J. Insect Physiol. Vol. 39, No. 9, pp. 769-774, 1993 0022-1910/93 $6.00 + 0.00 Printed in Great Britain. All rights reserved Copyright Q 1993 Pergamon Press Ltd Changes in Responsiveness to Factors Inducing Diapause Termination in Bruchus rufimanus (Boh.) (Coleoptera: Bruchidae) B. TRAN, J. DARQUENNE,* J. HUIGNARD* Received 7 December 1992; revised 15 February 1993 Bruclrusrufimunus undergoes a reproductive diapause from the emergence of the adults, in September, until April, when the adults enter the fields of their plant host, at the beginning of the flowering period. In a previous paper, we pointed out that long days and the consumption of host plant pollen are the main environmental factors which are required for the termination of the diapause. We now analyse the modifications of ovariole structure and changes in the responsiveness of the bruchid to these environmental factors during the course of diapause. In constant conditions (lO°C, constant darkness) in which diapause is maintained, the structure of the ovarioles does not vary during the first 5 months of diapause, but, after 6-7 months, previtellogenic phases are observed in the basal oiicytes. In September, a very small percentage of adults which have not experienced the exposure to lO*C, constant darkness, undergoes diapause termination when they are exposed to long days and consume pollen. The percentage of adults in which diapause terminates in conditions normally leading to termination increases after a period of 3-4 months in the diapausing conditions. We propose that horotelic processes take place during this initial phase of diapause. Longer periods in diapause do not result in greater rates of diapause termination when insects are exposed to long days and host plant pollen. The second phase of diapause, from the 5th month onwards, is interpreted as a facultative one, during which the insects are ready to respond to the factors inducing both the morphological and physiological development. The variations in responsiveness are important in the beetle’s life history. Reproductive diapause Diapause development Diapause termination Bruchids Ovetwintering INTRODUCTION Bruchus rufimanus is a bruchid whose larvae develop in the seeds of the broad bean, Vicia faba (L.), and consume the reserves of the cotyledons. The species is univoltine. The adults are in reproductive diapause from September until April. They overwinter under the bark of trees or in lichens (Speyer, 195 1; Middlekauf, 1961; DuPont, unpublished) and return to the growing crop in the spring, once the beans begin to flower. At the time of colonization, the males have active reproductive organs: their diapause has already terminated. The females remain in diapause and their sexual development begins in the fields. Experimental studies have shown that reproductive diapause termination depends, particularly in the case of females, on interactions between two factors: photo- period (long days, 18 h light-6 h dark) and a factor associated with the plant host (the consumption of its *Institut de Biocenotique des Afrosyst6mes-URA CNRS 1298, Facultb des Sciences de Tours, Part de Grandmont, 37200 Tours, France. pollen). In the absence of food, the proportion of females in which diapause terminates is small, even in long-day conditions. For the great majority of males, long pho- tophases alone are sufficient to terminate diapause (Tran and Huignard, 1992). Throughout reproductive dia- pause, in temperate climate, insects live at low tempera- tures and do not experience the factors which stimulate reproduction (long days and a supply of food). In some species, it has been reported that there is a change in the physiological state of the insects during diapause development, which renders them more recep- tive to those factors which stimulate the termination of diapause. The form of diapause development in which particular cues, whether or not the same as those which induced diapause, are necessary for diapause termin- ation, was termed tachytely by Hodek (1981, 1983). Tachytely has been observed in several different species of bruchids, including B. pisorum (Pesho and Van Houten, 1982), Bruchidius atrolineatus (Lenga and Huig- nard, 1992) and B. rgfimanus (Tran and Huignard, 1992). In the case of these insects, various climatic and plant host-related factors are necessary for diapause termination. In other cases, the duration of diapause is apparently 769

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J. Insect Physiol. Vol. 39, No. 9, pp. 769-774, 1993 0022-1910/93 $6.00 + 0.00
Printed in Great Britain. All rights reserved Copyright Q 1993 Pergamon Press Ltd
Changes in Responsiveness to Factors Inducing Diapause Termination in Bruchus rufimanus (Boh.) (Coleoptera: Bruchidae) B. TRAN, J. DARQUENNE,* J. HUIGNARD*
Received 7 December 1992; revised 15 February 1993
