. Cells at a density of 2 × 106 cells/ml were incubated in 24 well culture

7
plates. For dose-response study, cells were stimulated with bleomycin at concentrations ranging
from 0-100 mU/ml. Time-dependent secretion of cytokines following bleomycin treatment was
studied from 0-24h with 5 and 50 mU/ml of bleomycin. In experiments where the combined
effects of SP-A and bleomycin were examined, SP-A (10 µg/ml) and bleomycin (5 or 50 mU/ml)
were added to cells simultaneously, unless otherwise noted. After treatment, the culture medium
was collected at 4h or 6h for the ELISA assay of cytokine production and cells were harvested at
2h or 4h for cytokine mRNA analysis.
Cytosine arabinoside (Ara-C, Cytosar-U, Pharmacia & Upjohn, Kalamazoo, MI) was
included in some experiments to confirm the specificity of the effect of bleomycin on
proinflammatory cytokine production by THP-1 cells. Ara-C is another antineoplastic agent with
known cytotoxicity (39). Although it has not been associated with pulmonary fibrosis and
cytokine production, it has been shown to cause non-cardiogenic pulmonary edema (11). Ara-
C, at concentrations ranging from 0.1 to 5 mM in culture medium, was used for evaluating the
effect of Ara-C on TNF-α. The conditions employed in the Ara-C experiments were the same as
those in bleomycin studies except that only a 4h time point was done.
Infasurf Inhibition of cytokine production
Infasurf (Forest Pharmaceuticals, St. Louis, MO), an extract of natural surfactant from calf
lung, was used as a source of surfactant lipid. Infasurf was supplied by the manufacturer as a
suspension containing 35mg phospholipids/ml of sterile saline. Infasurf is predominately
phosphatidylcholine and contains ∼ 2% wt/wt protein that includes SP-B and SP-C, but no SP-A.
Infasurf in concentrations ranging from 100 to 800 µg/ml was used in the experiments for ELISA

8
assay of cytokine production, but only a single dose (400 µg/ml) of Infasurf was used in the
experiment for mRNA analysis. Infasurf was preincubated separately with SP-A (10 µg/ml),
bleomycin (5 mU/ml), and SP-A + bleomycin for 15min at 37oC before addition to the THP-1
cells. Cells were incubated for 4h after the treatment. Culture medium and cell pellets were then
collected for ELISA assay and mRNA analysis, respectively.
ELISA assay
The ELISA assays for TNF-α, IL-8, and IL-1β (OptEIA Human ELISA Sets, Pharmingen,
San Diego, CA) were performed according to the instructions recommended by the
manufacturer. The ELISA kits were capable of measuring levels of 7.8-500 pg/ml for TNF-α,
6.2-400 pg/ml for IL-8 and 20-1000 pg/ml for IL-1β. A reference curve for each of these
cytokines was obtained by plotting the concentration of several dilutions of standard protein
versus the corresponding absorbance.
Analysis of cytokine mRNA
Total RNA was isolated from THP-1 cells at 2h or 4h after treatment by using RNeasy Mini
Kits (QIAGEN, Valencia, CA) according to the protocol of the RNeasy Mini Handbook.
Cytokine mRNA quantification was performed by ribonuclease protection assay (RPA).
RiboQuantTM Ribonuclease RPA Starter Package and a Customized Human Template Set
(Pharmingen, San Diego, CA) were used to analyze TNF-α, IL-1β, and IL-8 mRNA in one
assay. The customized template set contains DNA templates that can be used for T7 RNA
polymerase–directed synthesis of α-32P-UTP-labeled, anti-sense RNA probes. These can be
hybridized to TNF-α, IL-1β, and IL-8 mRNA. Templates for the L32 and GAPDH

9
housekeeping genes were also included to allow for normalization of sampling or technical error.
Aliquots of 2µg of total RNA were hybridized with radiolabeled probes at 56oC for 16h. RNase
treatment followed, resulting in degradation of single-stranded RNA and free probes. After
inactivation and precipitation, protected probes were resolved by a 5% polyacrylamide-urea
sequence gel electrophoresis and visualized by autoradiography. Densities of the protected
bands were quantified by soft laser densitometry. The mRNA level is expressed as the ratio of
the densitometric value of each cytokine mRNA to that of the L32 or GAPDH mRNA.
Statistics
Values are presented as means ± SEM. Data were analyzed using SigmaStat statistical
software. For each experiment, statistical treatment included a one way analysis of variance
(ANOVA) followed by a Student-Newman-Keuls test for pairwise comparison and was judged
to be significantly different at p<0.05.
Results
Dose-response and time course studies of bleomycin effects on stimulation of cytokine
production by THP-1 cells.
To study the response of THP-1 cells to bleomycin stimulation, we first performed a dose-
response and a time course of bleomycin effects on TNF-α, IL-8, and IL-1β levels. The
concentrations of bleomycin for the dose-response study ranged from 0-100 mU/ml, which spans
a relevant pharmacological dose (33). As shown in Figure 1, a bleomycin concentration as low
as 0.5 mU/ml increased both TNF-α and IL-8 levels (panels A and B), but a higher concentration
of bleomycin (50 mU/ml) was needed to increase the IL-1β level significantly (panel C).

