Heterologous Prime–Boost Strategy to Immunize Very Young Infants against Measles: Pre-clinical Studies in Rhesus Macaques MF Pasetti 1,2 , A Resendiz-Albor 1 , K Ramirez 1 , R Stout 3 , M Papania 4 , RJ Adams 5 , FP Polack 5 , BJ Ward 6 , D Burt 7 , S Chabot 6 , J Ulmer 8 , EM Barry 1,9 and MM Levine 1,2,9 Infants in developing countries are at high risk of developing severe clinical measles if they become infected during the ‘‘window of vulnerability’’ (age 4–9 months), when declining maternal antibodies do not protect against wild virus, yet impede successful immunization by attenuated measles vaccine. We developed two Sindbis replicon-based DNA vaccines expressing measles virus hemagglutinin and fusion protein with the goal of priming young infants to respond safely and effectively to subsequent boosting with attenuated measles vaccine. Intradermal prime with DNA vaccines by needle-free injection followed by aerosol or parenteral boost with licensed measles vaccine was well tolerated by juvenile and young infant rhesus macaques, and protected against clinical measles and viremia on wild-type virus challenge. A proteosome-measles vaccine administered alone (three doses) or as a boost following DNA vaccine priming was also safe and protective. These promising results pave the way for clinical trials to assess this prime–boost strategy. Measles continues to cause unacceptable morbidity and mortality in infants and young children in the world’s least developed countries. 1 A limitation of the current licensed and generally efficacious attenuated measles virus (MV) vaccines is that a large proportion of measles deaths in developing countries occur among young infants during the ‘‘window of vulnerability,’’ between 4 and 9 months of age. 2 During this period, the falling titers of maternally derived measles antibodies no longer protect against wild-type MV. These low antibody titers, in conjunction with the infant’s immature immune system, nonetheless hamper successful immunization using the current licensed measles vaccines. Consequently, severe clinical measles with high case fatality rates (B10 to 25%) can occur when such young infants are exposed to wild-type virus. 1,3 We developed two Sindbis virus-based DNA vaccines encoding the MV hemagglutinin (H), 4 and MV H and fusion (F) glycoproteins. 5 Sindbis replicons represent a new generation of improved DNA vaccine in which cDNAs driven by eukaryotic promoters express self-replicating double-stranded RNAs that lead to high-level antigen expression. 6,7 The system also contributes immunostimula- tory signals including gene sequences from Sindbis virus with tropism for dendritic cells 8 and double-stranded RNA that stimulates toll-like receptors on antigen-presenting cells and secretion of proinflammatory cytokines. 9 The replicons also induce caspase-dependent apoptosis of transfected cells, further enhancing immune responses by antigen cross- presentation. 10 Thus, Sindbis-based DNA vaccines offer promise for engaging the immature immune system of young infants, even in the face of placentally transferred measles antibodies. 11 Earlier generations of measles DNA vaccines have conferred limited protection in rhesus macaques, 12,13 includ- ing newborns that were immunized with a conventional DNA vaccine in the presence of measles immunoglobulins. 14 Our strategy is to prime the young infant immune system by means of a measles DNA vaccine, so that it can respond safely and effectively to a subsequent dose of current attenuated measles vaccine administered at a much earlier age ARTICLES nature publishing group 1 Center for Vaccine Development, University of Maryland School of Medicine, Baltimore, Maryland, USA; 2 Department of Pediatrics, University of Maryland School of Medicine, Baltimore, Maryland, USA; 3 Bioject, Portland, Oregon, USA; 4 Centers for Disease Control and Prevention, Atlanta, Georgia, USA; 5 Bloomberg School of Hygiene and Public Health, Johns Hopkins University, Baltimore, Maryland, USA; 6 Department of Medicine, Tropical Diseases Center, McGill University, Montreal, Quebec, Canada; 7 GlaxoSmithKline Biologicals North America, Laval, Quebec, Canada; 8 Novartis Vaccines and Diagnostics, Emeryville, California, USA; 9 Department of Medicine, University of Maryland School of Medicine, Baltimore, Maryland, USA. Correspondence: MF Pasetti ([email protected]) Received 19 July 2007; accepted 10 September 2007; published online 31 October 2007. doi:10.1038/sj.clpt.6100420 672 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Heterologous Prime–Boost Strategy to ImmunizeVery Young Infants against Measles: Pre-clinicalStudies in Rhesus MacaquesMF Pasetti1,2, A Resendiz-Albor1, K Ramirez1, R Stout3, M Papania4, RJ Adams5, FP Polack5, BJ Ward6,
D Burt7, S Chabot6, J Ulmer8, EM Barry1,9 and MM Levine1,2,9
Infants in developing countries are at high risk of developing severe clinical measles if they become infected during the
‘‘window of vulnerability’’ (age 4–9 months), when declining maternal antibodies do not protect against wild virus, yet
impede successful immunization by attenuated measles vaccine. We developed two Sindbis replicon-based DNA
vaccines expressing measles virus hemagglutinin and fusion protein with the goal of priming young infants to respond
safely and effectively to subsequent boosting with attenuated measles vaccine. Intradermal prime with DNA vaccines by
needle-free injection followed by aerosol or parenteral boost with licensed measles vaccine was well tolerated by
juvenile and young infant rhesus macaques, and protected against clinical measles and viremia on wild-type virus
challenge. A proteosome-measles vaccine administered alone (three doses) or as a boost following DNA vaccine priming
was also safe and protective. These promising results pave the way for clinical trials to assess this prime–boost strategy.
Measles continues to cause unacceptable morbidity andmortality in infants and young children in the world’s leastdeveloped countries.1 A limitation of the current licensed andgenerally efficacious attenuated measles virus (MV) vaccinesis that a large proportion of measles deaths in developingcountries occur among young infants during the ‘‘window ofvulnerability,’’ between 4 and 9 months of age.2 During thisperiod, the falling titers of maternally derived measlesantibodies no longer protect against wild-type MV. Theselow antibody titers, in conjunction with the infant’simmature immune system, nonetheless hamper successfulimmunization using the current licensed measles vaccines.Consequently, severe clinical measles with high case fatalityrates (B10 to 25%) can occur when such young infants areexposed to wild-type virus.1,3
We developed two Sindbis virus-based DNA vaccinesencoding the MV hemagglutinin (H),4 and MV H andfusion (F) glycoproteins.5 Sindbis replicons represent a newgeneration of improved DNA vaccine in which cDNAsdriven by eukaryotic promoters express self-replicating
double-stranded RNAs that lead to high-level antigenexpression.6,7 The system also contributes immunostimula-tory signals including gene sequences from Sindbis virus withtropism for dendritic cells8 and double-stranded RNA thatstimulates toll-like receptors on antigen-presenting cells andsecretion of proinflammatory cytokines.9 The replicons alsoinduce caspase-dependent apoptosis of transfected cells,further enhancing immune responses by antigen cross-presentation.10 Thus, Sindbis-based DNA vaccines offerpromise for engaging the immature immune system ofyoung infants, even in the face of placentally transferredmeasles antibodies.11
Earlier generations of measles DNA vaccines haveconferred limited protection in rhesus macaques,12,13 includ-ing newborns that were immunized with a conventional DNAvaccine in the presence of measles immunoglobulins.14 Ourstrategy is to prime the young infant immune system bymeans of a measles DNA vaccine, so that it can respond safelyand effectively to a subsequent dose of current attenuatedmeasles vaccine administered at a much earlier age
ARTICLES nature publishing group
1Center for Vaccine Development, University of Maryland School of Medicine, Baltimore, Maryland, USA; 2Department of Pediatrics, University of Maryland Schoolof Medicine, Baltimore, Maryland, USA; 3Bioject, Portland, Oregon, USA; 4Centers for Disease Control and Prevention, Atlanta, Georgia, USA; 5Bloomberg School ofHygiene and Public Health, Johns Hopkins University, Baltimore, Maryland, USA; 6Department of Medicine, Tropical Diseases Center, McGill University, Montreal,Quebec, Canada; 7GlaxoSmithKline Biologicals North America, Laval, Quebec, Canada; 8Novartis Vaccines and Diagnostics, Emeryville, California, USA; 9Departmentof Medicine, University of Maryland School of Medicine, Baltimore, Maryland, USA. Correspondence: MF Pasetti ([email protected])
Received 19 July 2007; accepted 10 September 2007; published online 31 October 2007. doi:10.1038/sj.clpt.6100420
672 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
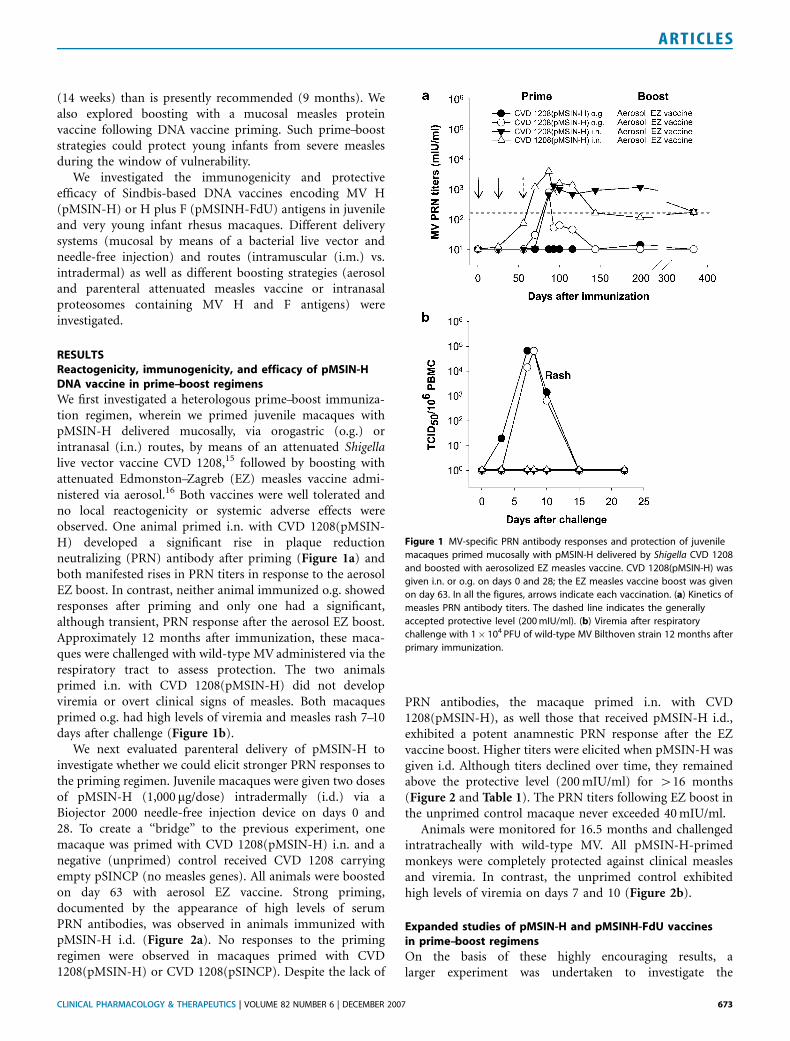
(14 weeks) than is presently recommended (9 months). Wealso explored boosting with a mucosal measles proteinvaccine following DNA vaccine priming. Such prime–booststrategies could protect young infants from severe measlesduring the window of vulnerability.