Bruclrus rufimunus undergoes a reproductive diapause from the emergence of the adults, in September, until April, when the adults enter the fields of their plant host, at the beginning of the flowering period. In a previous paper, we pointed out that long days and the consumption of host plant pollen are the main environmental factors which are required for the termination of the diapause. We now analyse the modifications of ovariole structure and changes in the responsiveness of the bruchid to these environmental factors during the course of diapause. In constant conditions (lO°C, constant darkness) in which diapause is maintained, the structure of the ovarioles does not vary during the first 5 months of diapause, but, after 6-7 months, previtellogenic phases are observed in the basal oiicytes. In September, a very small percentage of adults which have not experienced the exposure to lO*C, constant darkness, undergoes diapause termination when they are exposed to long days and consume pollen. The percentage of adults in which diapause terminates in conditions normally leading to termination increases after a period of 3-4 months in the diapausing conditions. We propose that horotelic processes take place during this initial phase of diapause. Longer periods in diapause do not result in greater rates of diapause termination when insects are exposed to long days and host plant pollen. The second phase of diapause, from the 5th month onwards, is interpreted as a facultative one, during which the insects are ready to respond to the factors inducing both the morphological and physiological development. The variations in responsiveness are important in the beetle’s life history.
Reproductive diapause Diapause development Diapause termination Bruchids Ovetwintering
INTRODUCTION
Bruchus rufimanus is a bruchid whose larvae develop in the seeds of the broad bean, Vicia faba (L.), and consume the reserves of the cotyledons. The species is univoltine. The adults are in reproductive diapause from September until April. They overwinter under the bark of trees or in lichens (Speyer, 195 1; Middlekauf, 1961; DuPont, unpublished) and return to the growing crop in the spring, once the beans begin to flower. At the time of colonization, the males have active reproductive organs: their diapause has already terminated. The females remain in diapause and their sexual development begins in the fields.
Experimental studies have shown that reproductive diapause termination depends, particularly in the case of females, on interactions between two factors: photo- period (long days, 18 h light-6 h dark) and a factor associated with the plant host (the consumption of its
*Institut de Biocenotique des Afrosyst6mes-URA CNRS 1298, Facultb des Sciences de Tours, Part de Grandmont, 37200 Tours, France.
pollen). In the absence of food, the proportion of females in which diapause terminates is small, even in long-day conditions. For the great majority of males, long pho- tophases alone are sufficient to terminate diapause (Tran and Huignard, 1992). Throughout reproductive dia- pause, in temperate climate, insects live at low tempera- tures and do not experience the factors which stimulate reproduction (long days and a supply of food).
In some species, it has been reported that there is a change in the physiological state of the insects during diapause development, which renders them more recep- tive to those factors which stimulate the termination of diapause. The form of diapause development in which particular cues, whether or not the same as those which induced diapause, are necessary for diapause termin- ation, was termed tachytely by Hodek (1981, 1983). Tachytely has been observed in several different species of bruchids, including B. pisorum (Pesho and Van Houten, 1982), Bruchidius atrolineatus (Lenga and Huig- nard, 1992) and B. rgfimanus (Tran and Huignard, 1992). In the case of these insects, various climatic and plant host-related factors are necessary for diapause termination.
In other cases, the duration of diapause is apparently
769

770 B. TRAN ef al.
predetermined and its termination is not materially affected by any external factors. Such diapause develop- ment was termed horotely by Hodek (1981, 1983) and has been described in the case of Sitona cylindricollis (Hans, 1961) and Riptortus clavatus (Numata and Hi- daka, 1984). Danks (1987) reported that there are large numbers of horotelic species living in temperate climates. It is not known whether B. rufimanus ever exhibits this form of diapause development. It is not known how insects not exposed, or exposed for different periods to environmental conditions in which diapause is normally maintained, would respond to those factors which termi- nate diapause. In this paper, we consider the responsive- ness of B. rujimanus to photoperiod and to factor(s) from the plant host which may affect the course of diapause development.
