Hydrobiologia 453/454: 295–308, 2001. R.M. Lopes, J.W. Reid & C.E.F.Rocha (eds), Copepoda: Developments in Ecology, Biology and Systematics. © 2001 Kluwer Academic Publishers. Printed in the Netherlands. 295 Diapause in copepods (Crustacea) from ephemeral habitats with different hydroperiods in Everglades National Park (Florida, U.S.A.) Maria Cristina Bruno 1 , William F. Loftus 2 , Janet W. Reid 3 & Sue A. Perry 1 1 South Florida Natural Resources Center, Everglades National Park, Homestead, FL 33034, U.S.A. 2 U.S. Geological Survey, Biological Resources Division, Everglades National Park, Homestead, FL 33034, U.S.A. 3 Department of Invertebrate Zoology, National Museum of Natural History, Smithsonian Institution, Washington, DC 20560-0163, U.S.A. Key words: copepods, diapause, ephemeral habitats, pre-fertilized females, Everglades, wetlands Abstract Water management practices in the Everglades have severely stressed the natural system, particularly by reducing the hydroperiods of much of the region. During the dry season of 1999, we investigated the influence of hydroperiod on the species composition and dormancy patterns of freshwater copepod communities in seasonal wetlands of Everglades National Park, Florida, U.S.A. The habitats were characterized by an annual dry season, from December through June. We sampled at two locations: the Long Pine Key area of the Rocky Glades region (short hydroperiod, ca. 4–5 months), and western Taylor Slough (intermediate hydroperiod, ca. 8–10 months). Both areas have experienced a reduction in natural hydroperiods and an increase in the frequency of dry-down. We collected weekly plankton samples from Rocky Glades solution holes to assess the potential species pool of copepods. To document the taxa capable of surviving dry-down by resting, we performed three immersion trials in which we rehydrated, in laboratory aquaria, sediment patches from solution holes and surface soils from all stations. Only a subset of the planktonic species collected emerged from the dried sediments. The cyclopoids Microcyclops rubellus and Paracyclops poppei were dominant. This is the first record of diapause for P. poppei. Species distributions from the different hydroperiod soil patches indicated that more diapausing species occurred at the sites that dried for shorter periods. Emerging individuals of M. rubellus and P. poppei were mainly ovigerous females, demonstrating a resting strategy seldom before recorded. The cyclopoid Diacyclops nearcticus had not been previously reported to diapause, but they emerged from the dried sediments in our trials. Our collections included six new records for Flor- ida: Diacyclops nearcticus, Megacyclops latipes, Orthocyclops modestus, Elaphoidella marjoryae, Bryocamptus sp. and Bryocamptus cf. newyorkensis. Paracyclops poppei, Macrocyclops fuscus and Arctodiaptomus floridanus are new records for Everglades National Park. Clearly, diapause is an important strategy for the persistence of copepods in short-hydroperiod wetlands. The duration of the dry period appears to be inversely related to the number of species that emerge from diapause. Introduction The Florida Everglades is an extensive subtropical wetland ecosystem formed during the past 5000 years, when prolonged inundation resulted in peat and marl accretion within the pre-existing limestone depres- sion (Gleason & Stone, 1994). The Everglades once covered over 10 000 km 2 of the southern Florida pen- insula, but half the original area has been developed for agricultural and urban use (Gunderson & Loftus, 1993). In the southern Everglades, marsh hydrology was changed during the 1950s and 1960s by wa- ter management projects that impounded and diverted water from the north, and drained water eastward by canals through the Atlantic Coastal Ridge. Water depths and hydroperiods (duration of surface water flooding) in Everglades National Park (ENP) were re- duced markedly (Loftus et al., 1992). Hydroperiod is the major factor influencing the unique vegeta- tion patterns of the Everglades (Gunderson, 1994),

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hydrobiologia 453/454: 295–308, 2001.R.M. Lopes, J.W. Reid & C.E.F. Rocha (eds), Copepoda: Developments in Ecology, Biology and Systematics.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
295
Diapause in copepods (Crustacea) from ephemeral habitats with differenthydroperiods in Everglades National Park (Florida, U.S.A.)
Maria Cristina Bruno1, William F. Loftus2, Janet W. Reid3 & Sue A. Perry1
1South Florida Natural Resources Center, Everglades National Park, Homestead, FL 33034, U.S.A.2U.S. Geological Survey, Biological Resources Division, Everglades National Park, Homestead,FL 33034, U.S.A.3Department of Invertebrate Zoology, National Museum of Natural History, Smithsonian Institution,Washington, DC 20560-0163, U.S.A.
Key words: copepods, diapause, ephemeral habitats, pre-fertilized females, Everglades, wetlands
Abstract
Water management practices in the Everglades have severely stressed the natural system, particularly by reducingthe hydroperiods of much of the region. During the dry season of 1999, we investigated the influence of hydroperiodon the species composition and dormancy patterns of freshwater copepod communities in seasonal wetlandsof Everglades National Park, Florida, U.S.A. The habitats were characterized by an annual dry season, fromDecember through June. We sampled at two locations: the Long Pine Key area of the Rocky Glades region (shorthydroperiod, ca. 4–5 months), and western Taylor Slough (intermediate hydroperiod, ca. 8–10 months). Both areashave experienced a reduction in natural hydroperiods and an increase in the frequency of dry-down. We collectedweekly plankton samples from Rocky Glades solution holes to assess the potential species pool of copepods. Todocument the taxa capable of surviving dry-down by resting, we performed three immersion trials in which werehydrated, in laboratory aquaria, sediment patches from solution holes and surface soils from all stations. Only asubset of the planktonic species collected emerged from the dried sediments. The cyclopoids Microcyclops rubellusand Paracyclops poppei were dominant. This is the first record of diapause for P. poppei. Species distributions fromthe different hydroperiod soil patches indicated that more diapausing species occurred at the sites that dried forshorter periods. Emerging individuals of M. rubellus and P. poppei were mainly ovigerous females, demonstratinga resting strategy seldom before recorded. The cyclopoid Diacyclops nearcticus had not been previously reported todiapause, but they emerged from the dried sediments in our trials. Our collections included six new records for Flor-ida: Diacyclops nearcticus, Megacyclops latipes, Orthocyclops modestus, Elaphoidella marjoryae, Bryocamptussp. and Bryocamptus cf. newyorkensis. Paracyclops poppei, Macrocyclops fuscus and Arctodiaptomus floridanusare new records for Everglades National Park. Clearly, diapause is an important strategy for the persistence ofcopepods in short-hydroperiod wetlands. The duration of the dry period appears to be inversely related to thenumber of species that emerge from diapause.
Introduction
The Florida Everglades is an extensive subtropicalwetland ecosystem formed during the past 5000 years,when prolonged inundation resulted in peat and marlaccretion within the pre-existing limestone depres-sion (Gleason & Stone, 1994). The Everglades oncecovered over 10 000 km2 of the southern Florida pen-insula, but half the original area has been developedfor agricultural and urban use (Gunderson & Loftus,
1993). In the southern Everglades, marsh hydrologywas changed during the 1950s and 1960s by wa-ter management projects that impounded and divertedwater from the north, and drained water eastwardby canals through the Atlantic Coastal Ridge. Waterdepths and hydroperiods (duration of surface waterflooding) in Everglades National Park (ENP) were re-duced markedly (Loftus et al., 1992). Hydroperiodis the major factor influencing the unique vegeta-tion patterns of the Everglades (Gunderson, 1994),
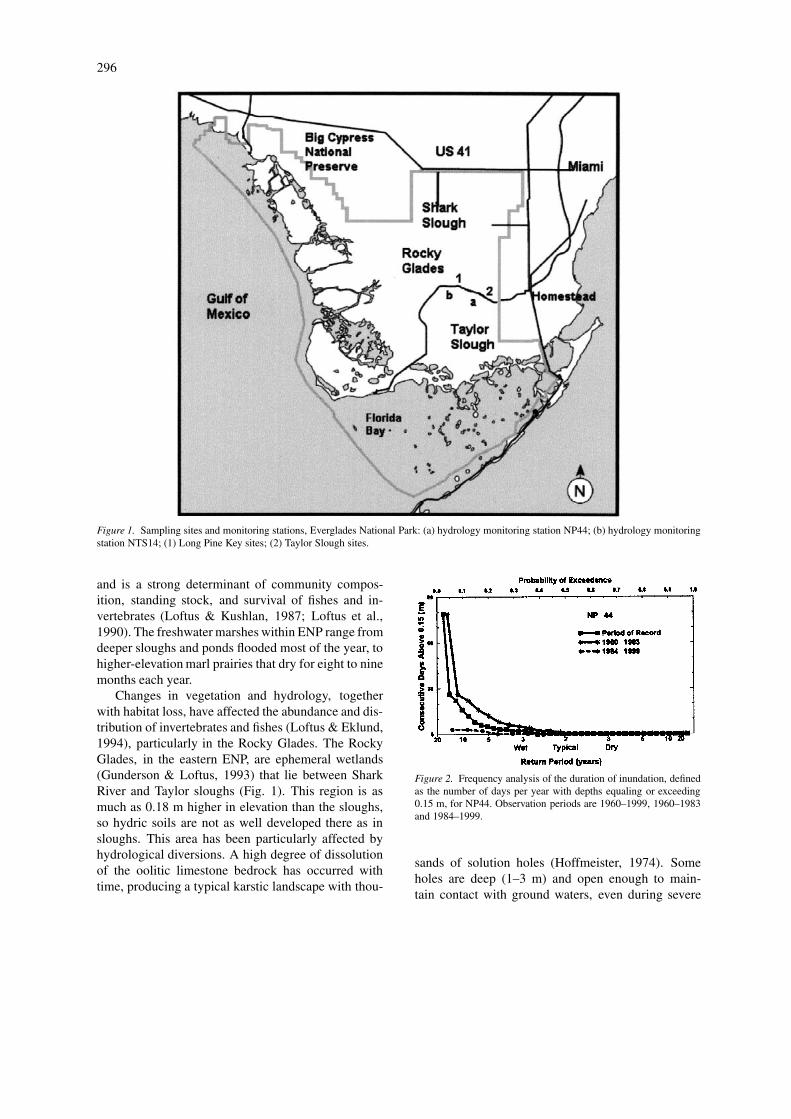
296
Figure 1. Sampling sites and monitoring stations, Everglades National Park: (a) hydrology monitoring station NP44; (b) hydrology monitoringstation NTS14; (1) Long Pine Key sites; (2) Taylor Slough sites.
and is a strong determinant of community compos-ition, standing stock, and survival of fishes and in-vertebrates (Loftus & Kushlan, 1987; Loftus et al.,1990). The freshwater marshes within ENP range fromdeeper sloughs and ponds flooded most of the year, tohigher-elevation marl prairies that dry for eight to ninemonths each year.