We investigated the immunogenicity and protectiveefficacy of Sindbis-based DNA vaccines encoding MV H(pMSIN-H) or H plus F (pMSINH-FdU) antigens in juvenileand very young infant rhesus macaques. Different deliverysystems (mucosal by means of a bacterial live vector andneedle-free injection) and routes (intramuscular (i.m.) vs.intradermal) as well as different boosting strategies (aerosoland parenteral attenuated measles vaccine or intranasalproteosomes containing MV H and F antigens) wereinvestigated.
RESULTSReactogenicity, immunogenicity, and efficacy of pMSIN-HDNA vaccine in prime–boost regimens
We first investigated a heterologous prime–boost immuniza-tion regimen, wherein we primed juvenile macaques withpMSIN-H delivered mucosally, via orogastric (o.g.) orintranasal (i.n.) routes, by means of an attenuated Shigellalive vector vaccine CVD 1208,15 followed by boosting withattenuated Edmonston–Zagreb (EZ) measles vaccine admi-nistered via aerosol.16 Both vaccines were well tolerated andno local reactogenicity or systemic adverse effects wereobserved. One animal primed i.n. with CVD 1208(pMSIN-H) developed a significant rise in plaque reductionneutralizing (PRN) antibody after priming (Figure 1a) andboth manifested rises in PRN titers in response to the aerosolEZ boost. In contrast, neither animal immunized o.g. showedresponses after priming and only one had a significant,although transient, PRN response after the aerosol EZ boost.Approximately 12 months after immunization, these maca-ques were challenged with wild-type MV administered via therespiratory tract to assess protection. The two animalsprimed i.n. with CVD 1208(pMSIN-H) did not developviremia or overt clinical signs of measles. Both macaquesprimed o.g. had high levels of viremia and measles rash 7–10days after challenge (Figure 1b).
We next evaluated parenteral delivery of pMSIN-H toinvestigate whether we could elicit stronger PRN responses tothe priming regimen. Juvenile macaques were given two dosesof pMSIN-H (1,000 mg/dose) intradermally (i.d.) via aBiojector 2000 needle-free injection device on days 0 and28. To create a ‘‘bridge’’ to the previous experiment, onemacaque was primed with CVD 1208(pMSIN-H) i.n. and anegative (unprimed) control received CVD 1208 carryingempty pSINCP (no measles genes). All animals were boostedon day 63 with aerosol EZ vaccine. Strong priming,documented by the appearance of high levels of serumPRN antibodies, was observed in animals immunized withpMSIN-H i.d. (Figure 2a). No responses to the primingregimen were observed in macaques primed with CVD1208(pMSIN-H) or CVD 1208(pSINCP). Despite the lack of
PRN antibodies, the macaque primed i.n. with CVD1208(pMSIN-H), as well those that received pMSIN-H i.d.,exhibited a potent anamnestic PRN response after the EZvaccine boost. Higher titers were elicited when pMSIN-H wasgiven i.d. Although titers declined over time, they remainedabove the protective level (200 mIU/ml) for 416 months(Figure 2 and Table 1). The PRN titers following EZ boost inthe unprimed control macaque never exceeded 40 mIU/ml.
Animals were monitored for 16.5 months and challengedintratracheally with wild-type MV. All pMSIN-H-primedmonkeys were completely protected against clinical measlesand viremia. In contrast, the unprimed control exhibitedhigh levels of viremia on days 7 and 10 (Figure 2b).
Expanded studies of pMSIN-H and pMSINH-FdU vaccinesin prime–boost regimens
On the basis of these highly encouraging results, alarger experiment was undertaken to investigate the
Figure 1 MV-specific PRN antibody responses and protection of juvenile
macaques primed mucosally with pMSIN-H delivered by Shigella CVD 1208
and boosted with aerosolized EZ measles vaccine. CVD 1208(pMSIN-H) was
given i.n. or o.g. on days 0 and 28; the EZ measles vaccine boost was given
on day 63. In all the figures, arrows indicate each vaccination. (a) Kinetics of
measles PRN antibody titers. The dashed line indicates the generally
accepted protective level (200 mIU/ml). (b) Viremia after respiratory
challenge with 1� 104 PFU of wild-type MV Bilthoven strain 12 months after
primary immunization.
CLINICAL PHARMACOLOGY & THERAPEUTICS | VOLUME 82 NUMBER 6 | DECEMBER 2007 673
ARTICLES
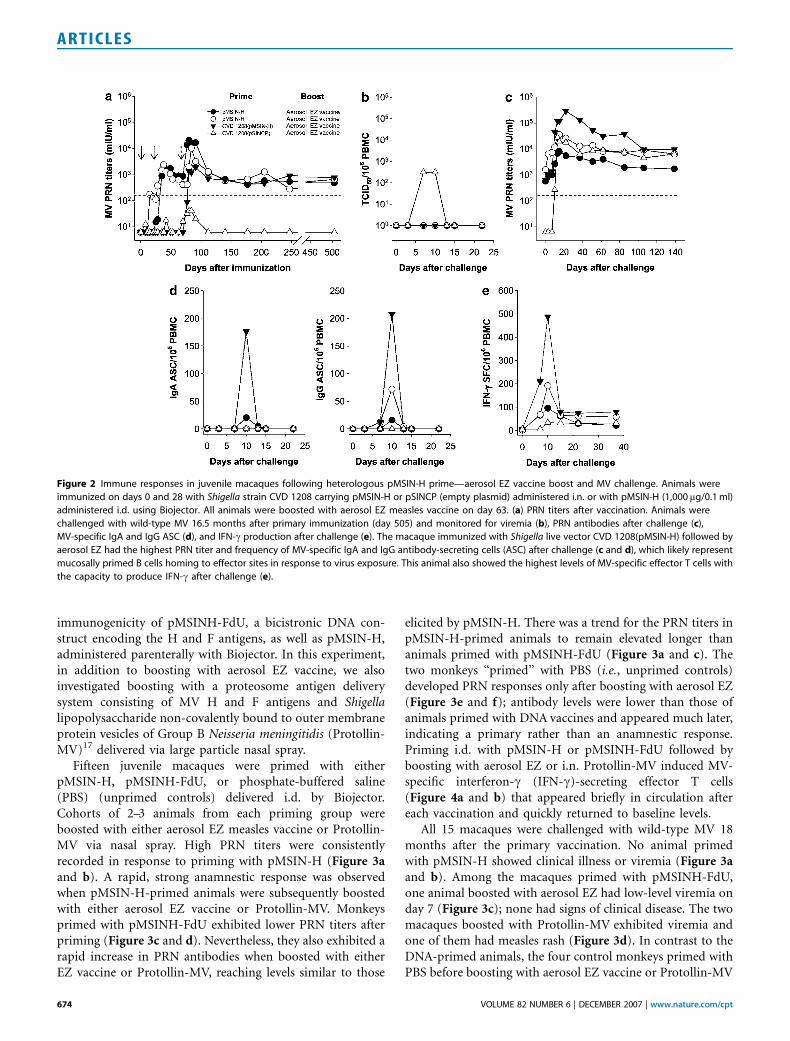
immunogenicity of pMSINH-FdU, a bicistronic DNA con-struct encoding the H and F antigens, as well as pMSIN-H,administered parenterally with Biojector. In this experiment,in addition to boosting with aerosol EZ vaccine, we alsoinvestigated boosting with a proteosome antigen deliverysystem consisting of MV H and F antigens and Shigellalipopolysaccharide non-covalently bound to outer membraneprotein vesicles of Group B Neisseria meningitidis (Protollin-MV)17 delivered via large particle nasal spray.
Fifteen juvenile macaques were primed with eitherpMSIN-H, pMSINH-FdU, or phosphate-buffered saline(PBS) (unprimed controls) delivered i.d. by Biojector.Cohorts of 2–3 animals from each priming group wereboosted with either aerosol EZ measles vaccine or Protollin-MV via nasal spray. High PRN titers were consistentlyrecorded in response to priming with pMSIN-H (Figure 3aand b). A rapid, strong anamnestic response was observedwhen pMSIN-H-primed animals were subsequently boostedwith either aerosol EZ vaccine or Protollin-MV. Monkeysprimed with pMSINH-FdU exhibited lower PRN titers afterpriming (Figure 3c and d). Nevertheless, they also exhibited arapid increase in PRN antibodies when boosted with eitherEZ vaccine or Protollin-MV, reaching levels similar to those
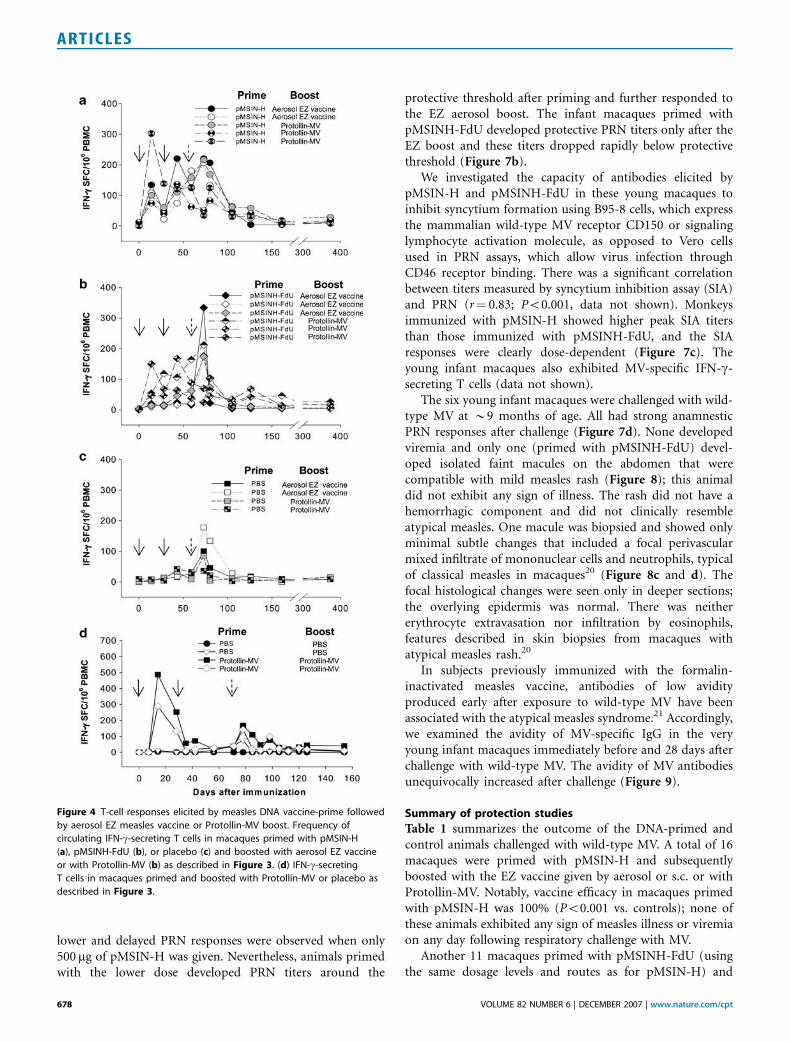
elicited by pMSIN-H. There was a trend for the PRN titers inpMSIN-H-primed animals to remain elevated longer thananimals primed with pMSINH-FdU (Figure 3a and c). Thetwo monkeys ‘‘primed’’ with PBS (i.e., unprimed controls)developed PRN responses only after boosting with aerosol EZ(Figure 3e and f); antibody levels were lower than those ofanimals primed with DNA vaccines and appeared much later,indicating a primary rather than an anamnestic response.Priming i.d. with pMSIN-H or pMSINH-FdU followed byboosting with aerosol EZ or i.n. Protollin-MV induced MV-specific interferon-g (IFN-g)-secreting effector T cells(Figure 4a and b) that appeared briefly in circulation aftereach vaccination and quickly returned to baseline levels.
