1167 ISSN 0031-0301, Paleontological Journal, 2008, Vol. 42, No. 11, pp. 1167–1284. © Pleiades Publishing, Ltd., 2008. CONTENTS INTRODUCTION 1168 CHAPTER 1. FUNCTIONAL INTERPRETATION OF THE BODY PLAN IN TAXA OF THE ORDER RANK 1169 1.1. State of the Modern System of Cephalopods 1169 1.2. Functional Interpretation of Morphological Characters of the Cephalopod Shell 1169 1.3. Improving and Regulating of Buoyancy 1170 1.4. Orientation and Support of Orientated Position 1172 1.5. Body Plans and Major Evolutionary Trends in Cephalopod Orders 1172 CHAPTER 2. LIFE-FORMS OF CEPHALOPODS 1186 2.1. Concept of Life-Forms 1186 2.2. Life-Forms of Cephalopods 1188 2.3. Ecological (Adaptive) Significance of Constructive Differences of the Outer Shell in Fossil Cephalopods, Criteria and Methods of Their Assignment to Various Life-Forms 1189 2.4. Life-Forms of Cephalopods with a Curved Shell 1191 2.5. Life-Forms of Cephalopods with a Straight Shell 1192 2.6. Life-Forms of Cephalopods with a Planispiral Shell 1194 2.7. Life-Forms of Cephalopods with Planispiral Shell in which the Whorls Were Not in Contact 1209 2.8. Life-Forms of Cephalopods with a Conispiral Shell 1210 2.9. Life-Forms of Cephalopods with a Heteromorphic Shell 1211 CHAPTER 3. ECOLOGICAL SPECIALIZATION AND ECOGENESIS OF PALEOZOIC CEPHALOPODS 1214 3.1. Ecological Structure of the Modern Cephalopod Taxocoenosis 1214 3.2. Ecological Structure of Paleozoic Cephalopods 1215 3.2.1. Order Ellesmerocerida 1215 3.2.2. Order Endocerida 1216 3.2.3. Order Actinocerida 1216 3.2.4. Order Orthocerida 1218 3.2.5. Order Pseudorthocerida 1218 3.2.6. Order Tarphycerida 1220 3.2.7. Order Lituitida 1220 3.2.8. Order Barrandeocerida 1220 3.2.9. Order Discosorida 1222 3.2.10. Order Oncocerida 1222 3.2.11. Order Ascocerida 1223 3.2.12. Order Nautilida 1223 3.2.13. Order Anarcestida 1224 3.2.14. Order Tornoceratida 1225 3.2.15. Order Clymeniida 1225 3.2.16. Order Praeglyphioceratida 1226 3.2.17. Order Goniatitida 1226 3.2.18. Order Prolecanitida 1226 3.2.19. Order Ceratitida 1228 3.3. Morphological Diversity of Life-forms and Ecogenesis of Cephalopod Taxocoenosis in the Paleozoic 1228 CHAPTER 4. ECOLOGICAL STRUCTURE OF PALEOZOIC AMMONOID COMMUNITIES IN THE URALIAN PALEOBASIN 1240 4.1. General Background 1240 4.2. Ecological Structure of the Paleozoic Ammonoid Communities in the Urals 1242 4.3.1. Early Devonian 1243 4.3.2. Middle Devonian 1245 4.3.3. Late Devonian 1245 4.3.4. Mississippian (Early Carboniferous) 1253 4.3.5. Pennsylvanian (Middle and Late Carboniferous) 1262 4.3.6. Early Permian 1268 CONCLUSIONS 1275 REFERENCES 1277 Cephalopods in the Marine Ecosystems of the Paleozoic I. S. Barskov, M. S. Boiko, V. A. Konovalova, T. B. Leonova, and S. V. Nikolaeva Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya ul. 123, Moscow, 117997 Russia e-mail: [email protected], [email protected] Received February 17, 2008 DOI: 10.1134/S0031030108110014

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1167
ISSN 0031-0301, Paleontological Journal, 2008, Vol. 42, No. 11, pp. 1167–1284. © Pleiades Publishing, Ltd., 2008.
CONTENTS
INTRODUCTION 1168CHAPTER 1. FUNCTIONAL INTERPRETATION OF THE BODY PLAN IN TAXA OF THE ORDER RANK 1169
1.1. State of the Modern System of Cephalopods 11691.2. Functional Interpretation of Morphological Characters of the Cephalopod Shell 11691.3. Improving and Regulating of Buoyancy 11701.4. Orientation and Support of Orientated Position 11721.5. Body Plans and Major Evolutionary Trends in Cephalopod Orders 1172
CHAPTER 2. LIFE-FORMS OF CEPHALOPODS 11862.1. Concept of Life-Forms 11862.2. Life-Forms of Cephalopods 11882.3. Ecological (Adaptive) Significance of Constructive Differences of the Outer Shell
in Fossil Cephalopods, Criteria and Methods of Their Assignment to Various Life-Forms 11892.4. Life-Forms of Cephalopods with a Curved Shell 11912.5. Life-Forms of Cephalopods with a Straight Shell 11922.6. Life-Forms of Cephalopods with a Planispiral Shell 11942.7. Life-Forms of Cephalopods with Planispiral Shell in which the Whorls Were Not in Contact 12092.8. Life-Forms of Cephalopods with a Conispiral Shell 12102.9. Life-Forms of Cephalopods with a Heteromorphic Shell 1211
CHAPTER 3. ECOLOGICAL SPECIALIZATION AND ECOGENESIS OF PALEOZOIC CEPHALOPODS 1214
3.1. Ecological Structure of the Modern Cephalopod Taxocoenosis 12143.2. Ecological Structure of Paleozoic Cephalopods 1215
3.2.1. Order Ellesmerocerida 12153.2.2. Order Endocerida 12163.2.3. Order Actinocerida 12163.2.4. Order Orthocerida 12183.2.5. Order Pseudorthocerida 12183.2.6. Order Tarphycerida 12203.2.7. Order Lituitida 12203.2.8. Order Barrandeocerida 12203.2.9. Order Discosorida 12223.2.10. Order Oncocerida 12223.2.11. Order Ascocerida 12233.2.12. Order Nautilida 12233.2.13. Order Anarcestida 12243.2.14. Order Tornoceratida 12253.2.15. Order Clymeniida 12253.2.16. Order Praeglyphioceratida 12263.2.17. Order Goniatitida 12263.2.18. Order Prolecanitida 12263.2.19. Order Ceratitida 1228
3.3. Morphological Diversity of Life-forms and Ecogenesis of Cephalopod Taxocoenosis in the Paleozoic 1228CHAPTER 4. ECOLOGICAL STRUCTURE OF PALEOZOIC AMMONOID COMMUNITIES IN THE URALIAN PALEOBASIN 1240
4.1. General Background 12404.2. Ecological Structure of the Paleozoic Ammonoid Communities in the Urals 1242
4.3.1. Early Devonian 12434.3.2. Middle Devonian 12454.3.3. Late Devonian 12454.3.4. Mississippian (Early Carboniferous) 12534.3.5. Pennsylvanian (Middle and Late Carboniferous) 12624.3.6. Early Permian 1268
CONCLUSIONS 1275REFERENCES 1277
Cephalopods in the Marine Ecosystems of the Paleozoic
I. S. Barskov, M. S. Boiko, V. A. Konovalova, T. B. Leonova, and S. V. Nikolaeva
Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya ul. 123, Moscow, 117997 Russiae-mail: [email protected], [email protected]
Received February 17, 2008
DOI:
10.1134/S0031030108110014

1168
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
INTRODUCTION
Cephalopods are the largest and most diverse groupof the Paleozoic marine biota. By now about one and ahalf thousand valid genera and over four thousand spe-cies have been described. In the last century hundredsof papers have been written on Paleozoic cephalopods,including fundamental studies on all major groups andgeochronological intervals. The historical developmentof higher taxa, their morphological and taxonomicdiversity, have been relatively fully studied, their diver-sity dynamics have been tracked, and their biostrati-graphic significance, which was the basis for the stratig-raphy of the Late Paleozoic, has been demonstrated. Theontogenetic studies of Paleozoic ammonoids became thebasis of the onto-phylogenetic method in paleontologyand allowed the recognition of the major evolutionarypatterns, which became important across many fields ofbiology.
All this knowledge was to a large extent summa-rized half a century ago in
Osnovy paleontologii
(
Fun-damentals of Paleontology,
Ed. by V.E. Ruzhencev,1962) and
Treatise on Invertebrate Paleontology. Part K,Part L
, Ed. by R.C. Moore (1957, 1964). In recentdecades new data were accumulated in all the abovefields, and some new methods and approaches in Pale-ozoic cephalopod studies were proposed and success-fully tried (this includes microstructural studies andvarious sorts of mathematical methods).
All these numerous and diverse studies show that inthe Paleozoic cephalopods represented one of the majorelements of the ecological structure in marine ecosys-tems and played a significant role in the evolution of theentire biosphere.
At the same time, our knowledge of the ecology andlifestyle of fossil cephalopods are far from sufficient,and this field is the least explored in cephalopod stud-ies. Until now both professional and semipopular liter-ature widely promotes an image of all cephalopods asactive pelagic predators. This belief is based on a quiteunjustified transferal of the lifestyle and behavior of thebest known extant cephalopods to the whole of thismorphologically and taxonomically diverse group,including numerous fossil cephalopods; even amongthe modern cephalopods, active pelagic predators con-stitute less than half of the taxocoenosis.
The lifestyle of fossil cephalopods with an outershell have only been seriously studied in about ten or sopapers. These papers are generally those that addressgenera and species with unusual morphology (seeShevyrev, 2005). In
Treatise
(1964) only the data on themodern
Nautilus
are cited. In
Osnovy paleontologii
there is a small section with general views on the life-style of some cephalopods (Shimansky, 1962). Very
few papers discuss the general problems of adaptivespecialization of fossil cephalopods, ecogenesis ofgroups, which would be important for understandingtheir role in past ecosystems.
Louis Dollo (1922) was the first to attempt to pro-pose a general scheme of adaptive specialization of fos-sil cephalopods. Barskov (1976, 1988, 1989, etc.) sub-stantiated the recognition of life-forms based on shellgeometry and hydrostatics.
The main task facing the present authors was toapply functional and ecological approaches to the studyof the entire community of Paleozoic cephalopods. Thefirst chapter discusses the general state of the macrosys-tem of Paleozoic cephalopods. The considerable vari-ety of proposed systematic schemes and the lack ofunanimity in understanding of the rank and composi-tion of the higher taxa of cephalopods probably resultfrom the morphological approach being used in isola-tion. We attempt to substantiate the differences in thebody plans and ranks of the major groups of Paleozoiccephalopods based on the functional morphologicalmethod. Fundamental differences in the body plansallowing the recognition of orders are related to differ-ent mechanisms of manipulating buoyancy and orienta-tion within the framework of the archetype of this class.