Changes in vegetation and hydrology, togetherwith habitat loss, have affected the abundance and dis-tribution of invertebrates and fishes (Loftus & Eklund,1994), particularly in the Rocky Glades. The RockyGlades, in the eastern ENP, are ephemeral wetlands(Gunderson & Loftus, 1993) that lie between SharkRiver and Taylor sloughs (Fig. 1). This region is asmuch as 0.18 m higher in elevation than the sloughs,so hydric soils are not as well developed there as insloughs. This area has been particularly affected byhydrological diversions. A high degree of dissolutionof the oolitic limestone bedrock has occurred withtime, producing a typical karstic landscape with thou-
Figure 2. Frequency analysis of the duration of inundation, definedas the number of days per year with depths equaling or exceeding0.15 m, for NP44. Observation periods are 1960–1999, 1960–1983and 1984–1999.
sands of solution holes (Hoffmeister, 1974). Someholes are deep (1–3 m) and open enough to main-tain contact with ground waters, even during severe

297
droughts (Loftus et al., 1992). However, most areshallow (< 1 m) and partly filled by hydric soils thatsupport rooted aquatic macrophytes. During the wetseason (June–October), rainfall and groundwater re-charge fill the solution holes and reflood the wetlandsurface. In the dry season (November–May), surfacewater disappears when pumping operations that intro-duce water from canals stop. This, together with evap-oration, percolation and evapotranspiration, leavesonly the deepest holes with water. Re-establishmentof more natural hydrological conditions in the trans-itional wetlands of the Rocky Glades is an essentialcomponent of the ecosystem restoration program inENP. Average peak water-level reductions of 21 and32 cm occurred in wet and dry seasons, respectively,after change of management practices in the 1960s(Loftus et al., 1992). Modeling results indicated thatthe hydroperiod of the Rocky Glades has been reducedon average from 8–10 months to 3–7 months yr−1
at present (Olmsted et al., 1980). For a typical wetyear, hydrological analysis indicated that the numberof consecutive days above a water level of 0.15 m(a measure of hydroperiod) varied from 25 to 80 in1960–1983, but was reduced to less than 10 days in1984–1999 (Fig. 2).
Although major effects of altered hydropattern onsome aquatic communities have been demonstrated(Loftus et al. 1992; Loftus & Eklund, 1994), theeffects on zooplanktonic communities have not beenstudied. Copepods are the zooplanktonic group withthe highest density in ENP (Loftus et al., 1990).Earlier emergence trials by Loftus & Reid (2000),with copepods from a broader hydroperiod range ofENP marshes, showed that differences in speciesrichness and densities of individuals were related tohydroperiod. Those differences might be explainedby low levels of organic matter in short-hydroperiodmarsh soils, and/or by the frequency and severity ofdry-down events. There are no pre-management dataon copepod communities, but the low species rich-ness recorded in ENP (Reid, 1992) might be the resultof water management activities as well as historicalzoogeography.
The success of aquatic animals inhabiting tempo-rary ponds depends on survival through the dry season,and on their dynamics during the wet season (Wyn-gaard et al., 1991). These animals have a diversityof adaptations to life in ephemeral habitats, includingresting stages, dispersion to more suitable areas, andlife cycles with aquatic and terrestrial phases (Wig-gins et al., 1980). Temporary ponds are depauperate
Table 1. Solution holes dimensions and date of drying at theLong Pine Key, ENP location
Site Depth below ground Length (m) Width (m) Date of dryingsurface (m) (1999)
LP1S 0.25 1.50 1.40 February 2, 1999LP1M 0.42 1.60 1.50 February 9, 1999LP1D 0.82 1.30 1.10 March 30, 1999LP2S 0.30 2.50 1.40 February 9, 1999LP2M 0.38 3.40 1.80 February 9, 1999LP2D 0.52 1.52 1.00 April 4, 1999LP3S 0.28 1.50 1.40 January 19, 1999LP3M 0.37 1.30 1.30 January 19, 1999LP3D 0.89 3.00 2.50 April 4, 1999
in species, because few can adapt to the stress ofdesiccation (Wiggins et al., 1980). For those speciesthat have evolved dormant stages, those stages must beable to resist complete desiccation for weeks, monthsand even years (Roy, 1932; Rzoska, 1961; Champeau,1970a, b). Because the timing of dormancy and pat-terns of emergence determine community structureand dynamics (De Stasio, 1990), their assessmentis an essential starting point for studying copepodcommunities in ephemeral habitats.
The objectives of our research were to focus onlocations of short to intermediate hydroperiod, to in-vestigate the suite of strategies used by planktoniccopepods to survive episodes of drying, and to doc-ument which taxa use those adaptations to exploitEverglades wetlands during the wet season. In partic-ular, we wanted to assess if and how the hydroperiodclasses affected the species assemblages of copepodsemerging from diapause. This was done by contrastingthe numbers of planktonic species from water samplesto the number of species emerging from dry pools onthe marsh surface and from solution holes.
The restoration of the historical hydroperiods inthe Everglades is a major conservation goal. Data fromthis study will help to understand the responses ofcopepod communities to anticipated increases in hy-droperiod. Because copepods are important prey forcommon fish species in the marshes (Rader, 1999),information on copepod composition and dynamics indifferent habitats can be applied in modeling food-webchanges in the system.
Materials and methods
We collected samples from two locations (Fig. 1)with different hydroperiods. We used data from Ever-
298
Figure 3. Average daily temperatures (◦C), Everglades, NationalPark, from January to May 1999.
glades National Park hydrological monitoring stationsNP44 and NTS14 to calculate the hydroperiods at oursampling locations. Within the Rocky Glades region,the marl prairies at Long Pine Key locations (25◦ 25′10′′ N, 80◦ 39′ 7′′ W) had the shortest hydroperiod.We sampled there at three sites, named LP1, LP2,LP3. LP1 was about 800 m east of LP2, and LP3was about 300 m west of LP2. At each Long PineKey site, we selected a set of three adjacent solutionholes that, from previous research, did not appear to beconnected to groundwater, thereby drying soon afterthe end of the wet season. All solution holes were lessthan 90 cm deep, and at each site we selected holesof three different depths (deep, medium depth, andshallow), designated LP1D, LP1M, LP1S, etc. (Table1), and a surface-soil station (named LP1soil, LP2soil,LP3soil), within an area of about 100 m2.
We estimated from hydrological data that LongPine Key surface-soil stations dried on 1 December,and Taylor Slough stations on 6 December 1998. Wechecked water levels in the Long Pine Key solutionholes weekly, and they dried 1.5–4 months later, de-pending on their depths (Table 1). We started to collectplanktonic copepod samples from water, two weeksafter surface water disappeared around the holes. From12 January 1999 until the solution holes dried, we col-lected 20-l water samples weekly, with a Par Jabasco�
hand pump, filtered the water through an 80 µm meshplankton net, and fixed the samples in the field with5% buffered formalin. At LPK, we also collectedsoil samples for immersion from both the soil andsolution-hole sites after they had dried.
We performed three trials to test for emergencefrom dry soil during this study. In western TaylorSlough, an area of intermediate hydroperiod (25◦ 24′6′′ N, 80◦ 36′ 13′′ W), we selected three sites to col-
lect surface-soil samples for the emergence trials. Thesites TS1, TS2, TS3 were approximately 20 m apartfrom each other. Solution holes did not occur at theTaylor Slough locations. The first samples, taken on 12January 1999, included soils from these sites: LP1soil,LP2soil, LP3soil, TS1, TS2 and TS3. The second,on 17 February, included soil stations as in the pre-vious trial, plus solution holes LP1S and M, LP2Sand M, LP3S and M. The third trial, on 27 April, in-cluded all previous stations, plus solution holes LP1D,LP2D, and LP3D. The immersion-trial durations were25, 60 and 24 d, respectively, based on the length oftime that copepods continued to emerge. The samplesconsisted of plugs (about 40 × 40 cm wide, 5–7 cmdeep) taken from the substratum with a shovel, thatwere placed in individually numbered, plastic-coveredtrays for transport to the laboratory. We rehydratedthe samples within a few days of collection, afterthe soil was completely dry. Each sample was rehy-drated in the laboratory, in separate 60-l aquaria thathad been previously disinfected with chlorine bleach,then filled with well water filtered through an 80-µmmesh plankton net. We simulated temperature (Fig. 3)and photoperiod (approximately 11 h of light for thefirst trial, 12 for the second trial, and 13 for the thirdtrial), in the laboratory. Each day, at approximatelythe same time during the trials, we filtered the waterof the aquaria, first siphoning with a PVC pipe, andthen collecting all the remaining water with a pipette,checking visually that no individuals were left. Waterwas filtered through the 80-µm mesh plankton net,and returned to the aquaria. The samples were fixedin 5% buffered formalin for sorting. We continued tosample until we had not collected any animal for fiveconsecutive days.
