Changing Patterns of Mobility as a Response to Climatic Deterioration and Aridification in the Middle Holocene Southern Sahara Christopher M. Stojanowski* and Kelly J. Knudson Center for Bioarchaeological Research, School of Human Evolution and Social Change, PO Box 872402, Arizona State University, Tempe, AZ 85287 KEY WORDS bioarchaeology; radiogenic strontium isotope analysis; North Africa; Saharan prehistory; paleomobility ABSTRACT The African Humid Period witnessed a rapid human re-occupation of the Sahara as numerous lakes formed during the Holocene climatic optimum circa 10-5 kya. Permanent waters attracted a variety of aquatic and terrestrial fauna allowing for long-term occupation of specific paleolake basins. The Gobero pale- olake in central Niger was one such location that pre- serves a unique mortuary record from the southern Sahara. Here, we use radiogenic strontium isotope anal- ysis to investigate how human communities adapted to aridification throughout the Holocene. In particular, we examine the effects of increasing climate instability on patterns of human mobility. Results of radiogenic stron- tium isotope analysis of enamel and bone samples from Middle Holocene burials (7.2–4.9 kya) indicate predom- inantly local values with no evidence for sex-based vari- ation. Comparisons of radiogenic strontium isotope data with previously published (Stojanowski and Knudson: Am J Phys Anthropol 146 (2011) 49–61) Early Holocene burials (9.7–8.3 kya) indicate significant differences in both enamel and bone values. Middle Holocene individu- als demonstrate a predominantly non-local signature for enamel values and a predominantly local signature for bone values. Those individuals with non-local bone val- ues always demonstrated non-local enamel values; how- ever, the opposite was not the case. This suggests a divergence of mobility strategies during the Middle Hol- ocene with a minority of individuals maintaining a more mobile existence throughout their life and others main- taining a similar strategy as Early Holocene hunter– gatherers that was tied to the paleolake basin. The more mobile individuals likely lived during the terminal phase of the lake’s occupation. One response to aridifica- tion by Saharan peoples, then, was increasing mobility. Am J Phys Anthropol 154:79–93, 2014. V C 2014 Wiley Peri- odicals, Inc. Climate change is one of the most important external stimuli affecting levels of population stress and overall well-being. Adaptable and resilient populations are able to buffer periodic climate stress through diverse strat- egies, the result of which is improved health and overall quality of life. We argue that inferring adaptive approaches from past populations informs modern strat- egies of resilience in regions with marginal, sub-optimal climates, or during short-term periods of stress affecting subsistence practices and water security. This dynamic relationship between climate, aridity, and human adapt- ability is perhaps no more evident than in the Sahara where, for much of the Holocene, human communities adapted to climate degradation that reduced surface waters and modified ecosystems. Throughout the Holo- cene, Saharan populations witnessed the transformation from hunting–gathering–fishing economies to pastoral and eventually agricultural economies, demonstrating diverse solutions to the problem of aridity and food inse- curity. Importantly, a community’s subsistence adapta- tion impacts group structure and composition, which in turn determine patterns of mobility across the human lifespan. Mobility is a key aspect of a population’s rela- tionship with the landscape, and the ability to move and the strategies involved in spatially organizing people are important buffers to short and long term changes in cli- mate. One of the most visible approaches for inferring paleomobility is through radiogenic strontium isotope ratios (see Bentley, 2006), which can be used to deter- mine life course changes in residence and extra-local in- migration, both of which determine group composition and affect biocultural adaptations and patterns of genetic variability. Previous studies of paleomobility in North Africa have focused on the following three contexts: the New King- dom of the Nile Valley (Buzon et al., 2007; Buzon and Simonetti, 2013), the Early and Middle Holocene sequence from the Libyan Fezzan (Tafuri et al., 2006; di Lernia and Tafuri, 2013; di Lernia et al., 2013), and the Gobero region of central Niger (Sereno et al., 2008; Grant sponsor: National Science Foundation; Grant numbers: BCS-0636066, BCS-0820805; Grant sponsor: Wenner-Gren Founda- tion; Grant number: GR-7747; Grant sponsors: Arizona State University Institute for Social Science Research Catalyst Grant, and Arizona State University School of Human Evolution and Social Change. *Correspondence to: C.M. Stojanowski, Center for Bioarchaeologi- cal Research, School of Human Evolution and Social Change, PO Box 872402, Arizona State University, Tempe, AZ 85287. E-mail: [email protected] Received 26 September 2013; accepted 7 January 2014 DOI: 10.1002/ajpa.22474 Published online 22 January 2014 in Wiley Online Library (wileyonlinelibrary.com). Ó 2014 WILEY PERIODICALS, INC. AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 154:79–93 (2014)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Changing Patterns of Mobility as a Response toClimatic Deterioration and Aridification in the MiddleHolocene Southern Sahara
Christopher M. Stojanowski* and Kelly J. Knudson
Center for Bioarchaeological Research, School of Human Evolution and Social Change, PO Box 872402, ArizonaState University, Tempe, AZ 85287
KEY WORDS bioarchaeology; radiogenic strontium isotope analysis; North Africa;Saharan prehistory; paleomobility
ABSTRACT The African Humid Period witnessed arapid human re-occupation of the Sahara as numerouslakes formed during the Holocene climatic optimumcirca 10-5 kya. Permanent waters attracted a variety ofaquatic and terrestrial fauna allowing for long-termoccupation of specific paleolake basins. The Gobero pale-olake in central Niger was one such location that pre-serves a unique mortuary record from the southernSahara. Here, we use radiogenic strontium isotope anal-ysis to investigate how human communities adapted toaridification throughout the Holocene. In particular, weexamine the effects of increasing climate instability onpatterns of human mobility. Results of radiogenic stron-tium isotope analysis of enamel and bone samples fromMiddle Holocene burials (�7.2–4.9 kya) indicate predom-inantly local values with no evidence for sex-based vari-ation. Comparisons of radiogenic strontium isotope datawith previously published (Stojanowski and Knudson:
Am J Phys Anthropol 146 (2011) 49–61) Early Holoceneburials (�9.7–8.3 kya) indicate significant differences inboth enamel and bone values. Middle Holocene individu-als demonstrate a predominantly non-local signature forenamel values and a predominantly local signature forbone values. Those individuals with non-local bone val-ues always demonstrated non-local enamel values; how-ever, the opposite was not the case. This suggests adivergence of mobility strategies during the Middle Hol-ocene with a minority of individuals maintaining a moremobile existence throughout their life and others main-taining a similar strategy as Early Holocene hunter–gatherers that was tied to the paleolake basin. Themore mobile individuals likely lived during the terminalphase of the lake’s occupation. One response to aridifica-tion by Saharan peoples, then, was increasing mobility.Am J Phys Anthropol 154:79–93, 2014. VC 2014 Wiley Peri-
odicals, Inc.
Climate change is one of the most important externalstimuli affecting levels of population stress and overallwell-being. Adaptable and resilient populations are ableto buffer periodic climate stress through diverse strat-egies, the result of which is improved health and overallquality of life. We argue that inferring adaptiveapproaches from past populations informs modern strat-egies of resilience in regions with marginal, sub-optimalclimates, or during short-term periods of stress affectingsubsistence practices and water security. This dynamicrelationship between climate, aridity, and human adapt-ability is perhaps no more evident than in the Saharawhere, for much of the Holocene, human communitiesadapted to climate degradation that reduced surfacewaters and modified ecosystems. Throughout the Holo-cene, Saharan populations witnessed the transformationfrom hunting–gathering–fishing economies to pastoraland eventually agricultural economies, demonstratingdiverse solutions to the problem of aridity and food inse-curity. Importantly, a community’s subsistence adapta-tion impacts group structure and composition, which inturn determine patterns of mobility across the humanlifespan. Mobility is a key aspect of a population’s rela-tionship with the landscape, and the ability to move andthe strategies involved in spatially organizing people areimportant buffers to short and long term changes in cli-mate. One of the most visible approaches for inferringpaleomobility is through radiogenic strontium isotoperatios (see Bentley, 2006), which can be used to deter-
mine life course changes in residence and extra-local in-migration, both of which determine group compositionand affect biocultural adaptations and patterns ofgenetic variability.
Previous studies of paleomobility in North Africa havefocused on the following three contexts: the New King-dom of the Nile Valley (Buzon et al., 2007; Buzon andSimonetti, 2013), the Early and Middle Holocenesequence from the Libyan Fezzan (Tafuri et al., 2006; diLernia and Tafuri, 2013; di Lernia et al., 2013), and theGobero region of central Niger (Sereno et al., 2008;
Grant sponsor: National Science Foundation; Grant numbers:BCS-0636066, BCS-0820805; Grant sponsor: Wenner-Gren Founda-tion; Grant number: GR-7747; Grant sponsors: Arizona StateUniversity Institute for Social Science Research Catalyst Grant,and Arizona State University School of Human Evolution and SocialChange.
*Correspondence to: C.M. Stojanowski, Center for Bioarchaeologi-cal Research, School of Human Evolution and Social Change, POBox 872402, Arizona State University, Tempe, AZ 85287. E-mail:[email protected]
Received 26 September 2013; accepted 7 January 2014
DOI: 10.1002/ajpa.22474Published online 22 January 2014 in Wiley Online Library
(wileyonlinelibrary.com).
� 2014 WILEY PERIODICALS, INC.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 154:79–93 (2014)

Stojanowski and Knudson, 2011). The cemeteries atGobero preserve one of the largest series of humanremains from North Africa (Sereno et al., 2008; Garcea,2013a; Stojanowski, 2013). There, human communitieslived at the interface of the Sahara and Sahel around anow extinct paleolake beginning circa 10 kya at theonset of the African Humid Period (e.g., Gasse and VanCampo, 1994; Hassan, 1997; Gasse, 2000, 2002; Kuperand Kr€opelin, 2006). There was nearly continuous occu-pation of the basin until circa 4.5 kya when the Saharanclimate approached its modern arid status (deMenocalet al. 2000). The occupational sequence at Gobero wasdisrupted by several arid phases (Sereno et al., 2008;Giraudi, 2013), the most prolonged of which was the 8.2kya event (e.g., Alley et al., 1997; Kobashi et al., 2007;Lal et al., 2007), an 800 year arid phase preceding theintroduction of cattle pastoralism into the central andsouthern Sahara (Clark et al., 2008; Cremaschi and Zer-boni, 2011; Mercuri et al., 2011). Although regionalsequences of lake formation varied with basin hydrology,morphology, and aquifer characteristics (see Gasse,2000, 2002), more arid conditions generally prevailedafter the 8.2 kya event. The skeletal material fromGobero brackets this significant climatic event and pro-vides a unique opportunity to observe how humangroups responded to the changing nature of the cli-mate—an Early Holocene climatic optimum character-ized by high lake levels with relatively uninterruptedhumidity and a Middle Holocene phase characterized bylower lake levels, greater variation in humid–aridcycling, and a general trend toward increasing ariditythrough time.
Previous isotopic analyses at Gobero focused on theEarly Holocene burials with the goal of reconstructingmobility patterns among the hunting–gathering–fishingpopulation (Stojanowski and Knudson, 2011). In thisarticle, we present radiogenic strontium isotopic analy-ses for 18 Middle Holocene burials from Gobero, comple-menting previously reported data for 13 Early Holoceneburials. We use these data to characterize changingmobility practices within individuals throughout the lifecourse and to determine whether variability in isotopeprofiles among individuals increased through time asmight be expected if some or all of the population prac-ticed transhumant pastoralism. We then compare thesedata to those previously published from the Early Holo-cene occupation phase to reconstruct how mobility pat-terns changed through time.
BIOARCHAEOLOGICAL CONTEXT
The prehistory of Niger
The archaeology of Niger is poorly known (Haour,2003), with sites often best described as deflated pal-impsests of mixed time horizons. Most information onEarly and Middle Holocene peoples derives from earlyFrench and British surveys that helped define the dis-tinction between Early Holocene hunting–gathering–fishing peoples and Middle Holocene pastoralists(reviewed in Haour, 2003; Garcea, 2013b). In the EarlyHolocene, the culture most well-known archaeologicallyis the Kiffian named after the type site Adrar n0Kiffi atAdrar Bous (Clark et al., 2008), and known sporadicallyfrom other sites at Adrar Bous and throughout northernNiger (Roset, 1983, 1987; �Echallier and Roset, 1986;Roset et al., 1990; Smith, 2008a). The Kiffian is a hunt-ing–gathering–fishing economy focused on aquatic
resources and processing wild grasses, and associatedwith a microlithic tool industry; coarse, decorated wavy-line pottery; grinding stones and bone harpoons. Kiffiangroups were thought to have been nomadic, living inseasonal camps on dune sites near the edges of nowextinct lakes or at the margins of rocky massifs.Archaeologically ephemeral, the makers of the Kiffianwere unknown skeletally prior to our work at Gobero(Sereno et al., 2008; Stojanowski, 2013).