All 15 macaques were challenged with wild-type MV 18months after the primary vaccination. No animal primedwith pMSIN-H showed clinical illness or viremia (Figure 3aand b). Among the macaques primed with pMSINH-FdU,one animal boosted with aerosol EZ had low-level viremia onday 7 (Figure 3c); none had signs of clinical disease. The twomacaques boosted with Protollin-MV exhibited viremia andone of them had measles rash (Figure 3d). In contrast to theDNA-primed animals, the four control monkeys primed withPBS before boosting with aerosol EZ vaccine or Protollin-MV
Figure 2 Immune responses in juvenile macaques following heterologous pMSIN-H prime—aerosol EZ vaccine boost and MV challenge. Animals were
immunized on days 0 and 28 with Shigella strain CVD 1208 carrying pMSIN-H or pSINCP (empty plasmid) administered i.n. or with pMSIN-H (1,000 mg/0.1 ml)
administered i.d. using Biojector. All animals were boosted with aerosol EZ measles vaccine on day 63. (a) PRN titers after vaccination. Animals were
challenged with wild-type MV 16.5 months after primary immunization (day 505) and monitored for viremia (b), PRN antibodies after challenge (c),
MV-specific IgA and IgG ASC (d), and IFN-g production after challenge (e). The macaque immunized with Shigella live vector CVD 1208(pMSIN-H) followed by
aerosol EZ had the highest PRN titer and frequency of MV-specific IgA and IgG antibody-secreting cells (ASC) after challenge (c and d), which likely represent
mucosally primed B cells homing to effector sites in response to virus exposure. This animal also showed the highest levels of MV-specific effector T cells with
the capacity to produce IFN-g after challenge (e).
674 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
ARTICLES
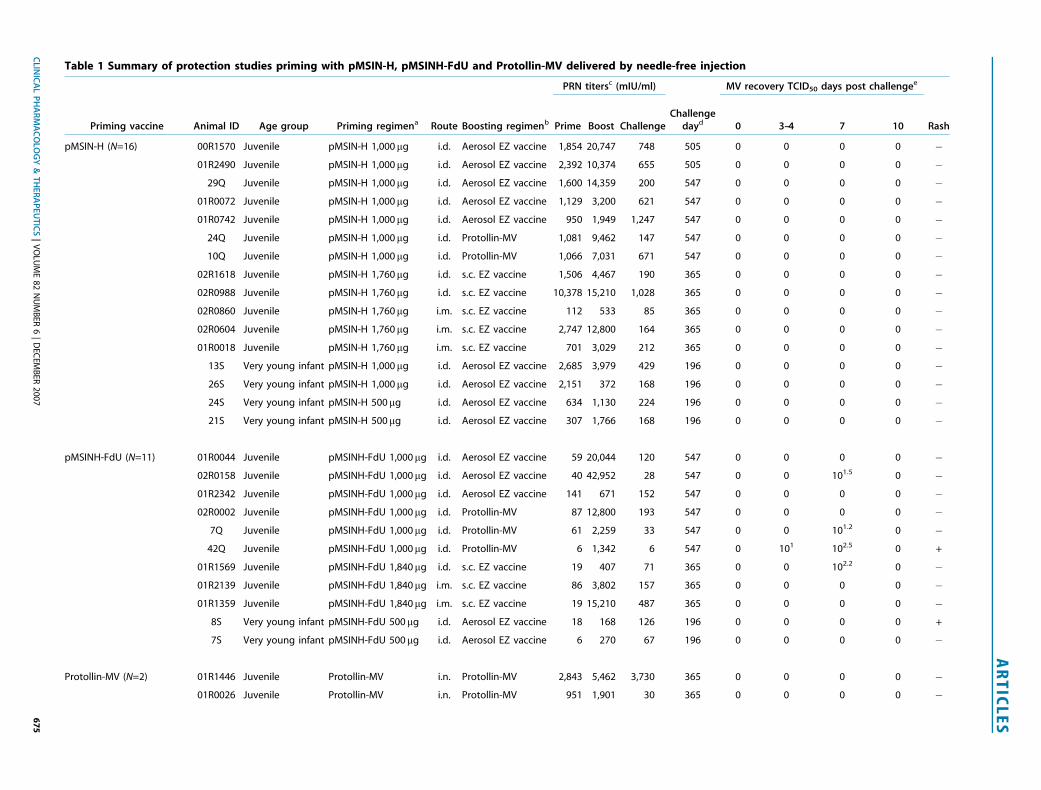
Table 1 Summary of protection studies priming with pMSIN-H, pMSINH-FdU and Protollin-MV delivered by needle-free injection
PRN titersc (mIU/ml) MV recovery TCID50 days post challengee
Priming vaccine Animal ID Age group Priming regimena Route Boosting regimenb Prime Boost ChallengeChallenge
dayd 0 3–4 7 10 Rash
pMSIN-H (N=16) 00R1570 Juvenile pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 1,854 20,747 748 505 0 0 0 0 �01R2490 Juvenile pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 2,392 10,374 655 505 0 0 0 0 �
29Q Juvenile pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 1,600 14,359 200 547 0 0 0 0 �01R0072 Juvenile pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 1,129 3,200 621 547 0 0 0 0 �01R0742 Juvenile pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 950 1,949 1,247 547 0 0 0 0 �
24Q Juvenile pMSIN-H 1,000mg i.d. Protollin-MV 1,081 9,462 147 547 0 0 0 0 �10Q Juvenile pMSIN-H 1,000mg i.d. Protollin-MV 1,066 7,031 671 547 0 0 0 0 �
02R1618 Juvenile pMSIN-H 1,760mg i.d. s.c. EZ vaccine 1,506 4,467 190 365 0 0 0 0 �02R0988 Juvenile pMSIN-H 1,760mg i.d. s.c. EZ vaccine 10,378 15,210 1,028 365 0 0 0 0 �02R0860 Juvenile pMSIN-H 1,760mg i.m. s.c. EZ vaccine 112 533 85 365 0 0 0 0 �02R0604 Juvenile pMSIN-H 1,760mg i.m. s.c. EZ vaccine 2,747 12,800 164 365 0 0 0 0 �01R0018 Juvenile pMSIN-H 1,760mg i.m. s.c. EZ vaccine 701 3,029 212 365 0 0 0 0 �
13S Very young infant pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 2,685 3,979 429 196 0 0 0 0 �26S Very young infant pMSIN-H 1,000mg i.d. Aerosol EZ vaccine 2,151 372 168 196 0 0 0 0 �24S Very young infant pMSIN-H 500mg i.d. Aerosol EZ vaccine 634 1,130 224 196 0 0 0 0 �21S Very young infant pMSIN-H 500mg i.d. Aerosol EZ vaccine 307 1,766 168 196 0 0 0 0 �
pMSINH-FdU (N=11) 01R0044 Juvenile pMSINH-FdU 1,000mg i.d. Aerosol EZ vaccine 59 20,044 120 547 0 0 0 0 �02R0158 Juvenile pMSINH-FdU 1,000mg i.d. Aerosol EZ vaccine 40 42,952 28 547 0 0 101.5 0 �01R2342 Juvenile pMSINH-FdU 1,000mg i.d. Aerosol EZ vaccine 141 671 152 547 0 0 0 0 �02R0002 Juvenile pMSINH-FdU 1,000mg i.d. Protollin-MV 87 12,800 193 547 0 0 0 0 �
7Q Juvenile pMSINH-FdU 1,000mg i.d. Protollin-MV 61 2,259 33 547 0 0 101.2 0 �42Q Juvenile pMSINH-FdU 1,000mg i.d. Protollin-MV 6 1,342 6 547 0 101 102.5 0 +
01R1569 Juvenile pMSINH-FdU 1,840mg i.d. s.c. EZ vaccine 19 407 71 365 0 0 102.2 0 �01R2139 Juvenile pMSINH-FdU 1,840mg i.m. s.c. EZ vaccine 86 3,802 157 365 0 0 0 0 �01R1359 Juvenile pMSINH-FdU 1,840mg i.m. s.c. EZ vaccine 19 15,210 487 365 0 0 0 0 �
8S Very young infant pMSINH-FdU 500mg i.d. Aerosol EZ vaccine 18 168 126 196 0 0 0 0 +
7S Very young infant pMSINH-FdU 500mg i.d. Aerosol EZ vaccine 6 270 67 196 0 0 0 0 �
Protollin-MV (N=2) 01R1446 Juvenile Protollin-MV i.n. Protollin-MV 2,843 5,462 3,730 365 0 0 0 0 �01R0026 Juvenile Protollin-MV i.n. Protollin-MV 951 1,901 30 365 0 0 0 0 �
CLIN
ICA
LP
HA
RM
AC
OLO
GY
&T
HE
RA
PE
UT
ICS
|V
OLU
ME
82
NU
MB
ER6
|D
ECEM
BER
200
76
75
AR
TIC
LE
S
exhibited high levels of viremia on several consecutive days,and one that received Protollin-MV developed clinical disease(Figure 3e and f). High levels of PRN antibodies and MV-specific IFN-g-secreting T cells, similar for all animals, wereobserved following MV challenge (data not shown).
Another arm of this study included four MV seronegativejuvenile macaques that received three doses of Protollin-MVor placebo (Figure 3g). One of the two animals immunizedwith Protollin-MV had a strong PRN response after the firstdose and both responded to subsequent doses. These animalswere protected when challenged with wild-type MV (Figure3g). No significant antibody responses were detected inplacebo recipients and when challenged they exhibited highviremia and measles rash. The Protollin-MV vaccine elicitedmeasles-specific IFN-g-secreting T cells (Figure 4d).