10
Cytokine production continued to increase as the bleomycin dose was increased to 100 mU/ml.
In contrast, the TNF-α level after Ara-C treatment did not differ from that of control (panel D).
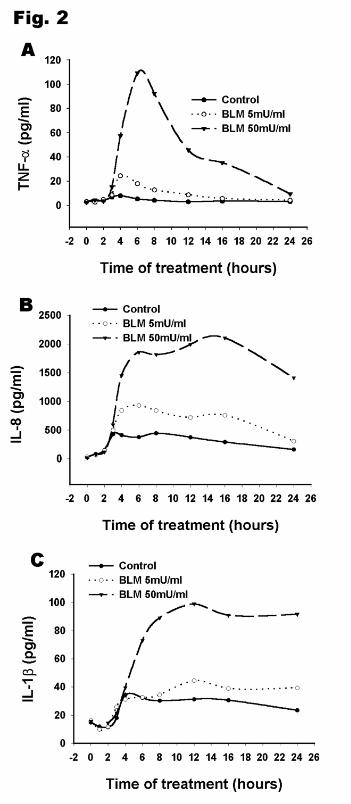
Figure 2 illustrates the time-dependent secretion (0-24h) of TNF-α, IL-8 and IL-1β by THP-
1 cells in the presence or absence of different bleomycin doses. The basal level of TNF-α at 0h
(starting point) was low. The increase of TNF-α levels was usually detected at 3h following
bleomycin treatment and quickly reached a maximum by 4 to 6h depending on the dose selected
(Fig. 2A). The content of TNF-α subsequently decreased and returned to background levels at
24h. Initially, IL-8 had a similar response pattern to TNF-α, with respect to its increase and peak
response time, but after reaching maximal level at around 5h, IL-8 didn’t show a significant
decline until 24h (Fig. 2B). The level of IL-1β increased much later (Fig. 2C) than that of TNF-
α and IL-8, and reached a peak at 10h after bleomycin treatment. Unlike that of TNF-α and IL-
8, the level of IL-1β then did not decline, but remained elevated over the 24h test period.
The combined effect of SP-A and bleomycin on cytokine production and mRNA expression by
THP-1 cells.
After testing the effects of bleomycin treatment on TNF-α, IL-1β and IL-8 production by
THP-1 cells, we examined the combined effects of SP-A and bleomycin on cytokine production.
A dose of 10 µg/ml of SP-A was chosen rather than the dose of 50 µg/ml we have used in
previous experiments (19; 45), as the low dose may better identify synergistic or additive effects
of the two substances. As shown in Figure 3A, TNF-α values induced by SP-A (10 µg/ml) alone
and bleomycin (5 mU/ml) alone were 86.6 ± 11.5 pg/ml and 45.9 ± 10.6 pg./ml, respectively.
But the combined treatment increased the level to 201.7 ± 34.3 pg/ml. High concentration of

11
bleomycin (50 mU/ml) alone induced a TNF-α level of 82.1 ± 17.3 pg/ml, while the value of the
combined effect was 416 ± 61.9 pg/ml. There was a similar response pattern for IL-8 when the
combined effects of SP-A and bleomycin were examined (Fig. 3B). Because IL-1β reached a
maximum value at a later time point than TNF-α and IL-8 did, we measured its level at 6h after
treatment. The means of IL-1β levels (Fig.3C) induced by the combined treatment were greater
than the sum of the separate means by SP-A or bleomycin alone as we saw with TNF-α and IL-
8. SP-A and bleomycin appear to have synergistic effects on TNF-α, IL-1β and IL-8 production
by THP-1 cells.
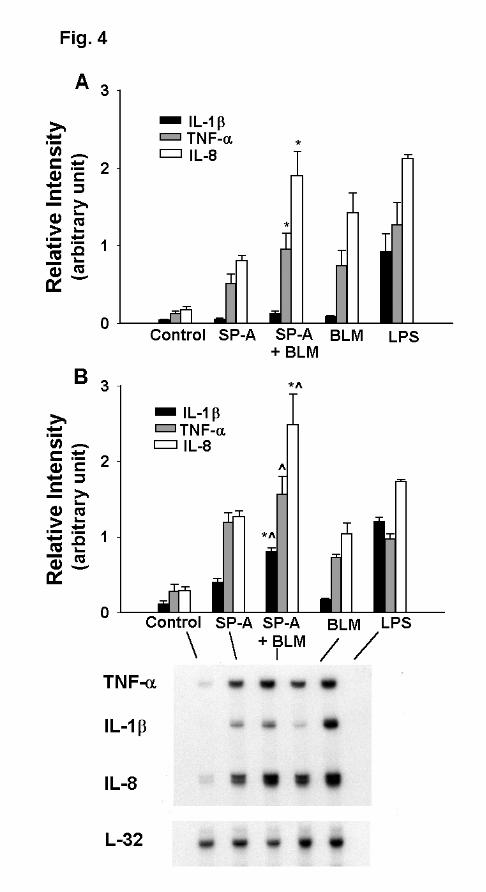
The mRNA levels of TNF-α, IL-1β, and IL-8 were measured by RPA. THP-1 cells were
treated with SP-A (10 µg/ml) and/or bleomycin (5 mU/ml) for 2h and 4h separately. LPS-treated
groups (0.1 ng/ml) were included as a positive control. After 2h of incubation (Fig.4A), TNF-α
and IL-8 mRNA significantly increased as compared to the control (p<0.05). When the cells
were treated with SP-A + bleomycin, the relative intensity of TNF-α mRNA increased from 0.51
± 0.27 (SP-A alone) to 0.95 ± 0.21 (p<0.05), but no significant difference was observed between
SP-A + bleomycin and bleomycin alone (0.95 ± 0.21 vs 0.74 ± 0.20). A similar response pattern
was also seen with IL-8 mRNA. Very low levels of IL-1β mRNA were detected in all SP-A-
and bleomycin-treated cells, but LPS significantly increased the level of IL-1β mRNA at 2h.
The effect of SP-A and bleomycin on IL-1β mRNA expression at 4h (Fig. 4B) were greater than
that at 2h. A significant increase of IL-1β mRNA expression was observed when SP-A
treatment was combined with bleomycin compared to bleomycin alone or SP-A alone. The IL-8
mRNA level (2.47 ± 0.75) following SP-A + bleomycin treatment was significantly higher