The Middle Holocene of Niger is associated with theappearance of cattle pastoralism. The most well-knownculture group is the Tenerian, now identified at a numberof sites throughout the region (see Reygasse, 1934; Jou-bert and Vaufrey, 1941-46; Tixier, 1962; Smith, 2008b;Garcea, 2013b). Much of the spatial–temporal chronologyremains abstract with definitional debates emphasizingdifferences in lithic technology and sorting deflated depos-its into distinct time horizons. The unusual vitric greenstone tools recovered from Tenerian sites have become adefining aspect of the complex. The presence of domesti-cated cattle (Bos), often burnt or intentionally interred, isimportant but not solely diagnostic of Tenerian sites sinceother pastoralist groups were likely present in Niger atthe same time (Garcea, 2013b). Although domesticatedcattle is the most visible addition to the diet, hunting,fishing, and wild plants remained important subsistencefoci (see Clark et al., 2008; Sereno et al., 2008). Dating ofthe Tenerian is controversial but appears to occur fromcirca 7.5 to 5 kya at sites present in northern Niger, south-ern Algeria, and as far east as the Borkou region of north-ern Chad. The long distances between roughly similartypes of sites suggests a nomadic pastoral existence inwhich different adaptive strategies were implementeddepending on the home ranges of distinct pastoral com-munities and the seasonal constraints imposed by differ-ent microenvironments. Unlike contemporaneouspastoralist sites in southwestern Libya (Biagetti and diLernia, 2013; di Lernia et al., 2013), Tenerian sites andother mid-Holocene sites in Niger lack evidence of domes-ticated caprines, reflecting a generally wetter climate incomparison to the central Sahara.
The Gobero site complex
Human activity at the Gobero site complex was cen-tered on a paleolake that formed during the Early Holo-cene climatic optimum circa 10 kya and lasted untilapproximately 4.5 kya. The Gobero basin is endorheic anddefined on its southern margin by an uplifted Cretaceousfault (Mazelet) over which connections with the NigerRiver system and Lake Chad may have occurred duringhigh lake stands (Sereno et al., 2008; Giraudi, 2013). Dur-ing the hyper-arid terminal Pleistocene, aeolian dunesformed within the basin on top of the Cretaceous bedrock.During the Holocene, human burials were placed withinthese dunes (Fig. 1). Much of the basin is littered withdeflated material culture and partially fossilized fauna,reflective of the degree of erosion that has occurred sinceits human occupants abandoned the region. In addition,the dune surfaces preserve a density of lithic, faunal, andceramic materials suggesting that people also lived on thedune top surfaces. While a succession of mortuary andresidential activities may have occurred at differenttimes, limited chronological control of deflated materialscomplicates this interpretation.
Archaeological recovery of burials occurred in 2000,2006, and 2011 (Sereno et al., 2008). Human remains
80 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology

exhibited distinct taphonomic signatures that allowedallocation of all burials into two major occupationphases, confirmed radiometrically (Sereno et al., 2008;Stojanowski, 2013). The Early Holocene occupation datesfrom circa 9.7 to 8.3 kya with a majority of burials clus-tering on one dune (site G3—Fig. 1) and within a nar-row time horizon (Sereno et al., 2008; Stojanowski,2013). Burials allocated to the Early Holocene presenteddarkly stained and fossilized skeletons, robust infracra-nial skeletons, with tall, linear builds, and tightly con-stricted burial postures suggesting the use of perishablegrave containers (Stojanowski, 2013). These individualsare contemporaneous with Kiffian sites further north,and there is little doubt that they were hunting–fishing–gathering peoples attracted to the variety of taxa livingin and around the lake at that time. Pottery and grind-ing equipment also suggests parallels with the Kiffianindustry (Smith, 2008a) and a broad subsistence basethat utilized wild plant materials.
Middle Holocene burials at Gobero post-date the 8.2kya arid event with radiocarbon dates indicating a rangefrom 7.2 to 4.9 kya (Sereno et al., 2008). These burialsnever experienced lake level submergence and, as aresult, the skeletons were unfossilized, unstained, andmore friable, making them easily distinguishable fromthe older material from the site (Stojanowski, 2013). Bio-logically, the Middle Holocene peoples were more diversecraniometrically (Sereno et al., 2008), shorter in staturewith a more gracile infracranial skeleton, and buried inloosely flexed and more formalized burial postures (Sto-janowski, 2013). Middle Holocene burials were docu-mented at several sites within the Gobero site complexwith the greatest density occurring on sites G1 and G3(see Fig. 1). Although contemporaneous with Teneriansites further north, Garcea (2013b) notes that it may beinappropriate to apply this cultural label to Goberobecause not all Middle Holocene pastoralist sites inNiger have been attributed to the Tenerian. Still, Garcea(2013b) does consider the Middle Holocene inhabitantsof Gobero to be pastoralists, despite the limited directevidence for domesticated cattle at the site. The issue ofwhether these individuals were pastoralists remainscontroversial (see discussion in Sereno et al., 2008; Sto-janowski and Carver, 2011). Excavated Early and MiddleHolocene middens at Gobero show little change in thetype of taxa utilized through time (see Sereno et al.,
2008). It is possible the lake’s waters continued to pro-vide key resources throughout its occupation, perhapsobviating the adoption of pastoralism as an adaptivestrategy.
Climatic variation at Gobero
Climatic variation at Gobero is reconstructed based ongeneral climate trends in Niger (Gasse and Van Campo,1994; Gasse, 2000, 2002) and specific information onGobero’s basin hydrology and lake level transgressionscombined with the succession of aeolian (arid phase) andlacustrine (humid phase) sediments and pollen analysis(Sereno et al., 2008; Giraudi, 2013; Giraudi and Mercuri,2013; Mercuri et al., 2013). The Early and Middle Holo-cene are divided by the 8.2 kya arid event, dated atGobero based on OSL and radiocarbon determinationsfrom approximately 8.3 to 7.5 kya (Sereno et al., 2008).The period prior to this arid phase defines the EarlyHolocene occupation phase. The lake level reached itshighest extent during the Early Holocene with a highlake stand (>9 m) recorded around 8.7 kya (Serenoet al., 2008). The high water level reflects the climaticoptimum, and it was during this phase that hydrologicalconnections were made at different times with the NigerRiver system and with Lake Chad (Giraudi, 2013). Whilethere is little evidence for climatic instability during theEarly Holocene occupation phase, the onset of arid con-ditions circa 8.2 kya must have been rapid. The areawas completely abandoned for close to 800 years.
In contrast, the Middle Holocene occupation occurredduring less humid conditions with more rapid changesin humid-arid cycles. For example, during the earliestphase of the Middle Holocene (circa 7.5–6.8 kya), Gir-audi and Mercuri (2013) recorded four different phasesof aridity and hydrological balance within the basin inshort succession. Following an arid phase from 6.9 to 6.6kya, the lake formed once again, characterized as, “acomplex evolution with lake levels hard to define, butcharacterized by a barely positive [emphasis added]hydrological balance. . .much drier environmental condi-tions than previously. . .with semi-desert grassland anddesert vegetation (Giraudi and Mercuri, 2013:120).” Thelower lake levels (<4 m) also affected the faunal ecologywith large species of fish (such as Lates niloticus) unableto thrive in the shallow, low energy lacustrine
Fig. 1. Map of Africa showing the location of Niger (A) with major archaeological sites, geological features, and modern townsindicated. (B) Plan-view map of archaeological features within the Gobero paleolake basin with human burial sites (G1-7) andintact lakebed deposits (shaded gray areas) indicated. Figure modified after Sereno et al. (2008).
MID-HOLOCENE SAHARAN MOBILITY 81
American Journal of Physical Anthropology

environment. In general, once reformed after 6.6 kya thelake progressively declined into a marshy habitat untilit was abandoned by humans around 4.5 kya.
Previous studies of paleomobility in North Africa
Previous isotopic research at Gobero focused solely onthe Early Holocene forager population (Stojanowski andKnudson, 2011). That research established baseline radio-genic strontium isotopic data from soil and modern faunalsamples collected at Gobero; reconstructed underlyingbedrock geology for the Gobero basin, the Chad drainage,and the A€ır Massif; and evaluated the radiogenic stron-tium isotope data for diagenetic contamination. Resultsindicated “local” 87Sr/86Sr values among all individuals,limited life course variation, and significantly less inter-individual variation at Gobero in comparison to contempo-raneous hunter–gatherer populations (Tafuri et al., 2006).These data suggested a high degree of sedentism amongthe hunting–gathering–fishing population at Gobero (Sto-janowski and Knudson, 2011).
To help inform our interpretations of observed tempo-ral changes at Gobero, we utilized isotopic data fromcontemporaneous sites from southwestern Libya (Bia-getti and di Lernia, 2007, 2013; di Lernia et al., 2013;Tafuri et al., 2006). There, isotopic data reflected infer-ences about “local” versus “non-local” signatures at theinter-individual level within a cultural trajectory that isbroadly similar to that observed in Niger—that is,hunter–gatherer populations define the earliest occupa-tion phases (Early and Late Acacus) and pastoralist pop-ulations define the later occupation phases. Becauseexploring life course variability was not an option due topoor burial preservation at the Libyan sites, thisresearch focused on a “territorial scale of analysis” (diLernia et al., 2013:3) that produced inferences abouthow Saharan peoples moved between ecological nicheswithin their landscape. Results indicated that Late Aca-cus hunter–gatherers presented heterogeneity in radio-genic strontium isotope values, suggesting highresidential mobility (Tafuri et al., 2006). Inter-individualvariability decreased during the Early and Middle Pasto-ral periods and demonstrated predominantly “local”87Sr/86Sr values (Tafuri et al., 2006). The combinedarchaeological and isotopic data suggests seasonal move-ments within a relatively restricted range as part of themaintenance of cattle herds and their water needs (Bia-getti and di Lernia, 2013; di Lernia et al., 2013; Tafuriet al., 2006). Patterns of movement may have assumedan upland–lowland organization (Tafuri et al., 2006).Analyses of cattle dentition using microsampling techni-ques suggested domesticated cattle were of local origin,but confirmed a pattern of annual movement suggestingthe herds were managed successfully within the regionalenvironment (di Lernia et al., 2013). As more arid condi-tions prevailed, cattle were replaced with drought resist-ant caprines, and radiogenic strontium isotope datasuggest a shift to a more intense form of mobility withperhaps a change in residence pattern and social organi-zation (Tafuri et al., 2006).
RADIOGENIC STRONTIUM ISOTOPE ANALYSISAND PALEOMOBILITY
Radiogenic strontium isotope analysis utilizes geologicvariability to infer paleomobility from 87Sr/86Sr values inarchaeological enamel and bone (see overview in Bent-ley, 2006). Since 87Sr decays from 87Rb, 87Sr/86Sr values
vary according to the age and initial composition of thebedrock (Faure and Powell, 1972). Throughout an eco-system, 87Sr/86Sr does not fractionate appreciably asstrontium cycles through soil, groundwater, and plantsand animals (e.g., Aberg, 1995), so that the 87Sr/86Sr val-ues in enamel and bone reflects the 87Sr/86Sr values inthe strontium sources in the diet (Ericson, 1985; Priceet al., 1994). If the strontium sources in the diet arelocal, observed 87Sr/86Sr values will reflect the geologicalregion in which an individual lived during enamel orbone formation (see overview in Bentley, 2006).
A key aspect of paleomobility studies is identifyingand minimizing diagenetic, or post-depositional, contam-ination in archaeological samples. Bone is more suscepti-ble than enamel to diagenesis and there are variousapproaches to minimizing and identifying diagenetic
Fig. 2. Map of the central and southern Sahara showingunderlying bedrock geology and major archaeological sites dis-cussed in text. Libyan Sahara sites are indicated by the boxedarea (upper right corner). Lake Chad is in the lower right cor-ner. Geological data based on Schl€uter (2006) and Cratchleyet al. (1984). Expected radiogenic strontium isotope valuesbased on geological and faunal baseline data are listed. Sites:(1) Adar Bous, (2) Iwelen, (3) In Tuduf, (4) Chin Tafidet, (5)Afunfun. Site locations approximate to scale of the map, withlocations based on Haour (2003, Fig. 5). The approximateboundary of the Chad drainage is indicated by the white linethat divides the A€ır.
82 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology
contamination (e.g., Nelson et al., 1986; Lee-Thorp,2002, 2008; Lee-Thorp and Sponheimer, 2003). In addi-tion to using a weak acid wash to remove diageneticstrontium (Sillen, 1989; Sillen and Sealy, 1995), major,minor, and trace element concentrations can identifyhydroxyapatite in which elements like strontium havelikely been altered (e.g., Williams and Marlowe, 1987;Price et al., 2002; Gr€un et al., 2008; T€utken et al., 2008).