During rehydration, the soil completely coveredthe bottom of the aquaria. Therefore, we standardizedthe density of individuals that emerged by dividingthe total number of individuals by the bottom areaof the aquaria (0.165 m2). We sorted all copepodsusing a Leica� Stereoscope. We mounted the anim-als on permanent slides with Faure’s medium, andidentified them with a Leica� DMLS phase-contrastmicroscope. All the adults, and the stage III, IV, Vcopepodids, were identified to species, sex, and devel-opmental stage. Here we refer to the copepodid stagesas CIII, CIV, CVF, and CVM for copepodid stages III,IV, V female and V male, respectively. We used SPSSfor Windows© (SPSS Inc.) for all statistical analyses.
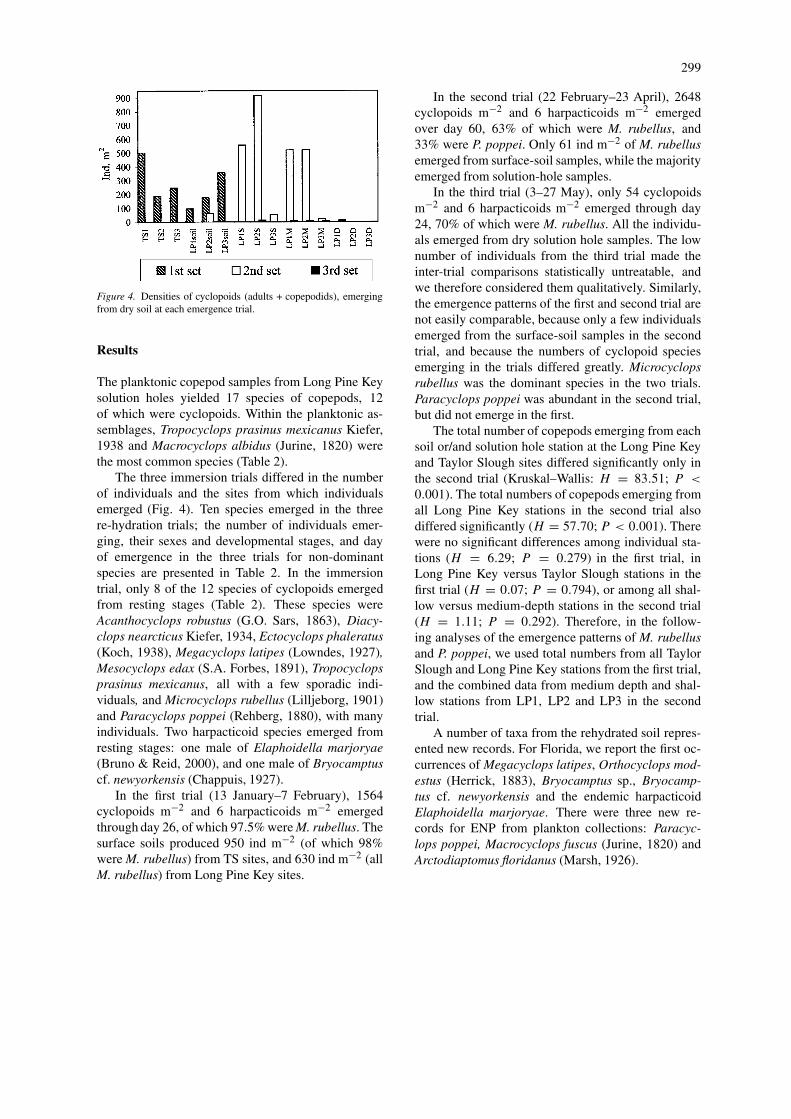
299
Figure 4. Densities of cyclopoids (adults + copepodids), emergingfrom dry soil at each emergence trial.
Results
The planktonic copepod samples from Long Pine Keysolution holes yielded 17 species of copepods, 12of which were cyclopoids. Within the planktonic as-semblages, Tropocyclops prasinus mexicanus Kiefer,1938 and Macrocyclops albidus (Jurine, 1820) werethe most common species (Table 2).
The three immersion trials differed in the numberof individuals and the sites from which individualsemerged (Fig. 4). Ten species emerged in the threere-hydration trials; the number of individuals emer-ging, their sexes and developmental stages, and dayof emergence in the three trials for non-dominantspecies are presented in Table 2. In the immersiontrial, only 8 of the 12 species of cyclopoids emergedfrom resting stages (Table 2). These species wereAcanthocyclops robustus (G.O. Sars, 1863), Diacy-clops nearcticus Kiefer, 1934, Ectocyclops phaleratus(Koch, 1938), Megacyclops latipes (Lowndes, 1927),Mesocyclops edax (S.A. Forbes, 1891), Tropocyclopsprasinus mexicanus, all with a few sporadic indi-viduals, and Microcyclops rubellus (Lilljeborg, 1901)and Paracyclops poppei (Rehberg, 1880), with manyindividuals. Two harpacticoid species emerged fromresting stages: one male of Elaphoidella marjoryae(Bruno & Reid, 2000), and one male of Bryocamptuscf. newyorkensis (Chappuis, 1927).
In the first trial (13 January–7 February), 1564cyclopoids m−2 and 6 harpacticoids m−2 emergedthrough day 26, of which 97.5% were M. rubellus. Thesurface soils produced 950 ind m−2 (of which 98%were M. rubellus) from TS sites, and 630 ind m−2 (allM. rubellus) from Long Pine Key sites.
In the second trial (22 February–23 April), 2648cyclopoids m−2 and 6 harpacticoids m−2 emergedover day 60, 63% of which were M. rubellus, and33% were P. poppei. Only 61 ind m−2 of M. rubellusemerged from surface-soil samples, while the majorityemerged from solution-hole samples.
In the third trial (3–27 May), only 54 cyclopoidsm−2 and 6 harpacticoids m−2 emerged through day24, 70% of which were M. rubellus. All the individu-als emerged from dry solution hole samples. The lownumber of individuals from the third trial made theinter-trial comparisons statistically untreatable, andwe therefore considered them qualitatively. Similarly,the emergence patterns of the first and second trial arenot easily comparable, because only a few individualsemerged from the surface-soil samples in the secondtrial, and because the numbers of cyclopoid speciesemerging in the trials differed greatly. Microcyclopsrubellus was the dominant species in the two trials.Paracyclops poppei was abundant in the second trial,but did not emerge in the first.
The total number of copepods emerging from eachsoil or/and solution hole station at the Long Pine Keyand Taylor Slough sites differed significantly only inthe second trial (Kruskal–Wallis: H = 83.51; P <
0.001). The total numbers of copepods emerging fromall Long Pine Key stations in the second trial alsodiffered significantly (H = 57.70; P < 0.001). Therewere no significant differences among individual sta-tions (H = 6.29; P = 0.279) in the first trial, inLong Pine Key versus Taylor Slough stations in thefirst trial (H = 0.07; P = 0.794), or among all shal-low versus medium-depth stations in the second trial(H = 1.11; P = 0.292). Therefore, in the follow-ing analyses of the emergence patterns of M. rubellusand P. poppei, we used total numbers from all TaylorSlough and Long Pine Key stations from the first trial,and the combined data from medium depth and shal-low stations from LP1, LP2 and LP3 in the secondtrial.
A number of taxa from the rehydrated soil repres-ented new records. For Florida, we report the first oc-currences of Megacyclops latipes, Orthocyclops mod-estus (Herrick, 1883), Bryocamptus sp., Bryocamp-tus cf. newyorkensis and the endemic harpacticoidElaphoidella marjoryae. There were three new re-cords for ENP from plankton collections: Paracyc-lops poppei, Macrocyclops fuscus (Jurine, 1820) andArctodiaptomus floridanus (Marsh, 1926).
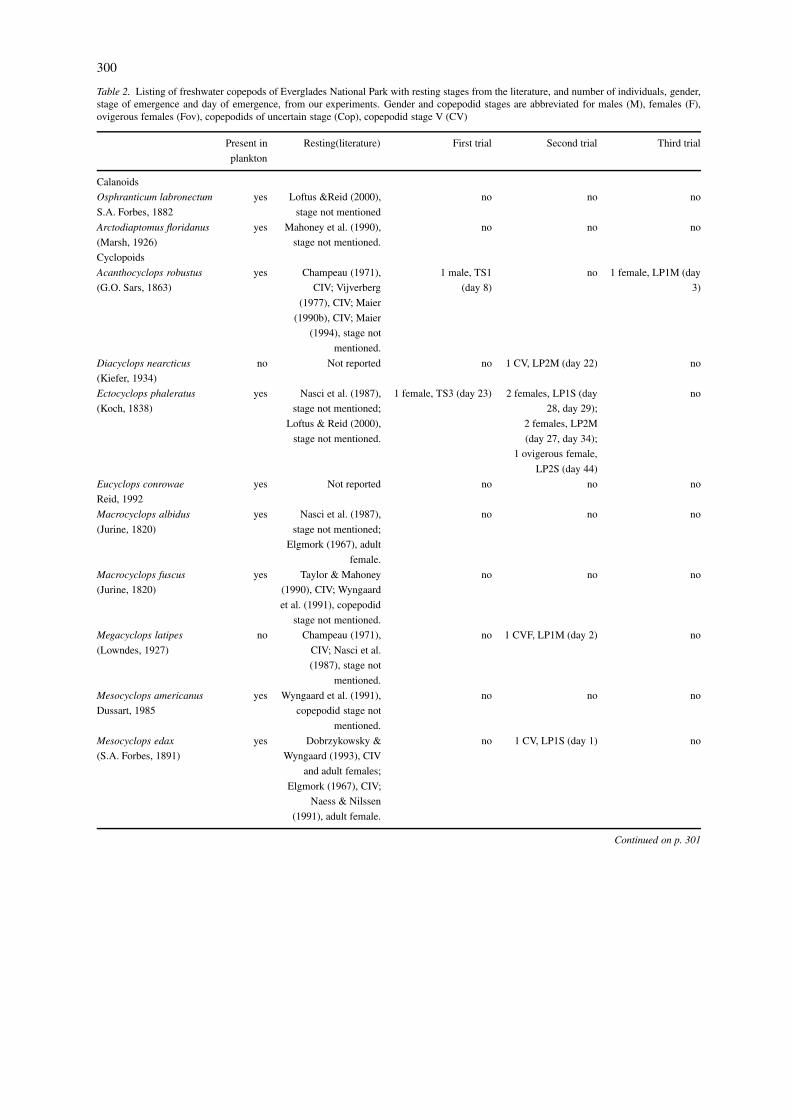
300
Table 2. Listing of freshwater copepods of Everglades National Park with resting stages from the literature, and number of individuals, gender,stage of emergence and day of emergence, from our experiments. Gender and copepodid stages are abbreviated for males (M), females (F),ovigerous females (Fov), copepodids of uncertain stage (Cop), copepodid stage V (CV)
Present in Resting(literature) First trial Second trial Third trial
plankton
Calanoids
Osphranticum labronectum yes Loftus &Reid (2000), no no no
S.A. Forbes, 1882 stage not mentioned
Arctodiaptomus floridanus yes Mahoney et al. (1990), no no no
(Marsh, 1926) stage not mentioned.