In the second chapter, the functional morphologicalapproach is used to substantiate adaptive types, i.e.,life-forms of cephalopods with different morphology.They are based on the evaluation of cephalopod poten-tial to inhabit different adaptive zones of the sea, whichis largely dependent on shell architecture and geometry,including mechanisms for maintaining orientation, andother features important for an animal with a shell oper-ating as a buoyancy device.
In the third chapter, the life-forms are recognized ineach of 23 cephalopod orders based on the criteria pro-posed in the second chapter, and the ecogenesis of theseorders is discussed throughout their evolutionary his-tory. The ecological structure of the entire cephalopodtaxocoenosis is discussed and its changes and thechanges in the taxonomic composition if each life-formthroughout the Paleozoic in relation to changes in abi-otic factors are considered.
The fourth chapter discusses in detail the evolutionof the ecological structure of ammonoid communitiesin the Uralian Paleobasin from the Early Devonian tothe Early Permian inclusive. Changes in the ecologicalstructure and taxonomic composition are discussed inthe context of their potential connection with the geo-logical history of the basin.
The authors dedicate this paperto the memory of A.A. Shevyrev

PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1169
CHAPTER 1. FUNCTIONAL INTERPRETATION OF THE BODY PLAN IN TAXA
OF THE ORDER RANK
1.1. State of the Modern System of Cephalopods
Although a sensible system of cephalopods at themegataxon level has been intensively sought since themid-20th century, no agreement has been achieved onthe number of orders and subclasses, i.e., on the bodyplans within the archetype of the class (Flower andKümmel, 1950; Flower, 1964; Shimansky and Zhurav-leva, 1961, Donovan, 1964; Teichert, 1967, 1988;Zeiss, 1969; Zhuravleva, 1972; Salvini-Plawen, 1980;House, 1981; Drushchits and Shimansky, 1982; Staro-bogatov, 1983; Leonova, 2002; Shevyrev, 2005, 2006a,2006b; etc.). Approaches to classification have beenvery varied, which is reflected in the subdivision of theclass into between two and eight subclasses. There ismore consistency in the understanding of the orders,although the number of recognized orders varies from15 to 30. The latest review of the macrosystem wasundertaken by Shevyrev (2005, 2006a, 2006b). Unfor-tunately, this system, like those proposed earlier, isnothing more than another shuffling of orders, some ofwhich is widely accepted, and some is only accepted bya few authors. However, even the widely acceptedorders have been placed in all subclasses in the systemsproposed by various authors. In our opinion, the mainreason for the lack of success in reconstructing the mac-rosystem of cephalopods was that when the orders wereestablished and united in the subclasses, the compara-tive taxonomic analysis was usually based on the mor-phological characters of the shell and/or its parts withan estimation of their functional or ecological roles.The disparity and taxonomic value of characters areunderstood differently by different authors. This makesan agreement on the number of orders of fossil cepha-lopods and on their combinations in yet higher taxa,virtually impossible. In our opinion, functional inter-pretation of morphological characters of the cephalo-pod shell is the criterion on which the evaluation of dis-parities can be based. In recent cephalopod taxa, thedescription of the morphological state of the soft bodyanatomy, which has experimentally testable physiolog-ical and functional significance, may characterize theirbody plan, and consequently, their affinity to differenthigher rank taxa. In fossil cephalopods (which consti-tute three-quarters of the entire taxonomic diversity ofcephalopods), a simple description of morphologicalcharacters is insufficient to form an opinion on funda-mental differences in the body plan, and cannot lead toanything but to another shuffling the possible interpre-tations. At present, to make progress in understandingthe disparity of the body plan, the evaluation of dispar-ity of morphological characters of the cephalopod shelland its parts has to be supplemented by interpretation ofthe functional significance of this disparity and of thedegree of its functional realization.
Therefore, the orders recognized in this study willbe discussed based on the explanation of fundamentaldifferences in the shell morphology from the functionalinterpretation of a particular body plan specificity in themembers of an order at the time when this orderevolved, and on the major trend in the evolution of thisspecificity (referred to as “the main cluster of develop-ment” by Ruzhencev (1960)).
1.2. Functional Interpretation of Morphological Characters of the Cephalopod Shell
Cephalopoda are the only class of mollusks whoseorigin can be traced from fossil material. Confirmedcephalopod remains are presently known only from theend of the Cambrian. The Early Cambrian genera
Volbortella
and
Salterella
, and the Middle Cambrian
Vologdinella
and
Olenecoceras
, previously assigned tocephalopods, are now excluded from this class of mol-lusks. Hence, in contrast to all other molluscan classes,whose fossil remains are known as early as the LowerCambrian, the first confirmed cephalopods are knownonly from the topmost Cambrian (genera
Plectrono-ceras
,
Paleoceras
, and
Ectenolites
). Therefore, there ismore chance of finding taxa which may be cephalopodancestors. It is currently widely accepted that cephalo-pods evolved from monoplacophorans with a relativelyhigh-coned shell. The origin of cephalopods was dis-cussed in most detail by Kobayashi (1987) and Dzik(1981). The origin of the cephalopodan archetypebegan with the appearance of the septa and siphuncle inthe apical part of the shell, the features that allowed thedevelopment of the gaseous-fluid float and colonizationof the pelagic zone, then a new adaptive zone, at thattime inaccessible for other groups of mollusks. Contin-uous septa are present in the apical parts of the shell ofmany groups of gastropods, and also in the fossilgroups Hyolitha and Tentaculita, which are sometimesplaced together in the molluscan class Coniconchia,and in the Cambrian monoplacophorans
Helcionella
and
Knightoconus
) (Yochelson et al., 1973; Kobayashi,1987). According to Kobayashi (1987),
Helcionella
,with its taller high-coned shell is a more appropriatecandidate for the role of a morphological ancestor ofcephalopods than
Knightoconus
, with its low-conedshell. Thus, the development of septa is not a featureunique to cephalopods, whereas the appearance of thesiphuncle is a fundamentally new character, as of a partof the body that remains in the chambers and is capableof controlling the buoyancy of the animal. According toDzik (1981), the development of septa happens at thelarval stage and begins with the retention of a bubblebetween the posterior mantle and the shell filled withliquid of a density less than that of the molluscan bodyor of seawater. Functionally, this is necessary to facili-tate the existence of the larva in the pelagic zone. At thenext stage, the fluid-filled sac is separated by a solidcarbonate or organic septum. Rhythmical alternation ofthese two processes led to the development of septa in

1170
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
septate monoplacophorans and tentaculites. In Dzik’sinterpretation, the siphuncle of cephalopods, responsi-ble for calibration of buoyancy, is not homologous toany structures in other mollusks and may have origi-nated from the part of the larval shell that was involvedin soft body attachment to the shell.
Kobayashi (1987) suggested that in the earliestCambrian cephalopods, septa in the apical region of theshell were imperforate, whereas the siphuncle appearedlater, at the adult stages.
Starobogatov (1974) suggested that the appearanceof the imperforate septa is the first step toward thedecollation of the apical parts of a high-conical shell.Functionally, this is related to the necessity of maintain-ing compactness and stability of the shell through thelowering of the center of gravity. Decollation is widelydeveloped in other molluscan groups. The developmentof septa and siphuncles is a result of incomplete andunfinished decollation, i.e., the septa are formed on theconvex, anatomically anterior side, and lateral sides ofthe endogastric shell, whereas the posterior half of thebody sac, which extended up to the apex, remains freeof septa. This resulted in the functional reorientation ofthe body and shell of the animal. The anterior sidebecame functionally the dorsal side, whereas the poste-rior side, where the siphuncle is located became theventral side. The previously entire mantle epitheliumsecreting the shell was subdivided into three indepen-dent sections: shell walls, septa, and posterior siphun-cular section. Later, another zone of secretion formed,that was responsible for the development of endosi-phuncular deposits. According to this interpretation, thecephalopod siphuncle is homologous to the apical partof the body sac. This interpretation is, in our opinion,more realistic than Dzik’s (1981) hypothesis of the ori-gin of the siphuncle from the larval muscle cord. Thesiphuncle of the earliest cephalopods is wide. Its diam-eter in many Ellesmerocerids, Endocerida, and Acti-nocerida is over a third and even more than half thediameter of the soft body in the body chamber. Thestructure of the soft tissue of the siphuncle with a well-developed system of blood vessels is impossible toexplain using Dzik’s hypothesis.
The gas-fluid float in the shell presented at least twomajor problems: necessity to develop and regulate thebuoyancy and orientate and stabilize the body in spacein a position comfortable for life.
1.3. Improving and Regulating of Buoyancy
Different ways of regulating buoyancy are reflectedin the shape and structure of the connecting rings of thesiphuncle. Because the epithelium of the siphuncularzone derived from the external epithelium of the man-tle, three layers homologous to the three successive lay-ers in the shell wall and septa (spherulite-prismatic,nacreous, and semi-prismatic) are secreted when thesiphuncular sheath (connecting rings) are formed. The
earliest cephalopods of the order Ellesmerocerida hadthick three-layered, apparently strongly mineralizedconnecting rings, the ends of which directly correspondto three layers of the shell walls. This structure suggestslow permeability of the siphuncular cover and, hence,low ability to regulate buoyancy. Demineralizationfacilitates the intensification of this process, which isshown by their varying degrees of thinning, mineraliza-tion, and differentiation in the longitudinal and trans-verse directions. In modern
Nautilus
, the main part of aconnective ring is a demineralized homologue of thenacreous layer, with highly porous organic membranes(Gregoire, 1968, Mutvei, 1980). Denton and Gilpin-Brown (1961) showed that the change in buoyancy in
Nautilus
occurs by osmotic pumping of the fluid fromthe chamber of the phragmocone through the special-ized epithelial cells, with cytoplasm containing highconcentrations of salts. A similar mechanism is used forchanging buoyancy in
Sepia
and
Spirula
(Denton andGilpin-Brown, 1961, 1971; Denton et al., 1961). Wardand Martin (1978) showed that for
Nautilus
, living atdepths of 200 m and deeper, the simple osmotic mech-anism is insufficient to regulate buoyancy, and the pres-ence of other mechanisms, e.g., a partial osmotic pump,is possible. Strictly speaking, the osmotic mechanismof buoyancy regulation works in one direction only, i.e.,to increase buoyancy by pumping liquid from thechambers. The decrease in buoyancy, i.e., filling thechambers with liquid is not supported by osmoticmechanisms. To explain this phenomenon, Barskov(1999) suggested capillary transfer of the liquid in thereverse direction. It is most likely that the regulation ofbuoyancy is controlled by both processes, while theseprocesses are restricted to different zones of the siphun-cular cover, as these are derived from different layers oforiginally trifoliate connecting rings of ancient Elles-merocerida. It is possible that the osmotic regulation ofbuoyancy is controlled by those zones of the siphuncu-lar cover which are homologous to the nacreous layer,whereas capillary transport is controlled by the vari-ously mineralized homologues to the spherulite and/orsemiprismatic layers. In addition, the capillary regula-tion of buoyancy may be also controlled by the zones ofsepta and septal necks contacting the connecting rings.Judging from the existing data on the structure of theconnecting rings in various groups of fossil cephalo-pods (Flower, 1957; Hewitt, 1982; Mutvei, 1972, 1997;Druschits et al., 1976), there are several possible typesof the structure of the siphuncular cover suggesting dif-ferent ways of controlling buoyancy.