Radiogenic strontium isotope signatures in thesouthern Sahara
Gobero is located in the greater Chad Basin Drainagenear the division with the Iullemeden Basin to the east(Fig. 2). The Chad Basin is composed of Jurassic-Cretaceous (Mesozoic-Cenozoic) sandstones (Tegama For-mation—Wright et al., 1985; Begg et al., 2009), whichwere exposed at Gobero but overlain sporadically withPleistocene (Quaternary) dunes directly on Cretaceousbedrock (Sereno et al., 2008). These dunes formed duringthe Ogolian hyper-arid, are <20,000 years old, and werethe matrix within which the human burials were placed.
Based on the age and type of bedrock, we hypothesizethat individuals who obtained strontium near Goberowill likely exhibit 87Sr/86Sr �0.710 based on radiogenicstrontium isotope values from similar exposed bedrockand archaeological human remains in western Egyptand Libya (Schaaf and M€uller-Sohnius, 2002; Tafuriet al., 2006). However, radiogenic strontium isotope anal-yses of soil samples collected from burial contexts exhibitmean 87Sr/86Sr 5 0.71290 6 0.00064 (n 5 7, 1r) (Stoja-nowski and Knudson, 2011). As such, the dune sedi-ments could include some reworked, weathered materialfrom the A€ır Massif, as discussed below. Furthermore, asa proxy for bioavailable strontium, we previouslyreported data from modern and archaeological faunalsamples collected from Gobero; these data exhibit mean87Sr/86Sr 5 0.71261 6 0.00116 (n 5 16, 1r) (Stojanowskiand Knudson, 2011). Additional archaeological faunalsamples were collected and analyzed here, and presentedand discussed in subsequent sections.
Reconstructing paleomobility using radiogenic stron-tium isotope ratios requires variation in underlying bed-rock geology and an understanding of potential mobility
TABLE 1. Biogeochemical data from archaeological and modern faunal samples
Laboratory no. Specimen no. Species (material) 87Sr/86Sr
Gobero site complexACL-0548 GOB-BCS4 Antilopini sp.? (modern, tooth) 0.71157a
ACL-0549 GOB-BCS5 Rodent (modern, cranium) 0.71152a
ACL-0560 GOB-BCS16 Snail (modern, shell) 0.71120a
ACL-0561 GOB-BCS17 Hippopotamus sp. (archaeological, enamel) 0.71122a
ACL-0562 GOB-BCS18 Antilopini sp.? (modern, cranium) 0.71251a
ACL-4014 GOB-106N-130E Hippopotamus sp. (archaeological, enamel) 0.71119ACL-4015 GOB-151N-161N Hippopotamus sp. (archaeological, enamel) 0.71181ACL-4016 GOB-108-N-132E Pelomedusidae sp. (archaeological, bone) 0.71131ACL-4018 GOB-150N-161E-1 Crocodylus niloticus (archaeological, bone) 0.71140ACL-4020 GOB-150N-161E-2 Trionychidae sp. (archaeological, bone) 0.71166ACL-4025 GOB-84N-138E Kobus kob (archaeological, enamel) 0.71103ACL-4030 GOB-107N-132E-2 Lates niloticus (archaeological, bone) 0.71140ACL-4031 GOB-108N-132E-1 Lates niloticus (archaeological, bone) 0.71140ACL-4033 GOB-107N-132E-3 Clarias sp. (archaeological, bone) 0.71145ACL-4035 GOB-108N-132E-3 Tilapinii sp. (archaeological, bone) 0.71136ACL-4045 GOB-108N-130E-1 Pelomedusidae sp. (archaeological, bone) 0.71165ACL-4047 GOB-108N-130E-3 Crocodylus niloticus (archaeological, bone) 0.71166ACL-4048 GOB-107N-131E-1 Tilapinii sp. (archaeological, bone) 0.71150ACL-4050 GOB-107N-131E-3 Clarias sp. (archaeological, bone) 0.71137ACL-4051 GOB-107N-131E-4 Claridae sp. (archaeological, bone) 0.71144ACL-4052 GOB-107N-131E-5 Lates niloticus (archaeological, bone) 0.71149ACL-4053 GOB-107N-131E-6 Lates niloticus (archaeological, bone) 0.71137Air MassifACL-0545 GOB-BCS1 Capra hircus (modern, mandible) 0.71470a
ACL-0546 GOB-BCS2 Camelus dromedarius (modern, enamel) 0.71308a
ACL-0547 GOB-BCS3 Camelus dromedarius (modern, enamel) 0.71225a
ACL-0566 GOB-BCS2 Camelus dromedarius (modern, enamel) 0.71306a
ACL-0567 GOB-BCS2 Camelus dromedarius (modern, enamel) 0.71334a
Mazelet FaultACL-0550 GOB-BCS6 Bubo sp.? pellet (modern) 0.71261a
ACL-0551 GOB-BCS7 Bubo sp.? pellet (modern) 0.71217a
ACL-0552 GOB-BCS8 Bubo sp.? pellet (modern) 0.71238a
ACL-0553 GOB-BCS9 Bubo sp.? pellet (modern) 0.71444a
ACL-0554 GOB-BCS10 Bubo sp.? pellet (modern) 0.71160a
ACL-0555 GOB-BCS11 Bubo sp.? pellet (modern) 0.71477a
ACL-0556 GOB-BCS12 Bubo sp.? pellet (modern) 0.71080a
ACL-0557 GOB-BCS13 Antilopini sp.? Capra sp.? (modern, vertebra) 0.71354a
ACL-0558 GOB-BCS14 Caprine or ovine (modern, mandible) 0.71371a
ACL-0559 GOB-BCS15 Caprine or ovine (modern, mandible) 0.71176a
ACL-0563 GOB-BCS19 Antilopini sp.? (modern, cranium) 0.71252a
ACL-0564 GOB-BCS20 Antilopini sp.? (modern, horns) 0.71345a
a These samples have been previously published and discussed in detail elsewhere (Sereno et al., 2008; Stojanowski and Knudson, 2011).
MID-HOLOCENE SAHARAN MOBILITY 83
American Journal of Physical Anthropology

structuring features of the landscape. However, majortransformation of the North African climate since themid-Holocene complicates this analysis. Many lake andriver systems that may have organized human popula-tions on the landscape have disappeared over the last5,000 years and Holocene living surfaces have been sig-nificantly eroded. Therefore, we focus on enduring fea-tures of the landscape that may have attracted humanpopulations, including the A€ır Massif northwest ofGobero and Lake Chad located to the southeast. Thequality of baseline comparative data is variable for theseregions, particularly the Lake Chad region, whose shore-lines varied considerably within the Early and MiddleHolocene (Drake and Bristow, 2006; Drake et al., 2011).Modern proxies may be problematic because ofthe potential homogenizing effect of Saharan dust trans-port (Frumkin and Stein, 2004; Grousset et al., 1992;Rognon et al., 1996; Krom et al., 1999). Saharan dustexhibits mean 87Sr/86Sr 5 0.722 (Weldeab et al., 2002)and varies depending on particle size (Grousset et al.,1992, 1998; Rognon et al., 1996), from 87Sr/86Sr 5 0.713to 87Sr/86Sr 5 0.737 (Grousset et al., 1998). Significantinputs from modern dust may therefore increase meas-ured 87Sr/86Sr values in some of the baseline samples,particularly modern faunal samples.
The Chad Drainage is a large, intra-continental basinformed by uplifting events at its margins (Burke, 1976).Its interior sediments are post Pan-African in age (<550mya), which contrast with the circumferential, upliftedPrecambrian massifs. Today, most of the Chad Basin iscovered with Quaternary sediments such as the T�en�er�eDesert in northern Niger. While not relevant to paleomo-bility reconstructions at Gobero, Holocene dunes docover Pleistocene material that is visible throughout thesouthern basin (Burke, 1976; Drake and Bristow, 2006;Bouchette et al., 2010). Although the drainage basementconsists of Cretaceous sandstones, these are onlyexposed in the western drainage near the A€ır Massifand Gobero (The Tegama Formation). Genik (1992) andHartley and Allen (1994) date sandstones in the Nigersegment of the Chad Basin (Termit and T�en�er�e riftstructures) to the Miocene. Importantly, sediments thatwould have been present during the Middle Holocenetrend from younger near Lake Chad (due to greateraccumulation of Pleistocene sand and clays in the ChadFormation) to older near the A€ır Massif; however, mostof this variation manifests only near the A€ır Massifitself. These older bedrocks yield higher radiogenicstrontium isotope values and samples of Camelus drome-darius collected on the Agadez-Bilma piste betweenGobero and the A€ır Massif exhibited mean 87Sr/86Sr 50.71293 6 0.00047 (n 5 4, 1r) (Table 1) (Stojanowskiand Knudson, 2011).
Unlike the relatively homogenous Chad Basin, the A€ırMassif is geologically complex. It is part of the TuaregShield located on the West African Mobile belt that wasdeformed during the Pan-African orogeny circa 550 mya(Black et al., 1994). Mapped exposures include a pre-dominance of Precambrian basement basalts, Paleozoic(Silurian and Devonian) subvolcanic ring structures, andCenozoic volcanic rocks (Black et al., 1994; Li�egeoiset al., 1994; Schl€uter, 2006; Williams, 2008). There is amajor sheer zone that divides the A€ır Massif along itsnorth–south axis reflecting more recent volcanism. Sedi-ments of Devonian and Carboniferous age (Paleozoic)outcrop along the massif ’s western side, while the east-ern terrane (Aouzeguer) surrounding the Tafidet Basin
(an extension of the Tegama Formation) consists ofProche-T�en�er�e molassics (Fig. 2) (Li�egeois et al., 1994).Therefore, the eastern and western sides of the A€ır Mas-sif should exhibit different 87Sr/86Sr values. Youngerbasaltic flows are predominantly found in the southernhalf of the massif. Paleozoic anorogenic ring structuresare expected to yield 87Sr/86Sr 5 0.705–0.706 (Brownet al., 1989; Demaiffe et al., 1991), while Cenozoic vol-canic rocks should yield lower values of 87Sr/86Sr 50.703–0.706 (Leger, 1985; Petters, 1991; Dupuy et al.,1993; Black et al., 1994) and Precambrian granitesshould exhibit 87Sr/86Sr > 0.720 (Price et al., 2006). InStojanowski and Knudson (2011), we reported a juvenileCapra hircus sample from the A€ır Massif with 87Sr/86Sr5 0.71470, consistent with higher expected values forPrecambrian sediments. The complexity of geologicalfeatures and inputs of modern Saharan dust confirmsthe importance of generating baseline data fromarchaeological samples.
MATERIALS AND METHODS
Sampling strategy
We previously generated baseline radiogenic strontiumisotope data from soil samples collected from grave con-texts (n 5 7); from archaeological and modern faunalsamples from Gobero, the A€ır Massif, and along the pistebetween Gobero and Agadez (located at the southernend of the A€ır Massif) (n 5 22); and from human enameland bone samples from the Early Holocene burials atGobero (n 5 13) (Fig. 3) (Stojanowski and Knudson,2011). Here we present new data from the human buri-als that date to the Middle Holocene occupation phase(n 5 18) as well as new archaeological faunal samples(n 5 17). Both archaeological human and faunal enameland bone samples were selected based on observed pres-ervation and degree of exposure prior to excavation. Tounderstand changes in paleomobility throughout the lifecourse, adult first, second, and third molars, as well as anon-diagnostic segment of long bone or rib were collectedto provide isotopic evidence from dental and skeletal ele-ments that formed at different times in that individual’slife. Significantly eroded burials were not incorporatedinto the sampling strategy. Elements treated with bulk-ing agents (such as Rhoplex) or consolidants (such asAcryloid B-72) either in the field or laboratory were notselected for isotopic analysis.
Osteological analyses
In addition to generating radiogenic strontium isotopicdata, we also compared differences across sexes andthroughout the life course. Sex estimates were generatedfrom cranial and pelvic features following the protocol inBuikstra and Ubelaker (1994). Instead of direct age-at-death estimates based on skeletal indicators, we usedaverage M1 wear scores (maxillary and mandibular,antimeres) as a proxy for adult age at death. Juvenileage estimates were based on dental eruption and devel-opment, supplemented with long bone lengths and decid-uous and permanent dental wear scores (Buikstra andUbelaker, 1994).