12
(p<0.05) than that with SP-A alone (1.26 ± 0.14) or bleomycin alone (1.02 ± 0.16). For TNF-α at
4h, a significant difference was observed between SP-A + bleomycin and bleomycin alone, but
not between the combined treatment and SP-A alone. These results together indicate an additive
effect of SP-A and bleomycin on the TNF-α, IL-1β, and IL-8 mRNA expression.
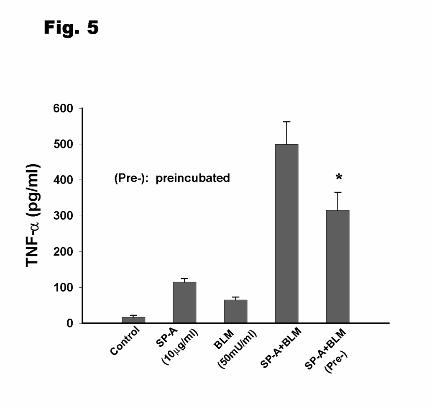
When SP-A (10 µg/ml) was preincubated with bleomycin (50mU/ml) 15min before addition
to the cells, the TNF-α level was significantly lower than that without preincubation (Fig.5).
However, the synergistic effects on TNF-α production remained. There was a similar response
with respect to IL-8 production (Data not shown).
The inhibitory effect of Infasurf on SP-A and bleomycin-induced cytokine production and
mRNA expression.
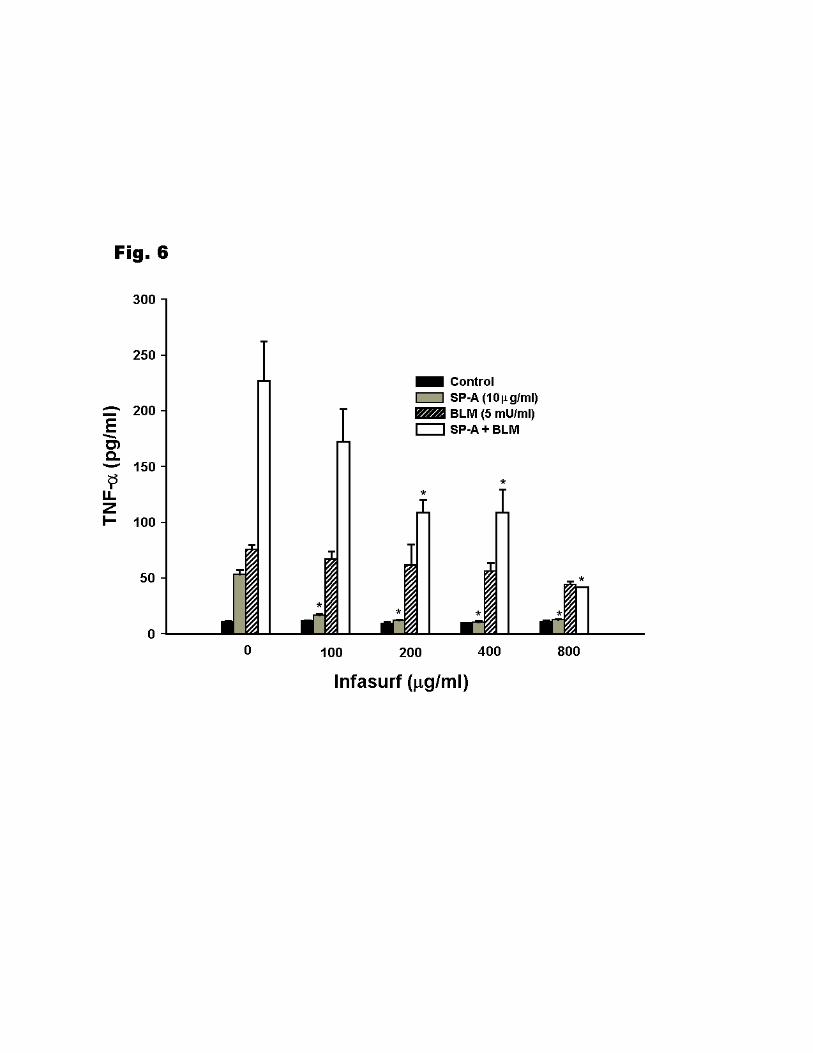
We examined the ability of surfactant lipids to modulate cytokine level. As shown in Fig. 6,
Infasurf had no effect on TNF-α level in the absence of SP-A and bleomycin. SP-A-induced
TNF-α level was significantly reduced by Infasurf at 100 µg/ml and was totally inhibited with a
higher dose of Infasurf. In contrast, the bleomycin effect was not significantly reduced by
Infasurf even at 800µg/ml. Infasurf decreased the TNF-α level induced by SP-A + bleomycin in
a dose-dependent pattern. The TNF-α level was significantly decreased from 226.8 ± 35.7 pg/ml
in the absence of Infasurf to 109 ± 19.3 pg/ml and 41.5 ± 0.7 pg/ml at 200 µg/ml and 800 µg/ml
of Infasurf, respectively.
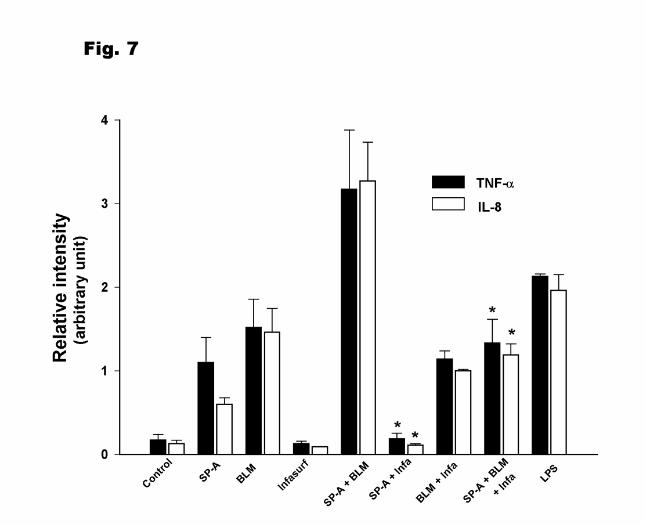
Similar results were obtained when we measured TNF-α and IL-8 mRNA expression by
RPA (Fig.7). TNF-α mRNA expression induced by SP-A was totally inhibited by Infasurf (400