Cyclopoids
Acanthocyclops robustus yes Champeau (1971), 1 male, TS1 no 1 female, LP1M (day
(G.O. Sars, 1863) CIV; Vijverberg (day 8) 3)
(1977), CIV; Maier
(1990b), CIV; Maier
(1994), stage not
mentioned.
Diacyclops nearcticus no Not reported no 1 CV, LP2M (day 22) no
(Kiefer, 1934)
Ectocyclops phaleratus yes Nasci et al. (1987), 1 female, TS3 (day 23) 2 females, LP1S (day no
(Koch, 1838) stage not mentioned; 28, day 29);
Loftus & Reid (2000), 2 females, LP2M
stage not mentioned. (day 27, day 34);
1 ovigerous female,
LP2S (day 44)
Eucyclops conrowae yes Not reported no no no
Reid, 1992
Macrocyclops albidus yes Nasci et al. (1987), no no no
(Jurine, 1820) stage not mentioned;
Elgmork (1967), adult
female.
Macrocyclops fuscus yes Taylor & Mahoney no no no
(Jurine, 1820) (1990), CIV; Wyngaard
et al. (1991), copepodid
stage not mentioned.
Megacyclops latipes no Champeau (1971), no 1 CVF, LP1M (day 2) no
(Lowndes, 1927) CIV; Nasci et al.
(1987), stage not
mentioned.
Mesocyclops americanus yes Wyngaard et al. (1991), no no no
Dussart, 1985 copepodid stage not
mentioned.
Mesocyclops edax yes Dobrzykowsky & no 1 CV, LP1S (day 1) no
(S.A. Forbes, 1891) Wyngaard (1993), CIV
and adult females;
Elgmork (1967), CIV;
Naess & Nilssen
(1991), adult female.
Continued on p. 301
301
Table 2. contd.
Present in Resting(literature) First trial Second trial Third trialplankton
Microcyclops rubellus yes Hebert &Hann (1986), yes (see text) yes (see text) yes (see text)(Lilljeborg, 1901) stage not mentioned;
Wyngaard et al. (1991),stage not mentioned;
Nasci et al. (1987),stage not mentioned;
King et al. (1996),stage not mentioned;
Loftus &Reid (2000),stage not mentioned.
Microcyclops varicans yes Champeau (1971), CIII no no no(G.O. Sars, 1862) and CIV; Taylor &
Mahoney (1990), stagenot mentioned.
Orthocyclops modestus yes Taylor & Mahoney no no noHerrick, 1883) (1990), CIV; Wyngaard
et al. (1991), copepodidstage not mentioned.
Paracyclops poppei yes Not reported no yes (see text) no(Rehberg, 1880)Thermocyclops parvus yes Not reported no no noReid, 1989Tropocyclops prasinus yes Wyngaard et al. (1991), no 1 female, LP3M (day nomexicanus copepodid stage not 29);Kiefer, 1938 mentioned; data for T. 1 CVF, LP2 soil
prasinus, Nasci et al. (day2)(1987), stage not
mentioned; Taylor &Mahoney (1990), CIV.
HarpacticoidsElaphoidella marjoryae yes Not reported no 1 male, LP2S (day noBruno & Reid, 2000. 36)Bryocamptus sp. yes Not reported no no noBryocamptus cf. no Not reported no no 1 male, TS1 (day 1)newyorkensis(Chappuis, 1927)
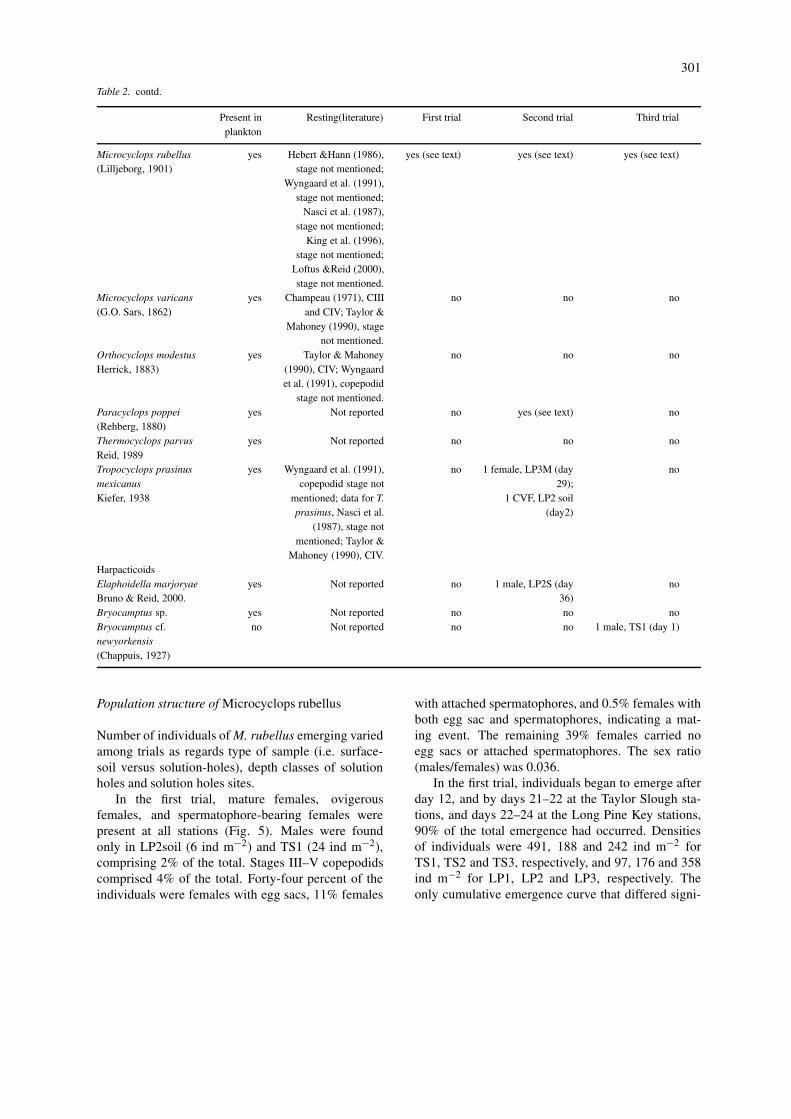
Population structure of Microcyclops rubellus
Number of individuals of M. rubellus emerging variedamong trials as regards type of sample (i.e. surface-soil versus solution-holes), depth classes of solutionholes and solution holes sites.
In the first trial, mature females, ovigerousfemales, and spermatophore-bearing females werepresent at all stations (Fig. 5). Males were foundonly in LP2soil (6 ind m−2) and TS1 (24 ind m−2),comprising 2% of the total. Stages III–V copepodidscomprised 4% of the total. Forty-four percent of theindividuals were females with egg sacs, 11% females
with attached spermatophores, and 0.5% females withboth egg sac and spermatophores, indicating a mat-ing event. The remaining 39% females carried noegg sacs or attached spermatophores. The sex ratio(males/females) was 0.036.
In the first trial, individuals began to emerge afterday 12, and by days 21–22 at the Taylor Slough sta-tions, and days 22–24 at the Long Pine Key stations,90% of the total emergence had occurred. Densitiesof individuals were 491, 188 and 242 ind m−2 forTS1, TS2 and TS3, respectively, and 97, 176 and 358ind m−2 for LP1, LP2 and LP3, respectively. Theonly cumulative emergence curve that differed signi-
302
Figure 5. Microcyclops rubellus emergence in the first trial. Densities of individuals and cumulative percentages for adults and copepodids atTaylor Slough (top) and Long Pine Key stations (bottom).
