Chronic morphine exposure causes pronounced virus replication in cerebral compartment and accelerated onset of AIDS in SIV/SHIV-infected Indian rhesus macaques Rakesh Kumar a , Suheydi Orsoni a , Lisa Norman a , Ashish S. Verma a , Grissell Tirado a , Luis D. Giavedoni b , Silvija Staprans c , Gregory M. Miller d , Shilpa J. Buch e , Anil Kumar a,f, ⁎ a Laboratory of Viral Immunology, AIDS Research Program and Department of Microbiology, Ponce School of Medicine, Ponce, PR 00732, Puerto Rico b Southwest National Primate Research Center, Southwest Foundation for Biomedical Research, P.O. Box 760549 San Antonio, TX 78245-0549, USA c Emory Vaccine Center, Emory University School of Medicine, Atlanta, GA, USA d Division of Neurochemistry, New England Primate Research Center, Southborough, MA 01772, USA e Department of Physiology, KU Medical Center, Kansas City, KS 66160, USA f Division of Pharmacology, School of Pharmacy, University of Missouri at Kansas City, Kansas City MO 64108, USA Received 12 May 2006; returned to author for revision 14 June 2006; accepted 19 June 2006 Available online 28 July 2006 Abstract Six morphine-exposed and 3 control male Indian rhesus macaques were intravenously inoculated with mixture of SHIV KU , SHIV 89.6 P and SIV/17E-Fr. These animals were followed for a period of 56 weeks in order to determine CD4 and CD8 profile, viral loads in plasma and cerebrospinal fluid (CSF), relative distribution of 3 pathogenic viruses in blood and brain, binding as well neutralizing antibody levels and cellular immune responses. Both morphine-exposed and control macaques showed a precipitous loss of CD4 + T cells; control animals, however, showed a greater tendency to recover these cells than did their morphine-exposed counterparts. The plasma and CSF viral loads were significantly higher in morphine-exposed group than those in the control group. Four morphine-exposed animals succumbed to SIV/SHIV-induced AIDS at week 18, 19, 20 and 51; post-infection with neurological disorders was found in 3 of the 4 animals. At the end of the 56-week observation period, 2 morphine- exposed and 3 control animals were still alive. All 3 viruses replicated in the blood of both morphine-exposed and control macaques, but the cerebral compartment showed a selection phenomenon; only SIV/17E-Fr and SHIV KU successfully crossed the blood brain barrier (BBB). The morphine-exposed macaques further favored viral migration through the blood brain barrier (BBB). SIV/17E-Fr crossed the BBB within 2 weeks in both morphine-exposed and control macaques, whereas SHIV KU crossed the BBB more rapidly in morphine-exposed than in control macaques. Three morphine-exposed macaques (euthanized at weeks 18, 19 and 20) did not develop cellular or humoral immune responses, whereas the other 3 morphine-exposed and 3 control macaques developed both cellular and humoral immune responses. © 2006 Elsevier Inc. All rights reserved. Keywords: SIV; SHIV; AIDS; Indian rhesus macaques, Morphine Introduction Current estimates indicate that injection drug users (IDU) constitute a large cohort among HIV-infected individuals. The recent CDC estimate indicated that as of 2002 more than 27% of total AIDS cases in the country were associated with IDU (CDC, 2002). However, the natural history and progression of HIV infection among IDU remain ambiguous (Alcabes and Friedland, 1995; Metzger et al., 1993). According to one prospective study, AIDS is the most frequent cause of death among IDU (Brancato et al., 1995), but conflicting reports of the mortality rate among HIV-infected IDU have demonstrated both a survival advantage for HIV-infected IDU and lower survival rates among IDU (Alcabes and Friedland, 1995; Selwyn et al., 1992). Nevertheless, it has been clearly demonstrated that HIV-infected IDU have substantial pre- AIDS morbidity and mortality, particularly from bacterial Virology 354 (2006) 192 – 206 www.elsevier.com/locate/yviro ⁎ Corresponding author. Laboratory of Viral Immunology, Department of Microbiology, Ponce School of Medicine, Ponce, PR 00732, Puerto Rico. Fax: +1 775 659 7298. E-mail address: [email protected] (A. Kumar). 0042-6822/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2006.06.020

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

6) 192–206www.elsevier.com/locate/yviro
Virology 354 (200
Chronic morphine exposure causes pronounced virus replication incerebral compartment and accelerated onset of AIDS in
SIV/SHIV-infected Indian rhesus macaques
Rakesh Kumar a, Suheydi Orsoni a, Lisa Norman a, Ashish S. Verma a, Grissell Tirado a,Luis D. Giavedoni b, Silvija Staprans c, Gregory M. Miller d, Shilpa J. Buch e, Anil Kumar a,f,⁎
a Laboratory of Viral Immunology, AIDS Research Program and Department of Microbiology, Ponce School of Medicine, Ponce, PR 00732, Puerto Ricob Southwest National Primate Research Center, Southwest Foundation for Biomedical Research, P.O. Box 760549 San Antonio, TX 78245-0549, USA
c Emory Vaccine Center, Emory University School of Medicine, Atlanta, GA, USAd Division of Neurochemistry, New England Primate Research Center, Southborough, MA 01772, USA
e Department of Physiology, KU Medical Center, Kansas City, KS 66160, USAf Division of Pharmacology, School of Pharmacy, University of Missouri at Kansas City, Kansas City MO 64108, USA
Received 12 May 2006; returned to author for revision 14 June 2006; accepted 19 June 2006Available online 28 July 2006
Abstract
Six morphine-exposed and 3 control male Indian rhesus macaques were intravenously inoculated with mixture of SHIVKU, SHIV89.6P andSIV/17E-Fr. These animals were followed for a period of 56 weeks in order to determine CD4 and CD8 profile, viral loads in plasma andcerebrospinal fluid (CSF), relative distribution of 3 pathogenic viruses in blood and brain, binding as well neutralizing antibody levels and cellularimmune responses. Both morphine-exposed and control macaques showed a precipitous loss of CD4+ T cells; control animals, however, showed agreater tendency to recover these cells than did their morphine-exposed counterparts. The plasma and CSF viral loads were significantly higher inmorphine-exposed group than those in the control group. Four morphine-exposed animals succumbed to SIV/SHIV-induced AIDS at week 18, 19,20 and 51; post-infection with neurological disorders was found in 3 of the 4 animals. At the end of the 56-week observation period, 2 morphine-exposed and 3 control animals were still alive. All 3 viruses replicated in the blood of both morphine-exposed and control macaques, but thecerebral compartment showed a selection phenomenon; only SIV/17E-Fr and SHIVKU successfully crossed the blood brain barrier (BBB). Themorphine-exposed macaques further favored viral migration through the blood brain barrier (BBB). SIV/17E-Fr crossed the BBB within 2 weeksin both morphine-exposed and control macaques, whereas SHIVKU crossed the BBB more rapidly in morphine-exposed than in control macaques.Three morphine-exposed macaques (euthanized at weeks 18, 19 and 20) did not develop cellular or humoral immune responses, whereas the other3 morphine-exposed and 3 control macaques developed both cellular and humoral immune responses.© 2006 Elsevier Inc. All rights reserved.
Keywords: SIV; SHIV; AIDS; Indian rhesus macaques, Morphine
Introduction
Current estimates indicate that injection drug users (IDU)constitute a large cohort among HIV-infected individuals. Therecent CDC estimate indicated that as of 2002 more than 27% oftotal AIDS cases in the country were associated with IDU
⁎ Corresponding author. Laboratory of Viral Immunology, Department ofMicrobiology, Ponce School of Medicine, Ponce, PR 00732, Puerto Rico. Fax:+1 775 659 7298.
E-mail address: [email protected] (A. Kumar).
0042-6822/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.virol.2006.06.020
(CDC, 2002). However, the natural history and progression ofHIV infection among IDU remain ambiguous (Alcabes andFriedland, 1995; Metzger et al., 1993). According to oneprospective study, AIDS is the most frequent cause of deathamong IDU (Brancato et al., 1995), but conflicting reports ofthe mortality rate among HIV-infected IDU have demonstratedboth a survival advantage for HIV-infected IDU and lowersurvival rates among IDU (Alcabes and Friedland, 1995;Selwyn et al., 1992). Nevertheless, it has been clearlydemonstrated that HIV-infected IDU have substantial pre-AIDS morbidity and mortality, particularly from bacterial

193R. Kumar et al. / Virology 354 (2006) 192–206
infections, suggesting that co-infection with other pathogensmay enhance HIV expression and assist in the spread of thedisease (Eisenstein et al., 2001; Hilburger et al., 1997;MacFarlane et al., 2000; Selwyn et al., 1992; Sepkowitz etal., 1995; Shattock et al., 1994). This was further confirmed by areduction in the rate of disease progression after the cessation ofinjection drug use (Ronald et al., 1994). However, the directevidence of a correlation between drug of abuse and increasedviral replication came from several in vitro studies, whereinmorphine and cocaine were shown to intensify both HIV andsimian immunodeficiency virus (SIV) (Peterson et al., 1990,1991, 1992, 1993, 1994; Li et al., 2003; Chuang et al., 1993a,1993b).
More recently, an in vivo study of an SCID mouse model ofHIV infection has shown that cocaine adversely affects thevirus replication (Roth et al., 2002; Tashkin, 2004), but theusefulness of this model remains limited as regards thepathogenesis and progression of the disease. The earlier studiesusing a monkey model of HIV/AIDS also provided conflictingevidence. In one set of studies, morphine dependence resultedin an exacerbation of the SIV infection in rhesus macaques.However, sample size in this study was too small to draw anymeaningful conclusions (Suzuki et al., 2002; Chuang et al.,1993a, 1993b). Furthermore, the increase in virus replication inmorphine-dependent macaques was documented only17 months after infection (Suzuki et al., 2002). In anotherstudy, morphine dependence exerted a protective effect in anSIVsmm9/rhesus monkey model (Donahoe et al., 1993).However, this study lacked concurrent control animals; there-fore, the virus replication in morphine-dependent animals wascompared with historical controls. Both of these studies, inaddition, utilized a challenge model where animals developrelatively slower disease. With the introduction of the simian/human immunodeficiency virus, it has been possible to developa model for HIV/AIDS wherein macaques not only develop ahighly productive infection in the lymphoid system (similar tothat seen in SIV infection) (Daniel et al., 1985; Nathanson et al.,1999), but also exhibit the near total elimination of the CD4+ Tcells in blood and lymphoid tissue during acute infection(Igarashi et al., 1999; Joag et al., 1996; Karlsson et al., 1997;Luciw et al., 1995; Reimann et al., 1996a, 1996b, 1999). Theanimals developing acute disease (i.e., AIDS within 6–8 months) caused by pathogenic SHIV do not develop anti-viral immune responses (Kumar et al., 2001). However, thosesurviving for longer periods develop both cellular and humoralimmune responses (Kumar et al., 2001; Silverstein et al., 2000;Stipp et al., 2000). Prolonged survival of the infected animalhas been correlated with humoral immune responses mediatedby neutralizing antibodies and cellular immune responsesmediated by CD8+ T cells. Such responses are similar to thosefound in long-term non-progressors of HIV infection. In one ofour previously reported studies, the mixture of 2 SHIVs(SHIVKU and SHIV89.6P) and a neuropathogenic SIV provedan excellent challenge system to develop rapid and reprodu-cible disease in the rhesus macaques (Kumar et al., 2002).Utilizing this model, we have shown that, although chronicmorphine exposure did not alter the peak viral loads in rhesus
macaques, it caused significantly higher viral replication at6 weeks post-infection (and thereafter) in the brain as well as8 weeks (and thereafter) in the blood (Kumar et al., 2004).
In view of this evidence, it appears that opiates enhancereplication of HIV and SIV, and opiate dependence may, infact, accelerate the onset of disease. However, no in vivostudies have been undertaken to evaluate the effect of opiatedependence on the onset of clinical disease. In this study, wesought to address this issue, and for the first time, weunequivocally show that chronic morphine exposure causedrapid onset of the disease in a majority of virus-infectedanimals and compromised the development of virus-specificimmune responses in at least 50% of the morphine-exposedanimals. Two-thirds of the morphine-exposed but none of thecontrol animals succumbed to SIV/SHIV-induced AIDS within51 weeks after infection.
Results
Morphine-exposed and control animals were monitoredfor complete blood counts at the time of every bleeding.None of the animals showed any significant change in anyof the different parameters except monocyte counts. Themonocyte counts in morphine-exposed animals were found tobe generally elevated; that coincided with the inception ofclinical disease. The increase ranged between 11- and 20-foldin 4 animals (1/28Q, 1/04L, 1/42N and 1/56L), those whichdeveloped disease within the course of the study (results notshown).
Morphine-exposed and control macaques showed a precipitousloss in CD4+ T cells
The morphine-exposed and control macaques were followedin order to determine CD4, CD8 counts and CD4/CD8 ratiothroughout the 56-week observation period. The results of theCD4+ T-cell counts and the CD4/CD8 ratio are shown in Fig. 1.Five of the 6 morphine-exposed animals lost >95% of theirCD4+ T cells within 3 weeks after infection, and levelsremained low throughout the observation period. The 6thanimal (1/52N) showed more than a 90% loss in CD4population at week 1 after infection; this loss was followed bya remarkable recovery, and the macaque attained its pre-infection level within 16 weeks. Since then, this animal hasmaintained a high CD4 count throughout the observationperiod. Alternately, control animals exhibited an 85–90% lossin CD4+ T-cell numbers at week 3 followed by a reasonablerecovery in all 3 animals. One macaque (2/02P) underwentanother round of CD4 loss (starting week 12, post-infection);this animal had maintained very low CD4 counts throughout thestudy. The other two animals have maintained reasonable CD4counts. There was no statistical difference observed betweenmorphine and control groups (p=0.5 and p=0.9 at weeks 8 and18, respectively). However, the CD4 loss was highly significantin the morphine group (after the values from macaque 1/52Nthat showed substantial CD4 recovery – p=0.02 and 0.06 atweeks 8 and 18, respectively – were excluded).
