1992;52:6297-6304. Cancer Res Ashutosh Banerjee, Hong-Ji Xu, Shi-Xue Hu, et al. Types by Microcell Transfer of Chromosome 13 Cell of Retinoblastoma Gene Function in Various Human Cancer Changes in Growth and Tumorigenicity following Reconstitution Updated version http://cancerres.aacrjournals.org/content/52/22/6297 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on January 16, 2014. © 1992 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on January 16, 2014. © 1992 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1992;52:6297-6304. Cancer Res Ashutosh Banerjee, Hong-Ji Xu, Shi-Xue Hu, et al. Types by Microcell Transfer of Chromosome 13
Cellof Retinoblastoma Gene Function in Various Human Cancer Changes in Growth and Tumorigenicity following Reconstitution
Updated version
http://cancerres.aacrjournals.org/content/52/22/6297
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 52, 6297-Õ304. November 15, 1992]
Changes in Growth and Tumorigenicity following Reconstitution of
Retinoblastoma Gene Function in Various Human Cancer CellTypes by Microcell Transfer of Chromosome 131
Ashutosh Banerjee, Hong-Ji Xu, Shi-Xue Hu, Diana Araujo, Rei Takahashi, Eric J. Stanbridge, andWilliam F. Benedict2
Center for Biotechnology, Baylor College of Medicine, The Woodlands, Texas 77381 ¡A.B., H-J. X., S-X. H., R. T., W. F. B.J, and Department of Microbiology andMolecular (ienetics, College of Medicine, University of California, Irvine, California 92717 (D. A., E. J. S.J
ABSTRACT
Functional loss of the retinoblastoma (RB) gene has been implicatedin the initiation or progression of several human tumor types includingcancer of the eye, bone, bladder, and prostate. To examine the consequence of adding one Rii alÃelecontaining its normal regulatory elements back into representative examples of each of these cancer types,as well as to compare the results to those previously reported usingvarious RB complementary DNA constructs, a neomycin resistantmarked 13 chromosome «astransferred by microcell fusion. Severalattempts to obtain RB positive osteosarcoma cells failed. In addition,only one RB positive retinoblastoma clone was isolated. This clonecontained many large cells, could not be maintained in long-term culture, and produced only RB negative tumors. Three RB positive bladdercancer cell clones were obtained, all of which grew slower in culture thantheir RB negative parental counterpart and did not form colonies in softagar. Tumorigenicity was markedly suppressed in these clones. Oneclone yielded no tumors, and the other 2 clones produced only one smalltumor each, both of which were RB negative. In contrast, the 2 RBpositive prostate cancer cell clones isolated had no differences in theircell culture growth properties, including growth in soft agar compared tothe parental cells. One of the clones was nontumorigenic, while the otherclone produced 4 small tumors, all of which were RB positive. Theseresults indicate that the transfer of one Rii alÃeleby microcell transferproduces different levels of growth inhibition as well as tumor suppression, depending on the cell type examined. In the case of prostate cancer,the function of the HI! gene in tumor suppression appears to be independent from its growth regulatory function, since no growth inhibitionin cell culture was noted in these cells, although tumor suppression wassignificant.
INTRODUCTION
The RB3 gene represents the first recessive human cancersusceptibility gene isolated (1-3). Loss of both RB alÃelesis acritical step in the development of retinoblastoma, togetherwith being one of the genetic factors involved in the initiationand/or progression of several other human cancers, includingosteosarcoma (1,2, 4-7), and carcinomas of the bladder (8-10)and prostate (11, 12). It was proposed, based on early cytoge-netic and molecular biological studies, that the functional lossof both RB alÃelesis responsible for retinoblastoma and wasthus termed a tumor "suppressor" gene (13). Recently, RB
cDNA constructs have been transferred through retroviral in-
Received 6/1/92; accepted 9/11/92.The costs of publication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This study was supported by NIH Grants EYO6195 and CA29401, a grantfrom the Retina Research Foundation, and The Council for Tobacco Research-United States of America.
2 To whom requests for reprints should be addressed, at the Center for Biotechnology, Baylor College of Medicine, 4000 Research Forest Drive, The Woodlands,TX 77381.
3 The abbreviations used are: RB, retinoblastoma; DMEM, Dulbecco's minimal
essential medium; FBS, fetal bovine serum; cDNA, complementary DNA.
fection or plasmid transfection into RB negative retinoblastoma, osteosarcoma, bladder, and prostate cell lines (11, 14-22). Although in general the results of these studies indicatethat there is both a significant growth inhibition in culture andtumor suppression following reintroduction of the RB cDNAinto such RB negative cancer cells, the results have not beentotally consistent, and it has been difficult to unequivocallyseparate the effects of RB expression on growth inhibition fromthat of tumor suppression. In addition, the differences observedmay reflect the type of promoter chosen or method of RBtransfer used.
The RB gene has been successfully transferred thus far onlyusing artificial constructs that included either the Moloney murine leukemia virus long terminal repeat promoter (11, 14-16,18, 19, 22), the ß-actinpromoter (17), the mouse metallothio-nein promoter (20), or the SV40 early promoter (21). We believed that it would be important, therefore, to compare theresults using such constructs to those obtained from transferring a normal RB alÃelein the presence of its natural promoterand other regulatory elements. This was accomplished by themicrocell fusion of a normal 13 chromosome, which also contained a neomycin selectable marker. Indeed, although manysimilarities were observed, some significant differences werefound between the results obtained using the chromosome 13 totransfer the RB alÃelerather than retroviral infection or plasmidtransfection of the RB cDNA.
MATERIALS AND METHODS
Cells Used for Microcell Fusion. The mouse-human microcell hybrid, MCH 204-4, which contained a single neomycin resistant markedchromosome 13 as its only human chromosome (23), was grown incomplete medium containing 200 Mg/ml of G418. The recipient cellsused were as follows: Y79, a retinoblastoma suspension cell line;Saos-2, an osteosarcoma cell line; HTB9 (5637), a bladder carcinomacell line; and DU 145, a prostate carcinoma cell line. All were obtainedfrom the American Type Culture Collection. They were maintained inDMEM with 10% FBS, supplemented with 1% penicillin and streptomycin.