MATERIALS AND METHODS
Maintenance of reproductive diapause in artijkial con- ditions
Seeds of V. faba (INRA Soravi cultivar) were har- vested in the Lignikres region, RCgion Centre of France, in July, and transported to the laboratory. Adults which emerged from these seeds in August and September were in reproductive diapause. They were placed in a tempera- ture-controlled room, at lO”C, in continuous darkness, where they were fed on a 10% sucrose solution. These conditions are referred to hereafter as condition A. In these conditions, all the adults remained in reproductive diapause, normal development being prevented by low temperature. The proportion surviving during the first few months was consistently high. After 6 months, the number surviving decreased rapidly. In these conditions, no termination of diapause was ever observed.
Experimental conditions
Diapausing adults, both males and females, were removed from the stock culture either at emergence, or after periods of l-8 months of maintenance under condition A. They were placed in plastic boxes (16 dm3, dimensions 30 x 21 x 25 cm), maintained at a constant temperature (25°C) and subjected to a photoperiod of 18 h light-6 h dark, henceforth known as condition B, which would normally result in the termination of diapause in a significant proportion of individuals (Tran and Huignard, 1992). In condition B, approximately half the insects were offered inflorescences of V. faba from which they consumed pollen (Tran and Huignard, 1992), renewed every 5 days; the remainder were given no flowers (Table 1). In every case, they were provided with a 10% sucrose solution.
Preliminary studies (Tran, unpublished) showed that insects respond to stimuli leading to termination of the diapause only after a minimal latency period of 10-15 days. Experiments in condition B could only last for a maximum of 30 days since mortality beyond this period was excessive. After 30 days of exposure to condition B,
diapause termination was determined in males by measurement of the diameter of the lateral accessory gland. The increased diameter of this gland, together with the presence of secretions in its lumen, is the most reliable criterion for characterizing the termination of the diapause (Tran and Huignard, 1992). The same phenomenon has been observed in another bruchid, B. atrolineatus (Glitho et al., 1991). In females, diapause termination is easily determined by the presence of vitellogenic oiicytes in the ovarioles and mature oiicytes in the lateral oviducts.
Histological studies
In order to follow the changes in reproductive organs during the course of diapause development, females were dissected after different periods of exposure in condition A. The ovarioles were removed and fixed in alcoholic Bouin. Sections of prepared blocks were cut at 7 pm, and stained in one of three different ways: Goldner’s variety of Masson trichrome, PAS or methyl green-py- ronine were all used following the techniques described by Martoja and Martoja (1967).
RESULTS
Changes in the ovarioles
B. rujimanus possesses 24 ovarioles opening into paired lateral oviducts (Tran, 1992). These ovarioles are of the meroistic telotrophic kind, as is found in other bruchids (Biining, 1972). In September [Fig. l(A)], the ovarioles of diapausing females consist of a germarium, and proximally, a single oiicyte measuring 6-7 pm in diameter. This oijcyte consists of a large central nucleus, and a small amount of peripheral cytoplasm. The oiicyte is surrounded by follicular epithelium.
During the first 5 months of diapause, the structure of the ovarioles remains the same [Fig. l(B)]. However, after 7 months had elapsed in diapausing conditions [Figs l(C) and (E)], in March, the diameter of the terminal oiicyte had increased to 11 pm. This is due
TABLE 1. Percentage diapause termination among males and females of B. rujmanus, in 25”C, 18 h light4 h dark, preceded by various
durations of exposure to lo”C, in constant darkness
Males Females Exposure to condition A Sucrose only Flowers Sucrose only Flowers (in months) (%) (%) (%) (%)
0 0 (26) 29 (35) 0 (21) 27 (30) 1 43 (30) 33 (30) 3 29 (89) 77 (26) 3 (78) 52 (29) 4 80 (25) 83 (30) 5 48 (31) 7 (29) 6 57 (44) 2 (43) 7 100 (31) 89 (37) 8 48 (25) 8 (26)
All bruchids were fed with 10% sucrose, and some (as indicated) were provided with V. faba flowers, from which they ate pollen. Sample sizes are given in parentheses.