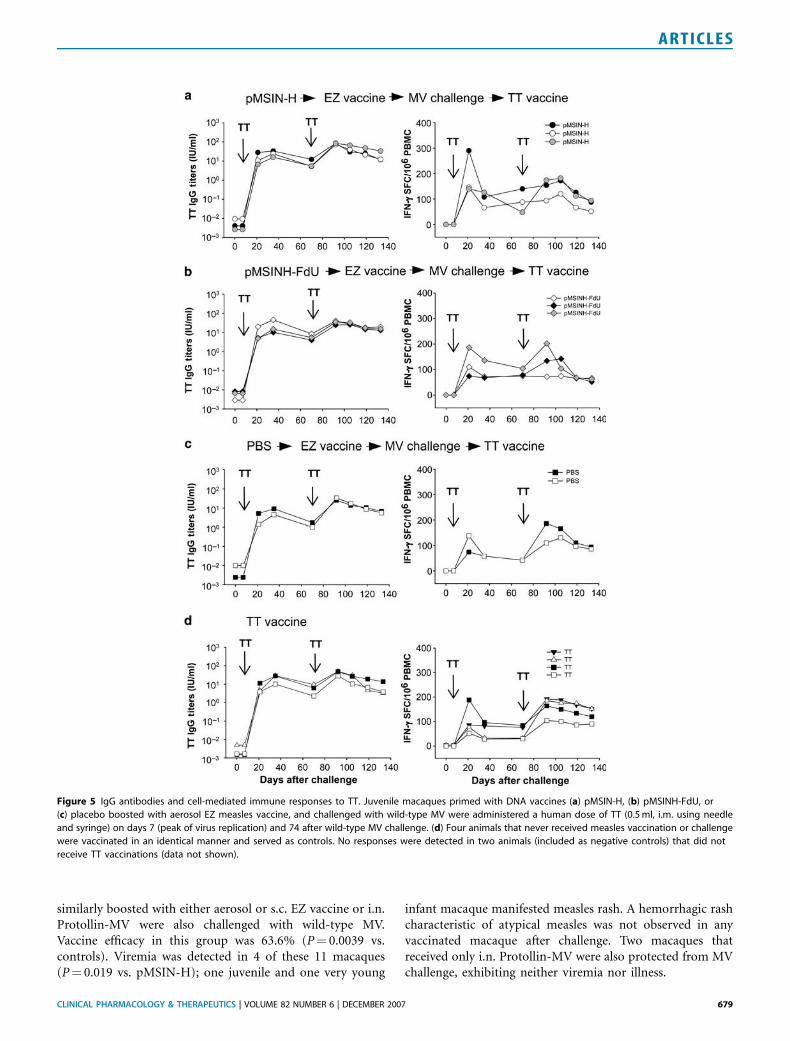
MV infection is known to be associated with immuno-suppression characterized by a shift to Th2-type responses,downregulation of interleukin-12, and impaired T-cellfunction that leads to increased susceptibility to secondaryinfections with other pathogens.18 Accordingly, we investi-gated the capacity of juvenile macaques that had been primedwith pMSIN-H, pMSINH-FdU, or PBS, boosted with aerosolEZ vaccine, and challenged with wild-type MV to respond toan unrelated vaccine antigen, tetanus toxoid (TT), given ondays 7 (peak of virus replication) and 74 after challenge(Figure 5a–c). A similar approach was described toinvestigate immunosuppression in MV DNA-vaccinatedand challenged infant macaques.14 Four control macaquesthat had not undergone either measles vaccination orchallenge were also given TT (Figure 5d). There was noapparent dampening of either antibody or IFN-g-secretingT-cell responses to TT in vaccinated/challenged animals vs.the unvaccinated/unchallenged controls.
Routes of inoculation for measles DNA vaccine priming
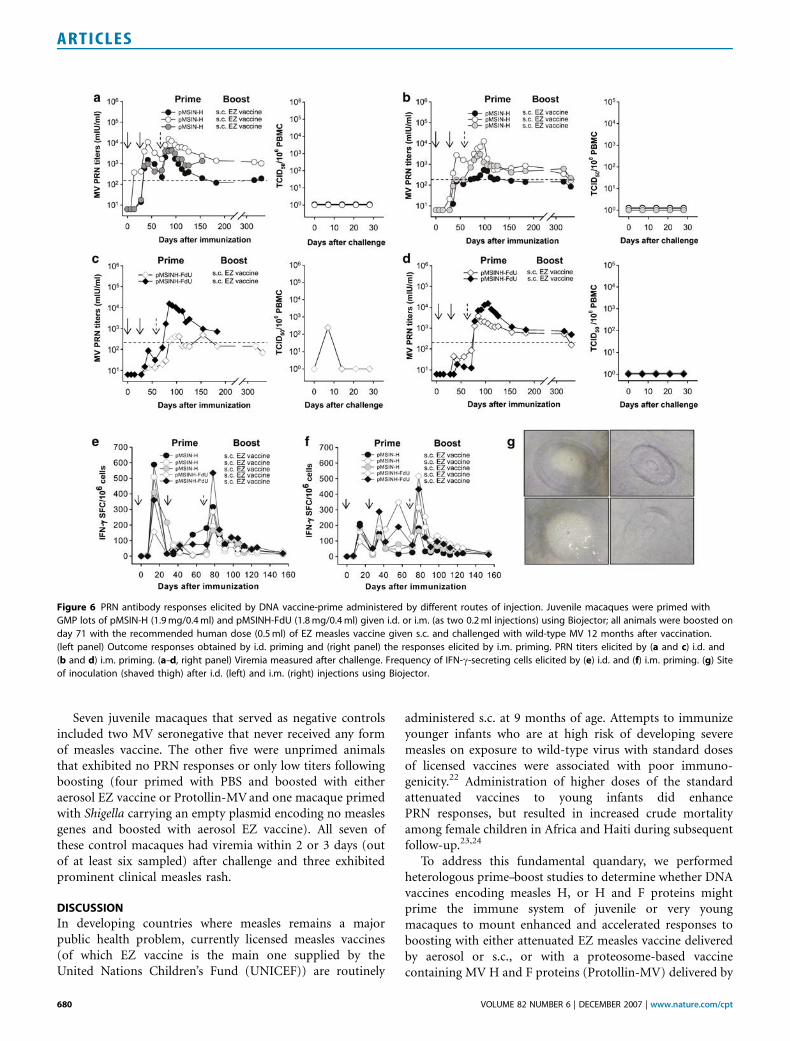
To compare parenteral routes of injection of measles DNAvaccines, 10 juvenile macaques were primed with eitherpMSIN-H or pMSINH-FdU administered i.d. or i.m. byBiojector, followed by boosting with the EZ vaccineadministered subcutaneously (s.c.). GMP pilot lots ofpMSIN-H and pMSINH-FdU manufactured for an impend-ing Phase 1 clinical trial were used. The vaccines andimmunization procedures were well tolerated by all animals.Although the numbers of macaques per group are small, i.d.priming with measles DNA vaccines appeared similarlyefficient as i.m. in eliciting PRN antibodies (Figure 6a–d).As observed previously, pMSIN-H elicited higher PRNtiters than pMSINH-FdU. All achieved protective titersafter boosting. IFN-g-secreting cells were also observedafter priming, regardless of the route, and after the boost(Figure 6e and f).
Protective efficacy was assessed by challenging eightmacaques 12 months after primary immunization. Unavail-able for challenge were two monkeys, one immunized i.d.with pMSIN-H and one i.d. with pMSINH-FdU, that died,respectively, on days 178 and 212 during follow-up forT
ab
le1
Co
nti
nu
ed
PR
Nti
ters
c(m
IU/m
l)M
Vre
cov
ery
TC
ID5
0d
ays
po
stch
all
en
ge
e
Pri
min
gv
acc
ine
An
ima
lID
Ag
eg
rou
pP
rim
ing
reg
ime
na
Ro
ute
Bo
ost
ing
reg
ime
nb
Pri
me
Bo
ost
Ch
alle
ng
eC
ha
llen
ge
da
yd0
3–4
71
0R
ash
Co
ntr
ols
(no
pri
min
g)
(N=
7)
99
R1
98
4Ju
ven
ileC
VD
12
08
(pSI
NC
P)
i.n.
Ae
roso
lEZ
vacc
ine
64
16
50
50
01
02
.81
02
.8�
01
R0
89
4Ju
ven
ileP
BS
i.d.
Ae
roso
lEZ
vacc
ine
61
68
39
54
70
10
1.2
10
2.5
0�
01
R0
65
0Ju
ven
ileP
BS
i.d.
Ae
roso
lEZ
vacc
ine
66
26
54
71
01
.21
04
.51
02
.2�
5Q
Juve
nile
PB
Si.d
.P
roto
llin
-MV
66
65
47
01
01
.81
04
.51
03
.8+
+
25
PJu
ven
ileP
BS
i.d.
Pro
tolli
n-M
V6
66
54
70
01
03
.21
04
.5+
03
R0
35
2Ju
ven
ileN
aive
—N
aive
66
63
65
01
01
.81
05
.11
05
.1+
+
10
20
Juve
nile
Nai
ve—
Nai
ve6
66
36
50
10
1.2
10
3.8
10
3.5
�
EZ,E
dm
on
sto
n–Z
agre
b;i
.d.,
intr
ade
rmal
;i.m
.,in
tram
usc
ula
r;i.n
.,in
tran
asal
;MV
,me
asle
svi
rus;
PB
S,p
ho
sph
ate
-bu
ffe
red
salin
e;P
RN
,pla
qu
e-r
ed
uct
ion
ne
utr
aliz
atio
n;s
.c.,
sub
cuta
ne
ou
s;T
CID
,tis
sue
cult
ure
infe
ctin
gd
ose
.PR
Nti
ters
pM
SIN
-Hvs
.co
ntr
ols
afte
rp
rim
e,
P=
0.0
00
2;
afte
rb
oo
st,
P=
0.0
00
2;
pre
-ch
alle
ng
e,
P=
0.0
00
2.
PR
Nti
ters
pM
SIN
H-F
dU
vs.
con
tro
lsaf
ter
pri
me
,P
=0
.00
3;
afte
rb
oo
st,
P=
0.0
00
6;p
re-c
hal
len
ge
,P
=0
.00
3.
PR
Nti
ters
pM
SIN
-Hvs
.p
MSI
NH
-Fd
Uaf
ter
pri
me
,Po
0.0
00
1;a
fte
rb
oo
st,P
=0
.57
;pre
-ch
alle
ng
e,
P=
0.0
01
.Vir
em
iain
mac
aqu
es
pri
me
dw
ith
pM
SIN
-H(0
/16
)ve
rsu
sco
ntr
ols
(7/7
),Po
0.0
00
1.V
ire
mia
inm
acaq
ue
sp
rim
ed
wit
hp
MSI
NH
-Fd
U(4
/11
)vs
.co
ntr
ols
(7/7
),P
=0
.01
3.V
ire
mia
inm
acaq
ue
sp
rim
ed
wit
hp
MSI
N-H
(0/1
6)
vs.a
nim
als
pri
me
dw
ith
pM
SIN
H-F
dU
(4/1
1),
P=
0.0
19
.aA
llp
rim
ing
reg
ime
ns
invo
lve
dtw
od
ose
sad
min
iste
red
28
day
sap
art.
bA
llb
oo
stin
gre
gim
en
sin
volv
ed
asi
ng
led
ose
adm
inis
tere
d2
8–4
3d
ays
afte
rp
rim
ing
.cP
eak
PR
Nti
ters
afte
rp
rim
e,
bo
ost
and
atth
eti
me
of
chal
len
ge
.dIn
rela
tio
nto
init
iati
on
of
imm
un
izat
ion
.eR
eco
very
of
MV
fro
mP
BM
Caf
ter
chal
len
ge
wit
hw
ild-t
ype
MV
.