Microcell Fusion and Hybrid Selection. The donor MCH 204-4cells were treated with Colcemid at a final concentration of 0.5 Mg/mlfor 48 h. Microcells were then obtained as described previously (24).Recipient Y-79 cells (1 x IO6) were centrifuged and washed with me
dium containing no serum. The cells were then recentrifuged in a flatbottomed centrifuge tube and exposed to the microcells in 2 ml ofphytohemagglutinin P (50 Mg/ml;Difco Laboratory, Detroit, MI) for 20min in an incubator at 37°C.The cells were then recentrifuged, the
supernatant removed, and 1 ml of polyethylene glycol (50% PEG 400)was added for 1 min. The cells were then washed twice with serum-freemedium and grown in complete medium for 24 h. Control Y-79 cellswere treated similarly except that no microcells were added. Both control Y-79 cells and Y-79 cells following microcell fusion were thengrown in regular medium or medium containing 300 or 600 Mg/ml of
6297
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
G418. The only cells that survived 2 weeks of G418 treatment were theY-79 cells, which had undergone microcell fusion and were selected in300ng/ml of G418. This mass culture was designated Y-79/13 and wasused for the further studies described in the text. The monolayer HTB9,DU 145, and Saos-2 recipient cells were treated similarly except that thepurified microcells were resuspended in 2 ml of serum-free DMEMcontaining 100/ig/ml of phytohemagglutinin. The microcells were thenattached to prewashed recipient cell monolayers by incubating for 15min at 37°C.The cells were fused by treatment with 2 ml of polyeth
ylene glycol for 60 s. They were then immediately rinsed 3 times inphosphate-buffered saline to remove excess polyethylene glycol andincubated in DMEM plus 10% FBS for 2 days. The cells were thentrypsinized and split into 3 100-mm tissue culture dishes containingDMEM and 10% FBS plus 200^g/ml ofG418. The resulting microcellhybrids were subsequently isolated and expanded. These microcell hybrids (3 from HTB9 fusion and 2 from DU 145 fusion) were thenexamined for their RB protein status and presence of the introducedchromosome 13. All cell lines were tested frequently for Mycoplasmacontamination and were negative.
Morphological Studies. Y-79 and Y-79/13 cells were grown in60-mm tissue culture dishes. They were examined a minimum of twiceweekly in both regular and G418 containing medium and photographedweekly using an inverted phase photomicroscope at x200. Other hybrids were examined also twice weekly using an inverted microscope.
Examination of the RB Protein. Immunoprecipitation was done aspreviously described using the RB-WL-1 antibody (25). For Y-79 singlecell studies, a drop of cells was placed in the center of a poly-i.-lysinecoated coverslip. The cells were allowed to attach for approximately 30min. They were then fixed with 10% formaldehyde in 0.1 Mphosphatebuffer. After rinsing in phosphate buffered saline, the coverslips wereblocked at room temperature for 4 h with 100 n\ of phosphate buffercontaining 5% bovine serum albumin and 4% normal goat serum, followed by adding the RB-WL-1 antibody to a final concentration of 8Mg/mland continuing the incubation overnight. A goat anti-rabbit IgG-.11kali IK-phosphatase conjugated antibody (1:1000 diluted in blocking
buffer) was used as the second antibody, and the color detection wasperformed using the Proto Blot AP system (Promega). Single cell staining of the monolayer cells was done as described previously (17).
Chromosomal Analysis. The cytogenetic status of the HTB9,DU 145, and microcell hybrids was evaluated in a minimum of 20metaphases each following Giemsa-trypsin banding as described previously (26). Since neither HTB9 nor DU 145 cells contain a normalchromosome 13 (27, 28), the presence of a normal chromosome 13 inthe microcell hybrids was considered conclusive evidence for the transfer of the normal 13 from the donor MCH 204-4 cells.
Intraocular and s.c. Injections. Y-79 and Y-79/13 cells were centri-fuged and resuspended in 1 ml of complete medium. Four- to 8-week-old nu/nu (athymic) mice were then given injections of these cells in theanterior chamber of the eye as described previously (29, 30). The micewere then examined under anesthesia twice weekly using a dissectingmicroscope. The injection was recorded as positive as soon as tumorswere visible. To obtain intraocular tumor cells from Y-79/13 injectedmice, they were sacrificed and the orbit was removed intact. The orbitwas then rinsed in 70% alcohol and placed in complete medium. It wasthen opened with a needle yielding a suspension of almost pure tumorcells. Tumorigenicity of the HTB9, DU 145, and microcell hybrids wasdetermined by s.c. injections of 1 x IO7 cells in the nu/nu mice as
described previously (26). The mice were then examined weekly for theappearance of tumors.
RESULTS
Retinoblastoma. Six separate attempts in 2 laboratories(W. B. and E. S.) were made to obtain RB positive retinoblas-toma cells by microcell fusion. On only one occasion was chromosome 13 transfer successful. In this case, following microcellfusion of Y-79 cells, the mass culture was grown in regularmedium for 24 h and then placed in selective medium contain
ing 300 or 600 Mg/ml of G418. No viable cells were seen after 2weeks in control Y-79 cell cultures at either concentration ofG418 or in the cell cultures after microcell fusion followed byselection in 600 Mg/ml of G418. A few viable cells, however,were observed in one of the dishes containing 300 Mg/ml ofG418. These appeared as small clumps of less than 10 cells,each having some cells 2 to several times larger than the sizeof normal Y-79 cells. A few individual clumps were placedinto separate wells of a microplate by pipetting them underdirect microscopic observation. All wells were inspected immediately after pipetting to ensure that the viable cells had beentransferred.
Regular medium was then added to each well. Three majordifferences were initially observed in these cells compared to theY-79 parental cells, (a) Although large cells are occasionallyseen in Y-79 cultures, the majority of the population was com
posed of uniformly smaller cells rapidly growing in clumps orchains. However, approximately 50% of the cells selected aftermicrocell fusion ranged in size from 2 to 4 times or more thanthe normal size of Y-79 cells, (b) A significantly higher numberof cells attached to the dish compared to the parental Y-79 cells,which were also placed in microcells at a similar concentration.(c) The cells proliferated at a markedly slower rate than theparental cells.
Once a sufficient number of cells were available (approximately 2 months after microcell fusion), a portion of the massculture was examined for the presence of normal RB protein atthe single cell level. The majority of cells subsequently termedY-79/13 showed RB nuclear staining (Fig. \A). The typicalnormal protein RB pattern (24) also was detectable in theY-79/13 cells by immunoprecipitation (Fig. 2, Lane 3), whereasno normal RB protein was present in the Y-79 parental cells.