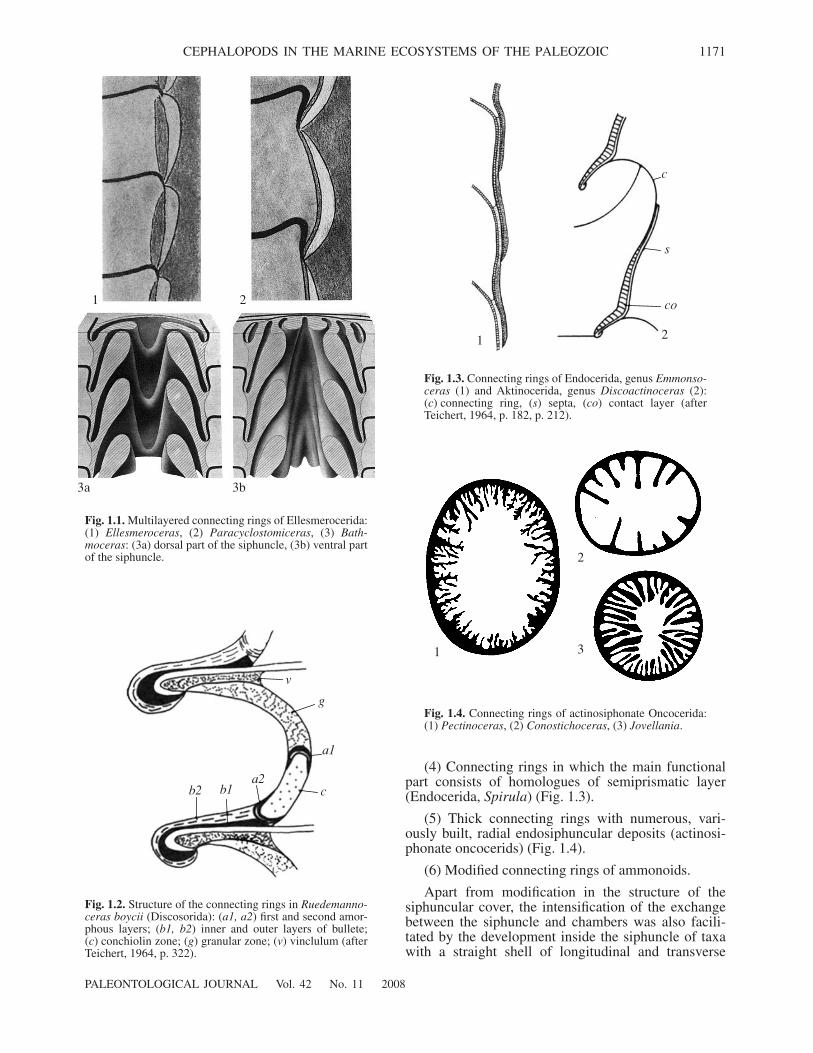
(1) Trifoliate, strongly calcified connecting rings(typically developed in Ellesmerocerida) (Fig. 1.1).
(2) Connecting rings with a transverse differentia-tion of layers (Discosorida) (Fig. 1.2).
(3) Connecting rings in which the main part consistsof homologues of the nacreous layer (Nautilida),whereas the homologues of other layers are locallyrestricted.
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1171
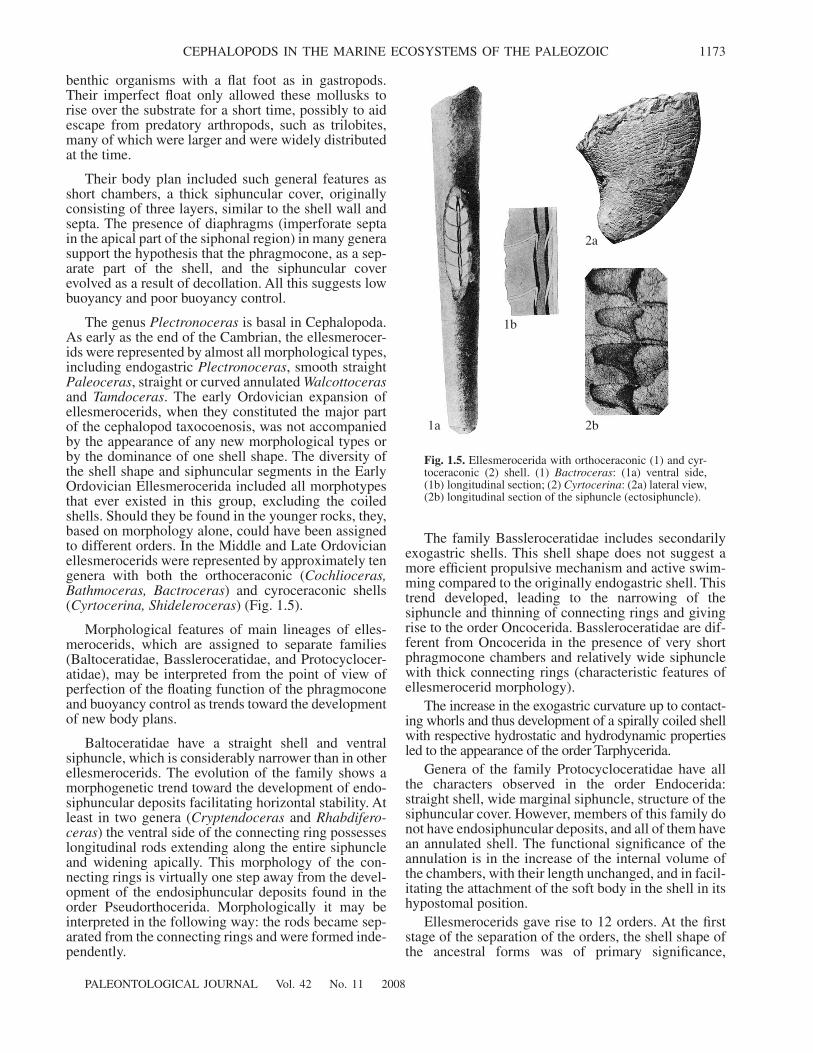
(4) Connecting rings in which the main functionalpart consists of homologues of semiprismatic layer(Endocerida,
Spirula
) (Fig. 1.3).
(5) Thick connecting rings with numerous, vari-ously built, radial endosiphuncular deposits (actinosi-phonate oncocerids) (Fig. 1.4).
(6) Modified connecting rings of ammonoids.
Apart from modification in the structure of thesiphuncular cover, the intensification of the exchangebetween the siphuncle and chambers was also facili-tated by the development inside the siphuncle of taxawith a straight shell of longitudinal and transverse
1 2
3‡ 3b
Fig. 1.1.
Multilayered connecting rings of Ellesmerocerida:(1)
Ellesmeroceras
, (2)
Paracyclostomiceras
, (3)
Bath-moceras
: (3a) dorsal part of the siphuncle, (3b) ventral partof the siphuncle.
v
g
a1
ca2
b1b2
Fig. 1.2.
Structure of the connecting rings in
Ruedemanno-ceras boycii
(Discosorida): (
a1, a2
) first and second amor-phous layers; (
b1, b2
) inner and outer layers of bullete;(
c
) conchiolin zone; (
g
) granular zone; (
v
) vinclulum (afterTeichert, 1964, p. 322).
c
s
co
1 2
Fig. 1.3.
Connecting rings of Endocerida, genus
Emmonso-ceras
(1) and Aktinocerida, genus
Discoactinoceras
(2):(
c
) connecting ring, (
s
) septa, (
co
) contact layer (afterTeichert, 1964, p. 182, p. 212).
1
2
3
Fig. 1.4.
Connecting rings of actinosiphonate Oncocerida:(1)
Pectinoceras
, (2)
Conostichoceras
, (3)
Jovellania
.

1172
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
organic membranes, separating the deposits (Endocer-ida, Intejocerida, Actinocerida).
1.4. Orientation and Support of Orientated Position
This is a second major problem that cephalopodshad to face after having acquired a gas-fluid float. Theproblem of stabilizing the shell in an oriented positioncomfortable for life was solved in a number of differentways:
(1) Using the weight of the wide ventral siphuncle.(2) Through the development of endosiphuncular
deposits.(3) Through the development of deposits inside the
chambers (cameral deposits).These strategies are mostly effective for forms with
a straight shell, since they can provide a horizontalposition, suitable for active swimming.
(4) Coiling in a flat spiral. This is one of the bestways to solve the problem of stability of the animal inthe water, because it results in the close approximationof the centers of gravity and buoyancy, and the animalis constantly in a state of unconditional balance, allow-ing any position in relation to the bottom or the surfaceof water with the minimal energy loss, only using thearms (tentacles), or the funnel.
(5) Decollation of the posterior end of the shell lead-ing to the approximation of the centers of gravity andbuoyancy.
(6) In the shell in the state of unconditional balancethe center of gravity in the phragmocone will be alwaysabove and on the same vertical line with the center ofgravity of the animal in the body chamber. In the shellmorphology this is reflected by a narrow or almostcompletely closed aperture.
Comments on the Origin of the Endosiphuncularand Cameral Deposits
Endosiphuncular deposits and the connecting ringswere formed by the siphuncular epithelium. It is notclear whether the epithelium secreting the endosiphun-cular deposits is a separate epithelial zone like one thatsecretes septa and unattached regions of the connectingrings, or the endosiphuncular deposits are formed in thefinal secretory phase by the same zone of the epitheliumthat forms the connecting rings. In the former case itcan be expected that the endosiphuncular deposits willcontain layers of all three secretory phases: spherulite,nacreous, and semiprismatic. The existing data on themicrostructure of these deposits showing homogenousfine-prismatic structure of the endosiphuncular depos-its and the absence of the repeated layers of variousstructures suggest that the endosiphuncular deposits areformed in the last, semiprismatic secretory phase by thesame epithelial region as the connecting rings. Evi-dently, the endosiphuncular deposits were not massive,
but porous, capable of retaining a large amount of liq-uid to increase the weight of the mollusk.