676 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
ARTICLES
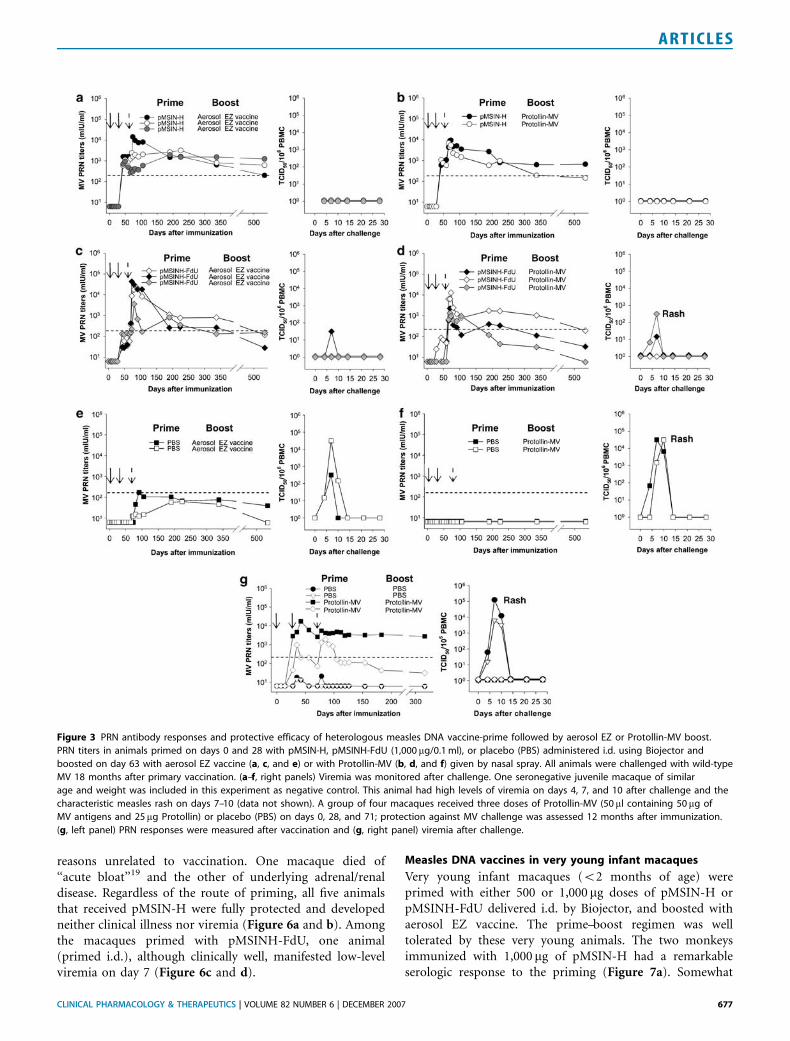
reasons unrelated to vaccination. One macaque died of‘‘acute bloat’’19 and the other of underlying adrenal/renaldisease. Regardless of the route of priming, all five animalsthat received pMSIN-H were fully protected and developedneither clinical illness nor viremia (Figure 6a and b). Amongthe macaques primed with pMSINH-FdU, one animal(primed i.d.), although clinically well, manifested low-levelviremia on day 7 (Figure 6c and d).
Measles DNA vaccines in very young infant macaques
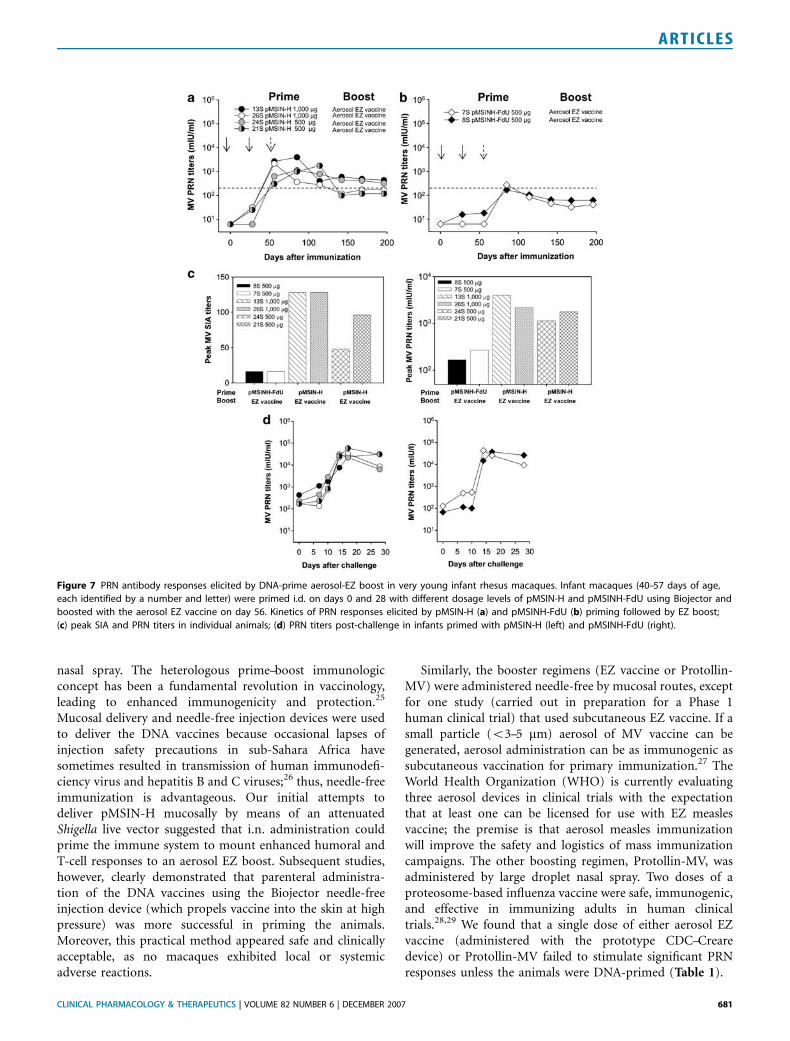
Very young infant macaques (o2 months of age) wereprimed with either 500 or 1,000 mg doses of pMSIN-H orpMSINH-FdU delivered i.d. by Biojector, and boosted withaerosol EZ vaccine. The prime–boost regimen was welltolerated by these very young animals. The two monkeysimmunized with 1,000 mg of pMSIN-H had a remarkableserologic response to the priming (Figure 7a). Somewhat
Figure 3 PRN antibody responses and protective efficacy of heterologous measles DNA vaccine-prime followed by aerosol EZ or Protollin-MV boost.
PRN titers in animals primed on days 0 and 28 with pMSIN-H, pMSINH-FdU (1,000 mg/0.1 ml), or placebo (PBS) administered i.d. using Biojector and
boosted on day 63 with aerosol EZ vaccine (a, c, and e) or with Protollin-MV (b, d, and f) given by nasal spray. All animals were challenged with wild-type
MV 18 months after primary vaccination. (a–f, right panels) Viremia was monitored after challenge. One seronegative juvenile macaque of similar
age and weight was included in this experiment as negative control. This animal had high levels of viremia on days 4, 7, and 10 after challenge and the
characteristic measles rash on days 7–10 (data not shown). A group of four macaques received three doses of Protollin-MV (50 ml containing 50 mg of
MV antigens and 25mg Protollin) or placebo (PBS) on days 0, 28, and 71; protection against MV challenge was assessed 12 months after immunization.
(g, left panel) PRN responses were measured after vaccination and (g, right panel) viremia after challenge.
CLINICAL PHARMACOLOGY & THERAPEUTICS | VOLUME 82 NUMBER 6 | DECEMBER 2007 677
ARTICLES
lower and delayed PRN responses were observed when only500 mg of pMSIN-H was given. Nevertheless, animals primedwith the lower dose developed PRN titers around the
protective threshold after priming and further responded tothe EZ aerosol boost. The infant macaques primed withpMSINH-FdU developed protective PRN titers only after theEZ boost and these titers dropped rapidly below protectivethreshold (Figure 7b).
We investigated the capacity of antibodies elicited bypMSIN-H and pMSINH-FdU in these young macaques toinhibit syncytium formation using B95-8 cells, which expressthe mammalian wild-type MV receptor CD150 or signalinglymphocyte activation molecule, as opposed to Vero cellsused in PRN assays, which allow virus infection throughCD46 receptor binding. There was a significant correlationbetween titers measured by syncytium inhibition assay (SIA)and PRN (r¼ 0.83; Po0.001, data not shown). Monkeysimmunized with pMSIN-H showed higher peak SIA titersthan those immunized with pMSINH-FdU, and the SIAresponses were clearly dose-dependent (Figure 7c). Theyoung infant macaques also exhibited MV-specific IFN-g-secreting T cells (data not shown).
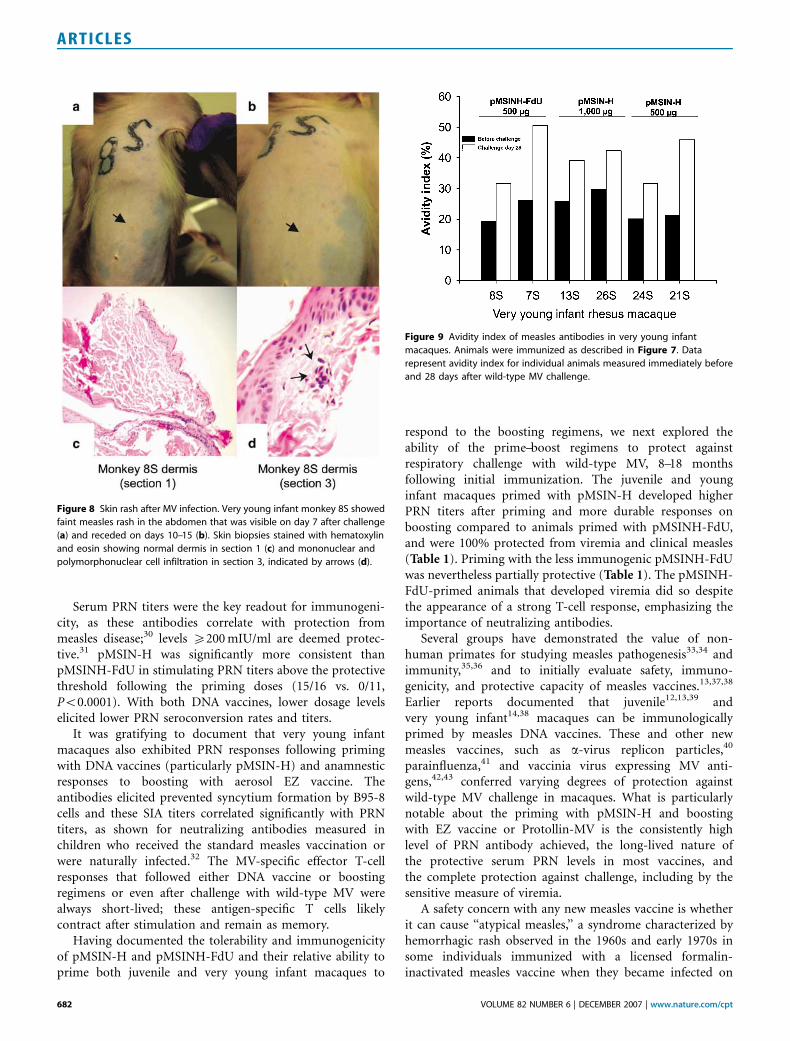
The six young infant macaques were challenged with wild-type MV at B9 months of age. All had strong anamnesticPRN responses after challenge (Figure 7d). None developedviremia and only one (primed with pMSINH-FdU) devel-oped isolated faint macules on the abdomen that werecompatible with mild measles rash (Figure 8); this animaldid not exhibit any sign of illness. The rash did not have ahemorrhagic component and did not clinically resembleatypical measles. One macule was biopsied and showed onlyminimal subtle changes that included a focal perivascularmixed infiltrate of mononuclear cells and neutrophils, typicalof classical measles in macaques20 (Figure 8c and d). Thefocal histological changes were seen only in deeper sections;the overlying epidermis was normal. There was neithererythrocyte extravasation nor infiltration by eosinophils,features described in skin biopsies from macaques withatypical measles rash.20
In subjects previously immunized with the formalin-inactivated measles vaccine, antibodies of low avidityproduced early after exposure to wild-type MV have beenassociated with the atypical measles syndrome.21 Accordingly,we examined the avidity of MV-specific IgG in the veryyoung infant macaques immediately before and 28 days afterchallenge with wild-type MV. The avidity of MV antibodiesunequivocally increased after challenge (Figure 9).