The Y-79/13 cells were then maintained on regular mediumfor approximately an additional month, while a second cultureremained on 200 Mg/ml of G418. Subsequently, the cells wereexamined for their tumorigenic potential following intraocularinjection into nu/nu mice. At the time of injection, a portion ofthe cells was again studied for RB content at the single cell level.Although most of the cells still showed normal nuclear RBstaining whether or not the cells had the typical Y-79 cell sizeor were larger (Fig. \B), clumps of Y-79 size cells were observedthat had no RB protein staining (Fig. \B, arrows). The proportion of large to small cells at the time of intraocular injection isshown in Fig. 1C. As illustrated in Table 1, these cells were lesstumorigenic than the parental Y-79 cells and had a longer latency period. All tumors within a given group occurred within1 week of each other and each Y-79 tumor appeared on thesecond week after injection. Once a Y-79/13 tumor was observed, it progressed as rapidly as the Y-79 controls and couldnot be distinguished histologically from Y-79 tumors. In addition, when the Y-79/13 tumors were removed, they containedprimarily cells of the parental Y-79 size, none of which was RBpositive (Fig. ID). These results strongly imply that Y-79 cellsin which a chromosome 13 had been transferred and normal RBprotein produced no longer had the ability to form intraoculartumors. However, we were not able to clearly distinguishgrowth inhibition from tumor suppression in this system, sinceby 5 months in culture following initial microcell fusion no RBpositive Y-79/13 could be found. Consequently, considerablegrowth inhibition must have occurred in cell culture similar tothat found in vivo.
6298
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from
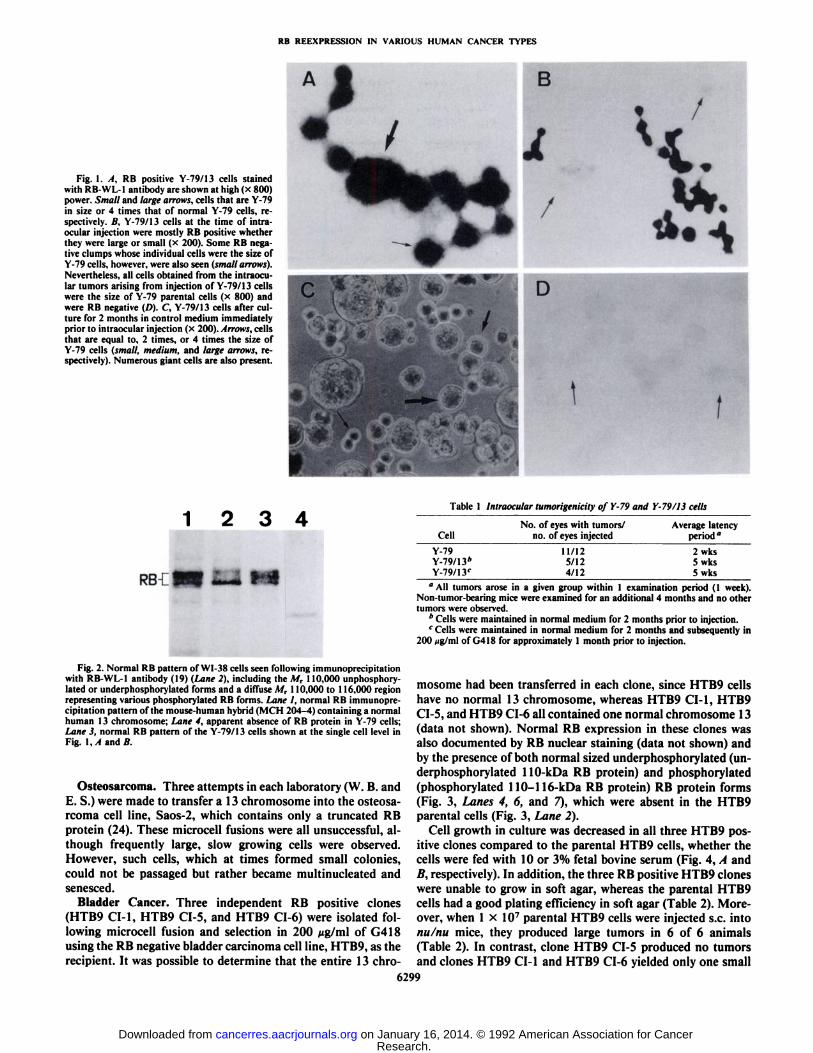
RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
Fig. 1. A, RB positive Y-79/13 cells stainedwith RB-WL-1 antibody are shown at high (x 800)power. Small and large arrows, cells that are Y-79in size or 4 times that of normal Y-79 cells, respectively. B, Y-79/13 cells at the time of intraocular injection were mostly RB positive whetherthey were large or small (x 200). Some RB negative clumps whose individual cells were the size ofY-79 cells, however, were also seen (small arrows).Nevertheless, all cells obtained from the intraocular tumors arising from injection of Y-79/13 cellswere the size of Y-79 parental cells (x 800) andwere RB negative (D). C, Y-79/13 cells after culture for 2 months in control medium immediatelyprior to intraocular injection (x 200). Arrows, cellsthat are equal to, 2 times, or 4 times the size ofY-79 cells (small, medium, and large arrows, respectively). Numerous giant cells are also present.
1234Table 1 Intraocular tumorigenicity of Y-79 and Y-79/13 cells
RBCI
Fig. 2. Normal RB pattern of WI-38 cells seen following immunoprecipitationwith RB-WL-1 antibody (19) (Lane 2), including the M, 110,000 unphosphory-lated or underphosphorylated forms and a diffuse M, 110,000 to 116,000 regionrepresenting various phosphorylated RB forms. Lane I, normal RB immunoprecipitation pattern of the mouse-human hybrid (M( 11204-4) containing a normalhuman 13 chromosome; Lane 4, apparent absence of RB protein in Y-79 cells;Lane 3, normal RB pattern of the Y-79/13 cells shown at the single cell level inFig. 1, I ami //
Osteosarcoma. Three attempts in each laboratory (W. B. andE. S.) were made to transfer a 13 chromosome into the osteosa-rcoma cell line, Saos-2, which contains only a truncated RBprotein (24). These microcell fusions were all unsuccessful, although frequently large, slow growing cells were observed.However, such cells, which at times formed small colonies,could not be passaged but rather became multinucleated andsenesced.