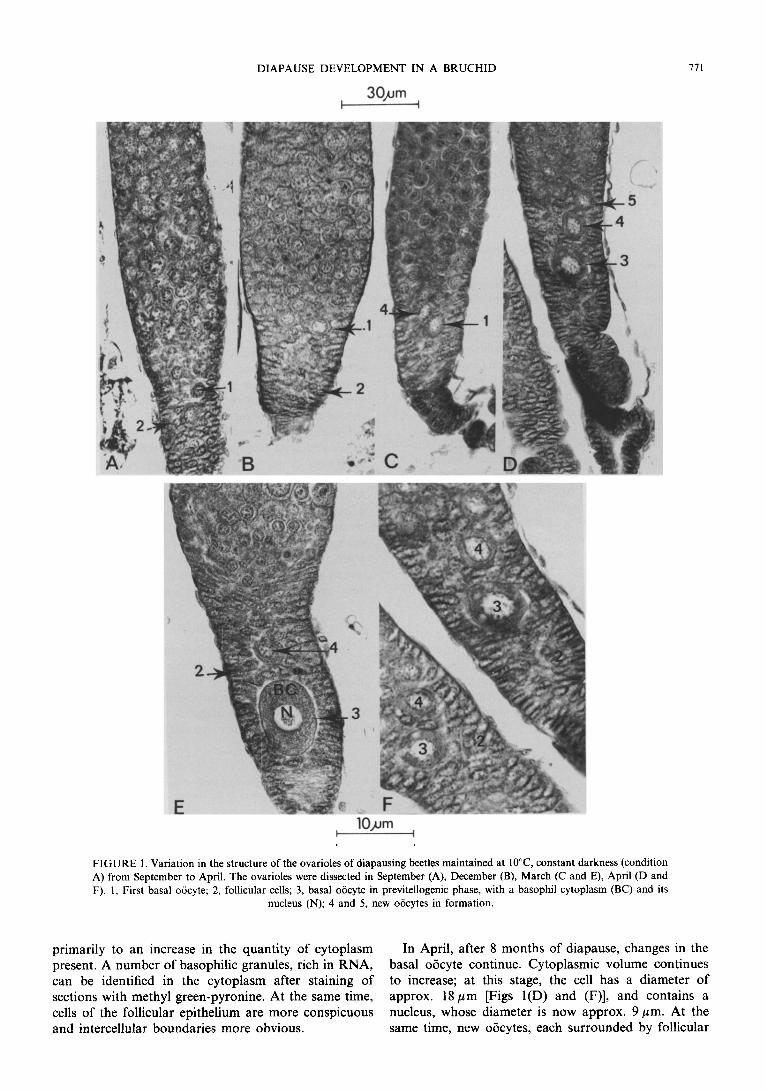
DIAPAUSE DEVELOPMENT IN A BRUCHID
30pm
1Op-n
FIGURE 1. Variation in the structure of the ovarioles of diapausing beetles maintained at 10°C constant darkness (condition A) from September to April. The ovarioles were dissected in September (A), December (B), March (C and E), April (D and F). 1, First basal oocyte; 2, follicular cells; 3, basal o6cyte in previtellogenic phase, with a basophil cytoplasm (BC) and its
nucleus (N); 4 and 5, new oiicytes in formation.
primarily to an increase in the quantity of cytoplasm In April, after 8 months of diapause, changes in the present. A number of basophilic granules, rich in RNA, basal oijcyte continue. Cytoplasmic volume continues can be identified in the cytoplasm after staining of to increase; at this stage, the cell has a diameter of sections with methyl green-pyronine. At the same time, approx. 18 pm [Figs l(D) and (F)], and contains a cells of the follicular epithelium are more conspicuous nucleus, whose diameter is now approx. 9 pm. At the and intercellular boundaries more obvious. same time, new oocytes, each surrounded by follicular

712 B. TRAN et al.
cells begin to separate from the germarium, behind the Additional eflects of stimulation by pollen from the host first odcyte. plant
We interpret the increase in the diameter of the basal oGcyte, after about 7 months of diapause, as due to previtellogenic processes. Electrophoretic studies re- vealed that no protein bands characteristic of vitel- logenin are present in the haemolymph at this time. These bands do not appear until after the end of the diapause.