Summary of protection studies
Table 1 summarizes the outcome of the DNA-primed andcontrol animals challenged with wild-type MV. A total of 16macaques were primed with pMSIN-H and subsequentlyboosted with the EZ vaccine given by aerosol or s.c. or withProtollin-MV. Notably, vaccine efficacy in macaques primedwith pMSIN-H was 100% (Po0.001 vs. controls); none ofthese animals exhibited any sign of measles illness or viremiaon any day following respiratory challenge with MV.
Another 11 macaques primed with pMSINH-FdU (usingthe same dosage levels and routes as for pMSIN-H) and
Figure 4 T-cell responses elicited by measles DNA vaccine-prime followed
by aerosol EZ measles vaccine or Protollin-MV boost. Frequency of
circulating IFN-g-secreting T cells in macaques primed with pMSIN-H
(a), pMSINH-FdU (b), or placebo (c) and boosted with aerosol EZ vaccine
or with Protollin-MV (b) as described in Figure 3. (d) IFN-g-secreting
T cells in macaques primed and boosted with Protollin-MV or placebo as
described in Figure 3.
678 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
ARTICLES
similarly boosted with either aerosol or s.c. EZ vaccine or i.n.Protollin-MV were also challenged with wild-type MV.Vaccine efficacy in this group was 63.6% (P¼ 0.0039 vs.controls). Viremia was detected in 4 of these 11 macaques(P¼ 0.019 vs. pMSIN-H); one juvenile and one very young
infant macaque manifested measles rash. A hemorrhagic rashcharacteristic of atypical measles was not observed in anyvaccinated macaque after challenge. Two macaques thatreceived only i.n. Protollin-MV were also protected from MVchallenge, exhibiting neither viremia nor illness.
Figure 5 IgG antibodies and cell-mediated immune responses to TT. Juvenile macaques primed with DNA vaccines (a) pMSIN-H, (b) pMSINH-FdU, or
(c) placebo boosted with aerosol EZ measles vaccine, and challenged with wild-type MV were administered a human dose of TT (0.5 ml, i.m. using needle
and syringe) on days 7 (peak of virus replication) and 74 after wild-type MV challenge. (d) Four animals that never received measles vaccination or challenge
were vaccinated in an identical manner and served as controls. No responses were detected in two animals (included as negative controls) that did not
receive TT vaccinations (data not shown).
CLINICAL PHARMACOLOGY & THERAPEUTICS | VOLUME 82 NUMBER 6 | DECEMBER 2007 679
ARTICLES
Seven juvenile macaques that served as negative controlsincluded two MV seronegative that never received any formof measles vaccine. The other five were unprimed animalsthat exhibited no PRN responses or only low titers followingboosting (four primed with PBS and boosted with eitheraerosol EZ vaccine or Protollin-MV and one macaque primedwith Shigella carrying an empty plasmid encoding no measlesgenes and boosted with aerosol EZ vaccine). All seven ofthese control macaques had viremia within 2 or 3 days (outof at least six sampled) after challenge and three exhibitedprominent clinical measles rash.
DISCUSSION
In developing countries where measles remains a majorpublic health problem, currently licensed measles vaccines(of which EZ vaccine is the main one supplied by theUnited Nations Children’s Fund (UNICEF)) are routinely
administered s.c. at 9 months of age. Attempts to immunizeyounger infants who are at high risk of developing severemeasles on exposure to wild-type virus with standard dosesof licensed vaccines were associated with poor immuno-genicity.22 Administration of higher doses of the standardattenuated vaccines to young infants did enhancePRN responses, but resulted in increased crude mortalityamong female children in Africa and Haiti during subsequentfollow-up.23,24
To address this fundamental quandary, we performedheterologous prime–boost studies to determine whether DNAvaccines encoding measles H, or H and F proteins mightprime the immune system of juvenile or very youngmacaques to mount enhanced and accelerated responses toboosting with either attenuated EZ measles vaccine deliveredby aerosol or s.c., or with a proteosome-based vaccinecontaining MV H and F proteins (Protollin-MV) delivered by
Figure 6 PRN antibody responses elicited by DNA vaccine-prime administered by different routes of injection. Juvenile macaques were primed with
GMP lots of pMSIN-H (1.9 mg/0.4 ml) and pMSINH-FdU (1.8 mg/0.4 ml) given i.d. or i.m. (as two 0.2 ml injections) using Biojector; all animals were boosted on
day 71 with the recommended human dose (0.5 ml) of EZ measles vaccine given s.c. and challenged with wild-type MV 12 months after vaccination.
(left panel) Outcome responses obtained by i.d. priming and (right panel) the responses elicited by i.m. priming. PRN titers elicited by (a and c) i.d. and
(b and d) i.m. priming. (a–d, right panel) Viremia measured after challenge. Frequency of IFN-g-secreting cells elicited by (e) i.d. and (f) i.m. priming. (g) Site
of inoculation (shaved thigh) after i.d. (left) and i.m. (right) injections using Biojector.
680 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
ARTICLES
nasal spray. The heterologous prime–boost immunologicconcept has been a fundamental revolution in vaccinology,leading to enhanced immunogenicity and protection.25
Mucosal delivery and needle-free injection devices were usedto deliver the DNA vaccines because occasional lapses ofinjection safety precautions in sub-Sahara Africa havesometimes resulted in transmission of human immunodefi-ciency virus and hepatitis B and C viruses;26 thus, needle-freeimmunization is advantageous. Our initial attempts todeliver pMSIN-H mucosally by means of an attenuatedShigella live vector suggested that i.n. administration couldprime the immune system to mount enhanced humoral andT-cell responses to an aerosol EZ boost. Subsequent studies,however, clearly demonstrated that parenteral administra-tion of the DNA vaccines using the Biojector needle-freeinjection device (which propels vaccine into the skin at highpressure) was more successful in priming the animals.Moreover, this practical method appeared safe and clinicallyacceptable, as no macaques exhibited local or systemicadverse reactions.
Similarly, the booster regimens (EZ vaccine or Protollin-MV) were administered needle-free by mucosal routes, exceptfor one study (carried out in preparation for a Phase 1human clinical trial) that used subcutaneous EZ vaccine. If asmall particle (o3–5 mm) aerosol of MV vaccine can begenerated, aerosol administration can be as immunogenic assubcutaneous vaccination for primary immunization.27 TheWorld Health Organization (WHO) is currently evaluatingthree aerosol devices in clinical trials with the expectationthat at least one can be licensed for use with EZ measlesvaccine; the premise is that aerosol measles immunizationwill improve the safety and logistics of mass immunizationcampaigns. The other boosting regimen, Protollin-MV, wasadministered by large droplet nasal spray. Two doses of aproteosome-based influenza vaccine were safe, immunogenic,and effective in immunizing adults in human clinicaltrials.28,29 We found that a single dose of either aerosol EZvaccine (administered with the prototype CDC–Crearedevice) or Protollin-MV failed to stimulate significant PRNresponses unless the animals were DNA-primed (Table 1).
Figure 7 PRN antibody responses elicited by DNA-prime aerosol-EZ boost in very young infant rhesus macaques. Infant macaques (40–57 days of age,
each identified by a number and letter) were primed i.d. on days 0 and 28 with different dosage levels of pMSIN-H and pMSINH-FdU using Biojector and
boosted with the aerosol EZ vaccine on day 56. Kinetics of PRN responses elicited by pMSIN-H (a) and pMSINH-FdU (b) priming followed by EZ boost;
(c) peak SIA and PRN titers in individual animals; (d) PRN titers post-challenge in infants primed with pMSIN-H (left) and pMSINH-FdU (right).
CLINICAL PHARMACOLOGY & THERAPEUTICS | VOLUME 82 NUMBER 6 | DECEMBER 2007 681
ARTICLES
Serum PRN titers were the key readout for immunogeni-city, as these antibodies correlate with protection frommeasles disease;30 levels X200 mIU/ml are deemed protec-tive.31 pMSIN-H was significantly more consistent thanpMSINH-FdU in stimulating PRN titers above the protectivethreshold following the priming doses (15/16 vs. 0/11,Po0.0001). With both DNA vaccines, lower dosage levelselicited lower PRN seroconversion rates and titers.
It was gratifying to document that very young infantmacaques also exhibited PRN responses following primingwith DNA vaccines (particularly pMSIN-H) and anamnesticresponses to boosting with aerosol EZ vaccine. Theantibodies elicited prevented syncytium formation by B95-8cells and these SIA titers correlated significantly with PRNtiters, as shown for neutralizing antibodies measured inchildren who received the standard measles vaccination orwere naturally infected.32 The MV-specific effector T-cellresponses that followed either DNA vaccine or boostingregimens or even after challenge with wild-type MV werealways short-lived; these antigen-specific T cells likelycontract after stimulation and remain as memory.
Having documented the tolerability and immunogenicityof pMSIN-H and pMSINH-FdU and their relative ability toprime both juvenile and very young infant macaques to
respond to the boosting regimens, we next explored theability of the prime–boost regimens to protect againstrespiratory challenge with wild-type MV, 8–18 monthsfollowing initial immunization. The juvenile and younginfant macaques primed with pMSIN-H developed higherPRN titers after priming and more durable responses onboosting compared to animals primed with pMSINH-FdU,and were 100% protected from viremia and clinical measles(Table 1). Priming with the less immunogenic pMSINH-FdUwas nevertheless partially protective (Table 1). The pMSINH-FdU-primed animals that developed viremia did so despitethe appearance of a strong T-cell response, emphasizing theimportance of neutralizing antibodies.