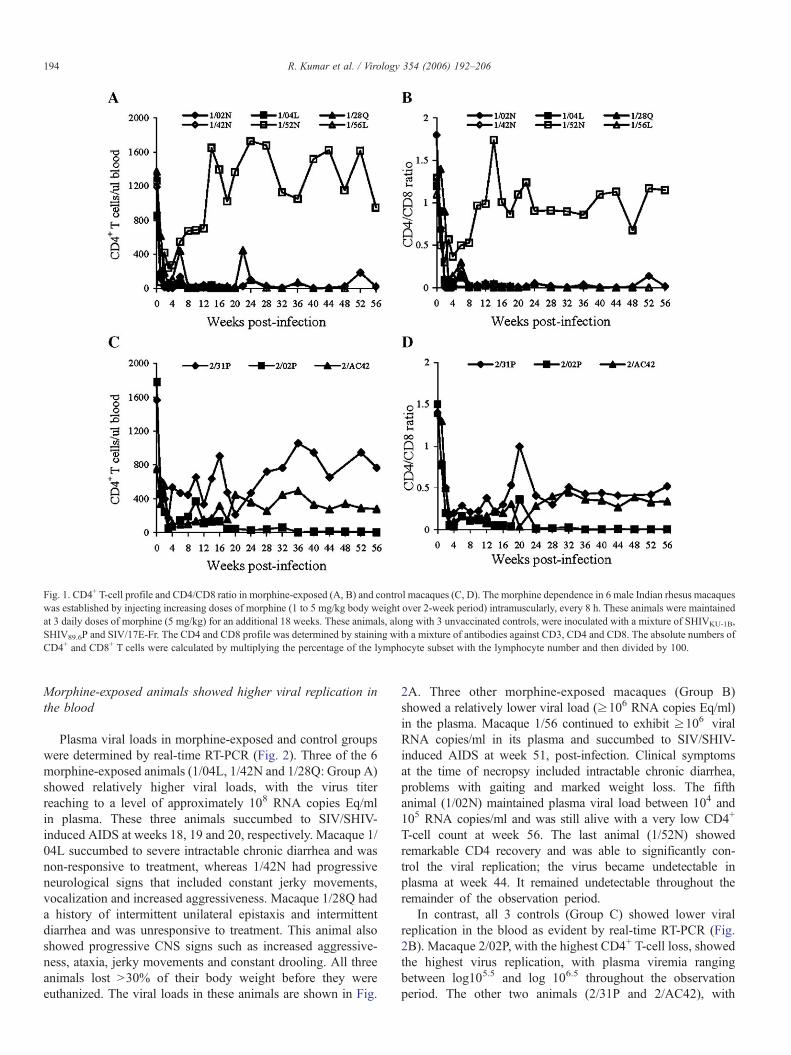
Fig. 1. CD4+ T-cell profile and CD4/CD8 ratio in morphine-exposed (A, B) and control macaques (C, D). The morphine dependence in 6 male Indian rhesus macaqueswas established by injecting increasing doses of morphine (1 to 5 mg/kg body weight over 2-week period) intramuscularly, every 8 h. These animals were maintainedat 3 daily doses of morphine (5 mg/kg) for an additional 18 weeks. These animals, along with 3 unvaccinated controls, were inoculated with a mixture of SHIVKU-1B,SHIV89.6P and SIV/17E-Fr. The CD4 and CD8 profile was determined by staining with a mixture of antibodies against CD3, CD4 and CD8. The absolute numbers ofCD4+ and CD8+ T cells were calculated by multiplying the percentage of the lymphocyte subset with the lymphocyte number and then divided by 100.
194 R. Kumar et al. / Virology 354 (2006) 192–206
Morphine-exposed animals showed higher viral replication inthe blood
Plasma viral loads in morphine-exposed and control groupswere determined by real-time RT-PCR (Fig. 2). Three of the 6morphine-exposed animals (1/04L, 1/42N and 1/28Q: Group A)showed relatively higher viral loads, with the virus titerreaching to a level of approximately 108 RNA copies Eq/mlin plasma. These three animals succumbed to SIV/SHIV-induced AIDS at weeks 18, 19 and 20, respectively. Macaque 1/04L succumbed to severe intractable chronic diarrhea and wasnon-responsive to treatment, whereas 1/42N had progressiveneurological signs that included constant jerky movements,vocalization and increased aggressiveness. Macaque 1/28Q hada history of intermittent unilateral epistaxis and intermittentdiarrhea and was unresponsive to treatment. This animal alsoshowed progressive CNS signs such as increased aggressive-ness, ataxia, jerky movements and constant drooling. All threeanimals lost >30% of their body weight before they wereeuthanized. The viral loads in these animals are shown in Fig.
2A. Three other morphine-exposed macaques (Group B)showed a relatively lower viral load (≥106 RNA copies Eq/ml)in the plasma. Macaque 1/56 continued to exhibit ≥106 viralRNA copies/ml in its plasma and succumbed to SIV/SHIV-induced AIDS at week 51, post-infection. Clinical symptomsat the time of necropsy included intractable chronic diarrhea,problems with gaiting and marked weight loss. The fifthanimal (1/02N) maintained plasma viral load between 104 and105 RNA copies/ml and was still alive with a very low CD4+
T-cell count at week 56. The last animal (1/52N) showedremarkable CD4 recovery and was able to significantly con-trol the viral replication; the virus became undetectable inplasma at week 44. It remained undetectable throughout theremainder of the observation period.
In contrast, all 3 controls (Group C) showed lower viralreplication in the blood as evident by real-time RT-PCR (Fig.2B). Macaque 2/02P, with the highest CD4+ T-cell loss, showedthe highest virus replication, with plasma viremia rangingbetween log105.5 and log 106.5 throughout the observationperiod. The other two animals (2/31P and 2/AC42), with
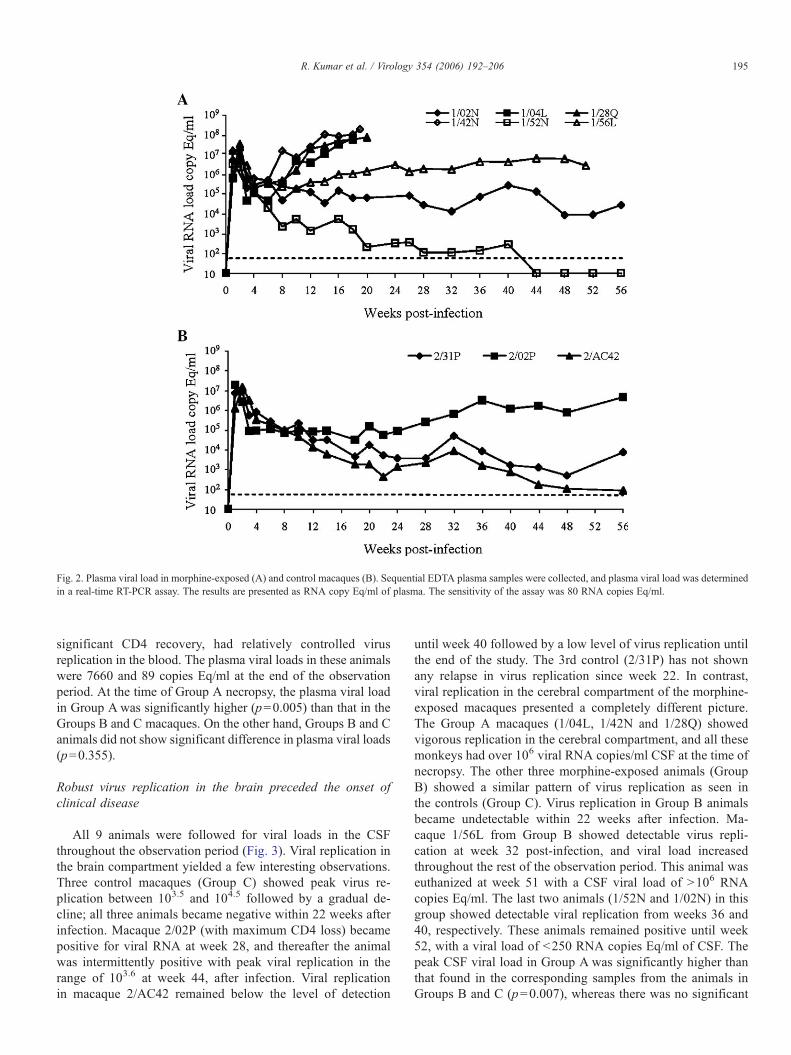
Fig. 2. Plasma viral load in morphine-exposed (A) and control macaques (B). Sequential EDTA plasma samples were collected, and plasma viral load was determinedin a real-time RT-PCR assay. The results are presented as RNA copy Eq/ml of plasma. The sensitivity of the assay was 80 RNA copies Eq/ml.
195R. Kumar et al. / Virology 354 (2006) 192–206
significant CD4 recovery, had relatively controlled virusreplication in the blood. The plasma viral loads in these animalswere 7660 and 89 copies Eq/ml at the end of the observationperiod. At the time of Group A necropsy, the plasma viral loadin Group A was significantly higher (p=0.005) than that in theGroups B and C macaques. On the other hand, Groups B and Canimals did not show significant difference in plasma viral loads(p=0.355).
Robust virus replication in the brain preceded the onset ofclinical disease
All 9 animals were followed for viral loads in the CSFthroughout the observation period (Fig. 3). Viral replication inthe brain compartment yielded a few interesting observations.Three control macaques (Group C) showed peak virus re-plication between 103.5 and 104.5 followed by a gradual de-cline; all three animals became negative within 22 weeks afterinfection. Macaque 2/02P (with maximum CD4 loss) becamepositive for viral RNA at week 28, and thereafter the animalwas intermittently positive with peak viral replication in therange of 103.6 at week 44, after infection. Viral replicationin macaque 2/AC42 remained below the level of detection
until week 40 followed by a low level of virus replication untilthe end of the study. The 3rd control (2/31P) has not shownany relapse in virus replication since week 22. In contrast,viral replication in the cerebral compartment of the morphine-exposed macaques presented a completely different picture.The Group A macaques (1/04L, 1/42N and 1/28Q) showedvigorous replication in the cerebral compartment, and all thesemonkeys had over 106 viral RNA copies/ml CSF at the time ofnecropsy. The other three morphine-exposed animals (GroupB) showed a similar pattern of virus replication as seen inthe controls (Group C). Virus replication in Group B animalsbecame undetectable within 22 weeks after infection. Ma-caque 1/56L from Group B showed detectable virus repli-cation at week 32 post-infection, and viral load increasedthroughout the rest of the observation period. This animal waseuthanized at week 51 with a CSF viral load of >106 RNAcopies Eq/ml. The last two animals (1/52N and 1/02N) in thisgroup showed detectable viral replication from weeks 36 and40, respectively. These animals remained positive until week52, with a viral load of <250 RNA copies Eq/ml of CSF. Thepeak CSF viral load in Group A was significantly higher thanthat found in the corresponding samples from the animals inGroups B and C (p=0.007), whereas there was no significant
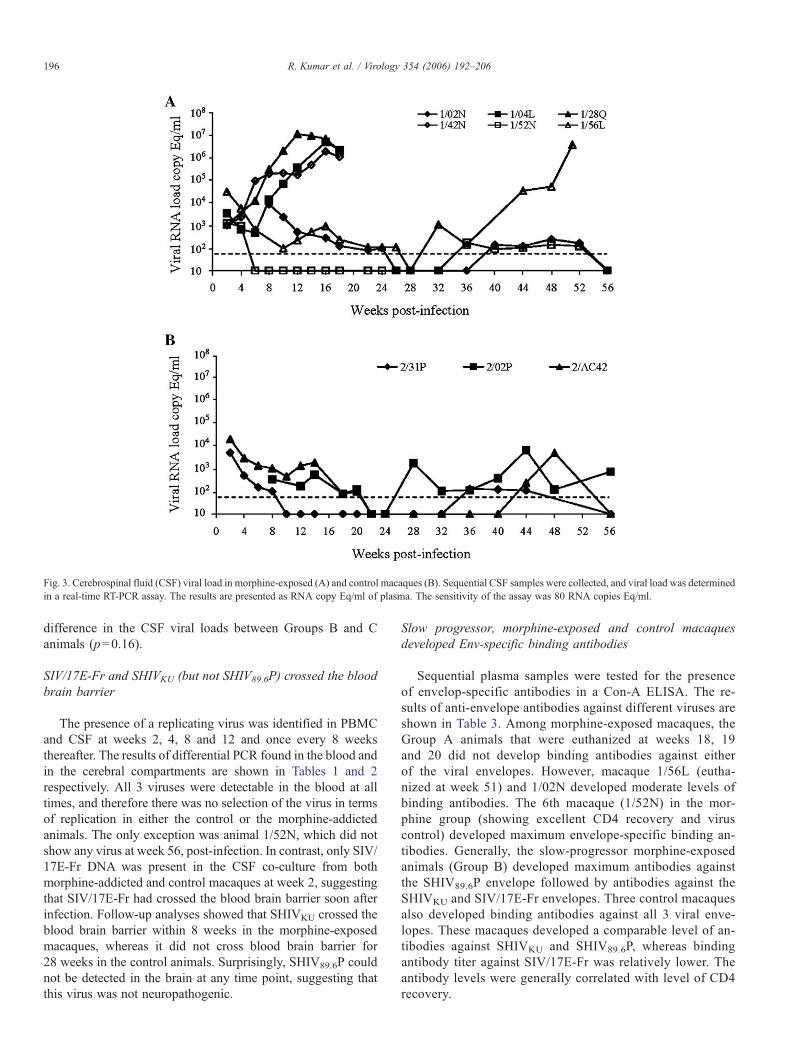
Fig. 3. Cerebrospinal fluid (CSF) viral load in morphine-exposed (A) and control macaques (B). Sequential CSF samples were collected, and viral load was determinedin a real-time RT-PCR assay. The results are presented as RNA copy Eq/ml of plasma. The sensitivity of the assay was 80 RNA copies Eq/ml.