Despite the recurrent discussion of the problem ofthe cameral deposits, there are still alternative views ontheir origin: (1) The deposits are formed by a mantlezone, which remains in the chambers after these areformed and continues functioning by secreting carbon-ate deposits in the apical part of the shell; (2) No mantleis present in the chambers, whereas the deposits aresecreted by the cameral extrapallial liquid (a substancebetween the mantle and carbonate layers at the anterioredge of the shell during its accretionary growth.Recently, those accepting the former point of viewbrought in new arguments supporting the existence ofthe cameral mantle. Klebaba (1999a; 1999b) suggestedthat the connecting rings are gradually resorbed in theapical zones of the shell in orthocerids, beginning fromthe dorsal side; the mantle grows over the cameral wallsand septa and secretes cameral deposits. Zhuravlevaand Doguzhaeva (1999) discovered structures in cham-bers of some pseudorthocerids and actinocerids, whichthey interpreted as remains of soft tissue and blood ves-sels, and discovered pores in the connecting rings,enclosing these vessels. In our view, all these data arenot confirmed, whereas the structures found are arti-facts that appeared due the incompleteness or poorpreservation of the material studied. Additionally, thehypothesis of the existence of live soft tissue inside theisolated chambers of the siphuncle is quite simplyagainst common sense. Nevertheless, cameral depositscertainly did develop in live mollusks. Their develop-ment was only possible to explain by suggesting that inthe live mollusk the cameral deposits were not strictlyspeaking “deposits,” as it was a series of porous organicmembranes, which served as reservoirs for the cameralliquid which entered the chambers through the porouszones of the connecting rings. It is possible that thechambers contained no unbound fluid at all, which,being highly inert, could have presented difficulties toactive swimming.
1.5. Body Plans and Major Evolutionary Trendsin Cephalopod Orders
(Functional morphological features of the bodyplans of Paleozoic cephalopods and major trends in thechanges in the geological history.)
Order Ellesmerocerida
Flower, 1950 (Late Cam-brian–Late Ordovician). The first cephalopods andknown from the Upper Cambrian beds of northeasternChina (Manzhou). At the end of the Cambrian, thisregion was situated slightly north of the equator, and,judging from paleotectonic reconstructions, occupied aspecific borderline position between the PanthalassaOcean and the just born Paleotethys Ocean, and was thecenter of origin and primary diversification of cephalo-pods. These earliest cephalopod taxa belong to theorder Ellesmerocerida. According to Schindewolf(1933), the earliest ellesmerocerids were crawling
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1173
benthic organisms with a flat foot as in gastropods.Their imperfect float only allowed these mollusks torise over the substrate for a short time, possibly to aidescape from predatory arthropods, such as trilobites,many of which were larger and were widely distributedat the time.
Their body plan included such general features asshort chambers, a thick siphuncular cover, originallyconsisting of three layers, similar to the shell wall andsepta. The presence of diaphragms (imperforate septain the apical part of the siphonal region) in many generasupport the hypothesis that the phragmocone, as a sep-arate part of the shell, and the siphuncular coverevolved as a result of decollation. All this suggests lowbuoyancy and poor buoyancy control.
The genus
Plectronoceras
is basal in Cephalopoda.As early as the end of the Cambrian, the ellesmerocer-ids were represented by almost all morphological types,including endogastric
Plectronoceras
, smooth straight
Paleoceras
, straight or curved annulated
Walcottoceras
and
Tamdoceras
. The early Ordovician expansion ofellesmerocerids, when they constituted the major partof the cephalopod taxocoenosis, was not accompaniedby the appearance of any new morphological types orby the dominance of one shell shape. The diversity ofthe shell shape and siphuncular segments in the EarlyOrdovician Ellesmerocerida included all morphotypesthat ever existed in this group, excluding the coiledshells. Should they be found in the younger rocks, they,based on morphology alone, could have been assignedto different orders. In the Middle and Late Ordovicianellesmerocerids were represented by approximately tengenera with both the orthoceraconic (
Cochlioceras,Bathmoceras, Bactroceras
) and cyroceraconic shells(
Cyrtocerina, Shideleroceras
) (Fig. 1.5).
Morphological features of main lineages of elles-merocerids, which are assigned to separate families(Baltoceratidae, Bassleroceratidae, and Protocyclocer-atidae), may be interpreted from the point of view ofperfection of the floating function of the phragmoconeand buoyancy control as trends toward the developmentof new body plans.
Baltoceratidae have a straight shell and ventralsiphuncle, which is considerably narrower than in otherellesmerocerids. The evolution of the family shows amorphogenetic trend toward the development of endo-siphuncular deposits facilitating horizontal stability. Atleast in two genera (
Cryptendoceras
and
Rhabdifero-ceras
) the ventral side of the connecting ring possesseslongitudinal rods extending along the entire siphuncleand widening apically. This morphology of the con-necting rings is virtually one step away from the devel-opment of the endosiphuncular deposits found in theorder Pseudorthocerida. Morphologically it may beinterpreted in the following way: the rods became sep-arated from the connecting rings and were formed inde-pendently.
The family Bassleroceratidae includes secondarilyexogastric shells. This shell shape does not suggest amore efficient propulsive mechanism and active swim-ming compared to the originally endogastric shell. Thistrend developed, leading to the narrowing of thesiphuncle and thinning of connecting rings and givingrise to the order Oncocerida. Bassleroceratidae are dif-ferent from Oncocerida in the presence of very shortphragmocone chambers and relatively wide siphunclewith thick connecting rings (characteristic features ofellesmerocerid morphology).
The increase in the exogastric curvature up to contact-ing whorls and thus development of a spirally coiled shellwith respective hydrostatic and hydrodynamic propertiesled to the appearance of the order Tarphycerida.
Genera of the family Protocycloceratidae have allthe characters observed in the order Endocerida:straight shell, wide marginal siphuncle, structure of thesiphuncular cover. However, members of this family donot have endosiphuncular deposits, and all of them havean annulated shell. The functional significance of theannulation is in the increase of the internal volume ofthe chambers, with their length unchanged, and in facil-itating the attachment of the soft body in the shell in itshypostomal position.
Ellesmerocerids gave rise to 12 orders. At the firststage of the separation of the orders, the shell shape ofthe ancestral forms was of primary significance,
1‡ 2b
2a
1b
Fig. 1.5.
Ellesmerocerida with orthoceraconic (1) and cyr-toceraconic (2) shell. (1)
Bactroceras
: (1a) ventral side,(1b) longitudinal section; (2)
Cyrtocerina
: (2a) lateral view,(2b) longitudinal section of the siphuncle (ectosiphuncle).

1174
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
although in later evolution the external shell morphologycould become considerably different. The discussionbelow begins with orders, which originally had a straightor slightly curved exogastrically longiconic shell.
The presence of a straight shell is adaptively usefulonly when it is accompanied by a mechanism allowingthe horizontal orientation of the shell.
Order Yanhecerida
Chen et Qi, 1979 (Late Cam-brian or Early Ordovician). Endemic Chinese taxa witha straight shell with all characteristic features of Elles-merocerida, including low chambers of the phragmo-cone and a wide siphuncle with tubular segments. Thepresence of siphonal deposits is a fundamental func-tional difference allowing their placement in a separateorder. The structure of these deposits is still unknown.However, the very presence of these deposits in theshell as a mechanism of orientation and stability of theshell certainly suggests a different body plan from thatof Ellesmerocerida, the active swimming ability, andpossible presence of an organ allowing this function. InEllesmerocerida these mechanisms were absent. Theorigin and further evolution of Yanhecerida remainunknown.
Order Protactinocerida
Chen et Qi, 1979 (LateCambrian or Early Ordovician of China). Like in Yan-hecerida, the appearance of the straight shell wasaccompanied by the siphonal deposits, which servedthe same function but were based on a different mor-phology of the siphuncle. In Protactinocerida, thesiphuncular segments expand into the phragmoconechambers, and the siphuncle is moniliform. The differ-ent morphology of the siphuncle in these orders suggestdifferent structures, which may be used as evidence oftheir taxonomic separation. The morphology and struc-ture of the endosiphuncular in Protactinocerida are alsounknown. The outline of the siphuncular segments aresimilar to Actinocerida, which appeared later, and as isseen from their name, were their supposed ancestors.However, a considerable stratigraphic gap betweenthese orders and the unknown structure of their endosi-phuncular deposits do not allow a positive link.
If the apomorphy that separated cephalopods fromother mollusks, which for the first time allowed macro-scopic shelled organisms to take off from the bottom,be compared to the invention of the hot air balloon, thenYanhecerida and Proactinocerida became the first zep-pelins capable of active directional movement.
Order Endocerida
Teichert, 1933 (Early–LateOrdovician). High phragmocone chambers, widesiphuncle, and diversity in the structure of the siphun-cular cover, all this suggests a more efficient means ofbuoyancy control. During the Ordovician, Endoceridawere the largest bottom-dwelling pelagic animals,active predators at the top of the trophic pyramid. Per-haps, from that time, the roles changed and the bottom-dwelling cephalopods instead of being the prey ofarthropods became their predators.
At the same time the mechanism of stability controlarose, which worked by filling the siphuncle with con-tinuous endosiphuncular deposits, coating the connect-ing rings, as in Yanchecerida. The continuous deposi-tion inside the siphuncle interrupted the communica-tions between the siphuncle and chambers andeventually prohibited buoyancy control in the zones ofthe phragmocone where they were present. Thus thisbody plan was seriously internally controversial. At theend of the Ordovician or at the very beginning of theSilurian Endocerida became extinct.
Order Intejocerida
Balashov, 1960 (Early Ordovi-cian). The attempts to find a compromise between theneed in the increasingly heavy apical end of the phrag-mocone by development of endosiphuncular depositsand retention of communications between the siphun-cle and the chambers through a series of longitudinalorganic membranes, which cut through the massivedeposits and worked as channels connecting thesiphuncular epithelium with the chambers (importantfor buoyancy control) were unsuccessful. This mor-phology was characteristic of the order Intejocerida,which branched off the Endocerida in the Early Ordov-ician. This approach was not efficient, and Intejoceridabecame extinct as early as the Early Ordovician. Never-theless, as shown below, this method of intensificationof the exchange between the siphuncle and the cham-bers was used on several occasions.