Laboratory methodology
Archaeological enamel and bone samples were pre-pared and analyzed at Arizona State University (ASU)using published procedures (Knudson and Price, 2007;
84 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology

Knudson et al., 2012). After mechanical and chemicalcleaning in the Archaeological Chemistry Laboratory,strontium was separated from the sample matrix withEiChrom SrSpec resin. In the W.M. Keck FoundationLaboratory for Environmental Biogeochemistry, radio-genic strontium isotopes were measured on a Thermo-Finnigan Neptune multi-collector inductively coupledplasma mass spectrometer (MC-ICP-MS), where SRM-987 has exhibited 87Sr/86Sr 5 0.710265 6 0.000010 (2r,n 5 25). In addition, samples were analyzed on aThermo-Finnigan quadrupole inductively-coupledplasma mass spectrometer (Q-ICP-MS) for major, minor,and trace element concentrations, including calcium,phosphorus, uranium, and neodymium, in order to iden-tify the presence of diagenetic contamination; mean Ca/P 5 2.18 6 0.01 (2r, n 5 5) for ACL standard CUE-0001.
RESULTS AND INTERPRETATIONS
Evaluating diagenetic contamination
Major, minor, and trace element concentrations inarchaeological human enamel were analyzed as a proxyfor identifying diagenetic contamination (e.g., Williamsand Marlowe, 1987; Price et al., 2002; Gr€un et al., 2008;T€utken et al., 2008). In biogenic samples, Ca/P 5 2.1; atGobero, mean Ca/P 5 2.12 6 0.11 (n 5 89, 1r), whichwe interpret as evidence of little diagenetic contamina-tion. In addition, mean concentrations of uranium (U)and neodymium (Nd) are low; mean U/Ca 5 5.0 3 1025
6 2.2 3 1024 (n 5 79, 1r) and mean Nd/Ca 5 3.9 31024 6 6.531024 (n 5 84, 1r). While there were foursamples that exhibited Ca/P 5 2.4 (ACL-1635, ACL-1693) and Ca/P 5 2.5 (ACL-1660, ACL-1641), the U/Caand Nd/Ca values were not anomalously high for these
samples, indicating at least some biogenic strontium inthe samples. These values are within the range of pub-lished U/Ca enamel data that were interpreted as uncon-taminated (Price et al., 2002). In addition, the boneradiogenic strontium isotope data do not reflect a trendtoward the 87Sr/86Sr values in soil samples from Gobero,as expected if bone was significantly affected by diage-netic contamination. Both the mean human enamel87Sr/86Sr (P 5 0.010) and mean human bone 87Sr/86Srdata (P 5 0.001) are significantly different from the bur-ial soil data. In fact, bone data are more divergent fromthe soil data than are the enamel data. Archaeologicalfaunal samples from Gobero are also significantly differ-ent from the soil values (P < 0.001); however, the mod-ern faunal samples from Gobero are not significantlydifferent (P 5 0.959). This is the opposite of what wouldone expect if diagenetic contamination were affecting thearchaeological faunal and human 87Sr/86Sr values andsupports the biogenic nature of the data.
Identifying bioavailable 87Sr/86Sr values atGobero
Previously published soil samples collected from burialsat Gobero exhibit mean 87Sr/86Sr 5 0.71290 6 0.00064(n 5 7, 1r) (Sereno et al., 2008). In addition, previousresearch at Gobero demonstrated that modern faunalsamples collected from the Mazelet Fault region of theGobero basin exhibited mean 87Sr/86Sr 5 0.71281 60.00019 (n 5 12, 1r) and that mean 87Sr/86Sr 5 0.71329 60.00089 (n 5 5, 1r) from the A€ır Massif and the surround-ing region (Table 1) (Sereno et al., 2008; Stojanowski andKnudson 2011). Given the available data, we had previ-ously considered these values reflective of the range of87Sr/86Sr for the Gobero basin as well as the surroundingregions. However, new data from archaeological faunal
Fig. 3. Radiogenic strontium isotopes values from individuals buried in Early Holocene contexts in the Gobero site complex. Inthis figure, previously published data points (Stojanowski and Knudson, 2011) from dental and skeletal elements from one individualare arranged from early to late forming enamel followed by skeletal elements, and data points from the same individual are labeledand grouped together. The horizontal bar indicates the two standard deviation “local” range based on archaeological faunal samples.
MID-HOLOCENE SAHARAN MOBILITY 85
American Journal of Physical Anthropology

samples suggest that these data are more indicative ofbioavailable strontium values in the region than the mod-ern faunal and soil samples. For example, new archaeo-logical faunal 87Sr/86Sr values from the Goberosite complex exhibit mean 87Sr/86Sr 5 0.71143 6 0.00019(n 5 18, 1r) with a range of 87Sr/86Sr 5 0.71105 to87Sr/86Sr 5 0.71180 (Table 1). Within the Gobero region,archaeological faunal data are significantly different fromthe soil data (P 5 0.001) as well as the modern faunaldata (P 5 0.005). When all faunal data from Gobero andthe A€ır Massif are combined, the modern and archaeologi-cal data are significantly different (P < 0.001). In particu-lar, modern faunal and soil values are significantly higherthan the archaeological faunal samples, consistent withthe expectations of the effects of modern Saharan dustincreasing the values of bioavailable 87Sr/86Sr values.
Many of the species collected from Gobero were lake-dwelling species, and thus reflect the bioavailable stron-tium in the paleolake. Interestingly, the one terrestrialspecies represented (Kobus) has the lowest radiogenicstrontium isotope value. There are no significant differ-ences between archaeological faunal enamel and bonevalues (P 5 0.451), suggesting limited diagenetic con-tamination. In addition, 87Sr/86Sr values did not differsignificantly between fish versus non-fish (includingmammalian and reptilian) taxa (P 5 0.680). Given theaveraging effects of faunal samples and the heterogene-ity in soil samples (e.g., Price et al., 2002; Evans andTatham, 2004; Bentley et al., 2004; Bentley, 2006; Evanset al, 2010), as well as the significant differencesobserved between modern and archaeological samplesfrom the Gobero region, we argue that the most conserv-ative and accurate definition of a “local” range at Goberois based on archaeological faunal data. Therefore, wedefine the “local” radiogenic strontium isotope signaturebased on the mean archaeological faunal 87Sr/86Sr valueplus or minus two standard deviations (Price et al.2002), which is 87Sr/86Sr 5 0.7112–0.7116. In addition,we had previously observed higher 87Sr/86Sr for samplesderiving from the region surrounding the A€ır Massifsuggesting moving into the A€ır Massif would result inhigher radiogenic strontium isotope values (Stojanowskiand Knudson, 2011). However, we currently only havemodern faunal data from the A€ır region and it is, there-fore, most appropriate to consider only differencesbetween Gobero and the A€ır Massif using modern sam-ples. We note that the average modern faunal valuefrom the A€ır Massif (mean 87Sr/86Sr 5 0.71329 60.00089 [n 5 5, 1r]) is larger than the correspondingmodern faunal value from Gobero (87Sr/86Sr 5 0.71254);however, this difference is not statistically significant (P5 0.170) owing to the large standard deviation of thelatter reflecting the inclusion of highly variable rodentdata extracted from modern owl pellets (see discussionin Stojanowski and Knudson, 2011).
Paleomobility at Gobero during the middleHolocene
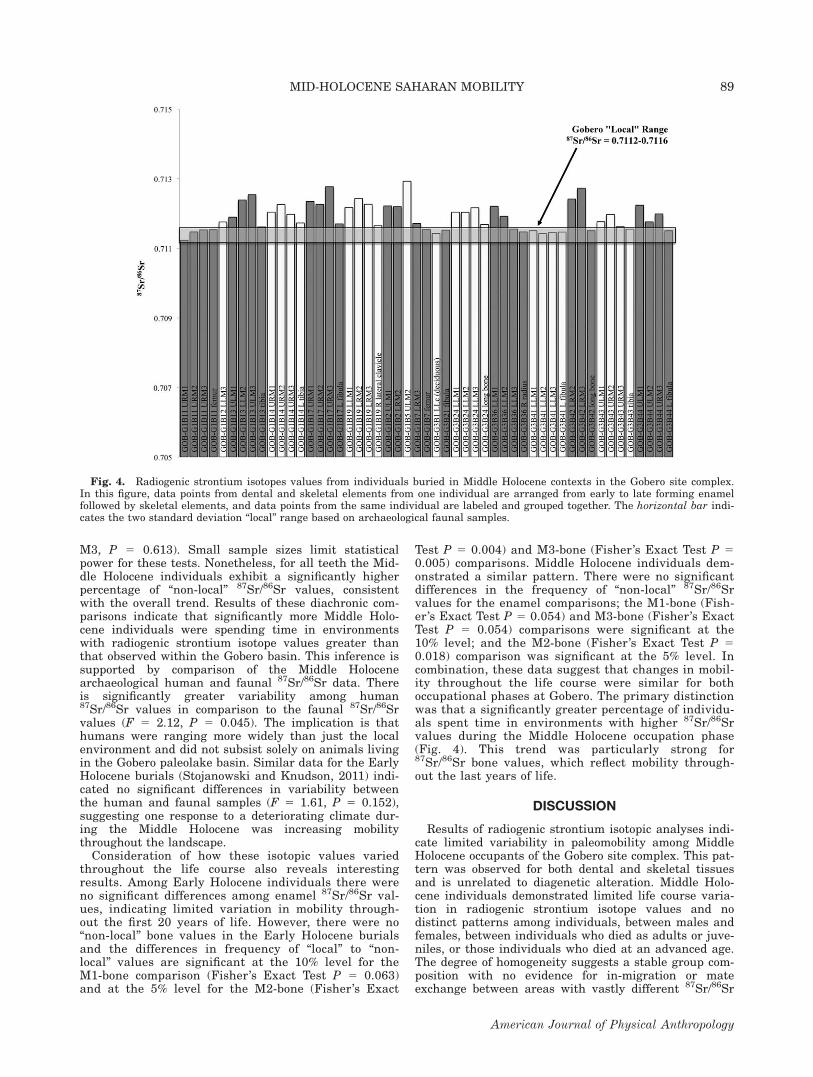
Results of radiogenic strontium isotope analyses ofarchaeological human tooth enamel and bone samplesfor Middle Holocene burials from Gobero are presentedin Tables 2 and 3 and Fig. 4. Mean archaeologicalhuman enamel and bone 87Sr/86Sr 5 0.71191 6 0.00040(1r, n 5 55) and ranged from87Sr/86Sr 5 0.71123to 87Sr/86Sr 5 0.71293. When separated, meanarchaeological human enamel 87Sr/86Sr 5 0.71202 6
0.00040 (1r, n 5 41) and mean archaeological humanbone 87Sr/86Sr 5 0.71157 6 0.00090 (1r, n 5 14). Thesevalues are significantly different from each other (P <0.0001), with the enamel data (87Sr/86Sr �0.712) closerto the soil values than the bone data (87Sr/86Sr �0.711).This result is not what would be expected if diagenesiswere affecting the bone values preferentially, and thedifference in the third decimal place likely reflectsanthropologically relevant data, as discussed below. Aswith our previous research at Gobero (Stojanowski andKnudson, 2011), bone data exhibit an order of magnitudeless variability than enamel data (bone 87Sr/86Sr sd 50.00009; tooth 87Sr/86Sr sd 5 0.00042), which reflects thelonger period of life that is averaged within the bulkbone data in comparison to the short crown formationtimes of human enamel (Reid and Dean, 2006).
There are no significant differences in 87Sr/86Sr valuesamong adult males, females, and juveniles (ANOVAP-values: M1 5 0.947, M2 5 0.933, M3 5 0.664, bone 50.374). These results do not change when only 87Sr/86Srvalues from adult males and females are compared (T-test P-values: M1 5 0.848, M2 5 0.907, M3 5 0.850,bone 5 0.428). Males and females show no significantdifferences in the average tooth-to-bone spacing (P 50.231), M1-to-M2 spacing (P 5 0.433), M3-to-bone spac-ing (P 5 0.552) or overall standard deviation among87Sr/86Sr values (P 5 0.289). There is no evidence forsex-specific patterning of life course variability in theMiddle Holocene burials at Gobero. Another measure oflife course variability is the correlation between observedage-at-death (indexed on M1 wear score) and tooth-to-bone isotope spacing. If individuals accumulated stron-tium from a greater variety of sources as they aged,then there should be a positive correlation betweenthese values. This correlation was positive, as expected,but not significantly different from zero (r 5 0.206, P 50.568) owing to the small sample size. When juvenilesand adults are considered separately, the adult correla-tion increases (r 5 0.433); however with a sample ofonly 10 individuals the P-value was not significant (P 50.211). These results suggest limited variation in mobil-ity patterns among Middle Holocene individuals, no evi-dence for extra-local in-migration from areas withdifferent underlying radiogenic strontium isotope values,and similar life course experiences for most Middle Holo-cene individuals at Gobero, regardless of age-at-death orsex.