196 R. Kumar et al. / Virology 354 (2006) 192–206
difference in the CSF viral loads between Groups B and Canimals (p=0.16).
SIV/17E-Fr and SHIVKU (but not SHIV89.6P) crossed the bloodbrain barrier
The presence of a replicating virus was identified in PBMCand CSF at weeks 2, 4, 8 and 12 and once every 8 weeksthereafter. The results of differential PCR found in the blood andin the cerebral compartments are shown in Tables 1 and 2respectively. All 3 viruses were detectable in the blood at alltimes, and therefore there was no selection of the virus in termsof replication in either the control or the morphine-addictedanimals. The only exception was animal 1/52N, which did notshow any virus at week 56, post-infection. In contrast, only SIV/17E-Fr DNA was present in the CSF co-culture from bothmorphine-addicted and control macaques at week 2, suggestingthat SIV/17E-Fr had crossed the blood brain barrier soon afterinfection. Follow-up analyses showed that SHIVKU crossed theblood brain barrier within 8 weeks in the morphine-exposedmacaques, whereas it did not cross blood brain barrier for28 weeks in the control animals. Surprisingly, SHIV89.6P couldnot be detected in the brain at any time point, suggesting thatthis virus was not neuropathogenic.
Slow progressor, morphine-exposed and control macaquesdeveloped Env-specific binding antibodies
Sequential plasma samples were tested for the presenceof envelop-specific antibodies in a Con-A ELISA. The re-sults of anti-envelope antibodies against different viruses areshown in Table 3. Among morphine-exposed macaques, theGroup A animals that were euthanized at weeks 18, 19and 20 did not develop binding antibodies against eitherof the viral envelopes. However, macaque 1/56L (eutha-nized at week 51) and 1/02N developed moderate levels ofbinding antibodies. The 6th macaque (1/52N) in the mor-phine group (showing excellent CD4 recovery and viruscontrol) developed maximum envelope-specific binding an-tibodies. Generally, the slow-progressor morphine-exposedanimals (Group B) developed maximum antibodies againstthe SHIV89.6P envelope followed by antibodies against theSHIVKU and SIV/17E-Fr envelopes. Three control macaquesalso developed binding antibodies against all 3 viral enve-lopes. These macaques developed a comparable level of an-tibodies against SHIVKU and SHIV89.6P, whereas bindingantibody titer against SIV/17E-Fr was relatively lower. Theantibody levels were generally correlated with level of CD4recovery.
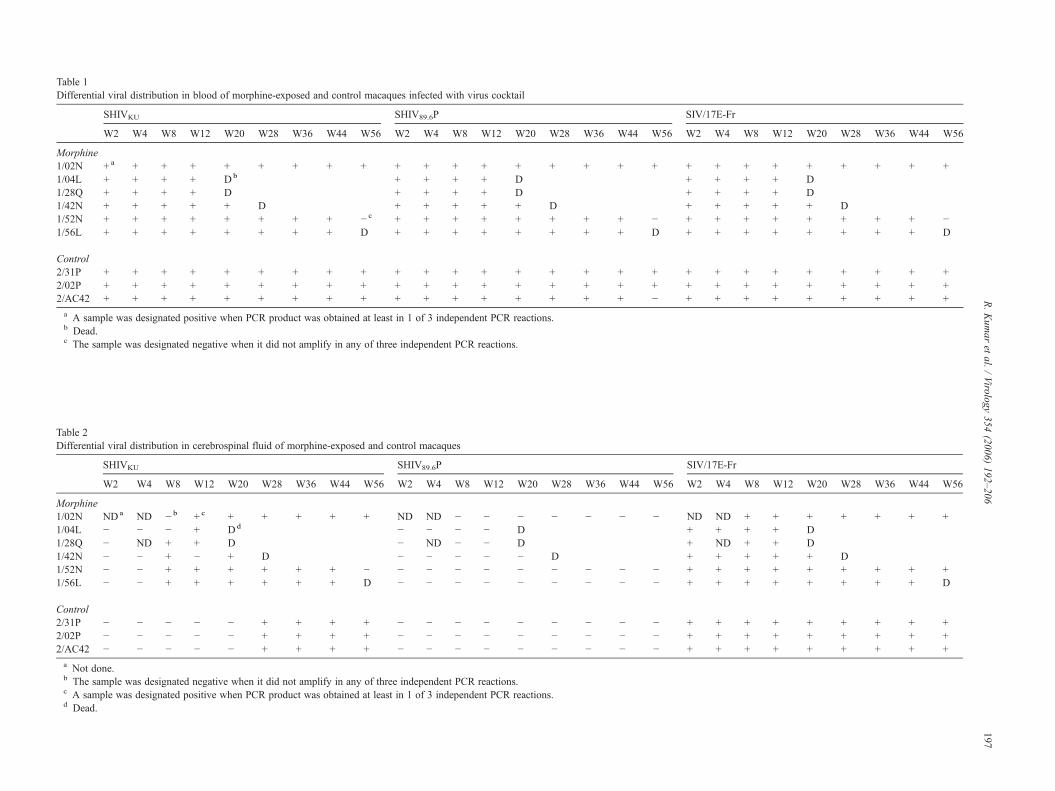
Table 1Differential viral distribution in blood of morphine-exposed and control macaques infected with virus cocktail
SHIVKU SHIV89.6P SIV/17E-Fr
W2 W4 W8 W12 W20 W28 W36 W44 W56 W2 W4 W8 W12 W20 W28 W36 W44 W56 W2 W4 W8 W12 W20 W28 W36 W44 W56
Morphine1/02N + a + + + + + + + + + + + + + + + + + + + + + + + + + +1/04L + + + + Db + + + + D + + + + D1/28Q + + + + D + + + + D + + + + D1/42N + + + + + D + + + + + D + + + + + D1/52N + + + + + + + + − c + + + + + + + + − + + + + + + + + −1/56L + + + + + + + + D + + + + + + + + D + + + + + + + + D
Control2/31P + + + + + + + + + + + + + + + + + + + + + + + + + + +2/02P + + + + + + + + + + + + + + + + + + + + + + + + + + +2/AC42 + + + + + + + + + + + + + + + + + − + + + + + + + + +
a A sample was designated positive when PCR product was obtained at least in 1 of 3 independent PCR reactions.b Dead.c The sample was designated negative when it did not amplify in any of three independent PCR reactions.
Table 2Differential viral distribution in cerebrospinal fluid of morphine-exposed and control macaques
SHIVKU SHIV89.6P SIV/17E-Fr
W2 W4 W8 W12 W20 W28 W36 W44 W56 W2 W4 W8 W12 W20 W28 W36 W44 W56 W2 W4 W8 W12 W20 W28 W36 W44 W56
Morphine1/02N NDa ND − b + c + + + + + ND ND − − − − − − − ND ND + + + + + + +1/04L − − − + Dd − − − − D + + + + D1/28Q − ND + + D − ND − − D + ND + + D1/42N − − + − + D − − − − − D + + + + + D1/52N − − + + + + + + − − − − − − − − − − + + + + + + + + +1/56L − − + + + + + + D − − − − − − − − − + + + + + + + + D
Control2/31P − − − − − + + + + − − − − − − − − − + + + + + + + + +2/02P − − − − − + + + + − − − − − − − − − + + + + + + + + +2/AC42 − − − − − + + + + − − − − − − − − − + + + + + + + + +a Not done.b The sample was designated negative when it did not amplify in any of three independent PCR reactions.c A sample was designated positive when PCR product was obtained at least in 1 of 3 independent PCR reactions.d Dead.
197R.Kum
aret
al./Virology
354(2006)
192–206
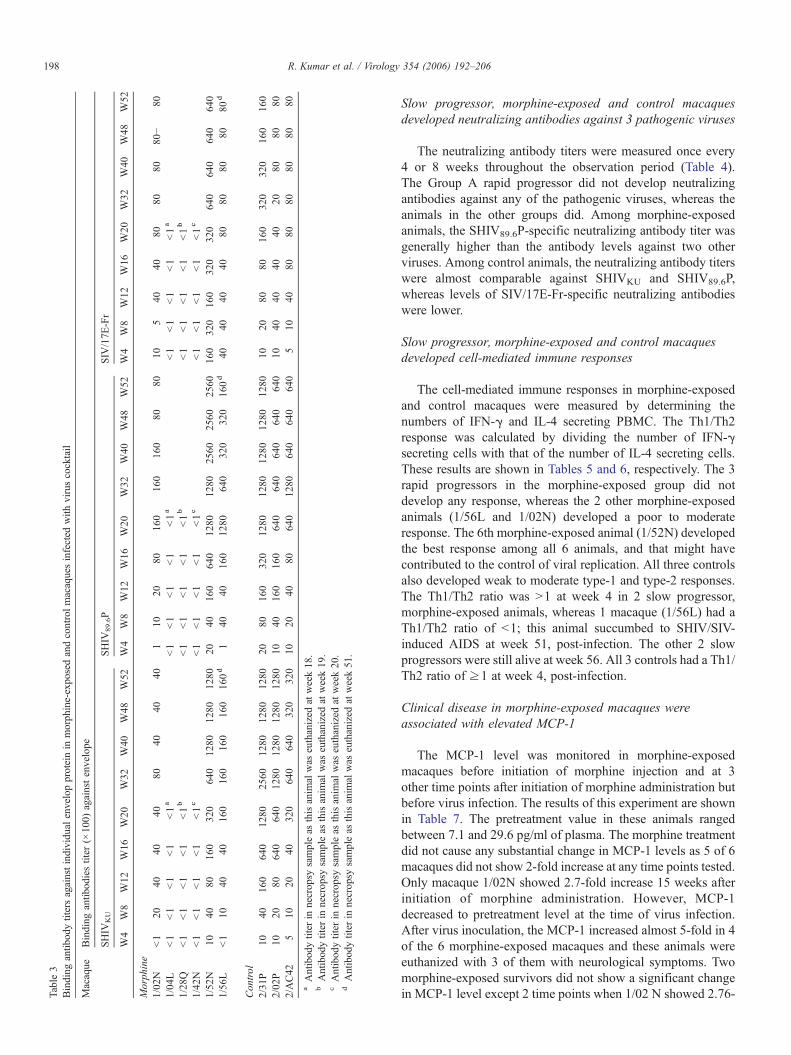
Table3
Binding
antib
odytitersagainstindividu
alenvelopproteinin
morph
ine-exposedandcontrolmacaquesinfected
with
viruscocktail
Macaque
Binding
antib
odiestiter
(×10
0)againstenvelope
SHIV
KU
SHIV
89.6P
SIV
/17E
-Fr
W4
W8
W12
W16
W20
W32
W40
W48
W52
W4
W8
W12
W16
W20
W32
W40
W48
W52
W4
W8
W12
W16
W20
W32
W40
W48
W52
Morph
ine
1/02
N<1
2040
4040
8040
4040
110
2080
160
160
160
8080
105
4040
8080
8080−
801/04
L<1
<1
<1
<1
<1a
<1
<1
<1
<1
<1a
<1
<1
<1
<1
<1a
1/28
Q<1
<1
<1
<1
<1b
<1
<1
<1
<1
<1b
<1
<1
<1
<1
<1b
1/42
N<1
<1
<1
<1
<1c
<1
<1
<1
<1
<1c
<1
<1
<1
<1
<1c
1/52
N10
4080
160
320
640
1280
1280
1280
2040
160
640
1280
1280
2560
2560
2560
160
320
160
320
320
640
640
640
640
1/56
L<1
1040
4016
016
016
016
016
0d
140
4016
012
8064
032
032
016
0d
4040
4040
8080
8080
80d
Con
trol
2/31
P10
4016
064
012
8025
6012
8012
8012
8020
8016
032
012
8012
8012
8012
8012
8010
2080
8016
032
032
016
016
02/02
P10
2080
640
640
1280
1280
1280
1280
1040
160
160
640
640
640
640
640
1040
4040
4020
8080
802/AC42
510
2040
320
640
640
320
320
1020
4080
640
1280
640
640
640
510
4080
8080
8080
80aAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek18.
bAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek19
.cAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek20.
dAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek51
.