Bladder Cancer. Three independent RB positive clones(HTB9 CI-1, HTB9 CI-5, and HTB9 CI-6) were isolated following microcell fusion and selection in 200 /ug/ml of G418using the RB negative bladder carcinoma cell line, HTB9, as therecipient. It was possible to determine that the entire 13 chro-
CellNo. of eyes with tumors/
no. of eyes injectedAverage latency
period"
Y-79Y-79/13*Y-79/1y
11/125/124/12
2 wks5 wks5 wks
"All tumors arose in a given group within 1 examination period (1 week).Non-tumor-bearing mice were examined for an additional 4 months and no othertumors were observed.
" Cells were maintained in normal medium for 2 months prior to injection.r Cells were maintained in normal medium for 2 months and subsequently in
200 ng/ml of G418 for approximately 1 month prior to injection.
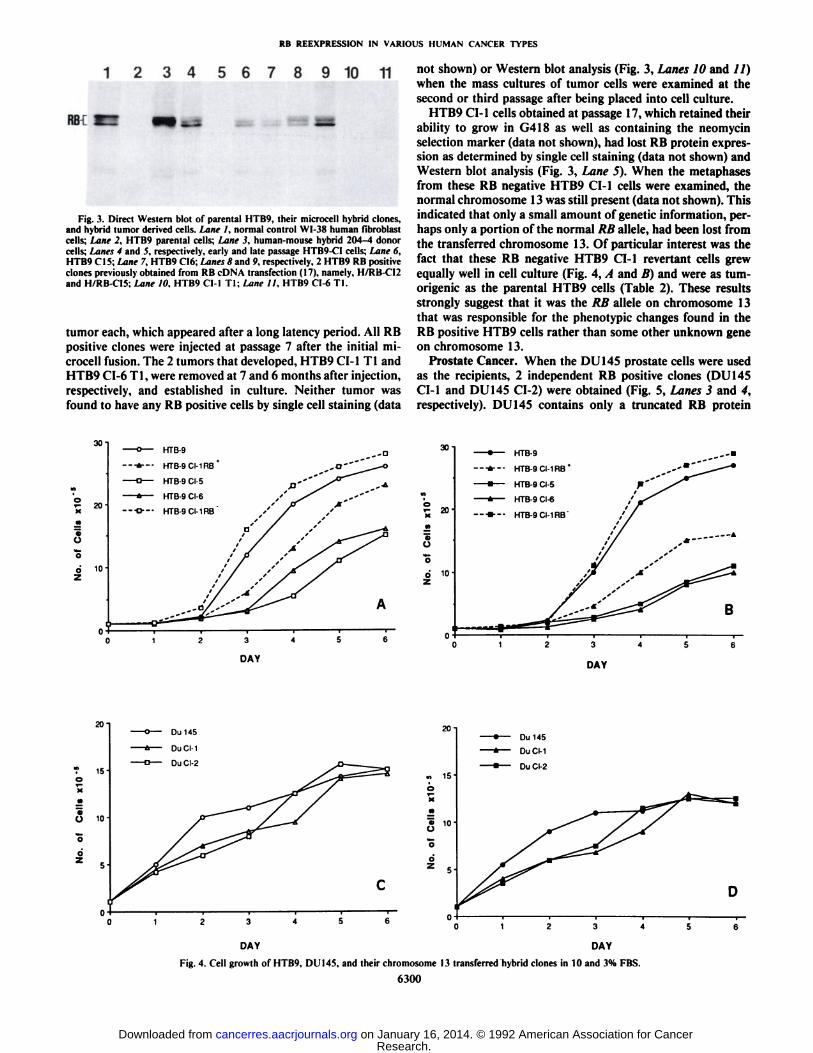
mosome had been transferred in each clone, since HTB9 cellshave no normal 13 chromosome, whereas HTB9 CI-1, HTB9CI-5, and HTB9 CI-6 all contained one normal chromosome 13(data not shown). Normal RB expression in these clones wasalso documented by RB nuclear staining (data not shown) andby the presence of both normal sized underphosphorylated (underphosphorylated 110-kDa RB protein) and phosphorylated(phosphorylated 110-116-kDa RB protein) RB protein forms(Fig. 3, Lanes 4, 6, and 7), which were absent in the HTB9parental cells (Fig. 3, Lane 2).
Cell growth in culture was decreased in all three HTB9 positive clones compared to the parental HTB9 cells, whether thecells were fed with 10 or 3% fetal bovine serum (Fig. 4, A andA, respectively). In addition, the three RB positive HTB9 cloneswere unable to grow in soft agar, whereas the parental HTB9cells had a good plating efficiency in soft agar (Table 2). Moreover, when 1 x IO7 parental HTB9 cells were injected s.c. into
nu/nu mice, they produced large tumors in 6 of 6 animals(Table 2). In contrast, clone HTB9 CI-5 produced no tumorsand clones HTB9 CI-1 and HTB9 CI-6 yielded only one small
6299
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from
RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
1 234 56789 10 11
RB
Fig. 3. Direct Western blot of parental HTB9, their microcell hybrid clones,and hybrid tumor derived cells. Lane I. normal control WI-38 human fibroblastcells; Lane 2, HTB9 parental cells; Lane 3. human-mouse hybrid 204-4 donorcells; Lanes 4 and 5, respectively, early and late passage HTB9-CI cells: Lane 6.HTB9 CMS;iMne 7, HTB9 CI6;'Lanes 8 and 9. respectively. 2 HTB9 RB positive
clones previously obtained from RB cDNA transfection (17), namely, H/RB-C12and H/RB-CÕ5:Lane IO, HTB9 Cl-1 TI: Lane 11. HTB9 CI-6 Tl.
tumor each, which appeared after a long latency period. All RBpositive clones were injected at passage 7 after the initial microcell fusion. The 2 tumors that developed, HTB9 CI-1 Tl andHTB9 CI-6 Tl, were removed at 7 and 6 months after injection,respectively, and established in culture. Neither tumor wasfound to have any RB positive cells by single cell staining (data
not shown) or Western blot analysis (Fig. 3, Lanes 10 and //)when the mass cultures of tumor cells were examined at thesecond or third passage after being placed into cell culture.