Table 3. Results of Kolmogorov-Smirnov tests, cumulative per-centage of Microcyclops rubellus, first trial. In each cell, Z (firstline) and p (second line) are given
LP1soil LP2soil LP3soil TS1 TS2
LP2soil 0.668
0.763
LP3soil 1.871 1.871
0.002 0.002
TS1 0.701 0.436 1.785
0.709 0.991 0.003
TS2 0.757 0.424 1.412 0.378
0.616 0.994 0.037 0.0999
TS3 0.535 0.401 1.604 0.333 0.403
0.938 0.997 0.012 1.000 0.997
ficantly from the others was LP3, when tested versusTS1, TS2, TS3, LP1, and LP2 (Table 3). The num-ber of individuals had a similar trend for all TaylorSlough stations, with a major peak for all reproductive
Table 4. Results of Kolmogorov-Smirnov tests, cumulative percent-age of Microcyclops rubellus, second trial. In each cell, Z (first line)and p (second line) are given
LP1S LP1M LP2S LP2M LP2soil LP3S
LP1M 1.0040.266
LP2S 1.643 0.8220.009 0.509
LP2M 1.643 1.187 1.2780.009 0.123 0.076
LP2soil 3.651 3.651 3.834 4.199<0.001 <0.001 <0.001 <0.001
LP3S 2.556 2.556 2.739 3.104 1.278<0.001 <0.001 <0.001 <0.001 0.076
LP3M 2.921 2.921 3.104 3.469 4.290 3.195<0.001 <0.001 <0.001 <0.001 <0.001 <0.0001
stages around day 18, plus another peak for all classesof females around day 21. Copepodids followed thisgeneral trend (Fig. 5). Long Pine Key sites showeda more complicated pattern, although a peak for all
303
Figure 7. Paracyclops poppei emergence in the second trial. Dens-ities of individuals and cumulative percentages for adults andcopepodids at Long Pine Key stations.
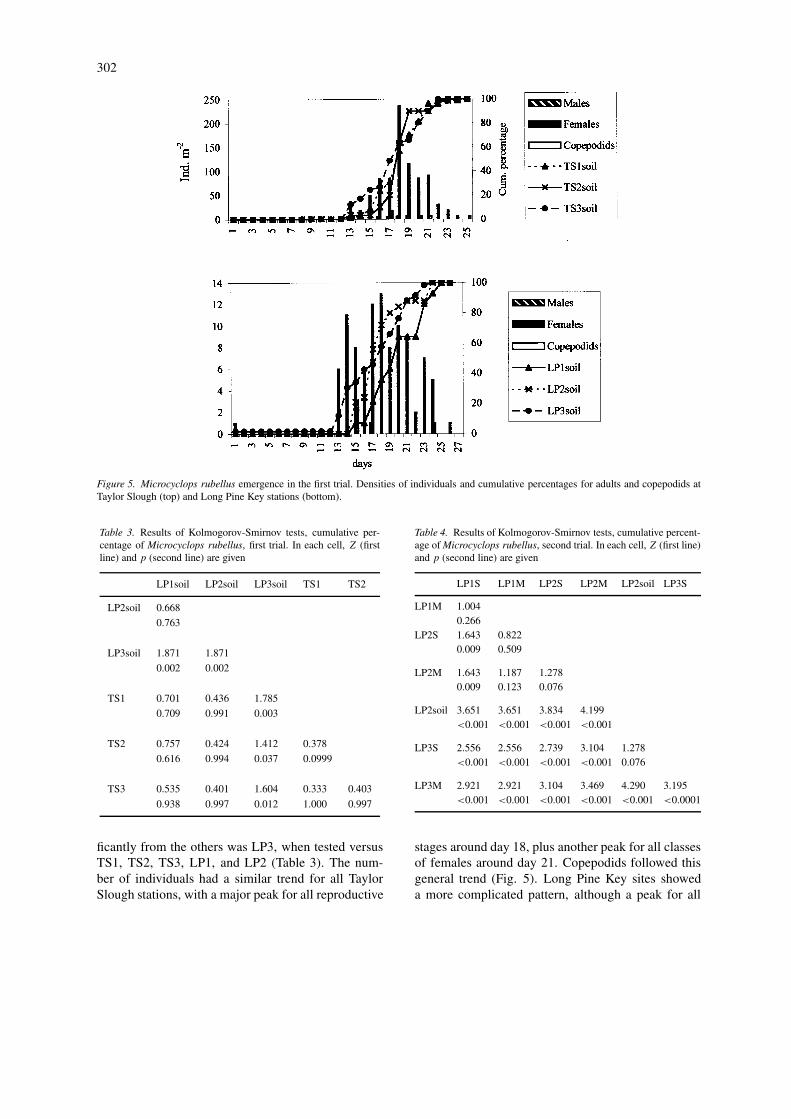
female stages occurred at all stations around days 17–19. Adult females were also numerous around days 12and 19 (Fig. 5).
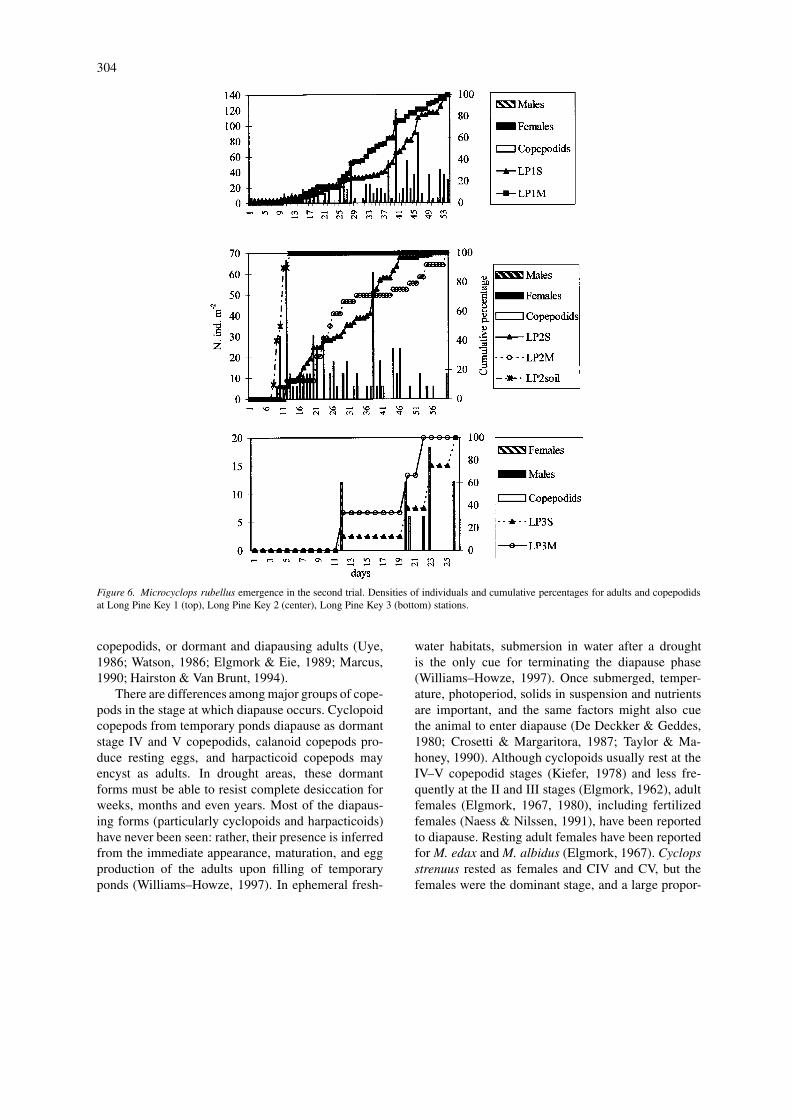
In the second trial, males emerged from all stationsexcept LP3S, but they were always rare compared withfemales (3% of total). Fewer females carried egg sacs(26% of the females) or spermatophores (6%). Sixty-one percent were mature females with no spermato-phores or egg sacs; 4% were stages III–V copepodids.The cumulative curves for the six stations differed sig-nificantly, except for LP1M against LP1S, LP2S andLP2M, LP2M against LP2S, and LP3S against LP2soil(Table 4). The sex ratio of males – females was 0.027.LP1 stations had the highest density of individuals(521 and 497 ind m−2 for the shallow and medium sta-tions), and LP2 stations had 61, 394 and 145 ind m−2
for the soil, shallow, and medium sites, respectively.At LP3, individuals emerged early and in low num-bers. Individuals began to emerge by day 7, and thepopulation grew much slower than in the first trial, asshown in Figure 6. The number of females, represen-ted by mature and ovigerous females, at stations LP1and LP2 was variable but peaked around days 38–40.Males and copepodids occurred in very low numbersat both stations. In surface-soil samples from LP2, thetotal number of animals was very low; a peak occurredbetween days 9 and 13. No copepodids emerged, andonly 6 males m−2 emerged by day 14. At LP3, indi-viduals emerged until day 27, with an irregular pattern,but the maximum number occurred between days 19and 26, with a peak in total females at day 23. Sixmales m−2 and 6 copepodids m−2 emerged as well.
In the third trial, only 42 females m−2 emerged,including 6 ovigerous females and 6 spermatophore-bearing females.