198 R. Kumar et al. / Virology 354 (2006) 192–206
Slow progressor, morphine-exposed and control macaquesdeveloped neutralizing antibodies against 3 pathogenic viruses
The neutralizing antibody titers were measured once every4 or 8 weeks throughout the observation period (Table 4).The Group A rapid progressor did not develop neutralizingantibodies against any of the pathogenic viruses, whereas theanimals in the other groups did. Among morphine-exposedanimals, the SHIV89.6P-specific neutralizing antibody titer wasgenerally higher than the antibody levels against two otherviruses. Among control animals, the neutralizing antibody titerswere almost comparable against SHIVKU and SHIV89.6P,whereas levels of SIV/17E-Fr-specific neutralizing antibodieswere lower.
Slow progressor, morphine-exposed and control macaquesdeveloped cell-mediated immune responses
The cell-mediated immune responses in morphine-exposedand control macaques were measured by determining thenumbers of IFN-γ and IL-4 secreting PBMC. The Th1/Th2response was calculated by dividing the number of IFN-γsecreting cells with that of the number of IL-4 secreting cells.These results are shown in Tables 5 and 6, respectively. The 3rapid progressors in the morphine-exposed group did notdevelop any response, whereas the 2 other morphine-exposedanimals (1/56L and 1/02N) developed a poor to moderateresponse. The 6th morphine-exposed animal (1/52N) developedthe best response among all 6 animals, and that might havecontributed to the control of viral replication. All three controlsalso developed weak to moderate type-1 and type-2 responses.The Th1/Th2 ratio was >1 at week 4 in 2 slow progressor,morphine-exposed animals, whereas 1 macaque (1/56L) had aTh1/Th2 ratio of <1; this animal succumbed to SHIV/SIV-induced AIDS at week 51, post-infection. The other 2 slowprogressors were still alive at week 56. All 3 controls had a Th1/Th2 ratio of ≥1 at week 4, post-infection.
Clinical disease in morphine-exposed macaques wereassociated with elevated MCP-1
The MCP-1 level was monitored in morphine-exposedmacaques before initiation of morphine injection and at 3other time points after initiation of morphine administration butbefore virus infection. The results of this experiment are shownin Table 7. The pretreatment value in these animals rangedbetween 7.1 and 29.6 pg/ml of plasma. The morphine treatmentdid not cause any substantial change in MCP-1 levels as 5 of 6macaques did not show 2-fold increase at any time points tested.Only macaque 1/02N showed 2.7-fold increase 15 weeks afterinitiation of morphine administration. However, MCP-1decreased to pretreatment level at the time of virus infection.After virus inoculation, the MCP-1 increased almost 5-fold in 4of the 6 morphine-exposed macaques and these animals wereeuthanized with 3 of them with neurological symptoms. Twomorphine-exposed survivors did not show a significant changein MCP-1 level except 2 time points when 1/02 N showed 2.76-
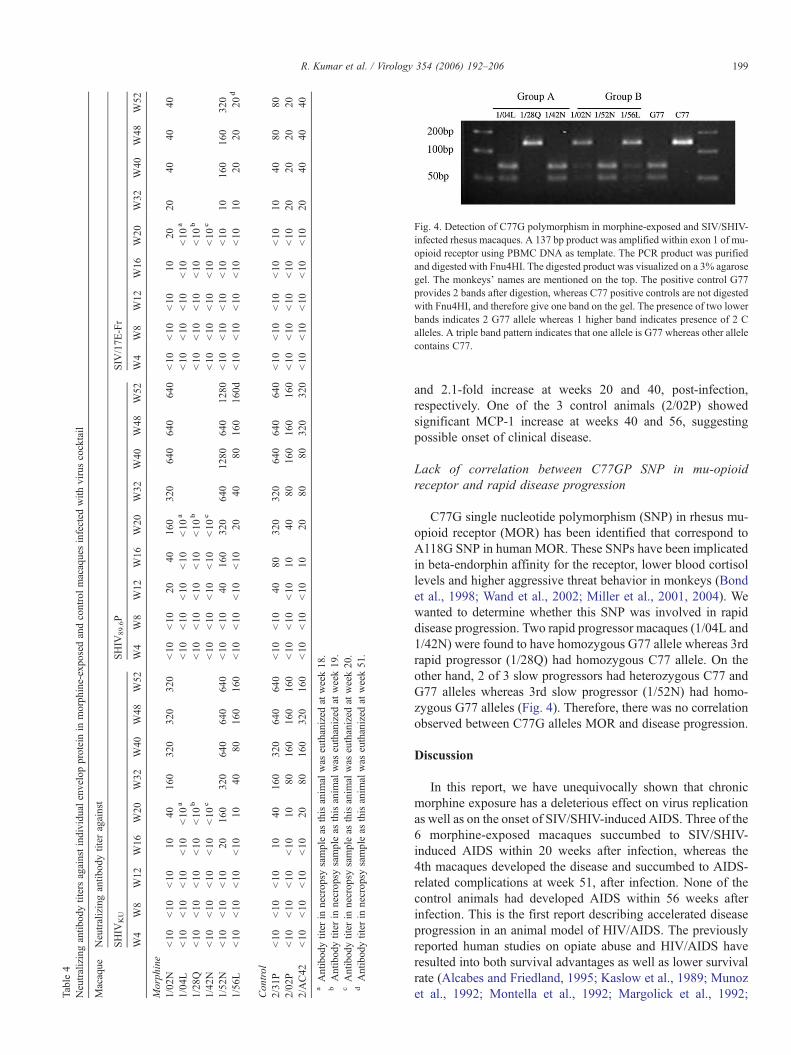
Table4
Neutralizingantib
odytitersagainstindividual
envelopproteinin
morphine-exposedandcontrolmacaquesinfected
with
viruscocktail
Macaque
Neutralizingantib
odytiter
against
SHIV
KU
SHIV
89.6P
SIV
/17E
-Fr
W4
W8
W12
W16
W20
W32
W40
W48
W52
W4
W8
W12
W16
W20
W32
W40
W48
W52
W4
W8
W12
W16
W20
W32
W40
W48
W52
Morph
ine
1/02
N<10
<10
<10
1040
160
320
320
320
<10
<10
2040
160
320
640
640
640
<10
<10
<10
1020
2040
4040
1/04
L<10
<10
<10
<10
<10
a<10
<10
<10
<10
<10
a<10
<10
<10
<10
<10
a
1/28
Q<10
<10
<10
<10
<10
b<10
<10
<10
<10
<10
b<10
<10
<10
<10
<10
b
1/42
N<10
<10
<10
<10
<10
c<10
<10
<10
<10
<10
c<10
<10
<10
<10
<10
c
1/52
N<10
<10
<10
2016
032
064
064
064
0<10
<10
4016
032
064
012
8064
012
80<10
<10
<10
<10
<10
1016
016
032
01/56
L<10
<10
<10
<10
1040
8016
016
0<10
<10
<10
<10
2040
8016
016
0d<10
<10
<10
<10
<10
1020
2020
d
Con
trol
2/31
P<10
<10
<10
1040
160
320
640
640
<10
<10
4080
320
320
640
640
640
<10
<10
<10
<10
<10
1040
8080
2/02
P<10
<10
<10
<10
1080
160
160
160
<10
<10
<10
1040
8016
016
016
0<10
<10
<10
<10
<10
2020
2020
2/AC42
<10
<10
<10
<10
2080
160
320
160
<10
<10
<10
1020
8080
320
320
<10
<10
<10
<10
<10
2040
4040
aAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek18.
bAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek19
.cAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek20.
dAntibodytiter
innecropsy
sampleas
thisanim
alwas
euthanized
atweek51
.
Fig. 4. Detection of C77G polymorphism in morphine-exposed and SIV/SHIV-infected rhesus macaques. A 137 bp product was amplified within exon 1 of mu-opioid receptor using PBMC DNA as template. The PCR product was purifiedand digested with Fnu4HI. The digested product was visualized on a 3% agarosegel. The monkeys' names are mentioned on the top. The positive control G77provides 2 bands after digestion, whereas C77 positive controls are not digestedwith Fnu4HI, and therefore give one band on the gel. The presence of two lowerbands indicates 2 G77 allele whereas 1 higher band indicates presence of 2 Calleles. A triple band pattern indicates that one allele is G77 whereas other allelecontains C77.
199R. Kumar et al. / Virology 354 (2006) 192–206
and 2.1-fold increase at weeks 20 and 40, post-infection,respectively. One of the 3 control animals (2/02P) showedsignificant MCP-1 increase at weeks 40 and 56, suggestingpossible onset of clinical disease.
Lack of correlation between C77GP SNP in mu-opioidreceptor and rapid disease progression
C77G single nucleotide polymorphism (SNP) in rhesus mu-opioid receptor (MOR) has been identified that correspond toA118G SNP in human MOR. These SNPs have been implicatedin beta-endorphin affinity for the receptor, lower blood cortisollevels and higher aggressive threat behavior in monkeys (Bondet al., 1998; Wand et al., 2002; Miller et al., 2001, 2004). Wewanted to determine whether this SNP was involved in rapiddisease progression. Two rapid progressor macaques (1/04L and1/42N) were found to have homozygous G77 allele whereas 3rdrapid progressor (1/28Q) had homozygous C77 allele. On theother hand, 2 of 3 slow progressors had heterozygous C77 andG77 alleles whereas 3rd slow progressor (1/52N) had homo-zygous G77 alleles (Fig. 4). Therefore, there was no correlationobserved between C77G alleles MOR and disease progression.
Discussion
In this report, we have unequivocally shown that chronicmorphine exposure has a deleterious effect on virus replicationas well as on the onset of SIV/SHIV-induced AIDS. Three of the6 morphine-exposed macaques succumbed to SIV/SHIV-induced AIDS within 20 weeks after infection, whereas the4th macaques developed the disease and succumbed to AIDS-related complications at week 51, after infection. None of thecontrol animals had developed AIDS within 56 weeks afterinfection. This is the first report describing accelerated diseaseprogression in an animal model of HIV/AIDS. The previouslyreported human studies on opiate abuse and HIV/AIDS haveresulted into both survival advantages as well as lower survivalrate (Alcabes and Friedland, 1995; Kaslow et al., 1989; Munozet al., 1992; Montella et al., 1992; Margolick et al., 1992;
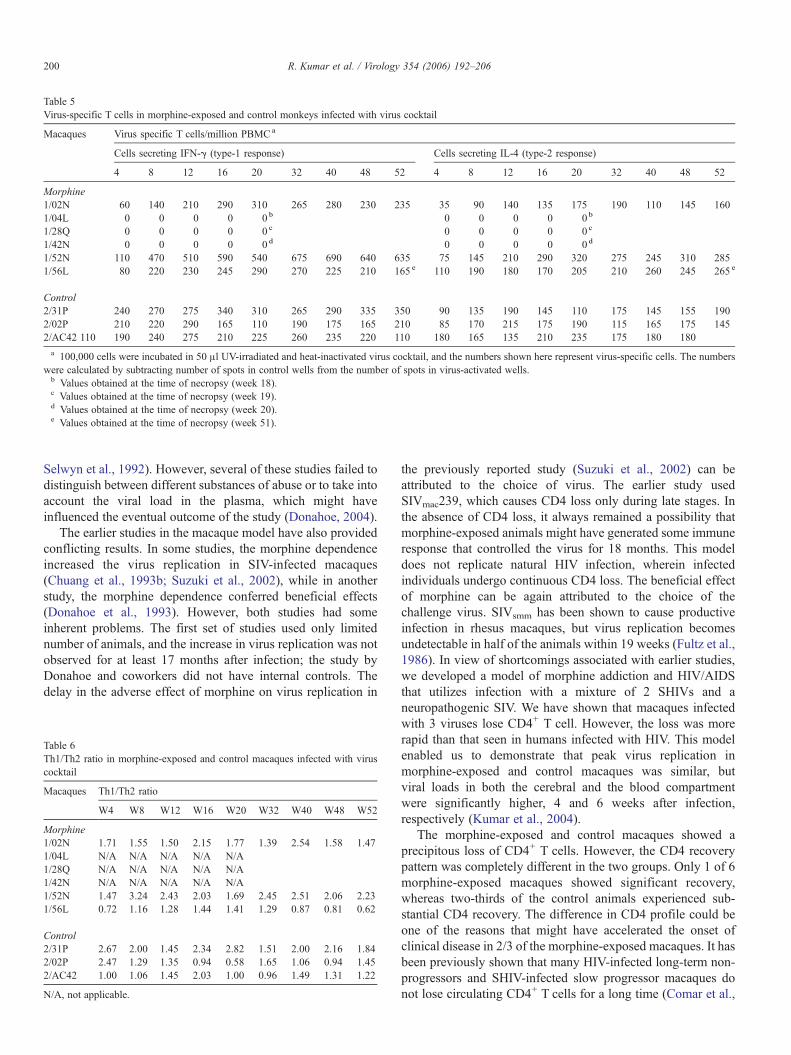
Table 5Virus-specific T cells in morphine-exposed and control monkeys infected with virus cocktail
Macaques Virus specific T cells/million PBMCa
Cells secreting IFN-γ (type-1 response) Cells secreting IL-4 (type-2 response)
4 8 12 16 20 32 40 48 52 4 8 12 16 20 32 40 48 52
Morphine1/02N 60 140 210 290 310 265 280 230 235 35 90 140 135 175 190 110 145 1601/04L 0 0 0 0 0 b 0 0 0 0 0 b
1/28Q 0 0 0 0 0 c 0 0 0 0 0 c
1/42N 0 0 0 0 0 d 0 0 0 0 0 d
1/52N 110 470 510 590 540 675 690 640 635 75 145 210 290 320 275 245 310 2851/56L 80 220 230 245 290 270 225 210 165 e 110 190 180 170 205 210 260 245 265 e
Control2/31P 240 270 275 340 310 265 290 335 350 90 135 190 145 110 175 145 155 1902/02P 210 220 290 165 110 190 175 165 210 85 170 215 175 190 115 165 175 1452/AC42 110 190 240 275 210 225 260 235 220 110 180 165 135 210 235 175 180 180
a 100,000 cells were incubated in 50 μl UV-irradiated and heat-inactivated virus cocktail, and the numbers shown here represent virus-specific cells. The numberswere calculated by subtracting number of spots in control wells from the number of spots in virus-activated wells.b Values obtained at the time of necropsy (week 18).c Values obtained at the time of necropsy (week 19).d Values obtained at the time of necropsy (week 20).e Values obtained at the time of necropsy (week 51).