Order Orthocerida
Kuhn, 1940 (Early Ordovi-cian–Triassic). Representatives of this order had astraight or weakly curved exogastric shell. In addition,orthocerids had two more fundamental features thatseparated them from all previously evolved orders:appearance of the mechanism controlling orientationand stability of the shell (cameral deposits) and changesin early ontogeny. The former was first tried in thecoiled and secondarily straight tarphycerids and lituit-ids, and the latter evolved for the first time in orthocer-ids, but was later inherited by the entire evolutionarylineage leading to ammonoids and coleoids. Morpho-logically, this can be observed as a formation of a smallspherical chamber (protoconch). The presence of theprotoconch suggests an incompletely formed hydro-static apparatus of one chamber only at the first post-embryonal stage. This means that in contrast to Elles-merocerida, a group with no protoconch and with aphragmocone of several chambers at the first post-embryonal stage, orthocerids were capable of earlieraccess to the pelagic zone (before they reached thephragmocone stage). This, on the one hand, promoted awider distribution, and, on the other hand, openedopportunities of evolutionary changes at early ontoge-netic stages. The realization of the wide distributionopportunities is supported by the known fact that in thesecond half of the Ordovician and in the Silurian, ortho-cerids were the most widespread cephalopod group andexisted for about 300 million years, until the end of theTriassic, and possibly longer. The possibilities of theevolutionary changes at the early stages were to a full

PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1175
extent used by bacritids (descendants of orthocerids,and further, by ammonoids and coleoids, which domi-nated in the Jurassic and Cretaceous). For a moredetailed description of major features of the evolutionof early cephalopods see Barskov (1989).
Order Pseudorthocerida
Barskov, 1968 (EarlyOrdovician–Triassic). The treatment of this taxon as anorder appears to have been a simple increase in the rankof the superfamily Pseudorthoceratacea (previously inthe order Orthocerida) (Barskov, 1968). There were twomajor features of pseudorthocerids that supported theidea of a higher rank for this taxon: (1) Absence of aprotoconch; (2) Presence of endosiphuncular deposits,different from pendant deposits, found in Actinocerids,from the endocones of endocerids, polyptychocones ofearly discosorids and from endosiphuncular deposits,which are present in some later genera of orthocerids(family Geisonoceratidae). In “typical” Late Paleozoicpseudorthocerids the endosiphuncular deposits are ini-tially formed in the septal foramen (as in Actinoceridaand Orthocerida), but later spread only adorally, form-ing the lining of the connecting rings of the next pre-ceding segment of the siphuncle, where they becomethicker, but do not merge with the deposits of this seg-ment. Thus, the deposits are formed in such a way thatthe communication between the siphuncle and cham-bers is maintained in almost all chambers of the phrag-mocone. A detailed study of the Early Paleozoic repre-sentatives showed that the development of the endosi-phuncular deposits in this cephalopod group toward thestage typical of the Late Paleozoic taxa was very grad-ual. Originally these deposits developed only adapi-cally, like endocones (opistoneckal deposits) later bothadorally and adapically (bilocal deposits). Only in theSilurian they acquired a typical shape spreading onlyadorally from their place of origin in the septal foramen(Barskov, 1972, 1989). Thus, the establishment of thetypical body plan of pseudorthocerids was a very grad-ual process.
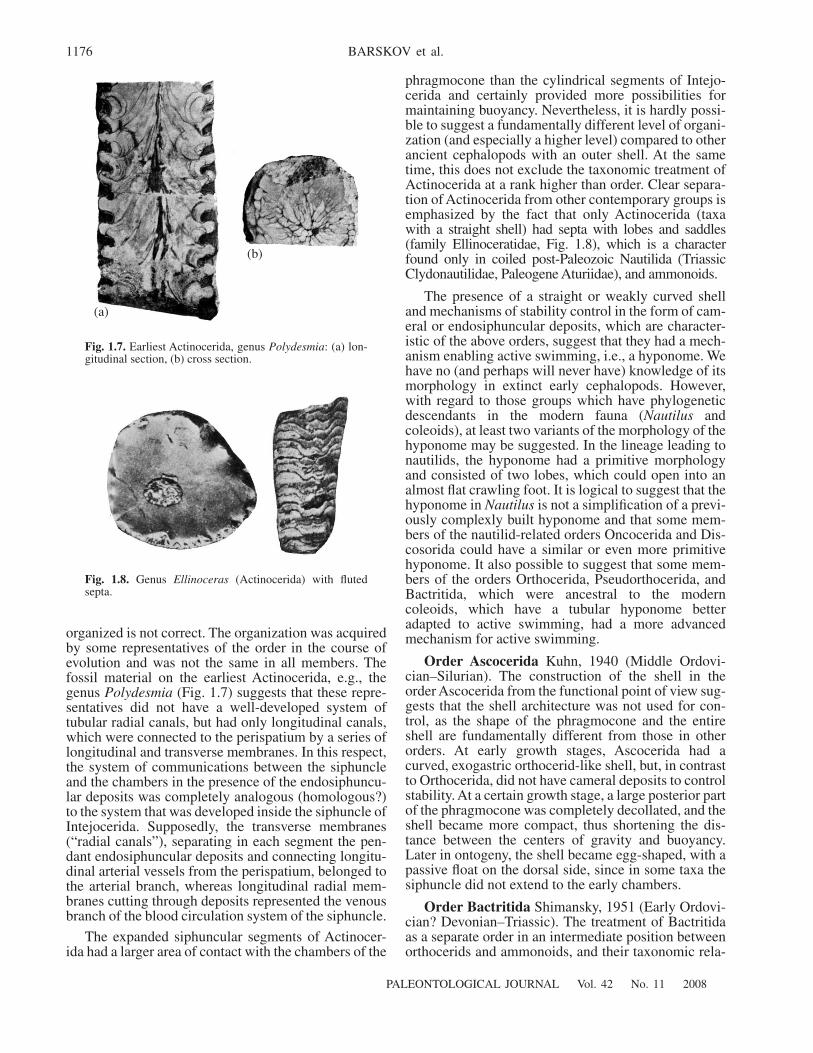
Order Actinocerida
Teichert, 1933 (Early Ordovi-cian–Middle Carboniferous). Morphologically, Acti-nocerida continue the evolutionary trend that beganwith the order Protactinocerida. Characteristic featuresof this body plan include siphonal segments expandinginto chambers, up to the development of rounded andnummuloid and development of endosiphunculardeposits. The expansion of the siphuncular segmentssuggests an increase in the exchange between thesiphuncle and the chambers by increasing the surfacearea of the exchange, and contributing towards horizon-tal stability comfortable for active swimming. The con-flict between the need for regulation of stability and itsunavoidable limitation by the development of depositsin the apical regions led to the development in Actinoc-erida of so-called pendant deposits. They did not linethe connecting rings, like endocones in endocerids, butwere formed discretely in each septal foramen andspread within the siphuncle pressing the connective tis-sues of the siphuncle and leaving longitudinal and
transverse spaces for blood vessels and deposit-freeperipheral space near connecting rings (perispatium) ineach segment, where the exchange between the siphun-cle and phragmocone chambers took place. Usually, thestructure of the siphuncle in Actinocerida is interpretedas follows. Longitudinal canals run along the entirelength of the siphuncle (they most certainly were part ofthe blood system), arterial (in Nautilus arteries withperistaltic epithelium) and venous (in Nautilus a systemof lacunas in the connective tissue lacking epithelium).Each segment also possessed radial canals providingthe connection with the perispatium, possessing epithe-lium responsible for emission of gas into the chambersof phragmocone and for liquid extraction and removalto maintain buoyancy. The reconstructions of thesiphuncle in Actinocerida led to interpretation of radialcanals as tubular structures, similar to longitudinalcanals (Fig. 1.6). This led to the conclusion that,because the both arterial and venous branches of theblood systems are tubular, the venous branch had bloodvessels similar to those in the arterial branch. Thus, thefundamental organization in Actinocerida was moredeveloped than even in the modern Nautilus, whichdoes not have true vessels (i.e., with their own walls) inthe venous branch of the blood circulatory system. Thisreconstructed endosiphuncular system and its interpre-tation are the basis for the separation of Actinocerida asa high-rank taxon, originally as a superorder (Shiman-sky and Zhuravleva, 1961) and later as a subclass(Teichert, 1964).
However, the interpretation of the intrasiphuncularsystem of Actinocerida as of being originally highly
Fig. 1.6. Reconstructed siphuncle in Actinocerida (afterTeichert, 1964, p. 196).
1176
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
organized is not correct. The organization was acquiredby some representatives of the order in the course ofevolution and was not the same in all members. Thefossil material on the earliest Actinocerida, e.g., thegenus Polydesmia (Fig. 1.7) suggests that these repre-sentatives did not have a well-developed system oftubular radial canals, but had only longitudinal canals,which were connected to the perispatium by a series oflongitudinal and transverse membranes. In this respect,the system of communications between the siphuncleand the chambers in the presence of the endosiphuncu-lar deposits was completely analogous (homologous?)to the system that was developed inside the siphuncle ofIntejocerida. Supposedly, the transverse membranes(“radial canals”), separating in each segment the pen-dant endosiphuncular deposits and connecting longitu-dinal arterial vessels from the perispatium, belonged tothe arterial branch, whereas longitudinal radial mem-branes cutting through deposits represented the venousbranch of the blood circulation system of the siphuncle.
The expanded siphuncular segments of Actinocer-ida had a larger area of contact with the chambers of the
phragmocone than the cylindrical segments of Intejo-cerida and certainly provided more possibilities formaintaining buoyancy. Nevertheless, it is hardly possi-ble to suggest a fundamentally different level of organi-zation (and especially a higher level) compared to otherancient cephalopods with an outer shell. At the sametime, this does not exclude the taxonomic treatment ofActinocerida at a rank higher than order. Clear separa-tion of Actinocerida from other contemporary groups isemphasized by the fact that only Actinocerida (taxawith a straight shell) had septa with lobes and saddles(family Ellinoceratidae, Fig. 1.8), which is a characterfound only in coiled post-Paleozoic Nautilida (TriassicClydonautilidae, Paleogene Aturiidae), and ammonoids.
The presence of a straight or weakly curved shelland mechanisms of stability control in the form of cam-eral or endosiphuncular deposits, which are character-istic of the above orders, suggest that they had a mech-anism enabling active swimming, i.e., a hyponome. Wehave no (and perhaps will never have) knowledge of itsmorphology in extinct early cephalopods. However,with regard to those groups which have phylogeneticdescendants in the modern fauna (Nautilus andcoleoids), at least two variants of the morphology of thehyponome may be suggested. In the lineage leading tonautilids, the hyponome had a primitive morphologyand consisted of two lobes, which could open into analmost flat crawling foot. It is logical to suggest that thehyponome in Nautilus is not a simplification of a previ-ously complexly built hyponome and that some mem-bers of the nautilid-related orders Oncocerida and Dis-cosorida could have a similar or even more primitivehyponome. It also possible to suggest that some mem-bers of the orders Orthocerida, Pseudorthocerida, andBactritida, which were ancestral to the moderncoleoids, which have a tubular hyponome betteradapted to active swimming, had a more advancedmechanism for active swimming.