HTB9 CI-1 cells obtained at passage 17, which retained theirability to grow in G418 as well as containing the neomycinselection marker (data not shown), had lost RB protein expression as determined by single cell staining (data not shown) andWestern blot analysis (Fig. 3, Lane 5). When the metaphasesfrom these RB negative HTB9 CI-1 cells were examined, thenormal chromosome 13 was still present (data not shown). Thisindicated that only a small amount of genetic information, perhaps only a portion of the normal RB alÃele,had been lost fromthe transferred chromosome 13. Of particular interest was thefact that these RB negative HTB9 CI-1 revenant cells grewequally well in cell culture (Fig. 4, A and B) and were as tum-origenic as the parental HTB9 cells (Table 2). These resultsstrongly suggest that it was the RB alÃeleon chromosome 13that was responsible for the phenotypic changes found in theRB positive HTB9 cells rather than some other unknown geneon chromosome 13.
Prostate Cancer. When the DU 145 prostate cells were usedas the recipients, 2 independent RB positive clones (DU 145CI-1 and DU 145 CI-2) were obtained (Fig. 5, Lanes 3 and 4,respectively). DU 145 contains only a truncated RB protein
30
Ü
O
-O— HTB-9
--*--- HTB-9CI-1RB
-O— HTB-9 CI-5
-a— HTB-9 CI-6
•¿�-D---HTB-9CI-1RB
30-1HTB-9
HTB-9CI-1RB
HTB-9CI-5
HTB-9CI-6
HTB-9CI-1RB
O
O
234
DAY DAY
Fig. 4. Cell growth of HTB9, DUI45, and their chromosome 13 transferred hybrid clones in 10 and 3% FBS.
6300
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
Table 2 Tumorigenicity and in vitro growth of RB+ and RB- HTB9and DUI45 cells 1 2345678
CelltypeHTB9HTB9CI-1,
RB+HTB9Cl-l.RB-HTB9Cl-5,RB+HTB9Cl-6,RB+DU
145D8/gLCI-9DU
145Cl-lDU145CI-2Plating
EfficiencyInsoftAgar6.80.0ND<0.00.07.8ND7.57.0No.ofcellsinjectedIxlO7IxlO7IxlO7IxlO7IxlO7IxlO7IxlO7IxlO7IxlO7Mice
withTumors/miceinjected6/61/186/60/121/186/65/54/120/12Av.
size(mm')314"75*264"ISO''625'684'76
/
" Tumors measured at 1 month after injection.* Tumor measured at 7 months after injection.c ND, not determined.d Tumor measured at 6 months after injection.' Tumors measured at 2 months after injection.•¿�^Threetumors measured at 2 months (76 mm3). 1 tumor measured at 6 months
was 150 mm3.
(Fig. 5, Lane 2) and also has no RB nuclear staining, whereasboth DU 145 CI-1 and CI-2 also showed strong RB nuclearstaining (data not shown). The lack of RB nuclear protein staining seen in the DU 145 cells likely reflects the fact that itstruncated RB protein does not enter the nucleus to any measurable extent. In addition, since parental DU 145 cells do nothave any normal chromosome 13, whereas both RB positive celllines contained one normal 13 chromosome, a complete chromosome 13 had been transferred in each clone (data notshown).
Unlike the HTB9 bladder carcinoma cell lines, however, neither RB positive DU 145 clones had any significant growthchanges in cell culture compared to the parental cells whethergrown in either 10 or 3% fetal bovine serum (Fig. 4, C and D,respectively). In addition, the RB positive DU 145 clonesformed colonies in soft agar with efficiency equal to that of theparental DU 145 cells (Table 2).
Clone DU 145 CI-2 was nontumorigenic, whereas the parental DU 145 produced large tumors rapidly in 6 of 6 of the nu/numice given injections (Table 2). However, 4 small tumors werefound following s.c. injection of the DU 145 CI-1 cell clone(Table 2). Three of these tumors were removed at 2 months andthe other tumor was removed 6 months after injection. None ofthese tumors grew progressively. Each tumor was established incell culture. All 4 cell lines obtained produced strong nuclearstaining by single cell analysis (data not shown) and showed asimilar normal RB protein pattern as found in the originalDU 145 CI-1 cell line (Fig. 5, Lanes 5 to 8). Thus, although
tumorigenicity was markedly suppressed, the small tumors thatformed remained RB positive.
DU 145 cells that had been transfected with a neomycin resistant plasmid and subsequently cloned in medium containing200 Mgof G418 (D8/gL) were used as a neomycin control forthe tumorigenicity studies. This was necessary since a RB negative revertant DU 145 clone could not be obtained from the RBpositive clones as was possible for the HTB9 CI-1 cells. Theinability to obtain such revenants was likely the result of thefact that no growth advantage existed for any potential RBnegative cells that might have been present in the DU 145 CI-1or CI-2 cultures. The neomycin resistant cells (D8/gL) were astumorigenic as the parental DU 145 cells (Table 2).
Fig. 5. Direct Western blot of parental DU145. their microcell hybrid clones,and hybrid tumor derived cells. Lane 1, normal control WI-38 human fibroblastcells; Lane 2, DU145 showing truncated pi 10RB present; Lane 3, DU145 CM;Lane 4, DU 145 CI-2; Lanes 5-8, respectively, tumors derived from DU 145 CI-2cells TI, T2, TV and T4.
DISCUSSION
Our results in general suggest that there is an even moremarked inhibition of cell growth and tumorigenicity followingthe transfer of a chromosome 13 containing a normal RB alÃelethan the introduction of RB cDNA by retroviral infection orplasmid transfection. This is summarized in Table 3. For example, only once were we able to obtain any RB positive retin-oblastoma cells by microcell fusion, and these RB positive cellscould only be passaged for a limited time. In contrast, RBpositive long-term retinoblastoma cultures have been obtainedby retroviral infection (16, 18, 20), although different tumori-genie potentials for these RB positive clones have been reportedwhen injected intraocularly (Table 3). Such differences maysuggest the advantage of a more "natural" regulation of the RB
gene, since presumably the normal RB promotor region andother regulatory elements are transferred by microcell fusion,whereas artificial promoters have been utilized for both theinfection and transfection studies. In addition, other 5' regula
tory sequences likely also have been eliminated from the variousRB cDNA constructs. In other chromosome transfer experiments, physiological levels of gene expression at the mRNAlevel have been observed (31 ).