Table 5. Results of Kolmogorov-Smirnov tests, cumulative per-centage of Paracyclops poppei, second trial. In each cell, Z (firstline) and p (second line) are given
LP1M LP2S LP2M
LP2S 0.2647
<0.001
LP2M 2.008 1.187
0.001 0.123
LP3S 4.473 4.473 4.382
<0.001 <0.001 <0.001
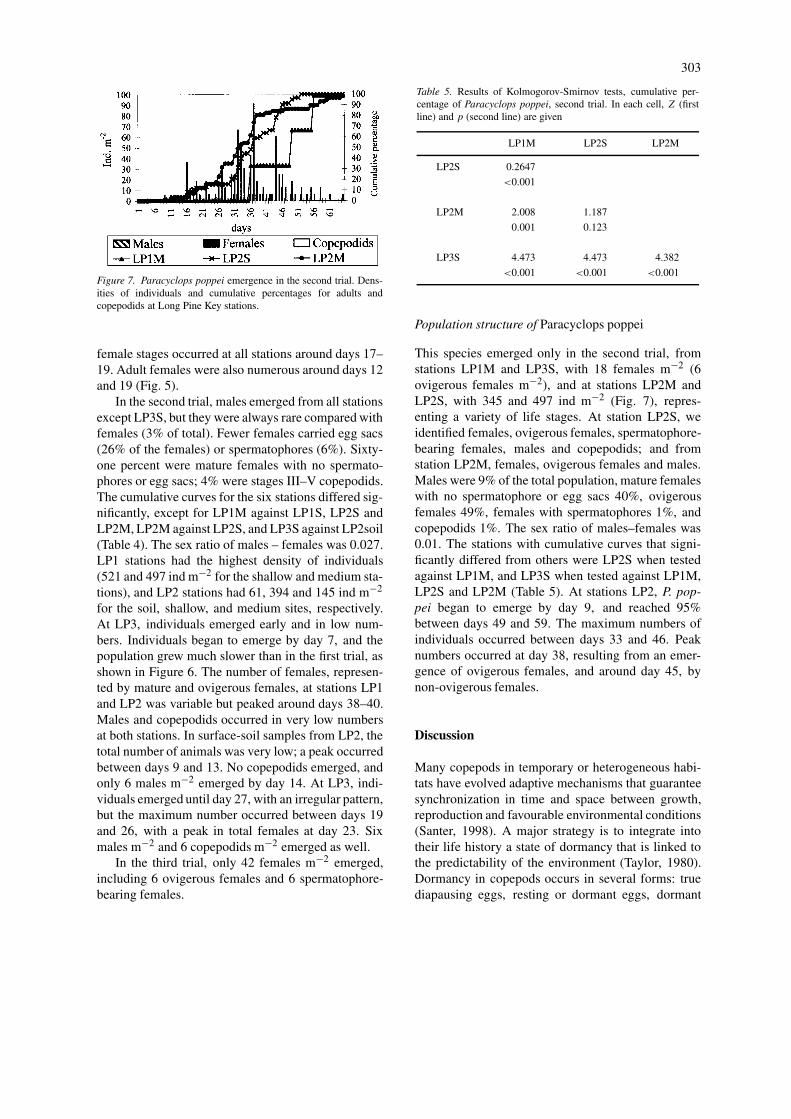
Population structure of Paracyclops poppei
This species emerged only in the second trial, fromstations LP1M and LP3S, with 18 females m−2 (6ovigerous females m−2), and at stations LP2M andLP2S, with 345 and 497 ind m−2 (Fig. 7), repres-enting a variety of life stages. At station LP2S, weidentified females, ovigerous females, spermatophore-bearing females, males and copepodids; and fromstation LP2M, females, ovigerous females and males.Males were 9% of the total population, mature femaleswith no spermatophore or egg sacs 40%, ovigerousfemales 49%, females with spermatophores 1%, andcopepodids 1%. The sex ratio of males–females was0.01. The stations with cumulative curves that signi-ficantly differed from others were LP2S when testedagainst LP1M, and LP3S when tested against LP1M,LP2S and LP2M (Table 5). At stations LP2, P. pop-pei began to emerge by day 9, and reached 95%between days 49 and 59. The maximum numbers ofindividuals occurred between days 33 and 46. Peaknumbers occurred at day 38, resulting from an emer-gence of ovigerous females, and around day 45, bynon-ovigerous females.
Discussion
Many copepods in temporary or heterogeneous habi-tats have evolved adaptive mechanisms that guaranteesynchronization in time and space between growth,reproduction and favourable environmental conditions(Santer, 1998). A major strategy is to integrate intotheir life history a state of dormancy that is linked tothe predictability of the environment (Taylor, 1980).Dormancy in copepods occurs in several forms: truediapausing eggs, resting or dormant eggs, dormant
304
Figure 6. Microcyclops rubellus emergence in the second trial. Densities of individuals and cumulative percentages for adults and copepodidsat Long Pine Key 1 (top), Long Pine Key 2 (center), Long Pine Key 3 (bottom) stations.
copepodids, or dormant and diapausing adults (Uye,1986; Watson, 1986; Elgmork & Eie, 1989; Marcus,1990; Hairston & Van Brunt, 1994).
There are differences among major groups of cope-pods in the stage at which diapause occurs. Cyclopoidcopepods from temporary ponds diapause as dormantstage IV and V copepodids, calanoid copepods pro-duce resting eggs, and harpacticoid copepods mayencyst as adults. In drought areas, these dormantforms must be able to resist complete desiccation forweeks, months and even years. Most of the diapaus-ing forms (particularly cyclopoids and harpacticoids)have never been seen: rather, their presence is inferredfrom the immediate appearance, maturation, and eggproduction of the adults upon filling of temporaryponds (Williams–Howze, 1997). In ephemeral fresh-
water habitats, submersion in water after a droughtis the only cue for terminating the diapause phase(Williams–Howze, 1997). Once submerged, temper-ature, photoperiod, solids in suspension and nutrientsare important, and the same factors might also cuethe animal to enter diapause (De Deckker & Geddes,1980; Crosetti & Margaritora, 1987; Taylor & Ma-honey, 1990). Although cyclopoids usually rest at theIV–V copepodid stages (Kiefer, 1978) and less fre-quently at the II and III stages (Elgmork, 1962), adultfemales (Elgmork, 1967, 1980), including fertilizedfemales (Naess & Nilssen, 1991), have been reportedto diapause. Resting adult females have been reportedfor M. edax and M. albidus (Elgmork, 1967). Cyclopsstrenuus rested as females and CIV and CV, but thefemales were the dominant stage, and a large propor-

305
tion of resting females mated before entering diapause(Naess & Nilssen, 1991). This was the only record ofdiapause in the fertilized female stage.
Several of the Everglades species have been repor-ted to diapause in temporary waters (Table 2). Theseare A. floridanus, A. robustus, E. phaleratus, M. al-bidus, M. latipes, M. rubellus, Microcyclops varicans(G.O. Sars, 1862), O. modestus, T. prasinus, and/or inpermanent ponds: M. fuscus, Mesocyclops americanusDussart, 1985 and M. edax. Moreover, Loftus & Reid(2000) found O. labronectum, M. varicans and M.rubellus emerging from re-hydrated soil from SharkSlough short-hydroperiod sites, and M. rubellus, E.phaleratus and Paracyclops chiltoni (Thomson, 1882)from Shark Slough long-hydroperiod sites. We sur-mise that those planktonic species that did not emergefrom our ENP re-hydrated soil samples either did notdiapause, did not have the appropriate cues for emer-gence, or were missed in our collections because oftheir patchy distribution. It is also possible that, whenwater management practices produce abrupt water-recession rates at the end of the wet season, speciesmight not have time to mate before entering diapause.
One species not previously reported to diapause,D. nearcticus, emerged in our experiments as CV,the most common stage for copepods. Diacyclopsnearcticus was formerly considered a rare, mainlysubterranean species (Reid, 1992b), but is actuallycommon and widespread in stream hyporheic zones(Strayer & Reid, 1999). In ENP, it has been collec-ted mainly in groundwater (Bruno et al., 2000). Thisrecord from surface water, and with resting stages (acommon feature for several congeners; see Williams-Howze, 1997), suggests that this species is euryoe-cious and may have colonized ENP surface waterthrough connections with ground water.
Although reports of dormancy are rare forharpacticoids, the two individuals collected seemed tomatch the normal resting strategies based on the en-cystment of adults. Eight of the 9 resting freshwaterspecies recorded in the literature for permanent wa-ters, and 8 of 10 of the species reported for ephemeralwaters (Williams-Howze, 1997; Santer, 1998), belongto the Canthocamptidae, as do the genera Elaphoidellaand Bryocamptus found in our study. Dormancy in thegenus Elaphoidella has never been reported before,but it has been reported for Bryocamptus washing-tonensis Wilson, 1958 (King et al., 1996) and for anunidentified Bryocamptus sp. (Wiggins et al., 1980).
Most of the other cyclopoids followed the patternof emergence recorded in the literature. Mesocyclops
edax was reported to diapause in nature as CIV and asadult females (Elgmork, 1967; Dobrzykowski & Wyn-gaard, 1993), and in the laboratory as CIII and CIV(Wyngaard, 1988). In the present study, this speciesdiapaused as CV. Tropocyclops prasinus mexicanusemerged as CV, as was reported for T. prasinus (Fisher,1860) (Taylor & Mahoney, 1990). On the other hand,A. robustus, which has been reported emerging as CIV(Champeau, 1971; Vijverberg, 1977; Maier, 1990b),emerged here as adult males and females.
Microcyclops rubellus is known to diapause intemporary habitats (Herbert & Hann, 1986; Taylor& Mahoney, 1990; Wyngaard et al., 1991), but dataon its life cycle and resting strategies have not beenpublished. Our emergence trials suggest a complexstrategy, with individuals undergoing diapause in boththe copepodid and adult phases. In our study, copepo-dids at stages III, IV and V appeared to diapause, evenif in low number. Adult females were dominant, al-though a few adult males diapaused as well. Moreover,many females were probably fertilized prior to dia-pause. The same strategy seemed to be used by P.poppei, evidenced by the high percentage of ovigerousfemales that emerged from diapause. In addition, E.phaleratus emerged as adult females, one of them withegg sacs.