However, there are statistically significant differenceswhen comparing radiogenic strontium isotope data fromEarly and Middle Holocene contexts. Although Earlyand Middle Holocene burials demonstrated little intra-and inter-individual variability, subtle differences doemerge when data are compared diachronically (Figs. 3and 4). First, bone 87Sr/86Sr values are significantly dif-ferent between the Early and Middle Holocene phases(P < 0.001). This reflects the extremely low standarddeviations; however, the average values differ only inthe fourth decimal place and are likely not anthropologi-cally relevant. Second, temporal differences in averageenamel 87Sr/86Sr values were also significantly different(P 5 0.009), with Middle Holocene individuals present-ing higher mean 87Sr/86Sr values in the third decimalplace (Middle Holocene mean 87Sr/86Sr 5 0.7120; EarlyHolocene mean 87Sr/86Sr 5 0.7117). Therefore, we arguethese differences reflect anthropologically relevant val-ues. Tooth-specific comparisons were significant at the10% level (M1 P 5 0.059, M2 P 5 0.067, M3 P 5 0.089).
86 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology
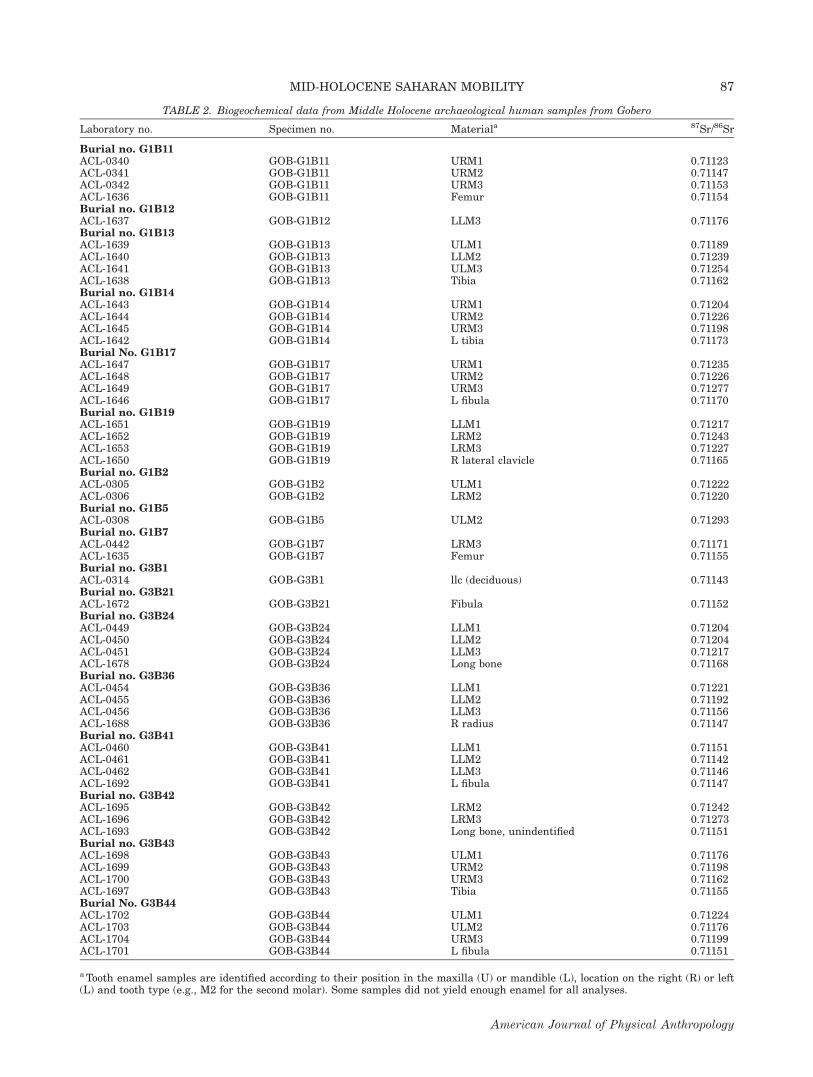
TABLE 2. Biogeochemical data from Middle Holocene archaeological human samples from Gobero
Laboratory no. Specimen no. Materiala 87Sr/86Sr
Burial no. G1B11ACL-0340 GOB-G1B11 URM1 0.71123ACL-0341 GOB-G1B11 URM2 0.71147ACL-0342 GOB-G1B11 URM3 0.71153ACL-1636 GOB-G1B11 Femur 0.71154Burial no. G1B12ACL-1637 GOB-G1B12 LLM3 0.71176Burial no. G1B13ACL-1639 GOB-G1B13 ULM1 0.71189ACL-1640 GOB-G1B13 LLM2 0.71239ACL-1641 GOB-G1B13 ULM3 0.71254ACL-1638 GOB-G1B13 Tibia 0.71162Burial no. G1B14ACL-1643 GOB-G1B14 URM1 0.71204ACL-1644 GOB-G1B14 URM2 0.71226ACL-1645 GOB-G1B14 URM3 0.71198ACL-1642 GOB-G1B14 L tibia 0.71173Burial No. G1B17ACL-1647 GOB-G1B17 URM1 0.71235ACL-1648 GOB-G1B17 URM2 0.71226ACL-1649 GOB-G1B17 URM3 0.71277ACL-1646 GOB-G1B17 L fibula 0.71170Burial no. G1B19ACL-1651 GOB-G1B19 LLM1 0.71217ACL-1652 GOB-G1B19 LRM2 0.71243ACL-1653 GOB-G1B19 LRM3 0.71227ACL-1650 GOB-G1B19 R lateral clavicle 0.71165Burial no. G1B2ACL-0305 GOB-G1B2 ULM1 0.71222ACL-0306 GOB-G1B2 LRM2 0.71220Burial no. G1B5ACL-0308 GOB-G1B5 ULM2 0.71293Burial no. G1B7ACL-0442 GOB-G1B7 LRM3 0.71171ACL-1635 GOB-G1B7 Femur 0.71155Burial no. G3B1ACL-0314 GOB-G3B1 llc (deciduous) 0.71143Burial no. G3B21ACL-1672 GOB-G3B21 Fibula 0.71152Burial no. G3B24ACL-0449 GOB-G3B24 LLM1 0.71204ACL-0450 GOB-G3B24 LLM2 0.71204ACL-0451 GOB-G3B24 LLM3 0.71217ACL-1678 GOB-G3B24 Long bone 0.71168Burial no. G3B36ACL-0454 GOB-G3B36 LLM1 0.71221ACL-0455 GOB-G3B36 LLM2 0.71192ACL-0456 GOB-G3B36 LLM3 0.71156ACL-1688 GOB-G3B36 R radius 0.71147Burial no. G3B41ACL-0460 GOB-G3B41 LLM1 0.71151ACL-0461 GOB-G3B41 LLM2 0.71142ACL-0462 GOB-G3B41 LLM3 0.71146ACL-1692 GOB-G3B41 L fibula 0.71147Burial no. G3B42ACL-1695 GOB-G3B42 LRM2 0.71242ACL-1696 GOB-G3B42 LRM3 0.71273ACL-1693 GOB-G3B42 Long bone, unindentified 0.71151Burial no. G3B43ACL-1698 GOB-G3B43 ULM1 0.71176ACL-1699 GOB-G3B43 URM2 0.71198ACL-1700 GOB-G3B43 URM3 0.71162ACL-1697 GOB-G3B43 Tibia 0.71155Burial No. G3B44ACL-1702 GOB-G3B44 ULM1 0.71224ACL-1703 GOB-G3B44 ULM2 0.71176ACL-1704 GOB-G3B44 URM3 0.71199ACL-1701 GOB-G3B44 L fibula 0.71151
a Tooth enamel samples are identified according to their position in the maxilla (U) or mandible (L), location on the right (R) or left(L) and tooth type (e.g., M2 for the second molar). Some samples did not yield enough enamel for all analyses.
MID-HOLOCENE SAHARAN MOBILITY 87
American Journal of Physical Anthropology
Small samples sizes are clearly an issue and power waslimited. For all tooth comparisons, the Middle Holoceneindividuals exhibited higher mean 87Sr/86Sr values sug-gesting greater incorporation of bioavailable strontiumfrom regions with higher 87Sr/86Sr values. This is con-sistent with the expectations of greater movement intothe A€ır Massif, an observation supported by the presenceof greenstone tools provisionally sourced to its easternside (Sereno et al., 2008).
The clearest temporal difference presented in Table 3is that individuals from Middle Holocene contextsexhibit a far greater percentage of radiogenic strontiumisotope values of 87Sr/86Sr >0.712. However, for consis-tency with other published datasets and utilizing stand-ard practice in paleomobility studies (e.g., Price et al.,2002; Evans and Tatham, 2004; Bentley et al., 2004;Bentley, 2006; Evans et al., 2010), we considered thepercentage of individuals from each time period whose87Sr/86Sr values were beyond the upper limit (none werebelow the lower limit) of the “local” radiogenic strontiumisotope baseline defined by archaeological faunal
87Sr/86Sr values (>0.7116). When bone and enamel dataare combined, there was a significantly higher percent-age of Middle Holocene individuals that exhibited87Sr/86Sr values above the local baseline (Early Holo-cene: 12/32 5 0.375; Middle Holocene; 38/55 5 0.691;Fisher’s Exact Test P 5 0.007). While this difference isnot statistically significant when only enamel 87Sr/86Srvalues are considered (Early Holocene: 12/20 5 0.600;Middle Holocene; 32/41 5 0.805; Fisher’s Exact Test P 50.123), the pattern is identical in that the Middle Holo-cene has more “non-local” 87Sr/86Sr values. Considera-tion of these data divided by tooth type reveals that thesignificant differences are being driven by variation inthe bone values. For example, the difference betweenthe percentage of Early and Middle Holocene “non-local”87Sr/86Sr values for the M1 (Early Holocene: 3/6 50.500; Middle Holocene: 10/13 5 0.769), M2 (Early Holo-cene: 5/8 5 0.625; Middle Holocene: 12/14 5 0.857),and M3 (Early Holocene: 4/6 5 0.666; Middle Holocene:11/14 5 0.786) were not significantly different (Fisher’sExact Test P-values are: M1, P 5 0.329; M2, P 5 0.309;
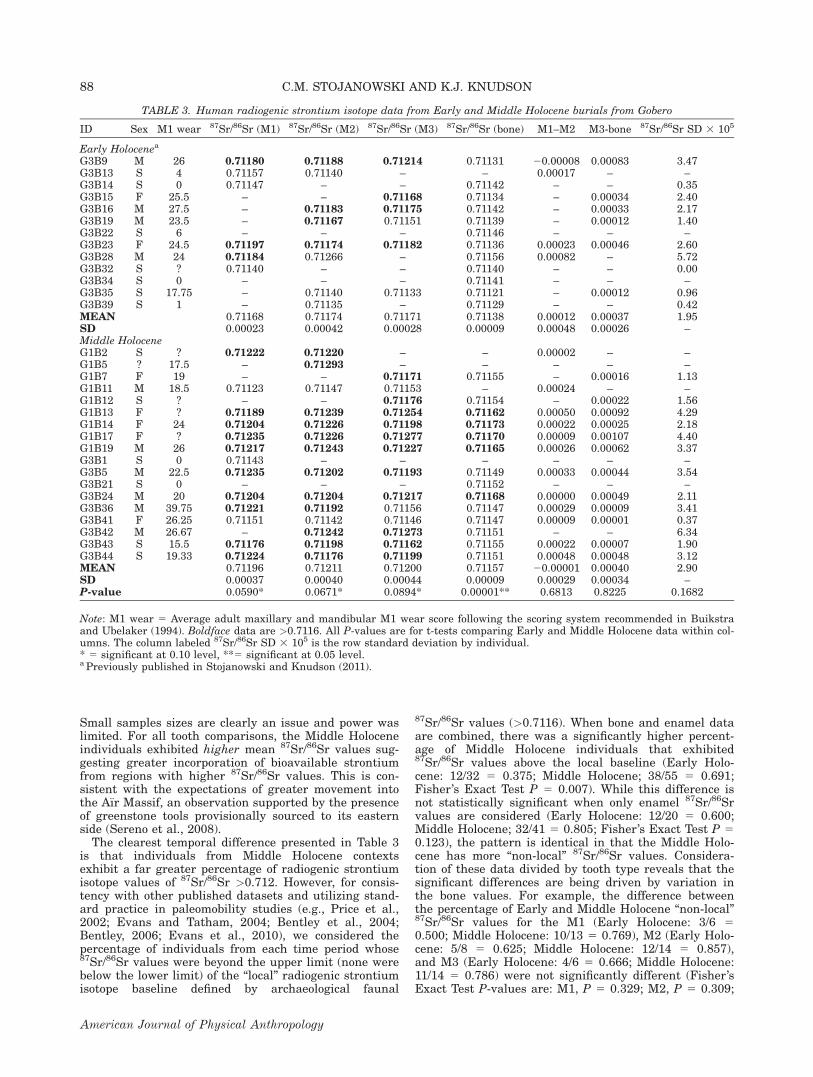
TABLE 3. Human radiogenic strontium isotope data from Early and Middle Holocene burials from Gobero
ID Sex M1 wear 87Sr/86Sr (M1) 87Sr/86Sr (M2) 87Sr/86Sr (M3) 87Sr/86Sr (bone) M1–M2 M3-bone 87Sr/86Sr SD 3 105
Early Holocenea
G3B9 M 26 0.71180 0.71188 0.71214 0.71131 20.00008 0.00083 3.47G3B13 S 4 0.71157 0.71140 – – 0.00017 – –G3B14 S 0 0.71147 – – 0.71142 – – 0.35G3B15 F 25.5 – – 0.71168 0.71134 – 0.00034 2.40G3B16 M 27.5 – 0.71183 0.71175 0.71142 – 0.00033 2.17G3B19 M 23.5 – 0.71167 0.71151 0.71139 – 0.00012 1.40G3B22 S 6 – – – 0.71146 – – –G3B23 F 24.5 0.71197 0.71174 0.71182 0.71136 0.00023 0.00046 2.60G3B28 M 24 0.71184 0.71266 – 0.71156 0.00082 – 5.72G3B32 S ? 0.71140 – – 0.71140 – – 0.00G3B34 S 0 – – – 0.71141 – – –G3B35 S 17.75 – 0.71140 0.71133 0.71121 – 0.00012 0.96G3B39 S 1 – 0.71135 – 0.71129 – – 0.42MEAN 0.71168 0.71174 0.71171 0.71138 0.00012 0.00037 1.95SD 0.00023 0.00042 0.00028 0.00009 0.00048 0.00026 –Middle HoloceneG1B2 S ? 0.71222 0.71220 – – 0.00002 – –G1B5 ? 17.5 – 0.71293 – – – – –G1B7 F 19 – – 0.71171 0.71155 – 0.00016 1.13G1B11 M 18.5 0.71123 0.71147 0.71153 – 0.00024 – –G1B12 S ? – – 0.71176 0.71154 – 0.00022 1.56G1B13 F ? 0.71189 0.71239 0.71254 0.71162 0.00050 0.00092 4.29G1B14 F 24 0.71204 0.71226 0.71198 0.71173 0.00022 0.00025 2.18G1B17 F ? 0.71235 0.71226 0.71277 0.71170 0.00009 0.00107 4.40G1B19 M 26 0.71217 0.71243 0.71227 0.71165 0.00026 0.00062 3.37G3B1 S 0 0.71143 – – – – – –G3B5 M 22.5 0.71235 0.71202 0.71193 0.71149 0.00033 0.00044 3.54G3B21 S 0 – – – 0.71152 – – –G3B24 M 20 0.71204 0.71204 0.71217 0.71168 0.00000 0.00049 2.11G3B36 M 39.75 0.71221 0.71192 0.71156 0.71147 0.00029 0.00009 3.41G3B41 F 26.25 0.71151 0.71142 0.71146 0.71147 0.00009 0.00001 0.37G3B42 M 26.67 – 0.71242 0.71273 0.71151 – – 6.34G3B43 S 15.5 0.71176 0.71198 0.71162 0.71155 0.00022 0.00007 1.90G3B44 S 19.33 0.71224 0.71176 0.71199 0.71151 0.00048 0.00048 3.12MEAN 0.71196 0.71211 0.71200 0.71157 20.00001 0.00040 2.90SD 0.00037 0.00040 0.00044 0.00009 0.00029 0.00034 –P-value 0.0590* 0.0671* 0.0894* 0.00001** 0.6813 0.8225 0.1682
Note: M1 wear 5 Average adult maxillary and mandibular M1 wear score following the scoring system recommended in Buikstraand Ubelaker (1994). Boldface data are >0.7116. All P-values are for t-tests comparing Early and Middle Holocene data within col-umns. The column labeled 87Sr/86Sr SD 3 105 is the row standard deviation by individual.* 5 significant at 0.10 level, **5 significant at 0.05 level.a Previously published in Stojanowski and Knudson (2011).
88 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology
M3, P 5 0.613). Small sample sizes limit statisticalpower for these tests. Nonetheless, for all teeth the Mid-dle Holocene individuals exhibit a significantly higherpercentage of “non-local” 87Sr/86Sr values, consistentwith the overall trend. Results of these diachronic com-parisons indicate that significantly more Middle Holo-cene individuals were spending time in environmentswith radiogenic strontium isotope values greater thanthat observed within the Gobero basin. This inference issupported by comparison of the Middle Holocenearchaeological human and faunal 87Sr/86Sr data. Thereis significantly greater variability among human87Sr/86Sr values in comparison to the faunal 87Sr/86Srvalues (F 5 2.12, P 5 0.045). The implication is thathumans were ranging more widely than just the localenvironment and did not subsist solely on animals livingin the Gobero paleolake basin. Similar data for the EarlyHolocene burials (Stojanowski and Knudson, 2011) indi-cated no significant differences in variability betweenthe human and faunal samples (F 5 1.61, P 5 0.152),suggesting one response to a deteriorating climate dur-ing the Middle Holocene was increasing mobilitythroughout the landscape.
Consideration of how these isotopic values variedthroughout the life course also reveals interestingresults. Among Early Holocene individuals there wereno significant differences among enamel 87Sr/86Sr val-ues, indicating limited variation in mobility through-out the first 20 years of life. However, there were no“non-local” bone values in the Early Holocene burialsand the differences in frequency of “local” to “non-local” values are significant at the 10% level for theM1-bone comparison (Fisher’s Exact Test P 5 0.063)and at the 5% level for the M2-bone (Fisher’s Exact
Test P 5 0.004) and M3-bone (Fisher’s Exact Test P 50.005) comparisons. Middle Holocene individuals dem-onstrated a similar pattern. There were no significantdifferences in the frequency of “non-local” 87Sr/86Srvalues for the enamel comparisons; the M1-bone (Fish-er’s Exact Test P 5 0.054) and M3-bone (Fisher’s ExactTest P 5 0.054) comparisons were significant at the10% level; and the M2-bone (Fisher’s Exact Test P 50.018) comparison was significant at the 5% level. Incombination, these data suggest that changes in mobil-ity throughout the life course were similar for bothoccupational phases at Gobero. The primary distinctionwas that a significantly greater percentage of individu-als spent time in environments with higher 87Sr/86Srvalues during the Middle Holocene occupation phase(Fig. 4). This trend was particularly strong for87Sr/86Sr bone values, which reflect mobility through-out the last years of life.
DISCUSSION
Results of radiogenic strontium isotopic analyses indi-cate limited variability in paleomobility among MiddleHolocene occupants of the Gobero site complex. This pat-tern was observed for both dental and skeletal tissuesand is unrelated to diagenetic alteration. Middle Holo-cene individuals demonstrated limited life course varia-tion in radiogenic strontium isotope values and nodistinct patterns among individuals, between males andfemales, between individuals who died as adults or juve-niles, or those individuals who died at an advanced age.The degree of homogeneity suggests a stable group com-position with no evidence for in-migration or mateexchange between areas with vastly different 87Sr/86Sr
Fig. 4. Radiogenic strontium isotopes values from individuals buried in Middle Holocene contexts in the Gobero site complex.In this figure, data points from dental and skeletal elements from one individual are arranged from early to late forming enamelfollowed by skeletal elements, and data points from the same individual are labeled and grouped together. The horizontal bar indi-cates the two standard deviation “local” range based on archaeological faunal samples.
MID-HOLOCENE SAHARAN MOBILITY 89
American Journal of Physical Anthropology

values. Using enamel data values as a proxy for pre-marital residence, juveniles did not appear to have livedextended periods of time in regions with higher or lowerradiogenic strontium isotope values. At first glance,then, the radiogenic strontium isotopic data from theMiddle Holocene occupation at Gobero suggest a remark-able degree of homogeneity.
However, careful consideration of the patterning ofvariation does reveal differences in mobility patterningbetween occupation phases. We had previously inter-preted the consistent M1, M2, M3 and bone values of87Sr/86Sr �0.711 among Early Holocene individuals asreflecting a “local” population with little evidence formovement into the A€ır Massif where 87Sr/86Sr valuesare expected to be higher (Stojanowski and Knudson,2011), or further north to the Libyan Fezzan or west andsouth toward Lake Chad where the 87Sr/86Sr values areexpected to be lower (87Sr/86Sr �0.710). New archaeolog-ical faunal baseline data presented here modify thisinterpretation. Therefore, prior to considering temporaldifferences in paleomobility it is useful to further refineour previous interpretations of the Early Holocene data(Stojanowski and Knudson, 2011).
Using the archaeological faunal data as the baseline,some Early Holocene enamel values do fall beyond thetwo standard deviation range based on archaeologicalfauna, while all bone data fall within the “local” range(see Table 2). However, “non-local” enamel values arenot randomly distributed among individuals. Rather, all“non-local” radiogenic strontium isotope enamel valuesare found in individuals who died as adults, whereasindividuals who died as juveniles (<15 years of age)exhibited radiogenic strontium isotope values that fellwithin the two standard range of the local baseline. Thispattern is quite stark, and statistically significant (t-testcomparing values, P < 0.001; Fisher’s Exact Test com-paring frequency of “local”: “non-local” values, P <0.001). In addition, individuals who died as juvenilesexhibited less overall variation among radiogenic stron-tium isotope values when measured using the standarddeviation of individual values (P 5 0.007), an interpreta-tion also supported by consideration of the range (seeTable 3). Individuals that died as juveniles exhibit exclu-sively “local” radiogenic strontium isotope values andless overall variability in life course mobility experience.The patterning is interesting but difficult to interpretwith confidence. Such a pattern is consistent with thepresence of first generation migrants to the region;adults with “non-local” enamel values were born in areaswith different underlying bedrock geology but migratedto the Gobero basin and remained there for the remain-der of their lives, thus establishing the “local” bone val-ues for adults. Juveniles in the Early Holocene samplemay have been those individuals who were born anddied within the “local” Gobero environment and, there-fore, present a homogenous series of radiogenic stron-tium isotope values indicating sedentism within thepaleolake basin. Other possible explanations of this pat-tern are also difficult to confirm. For example, given therelationship between childhood frailty and early mortal-ity, the “local” radiogenic strontium isotope enamel andbone values for juveniles may reflect heterogeneity offrailty that impacted patterns of individual mobility.Those individuals who died as juveniles may haveremained comparatively sedentary with respect to theircontemporaries due to issues with poor health andfrailty. That juveniles during the Early Holocene suf-
fered poor health is suggested by the high frequency oflocalized deciduous canine hypoplasias, which are die-tary stress indicators that were significantly less fre-quent among Middle Holocene juveniles from Gobero(Stojanowski and Carver, 2011). Finally, another possi-bility is that increased mobility was reflective ofenhanced survivorship; individuals that responded to cli-mate and subsistence challenges with increasing mobil-ity lived longer into adulthood.
Initial comparison of the Early and Middle Holocenedata suggests very similar early life experiences with amix of “local” and “non-local” enamel values and bonevalues that exhibit a “local” modal value. There wereno significant temporal differences for enamel values,while significantly different bone values were only inthe fourth decimal place and likely not relevant withrespect to underlying variation in bedrock geology.Nonetheless, the Middle Holocene dataset does reveal atendency toward a greater number of “non-local” radio-genic strontium isotope values. For all comparisons(M1, M2, M3, bone), there were more “non-local” valuesfor Middle Holocene individuals, which suggests atrend toward greater mobility through time. This pat-tern was particularly evident for the radiogenic stron-tium isotope bone data for which the Middle Holoceneoccupation phase exhibits a significantly higher fre-quency of “non-local” radiogenic strontium values.Therefore, one interpretation of these data is thathuman movement across the landscape increased dur-ing the Middle Holocene in response to increasingaridity.