200 R. Kumar et al. / Virology 354 (2006) 192–206
Selwyn et al., 1992). However, several of these studies failed todistinguish between different substances of abuse or to take intoaccount the viral load in the plasma, which might haveinfluenced the eventual outcome of the study (Donahoe, 2004).
The earlier studies in the macaque model have also providedconflicting results. In some studies, the morphine dependenceincreased the virus replication in SIV-infected macaques(Chuang et al., 1993b; Suzuki et al., 2002), while in anotherstudy, the morphine dependence conferred beneficial effects(Donahoe et al., 1993). However, both studies had someinherent problems. The first set of studies used only limitednumber of animals, and the increase in virus replication was notobserved for at least 17 months after infection; the study byDonahoe and coworkers did not have internal controls. Thedelay in the adverse effect of morphine on virus replication in
Table 6Th1/Th2 ratio in morphine-exposed and control macaques infected with viruscocktail
Macaques Th1/Th2 ratio
W4 W8 W12 W16 W20 W32 W40 W48 W52
Morphine1/02N 1.71 1.55 1.50 2.15 1.77 1.39 2.54 1.58 1.471/04L N/A N/A N/A N/A N/A1/28Q N/A N/A N/A N/A N/A1/42N N/A N/A N/A N/A N/A1/52N 1.47 3.24 2.43 2.03 1.69 2.45 2.51 2.06 2.231/56L 0.72 1.16 1.28 1.44 1.41 1.29 0.87 0.81 0.62
Control2/31P 2.67 2.00 1.45 2.34 2.82 1.51 2.00 2.16 1.842/02P 2.47 1.29 1.35 0.94 0.58 1.65 1.06 0.94 1.452/AC42 1.00 1.06 1.45 2.03 1.00 0.96 1.49 1.31 1.22
N/A, not applicable.
the previously reported study (Suzuki et al., 2002) can beattributed to the choice of virus. The earlier study usedSIVmac239, which causes CD4 loss only during late stages. Inthe absence of CD4 loss, it always remained a possibility thatmorphine-exposed animals might have generated some immuneresponse that controlled the virus for 18 months. This modeldoes not replicate natural HIV infection, wherein infectedindividuals undergo continuous CD4 loss. The beneficial effectof morphine can be again attributed to the choice of thechallenge virus. SIVsmm has been shown to cause productiveinfection in rhesus macaques, but virus replication becomesundetectable in half of the animals within 19 weeks (Fultz et al.,1986). In view of shortcomings associated with earlier studies,we developed a model of morphine addiction and HIV/AIDSthat utilizes infection with a mixture of 2 SHIVs and aneuropathogenic SIV. We have shown that macaques infectedwith 3 viruses lose CD4+ T cell. However, the loss was morerapid than that seen in humans infected with HIV. This modelenabled us to demonstrate that peak virus replication inmorphine-exposed and control macaques was similar, butviral loads in both the cerebral and the blood compartmentwere significantly higher, 4 and 6 weeks after infection,respectively (Kumar et al., 2004).
The morphine-exposed and control macaques showed aprecipitous loss of CD4+ T cells. However, the CD4 recoverypattern was completely different in the two groups. Only 1 of 6morphine-exposed macaques showed significant recovery,whereas two-thirds of the control animals experienced sub-stantial CD4 recovery. The difference in CD4 profile could beone of the reasons that might have accelerated the onset ofclinical disease in 2/3 of the morphine-exposed macaques. It hasbeen previously shown that many HIV-infected long-term non-progressors and SHIV-infected slow progressor macaques donot lose circulating CD4+ T cells for a long time (Comar et al.,
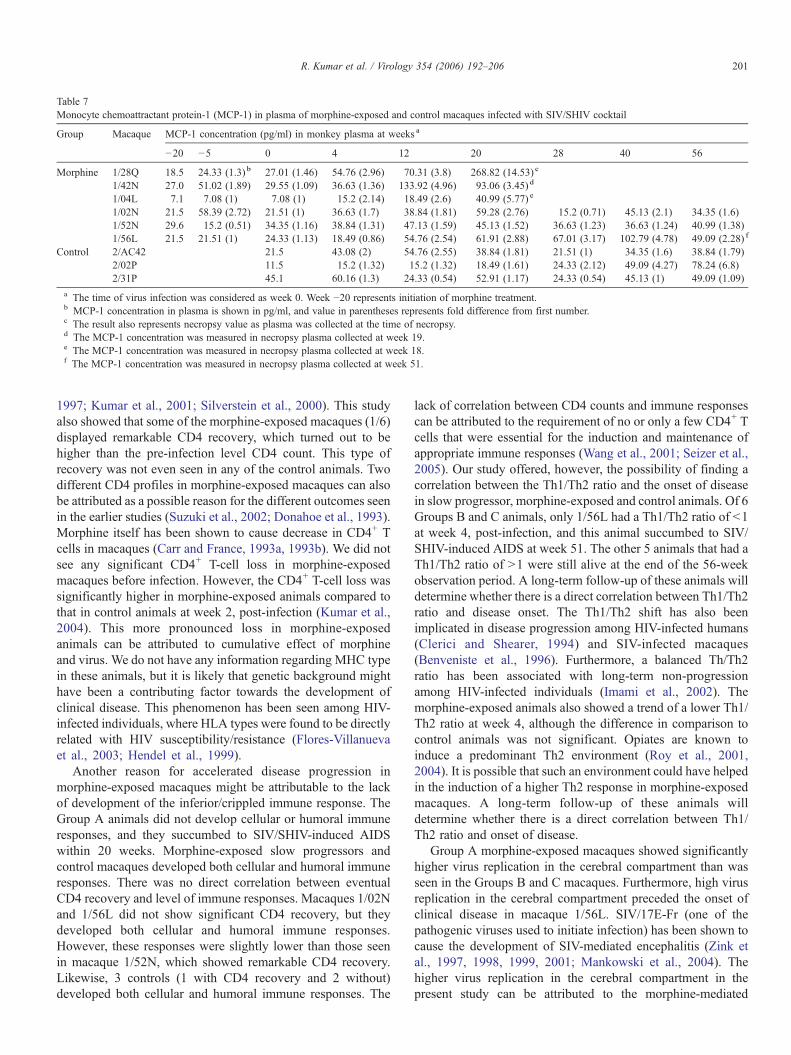
Table 7Monocyte chemoattractant protein-1 (MCP-1) in plasma of morphine-exposed and control macaques infected with SIV/SHIV cocktail
Group Macaque MCP-1 concentration (pg/ml) in monkey plasma at weeks a
−20 −5 0 4 12 20 28 40 56
Morphine 1/28Q 18.5 24.33 (1.3) b 27.01 (1.46) 54.76 (2.96) 70.31 (3.8) 268.82 (14.53) c
1/42N 27.0 51.02 (1.89) 29.55 (1.09) 36.63 (1.36) 133.92 (4.96) 93.06 (3.45) d
1/04L 7.1 7.08 (1) 7.08 (1) 15.2 (2.14) 18.49 (2.6) 40.99 (5.77) e
1/02N 21.5 58.39 (2.72) 21.51 (1) 36.63 (1.7) 38.84 (1.81) 59.28 (2.76) 15.2 (0.71) 45.13 (2.1) 34.35 (1.6)1/52N 29.6 15.2 (0.51) 34.35 (1.16) 38.84 (1.31) 47.13 (1.59) 45.13 (1.52) 36.63 (1.23) 36.63 (1.24) 40.99 (1.38)1/56L 21.5 21.51 (1) 24.33 (1.13) 18.49 (0.86) 54.76 (2.54) 61.91 (2.88) 67.01 (3.17) 102.79 (4.78) 49.09 (2.28) f
Control 2/AC42 21.5 43.08 (2) 54.76 (2.55) 38.84 (1.81) 21.51 (1) 34.35 (1.6) 38.84 (1.79)2/02P 11.5 15.2 (1.32) 15.2 (1.32) 18.49 (1.61) 24.33 (2.12) 49.09 (4.27) 78.24 (6.8)2/31P 45.1 60.16 (1.3) 24.33 (0.54) 52.91 (1.17) 24.33 (0.54) 45.13 (1) 49.09 (1.09)
a The time of virus infection was considered as week 0. Week −20 represents initiation of morphine treatment.b MCP-1 concentration in plasma is shown in pg/ml, and value in parentheses represents fold difference from first number.c The result also represents necropsy value as plasma was collected at the time of necropsy.d The MCP-1 concentration was measured in necropsy plasma collected at week 19.e The MCP-1 concentration was measured in necropsy plasma collected at week 18.f The MCP-1 concentration was measured in necropsy plasma collected at week 51.
201R. Kumar et al. / Virology 354 (2006) 192–206
1997; Kumar et al., 2001; Silverstein et al., 2000). This studyalso showed that some of the morphine-exposed macaques (1/6)displayed remarkable CD4 recovery, which turned out to behigher than the pre-infection level CD4 count. This type ofrecovery was not even seen in any of the control animals. Twodifferent CD4 profiles in morphine-exposed macaques can alsobe attributed as a possible reason for the different outcomes seenin the earlier studies (Suzuki et al., 2002; Donahoe et al., 1993).Morphine itself has been shown to cause decrease in CD4+ Tcells in macaques (Carr and France, 1993a, 1993b). We did notsee any significant CD4+ T-cell loss in morphine-exposedmacaques before infection. However, the CD4+ T-cell loss wassignificantly higher in morphine-exposed animals compared tothat in control animals at week 2, post-infection (Kumar et al.,2004). This more pronounced loss in morphine-exposedanimals can be attributed to cumulative effect of morphineand virus. We do not have any information regarding MHC typein these animals, but it is likely that genetic background mighthave been a contributing factor towards the development ofclinical disease. This phenomenon has been seen among HIV-infected individuals, where HLA types were found to be directlyrelated with HIV susceptibility/resistance (Flores-Villanuevaet al., 2003; Hendel et al., 1999).
Another reason for accelerated disease progression inmorphine-exposed macaques might be attributable to the lackof development of the inferior/crippled immune response. TheGroup A animals did not develop cellular or humoral immuneresponses, and they succumbed to SIV/SHIV-induced AIDSwithin 20 weeks. Morphine-exposed slow progressors andcontrol macaques developed both cellular and humoral immuneresponses. There was no direct correlation between eventualCD4 recovery and level of immune responses. Macaques 1/02Nand 1/56L did not show significant CD4 recovery, but theydeveloped both cellular and humoral immune responses.However, these responses were slightly lower than those seenin macaque 1/52N, which showed remarkable CD4 recovery.Likewise, 3 controls (1 with CD4 recovery and 2 without)developed both cellular and humoral immune responses. The
lack of correlation between CD4 counts and immune responsescan be attributed to the requirement of no or only a few CD4+ Tcells that were essential for the induction and maintenance ofappropriate immune responses (Wang et al., 2001; Seizer et al.,2005). Our study offered, however, the possibility of finding acorrelation between the Th1/Th2 ratio and the onset of diseasein slow progressor, morphine-exposed and control animals. Of 6Groups B and C animals, only 1/56L had a Th1/Th2 ratio of <1at week 4, post-infection, and this animal succumbed to SIV/SHIV-induced AIDS at week 51. The other 5 animals that had aTh1/Th2 ratio of >1 were still alive at the end of the 56-weekobservation period. A long-term follow-up of these animals willdetermine whether there is a direct correlation between Th1/Th2ratio and disease onset. The Th1/Th2 shift has also beenimplicated in disease progression among HIV-infected humans(Clerici and Shearer, 1994) and SIV-infected macaques(Benveniste et al., 1996). Furthermore, a balanced Th/Th2ratio has been associated with long-term non-progressionamong HIV-infected individuals (Imami et al., 2002). Themorphine-exposed animals also showed a trend of a lower Th1/Th2 ratio at week 4, although the difference in comparison tocontrol animals was not significant. Opiates are known toinduce a predominant Th2 environment (Roy et al., 2001,2004). It is possible that such an environment could have helpedin the induction of a higher Th2 response in morphine-exposedmacaques. A long-term follow-up of these animals willdetermine whether there is a direct correlation between Th1/Th2 ratio and onset of disease.