Order Ascocerida Kuhn, 1940 (Middle Ordovi-cian–Silurian). The construction of the shell in theorder Ascocerida from the functional point of view sug-gests that the shell architecture was not used for con-trol, as the shape of the phragmocone and the entireshell are fundamentally different from those in otherorders. At early growth stages, Ascocerida had acurved, exogastric orthocerid-like shell, but, in contrastto Orthocerida, did not have cameral deposits to controlstability. At a certain growth stage, a large posterior partof the phragmocone was completely decollated, and theshell became more compact, thus shortening the dis-tance between the centers of gravity and buoyancy.Later in ontogeny, the shell became egg-shaped, with apassive float on the dorsal side, since in some taxa thesiphuncle did not extend to the early chambers.
Order Bactritida Shimansky, 1951 (Early Ordovi-cian? Devonian–Triassic). The treatment of Bactritidaas a separate order in an intermediate position betweenorthocerids and ammonoids, and their taxonomic rela-
(a)
(b)
Fig. 1.7. Earliest Actinocerida, genus Polydesmia: (a) lon-gitudinal section, (b) cross section.
Fig. 1.8. Genus Ellinoceras (Actinocerida) with flutedsepta.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1177
tionship to coleoids are generally agreed. However, themorphological traits of Bactritida, shared with the orderOrthocerida, from which bactritids evolved, are lessfrequently discussed. Characters that are different fromorthocerids include: marginal position of the siphuncle,which is in contact with the shell wall (typical orthocer-ids have a central or almost central siphuncle), and theabsence of the cameral and endosiphuncular deposits inAscocerida. Shared characters include a straight orweakly curved shell and the presence of a protoconch.Many Late Silurian orthocerids had eccentric siphuncleclosely approximating to the ventral side of the shell,but not touching it. This morphology makes the rele-vant genera more similar to Bactritida than to Orthocer-ida. Hence, it is necessary to determine fundamentalfunctional consequences of the marginal siphuncle, andof the absence (as in some other straight non-ammonoid cephalopods) of cameral deposits. Thesefunctional differences are so highly specific that it isquite impossible to assign Bactritida to Orthocerida, oreven to Ammonoidea.
The absence of the cameral and endosiphunculardeposits in Bactritida with a straight shell suggests thatthey did not have a mechanism for orientation of stabil-ity control for the horizontal position, which was typi-cally present in all earlier cephalopod groups with astraight shell (orthocerids, pseudorthocerids, actinocer-ids, and endocerids). Hence, bactritids either could notorientate the shell and body horizontally, or they had adifferent mechanism to control stability. In the first caseit would be necessary to assume that these animals’ liveorientation in water was hypostomic, i.e., with the api-cal end facing up, or at an angle. The shell of bactritidsis longiconic, i.e., the animal had high buoyancy andtherefore bactritids apparently inhabited the upperpelagic zone. The pelagic affinity of bactritids is sup-ported by their small subspherical protoconch, suggest-ing a large number of small-sized eggs and, hence,direct or larval development. Hatchlings were pelagic.
The near-wall position of the siphuncle has impor-tant functional consequences. The siphuncle in ortho-cerids was enclosed in a hard tube composed of con-necting rings and was central or subcentral. Apart fromits main function, this siphuncular tube certainly serveda structure connecting and enforcing the unattachedpart of the septum. The transition of the siphuncle to theventral wall and its contact with the shell wall made thefree parts of the septa unconnected. As a natural conse-quence, the septum became curved to form a wideomnilateral lobe, morphological structure characteris-tic of bactritids that was inherited by ammonoids andinitiated the process of increasing sutural complexity.The attachment of the siphuncle to the shell wall natu-rally led the suture to break at this spot to form a necklobe, which was also inherited by ammonoids. In addi-tion, the ventral position of the siphuncle changed themeans of buoyancy and stability control and shell ori-entation. It is known that Nautilus controls its buoyancyby filling the chambers with liquid, and its osmotic
removal through the siphuncle. Reyment (1973) exper-imentally showed that the amount of liquid in thephragmocone chambers sufficient for neutral buoyancyis minimal in the involute Nautilus-like shells, andincreased in more evolute taxa. The presence of liquidin the chambers imposes significant constraints toactive swimming due to inertia of this liquid. Whileslowing down and stopping the liquid would moveinside the chamber. Cameral deposits are one of theways to reduce this effect. It is quite evident that in thelive animal these were not massive, heavy deposits, asare found in fossils, but a system of organic or weaklycalcified membranes capable of absorbing and retain-ing significant amounts of liquid. The porous skeletonsof extant sepiids are a functional analogue of such astructure. Liquid tied within the micropores does nothave its own inertia and does not impede active move-ment. In straight shells with a non-marginal siphuncle(Orthocerida, Pseudorthocerida), in which the cameraldeposits were present on the ventral side, the siphuncleretained its function. In bactritids, which had a siphun-cle in contact with the ventral wall, the development ofthe cameral would have precluded the siphuncle fromfunctioning normally.
Evidently, in the horizontally orientated bactritidshell, in which the chambers are filled with liquid, thesiphuncle would have also become dysfunctional.
A straight longiconic shell has very high buoyancy,which increases as the shell grows. An animal lacking amechanism for increasing the weight of the shell wouldhave to have lived very near the water surface, possiblyeven with the apical end sticking out of the water, or tofloat on the surface in a non-orientated position.Clearly, this situation is highly improbable. Therefore,a mechanism for increasing the shell weight and forbuoyancy control in bactritids must have existed, butwas apparently different from those discussed above.
A marginal siphuncle is different in that the func-tional epithelium of the siphuncle responsible for fillingthe chambers with liquid and its removal is directlyconnected with the organic membranes covering theshell wall and septa (in taxa with a non-marginalsiphuncle this connection is performed through con-necting rings, which in primitive forms are quite com-plexly built).
Soaking of liquid by the organic lining of the shellwalls and the septa may decrease buoyancy, whereasthe direct contact of the porous organic membraneswith the siphunclular epithelium makes this controleasier.
The curvature of septa resulting in development oflobes and saddles increase their surface and potentialfor buoyancy control. The increase in weight of thephragmocone resulting from the liquid tied in the mem-branes and the minimal quantity of free liquid in thephragmocone chambers, which, due to its inertia, couldhave hampered active movement, promoted the propul-sive swimming. It is possible that this means of buoy-
1178
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
ancy control was inherited by ammonoids from theancestral bactritids and is the most probable reason forthe septa (and sutures) of ammonoids and nautiloidsbeing so different (Barskov, 1999). In this respect, bac-tritids are closer to ammonoids than to orthocerids, andcan justifiably be assigned to the same superorder asammonoids (as in most western literature), or be recog-nized as a separate taxon of the same taxonomic rank asammonoids (as in the Russian cephalopod literature;see Zhuravleva and Shimansky, 1961; Shimansky,1979; Shevyrev, 2005).
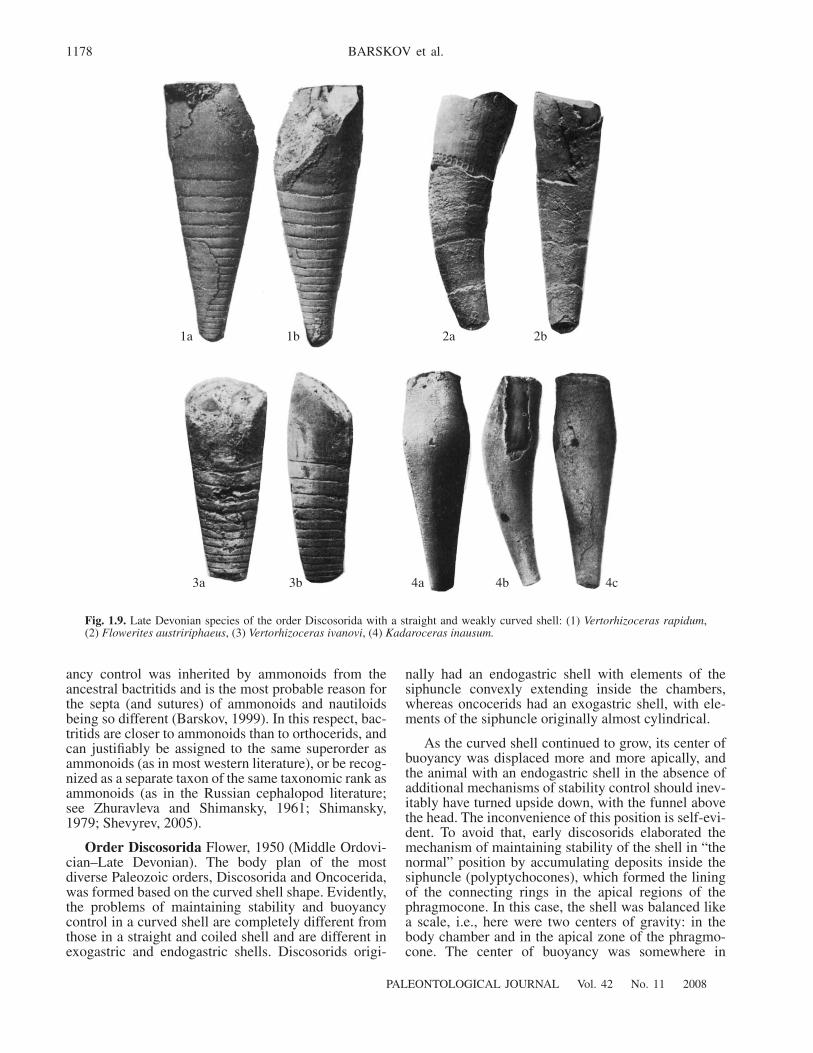
Order Discosorida Flower, 1950 (Middle Ordovi-cian–Late Devonian). The body plan of the mostdiverse Paleozoic orders, Discosorida and Oncocerida,was formed based on the curved shell shape. Evidently,the problems of maintaining stability and buoyancycontrol in a curved shell are completely different fromthose in a straight and coiled shell and are different inexogastric and endogastric shells. Discosorids origi-
nally had an endogastric shell with elements of thesiphuncle convexly extending inside the chambers,whereas oncocerids had an exogastric shell, with ele-ments of the siphuncle originally almost cylindrical.
As the curved shell continued to grow, its center ofbuoyancy was displaced more and more apically, andthe animal with an endogastric shell in the absence ofadditional mechanisms of stability control should inev-itably have turned upside down, with the funnel abovethe head. The inconvenience of this position is self-evi-dent. To avoid that, early discosorids elaborated themechanism of maintaining stability of the shell in “thenormal” position by accumulating deposits inside thesiphuncle (polyptychocones), which formed the liningof the connecting rings in the apical regions of thephragmocone. In this case, the shell was balanced likea scale, i.e., here were two centers of gravity: in thebody chamber and in the apical zone of the phragmo-cone. The center of buoyancy was somewhere in
1‡ 1b 2‡ 2b
3‡ 3b 4‡ 4b 4c
Fig. 1.9. Late Devonian species of the order Discosorida with a straight and weakly curved shell: (1) Vertorhizoceras rapidum,(2) Flowerites austririphaeus, (3) Vertorhizoceras ivanovi, (4) Kadaroceras inausum.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1179
between. This organization imposed serious limitationson the possibility of active swimming. The shell couldonly function as a passive float. A normal position ofthese animals is hypostomic, and many such forms hada narrow aperture. Two trends may be recognized in theevolution of Discosorids: maintenance of structures forpassive floating and an inactive mode of life, whichsuggest a weak propulsive mechanism. The shellbecame widely conical (the apical angle increased).The second trend lead to the acquisition of the weaklycurved, almost straight shell, more adapted for activeswimming. The latter approach was realized at the endof the order’s existence in the Late Devonian andresulted in the appearance of a number of genera withvarious kinds of straight shell (Fig. 1.9).