Several groups have demonstrated the value of non-human primates for studying measles pathogenesis33,34 andimmunity,35,36 and to initially evaluate safety, immuno-genicity, and protective capacity of measles vaccines.13,37,38
Earlier reports documented that juvenile12,13,39 andvery young infant14,38 macaques can be immunologicallyprimed by measles DNA vaccines. These and other newmeasles vaccines, such as a-virus replicon particles,40
parainfluenza,41 and vaccinia virus expressing MV anti-gens,42,43 conferred varying degrees of protection againstwild-type MV challenge in macaques. What is particularlynotable about the priming with pMSIN-H and boostingwith EZ vaccine or Protollin-MV is the consistently highlevel of PRN antibody achieved, the long-lived nature ofthe protective serum PRN levels in most vaccines, andthe complete protection against challenge, including by thesensitive measure of viremia.
A safety concern with any new measles vaccine is whetherit can cause ‘‘atypical measles,’’ a syndrome characterized byhemorrhagic rash observed in the 1960s and early 1970s insome individuals immunized with a licensed formalin-inactivated measles vaccine when they became infected on
Figure 8 Skin rash after MV infection. Very young infant monkey 8S showed
faint measles rash in the abdomen that was visible on day 7 after challenge
(a) and receded on days 10–15 (b). Skin biopsies stained with hematoxylin
and eosin showing normal dermis in section 1 (c) and mononuclear and
polymorphonuclear cell infiltration in section 3, indicated by arrows (d).
Figure 9 Avidity index of measles antibodies in very young infant
macaques. Animals were immunized as described in Figure 7. Data
represent avidity index for individual animals measured immediately before
and 28 days after wild-type MV challenge.
682 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
ARTICLES

subsequent exposure to wild-type MV (reviewed in refs.13,20, 21, 44). Polack et al.20,21 demonstrated that this formalin-inactivated vaccine induces high levels of complement-fixing,low-avidity antibodies that fail to neutralize MV in vivo anddeposit as immunocomplexes. It was hypothesized that theformalin-inactivated vaccine fails to elicit T-cell helpnecessary for affinity maturation and for selection andmaturation of long-lived antibody-secreting B cells.21 Thesustained levels of PRN antibodies induced by our DNAvaccines, particularly pMSIN-H, are consistent with adequateT-cell help for the development of long-lived plasma andmemory B cells. We found no evidence of atypical measles inany of our monkeys primed with DNA vaccines and boostedwith EZ or Protollin-MV, or in the animals primed andboosted with Protollin-MV.
pMSINH-FdU was in fact constructed on the hypothesisthat immunizing with MV F as well as H protein mightenhance immunogenicity and safety by avoiding the‘‘glycoprotein imbalance’’ and lack of stimulation of anti-bodies against F (necessary to prevent cell-to-cell virusspread), previously presumed to be associated with atypicalmeasles.44,45 The addition of F did not provide any furtherimprovement to the already excellent safety profile of the H-encoding vaccine. Moreover, immunization with pMSINHF-dU not only failed to enhance protective immunity, but alsothe resulting PRN titers and protective efficacy wereconsistently lower compared with pMSIN-H. Earlier, Polacket al.13 had shown that co-administration of two DNAvaccines encoding H and F, respectively, resulted in lowerantibody responses than immunization with a single DNAvaccine encoding H.
One additional experiment that we would like to performis to compare the immunogenicity and efficacy of primingwith pMSIN-H and boosting with EZ vaccine in very younginfant macaques with and without maternally derivedmeasles antibody. Nevertheless, on the basis of the studiesperformed to date, in particular, the strategy of priming withpMSIN-H and boosting with EZ vaccine (either via aerosolor s.c. routes) appears to be a rational, apparently safe,practical, and efficacious regimen worthy of evaluation inhigh-risk young human infants. Data from future clinicaltrials of this regimen in the target age group in measlesendemic areas will determine the usefulness of this potentialadjunct tool in the struggle to eliminate mortality and severemorbidity as a result of measeles among young infants indeveloping countries.
METHODS
Vaccines. Sindbis virus-derived replicons, pMSIN-H andpMSINH-FdU, encoding full-length MV H, or H and F proteinswere developed at CVD and purified as described previously.4,5
GMP lots of pMSIN-H and pMSINH-FdU containing 4.4 and4.6 mg/ml, respectively, were produced by Althea (San Diego, CA).Shigella CVD 1208(pMSIN-H) and CVD 1208(pSINCP) wereprepared as described previously.4 Monovalent EZ measles vaccinewas produced by the Serum Institute of India (Pune, India; lots2259, 2259, EU2617). The Protollin-MV was produced as described
previously.17 Tetanus and diphtheria toxoid vaccine (DECAVAC)was produced by Sanofi Pasteur (Swiftwater, PA).
Rhesus macaques, immunizations, and sample collection. Juvenilerhesus macaques (Macaca mulatta; 2- to 4-year-old, weighing2.5–3.5 kg) seronegative for MV, as determined by PRN assay, wereobtained from the Johns Hopkins University (JHU) primate facilityor from Three Spring Scientific (Perkasie, PA). Very young infantmacaques (40- to 57-day-old, weighing 550–850 g) seronegative forMV were bred at the JHU primate facility. Animals wereanesthetized i.m. with ketamine (15 mg/kg, young infants) orketamine (10 mg/kg) and xylazine (0.5 mg/kg, juveniles). Shigellalive vector vaccines were delivered o.g. (1010 colony-forming unit(CFU) in 1 ml) following sodium bicarbonate via a pediatric feedingtube (Neo Care; Klein-Baker Medical, San Antonio, TX) or i.n.(1� 1010 CFU/50 ml, 25ml/naris) delivered by pipette. DNA vaccineswere administered using Biojector 2000 (Bioject, Portland, OR) in0.1 or 0.2 ml by i.d. or i.m injection in different sites of the shavedthighs. The EZ vaccine was delivered via nasal prong (0.15 ml over30 s and 0.5 ml over 90 s for juveniles, and 5 ml over 2 min for veryyoung infants) using a vibrating mesh nebulizer developed by theCDC and Creare (Hanover, NH)16 or s.c. (0.5 ml) using needle andsyringe. The Protollin-MV was delivered in 50 ml volume (25ml/naris) containing 50 mg of MV antigens and 25 mg of Protollin bynasal spray device (Valois SA, Marly le Roi, France). Heparinizedblood samples were collected at different time points for isolation ofplasma and peripheral blood mononuclear cells (PBMCs). Animalstudies were approved by the University of Maryland and the JHUAnimal Care and Use Committees.
Virus challenge. Animals were inoculated intratracheally with90–100 ml containing 0.9–1� 104 tissue culture infecting dose 50%(TCID50) of wild-type MV Bilthoven strain, as described pre-viously.13,46
MV titration. Viremia was assessed by incubating serially dilutedPBMCs with B958 cells and scoring for the presence of syncytia bylight microscopy.47 The number of cells resulting in 50% of thecultures becoming infected (TCID50) was calculated by Reed andMuench.
PRN and SIA. PRN titers were measured as described4 andexpressed in mIU/ml (WHO standard 66/202). SIA was performedas described previously.32,48
TT-specific antibodies. TT-specific serum IgG was measured byenzyme-linked immunosorbent assay.49 Horseradish peroxidase-labeled anti-monkey IgG (KPL, Gaithersburg, MD) was used as aconjugate. Titers were expressed in IU/ml (WHO Tetanus AntitoxinHuman Immunoglobulin Reference NIBSC no. 76/589, UK).
Avidity of MV-specific antibodies. Avidity of MV-specific IgG wasdetermined by enzyme-linked immunosorbent assay with anadditional 6 M urea wash.5,48,50 Horseradish peroxidase-anti-monkeyIgG (KPL) was used as a conjugate. Avidity indices were calculatedas percentage of residual activity (end-point titer) after ureatreatment.
MV-specific ASC. PBMCs were purified by density gradientcentrifugation using Lymphocyte Separation Medium (ICN, Bio-medicals, Aurora, OH). Cells were washed and re-suspended inRPMI containing 10% fetal calf serum, 200 mM L-glutamine, 10 mM
HEPES ((4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid)), and50 mg of gentamicin (Invitrogen, Grand Island, NY). The frequencyof MV-specific ASC was measured as described previously.48 Cells
CLINICAL PHARMACOLOGY & THERAPEUTICS | VOLUME 82 NUMBER 6 | DECEMBER 2007 683
ARTICLES

were added in serial dilutions to nitrocellulose plates (Millipore,Billerica, MA) coated with 5 mg of MV lysate (AdvancedBiotechnologies, Columbia, MD). Antibodies were detected withbiotin-labeled goat anti-monkey IgG and IgA (Nordic Labs, Tilburg,The Netherlands) followed by streptavidin–horseradish peroxidase(Sigma-Aldrich) and TrueBlue substrate (KPL). Results are reportedas mean SFC per 106 PBMCs from replicate wells after subtraction ofSFC in control wells.
IFN-g-secreting cells. MV- and TT-specific IFN-g-secreting T cellswere measured by ELISpot as described previously.48 Serially dilutedPBMCs were incubated for 36 h with 10mg/ml of MV lysate(Advanced Biotechnologies) or 5 mg/ml of TT (Statens SerumInstitute, Copenhagen, Denmark) in nitrocellulose plates (Milli-pore) coated with 15 mg/ml of anti-monkey IFN-g (Mabtech,Stockholm, Sweden) followed by biotin-anti-monkey IFN-g at1 mg/ml and streptavidin–horseradish peroxidase (Mabtech) andTrueBlue substrate (KPL). Results are expressed as mean IFN-g SFCper 106 PBMCs from replicate cultures after subtraction of SFC incontrol wells.
Statistical analysis. The association between PRN and SIA titerswas assessed by Pearson’s correlation. Differences in PRN titers forthe different groups and time points were examined by two-tailedWilcoxon rank-sum test. Protective efficacy was analyzed by two-tailed Fisher’s exact test. Differences with Po0.05 were consideredsignificant. Statistical analysis was performed using SigmaStat 3.1(Systat, San Jose, CA) and NCSS (Kaysville, UT) software.
ACKNOWLEDGMENTSWe thank Dr Diane Griffin (Bloomberg School of Hygiene and Public
Health, Johns Hopkins University, Baltimore) for her support and
assistance throughout the studies, Dr Karen Kotloff (CVD) and Dr Marcelo
B Sztein (CVD) for helpful discussions, Dr Paul Rota (CDC Atlanta) for
providing the EZ live attenuated measles vaccine, Dr Cinthia
Drachemberg (Department of Pathology, University of Maryland) for
analysis of measles rash biopsies, Dr William Blackwelder (CVD) for
assistance with statistical analysis, Dr Jakub Simon (CVD), Dr James
Campbell (CVD), Ms Kim Rincavage (CVD), and Ms Janine Linden (GSK
Biologicals of NA) for assistance during immunization of young infants
and juvenile macaques, and Mr Yu Lim, Marlen Martinez, Ivonne
Arciniega-Martinez, Suhlan Wu, and Mardi Reymann (CVD Applied
Immunology Section) for outstanding technical support. We also thank Dr
Ivan Tatarov and Ms Theresa M Alexander (Veterinary Resources,
University of Maryland) for assistance in challenge experiments and Dr
Wilbur Chen (CVD) for serving as medical monitor during challenge
studies. This work was supported by a grant from the Bill and Melinda
Gates Foundation to M.M. Levine.