13
µg/ml), but at the same time bleomycin-induced TNF-α mRNA level was not significantly
changed. In SP-A + bleomycin treatment, the relative intensity of TNF-α mRNA was decreased
from 3.17 ± 0.71 in the absence of Infasurf to 1.14 ± 0.10 in the presence of Infasurf. A similar
response pattern was also seen for IL-8 mRNA after Infasurf treatment.
Discussion
In the present study, we investigated whether SP-A plays a role in bleomycin-induced
inflammation and whether surfactant lipids modulate this process. With the macrophage-like
THP-1 cell line we used ribonuclease protection assay and ELISA and observed the following:
Bleomycin (as has been shown for SP-A) enhances proinflammatory cytokine production by
THP-1 cells. The combined bleomycin/SP-A effect on cytokine production is additive by
ribonuclease protection assay and synergistic by ELISA. No effect on cytokine production is
observed by Ara-C, a chemotherapeutic agent that has not been associated with lung
inflammation and fibrosis, suggesting that the effect is specific to bleomycin and/or to agents
associated with lung inflammation and fibrosis. The surfactant lipids significantly suppress the
additive or synergistic effect on cytokine production observed in the presence of both SP-A and
bleomycin. These data indicate that surfactant lipids may be useful in the suppression of
inflammatory processes induced by SP-A and bleomycin in the lung. This in turn could prevent
lung fibrosis, a serious complication of chemotherapeutic agents such as bleomycin.
Bleomycin stimulates THP-1 cells, to secrete cytokines in dose- and time-dependent patterns
in the present study, as we have observed previously in these cells after treatment with SP-A (19;
45). THP-1 cells are of a monocytic origin that upon vitamin D3 differentiation (as described in
methods) acquire macrophage-like phenotype. Undifferentiated THP-1 cells, on the other hand,

14
respond minimally to SP-A (19) and to bleomycin (data not shown). The response pattern of
TNF-α time course observed in this study is similar but not identical to that of clinical
observations about circulating TNF-α level after bleomycin treatment (35). In THP-1 cells the
TNF-α response is transient, whereas in vivo (35) although it decreases with time it does not
return to basal levels. This may reflect the simple nature of the THP-1 system compared to that
in the intact organism. The transient (4-8 h) response of TNF-α is also seen with SP-A and LPS,
although other proinflammatory cytokines (IL-1β, IL-8) show a sustained increase (19). Because
some differences between the THP-1 cell line and alveolar macrophage were apparent in the
kinetics and level of TNF-α and IL-1β induced by bleomycin treatment (33), it is possible that
the THP-1 cells response may not entirely reflect that of alveolar macrophages. However, the
data presented in this report demonstrate the usefulness of the THP-1 cell line as a model system
for the study of bleomycin-induced cytokine production. Moreover, the THP-1 cells have the
advantage of providing a homogeneous cell population for study, whereas primary alveolar
macrophages, even from a single subject, vary significantly from one another depending on what
they have been exposed to in vivo and the length of time they have been in the alveolus. Ara-C,
another chemotherapeutic agent, can also cause pulmonary complications, but this is typically
non-cardiogenic pulmonary edema (11) rather than inflammation and fibrosis. No effect of Ara-
C on cytokine production was observed in THP-1 cells, confirming the specificity of the effect of
bleomycin on cytokine production by THP-1 cells and pulmonary toxicity.
It has been previously reported that native human SP-A can stimulate cytokine production in
macrophage-like THP-1 cells and that this effect of SP-A can be inhibited by surfactant lipids
(16; 36). There have also been reports that SP-A can inhibit LPS-induced cytokine production
(4). These disparate results have led to a controversy as to whether SP-A is proinflammatory or