Consideration of variability within the Middle Holo-cene data and throughout the lifecourse qualifies thisinterpretation. For example, some Middle Holocene indi-viduals exhibit enamel and bone radiogenic strontiumisotope values and patterns that are identical to EarlyHolocene individuals (i.e., “local” bone values with a mixof “local” and “non-local” enamel values). The trulyanomalous results with respect to this entire datasetare the five Middle Holocene individuals that demon-strate “non-local” radiogenic strontium isotope bone val-ues. All five individuals were adults when they died.Four of five individuals were buried on site G1 (see Fig.1). What is perhaps most interesting is that all five indi-viduals exhibited radiogenic strontium isotope enamelvalues that were, without exception, also beyond the“local” range of variation. Our interpretation of thesedata is that there were some individuals at Gobero dur-ing the Middle Holocene period that maintained a com-paratively mobile existence throughout the lifecourse,while others exhibited a more sedentary pattern thatwas similar to that observed for all Early Holocene indi-viduals. This suggests different individuals utilized dif-ferent mobility strategies during the Middle Holoceneoccupation of Gobero. That four of five of these individu-als were buried in cemetery G1 in the Gobero site com-plex may further explain this pattern as a temporal one.Although all individuals were buried during the MiddleHolocene, there was considerable temporal variabilityrepresented in this dataset. In particular, Stojanowski(2013) suggests, based on taphonomic and stratigraphicdata, that site G1 was used in the latter part of theMiddle Holocene occupational phase. This is supportedby radiocarbon dates (Stojanowski, 2013). Excluding thepartially fossilized G1B11, the light stained burialsrecovered from site G3 returned an average calibrateddate BP of 5,632 years (n 5 5), while burials from site
90 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology

G1 returned an average calibrated data BP of4,971years (n 5 5) (see Stojanowski, 2013; Table 5.3).This difference is statistically significant (P 5 0.0008).This could indicate that increasing mobility was adoptedonly in the terminal stages of Gobero’s occupation whenthe lake levels were their lowest and more ephemeral.Another interpretation of these data is that the MiddleHolocene was characterized by individuals practicingdistinct forms of subsistence that modified their rela-tionship to the landscape. Whether the more mobileindividuals were hunter–gatherers or pastoralists isimpossible to know using these data alone; however, theoverall interpretation is that climatic deterioration andincreasing aridity was accompanied by subtle shifts inmobility that may only have been adopted by the termi-nal occupants of the region immediately prior to thefinal abandonment of the area.
In closing, we note the importance of generatingadditional baseline data for radiogenic strontium iso-tope analyses in the central and southern Sahara.Although we have come a long way in the last sevenyears when the first human biogenic data were pub-lished (Tafuri et al., 2006), we currently lack sufficientdata to parse patterns of mobility throughout the ChadBasin or even into and throughout the A€ır Massif. Forexample, geological data suggests a higher 87Sr/86Srvalue for the A€ır Massif compared with Gobero andlower 87Sr/86Sr values for regions closer to the center ofthe Chad Basin. This interpretation is supported bythe limited modern faunal data but these data are sig-nificantly higher than archaeological faunal data sug-gesting the effects of modern dust transport onmeasured values. The significantly lower 87Sr/86Srvalue for the Libyan Fezzan (see Stojanowski andKnudson, 2011) suggests humans were not perma-nently migrating between Gobero and the Fezzan in away that would affect enamel values for lower ageranges. This, in itself, is noteworthy. Data from Libyaand Niger are not of the same scale; the latter presentdata on life course mobility while the former provideless fine-grain inferences about individual paleomobil-ity but can be interpreted within an archaeologicalresearch program in which the relationship betweenhumans and their landscape is more holistic andregional in focus (di Lernia et al., 2013). However, theimpression of Holocene Saharan peoples is not one ofwidespread nomadism, but one of regional adaptationto local conditions, which varied as much then asSaharan microclimates do today. Just as not all EarlyHolocene foragers accurately fall within the rubric ofSutton’s (1974, 1977) “aqualithic” (see Holl, 2005), Mid-dle Holocene groups are not easily subsumed within asimple subsistence-based model of biocultural adapta-tion that emphasizes a shift to pastoralism. At Gobero,the near permanent existence of the paleolake mayhave obviated a shift to pastoralism, or at least cur-tailed its relevance to the individuals who lived there.It is only after Gobero’s waters disappeared that wesee evidence for cattle pastoralism as an importantaspect of ideology and group identity in the southernSahara. At sites such as Chin Tafidet (�3,300–4,500bp), In Tuduf (�3,400–4,400 bp), Afunfun (�3,500–4,100 bp), and Iwelen (�2,500–5,000 bp), cattle (Bos)are buried with and alongside humans (Lhote, 1976;Paris, 1996, 1997, 2000), suggesting a diminishing dis-tinction between human and animal as a pastoral ide-ology took hold in the desiccating Sahara.
ACKNOWLEDGMENTS
We would like to thank members of the Gobero exca-vations and subsequent analyses for invaluable collabo-rations. In the Archaeological Chemistry Laboratory,Niamh Daly, Meridith Masoner, Katie Miller, andKathleen Paul performed invaluable laboratory assis-tance in sample preparation. In the W.M. Keck Founda-tion Laboratory for Environmental Biogeochemistry, weare extremely grateful for the expertise of Dr. GwynethGordon and laboratory access granted by Dr. ArielAnbar. Charisse Carver provided valuable assistancewith the construction of the figures. Preliminary inter-pretations of some of these data were presented at the19th Biennial Meeting of the Society of AfricanistArchaeologists in Frankfurt, Germany on September 8–11, 2008 in a presentation entitled, “Recent Bioarchaeo-logical and Biogeochemical Research at Gobero: Paleo-diet and Residential Mobility in the Early and MiddleHolocene” by Kelly J. Knudson and Christopher M. Sto-janowski; we thank the audience members, particularlyDr. Judith Sealy, for valuable input.
LITERATURE CITED
Aberg G. 1995. The use of natural strontium isotopes as tracersin environmental studies. Water Air Soil Poll 79:309–322.
Alley RB, Mayewski PA, Sowers T, Stuiver M, Taylor KC, ClarkPU. 1997. Holocene climatic instability: a prominent, wide-spread event 8200 years ago. Geology 25:483–486.
Begg GC, Griffin WL, Natapov LM, O0Reilly SY, Grand SP,O0Neill CJ, Hronsky JMA, Poudjom Djomani Y, Swain CJ,Deen T, Bowden P. 2009. The lithospheric architecture ofAfrica: seismic tomography, mantle petrology, and tectonicevolution. Geosphere 5:23–50.
Bentley RA. 2006. Strontium isotopes from the earth to thearchaeological skeleton: a review. J Arch Meth Theory 13:135–187.
Bentley RA, Price TD, Stephan E. 2004. Determining the “local”87Sr/86Sr range for archaeological skeletons: a case studyfrom Neolithic Europe. J Archaeol Sci 31:365–375.
Biagetti S, di Lernia S. 2007. Reflections on the Takarkori rock-shelter (Fezzan, Libyan Sahara). In: Kornfield M, Vasil’ev S,Miotti L, editors. On shelter’s ledge: histories, theories andmethods of rockshelter research. Oxford: Archaeopress BARS1655. p 125–132.
Biagetti S, di Lernia S. 2013. Holocene deposits of Saharan rockshelters: the case of Takarkori and other sites from theTadrart Acacus mountains (southwest Libya). Afr ArchaeolRev 30:305–338.
Black R, Latouche L, Li�egeois JP, Caby R, Bertrand JM. 1994.Pan-African displaced terranes in the Tuareg Shield (centralSahara). Geology 22:641–644.
Bouchette F, Schuster M, Ghienne J-F, Denamiel C, Roquin C,Moussa A, Marsaleix P, Duringer P. 2010. Hydrodynamics inHolocene Lake Mega-Chad. Quat Res 73:226–236.
Brown WL, Moreau C, Demaiffe D. 1989. An anorthosite suitein a ring-complex: crystallization and emplacement of ananorogenic type from Abontorok, A€ır, Niger. J Petrol 30:1501–1540.
Buikstra JE, Ubelaker DH. 1994. Standards for data collectionfrom human skeletal remains: proceedings of a seminar at theField Museum of Natural History. Research Series, no. 44.Fayetteville: Arkansas Archaeological Report Research Series.
Burke K. 1976. The Chad Basin: an active intra-continentalbasin. Tectonophysics 36:197–206.
Buzon MR, Simonetti A. 2013. Strontium isotope (87Sr/86Sr)variability in the Nile Valley: identifying residential mobilityduring ancient Egyptian and Nubian sociopolitical changes inthe New Kingdom and Napatan period. Am J Phys Anthropol151:1–9.
MID-HOLOCENE SAHARAN MOBILITY 91
American Journal of Physical Anthropology

Buzon MR, Simonetti A, Creaser RA. 2007. Migration in theNile Valley during the New Kingdom period: a preliminarystrontium isotope study. J Arch Sci 34:1391–1401.
Clark JD, Agrilla EJ, Crader DC, Galloway A, Garcea EAA,Gifford-Gonzalez D, Hall DN, Smith AB, Williams MAJ. 2008.Adrar Bous. Archaeology of a central Saharan granitic ringcomplex in Niger. Tervuren: Royal Museum for Central Africa.
Cratchley CR, Louis P, Ajakaiye DE. 1984. Geophysical and geo-logical evidence for the Benue-Chad Basin Cretaceous rift val-ley system and its tectonic implications. J Afr Earth Sci 2:141–150.
Cremaschi M, Zerboni A. 2011. Human communities in a dryinglandscape: holocene climate change and cultural response inthe central Sahara. In: Martini IP, Chesworth W, editors.Landscape and societies: selected cases. New York: Springer.p 67–89.
Demaiffe D, Moreau C, Brown WL, Weis D. 1991. Geochemicaland isotopic (Sr, Nd and Pb) evidence on the origin of theanorthosite-bearing anorogenic complexes of the A€ır province,Niger. Earth Planet Sci Lett 105:28–46.
deMenocal P, Ortiz J, Guilderson T, Adkins J, Sarnthein M,Baker L, Yarusinky M. 2000. Abrupt onset and termination ofthe African Humid Period: rapid climate responses to gradualinsolation forcing. Quat Sci Rev 19:347–361.
di Lernia S, Tafuri MA. 2013. Persistent deathplaces andmobile landmarks: the Holocene mortuary and isotopic recordfrom Wadi Takarkori (SW Libya). J Anthropol Archaeol 32:1–15.
di Lernia S, Tafuri MA, Gallinaro M, Alhaique F, Balasse M,Cavorsi L, Fullager PD, Mercuri AM, Monaco A, Perego A,Zerboni A. 2013. Inside the “African Cattle Complex”: animalburials in the Holocene central Sahara. PLoS One 8(2):1–28.
Drake N, Bristow C. 2006. Shorelines in the Sahara: geomor-phological evidence for an enhanced monsoon from palaeolakeMegchad. Holocene 16:901–911.
Drake NA, Blench RM, Armitage SJ, Bristow CS, White KH.2011. Ancient watercourses and biogeography of the Saharaexplain the peopling of the desert. Proc Nat Acad Sci USA108:458–462.
Dupuy C, Dostal J, Chikhaoui M. 1993. Trace element and iso-topic geochemistry of Cenozoic alkali basaltic lavas from Ata-kor (central Sahara). Geochem J 27:131–145.
�Echallier J-C, Roset J-P. 1986. La c�eramique des gisements deTagalagal et de l0Adrar Bous 10 (Air, R�epublique du Niger).Cah Sci Hum 22:151–158.
Ericson JE. 1985. Strontium isotope characterization in thestudy of prehistoric human ecology. J Hum Evol 14:503–514.
Evans JA, Montgomery J, Wildman G, Boulton N. 2010. Spatialvariations in biosphere 87Sr/86Sr in Britain. J Geol Soc Lon-don 167:1–4.
Evans JA, Tatham S. 2004. Defining ‘local signature’ in termsof Sr isotope composition using a tenth-to twelfth-centuryAnglo-Saxon population living on a Jurassic clay-carbonateterrain, Rutland, UK. For Geosci 232:237–248.
Faure G, Powell JL. 1972. Strontium isotope geology. New York:Springer-Verlag.
Frumkin A, Stein M. 2004. The Sahara-east Mediterraneandust and climate connection revealed by strontium and ura-nium isotopes in a Jerusalem speleothem. Earth Planet SciLett 217:451–464.
Garcea EAA. 2013a. Gobero: the no-return frontier. Archaeologyand landscape at the Saharo-Sahelian borderland. Journal ofAfrican Archaeology Monograph Series 9. Frankfurt: AfricaMagna Verlag.
Garcea EAA. 2013b. The archaeological significance of Gobero.In: Garcea EAA, editor. Gobero: the no-return frontier.Archaeology and landscape at the Saharo-Sahelian border-land. Journal of African Archaeology Monograph Series 9.Frankfurt: Africa Magna Verlag. p 5–18.
Gasse F. 2000. Hydrological changes in the African tropics sincethe Last Glacial Maximum. Quat Sci Rev 19:189–211.
Gasse F. 2002. Diatom-inferred salinity and carbonate oxygenisotopes in Holocene waterbodies of the western Sahara andSahel (Africa). Quat Sci Rev 21:737–767.
Gasse F, Van Campo E. 1994. Abrupt post-glacial climate eventsin West Asia and North Africa monsoon domains. EarthPlanet Sci Lett 126:435–456.
Genik GJ. 1992. Regional framework, structural and petroleumaspects of rift basins in Niger, Chad and the Central AfricanRepublic (C.A.R.). Tectonophysics 213:169–185.