Males. Twenty-nine % of males placed after emer- gence, for 30 days, in condition B, with flowers of V. faba, had functional reproductive organs (Table 1). After 1 month in condition A, the proportion of males in which diapause terminated when transferred to con- dition B, did not significantly increase (x2 = 1.5, P > 0.05), to 43%.
Variation in responsiveness of insects to photoperiod during the course of diapause
Males. In the case of males emerging from seeds and then immediately placed for 30 days in condition B, there were no cases of diapause termination (Table 1). After 3 months’ exposure to condition A, and subsequent exposure to condition B, 29% of males began reproductive maturation (Table 1). Increase in the length of exposure to cold beyond 5 months (Table 1) did not significantly increase the proportion of males in which diapause terminated (x:~,~, = 0.73, P > 0.05).
When 3, 4 or 7 months in condition A preceded exposure to condition B with flowers of V. faba, the proportion of males in which diapause terminated was always greater than 70% (Table 1). It reached 100% in the 7 months’ sample. The proportion observed in these three groups did not significantly differ ( x:~,~, = 1.04, P > 0.05).
Comparative measurements of the diameter of the lateral accessory gland (Table 2), in the case of those adults whose diapause had terminated upon exposure to condition B, did not reveal any significant differences among the five groups tested (H, = 4.1, P > 0.05).
The diameter of the lateral accessory gland, in groups of diapausing males, exposed for different lengths of time to condition A before exposure to condition B, were measured (Table 2). Application of a Kruskall-Wallis test revealed no significant differences between the groups (H, = 3.01, P > 0.05).
In the males in which diapause had terminated, the diameter of the lateral accessory glands was smaller in those which had experienced longer exposure to cold conditions. This was shown statistically, using a Spear- man’s rank correlation coefficient (p, = -0.3, t = -2.8, 76 d.f., P < 0.01).
Females. The proportion of females in which diapause terminated upon exposure to condition B in the presence of flowers of V. faba, after a month or less in cold conditions, was relatively small (27 and 33%) (Table 1). After 3 months in diapause, then exposure to condition B and the presence of flowers, 52% of the females became reproductive (Table 1). This proportion amounts to 83 and 89% in the case of those females which were kept under condition A for 4 or 7 months, respectively (Table 1). Values obtained in the five groups were significantly different (x$,~, = 37.86, P c 0.001).
Females. There is no evidence of diapause termination in females that were placed immediately after emergence in condition B (Table 1). Increase in the period of exposure to cold did not appear to increase the respon- siveness of females to photoperiod. Among four groups tested, the proportion of females in which diapause was terminated never exceeded 10% (Table 1).
DISCUSSION
These studies have shown that, in B. ru$manus, there are developmental changes during the period of dia- pause. There are changes in the structure of the ovari- oles, such as the commencement of previtellogenesis, and also in the responsiveness of insects to factors which may lead to the termination of diapause, photoperiod and the
TABLE 2. Average diameters @m, & SEM) of the lateral accessory gland of males of B. rujimanus, after 30 days in 25”C, 18 h light-6 h dark, preceded by various durations
of exposure to lO”C, in constant darkness
Exposure to condition A (in months)
Males with inactive glands
Sucrose only Flowers
Males with active glands
Sucrose only Flowers
0 55 (* 1.4) 69 (k 1.3) 1 64 (k2.1) 3 57 (kO.5) 61 (f2.9)
4 56 (+O) 5 57 (+ 1.5) 6 54 (h 1.6) 7 -
8 56 (+O)
- 193 (_f11.8) 221 (kS.3)
144 ((k7.8) 196 (&- 12.1) 214 (k8.9)
131 (& 10.1) 119 (k5.3)
214 (f 13.9) 113 (+3.2)
All bruchids were fed with 10% sucrose, and some (as indicated) provided with V. faba flowers, from which they ate pollen. Activity of the glands was recognized by the presence of secretion in their lumen.