15
anti-inflammatory. In several animal models there are increases in SP-A early in inflammatory
processes. These models include neonatal hyperoxia (7), sepsis (23), and preterm ventilated
sheep (15). On the other hand, in the SP-A knockout mouse (21) and in a very premature baboon
(2) the lack of SP-A appears to cause increased inflammation. In both of these models there is a
significant delay in pathogen clearance, which is likely to prolong the proinflammatory stimulus
provided by the pathogen. However, in the presence of SP-A, pathogen clearance is enhanced
and this may result in reduced inflammation due to removal of the proinflammatory stimulus
provided by the pathogen.
While SP-A clearly enhances many aspects of host-defense function, these different lines
of evidence prevent it from being easily classified as pro-inflammatory or anti-inflammatory. It
is possible that its role changes at different stages of the inflammatory response, as has been
proposed recently for NF-kB (20).
In the present study, we showed that SP-A in low dose (10 µg/ml) significantly increased
cytokines at both the protein and mRNA levels. LPS (0.1 ng/ml) was used as a positive control
in the present study. A striking difference in IL-1β mRNA expression between LPS and SP-A
treated cells was observed, especially at 2h after treatment (Fig. 4A). The relative intensity of
IL-1β mRNA induced by LPS was 18-fold greater than that induced by SP-A (0.92 ± 0.23 vs
0.05 ± 0.02), while the differences between LPS and SP-A in TNF-α or IL-8 mRNA are only
around 2-fold. These data may provide additional evidence that the regulation of
proinflammatory cytokine production by SP-A in THP-1 cells occurs by a different pathway than
that utilized by LPS (36).

16
Under normal physiological conditions, most of the SP-A in the alveoli is combined with
surfactant lipids in the form of a surfactant lipoprotein complex. Our data suggest that these SP-
A/lipid complexes do not affect cytokine production, perhaps because the complexed SP-A is
unable to interact directly with immune cells (36). Therefore, it is possible that if the lipids are
reduced in quantity or quality, the stimulatory influence of SP-A could be enhanced. In fact, in
bleomycin-induced pulmonary fibrosis, changes in surfactant composition and function have
been revealed in animal models (30; 38). Studies in rat indicate that there is significant increase
of SP-A but not of surfactant phospholipids in response to bleomycin treatment (32; 44). We
showed that although surfactant lipids by themselves had no effect on cytokine production,
Infasurf completely inhibited SP-A proinflammatory function observed in both, ELISA and RPA
assays of cytokine protein and mRNA, respectively. This result was comparable to that reported
previously with Survanta (19). Although both Infasurf and Survanta contain the hydrophobic
surfactant proteins, SP-B and SP-C, several studies comparing these preparations to either
protein-free synthetic surfactant or to pure lipids suggest that SP-B and SP-C do not affect
cytokine expression (1; 40; 41; 46; 47). Infasurf could significantly inhibit the combined effects
of SP-A and bleomycin on cytokines, suggesting involvement of complex mechanisms. This
result may be due to the association of “lipid-free” SP-A with surfactant lipids since the
bleomycin effect on cytokine was not significantly changed by Infasurf even at the highest dose
of 800 µg/ml, further suggesting that these two agents (SP-A and bleomycin) operate through
different mechanisms. Infasurf appears to significantly inhibit the LPS and the LPS + bleomycin
effect on TNF-α production (not shown).

17
The mechanism of bleomycin-induced cytokine production has not been fully elucidated. It
is generally believed to be related to DNA damage (25), apoptosis (29) and activation of NF-kB
(50). It has also been demonstrated that bleomycin-induced injury is associated with the
generation of reactive oxygen species, particularly superoxide anion (26; 37). We speculate that
this mechanism involves the production of reactive oxidants by the bleomycin-treated cells,
which in turn activate NF-kB and increase transcription of cytokine genes. Although the
mechanism of action of SP-A is not known either, it is likely that it involves interaction with a
cell membrane molecule, possibly the C1q receptor (22), activating intracellular events including
the eventual activation of NF-kB (14). When SP-A and bleomycin were added to the cell at the
same time, the levels of both TNF-α and IL-8 were higher than the sum of each cytokine induced
by SP-A or bleomycin alone. Analysis of mRNA showed that the combined treatment of SP-A
and bleomycin exhibited an additive effect on the expression of TNF-α, IL-1β, and IL-8 mRNA.
Although bleomycin itself did not induce a significant increase of IL-1β mRNA even at 4h after
treatment, it greatly enhanced the level of IL-1β mRNA after being combined with SP-A. The
fact that the combined effect of SP-A + bleomycin shown in cytokine protein production was
much greater than that observed in mRNA level indicates that various post-transcriptional and
post-translational mechanisms may be involved in bleomycin-induced proinflammatory cytokine
production by THP-1 cells in response to SP-A. The details of these mechanisms remain to be
determined. The synergistic effect of SP-A and bleomycin on cytokine production in THP-1
cells raises the possibility that SP-A plays a role in bleomycin-induced pulmonary fibrosis.
The underlying mechanism of the combined SP-A and bleomycin effect on cytokine
production by THP-cells is unclear. The carbohydrate recognition domain (CRD) of SP-A may

18
interact with bleomycin which is a group of glycopeptides. We speculate that in experiments
where SP-A and bleomycin are added to the cells simultaneously, these agents exert their effect
through different moieties and that the different pathways converge and cause increased cytokine
gene expression. To distinguish whether the synergistic effect is due to the binding of SP-A to
bleomycin or to independent action of SP-A and bleomycin, we performed preincubation
experiments. We observed that the TNF-α level was decreased following 15 min preincubation
of SP-A with bleomycin before addition to cells. The reduced effect seen when SP-A and
bleomycin are pre-incubated may be the result of bleomycin binding to the CRD of SP-A. This
may in turn compromise the binding of one or the other of these agents to THP-1 cells and thus
interfere with the stimulatory effects.
In summary, we have demonstrated that both SP-A and bleomycin can stimulate production
of inflammatory cytokines by THP-1 cells and that there is a synergistic effect when both agents
are used. Surfactant lipids significantly suppress the synergistic SP-A/bleomycin effect on
cytokine production. We speculate that the significantly elevated cytokine levels resulting from
this synergism are responsible for bleomycin-induced pulmonary fibrosis and that surfactant
lipids can help ameliorate pulmonary complication observed during chemotherapy.