Giraudi C. 2013. Late Upper Pleistocene and Holocene stratig-raphy of the palaeolakes in the Gobero basin. In: GarceaEAA, editor. Gobero: the no-return frontier. Archaeology andlandscape at the Saharo-Sahelian borderland. Journal of Afri-can Archaeology Monograph Series 9. Frankfurt: AfricaMagna Verlag. p 67–80.
Giraudi C, Mercuri AM. 2013. Early to Middle Holocene envi-ronmental variations in the Gobero basin. In: Garcea EAA,editor. Gobero: the no-return frontier. Archaeology and land-scape at the Saharo-Sahelian borderland. Journal of AfricanArchaeology Monograph Series 9. Frankfurt: Africa MagnaVerlag. p 114–126.
Grousset FE, Rognon P, Coud�e-Gaussen G, P�edemay P. 1992.Origins of peri-Saharan dust deposits traced by their Nd andSr isotopic composition. Palaeogeogr Palaeocl Palaeoecol 93:203–212.
Grousset FE, Parra M, Bory A, Martinez P, Bertrand P,Shimmield G, Ellam RM. 1998. Saharan wind regimes tracedby the Sr-Nd isotopic composition of subtropical Atlantic sedi-ments: last glacial maximum vs today. Quat Sci Rev 17:395–409.
Gr€un R, Aubert M, Joannes-Boyau R, Moncel M-H. 2008. Highresolution analysis of uranium and thorium concentration aswell as U-series isotope distributions in a Neanderthal toothfrom Payre (Ardeche, France) using laser ablation ICP-MS.Geochim Cosmochim Acta 72:5278–5290.
Haour AC. 2003. One hundred years of archaeology in Niger.J World Prehist 17:181–234.
Hartley RW, Allen PA. 1994. Interior cratonic basins of Africa:relation to continental break-up and role of marine convec-tion. Basin Res 6:95–113.
Hassan F. 1997. Holocene Palaeoclimates of Africa. Afr ArchaeolRev 14:213–230.
Holl AFC. 2005. Holocene “aquatic” adaptations in north tropi-cal Africa. In: Stahl AB, editor. African archaeology. Malden,MA: Blackwell Publishing. p. 174–186.
Joubert G. Vaufrey R. 1941-46. Le n�eolithique du T�en�er�e.L0Anthropologie 50:325–330.
Kobashi T, Severinghaus JP, Brook EJ, Barnola J-M, GrachevAM. 2007. Precise timing and characterization of abrupt cli-mate change 8200 years ago from air trapped in polar ice.Quat Sci Rev 26:1212–1222.
Knudson KJ. 2007. La influencia de Tiwanaku en San Pedro deAtacama: una investigaci�on por los is�otopos del estroncio.Estudios Atacame~nos 33:7–24.
Knudson KJ, Price TD. 2007. Utility of multiple chemical tech-niques in archaeological residential mobility studies: casestudies from Tiwanaku- and Chiribaya-affiliated sites in theAndes. Am J Phys Anthropol 132:25–39.
Knudson KJ, Gardella KR, Yaeger J. 2012. ProvisioningInka feasts at Tiwanaku, Bolivia: the geographic originsof camelids in the Pumapunku Complex. J Arch Sci 39:479–491.
Krom MD, Cliff RA, Eijsink LM, Herut B, Chester R. 1999. Thecharacterisation of Saharan dusts and Nile particulate matterin surface sediments from the Levantine basin using Sr iso-topes. Marine Geol 155:319–330.
Kuper R, Kr€opelin S. 2006. Climate-controlled Holocene occupa-tion in the Sahara: motor of Africa’s evolution. Sci 313:803–807.
Lal D, Large WG, Walker SG. 2007. Climatic forcing before,during, and after the 8.2 Kyr B.P. global cooling event.J Earth Syst Sci 116:171–177.
Lee-Thorp J. 2002. Two decades of progress towards under-standing fossilization processes and isotopic signals in calci-fied tissue minerals. Archaeometry 44:435–446.
Lee-Thorp JA. 2008. On isotopes and old bones. Archaeometry50:925–950.
92 C.M. STOJANOWSKI AND K.J. KNUDSON
American Journal of Physical Anthropology

Lee-Thorp J, Sponheimer M. 2003. Three case studies used toreassess the reliability of fossil bone and enamel isotope sig-nals for paleodietary studies. J Anthropol Arch 22:208–216.
Leger JM. 1985. G�eologie et �evolution magmatique du complexeplutonique d’Iskou (A€ır, Niger). J Afr Earth Sci 3:89–96.
Lhote H. 1976. Vers d’autres Tassili. Nouvelles d�ecouvertes auSahaha. M�emoire du CRAP XXIX, Algeria. S.N.E.D.
Li�egeois JP, Black R, Navez J, Latouche L. 1994. Early and latePan-African orogenies in the A€ır assembly terranes (Tuaregshield, Niger). Precamb Res 67:59–88.
Mercuri AM, Massamba N’siala I, Florenzano A. 2013. Environ-mental and ethnobotanical data inferred from pollen ofGobero and the dried lakebeds in the surrounding area. In:Garcea EAA, editor. Gobero: the no-return frontier. Archaeol-ogy and landscape at the Saharo-Sahelian borderland. Jour-nal of African Archaeology Monograph Series 9. Frankfurt:Africa Magna Verlag. p 81–104.
Mercuri AM, Sadori L, Uzquiano Ollero P. 2011. Mediterraneanand north-African cultural adaptations to mid-Holocene envi-ronmental and climatic changes. Holocene 21:189–206.
Nelson BK, DeNiro MJ, Schoeninger MJ, DePaolo DJ, Hare PE.1986. Effects of diagenesis on strontium, carbon, nitrogen,and oxygen concentration and isotopic concentration of bone.Geochim Cosmochim Acta 50:1941–1949.
Paris F. 1996. Les s�epultures du Sahara Nig�erien duN�eolithique �a l’Islamisation: coutumes fun�eraires, chronolo-gie, civilisation. ORSTROM, Paris.
Paris F. 1997. Les inhumations de Bos au Sahara m�eridional auN�eolithique. Archaeozoologia 9:113–122.
Paris F. 2000. African livestock remains from Saharan mortu-ary contexts. In: Blench RM, Macdonald KC, editors. The ori-gins and development of African livestock: archaeology,genetics, linguistics, and ethnography. London: UniversityCollege London Press. p. 111–126.
Petters SW. 1991. Regional geology of Africa. New York:Springer-Verlag.
Price TD, Burton JH, Bentley RA. 2002. The characterization ofbiologically available strontium isotope ratios for the study ofprehistoric migration. Archaeometry 44:117–135.
Price TD, Johnson CM, Ezzo JA, Ericson J, Burton JH. 1994.Residential mobility in the prehistoric Southwest UnitedStates: a preliminary study using strontium isotope snalysis.J Arch Sci 21:315–330.
Price TD, Tiesler V, Burton JH. 2006. Early African diaspora incolonial Campeche, Mexico: strontium isotopic evidence. Am JPhys Anthropol 130:485–490.
Reid DJ, Dean MC. 2006. Variation in modern human enamelformation times. J Hum Evol 50:329–346.
Reygasse M. 1934. Le T�en�er�een. Observations sur un faciesnouveau di N�eolithique des confins alg�erosoudanais. In:Compte rendu de la session du X Congres pr�ehistorique deFrance. P�erigueux. p. 577–584.
Rognon P, Coud�e-Gaussen G, Revel M, Grousset FE, PedemayP. 1996. Holocene Saharan dust deposition on the Cape VerdeIslands: sedimentological and Nd-Sr isotopic evidence. Sedi-mentology 43:359–366.
Roset J-P. 1983. Nouvelles donn�ees sur le probleme de lan�eolithisation du Sahara m�eridional: A€ır et T�en�er�e, au Niger.Cah ORSTROM ser Geol 13:119–142.
Roset J-P. 1987. Paleoclimatic and cultural conditions of Neo-lithic development in the Early Holocene of northern Niger(Air and Tenere). In: Close AE, editor. Prehistory of aridnorth Africa. Dallas: Southern Methodist University Press.p 211–234.
Roset J-P, de Broin F, Faure M, Gayet M, Gu�erin C, Mouchet F.1990. La faune de Tin Ouaffadene et d0Adrar Bous 10, deuxgisements arch�eologiques de l0Holocene ancien au Niger nord-oriental. G�eodynamique 5:67–89.
Schaaf P, M€uller-Sohnius D. 2002. Strontium and neodymiumisotopic study of Libyan desert glass: inherited pan-Africanage signatures and new evidence for target material. Meteror-itics Planetary Sci 37:565–576.
Schl€uter T. 2006. Geological atlas of Africa. Berlin: Springer-Verlag.
Sereno PC, Garcea EAA, Jousse H, Stojanowski CM, SaliegeJ-F, Maga A, Ide OA, Knudson KJ, Mercuri AM, Stafford TW,Jr., Kaye TG, Giraudi C, Massamba N’siala I, Cocca E, MootsHM, Dutheil DB, Stivers JP. 2008. Lakeside cemeteries in theSahara: 5000 years of Holocene population and environmen-tal change. PLoS One 3(8):1–22.
Sillen A. 1989. Diagenesis of the inorganic phase of corticalbone. In: Price TD, editor. The chemistry of prehistorichuman bone. Cambridge: Cambridge University Press.p 211–228.
Sillen A, Sealy JC. 1995. Diagenesis of strontium in fossil bone:a reconsideration of Nelson et al. (1986). J Arch Sci 22:313–320.
Smith AB. 2008a. The Kiffian. In: Clark JD, Gifford-GonzalezD, editors. Adrar Bous. Archaeology of a central Saharan gra-nitic ring complex in Niger. Tervuren: Royal Museum for Cen-tral Africa. p 179–199.
Smith AB. 2008b. The Tenerian. In: Clark JD, Gifford-GonzalezD, editors. Adrar Bous. Archaeology of a central Saharan gra-nitic ring complex in Niger. Tervuren: Royal Museum for Cen-tral Africa. p 201–243.
Stojanowski CM. 2013. An archaeological perspective on theburial record at Gobero. In: Garcea EAA, editor. Gobero: Theno return frontier. Archaeology and landscape at the Saharo-Sahelian borderland. The Journal of African ArchaeologyMonograph series number 9. Frankfurt: Africa Magna Verlag.p 44–64.
Stojanowski CM, Carver CL. 2011. Inference of emergent cattlepastoralism in the southern Sahara desert based on localizedhypoplasia of the primary canine. Int J Paleopathol 1:89–97.
Stojanowski CM, Knudson KJ. 2011. Biogeochemical inferencesof mobility of Early Holocene fisher-foragers from the south-ern Sahara desert. Am J Phys Anthropol 146:49–61.
Sutton JEG. 1974. The aquatic civilization of middle Africa.J African Hist 15:527–546.
Sutton JEG. 1977. The African aqualithic. Antiquity 51:25–34.Tafuri MA, Bentley RA, Manzi G, and di Lernia S. 2006. Mobil-
ity and kinship in the prehistoric Sahara: strontium isotopeanalysis of Holocene human skeletons from the Acacus Mts.(Southwestern Libya). J Anthropol Arch 25:390–402.
Tixier J. 1962. Le T�en�er�en de l0Adrar Bous III. In: Hugot, H-J,editor. Missions Berliet T�en�er�e-Tchad. Arts et m�etiers graphi-ques. Paris. 333–348.
T€utken T, Vennemann TW, Pfretzschner HU. 2008. Early dia-genesis of bone and tooth apatite in fluvial and marine set-tings: constraints from combined oxygen isotope, nitrogen andREE analysis. Palaeogeogr Palaeocl Palaeoecol 266:254–268.
Weldeab S, Emeis K-C, Hemleben C, Siebel W. 2002. Prove-nance of lithogenic surface sediments and pathways of river-ine suspended matter in the Eastern Mediterranean Sea:evidence from 143Nd/144Nd and 87Sr/86Sr ratios. Chem Geol186:139–149.
Williams MAJ. 2008. Geology, geomorphology and prehistoricenvironments. In: Clark JD, Gifford-Gonzalez D, editors. AdrarBous. Archaeology of a central Saharan granitic ring complexin Niger. Tervuren: Royal Museum for Central Africa. p 25–54.
Williams CT, Marlowe CA. 1987. Uranium and thorium distri-butions in fossil bones from Olduvai Gorge, Tanzania andKanam, Kenya. J Arch Sci 14:297–309.
Wright JB, Hastings DA, Jones WB, Williams HR. 1985. Geol-ogy and mineral resources of West Africa. London: Allen andUnwin.
MID-HOLOCENE SAHARAN MOBILITY 93
American Journal of Physical Anthropology
Related Documents