Group A morphine-exposed macaques showed significantlyhigher virus replication in the cerebral compartment than wasseen in the Groups B and C macaques. Furthermore, high virusreplication in the cerebral compartment preceded the onset ofclinical disease in macaque 1/56L. SIV/17E-Fr (one of thepathogenic viruses used to initiate infection) has been shown tocause the development of SIV-mediated encephalitis (Zink etal., 1997, 1998, 1999, 2001; Mankowski et al., 2004). Thehigher virus replication in the cerebral compartment in thepresent study can be attributed to the morphine-mediated

202 R. Kumar et al. / Virology 354 (2006) 192–206
increase in TNF-α production by microglia that has also beenpreviously implicated in increased viral replication in promo-nocyte line (Peterson et al., 1994, 1998). Furthermore, mor-phine has also been shown to potentiate apoptosis and therelease of various cytokines by astrocytes and neurons that mayeventually increase the incidence of HIV-associated dementia(HAD) (Gurwell et al., 2001; El Hage et al., 2005). This wasconfirmed in this study where most of the morphine-exposedmacaques developed neurological symptoms.
The cerebral compartment of morphine-exposed macaquesturned out to be a preferential target for SIV/17E-Fr andSHIVKU replication. The SIV/17E-Fr crossed the blood brainbarrier (BBB) within 2 weeks in both morphine-exposed andcontrol macaques. However, while SHIVKU crossed the BBBwithin 8 weeks in morphine-exposed macaques, it crossed theBBB within 28 weeks in control macaques. The systemicimmune response did not seem to play any role in themigration of the virus from the blood to the cerebral com-partment. Macaque 1/52N (Group B; slow progressor andmorphine-exposed) developed better antibody and cellularimmune responses than those seen in any of the controls, butthis did not cause a delay in the migration of SHIVKU fromthe systemic to the cerebral compartment. The exact reasonfor this preferential crossover in morphine-exposed macaquesis not known, but chronic morphine administration is knownto induce inflammatory responses by disrupting IL-1β mod-ulation of the hypothalamic–pituitary–adrenal axis (HPA)(House et al., 2001), and that might have played some rolein the early BBB crossing of SHIVKU in morphine-exposedmacaques.
Morphine is known to increase HIV replication in monocyte/macrophages (Guo et al., 2002; Li et al., 2003; Ho et al., 2003;Peterson et al., 1994). Furthermore, morphine is also known tomodulate monocyte migration (Singhal et al., 1996; Hatsukari etal., 2006) and MCP-1 is thought to be one of the factorsresponsible for monocyte migration (Maslin et al., 2005; Kim etal., 2006). MCP-1 has been implicated in HIV encephalitis andalso in SIV-associated CNS complications (Conant et al., 1998;Kelder et al., 1998; McManus et al., 2000; Sanders et al., 1998;Sasseville et al., 1996; Zink et al., 2001). In the present study,4 morphine-exposed monkeys developed clinical disease du-ring observation period. These animals showed 11- to 20-fold increase in monocyte number as well as 4.8- to 14.5-foldincrease in MCP-1 concentration in plasma, suggesting thatincreased MCP-1 might be responsible for infected monocytemigration to brain (as evident by high viral loads in CSFcompartment) and eventual neurological symptoms in 3 of 4macaques.
In summary, we have shown that chronic morphine exposurecauses higher virus replication in the systemic and cerebralcompartments, accelerated crossover of one of the pathogenicviruses into the cerebral compartment and speedy onset of SIV/SHIV-induced AIDS in Indian rhesus macaques. Developmentof the systemic immune response also coincided with at least atemporary suppression of virus replication in the brain.However, these immune responses did not affect the viralcrossover from the systemic to the cerebral compartment.
Materials and methods
Viruses
SHIVKU-1B was derived from non-pathogenic SHIV-4 bypassaging into pigtailed macaques (Joag et al., 1996).SHIV89.6P and SIV/17E-Fr were kindly provided by Profs. N.Letvin, Boston, MA and J. Clements, Baltimore, MD, res-pectively (Reimann et al., 1996a; Flaherty et al., 1997). Stockpreparations of these viruses were propagated in macaquePBMC. Infectivity of stocks of the three viruses was assayed inC8166 cell culture and had titers of 104 (SHIVKU), 1.4×10
4
(SHIV89.6P) and 4.3×104 TCID50/ml (SIV/17E-Fr).
Establishment of chronic morphine exposure and infection ofanimals
Nine 1.5- to 2.5-year-old male rhesus macaques (Macacamulatta) were obtained from the Caribbean Research PrimateCenter and maintained in the AAALAC-approved AnimalFacility at the University of Puerto Rico Medical School. Thebody weight of these animals ranged between 3 and 4.2 kg atthe commencement of the study. The animals were tested fortuberculosis, simian T-cell leukemia virus type-1 and simianretrovirus and found negative in all these tests. Theexperimental protocol was approved by the InstitutionalAnimal Care and Use Committee, and the research wasperformed in accordance with the Guide for the Care andUse of Laboratory Animals. These animals were divided intotwo groups (morphine-exposed and control) of 6 and 3,respectively. The chronic morphine exposure was establishedby injecting (intramuscularly) increasing doses of morphine(1 to 5 mg/kg of body weight over a 2-week period) every8 h (7:00 AM, 3:00 PM and 11:00 PM). Morphine wasadministered for an additional 18 weeks before virus infectionand also continued throughout the study period. Sixmorphine-exposed and three control animals were infectedwith SHIVKU-1B, SHIV89.6P and SIV/17E-Fr at week 20 afterthe initiation of the morphine injection (also known as week0, post-infection, and referred to as such hereafter). Theanimals were injected intravenously with 2 ml inoculumcontaining 104 50% tissue culture infectious doses (TCID50)of each virus.
Flow cytometry
Lymphocyte subset cell profiles were determined by stainingfor CD3, CD4 and CD8 cell surface markers using the wholeblood lysis technique (Wyand et al., 1999). Briefly, 10 μl of theantibody mix against CD3, CD4 and CD8 (Becton Dickinson)was added to 100 μl of whole blood and incubated for 1 h in thedark. Lysing solution (Becton Dickinson) was then added, andthe samples were incubated for another 10 min at roomtemperature. Stained cells were fixed with 0.5% paraformalde-hyde and analyzed in a flow cytometer (Becton DickinsonFACSCalibur). The absolute CD4+ CD8+ T-cell counts weredetermined by multiplying the percentage (obtained by flow

203R. Kumar et al. / Virology 354 (2006) 192–206
cytometric analysis) with the absolute lymphocyte numberobtained from a complete blood count.
Viral load measurements
Cell-associated viral loads in blood and brain weredetermined in an infectious cell assay (Kumar et al., 2001),and cell-free viral loads in plasma and cerebrospinal fluid (CSF)were determined using real-time RT-PCR (Amara et al., 2001).The viral RNA copy number in monkey samples wasdetermined by comparison with an external standard curveconsisting of an in vitro transcript representing bases 211–2101of the SIVMAC239 genome. This assay had a sensitivity of 80copies/ml of plasma or CSF.
Detection of challenge virus DNA in PBMC and CSF
For the purpose of virus identification in the brain, CSFwas co-cultured with CEMx174 cells for 1 week. ProviralDNAwas extracted from the PBMC and CSF-CEM co-culture,using the QIAamp DNA Blood Kit (QIAGEN, Inc., Valencia,California). Total cellular DNA (extracted from differentanimals) was screened for the presence of different proviralDNA by PCR using primers specific for the envelopesequences of SHIVKU, SHIV89.6P and SIV/17E-Fr. Thesequence of the primers for 1st and 2nd round amplifications,the location of the primers in the virus genome and theexpected size of the PCR products have been previouslydescribed (Kumar et al., 2002). Briefly, 0.5 μg of genomicDNA was used in the PCR mixture containing 4 mM MgCl2,200 μM each of the four deoxynucleoside triphosphates, 100pM each of the oligonucleotide primers and 2.5 U of Taqpolymerase (Perkin-Elmer Cetus, Norwalk, CN). The templatewas denatured at 95 °C for 3 min, and PCR amplificationperformed with an automated DNA thermal cycler (Perkin-Elmer Cetus) for 35 cycles of denaturation at 92 °C for 1 min;annealing at 55 °C for 1 min; and primer extension at 72 °C for3 min. The reaction was completed after 10 min of primerextension at 72 °C. Two microliters of the resultant product wasused in the 2nd round of PCR, using previously describedprimers (Kumar et al., 2002) and conditions (as described abovefor the 1st round.). Following the second round of amplifica-tion, a 5-μl aliquot was electrophoresed on a 1.5% agarose gel,and the bands were visualized by staining with ethidiumbromide. Three independent PCR reactions were performedseparately for each sample, and the sample was consideredpositive for a given virus with signal in any of the 3 independentPCR reactions.
Measurement of binding antibody titer against viral envelope
Plasma samples from both the morphine-exposed and thecontrol monkeys were analyzed for their reactivity to nativeviral glycoprotein in a concanavalin-A (Con-A) ELISA,developed by Robinson et al. (1990) and adapted to themacaque system (Cole et al., 1997; Kumar et al., 2000).Preparation of SHIVKU, SHIV89.6P, or SIV/17E-Fr, purified
by density gradient centrifugation and disrupted with 1%triton X-100, was used as source of viral glycoproteins.Immulon II microtiter plates (Dynatech Laboratories) wereincubated with 5 μg of Con-A (Sigma, St. Louis, MO) perwell in 100 μl of phosphate-buffered saline (PBS, pH 7.2) for90 min at 37 °C and then washed twice with Normal SalineTween (NST—0.85% NaCl and 0.1% Tween-20). Threehundred nanograms of viral proteins in 100 μl of PBS wasthen added to each well, and the plate was again incubated at37 °C for 90 min. The plate was again washed twice withNST and blocked with R-10 by adding 300 μl of R-10 intoeach well. After the blocking solution was removed, 100 μl ofserial two-fold diluted monkey plasma was added to eachwell in duplicate and the plate was again incubated at 37 °Cfor 90 min. The plate was washed again, thrice, with NST,and 100 μl of peroxidase-conjugated anti-monkey IgG(Sigma-diluted 1:10,000 in PBS containing 1% FBS) wasadded. The plate was again incubated at 37 °C followed by 4washings with NST. The reaction was developed by adding100 μl of citrate–phosphate buffer (pH 5.0) containing0.5 mg/ml of orthophenylenediamine (OPD) and 0.4 μl/mlof 30% H2O2. The reaction was stopped by adding 50 μl of1 N HCl, and optical density was read in a plate reader(Dynatech) at 490 nm wavelength. Endpoint titers weredetermined by the last dilution, whose OD was twice that ofnormal monkey plasma at 1:1000 dilution or an OD of 0.1,whichever value was greater.
Measurement of the neutralizing antibody titer against SHIVand SIV
The neutralizing antibody titer in plasma was determined asdescribed previously (Kumar et al., 2000). Briefly, two-foldserial dilutions of heat-inactivated plasma in R-10 wereprepared in quadruplicate in round-bottom 96-well plates, and16 TCID50 of the test virus (SHIVKU, SHIV89.6P or SIV/17E-Fr) was added to each well. Plates were incubated 1 h at 37 °C,and 104 C-8166 cells were added to each well. Each well wasscored individually for the virus-induced cytopathic effect aftera 7-day incubation at 37 °C. The endpoint titer was defined bythe ability of a plasma dilution to neutralize the virus in all 4wells.
ELISPOT assays
Cellular immune responses were monitored by measuringIFN-γ and the IL-4 secretion of morphine-exposed and controlmonkeys' PBMC, stimulated with a mixture of UV-irradiatedand heat-inactivated SHIVKU, SHIV89.6P or SIV/17E-Fr (UV-HI virus cocktail). The UV-HI virus cocktail was prepared bymixing an equal volume of SHIVKU, SHIV89.6P or SIV/17E-Frfollowed by UV irradiation for 1 h and heat inactivation at 56 °Cfor 3 h. The control stimulant was prepared by processing R-10(RPMI-1640 containing 10% fetal bovine serum) in the similarmanner. The ELISPOT assay was performed using IFN-γ andIL-4 ELISPOT kits that were procured from U-CyTechBiosciences, Utrecht, Netherlands. The assay was performed

204 R. Kumar et al. / Virology 354 (2006) 192–206
using the manufacturer's protocol except that the PBMCs wereincubated for 6 h at 37 °C in a humidified atmospherecontaining 5% CO2. One hundred thousand PBMCs werestimulated with 50 μl of UV-HI virus cocktail in duplicate. TheELISPOTs were counted by using an automated AID EliSPotreader system (Strassberg, Germany). The results are expressedas spot forming cells/106 PBMC which were obtained bysubtracting the number of spots in the control wells from thenumber of spots in the UV-HI virus cocktail-stimulated wells.The control numbers were obtained by incubating 105 of PBMCwith 50 μl of control stimulant. The Th1/Th2 ratio wascalculated by dividing the number of IFN-γ secreting cells bythe number of IL-4 secreting cells.
Determination of plasma MCP-1 using luminex technology
Monocyte chemoattractant protein-1 (MCP-1) was measuredin plasma as part of simultaneous detection of multiplecytokines and chemokines using luminex technology (Giave-doni, 2005). MCP-1 antibody pair was received as gift fromUpstate USA, Inc (Chicago, IL). The MCP-1 was measured atweeks −20, −5, 0, 4, 12, 20, 28, 40 and 56 in morphine-exposedmacaques and at weeks 0, 4, 12, 20, 28, 40 and 56 in controlmacaques. The results are presented as concentration in pg/mland also as fold difference compared to normal values. Morethan 2-fold difference in plasma MCP-1 was consideredsignificant.