19
Acknowledgments
The authors thank Susan DiAngelo and Todd M. Umstead for their expert technical
assistance. This work was supported by NIH 1R01 ES09882-01, R37 HL34788 and the Julia
Cotler Hematology Research Fund

20
References
1. Antal JM, Divis LT, Erzurum SC, Wiedemann HP and Thomassen MJ. Surfactant suppresses
NF-kappa B activation in human monocytic cells. Am J Respir Cell Mol Biol 14: 374-379, 1996.
2. Awasthi S, Coalson JJ, Yoder BA, Crouch E and King RJ. Deficiencies in lung surfactant
proteins A and D are associated with lung infection in very premature neonatal baboons. Am J
Respir Crit Care Med 163: 389-397, 2001.
3. Batenburg JJ and Haagsman HP. The lipids of pulmonary surfactant: dynamics and
interactions with proteins. Prog Lipid Res 37: 235-276, 1998.
4. Borron P, McIntosh JC, Korfhagen TR, Whitsett JA, Taylor J and Wright JR. Surfactant-
associated protein A inhibits LPS-induced cytokine and nitric oxide production in vivo. Am J
Physiol Lung Cell Mol Physiol 278: L840-847, 2000.
5. Chandler DB. Possible mechanisms of bleomycin-induced fibrosis. Clin Chest Med 11: 21-30,
1990.
6. Crouch EC. Collectins and pulmonary host defense. Am J Respir Cell Mol Biol 19: 177-201,
1998.
7. D'Angio CT, Finkelstein JN, Lomonaco MB, Paxhia A, Wright SA, Baggs RB, Notter RH and
Ryan RM. Changes in surfactant protein gene expression in a neonatal rabbit model of
hyperoxia-induced fibrosis. Am J Physiol 272: L720-730, 1997.
8. Denholm EM and Phan SH. The effects of bleomycin on alveolar macrophage growth factor
secretion. Am J Pathol 134: 355-363, 1989.
9. Denholm EM and Rollins SM. Expression and secretion of transforming growth factor-beta by
bleomycin-stimulated rat alveolar macrophages. Am J Physiol 264: L36-42., 1993.

21
10. Floros J and Phelps DS. Pulmonary surfactant. In: Anesthesia: Biologic Foundations, edited
by Biebuck J, Lynch III C, Maze M, Saidman LJ, Yaksh TL and Zapol WM: Lippincott-Raven,
1997, p. 1259-1279.
11. Haupt HM, Hutchins GM and Moore GW. Ara-C lung: noncardiogenic pulmonary edema
complicating cytosine arabinoside therapy of leukemia. Am J Med 70: 256-261, 1981.
12. Hawgood S, Benson BJ and Hamilton RL, Jr. Effects of a surfactant-associated protein and
calcium ions on the structure and surface activity of lung surfactant lipids. Biochemistry 24: 184-
190, 1985.
13. Khalil N, Whitman C, Zuo L, Danielpour D and Greenberg A. Regulation of alveolar
macrophage transforming growth factor-beta secretion by corticosteroids in bleomycin-induced
pulmonary inflammation in the rat. J Clin Invest 92: 1812-1818, 1993.
14. Koptides M, Umstead TM, Floros J and Phelps DS. Surfactant protein A activates NF-kappa
B in the THP-1 monocytic cell line. Am J Physiol 273: L382-388, 1997.
15. Kramer BW, Jobe AH, Bachurski CJ and Ikegami M. Surfactant protein A recruits
neutrophils into the lungs of ventilated preterm lambs. Am J Respir Crit Care Med 163: 158-165,
2001.
16. Kremlev SG and Phelps DS. Effect of SP-A and surfactant lipids on expression of cell
surface markers in the THP-1 monocytic cell line. Am J Physiol 272: L1070-1077, 1997.
17. Kremlev SG and Phelps DS. Surfactant protein A stimulation of inflammatory cytokine and
immunoglobulin production. Am J Physiol 267: L712-719., 1994.
18. Kremlev SG, Umstead TM and Phelps DS. Effects of surfactant protein A and surfactant
lipids on lymphocyte proliferation in vitro. Am J Physiol 267: L357-364, 1994.