We reported that RB positive WERI-27 cells obtained byretroviral infection could produce RB positive tumors wheninjected intraocularly, although with a longer latency periodand decreased frequency than the parental RB negative cells(16). Using the same construct and parental cell type, however,others have found that their RB positive WERI-27 cells werenontumorigenic (18). It had been stated that our first study didnot use RB positive clonally derived cells for intraocular injections but rather that mass cultures of RB positive cells wereused. It should be noted that their original report also used masscultures rather than clonal derivatives for tumorigenicity testing(14). In addition it was stated that RB protein expression wasnot analyzed immediately after tumor removal in our studies(16). However, RB expression was indeed examined both at thesingle cell level and by direct Western blotting in first passagecells after their removal from the eye. In addition, several independent RB positive WERI-27 clones have been subsequently isolated in our laboratory that have produced RB positive intraocular tumors as documented by both single cellstaining and Western blot analysis of the primary tumors.4 The
basis for these differences is yet unknown.
4Y. Zhou, K. Xu, S-X. Hu, W. F. Benedict, and H-J. Xu. unpublishedobservations.
6301
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
Table 3 Comparison of results ofmicrocell transfer to RB cDNA infection or transfection studies
Chromosomemicrocell transfer (this study)
RB cDNA transfer by retroviralinfection or plasmid transfection
Tumor type Cell culture Tumorigenicity Cell culture Tumorigenicity
Retinoblastoma 1. Large, slowly growingRB positive cells,unable to grow inlong-term culture
1. RB negative intraoculartumors produced only
Osteosarcoma
Bladder cancer
Prostate cancer
1. Large, slowly growingcells which could notbe subculturedand too few to analyzeRB protein expression
1. No morphologicalchanges; decreased growthin both 10% and 3% FBS;no growth in soft agar
1. No change includingmorphology, growthrate, or growth in soft agar
I. Insufficient number of cellsto test for tumorigenicity
1. Marked tumor suppression;only 2 small RB negativetumors produced in 48mice injected
Marked tumor suppression;4 small tumors from 1 clonewere all RB positive
1. Large, slowly growing RBpositive cells; initially long-termcultures not obtained (14, IS)
2. Long-term RB positive culturesobtained of normal size whichgrew well in culture (16)
3. RB positive long-term cellsproduced with no change inmorphology (19, 20)
4. RB positive cells producedwith no decrease in growthrate or morphologicalchanges(18)
1. Large, slowly growing RBpositive cells (14)
2. Large, rapidly senescentcells (21)
No morphological changes;normal growth in 10% FBS butdecreased growth in 3% FBS;no growth in soft agar ( 17)No morphological changes;normal growth in 10% FBS(22)
1. No change of morphologyor growth rate (11)
1. No tumors produceds.c. (14, 15)"
2. Growth of RB positivetumors intraocularly althoughtumor suppression occurred(16); no s.c. tumorsformed (16)
3. No RB positive s.c.tumors formed (19);marked tumor suppressionand decrease in RBpositive cells in fewintraocular tumors seen(20)
4. RB positive tumorsformed intraocularly asfrequently as RBnegative tumors
1. Insufficient number ofcells to test fortumorigenicity (14, 21)
1. Two largest tumorsproduced were RB negative;6 other smallertumors examined wereRB positive (17)
2. 21 tumors formed;the 3 tumors analyzed wereRB negative (22)
1. Decreased tumorigenicity;all tumors analyzed wereRB negative (11)
" Numbers in parentheses, references.
A third study using a different RB cDNA construct thatincluded an inducible mouse metallothionein promoter foundno decrease in tumorigenicity in the RB positive retinoblastomaclones isolated when these clones were compared to the tumorsformed intraocularly by the RB negative parental cells (20).However, it is likely that only low levels of RB expression couldhave been produced under the noninducible in vivo conditionsin which these studies were performed (20). Therefore, the promoter chosen for such experiments and the level of RB expression achieved appear to be critical to the degree of growthinhibition and tumor suppression that is produced using theseconstructs.
In the case of osteosarcoma, introduction of the RB gene byretroviral-mediated gene transfer of Saos-2 cells resulted in cellcultures having normal nuclear RB staining (14). The majorityof these cells were larger than the parental cells and grew veryslowly (14). No long-term cultures of Saos-2 cells with normalRB protein expression were isolated and, therefore, tumorigenicity studies could not be undertaken. Others have found similar results following transfection of a different RB cDNA construct containing an SV40 early promoter (21). In fact, therapid appearance of senescent cells resulting from introductionof a wild-type RB into Saos-2 cells was used as an assay toevaluate the properties of mutant forms of RB (21). Theseresults are similar to those found by us after microcell fusion ofa chromosome 13 into Saos-2 cells. In this case, the large cellsobtained appeared to grow even slower and could not be passedin culture in order to determine their RB protein expression.
When a RB cDNA using the ß-actingene promoter was
transfected into the bladder carcinoma cell line, HTB-9, nodecrease in growth rate was seen in the resultant 2 RB positiveclones obtained when they were cultured in 10% FBS, althoughdecrease in growth was observed when the concentration ofFBS was reduced to 3% ( 17). In addition, the RB positive clonesdid not grow in soft agar. They also in general produced tumorsthat were markedly smaller and that appeared to have a longerlatency period than the parental RB negative cells (17). The 2large tumors that occurred rapidly were RB negative, whereaseach of the 6 small tumors remained RB positive (17). In contrast, the 3 RB positive clones obtained by microcell fusionshowed a decreased growth rate even when cultured in 10%FBS, as well as not forming colonies in soft agar. Furthermore,only 2 tumors were produced in 48 animals given injectionsusing these 3 RB positive clones, and both tumors were foundto be RB negative. Therefore, again the RB positive clonesisolated after microcell fusion had a significantly decreasedgrowth rate in culture and did not form RB positive tumors,compared to RB positive clones obtained after transfection ofan RB cDNA plasmid into the same parental cells (17). The factthat we were able to obtain an RB negative revenant from theHTB9 CM cells that had retained the entire transferred 13chromosome, including the neomycin marker, and had the parental HTB9 phenotype also strongly suggests that our resultsare directly related to the transfer of a normal RB alÃeleratherthan some additional genes on chromosome 13.