Order Oncocerida Flower, 1950 (Middle Ordovi-cian–Early Carboniferous). The original shell shape isan exogastric cyrtoceracone. The continuing growthwith an absence of additional mechanisms of stabilitycontrol, an animal with such a shell shape had a liveposition promoting better possibilities for swimmingthan endogastric shells. However, even with this orga-nization and shell shape, the possibilities of improve-ment of swimming and, hence, the development of thepropulsive mechanism were limited. As early as thevery beginning of their evolution in the Middle Ordov-ician, several major lineages were separated withinoncocerids. These lineages had different morphology sug-gesting that they approached the problems of stability con-trol and of active swimming differently (or rejected theactive swimming). There are four initial morphologicalvarieties (Sweet, 1964a; Zhuravleva, 1994):
(1) Graciloceratidae–Oncoceratidae—exogastric cyr-toceracones with a narrow deposit-free siphuncle.
(2) Tripteroceratidae—orthoceracones with a nar-row deposit-free siphuncle and a characteristically sub-triangular shell cross-section.
(3) Valcouroceratidae—exogastric cyrtoceraconeswith a wider siphuncle and actinosiphonate structureswithin it.
(4) Diestoceratidae—endogastric shell with an acti-nosiphonate siphuncle.
The above shows that there are two groups of onco-cerids with a varying siphuncle structure: the twoformer families have a relatively narrow siphuncle,with no endosiphuncular deposits, whereas the lattertwo possess actinosiphonate deposits. These structuresrepresent radial outgrowths of connecting rings fromthe siphuncular wall to its middle. There are at least fivetypes of actinosiphonate structures, different in thenumber, thickness, and direction of these outgrowths(Sweet, 1964a). From the formal morphological posi-tion, the differences between the actinosiphonate andnonactinosiphonate groups may suggest significant dif-ferences in the body plan, which supports their separa-tion at the order level, as was suggested by Teichert(1933, 1939), who proposed the order Cyrtoceroideafor actinosiphonate taxa and the order Gomphoceroidea
for nonactinosiphonate taxa. However, as was alreadysuggested by Hyatt (1900), who separated cyrtoconiccephalopod shells from orthoconic and coiled shells,which were later recognized as the order Oncocerida,actinosiphonate structures develop independently, andare not a high ranked character. Similar structures arealso observed in later representatives of another order(Discosorida).
Three morphogenetic trends are observed in almostall of these original branches: straightening of the shell,coiling of the shell, and the development of widely con-ical rapidly expanding shells with narrowing and closedaperture. In the branch beginning with the Ordoviciancyrtoceraconic Valcouroceratidae with an actinosipho-nate siphuncle, the tendency to straightening of theshell is clearly tracked in the lineage Jovellaniidae (Sil-urian)–Tripleuroceratidae (Early Devonian–Carbonif-erous?)–Aktjubochilidae (Late Devonian). The ten-dency towards coiling the shell in this lineage lead tothe separation of the family Naedyceratidae with agyroceraconic and a low-trochoid shell and the Devo-nian genera Notoceras, Kotelnikoceras, and Loriero-ceras from the family Notoceratidae.
Another initial branch related to the persistent fam-ily Oncoceratidae, in the Devonian, gave rise to thefamily Ptenoceratidae, which included ornamentedtaxa with a gyroceraconic, trochoceraconic, and natulil-iconic shell, which are assigned by some authors to theorder Nautilida. In the Devonian this lineage alsoexhibited a third tendency, i.e., development of widelyconical shells with a closed multilobed aperture (Tri-meroceratidae).
Somewhat earlier, in the Silurian, the morphologi-cally similar forms (family Hemiphragmoceratidae)appeared in the other actinosuphuncular branch fromthe exogastric rather than endogastric taxa, related tothe initial family Diestoceratidae.
In the course of subsequent evolution the majority ofthe genera became adapted to a passive floating way oflife by developing a short-conical shell functioning as apassive float, similar to that of discosorids. The live ori-entation of the animal was also hypostomic. In theadults of some taxa, the aperture was almost completelyclosed (Fig. 1.10), leaving only small openings for thefunnel, arms, and eyes. Another change also related tothe permanently hypostomic orientation was polymer-ization of the attachment muscles at the base of thebody chamber. At the same time, taxa with a straightorthoceraconic shell appeared many times amongoncocerids, suggesting that active swimming wasbecoming more important.
The acquisition of the coiled planispiral shell incephalopods solved two problems: it retained the com-pactness of the shell during the long period of growth,and maintained the more or less stable position of thecenters of buoyancy and gravity, which facilitatedmaintenance of the orientated position in the water. Inthe evolution of cephalopods, taxa with a coiled shell

1180
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
appeared at least five or six times, at various taxonomiclevels. The appearance of a few taxa with a coiled shellin several lineages in the order Oncocerida did notresult from the initial body plan, in contrast to thegroups discussed below, in which the coiled shell was abasis for their subsequent evolution. Later these groupscould also include forms with other types of organiza-tions, including orthoconic and cyrtococnic, but the ini-tial morphology of those was, in contrast to Oncocer-ida, a coiled shell.
Paleozoic taxa with the originally coiled shell, areassigned either to one order Tarphycerida (Balashov,1962), or to two orders Tarphycerida and Barrandeo-cerida (Moore, 1964), or Tarphycerida and Lituitida(Starobogatov, 1983, Shevyrev, 2006a). Are the func-tional features of members of the three orders suffi-ciently different for their morphological implications tobe recognized as separate body plans and for the taxo-nomic substantiation of the order rank?
The structure of the siphunclular cover (thick, mul-tilayered connecting rings) and its relatively largediameter at least in early representatives of in early Tar-phycerida and Lituitida, was inherited from primitiveEllesmerocerida, suggesting an incomplete exchangefunction of the siphuncle. In this respect these groupswere at an “Ellesmerocerid” stage of buoyancy control.This imperfection of the exchange function of thesiphuncle became an impediment offsetting the advan-tages of the coiled shell. In the process of evolution,both orders experienced reversed morphological evolu-tion and returned to the morphology of their ancestors:to the curved or even straight shell (genus Rhyncho-rthoceras), which is particularly distinctly observed inthe morphological changes throughout shell ontogeny.At early stages the shell is coiled, later it becomes pro-gressively uncoiled and eventually almost straight for amost of its length. This is especially clearly observed inLituitida (Fig. 1.11). In Tarphycerida the shell uncoilsat the end of the last volution. In all tarphycerids, pres-ervation permitting, the apertural part does not overlapthe last whorl. This morphology suggests that theseammonoids, because of the way their shells were built,could not maintain a position when the axis of the jetwould be on the same horizontal axis as the center ofgravity of the animal. This excluded the possibility ofjet propulsion in their movement. The coiled part of theshell is solely a compact float. Thus, members of Tar-phycerida and Lituitida, if they did not have additionalmechanisms for stability control, could have only beenpassively floaters that were hypostmously orientated(aperture facing down).
The presence in many such taxa of a narrowed orclosed hypostome confirm this very clearly (Fig. 1.12).However, at least some of them developed cameraldeposits which provided the animal with an additionalstability and orientated position suitable for propulsion(Fig. 1.13). This mechanism of orientation is the mostefficient for taxa with a straight shell and its appearancein coiled cephalopods is a unique feature, which is onlyfound in Tarphycerida and Lituitida, and is unknown inlater taxa with a coiled shell (Natulida, Ammonoida,some families of Oncocerida) and even in the orderBarrandeocerida, direct descendants of Tarphycerida.
1‡2‡
1b 2b
Fig. 1.10. Oncocerida with an almost closed aperture:(1) genus Octamerella, (2) genus Inversoceras; (a) aper-tural and (b) lateral views.
Fig. 1.11. Heteromorphic shell of the genus Ancistroceras (Lituitida).

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1181
Representatives of the order BarrandeoceridaFlower, 1950 (Middle Ordovician–Late Devonian),which had a coiled shell (like their ancestral group Tar-phycerida) and Lituitida display the following funda-mental features of their organization. The connectingrings are thin, and have a diameter and structure similar
to those in the later taxa, suggesting a more advancedexchange function of the siphuncle. Originally, theshell of Barrandeocerida had a larger expansion rate.This is essential because in this case the body chamber,given all other variables unchanged, has a larger vol-ume and is isometric, rather than vermiculate in shape.This suggest a larger mantle cavity, allowing a largemass of propulsive muscles. These features suggest thatBarrandeocerida was a more advanced group. The adap-tive radiation of this order at the end of the Ordovician andduring the Silurian and Devonian led to the appearance oftaxa with a heteromoph shell morphologically similar tothat in the earlier genera of Tarphycerida and Lituitida. Inaddition, these were the first cephalopods with a trochoid,rather planispiral shell (Fig. 1.14).
A trend toward coiling into a compact flat spiral to afull extent was realized in the order Nautilida Agassiz,
Fig. 1.12. Genus Ophioceras (Tarphycerida) with a stronglynarrowed aperture.
1‡
1b
2
4
5
3
6
Fig. 1.13. Cameral deposits of Lituitida. (1) Lituites andTarphycerida: (1a) longitudinal section and (1b) lateralview. (2) Centrotarphyceras, (3) Aphetoceras, (4, 5) Curto-ceras, (6) Campbelloceras.
1‡
1b
2‡ 2b
Fig. 1.14. Barrandeocerida with a trochoid shell:(1) Sphyradoceras, (2) Piesmoceras; (a) apertural and(b) lateral views.

1182
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
1847 (Early Devonian–Recent), the major families ofwhich directly or indirectly evolved from Oncocerida(Kümmel, 1964, p. K412). However, in contrast to allother orders, the principle characters of the nautilidbody plan (compact planispiral shell, thin subcentralsiphuncle) were not acquired instantaneously. Theiracquisition was a very long process, which was eventu-ally completed in the Late Paleozoic (Shimansky,1979).
The assignment of the Devonian genera with anuncoiled shell to Nautilida is not unequivocally based,whereas the origin of the Late Paleozoic true Nautilidais unknown (Kümmel, 1964, p. K412). The earliestmembers had a cyrtoconic, gyroconic, loosely trochoid,and almost straight shells, which are not characteristicof typical Nautilida. The siphuncle in most nautilidswas ventral. All these parameters position these taxacloser to their contemporary Oncocerida. Essentially,the only character that unites all these genera and dis-tinguishes them from Oncocerida is the presence ofspecific ornamentation in the form of wing-like lateralprotrusions.