22
19. Kremlev SG, Umstead TM and Phelps DS. Surfactant protein A regulates cytokine
production in the monocytic cell line THP-1. Am J Physiol 272: L996-1004, 1997.
20. Lawrence T, Gilroy DW, Colville-Nash PR and Willoughby DA. Possible new role for NF-
kappaB in the resolution of inflammation. Nat Med 7: 1291-1297, 2001.
21. LeVine AM, Bruno MD, Huelsman KM, Ross GF, Whitsett JA and Korfhagen TR.
Surfactant protein A-deficient mice are susceptible to group B streptococcal infection. J Immunol
158: 4336-4340, 1997.
22. Malhotra R, Haurum J, Thiel S and Sim RB. Interaction of C1q receptor with lung surfactant
protein A. Eur J Immunol 22: 1437-1445, 1992.
23. Malloy J, McCaig L, Veldhuizen R, Yao LJ, Joseph M, Whitsett J and Lewis J. Alterations of
the endogenous surfactant system in septic adult rats. Am J Respir Crit Care Med 156: 617-623,
1997.
24. Mason RJ, Greene K and Voelker DR. Surfactant protein A and surfactant protein D in
health and disease. Am J Physiol 275: L1-13, 1998.
25. Mishra A, Doyle NA and Martin WJc. Bleomycin-mediated pulmonary toxicity: evidence for
a p53-mediated response. Am J Respir Cell Mol Biol 22: 543-549, 2000.
26. Moseley PL, Shasby DM, Brady M and Hunninghake GW. Lung parenchymal injury
induced by bleomycin. Am Rev Respir Dis 130: 1082-1086, 1984.
27. Ortiz LA, Lasky J, Hamilton RF, Jr., Holian A, Hoyle GW, Banks W, Peschon JJ, Brody AR,
Lungarella G and Friedman M. Expression of TNF and the necessity of TNF receptors in
bleomycin- induced lung injury in mice. Exp Lung Res 24: 721-743, 1998.
28. Ortiz LA, Lasky J, Lungarella G, Cavarra E, Martorana P, Banks WA, Peschon JJ, Schmidts
HL, Brody AR and Friedman M. Upregulation of the p75 but not the p55 TNF-alpha receptor

23
mRNA after silica and bleomycin exposure and protection from lung injury in double receptor
knockout mice. Am J Respir Cell Mol Biol 20: 825-833, 1999.
29. Ortiz LA, Moroz K, Liu JY, Hoyle GW, Hammond T, Hamilton RF, Holian A, Banks W,
Brody AR and Friedman M. Alveolar macrophage apoptosis and TNF-alpha, but not p53,
expression correlate with murine response to bleomycin. Am J Physiol 275: L1208-1218, 1998.
30. Osanai K, Takahashi K, Sato S, Iwabuchi K, Ohtake K, Sata M and Yasui S. Changes of lung
surfactant and pressure-volume curve in bleomycin- induced pulmonary fibrosis. J Appl Physiol
70: 1300-1308, 1991.
31. Phelps DS. Surfactant regulation of host defense function in the lung: a question of balance.
Pediatr Pathol Mol Med 20: 269-292, 2001.
32. Savani RC, Godinez RI, Godinez MH, Wentz E, Zaman A, Cui Z, Pooler PM, Guttentag SH,
Beers MF, Gonzales LW and Ballard PL. Respiratory distress after intratracheal bleomycin:
selective deficiency of surfactant proteins B and C. Am J Physiol Lung Cell Mol Physiol 281:
L685-696, 2001.
33. Scheule RK, Perkins RC, Hamilton R and Holian A. Bleomycin stimulation of cytokine
secretion by the human alveolar macrophage. Am J Physiol 262: L386-391, 1992.
34. Sleijfer S. Bleomycin-induced pneumonitis. Chest 120: 617-624, 2001.
35. Sleijfer S, Vujaskovic Z, Limburg PC, Schraffordt Koops H and Mulder NH. Induction of
tumor necrosis factor-alpha as a cause of bleomycin- related toxicity. Cancer 82: 970-974, 1998.
36. Song M and Phelps DS. Comparison of SP-A and LPS effects on the THP-1 monocytic cell
line. Am J Physiol Lung Cell Mol Physiol 279: L110-117, 2000.

24
37. Suwabe A, Otake K, Yakuwa N, Suzuki H, Ito M, Tomoike H, Saito Y and Takahashi K.
Artificial surfactant (Surfactant TA) modulates adherence and superoxide production of
neutrophils. Am J Respir Crit Care Med 158: 1890-1899, 1998.
38. Swendsen CL, Skita V and Thrall RS. Alterations in surfactant neutral lipid composition
during the development of bleomycin-induced pulmonary fibrosis. Biochim Biophys Acta 1301:
90-96, 1996.
39. Tanoue LT. Pulmonary toxicity associated with chemotherapeutic agents. In: Fishman's
Pulmonary Diseases and Disorders, edited by Fishman AP, Elias JA, Fishman JA, Grippi MA,
Kaiser LR and Senior RM. New York: McGraw-Hill, 1998, p. 1003-1016.
40. Thomassen MJ, Antal JM, Connors MJ, Meeker DP and Wiedemann HP. Characterization of
exosurf (surfactant)-mediated suppression of stimulated human alveolar macrophage cytokine
responses. Am J Respir Cell Mol Biol 10: 399-404, 1994.
41. Thomassen MJ, Meeker DP, Antal JM, Connors MJ and Wiedemann HP. Synthetic
surfactant (Exosurf) inhibits endotoxin-stimulated cytokine secretion by human alveolar
macrophages. Am J Respir Cell Mol Biol 7: 257-260, 1992.
42. Tino MJ and Wright JR. Surfactant protein A stimulates phagocytosis of specific pulmonary
pathogens by alveolar macrophages. Am J Physiol 270: L677-688, 1996.
43. van Iwaarden F, Welmers B, Verhoef J, Haagsman HP and van Golde LM. Pulmonary
surfactant protein A enhances the host-defense mechanism of rat alveolar macrophages. Am J
Respir Cell Mol Biol 2: 91-98, 1990.
44. Viviano CJ, Bakewell WE, Dixon D, Dethloff LA and Hook GE. Altered regulation of
surfactant phospholipid and protein A during acute pulmonary inflammation. Biochim Biophys
Acta 1259: 235-244, 1995.