Recently, Goodrich et al. (22) also examined phenotypicchanges following introduction of the RB gene by retrovirusmediated gene transfer into 3 bladder tumor cell lines that had
6302
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
lost normal RB function, namely, 5637 (HTB9). HT 1376, andJ-82. Since J-82 itself was not tumorigenic, no conclusions canbe drawn comparing growth rate changes and tumorigenic potential using this cell line. No significant changes were seen ingrowth rate when various RB positive 5637 and HT 1376 cloneswere analyzed, although unfortunately no studies on the growthof these clones in soft agar were reported (22), so that theresults could not be directly compared to previous experimentsin which the RB gene was transferred by transfection into one ofthe same cell lines, namely 5637 (HTB9) (17). In addition,although tumorigenicity was in general suppressed followinginjection of the RB positive clones, 6 of 7 of the RB positive1376 clones isolated produced tumors (16 total tumors formed)and 4 of 4 of the 5637 RB positive clones produced at least onetumor (5 total tumors). These investigators state that the tumors were RB negative (22). However, results from only two1376 tumors and one 5637 tumor are given (22). Since in aprevious study using RB positive 5637 clones, it was found thatthe 2 largest tumors formed were RB negative, whereas theother 6 smaller tumors were RB positive (17), it would havebeen helpful for comparison if all tumors produced in theGoodrich et al. (22) study had been examined for RB proteinexpression and reported in their paper. Nevertheless, tumorigenicity was decreased in general in these RB positive clonescompared to their RB negative parental counterparts (22).
Finally, either RB cDNA retroviral infection (11) or transferby microcell fusion of a normal 13 chromosome into the prostate cancer cells, DU 145, yielded clones with a normal proteinpattern, all of which showed no phenotypic differences in culture compared to the parental DU 145 cells. Numerous tumorswere produced following s.c. injection of the retroviral transferred RB positive clones, although they were all found to be RBnegative (11), whereas considerably fewer tumors developedafter injection of the 2 RB positive clones obtained from microcell fusion (0 of 12 and 4 of 12). In contrast, these latter 4small tumors remained RB positive, indicating that tumorigenicity of the RB positive cells was not completely suppressed,although tumorigenicity was markedly decreased. Since therewas no decrease in growth rate of our 2 RB positive prostatecancer clones in culture, although tumor formation was greatlysuppressed, it appears that we were able to separate the inhibition of cellular growth produced by transfer of normal RB geneexpression into this particular tumor type from its tumor sup-pressive function.
There also now appears to be a trend in the phenotypicchanges observed following RB transfer that correlates with thepresumed role of the RB gene in each of the tumor types analyzed (Table 3). Cells from those tumors in which the loss of RBfunction is thought to be directly involved as a causative factorin tumor development, namely retinoblastoma and osteosar-coma, show considerably more growth inhibition and morphological changes initially after transfer of a normal RB gene thansimilar transfer in bladder or prostate cells, both of which aretumors where the RB gene likely has a role in tumor progression rather than initiation (12, 24-26). In fact, no long-termcultures of RB positive osteosarcoma cells have yet been reported that were obtained following introduction of a normalRB construct into osteosarcoma cells in which RB function hadinitially been lost.
These differences may also reflect the total number of mutations that are required for retinoblastoma or osteosarcoma development (tumors that occur in childhood and adolescence or
early adult life, respectively) compared to bladder and prostatecancer (tumors that occur in later adult life). It is of particularinterest, however, even in those adult tumor types that havenumerous other genetic changes associated with their tumorigenicity and where functional RB loss likely reflects tumorprogression rather than causation, reexpression of normal RBprotein production results in marked, although not always complete, tumor suppression. Such findings may offer some hopethat in the future safe and efficient vectors can be produced thatwill enable normal RB gene transduction to occur in vivo, providing a new approach to therapy for some of the tumor typesin which loss of RB function has a major role.
REFERENCES
1.
to.
11.
12.
13.
14.
15.
17.
18.
19.
20.
Friend, S. H.. Bernards. R., Rogeli. S., Weinberg. R. A.. Rapaport. J. M.,Albert, D. M.. and Dryja, T. B. A human DNA segment with properties of thegene that predisposes to retinoblastoma and osteosarcoma. Nature (Lond.),323: 643-646, 1986.Fung. Y-K. T., Murphree, A. L., T'Ang. A., Qian, J.. Hinrichs, S. H., and