In our study, the sex ratio of the emerging cope-pods was strongly biased towards females, at leastfor the dominant species. Our results agreed with theresults of Dobrzykowski & Wyngaard (1993) for M.edax, which diapaused only as copepodids or adultfemales, and Naess & Nilssen (1991) for Cyclopsstrenuus Fischer, 1851, which diapaused mainly asadult females. Although males and females mightbe expected to be equally abundant in obligate go-nocoric copepods, skewed sex ratios are frequent infreshwater cyclopoids. Diapause can affect sex ratio(Smyly, 1961; George, 1973; Vijverberg, 1977). Ingeneral, more males than females are found duringdiapause (Smyly, 1961; George, 1976) and followingemergence (Maier, 1990b; Williams-Howze, 1997),and it is usually assumed that the early emergenceof a high number of males would allow them tobe readily available to mate with the emerging fe-males (Nilssen, 1980). Dormant females can storesperm, and thus may produce the post-dormant gen-eration more quickly than dormant copepodids (Naess& Nilssen, 1991), as confirmed by the large proportionof females emerging with attached spermatophores orwith egg sacs in our experiments. Sperm can sur-vive and remain functional inside a dormant female.

306
Repeated insemination is not necessary for egg pro-duction and, therefore, males are not necessary forthe survival of the resting populations. Quick re-establishment of a reproducing population gives anadvantage to the emerging species, which can rapidlyrecolonize habitats at the onset of the wet season inENP seasonal marshes.
In the rehydration experiments, two species in par-ticular were dominant, M. rubellus and P. poppei. Inearlier trials by two of us (Loftus & Reid, 2000), M.rubellus was common in both short- and long- hy-droperiod sites (i.e. Taylor Slough and Central SharkSlough) where it was persistent in the soil during theentire dry period. It became less abundant in samplesfollowing the longest dry period. Microcyclops rubel-lus may, therefore, succeed in habitats flooded for alonger time, such as the solution holes, rather thanthe surface soil from both Long Pine Key and TaylorSloughs. The dominance and high total number ofindividuals in our first trial, confirms the dominanceof this species at the beginning of the dry season,as observed by Loftus & Reid (2000). In the secondand third trials, this species became progressivelyless abundant. Although M. rubellus seems to exploitephemeral habitats quite well, there is an apparentlimit to its ability to extend diapause. The second dom-inant species, P. poppei, seemed to be better adaptedto sites that were wet for the longest period of time,such as the solution holes, because it emerged onlyduring the second trial from February to April, andnever from any dry soil sample.
Although we may not have exhaustively sampledemerging copepods, assuming some individuals re-mained buried in the substrate when we filtered waterin the aquaria, it appears that potential bias had min-imal effects on our interpretation of the data. Forexample, once the copepods began to emerge in theaquaria, we collected high number of individuals atthe same development stage only from certain typesof solution holes.
In the first trial, when only surface-soil sampleswere immersed, a lower number of species and in-dividuals emerged, and over a relatively short time.In the second trial, in which copepods emerged al-most exclusively from solution-hole samples, morespecies and individuals emerged, and over a longertime. By the third trial, very few species and individu-als emerged, and no cyclopoids emerged from the soilsamples.
Our results suggest that sites that dried for shorterperiods (such as the shallow and medium depth solu-
tion holes in the second trial) offered better habitats forcopepods. In those samples, cyclopoids emerged aftera longer time but in high numbers of both species andindividuals, with approximately the same temporalemergence pattern. Conversely, lower numbers of spe-cies and individuals were present in the sites that weredry for the longest time (surface soil samples, or allsamples in the third set). Solution holes might sustainmore complex communities than the open waters ofthe marshes, which dried sooner. Comparison betweensites showed that sampling site was responsible forthe main differences in emergence patterns among thestations: there were no significant differences in totalnumbers of copepods between different depth classesin the second trial. Positive correlations between spe-cies richness and both surface area and depth in vernalpools were reported (King et al., 1996), possibly re-lated to hydroperiod and spatial heterogeneity, but theauthors also reported that species composition variedamong pools of the same habitat and among poolsat the same site. In the marshes in ENP, where hy-droperiod acts to structure all animal and vegetationcommunities, predation on copepods can lead to a shiftin species composition, a phenomenon observed inseveral permanent and ephemeral water bodies (Maly,1970; Williamson, 1981, 1983; Wynfield & Town-send, 1983; Brandl & Fernando, 1984; Williamson,1986; Hairston, 1987; Adrian & Frost, 1992; Wickam,1995; Yang, 1996).
The seasonal repetition of flooding and drying inephemeral Everglades habitats might also account forco-occurrence of congeners (King et al., 1996), be-cause crustacean communities in temporary pools arere-established each year from dormant propagules.In the solution holes of the Rocky Glades, re-colonization from resting stages in the soil is prob-ably supplemented by dispersal from ground waters,from aerial propagules, and from permanent wa-ters when surface-water connections occur during thewet season. The management of water affecting theRocky Glades might be responsible for patchy, dis-junct communities of zooplanktonic copepods. Theco-occurrence of all these factors might have led to thedifferent emergence patterns of copepods recorded inthis study for groups of solution holes that were similarin hydroperiod.
The Rocky Glades area of ENP has been partic-ularly affected by drainage and extreme fluctuationsin water level, and this might explain the peculiarcomposition of resting copepod communities, andthe unusual strategies recorded here. The particularly

307
severe conditions occurring in the Rocky Glades mighthave been the selective force leading to new rest-ing strategies in copepods. Since dormant stages arelinked to the predictability of the environment (Taylor,1980), water management operations should be care-fully planned, in order not to exceed the ability ofplanktonic copepods to respond to critical conditions.
Although the study of the composition and lifehistory of the Everglades copepods is recent and willrequire further field and experimental work, our dataprovide information about the occurrence patterns andthe biodiversity of the zooplanktonic copepod com-munities. Detailed data on the ecology of taxa, suchas the copepods, near the base of the marsh foodweb (Rader, 1999), are essential to understand theecosystem changes produced by alterations in naturalhydropatterns. The data will also provide a scientificbasis for guiding and evaluating the difficult processof the restoration of the Everglades.
Acknowledgements
The research was supported by a South Florida Crit-ical Ecosystem Science Initiative (CESI) funding tothe Principal Investigator, Dr S. A. Perry. Fundingfor M. C. Bruno was provided through a Cooperat-ive Agreement with Florida International University,with the assistance of Dr Kelsey Downum. We wish tothank the Everglades National Park volunteers DanielLevitis and Thomas Muller, and U.S. Geological Sur-vey technician Victoria Foster, for helping to collectthe field samples and to monitor the aquarium immer-sion trials. Freddie James, Everglades National Park,provided the hydroperiod data. Two anonymous refer-ees improved the manuscript with their comments.
References
Adrian, R. & T. M. Frost, 1992. Comparative feeding ecology ofTropocyclops prasinus mexicanus (Copepoda, Cyclopoida). J.Plankton Res. 14: 1369–1382.
Brandl, Z. & C. H. Fernando, 1986. Feeding and food consumptionby Mesocyclops edax. Syllogeus 58: 254–258.
Bruno, M. C., J. W. Reid & S. A. Perry, 2000. New records of cope-pods from Everglades National Park (Florida): description of twonew species of Elaphoidella (Harpacticoida, Canthocamptidae),and supplementary description of Diacyclops nearcticus Kiefer(Cyclopoida, Cyclopidae). Crustaceana 73: 1171–1205.
Champeau, A., 1970a. Étude de la vie latente chez les calanoides(Copépodes) characteristiques des eaux temporaires de Basse-Provence. Ann. Fac. Sci. Marseille 44: 155–189.
Champeau, A., 1970b. Recherches sur l’écologie et l’adaptation àla vie latente des copépodes des eaux temporaires provençaleset corses. Thèse, Faculté des Sciences, Université Aix-en-Provence, Marseilles: 360 pp.
Champeau, A., 1971. Recherches sur l’adaptation à la vie latente descopépodes cyclopoïdes et harpacticoïdes des eaux temporairesprovençales. Bull. Soc. Écol. 2: 151–167.
Crosetti, D. & F. G. Margaritora, 1987. Distribution and life cyclesof cladocerans in temporary pools from central Italy. Freshwat.Biol. 18: 165–175.
De Deckker, P. & M. C. Geddes, 1989. Seasonal fauna of ephemeralsaline lakes near the Coorong Lagoon, South Australia. Aust. J.mar. Freshwat. Res. 31: 677–699.
De Stasio, B. T. Jr., 1990. The role of dormancy and emergencepatterns in the dynamics of a freshwater zooplankton community.Limnol. Oceanogr. 35: 1079–1090.
Dobrzykowsky, A. E. & G. A. Wyngaard, 1993. Phenology ofdormancy in a Virginia population of Mesocyclops edax (Crusta-cea: Copepoda). Hydrobiologia 250: 167–171.
Elgmork, K., 1959. Seasonal occurrence of Cyclops strenuusstrenuus in relation to environment in small water bodies insouthern Norway. Folia limnol. scand. 11: 1–196.
Elgmork, K., 1962. A bottom resting stage in the planktonicfreshwater copepod Cyclops scutifer Sars. Oikos 13: 306–310.
Elgmork, K., 1967. Ecological aspects of diapause in copepods.Proc. Symp. Crust. 3: 947–954.
Elgmork, K., 1980. Evolutionary aspects of diapause in freshwatercopepods. In Kerfoot, W. C. (ed.), Evolution and Ecology of Zo-oplankton Communities. The University Press of New England,Hanover (N.H.): 411–417.
Elgmork, K. & J. A. Eie, 1989. Two- and three-year life cycles inthe planktonic copepod Cyclops scutifer in two high mountainslakes. Holarct. Ecol. 12: 60–69.
George, D. G., 1973. Diapause in Cyclops vicinus. Oikos 24: 136-142.
George, D. G., 1976. Life cycle of Cyclops vicinus in a shalloweutrophic reservoir. Oikos 27: 101–110.
Gleason, P. J. & P. Stone, 1994. Age, origin and evolution of theEverglades peatlands. In Davis, S. M. & J. C. Odgen (eds),Everglades: the Ecosystem and Its Restoration. St. Lucie Press,Delray Beach, Florida: 149–197.