DIAPAUSE DEVELOPMENT IN A BRUCHID 773
presence of the plant host flowers. Diapausing insects which were not placed in cold conditions, but exposed directly on emergence to factors favourable to the termination of reproductive diapause, were not respon- sive to long days and high temperature, but remained in diapause. In the presence of V. fuba flowers immediately after emergence, 27% of the females and 29% of the males began to show reproductive activity (Table 1). However, in these conditions, even after consumption of I’. faba pollen, the majority of the insects were still in diapause.
It was only after at least 3 months in total darkness, at 10°C that the proportion of insects responsive to factors terminating the diapause began to increase. Experimental results showed that, after this 334 month period, there are no differences in responsiveness to the factors under consideration, at least in the conditions which we have employed.
According to Tauber and Tauber (1976), Hodek (1983, 1988) and Danks (1987) there is a progressive decrease in the intensity of diapause or, in other words, an increase in the likelihood of diapause termination under the influence of environmental factors. This is shown in a number of species. Processes underlying this responsiveness to external factors are characteristic of the horotelic type of diapause development (Hodek, 1983), and if they continue to their natural end, they would lead to the spontaneous termination of diapause. According to Tauber and Tauber (1976) such processes are very frequent among temperate insects. Our results on B. rufimanus show that the situation is complex and that the decrease in the intensity of diapause is not uniform. Diapause may even be biphasic in this species.
The first phase of diapause is characterized by its depth in the great majority of individuals. During this period, no external factor can lead to the termination of the diapause. The duration of this period, under our experimental conditions, is around 3 months, in about 60% of males and 70% of females.
The second phase is less deep, in both males and females. During this period, a high percentage of the insects are capable of having their diapause terminated if exposed to appropriate conditions. We suggest that horotelic processes take place during the first phase. In & ~ufinanus, they would not lead to the termination of the diapause, but only render the insects able to respond to environmental factors. During the second phase of the diapause, the insects would only be “waiting” for the environmental stimuli which will induce the termination of diapause. The termination of diapause is attained after the succession, therefore, of horotely and then tachytely. According to Tauber and Tauber (1976), a similar situation is found in a few species which require a specific stimulus for the termination of diapause. One such case is that of Ostrinia nubilalis, in which, according to Beck (1967), there is: “an initial phase in which considerable diapause development occurs (i.e. as measured by a progressive reduction in diapause inten-
sity), and a second phase that is somewhat static, until the terminating stimulus occurs”.
A number of studies, in various diapausing species, have revealed the importance of a period of low tem- perature exposure for the completion of diapause devel- opment (Danks, 1987). This period of cold, of variable duration, enables metabolic reserves to be used more slowly, and can also have an effect on the endocrine system. In B. rujimanus, exposure to condition A cer- tainly enables the insects to use their reserves more slowly. But the effects of this relatively cold exposure on the horotelic processes are not known.
Our experiments have revealed significant variation among individual insects. A certain number of individ- uals exhibited termination of diapause without exposure to cold, some with longer and some with shorter ex- posure at 10°C. This variability is particularly obvious when insects have been exposed to a short period of continuous darkness, at 10°C. It would appear that, within the population, individuals vary in the intensity of their diapause at any one point in time. This variation may have a genetic component, but could also be attributable to differences in the conditions experienced during larval development, i.e. have an environmental basis. Studies on B. atrolineatus (Glitho et al., 1991) have shown the importance of conditions during post-embry- onic growth on the intensity of diapause. In B. ruji- manus, the observed variability has not been explained and deserves further study. In particular, a comparative approach should be undertaken with strains from the region of Meknes, Morocco, which seem to undergo a longer larval development, and shallower diapause (El Shazly, personal communication).
In France, the period of deepest diapause, during the first few months, during the late summer and early autumn, is probably very important in the life history of B. rufimanus. It prevents reproductive development and sexual activity in a period where resources, including both flowers and pods of the host, may be available, and climatic conditions are apparently favourable for repro- duction, but which would lead to the death of immature insects in the winter.