Benedict, W. F. Structural evidence for the authenticity of the human retinoblastoma gene. Science (Washington DC), 236: 1657-1661. 1987.Lee, W-H., Bookstein, R., Hong, F.. Young. L-J., Shew, J-Y., and Lee, Y. H.P. Human retinoblastoma susceptibility gene: cloning, identification and sequence. Science (Washington DC), 236: 1394-1399. 1987.Friend. S. H., Horowitz, J. M., Gerber, M. R., Wang. X. F., Bogenmann. E.,Li, F. P.. and Weinberg, R. A. Deletions of a DNA sequence in rctinoblas-tomas and mesenchymal tumors: organization of the sequence and its encoded protein. Proc. Nati. Acad. Sci. USA, 84: 9059-9063, 1987.Weichselbaum. R., Beckett, M., and Diamond, A. Some retinoblastomas,osteosarcomas and soft tissue sarcomas may share a common etiology. Proc.Nati. Acad. Sci. USA, 85: 2106-2109, 1988.Reissman, P. T., Simon. M. A., Lee, W-H., and Slamon. D. J. Studies of theretinoblastoma gene in human sarcomas. Oncogene. 4: 839-843. 1989.Toguchida, J., Ishizaki, K., Sasaki, M. S.. Ikenaga. M., Sugimoto, M., Kot-oura, Y., and Yamamuro, T. Chromosomal reorganization for the expressionof recessive mutation of retinoblastoma susceptibility gene in the development of osteosarcoma. Cancer Res.. 48: 3939-3943, 1988.Horowitz, J. M., Yandell, D. W., Park, S-H., Canning. S., Shyte, P., Buch-kovich. K. J., Harlow, E., Weinberg, R. A., and Dryja, T. P. Point mutationalinactivation of the retinoblastoma antioncogene. Science (Washington DC),2^:937-940, 1989.Presti, J. C., Reuter, V. E.. Galan, T., Fair, W. R., and Cordon-Cardo, C.Molecular genetic alterations in superficial and locally advanced human bladder cancer. Cancer Res., 51: 5405-5409, 1991.Ishikawa, J.. Xu, H-J., Hu, S-X., Yandell, D. W., Maeda, S., Kamidono, S.,Benedict. W. F.. and Takahashi, R. Inactivation of the retinoblastoma gene inhuman bladder and renal cell carcinomas. Cancer Res., 51:5736-5743, 1991.Bookstein, R., Shew, J-Y., Chen, P-L.. Scully, P., and Lee, W-H. Suppression of tumorigenicity of human prostate carcinoma cells by replacing amutated RB gene. Science (Washington DC), 247: 712-715. 1990.Bookstein, R., Rio. P., Madreperla, S. A.. Hong, F., Allred, C, Grizzle, W.E., and Lee, W-H. Promoter deletion and loss of retinoblastoma gene expression in human prostate carcinoma. Proc. Nati. Acad. Sci. USA, 87:7762-7766, 1990.Murphree, A. L., and Benedict, W. F. Retinoblastoma: clues for humanoncogenesis. Science (Washington DC), 223: 1028-1033, 1984.Huang, H-J. S., Yee, J-K., Shew, J-Y.. Chen, P-L.. Bookstein, R.. Fried-mann. T.. Lee, E. Y-H. P., and Lee, W-H. Suppression of the neoplasticphenotype by replacement of the RB gene in human cancer cells. Science(Washington DC), 242: 1563-1566. 1988.Sumegi, J., Uzvolgyi, E., and Klein, G. Expression of the RB-gene under thecontrol of MuLV-LTR suppresses tumorigenicity of WERI-RB-27 retinoblastoma cells in immunodefective mice. Cell Growth & Differ., /: 247-250,1990.Xu. H-J., Sumegi, J., Hu, S-X., Bancrjcc. A., Uzvolgyi, E., Klein, G., andBenedict, W. F. Intraocular tumor formation of RB reconstituted retinoblastoma cells. Cancer Res.. 51: 4481-4485. 1991.Takahashi, R., Hashimoto. T.. Xu. H-J., Hu. S-X., Matsui, T., Miki, T.,Bigo-Marshall, H., Aaronson, S. A., and Benedict, W. F. The retinoblastomagene functions as a growth and tumor suppressor in human bladder carcinoma cells. Proc. Nati. Acad. Sci. USA, 88: 5257-5261, 1991.Madreperla, S. A., Whittum-Hudson, J. A., Prendergast, R. A., Chen, P-L.,and Lee, W-H. Intraocular tumor suppression of retinoblastoma gene-reconstituted retinoblastoma cells. Cancer Res., 51: 6381-6384, 1991.Chen. P-L.. Chen, Y.. Shan, B., Bookstein. R., and Lee. W-H. Stability ofretinoblastoma gene expression determines the tumorigenicity of reconstituted retinoblastoma cells. Cell Growth & Differ.. 3: 119-125, 1992.Muncaster. M. M.. Cohen, B. L., Phillips, R. A., and Gallic, B. L. Failure ofRBI to reverse the malignant phenotype of human tumor cell lines. Cancer
6303
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from

RB REEXPRESSION IN VARIOUS HUMAN CANCER TYPES
Res., 52:654-661, 1992.21. Templcton, D. J., Park, S. H., Lanier, L., and Weinberg, R. A. Nonfunctional
mutants of the retinoblastoma protein are characterized by defects in phos-phorylation, viral oncoprotein association, and nuclear tethering. Proc. Nati.Acad. Sci. USA, 88: 3033-3037, 1991.
22. Goodrich, D. W., Chen, Y., Scully, P., and Lee. W-H. Expression of theretinoblastoma gene product in bladder carcinoma cells associated with a lowfrequency of tumor formation. Cancer Res., 52: 1968-1973, 1992.
23. Stanbridge, E. J. The Genetic Basis of Tumor Suppression. CIBA Symposium 142, pp. 149-159. New York: John Wiley and Sons, 1989.
24. Saxon, P. J., Srivatsan, E. S., Leipzing, G. V., Sameshima, J. H.. and Stan-bridge. E. J. Selective transfer of individual human chromosomes to recipientcells. Mol. Cell Biol., 5: 140-146, 1985.
25. Xu, H-J., Hu. S-X., Hashimoto. T.. Takahashi, R., and Benedict, W. F. Theretinoblastoma susceptibility gene product: a characteristic pattern in normalcells and abnormal expression in malignant cells. Oncogene, 4: 807-812,1989.
26. Benedict, W. F., Weissman, B. E., Mark, C., and Stanbridge, E. Tumorige-nicity of human HT 1080 fibrosarcoma x normal fibroblast hybrids: chromo
some dosage dependency. Cancer Res., 44: 3471-3479, 1984.27. R. Hay, M. Macy, T. R. Chen, P. McClintock, and Y. Leid (eds.). American
Type Culture Collection Catalogue of Hybrid Lines and Hybridomas, Ed. 6,p. 196. Rockville, MD: American Type Culture Collection, 1988.
28. Hartley-Asp, B., Billstrom, A., and Kruse, E. Identification by c-banding oftwo human prostate tumour cell lines, 1013L and DU145. Int. J. Cancer, 44:161-164, 1989.
29. Benedict, W. F., Dawson, J. A., Banerjee, A., and Murphree, A. L. The nudemouse model for human retinoblastoma: a system for evaluation of retinoblastoma therapy. Med. Pediatr. Oncol., 8: 391-395, 1980.
30. Benedict, W. F., Lingua, R. W., Dolron. D. R., Dawson, J. A., and Murphree,A. L. Tumor regression of human retinoblastoma in the nude mouse following photoradiation therapy: a preliminary report. Med. Pediatr. Oncol., A'.-397-401, 1980.
31. Goyette, M. C., Cho, K., Lasching, C. L., Levy, D. B., Kinzler, K. W.,Paraskeva, C., Vogelstein, B., and Stanbridge, E. J. Progression of colorectalcancer is associated with multiple tumor suppressor gene defects but inhibition of tumorigenicity is accomplished by correction of any single defect viachromosome transfer. Mol. Cell. Biol., 12: 1387-1395, 1992.
6304
Research. on January 16, 2014. © 1992 American Association for Cancercancerres.aacrjournals.org Downloaded from
Related Documents