CONFLICT OF INTEREST
The authors declared no conflict of interest.
& 2007 American Society for Clinical Pharmacology and Therapeutics
1. Wolfson, L.J., Strebel, P.M., Gacic-Dobo, M., Hoekstra, E.J., McFarland,J.W. & Hersh, B.S. Measles initiative. Has the 2005 measles mortality
reduction goal been achieved? A natural history modelling study.
Lancet 369, 191–200 (2007).2. Tapia, M.D. et al. A serosurvey to identify the window of vulnerability
to wild-type measles among infants in rural Mali. Am. J. Trop. Med.
Hyg. 73, 26–31 (2005).3. Stein, C.E., Birmingham, M., Kurian, M., Duclos, P. & Strebel, P. The
global burden of measles in the year 2000—a model that uses
country-specific indicators. J. Infect. Dis. 187 (Suppl 1), S8–S14 (2003).4. Pasetti, M.F. et al. Attenuated Salmonella enterica serovar Typhi and
Shigella flexneri 2a strains mucosally deliver DNA vaccines encoding
measles virus hemagglutinin, inducing specific immune responses
and protection in cotton rats. J. Virol. 77, 5209–5217 (2003).
5. Song, M.K. et al. Characterization of immune responses induced byintramuscular vaccination with DNA vaccines encoding measles virushemagglutinin and/or fusion proteins. J. Virol. 79, 9854–9861 (2005).
6. Polo, J.M. et al. Alphavirus DNA and particle replicons for vaccinesand gene therapy. Dev. Biol. (Basel) 104, 181–185 (2000).
7. Leitner, W.W. et al. Alphavirus-based DNA vaccine breaksimmunological tolerance by activating innate antiviral pathways. Nat.Med. 9, 33–39 (2003).
8. Cella, M. et al. Maturation, activation, and protection of dendritic cellsinduced by double-stranded RNA. J. Exp. Med. 189, 821–829 (1999).
9. Kirman, J.R. et al. Enhanced immunogenicity to Mycobacteriumtuberculosis by vaccination with an alphavirus plasmid repliconexpressing antigen 85A. Infect. Immun. 71, 575–579 (2003).
10. Leitner, W.W. et al. Apoptosis is essential for the increased efficacy ofalphaviral replicase-based DNA vaccines. Vaccine 22, 1537–1544(2004).
11. Adkins, B., Leclerc, C. & Marshall-Clarke, S. Neonatal adaptiveimmunity comes of age. Nat. Rev. Immunol. 4, 553–564 (2004).
12. Stittelaar, K.J. et al. Priming of measles virus-specific humoral- andcellular-immune responses in macaques by DNA vaccination. Vaccine20, 2022–2026 (2002).
13. Polack, F.P. et al. Successful DNA immunization against measles:neutralizing antibody against either the hemagglutinin or fusionglycoprotein protects rhesus macaques without evidence of atypicalmeasles. Nat. Med. 6, 776–781 (2000).
14. Premenko-Lanier, M., Rota, P.A., Rhodes, G.H., Bellini, W.J. &McChesney, M.B. Protection against challenge with measles virus(MV) in infant macaques by an MV DNA vaccine administered in thepresence of neutralizing antibody. J. Infect. Dis. 189, 2064–2071(2004).
15. Kotloff, K.L. et al. Deletion in the Shigella enterotoxin genes furtherattenuates Shigella flexneri 2a bearing guanine auxotrophy in a phase1 trial of CVD 1204 and CVD 1208. J. Infect. Dis. 190, 1745–1754(2004).
16. de Swart, R.L. et al. Aerosol measles vaccination in macaques:preclinical studies of immune responses and safety. Vaccine 24,6424–6436 (2006).
17. Chabot, S. et al. A novel intranasal Protollin-based measlesvaccine induces mucosal and systemic neutralizing antibodyresponses and cell-mediated immunity in mice. Vaccine 23,1374–1383 (2005).
18. Moss, W.J., Ota, M.O. & Griffin, D.E. Measles: immune suppression andimmune responses. Int. J. Biochem. Cell Biol. 36, 1380–1385 (2004).
19. Smith, A.W., Casey, H.W., LaCroix, J.T. & Johnson, D.K. Acute bloatsyndrome (gastric dilatation) in Macaca mulatta. J. Am. Vet. Med.Assoc. 155, 1241–1244 (1969).
20. Polack, F.P. et al. Production of atypical measles in rhesus macaques:evidence for disease mediated by immune complex formation andeosinophils in the presence of fusion-inhibiting antibody. Nat. Med. 5,629–634 (1999).
21. Polack, F.P., Hoffman, S.J., Crujeiras, G. & Griffin, D.E. A role fornonprotective complement-fixing antibodies with low avidity formeasles virus in atypical measles. Nat. Med. 9, 1209–1213 (2003).
22. Cutts, F.T., Grabowsky, M. & Markowitz, L.E. The effect of dose andstrain of live attenuated measles vaccines on serological responses inyoung infants. Biologicals 23, 95–106 (1995).
23. Aaby, P. et al. Sex-specific differences in mortality after high-titremeasles immunization in rural Senegal. Bull. World Health Organ. 72,761–770 (1994).
24. Holt, E.A., Moulton, L.H., Siberry, G.K. & Halsey, N.A. Differentialmortality by measles vaccine titer and sex. J. Infect. Dis. 168,1087–1096 (1993).
25. Woodland, D.L. Jump-starting the immune system: prime-boostingcomes of age. Trends Immunol. 25, 98–104 (2004).
26. Kane, A., Lloyd, J., Zaffran, M., Simonsen, L. & Kane, M. Transmission ofhepatitis B, hepatitis C and human immunodeficiency viruses throughunsafe injections in the developing world: model-based regionalestimates. Bull. World Health Organ. 77, 801–807 (1999).
27. Valdespino-Gomez, J.L. et al. Measles aerosol vaccination. Curr. Top.Microbiol. Immunol. 304, 165–193 (2006).
28. Treanor, J. et al. Intranasal administration of a proteosome-influenzavaccine is well-tolerated and induces serum and nasal secretioninfluenza antibodies in healthy human subjects. Vaccine 24, 254–262(2006).
684 VOLUME 82 NUMBER 6 | DECEMBER 2007 | www.nature.com/cpt
ARTICLES

29. Langley, J.M. et al. Safety and immunogenicity of aproteosome-trivalent inactivated influenza vaccine, given nasallyto healthy adults. Vaccine 24, 1601–1608 (2006).
30. Chen, R.T. et al. Measles antibody: reevaluation of protective titers.J. Infect. Dis. 162, 1036–1042 (1990).
31. Cutts, F.T. Module7: Measles, The Immunological Basis forImmunization Series (World health Organization, Geneva, 1993).
32. Ward, B.J. et al. Measurement of measles virus-specific neutralizingantibodies: evaluation of the syncytium inhibition assay incomparison with the plaque reduction neutralization test. Diagn.Microbiol. Infect. Dis. 33, 147–152 (1999).
33. Auwaerter, P.G. et al. Measles virus infection in rhesus macaques:altered immune responses and comparison of the virulence of sixdifferent virus strains. J. Infect. Dis. 180, 950–958 (1999).
34. El Mubarak, H.S. et al. Infection of cynomolgus macaques (Macacafascicularis) and rhesus macaques (Macaca mulatta) with differentwild-type measles viruses. J. Gen. Virol. 88, 2028–2034 (2007).
35. Permar, S.R. et al. Limited contribution of humoral immunity to theclearance of measles viremia in rhesus monkeys. J. Infect. Dis. 190,998–1005 (2004).
36. van Binnendijk, R.S., van der Heijden, R.W., van, A.G., Uytdehaag, F.G.& Osterhaus, A.D. Viral replication and development of specificimmunity in macaques after infection with different measles virusstrains. J. Infect. Dis. 170, 443–448 (1994).
37. van Binnendijk, R.S., van der Heijden, R.W. & Osterhaus, A.D. Monkeysin measles research. Curr. Top. Microbiol. Immunol. 191, 135–148(1995).
38. Premenko-Lanier, M. et al. DNA vaccination of infants in the presenceof maternal antibody: a measles model in the primate. Virology 307,67–75 (2003).
39. Premenko-Lanier, M., Rota, P., Rhodes, G., Bellini, W. & McChesney, M.Prior DNA vaccination does not interfere with the live-attenuatedmeasles vaccine. Vaccine 22, 762–765 (2004).
40. Pan, C.H. et al. Inaugural article: modulation of disease T cellresponses, and measles virus clearance in monkeys vaccinated with
H-encoding alphavirus replicon particles. Proc. Natl. Acad. Sci. USA102, 11581–11588 (2005).
41. Skiadopoulos, M.H., Surman, S.R., Riggs, J.M., Collins, P.L. & Murphy,B.R. A chimeric human-bovine parainfluenza virus type 3 expressingmeasles virus hemagglutinin is attenuated for replication but is stillimmunogenic in rhesus monkeys. J. Virol. 75, 10498–10504 (2001).
42. Zhu, Y. et al. Evaluation of recombinant vaccinia virus—measlesvaccines in infant rhesus macaques with preexisting measlesantibody. Virology 276, 202–213 (2000).
43. Stittelaar, K.J. et al. Protective immunity in macaques vaccinated witha modified vaccinia virus Ankara-based measles virus vaccine in thepresence of passively acquired antibodies. J. Virol. 74, 4236–4243(2000).
44. Polack, F.P. Atypical measles and enhanced respiratory syncytial virusdisease (ERD) made simple. Pediatr. Res. 62, 111–115 (2007).
45. Merz, D.C., Scheid, A. & Choppin, P.W. Importance of antibodies to thefusion glycoprotein of paramyxoviruses in the prevention of spreadof infection. J. Exp. Med. 151, 275–288 (1980).
46. Polack, F.P., Hoffman, S.J., Moss, W.J. & Griffin, D.E. Differential effectsof priming with DNA vaccines encoding the hemagglutinin and/orfusion proteins on cytokine responses after measles virus challenge.J. Infect. Dis. 187, 1794–1800 (2003).
47. Kobune, F., Sakata, H. & Sugiura, A. Marmoset lymphoblastoid cells asa sensitive host for isolation of measles virus. J. Virol. 64, 700–705(1990).
48. Capozzo, A.V. et al. Neonatal immunization with a Sindbisvirus-DNA measles vaccine induces adult-like neutralizing antibodiesand cell-mediated immunity in the presence of maternal antibodies.J. Immunol. 176, 5671–5681 (2006).
49. Tapia, M.D. et al. Measurement of tetanus antitoxin in oral fluid: a toolto conduct serosurveys 24. Pediatr. Infect. Dis. J. 25, 819–825 (2006).
50. de Souza, V.A., Pannuti, C.S., Sumita, L.M. & Andrade Junior, H.F.Enzyme-linked immunosorbent assay-IgG antibody avidity test forsingle sample serologic evaluation of measles vaccines. J. Med. Virol.52, 275–279 (1997).
CLINICAL PHARMACOLOGY & THERAPEUTICS | VOLUME 82 NUMBER 6 | DECEMBER 2007 685
ARTICLES
Related Documents