Gunderson, L. H., 1994. Vegetation of the Everglades: Determinantsof community composition. In Davis, S. M. & J. C. Odgen (eds),Everglades: the Ecosystem and Its Restoration. St. Lucie Press,Delray Beach, Florida: 323–340.
Gunderson, L. H. & W. F. Loftus, 1993. The Everglades. In Martin,W. H., S. G. Boyce & A. C. Echternacht (eds), Biodiversity ofthe Southeastern United States: Terrestrial Communities. Wiley& Sons, New York, NY: 199–255.
Hairston, N. G. Jr., 1987. Diapause as a predator-avoidance adapt-ation. In Kerfoot, W. C. & A. Sih (eds), Predation: Direct andIndirect Impacts on Aquatic Communities. The University Pressof New England, Hanover, New Hampshire: 281–290.
Hairston, N. G. Jr. & R. A. Van Brunt, 1994. Diapause dynamicsof two diaptomid copepod species in a large lake. Hydrobiologia292/293: 209–218.
Hebert, P. D. N. & B. J. Hann, 1986. Patterns in composition ofArctic tundra pond microcrustacean communities. Can. J. Fish.aquat. Sci. 43: 1416–1425.
Hoffmeister, J. E., 1974. Land from the Sea. University of MiamiPress, Coral Gables, FL, 143 pp.
Kiefer, F., 1978. Das Zooplankton der Binnengewässer: FreilebendeCopepoda. In Elster, H. -J. & W. Ohle (eds), DieBinnengewässer, Bd. 16, Teil 2. Schweizerbart, Stuttgart: 1–343.

308
King, J. L., M. A. Simovich & R. C. Brusca, 1996. Species richness,endemism and ecology of crustacean assemblages in northernCalifornia vernal pools. Hydrobiologia 328: 85–116.
Loftus, W. F. & A. M. Eklund, 1994. Long-term dynamics of anEverglades small-fish assemblage. In Davis, S. M & J. C. Odgen(eds), Everglades: the Ecosystem and Its Restoration. St. LuciePress, Delray Beach, Florida: 461–483.
Loftus, W. F. & J. A. Kushlan, 1987. Freshwater fishes of southernFlorida. Bull. Florida St. Mus., biol. Sci. 31: 147–344.
Loftus, W. F. & J. W. Reid, 2000. Copepod (Crustacea) emergencefrom soils from Everglades marshes with different hydroperiods.J. Fresh. Ecol. 15: 515–523.
Loftus, W. F., J. D. Chapman & R. Conrow, 1990. Hydroperiodeffects on Everglades marsh food webs, with relation to marshrestoration efforts. In Larson, G. & M. Soukup (eds), Fisheriesand Coastal Wetland Research. Proceedings of the Conference onScience in the National Parks, Vol. 6. Colorado State University,Ft. Collins, July 1986: 1–22.
Loftus, W. F., R. A. Johnson & G. H. Anderson, 1992. Ecolo-gical impacts of the reduction of groundwater levels in short-hydroperiod marshes of the Everglades. In Stanford, J. A. &J. J. Simons (eds), Proceedings of the First International Con-ference on Groundwater Ecology. American Water ResourcesAssociation, Bethesda, Maryland: 199–208.
Maier, G., 1990a. The seasonal dynamics of Thermocyclops dy-bowskii (Lande, 1890) in a small pond (Copepoda, Cyclopoida).Crustaceana 59: 76–81.
Maier, G., 1990b. Coexistence of the predatory cyclopoids Acantho-cyclops robustus (Sars) and Mesocyclops leuckarti (Claus) in asmall eutrophic lake. Hydrobiologia 198: 185–203.
Maier, G., 1994. Patterns of life history among cyclopoid copepodsof central Europe. Freshwat. Biol. 31: 77–86.
Maly, E. J., 1970. The influence of predation on the adult sex ratiosof two copepod species. Limnol. Oceanogr. 15: 566–573.
Marcus, N. H., 1990. Calanoid copepod, cladoceran, and rotifereggs in sea-bottom sediments of northern Californian coastalwaters: identification, occurrence and hatching. Mar. Biol. 105:413–465.
Naess, T. & J. P. Nilssen, 1991. Diapausing fertilized adults: a newpattern of copepod life cycle. Oecologia 86: 368–371.
Nasci, R. S., S. G. F. Hare & M. Vecchione, 1987. Habitat associ-ations of mosquito and copepod species. J. am. Mosquito Cont.Ass. 3: 593–600.
Nilssen, J. P., 1980. When and how to reproduce: a dilemma forlimnetic cyclopid copepods. In Kerfoot, W. C. (ed.), Evolutionand Ecology of Zooplankton Communities. The University Pressof New England, Hanover, New Hampshire: 418–424.
Olmsted, I. C., L. L. Loope & R. E. Rintz, 1980. A survey andbaseline analysis of aspects of the vegetation of Taylor Slough,Everglades National Park. South Florida Research Center Tech.Rep. 7–586. Everglades National Park, Homestead, Florida: 24pp.
Rader, R. R., 1999. The Florida Everglades. Natural variability, in-vertebrate diversity, and foodweb stability. In Batzer, D. P., R.R. Rader & S. A. Wissinger (eds), Invertebrates in FreshwaterWetlands of North America: Ecology and Management. Wileyand Sons, Incorporated, New York, NY, U.S.A.: 25–53.
Reid, J. W., 1985. Chave de identificação e lista de referências bib-liográficas para as espécies continentais sulamericanas de vidalivre da ordem Cyclopoida (Crustacea, Copepoda). Bolm Zool.,Univ. S. Paulo 9: 17–143.
Reid, J. W., 1989. The distribution of the species of the genus Ther-mocyclops (Copepoda, Cyclopoida) in the western hemisphere,with description of T. parvus, new species. Hydrobiologia 175:149–174.
Reid, J. W., 1992a. Copepoda (Crustacea) from fresh waters ofthe Florida Everglades, U.S.A., with a description of Eucyclopsconrowae n. sp. Trans. am. microsc. Soc. 111: 229–254.
Reid, J. W., 1992b. Redescription of Diacyclops nearcticus (Kiefer,1934) and description of four similar new congeners from NorthAmerica, with comments on D. crassicaudis (G.O. Sars, 1863)and D. crassicaudis var. brachycercus (Kiefer, 1927) (Crustacea:Copepoda). Can. J. Zool. 70: 1445–1469.
Roy, J., 1932. Copépodes et cladocères de l’ouest de la France.Recherches biologiques et faunistiques sur le plancton d’eaudouce des vallées du Loir et de la Sarthe. Thèse, University ofParis: 222 pp.
Rzoska, J., 1961. Observations on tropical rainpools and generalremarks on temporary waters. Hydrobiologia 17: 265–287.
Santer, B., 1998. Life cycle strategies of free-living copepods infresh waters. J. mar. Syst. 15: 327–336.
Smyly, W. J. P., 1961. The life cycle of the freshwater copepodCyclops leuckarti Claus in Esthwaite Water. J. anim. Ecol. 30:153–169.
Smyly, W. J. P., 1970. Observations on the rate of develop-ment, longevity and fecundity of Acanthocyclops viridis (Jurine)(Copepoda, Cyclopoida) in relation to type of prey. Crustaceana18: 21–36.
Strayer, D. L. & J. W. Reid, 1999. Distribution of hyporheic cyclop-oids (Crustacea: Copepoda) in the eastern United States. Arch.Hydrobiol. 145: 79–92.
Taylor, F., 1980. Optimal switching to diapause in relation to theonset of winter. Theor. pop. Biol. 18: 125–133.
Taylor, B. E. & D. L. Mahoney, 1990. Zooplankton in RainbowBay, a Carolina Bay pond: population dynamics in a temporaryhabitat. Freshwat. Biol. 24: 597–612.
Uye, S., 1986. Resting egg production as a life history strategy ofmarine planktonic copepods. Bull. mar. Sci. 37: 440–449.
Vijverberg, J., 1977. Population structure, life histories and abund-ance of copepods in Tjeukemeer, The Netherlands. Freshwat.Biol. 7: 579–597.
Watson, N. H. F., 1986. Variability of diapause in copepods.Syllogeus 58: 509–512.
Wickham, S., 1995. Cyclops predation on ciliates: species-specificdifferences and functional responses. J. Plankton Res. 17: 1633–1646.
Wiggins, G. B., R. J. Mackay & I. M. Smith, 1980. Evolutionaryand ecological strategies of animals in annual temporary pools.Arch. Hydrobiol. Suppl. 58: 97–206.
Williams-Howze, J., 1997. Dormancy in the free-living copepodorders Cyclopoida, Calanoida and Harpacticoida. Ann. Rev.Oceanogr. mar. Biol. 35: 257–321.
Williamson, C. E., 1981. Foraging behaviour of a freshwater cope-pod: Frequency changes in looping behaviour at high and lowprey densities. Oecologia 50: 332–336.
Williamson, C. E., 1983. Behavioural interactions between acyclopoid copepod predator and its prey. J. Plankton Res. 5:701–711.
Williamson, C. E., 1986. The swimming and feeding behaviour ofMesocyclops edax. Hydrobiologia 134: 11–19.
Wyngaard, G. A., 1988. Geographical variation in dormancy ina copepod: evidence from population crosses. Hydrobiologia167/168: 367–374.
Wyngaard, G. A., B. E. Taylor & D. L. Mahoney, 1991. Emer-gence and dynamics of cyclopoid copepods in an unpredictableenvironment. Freshwat. Biol. 25: 219–232.
Yang, Y. F. & Z. Brandl, 1996. Feeding of Acanthocyclops robustuson zooplankton. Chin. J. Oceanol. Limnol. 14: 17–26.
Related Documents