Detection of MOR C77G SNP in morphine-exposed rhesusmacaques
Morphine-exposed and SIV/SHIV inoculated animals wereanalyzed for presence of C77G single nucleotide polymorph-ism (SNP) following a previously published protocol (Milleret al., 2004). Briefly, a 137 base fragment in mu-opioidreceptor was amplified from PBMC DNA. The PCR productwas purified using QiaquickTM PCR clean-up kit (Qiagen Inc.Valencia, CA, USA), and digested with Fnu4HI (New EnglandBiolabs, Beverly, MA, USA) for 1 h. The samples wereelectrophoresed on 3% agarose gels containing ethidiumbromide. The gels were visualized under UV transillumination(Fig. 4).
Acknowledgments
We thank Profs. Janice Clements, Norm Letvin, and EdwardStephens for providing SIV/17E-Fr, SHIV89.6P, and SHIVKU,respectively. We also thank Dr. Y. Yamamura for allowing us touse HIV core laboratory for use of Flowcytometer and Mr. BobRitchie for his critical reading and editing of the manuscript(supported by NCRR grant number RR003050). We appreciatehelpful discussion with Drs. Charles Sharp and Robert Donahoethat has immensely helped this project. The MCP-1 antibodypair was received as a gift from Upstate USA, Chicago, IL. Thiswork was supported by grants from the National Institute onDrug Abuse (DA015013) and National Institute on AlcoholAbuse and Alcoholism (AA015045).
References
Alcabes, P., Friedland, G., 1995. Injection drug use and human immunodefi-ciency virus infection. Clin. Infect. Dis. 20, 1467–1479.
Amara, R.R., Villinger, F., Altman, J.D., Lydy, S.L., O'Neil, S.P., Staprans, S.I.,Montefiori, D.C., Xu, Y., Herndon, J.G., Wyatt, L.S., Candido, M.A., Kozyr,N.L., Earl, P.L., Smith, J.M., Ma, H.L., Grimm, B.D., Hulsey, M.L., Miller,J., McClure, H.M., McNicholl, J.M., Moss, B., Robinson, H.L., 2001.Control of a mucosal challenge and prevention of AIDS by a multiproteinDNA/MVA vaccine. Science 292, 69–74.
Benveniste, O., Vaslin, B., Le Grand, R., Cheret, A., Matheux, F.,Theodoro, F., Cranage, M.P., Dormont, D., 1996. Comparativeinterleukin (IL-2)/interferon IFN-gamma and IL-4/IL-10 responsesduring acute infection of macaques inoculated with attenuated nef-truncated or pathogenic SICmac251 virus. Proc. Natl. Acad. Sci. U.S.A.93, 3658–3663.
Bond, C., LaForge, K.S., Tian, M., Melia, D., Zhang, S., Borg, L., Gong,J., Schluger, J., Strong, J.A., Leal, S.M., Tischfield, J.A., Kreek, M.J.,Yu, L., 1998. Single-nucleotide polymorphism in the human mu opioidreceptor gene alters beta-endorphin binding and activity: possibleimplications for opiate addiction. Proc. Natl. Acad. Sci. U.S.A. 95,9608–9613.
Brancato, V., Delvecchio, G., Simone, P., 1995. Survival and mortality in acohort of heroin addicts in 1985–1994. Minerva Med. 86, 97–99.
Carr, D.J., France, C.P., 1993a. Immune alterations in chronic morphine-treatedrhesus monkeys. Adv. Exp. Med. Biol. 335, 35–39.
Carr, D.J., France, C.P., 1993b. Immune alterations in morphine-treated rhesusmonkeys. J. Pharmacol. Exp. Ther. 267, 9–15.
CDC, 2002. Estimated numbers of diagnoses of HIV/AIDS, by year of diagnosisand selected characteristics of persons, 1999–2002. HIV/AIDS Surveill.Rep. 14, 10.
Chuang, L.F., Killam Jr., K.F., Chuang, R.Y., 1993a. Increased replication ofsimian immunodeficiency virus in CEM x174 cells by morphine sulfate.Biochem. Biophys. Res. Commun. 195, 1165–1173.
Chuang, R.Y., Blackbourn, D.J., Chuang, L.F., Liu, Y., Kilam, K.F., 1993b.Modulation of simian AIDS by opioids. Adv. Biosci. 86, 573–583.
Clerici, M., Shearer, G.M., 1994. The Th1–Th2 hypothesis of HIV infection:new insights. Immunol. Today 15, 575–581.
Cole, K.S., Rowles, J.L., Jagerski, B.A., Murphey-Corb, M., Unangst, T.,Clements, J.E., Robinson, J., Wyand, M.S., Desrosiers, R.C., Montelaro,R.C., 1997. Evolution of envelope-specific antibody responses in monkeysexperimentally infected or immunized with simian immunodeficiency virusand its association with the development of protective immunity. J. Virol.71, 5069–5079.
Comar, M., Simonelli, C., Zanussi, S., Paoli, P., Vaccher, E., Tirelli, U., Giacca,M., 1997. Dynamics of HIV-1 mRNA expression in patients with long-termnonprogressive HIV-1 infection. J. Clin. Invest. 100, 893–903.
Conant, K., Garzino-Demo, A., Nath, A., McArthur, J.C., Halliday, W., Power,C., Gallo, R.C., Major, E.O., 1998. Induction of monocyte chemoattractantprotein-1 in HIV-1 Tat-stimulated astrocytes and elevation in AIDSdementia. Proc. Natl. Acad. Sci. U.S.A. 95, 3117–3121.
Daniel, M.D., Letvin, N.L., King, N.W., Kannagi, M., Sehgal, P.K., Hunt, R.D.,Kanki, P.J., Essex, M., Desrosiers, R.C., 1985. Isolation of T-cell tropicHTLV-III-like retrovirus from macaques. Science 228, 1201–1204.
Donahoe, R.M., 2004. Multiple ways that drug abuse might influence AIDSprogression: clues from a monkey model. J. Neuroimmunol. 147, 28–32.
Donahoe, R.M., Byrd, L.D., McClure, H.M., Fultz, P., Brantley, M.,Marsteller, F., Ansari, A.A., Wenzel, D., Aceto, M., 1993. Consequencesof opiate-dependency in a monkey model of AIDS. Adv. Exp. Med. Biol.335, 21–28.
Eisenstein, L.K., MacFarland, A.S., Peng, X., Hilburger, M.E., Rahim, R.T.,Meissler Jr., L.J., Rogers, T.J., Wan, A.C., Adler, M.W., 2001. Effect ofopioids on oral Salmonella infection and immune function. Adv. Exp. Med.Biol. 493, 169–176.
El Hage, N., Gurwell, J.A., Singh, I.N., Knapp, P.E., Nath, A., Hauser, K.F.,2005. Synergistic increases in intracellular Ca2+, and the release of MCP-1,RANTES, and IL-6 by astrocytes treated with opiates and HIV-1 Tat. Glia50, 91–106.

205R. Kumar et al. / Virology 354 (2006) 192–206
Flaherty, M.T., Hauer, D.A., Mankowski, J.L., Zink, M.C., Clements, J.E., 1997.Molecular and biological characterization of a neurovirulent molecular cloneof simian immunodeficiency virus. J. Virol. 71, 5790–5798.
Flores-Villanueva, P.O., Hendel, H., Caillat-Zucman, S., Rappaport, J.,Burgos-Tiburcio, A., Bertin-Maghit, S., Ruiz-Morales, J.A., Teran, M.E.,Rodriguez-Tafur, J., Zagury, J.F., 2003. Associations of MHC ancestralhaplotypes with resistance/susceptibility to AIDS disease development.J. Immunol. 170, 1925–1929.
Fultz, P.N., McClure, H.M., Anderson, D.C., Swenson, R.B., Anand, R.,Srinivasan, A., 1986. Isolation of a T-lymphotropic retrovirus from naturallyinfected sooty mangabey monkeys (Cercocebus atys). Proc. Natl. Acad. Sci.U.S.A. 83, 5286–5290.
Giavedoni, L., 2005. Simultaneous detection of multiple cytokines andchemokines from nonhuman primates using luminex technology. J. Immunol.Methods 301 (1–2), 89–101 (Jun.).
Guo, C.J., Li, Y., Tian, S., Wang, X., Douglas, S.D., Ho, W.Z., 2002. Morphineenhances HIV infection of human blood mononuclear phagocytes throughmodulation of beta-chemokines and CCR5 receptor. J. Invest. Med. 50,435–442.
Gurwell, J.A., Nath, A., Sun, Q., Zhang, J., Martin, K.M., Chen, Y., Hauser,K.F., 2001. Synergistic neurotoxicity of opioids and human immunode-ficiency virus-1 Tat protein in striatal neurons in vitro. Neuroscience 102,555–563.
Hatsukari, I., Hitosugi, N., Dinda, A., Singhal, P.C., 2006. Morphine modulatesmonocyte–macrophage conversion phase. Cell. Immunol. (May 12).
Hendel, H., Caillat-Zucman, S., Lebuanec, H., Carrington, M., O'Brien, S.,Andrieu, J.M., Schachter, F., Zagury, D., Rappaport, J., Winkler, C., Nelson,G.W., Zagury, J.F., 1999. New class I and II HLA alleles strongly associatedwith opposite patterns of progression to AIDS. J. Immunol. 162, 6942–6946.
Hilburger, M.E., Adler, M.W., Truant, A.L., Meissler Jr., J.J., Satishchandran,V., Rogers, T.J., Eisenstein, T.K., 1997. Morphine induces sepsis in mice.J. Infect. Dis. 176, 183–188.
Ho, W.Z., Guo, C.J., Yuan, C.S., Douglas, S.D., Moss, J., 2003. Methylnaltrex-one antagonizes opioid-mediated enhancement of HIV infection of humanblood mononuclear phagocytes. J. Pharmacol. Exp. Ther. 307, 1158–1162.
House, S.D., Mao, X., Wu, G., Espinelli, D., Li, W.X., Chang, S.L., 2001.Chronic morphine potentiates the inflammatory response by disruptinginterleukin-1beta modulation of the hypothalamic–pituitary–adrenal axis.J. Neuroimmunol. 118, 277–285.
Igarashi, T., Endo, Y., Englund, G., Sadjadpour, R., Matano, T., Buckler, C.,Buckler-White, A., Plishka, R., Theodore, T., Shibata, R., Martin, M.,1999. Emergence of a highly pathogenic simian/human immunodeficiencyvirus in a rhesus macaque treated with anti-CD8 mAb during a primaryinfection with a nonpathogenic virus. Proc. Natl. Acad. Sci. U.S.A. 96,14049–14054.
Imami, N., Pires, A., Hardy, G., Wilson, J., Gazzard, B., Gotch, F., 2002. Abalanced type 1/type 2 response is associated with long-term nonprogressivehuman immunodeficiency virus type 1 infection. J. Virol. 76, 9011–9023.
Joag, S.V., Li, Z., Foresman, L., Stephens, E.B., Zhao, L.J., Adany, I., Pinson,D.M., McClure, H.M., Narayan, O., 1996. Chimeric simian/humanimmunodeficiency virus that causes progressive loss of CD4+ T cells andAIDS in pig-tailed macaques. J. Virol. 70, 3189–3197.
Karlsson, G.B., Halloran, M., Li, J., Park, I.W., Gomila, R., Reimann, K.A.,Axthelm, M.K., Iliff, S.A., Letvin, N.L., Sodroski, J., 1997. Characterizationof molecularly cloned simian–human immunodeficiency viruses causingrapid CD4+ lymphocyte depletion in rhesus monkeys. J. Virol. 71,4218–4225.
Kaslow, R.A., Blackwelder, W.C., Ostrow, D.G., Yerg, D., Palenicek, J.,Coulson, A.H., Valdiserri, R.O., 1989. No evidence for a role of alcohol orother psychoactive drugs in accelerating immunodeficiency in HIV-1-positive individuals. A report from the Multicenter AIDS Cohort Study.JAMA 261, 3424–3429.
Kelder, W., McArthur, J.C., Nance-Sproson, T., McClernon, D., Griffin, D.E.,1998. Beta-chemokines MCP-1 and RANTES are selectively increased incerebrospinal fluid of patients with human immunodeficiency virus-associated dementia. Ann. Neurol. 44, 831–835.
Kim, J.S., Kim, J.G., Moon, M.Y., Jeon, C.Y., Won, H.Y., Kim, H.J., Jeon, Y.J.,Seo, J.Y., Kim, J.I., Kim, J., Lee, J.Y., Kim, P.H., Park, J.B., 2006.
Transforming growth factor-{beta}1 regulates macrophage migration viaRhoA. Blood (online publication, May 16).
Kumar, A., Lifson, J.D., Silverstein, P.S., Jia, F., Sheffer, D., Li, Z., Narayan, O.,2000. Evaluation of immune responses induced by HIV-1 gp120 in rhesusmacaques: effect of vaccination on challenge with pathogenic strains ofhomologous and heterologous simian human immunodeficiency viruses.Virology 274, 149–164.
Kumar, A., Lifson, J.D., Li, Z., Jia, F., Mukherjee, S., Adany, I., Liu, Z., Piatak,M., Sheffer, D., McClure, H.M., Narayan, O., 2001. Sequential immuniza-tion of macaques with two differentially attenuated vaccines induced long-term virus-specific immune responses and conferred protection againstAIDS caused by heterologous simian human immunodeficiency virus(SHIV(89.6)P). Virology 279, 241–256.
Kumar, A., Mukherjee, S., Shen, J., Buch, S., Li, Z., Adany, I., Liu, Z., Zhuge,W., Piatak Jr., M., Lifson, J., McClure, H., Narayan, O., 2002. Immunizationof macaques with live simian human immunodeficiency virus (SHIV)vaccines conferred protection against AIDS induced by homologous andheterologous SHIVs and simian immunodeficiency virus. Virology 301,189–205.