25
45. Wang G, Phelps DS, Umstead TM and Floros J. Human SP-A protein variants derived from
one or both genes stimulate TNF-alpha production in the THP-1 cell line. Am J Physiol Lung
Cell Mol Physiol 278: L946-954, 2000.
46. Wilsher ML, Hughes DA and Haslam PL. Immunoregulatory properties of pulmonary
surfactant: influence of variations in the phospholipid profile. Clin Exp Immunol 73: 117-122,
1988.
47. Woerndle S and Bartmann P. The effect of three surfactant preparations on in vitro
lymphocyte functions. J Perinat Med 22: 119-128, 1994.
48. Yamamoto T, Katayama I and Nishioka K. Fibroblast proliferation by bleomycin stimulated
peripheral blood mononuclear cell factors. J Rheumatol 26: 609-615, 1999.
49. Zhang K, Gharaee-Kermani M, McGarry B, Remick D and Phan SH. TNF-alpha-mediated
lung cytokine networking and eosinophil recruitment in pulmonary fibrosis. J Immunol 158: 954-
959, 1997.
50. Zhang XY, Shimura S, Masuda T, Saitoh H and Shirato K. Antisense oligonucleotides to NF-
kappaB improve survival in bleomycin- induced pneumopathy of the mouse. Am J Respir Crit
Care Med 162: 1561-1568, 2000.

26
Legends:
Figure 1. Dose-response cytokine production after bleomycin treatment by THP-1 cells.
Differentiated THP-1 cells were stimulated with the indicated concentration of bleomycin for 4h
or 6h. TNF-α (Panel A) and IL-8 (Panel B) levels in culture medium at 4h incubation and IL-1β
(Panel C) at 6h were quantified by ELISA. The effect of Ara-C on TNF-α level was examined at
4h (panel D). Data are derived from 5 separate experiments except the Ara-C experiments which
are from two experiments in triplicate. Results are given as means ± SEM. The indicated values
(*) are significantly different (p<0.05) from points obtained in the absence of bleomycin.
Figure 2. Time course of cytokine production after bleomycin treatment. Differentiated THP-1
cells were incubated in the presence of 0, 5, 50 mU/ml bleomycin for the indicated time. TNF-α
(Panel A), IL-8 (Panel B) and IL-1β (Panel C) levels in culture medium were quantified by
ELISA.
Figure 3. The combined effect of SP-A and bleomycin on cytokine production by THP-1 cells.
Differentiated THP-1 cells were stimulated with SP-A (10µg/ml) and bleomycin (BLM 5, 50
mU/ml) simultaneously for 4h or 6h. TNF-α (Panel A) and IL-8 (Panel B) in culture medium at
4h incubation and IL-1β (Panel C) at 6h were quantified. Data are derived from 5 separate
experiments, and the results are given as means ± SEM. The indicated values (*) are
significantly different (p<0.05) from points with SP-A alone and points with the same dose of
bleomycin but without SP-A.

27
Figure 4. The combined effect of bleomycin and SP-A on cytokine mRNA levels.
Differentiated THP-1 cells were stimulated with SP-A (10 µg/ml) and/or bleomycin (BLM 5
mU/ml) simultaneously. After a 2 h (Panel A) or a 4 h (Panel B) incubation, the cells were
processed for quantification of cytokine mRNA by RNase protection assay (RPA). The mRNA
level for TNF-α, IL-1β, and IL-8 in 2µg of total cell RNA was normalized to mRNA for
ribosomal protein L32. Data shown in Panel A and B were derived from 4 and 3 separate
experiments, respectively. Results were given as means ± SEM. The indicated values are
significantly different (p<0.05) from points treated with SP-A (*) and bleomycin (^) alone,
respectively. LPS (0.1ng/ml) treatment was included as a positive control.
Figure 5. The effect of preincubation of SP-A and bleomycin (BLM) on TNF-α production.
Differentiated THP-1 cells were stimulated with SP-A (10 µg/ml) and bleomycin (50 mU/ml)
that were either preincubated together for 15 min at 37oC before adding them to the cells, or were
not preincubated. Data were derived from 4 separate experiments. Results were given as means
± SEM. The indicated values (*) are significantly different from points without preincubation.
Figure 6. The effect of surfactant lipids (Infasurf) on SP-A and bleomycin induced TNF-α
production. Infasurf at indicated concentration was preincubated separately with SP-A
(10µg/ml), bleomycin (5mU/ml), and SP-A + bleomycin for 15min at 37oC before addition to
THP-1 cells. Incubation was continued for 4 h. Cells were incubated for 4h after treatment.
Culture medium was then collected and TNF-α level was quantified by ELISA. Data shown are
from a single experiment in triplicate, but are representative of three experiments. Results are

28
given as means ± SEM. The indicated values (*) are significantly different (p<0.05) from points
obtained in the absence of Infasurf.
Figure 7. The effect of surfactant lipids (Infasurf) on SP-A and bleomycin induced TNF-α and
IL-8 mRNA expression. Infasurf (Infa; 400µg/ml) was preincubated separately with SP-A
(10µg/ml), bleomycin (5mU/ml), and SP-A + bleomycin 15min at 37oC before addition to THP-
1 cells. Cells were incubated for 4h after treatment. Cells were processed for quantification of
cytokine mRNA by RPA. The mRNA levels of TNF-α and IL-8 in 2µg of total RNA was
normalized to the mRNA of the housekeeping gene GAPDH. Data shown are from 3
experiments. Results are given as means ± SEM. The indicated values (*) are significantly
different (p<0.05) from points obtained in the absence of Infasurf. BLM: bleomycin.


Related Documents