REFERENCES
Beck S. D. (1967) Water intake and the termination of diapause in the European corn borer, Ostrinia nubilalis. J. Insecr Physiol. 13, 739-150.
Biining J. (1972) Untersuchungen am Ovar von Eruchidius obfecrus Say (Coleoptera Polyphaga) zur Kllrung der Oocytenwachstums in der Pravitellogenese. Z. Zellforsch. 128, 241-282.
Danks H. V. (1987) Insect Dormancy: an Ecological Perspective. Biological Survey of Canada Monograph Series 1. Ottawa.
Glitho I. A., Lenga A. and Huignard J. (1991) Intensity of the male reproductive diapause in Bruchidius arrolinearus (Pit) (Coleoptera: Bruchidae) is affected by induction conditions. Invert. Repr. Dev. 19, 233-243.
Hans H. (1961) Termination of diapause and continuous laboratory rearing of the sweet clover weevil, Sitona cylindrieofiis Fabr. Enro- mofogia exp. appl. 4, 4146.
Hodek I. (1981) Le role des signaux de l’environnement et des

774 B. TRAN et al.
processus endogenes dans la regulation de la reproduction par la diapause imaginale. Bull. Sot. Zool. I+. 106, 317-326.
Hodek I. (1983) Role of environmental factors and endogenous mechanisms in the seasonality of reproduction in insects diapausing as adults. In Diapause and Life Cycle Strategies in Insects (Eds Brown V. K. and Hodek I.), pp. 9-33. Junk, The Hague.
Hodek I. (1988) Photoperiod response and reproductive activity in Pyrrhocoris aprerus (Heteroptera) in relation to diapause develop- ment. Zool. Jb. Physiol. 92, 57-70.
Lenga A. and Huignard J. (1992) Effect of changes in the thermoperiod on reproductive diapause in Bruchidius atrolineatus Pit (Coleoptera: Bruchidae). Physiol. Ent. 17, 241-254.
Martoja R. and Martoja M. (1967) Initiation aux Techniques de
I’Histologie Animale. Masson et Cie, Paris. Middlekauf W. (1961) Field studies on the bionomics and control of
the broad bean weevil Bruchus rujimanus. J. Econ. Em. 44,240-243. Numata H. and Hidaka T. (1984) Photoperiodic control of adult
diapause in the bean bug Riptortus clavatus Thunberg (Heteroptera: Coreidae). III. Diapause development and temperature. Appl. Em.
Zool. 19, 3X-360.
Pesho G. R. and Van Houten R. J. (1982) Pollen and sexual matu- ration of the pea weevil (Coleoptera: Bruchidae). Ann. em. Sot. Am. 75, 439443.
Speyer W. (1951) Biologie und Beklmpfung des Pferdebohnenkiifers Bruchus ru$manus. Mitt. Biol. Zentralanst. 70, 122-126.
Tauber M. J. and Tauber C. A. (1976) Insect seasonality: diapause maintenance, termination and post-diapause development. A. Rev.
Em. 21, 81-107. Tran B. and Huignard J. (1992) Interactions between photoperiod and
food affect the termination of reproductive diapause in Bruchus rufmanus (Boh.) (Coleoptera: Bruchidae). J. Insect Physiol. 38,
633642.
Acknowledgements-This study was supported by a grant from the Conseil Regional de la Region Centre, France, to B. Tran. The authors wish to thank Dr P. F. Credland for his comments, and his help with the translation.
Related Documents











![AgrometScmme Oegrés ]ours [ Fonctlon d'é'ongatlon Besoin de froid pour lever la diapause Besoin de 250 DJ > CC 12 oc et pluie Maturation Diapause larvaire Larves matures Nymphose](https://static.cupdf.com/doc/110x72/6051b2e57e114a63fb30ebd7/agromet-scmme-oegrs-ours-fonctlon-dongatlon-besoin-de-froid-pour-lever.jpg)