Kumar, R., Torres, C., Yamamura, Y., Rodriguez, I., Martinez, M., Staprans, S.,Donahoe, R.M., Kraiselburd, E., Stephens, E.B., Kumar, A., 2004.Modulation by morphine of viral set point in rhesus macaques infectedwith simian immunodeficiency virus and simian–human immunodeficiencyvirus. J. Virol. 78, 11425–11428.
Li, Y., Merrill, J.D., Mooney, K., Song, L., Wang, X., Guo, C.J., Savani, R.C.,Metzger, D.S., Douglas, S.D., Ho, W.Z., 2003. Morphine enhances HIVinfection of neonatal macrophages. Pediatr. Res. 54, 282–288.
Luciw, P.A., Pratt-Lowe, E., Shaw, K.E., Levy, J.A., Cheng-Mayer, C., 1995.Persistent infection of rhesus macaques with T-cell-line-tropic andmacrophage-tropic clones of simian/human immunodeficiency viruses(SHIV). Proc. Natl. Acad. Sci. U.S.A. 92, 7490–7494.
MacFarlane, A.S., Peng, X., Meissler Jr., J.J., Rogers, T.J., Geller, E.B., Adler,M.W., Eisenstein, T.K., 2000. Morphine increases susceptibility to oralSalmonella typhimurium infection. J. Infect. Dis. 181, 1350–1358.
Mankowski, J.L., Queen, S.E., Clements, J.E., Zink, M.C., 2004. Cerebrospinalfluid markers that predict SIV CNS disease. J. Neuroimmunol. 157, 66–70.
Margolick, J.B., Munoz, A., Vlahov, D., Solomon, L., Astemborski, J., Cohn,S., Nelson, K.E., 1992. Changes in T-lymphocyte subsets in intravenousdrug users with HIV-1 infection. JAMA 267, 1631–1636.
Maslin, C.L., Kedzierska, K., Webster, N.L., Muller, W.A., Crowe, S.M., 2005.Transendothelial migration of monocytes: the underlying molecularmechanisms and consequences of HIV-1 infection. Curr. HIV Res. 3,303–317.
McManus, C.M., Weidenheim, K., Woodman, S.E., Nunez, J., Hesselgesser, J.,Nath, A., Berman, J.W., 2000. Chemokine and chemokine-receptorexpression in human glial elements: induction by the HIV protein, Tat,and chemokine autoregulation. Am. J. Pathol. 156, 1441–1453.
Metzger, D.S., Woody, G.E., McLellan, A.T., O'Brien, C.P., Druley, P.,Navaline, H., DePhilippis, D., Stolley, P., Abrutyn, E., 1993. Humanimmunodeficiency virus seroconversion among intravenous drug users in-and out-of-treatment: an 18-month prospective follow-up. J. AcquiredImmune Defic. Syndr. 6, 1049–1056.
Miller, G.M., De La Garza, M., Novak, R.D., Madras, M.A., 2001. Singlenucleotide polymorphisms distinguish multiple dopamine transporter allelesin primates: implications for association with attention deficit hyperactivitydisorder and other neuropsychiatric disorders. Mol. Psychiatry 6 (1), 50–58(Jan.).
Miller, G.M., Bendor, J., Tiefenbacher, S., Yang, H., Novak, M.A., Madras,B.K., 2004. A mu-opioid receptor single nucleotide polymorphism in rhesusmonkey: association with stress response and aggression. Mol. Psychiatry 9,99–108.
Montella, F., Di Sora, F., Perucci, C.A., Abeni, D.D., Recchia, O., 1992.T-lymphocyte subsets in intravenous drug users with HIV-1 infection.JAMA 268, 2516–2517.
Munoz, A., Vlahov, D., Solomon, L., Margolick, J.B., Bareta, J.C., Cohn, S.,Astemborski, J., Nelson, K.E., 1992. Prognostic indicators for developmentof AIDS among intravenous drug users. J. Acquired Immune Defic. Syndr.5, 694–700.

206 R. Kumar et al. / Virology 354 (2006) 192–206
Nathanson, N., Hirsch, V.M., Mathieson, B.J., 1999. The role of nonhumanprimates in the development of an AIDS vaccine. AIDS 13 (Suppl. A),S113–S120.
Peterson, P.K., Sharp, B.M., Gekker, G., Portoghese, P.S., Sannerud, K., BalfourJr., H.H., 1990. Morphine promotes the growth of HIV-1 in humanperipheral blood mononuclear cell cocultures. AIDS 4, 869–873.
Peterson, P.K., Gekker, G., Chao, C.C., Schut, R., Molitor, T.W., Balfour Jr.,H.H., 1991. Cocaine potentiates HIV-1 replication in human peripheralblood mononuclear cell cocultures. Involvement of transforming growthfactor-beta. J. Immunol. 146, 81–84.
Peterson, P.K., Gekker, G., Chao, C.C., Schut, R., Verhoef, J., Edelman, C.K.,Erice, A., Balfour Jr., H.H., 1992. Cocaine amplifies HIV-1 replication incytomegalovirus-stimulated peripheral blood mononuclear cell cocultures.J. Immunol. 149, 676–680.
Peterson, P.K., Gekker, G., Schut, R., Hu, S., Balfour Jr., H.H., Chao, C.C.,1993. Enhancement of HIV-1 replication by opiates and cocaine: thecytokine connection. Adv. Exp. Med. Biol. 335, 181–188.
Peterson, P.K., Gekker, G., Hu, S., Anderson, W.R., Kravitz, F., Portoghese,P.S., Balfour Jr., H.H., Chao, C.C., 1994. Morphine amplifies HIV-1expression in chronically infected promonocytes cocultured with humanbrain cells. J. Neuroimmunol. 50, 167–175.
Peterson, P.K., Molitor, T.W., Chao, C.C., 1998. The opioid–cytokineconnection. J. Neuroimmunol. 83, 63–69.
Reimann, K.A., Li, J.T., Veazey, R., Halloran, M., Park, I.W., Karlsson, G.B.,Sodroski, J., Letvin, N.L., 1996a. A chimeric simian/human immunodefi-ciency virus expressing a primary patient human immunodeficiency virustype 1 isolate env causes an AIDS-like disease after in vivo passage in rhesusmonkeys. J. Virol. 70, 6922–6928.
Reimann, K.A., Li, J.T., Voss, G., Lekutis, C., Tenner-Racz, K., Racz, P., Lin,W., Montefiori, D.C., Lee-Parritz, D.E., Lu, Y., Collman, R.G., Sodroski, J.,Letvin, N.L., 1996b. An env gene derived from a primary humanimmunodeficiency virus type 1 isolate confers high in vivo replicativecapacity to a chimeric simian/human immunodeficiency virus in rhesusmonkeys. J. Virol. 70, 3198–3206.
Reimann, K.A., Watson, A., Dailey, P.J., Lin, W., Lord, C.I., Steenbeke, T.D.,Parker, R.A., Axthelm, M.K., Karlsson, G.B., 1999. Viral burden anddisease progression in rhesus monkeys infected with chimeric simian–human immunodeficiency viruses. Virology 256, 15–21.
Robinson, J.E., Holton, D., Liu, J., McMurdo, H., Murciano, A., Gohd, R.,1990. A novel enzyme-linked immunosorbent assay (ELISA) for thedetection of antibodies to HIV-1 envelope glycoproteins based onimmobilization of viral glycoproteins in microtiter wells coated withconcanavalin A. J. Immunol. Methods 132, 63–71.
Ronald, P.J., Robertson, J.R., Elton, R.A., 1994. Continued drug use and othercofactors for progression to AIDS among injecting drug users. AIDS 8,339–343.
Roth, M.D., Tashkin, D.P., Choi, R., Jamieson, B.D., Zack, J.A., Baldwin,G.C., 2002. Cocaine enhances human immunodeficiency virusreplication in a model of severe combined immunodeficient miceimplanted with human peripheral blood leukocytes. J. Infect. Dis. 185,701–705.
Roy, S., Balasubramanian, S., Sumandeep, S., Charboneau, R., Wang, J.,Melnyk, D., Beilman, G.J., Vatassery, R., Barke, R.A., 2001. Morphinedirects T cells toward T(H2) differentiation. Surgery 130, 304–309.
Roy, S., Wang, J., Gupta, S., Charboneau, R., Loh, H.H., Barke, R.A., 2004.Chronic morphine treatment differentiates T helper cells to Th2 effector cellsby modulating transcription factors GATA 3 and T-bet. J. Neuroimmunol.147, 78–81.
Sanders, V.J., Pittman, C.A., White, M.G., Wang, G., Wiley, C.A., Achim, C.L.,1998. Chemokines and receptors in HIVencephalitis. AIDS 12, 1021–1026.
Sasseville, V.G., Du, Z., Chalifoux, L.V., Pauley, D.R., Young, H.L., Sehgal,P.K., Desrosiers, R.C., Lackner, A.A., 1996. Induction of lymphocyteproliferation and severe gastrointestinal disease in macaques by a nef genevariant SIVmac239. Am. J. Pathol. 149, 163–176.
Seizer, P., Riedl, P., Reimann, J., Schirmbeck, R., 2005. Different sources of“help” facilitate the antibody response to hepatitis D virus delta antigen.J. Mol. Med. 83, 225–234.
Selwyn, P.A., Alcabes, P., Hartel, D., Buono, D., Schoenbaum, E.E., Klein, R.S.,Davenny, K., Friedland, G.H., 1992. Clinical manifestations and predictorsof disease progression in drug users with human immunodeficiency virusinfection. N. Engl. J. Med. 327, 1697–1703.
Sepkowitz, K.A., Raffalli, J., Riley, L., Kiehn, T.E., Armstrong, D., 1995.Tuberculosis in the AIDS era. Clin. Microbiol. Rev. 8, 180–199.
Shattock, R.J., Friedland, J.S., Griffin, G.E., 1994. Phagocytosis of Mycobac-terium tuberculosismodulates human immunodeficiency virus replication inhuman monocytic cells. J. Gen. Virol. 75 (Pt4), 849–856.
Silverstein, P.S., Mackay, G.A., Mukherjee, S., Li, Z., Piatak Jr., M., Lifson, J.D., Narayan, O., Kumar, A., 2000. Pathogenic simian/human immunode-ficiency virus SHIV(KU) inoculated into immunized macaques causedinfection, but virus burdens progressively declined with time. J. Virol. 74,10489–10497.
Singhal, P.C., Shan, Z., Garg, P., Sharma, K., Sharma, P., Gibbons, N., 1996.Morphine modulates migration of monocytes. Nephron. 73, 526–531.
Stipp, H.L., Kumar, A., Narayan, O., 2000. Characterization of immuneescape viruses from a macaque immunized with live-virus vaccine andchallenged with pathogenic SHIVKU-1. AIDS Res. Hum. Retrovir. 16,1573–1580.
Suzuki, S., Chuang, A.J., Chuang, L.F., Doi, R.H., Chuang, R.Y., 2002.Morphine promotes simian acquired immunodeficiency syndrome virusreplication in monkey peripheral mononuclear cells: induction of CCchemokine receptor 5 expression for virus entry. J. Infect. Dis. 185,1826–1829.
Tashkin, D.P., 2004. Evidence implicating cocaine as a possible risk factor forHIV infection. J. Neuroimmunol. 147, 26–27.
Wand, G.S., McCaul, M., Yang, X., Reynolds, J., Gotjen, D., Lee, S., Ali, A.,2002. The mu-opioid receptor gene polymorphism (A118G) alters HPA axisactivation induced by opioid receptor blockade. Neuropsychopharmacology26 (1), 106–114 (Jan.).
Wang, B., Norbury, C.C., Greenwood, R., Bennink, J.R., Yewdell, J.W.,Frelinger, J.A., 2001. Multiple paths for activation of naive CD8+ T cells:CD4-independent help. J. Immunol. 167, 1283–1289.
Wyand, M.S., Manson, K., Montefiori, D.C., Lifson, J.D., Johnson, R.P.,Desrosiers, R.C., 1999. Protection by live, attenuated simian immunodefi-ciency virus against heterologous challenge. J. Virol. 73, 8356–8363.
Zink, M.C., Amedee, A.M., Mankowski, J.L., Craig, L., Didier, P., Carter, D.L.,Munoz, A., Murphey-Corb, M., Clements, J.E., 1997. Pathogenesis of SIVencephalitis. Selection and replication of neurovirulent SIV. Am. J. Pathol.151, 793–803.
Zink, M.C., Spelman, J.P., Robinson, R.B., Clements, J.E., 1998. SIV infectionof macaques-modeling the progression to AIDS dementia. J. NeuroVirol. 4,249–259.
Zink, M.C., Suryanarayana, K., Mankowski, J.L., Shen, A., Piatak Jr., M.,Spelman, J.P., Carter, D.L., Adams, R.J., Lifson, J.D., Clements, J.E.,1999. High viral load in the cerebrospinal fluid and brain correlates withseverity of simian immunodeficiency virus encephalitis. J. Virol. 73,10480–10488.
Zink, M.C., Coleman, G.D., Mankowski, J.L., Adams, R.J., Tarwater, P.M.,Fox, K., Clements, J.E., 2001. Increased macrophage chemoattractantprotein-1 in cerebrospinal fluid precedes and predicts simian immunodefi-ciency virus encephalitis. J. Infect. Dis. 184, 1015–1021.
Related Documents