AmmonoidsFunctionally, the origin of ammonoids is related to a
change in the mechanism of maintaining stability andorientation and in the change in buoyancy control. Thehypothesis of their origin from the order Bactritida(a small group of subclass rank or within the subclassAmmonoidea) is currently widely accepted.
The key features of the ammonoid body plan can bedescribed in three major morphological parameters:coiled shell, complex suture, and the presence of a pro-toconch. As shown above, coiling of the shell has anobvious function, i.e., retention of a compact shellthroughout the long growth period and facilitating sta-bility control by approximating the centers of gravityand buoyancy and the position of indifferent balance.The tightly coiled ammonoid shell was formed veryquickly (within a single zone in the Middle Emsian). Asmentioned above, the same process in Nautilida contin-ued through the most part of the Devonian and in theCarboniferous. The development of complexly flutedsepta (complex sutures) is the main cluster inammonoid evolution. Many explanations (includingmathematical) of the functional significance of thecomplex suture have been proposed, but the seeminglymost convincing are those connecting the complexsepta and, hence, sutures by enforcing the shell to resisthydrostatic pressure. It is likely that a complex septumand part of the septum next to the shell wall, which isobserved as a sutural outline reinforced the shell. How-ever this could only have a functional sense if the cham-bers of the phragmocone were empty or were filled withgas at a pressure lower than the pressure of the sur-rounding water. If the chambers, as in the modern Nau-tilus, contained gas and fluid, the development of suchcomplex septa would be superfluous. The shell of Nau-
tilus with its simple septa and suture was shown exper-imentally to be able to resist up to 50 hPa, whichequates to a depth of 500 m. Ammonoids were unlikelyto have inhabited great depths. The increased complex-ity of septa cannot be convincingly explained by thenecessity of shell reinforcement only and therefore theevolutionary trend to increased complexity displayedby most ammonoids cannot be explained from thispoint of view either.
Barskov (1999) proposed a functional explanationof the appearance of fluting in ammonoid as a measureof buoyancy control (major problem for cephalopodswith an outer shell), which was in various ways solvedin the above mentioned groups, that makes it possibleto understand why this particular character becameimportant in the evolution of ammonoids (main clusterin Ruzhencev’s terminology). The posteriorad curva-tures of the septa (lobes in the suture) played the role ofreservoirs for liquid that controlled the shell buoyancy.
Ammonoid orders are distinguished based on thedifferences in the incipient curvatures of the septum atthe first, postembryonic planktonic stage, mainly in thenumber of lobes in the primary suture and pathways oftheir subsequent differentiation, in the shape of spheri-cal or spindle-like protoconch and the body chamber ofa single low whorl. The number of the primary lobesand mode of their subsequent subdivision apparentlyhad functional significance only at these first stages ofpostembryonic development.
The type of suture is determined based on the pri-mary suture and several subsequent septa. The hatchinglarva formed the second septum (shown on the shellsurface as a primary suture). The number and arrange-ment of the lobes in the primary suture mainly definedthe body plan of a mollusk. The original type of thesuture was formed within the five first septa. The earli-est ammonoids had a two-lobed suture inherited frombactrites (VO—ventral and omnilateral lobes). At thefollowing stage the dorsal lobe appears (VO : D). Fur-ther on the omnilateral lobe is replaced by the umbilicallobe, and the sutural formula becomes VU : D. The twofollowing elements to appear are the external laterallobe L (initial formula VLU : D) and internal laterallobe I (VU : ID, VLU : ID, and later VL : ID), whichappear in different orders almost simultaneously. Allsubsequent modifications of the suture develop on thebasis of these five main lobes, which determine the sep-aration of the subclass into orders.
According to this classification (Bogoslovskayaet al., 1990; Leonova, 2002; Shevyrev, 2006b; etc.),Anarcestida Miller et Furnish, 1954 (Devonian) is theearliest ammonoid order. Anarcestida gave rise to otherorders of Ammonoidea. The first Anarcestida appearedin the Early Devonian (Emsian) and rapidly reachedconsiderable diversity (37 genera are known from theEmsian only). In general, the diversity of Anarcestidawas very high and included 114 genera, from theEmsian to the Famennian, inclusive. The order included
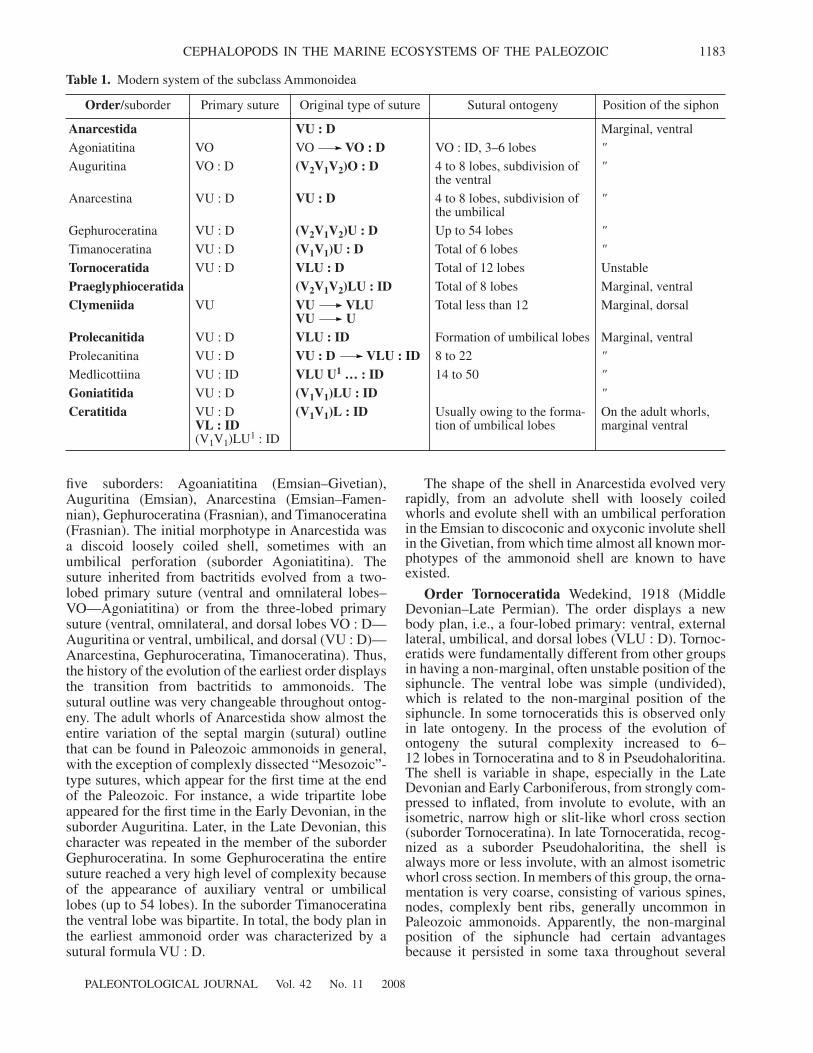
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1183
five suborders: Agoaniatitina (Emsian–Givetian),Auguritina (Emsian), Anarcestina (Emsian–Famen-nian), Gephuroceratina (Frasnian), and Timanoceratina(Frasnian). The initial morphotype in Anarcestida wasa discoid loosely coiled shell, sometimes with anumbilical perforation (suborder Agoniatitina). Thesuture inherited from bactritids evolved from a two-lobed primary suture (ventral and omnilateral lobes–VO—Agoniatitina) or from the three-lobed primarysuture (ventral, omnilateral, and dorsal lobes VO : D—Auguritina or ventral, umbilical, and dorsal (VU : D)—Anarcestina, Gephuroceratina, Timanoceratina). Thus,the history of the evolution of the earliest order displaysthe transition from bactritids to ammonoids. Thesutural outline was very changeable throughout ontog-eny. The adult whorls of Anarcestida show almost theentire variation of the septal margin (sutural) outlinethat can be found in Paleozoic ammonoids in general,with the exception of complexly dissected “Mesozoic”-type sutures, which appear for the first time at the endof the Paleozoic. For instance, a wide tripartite lobeappeared for the first time in the Early Devonian, in thesuborder Auguritina. Later, in the Late Devonian, thischaracter was repeated in the member of the suborderGephuroceratina. In some Gephuroceratina the entiresuture reached a very high level of complexity becauseof the appearance of auxiliary ventral or umbilicallobes (up to 54 lobes). In the suborder Timanoceratinathe ventral lobe was bipartite. In total, the body plan inthe earliest ammonoid order was characterized by asutural formula VU : D.
The shape of the shell in Anarcestida evolved veryrapidly, from an advolute shell with loosely coiledwhorls and evolute shell with an umbilical perforationin the Emsian to discoconic and oxyconic involute shellin the Givetian, from which time almost all known mor-photypes of the ammonoid shell are known to haveexisted.
Order Tornoceratida Wedekind, 1918 (MiddleDevonian–Late Permian). The order displays a newbody plan, i.e., a four-lobed primary: ventral, externallateral, umbilical, and dorsal lobes (VLU : D). Tornoc-eratids were fundamentally different from other groupsin having a non-marginal, often unstable position of thesiphuncle. The ventral lobe was simple (undivided),which is related to the non-marginal position of thesiphuncle. In some tornoceratids this is observed onlyin late ontogeny. In the process of the evolution ofontogeny the sutural complexity increased to 6–12 lobes in Tornoceratina and to 8 in Pseudohaloritina.The shell is variable in shape, especially in the LateDevonian and Early Carboniferous, from strongly com-pressed to inflated, from involute to evolute, with anisometric, narrow high or slit-like whorl cross section(suborder Tornoceratina). In late Tornoceratida, recog-nized as a suborder Pseudohaloritina, the shell isalways more or less involute, with an almost isometricwhorl cross section. In members of this group, the orna-mentation is very coarse, consisting of various spines,nodes, complexly bent ribs, generally uncommon inPaleozoic ammonoids. Apparently, the non-marginalposition of the siphuncle had certain advantagesbecause it persisted in some taxa throughout several
Table 1. Modern system of the subclass Ammonoidea
Order/suborder Primary suture Original type of suture Sutural ontogeny Position of the siphon
Anarcestida VU : D Marginal, ventral
Agoniatitina VO VO VO : D VO : ID, 3–6 lobes "
Auguritina VO : D (V2V1V2)O : D 4 to 8 lobes, subdivision of the ventral
"
Anarcestina VU : D VU : D 4 to 8 lobes, subdivision of the umbilical
"
Gephuroceratina VU : D (V2V1V2)U : D Up to 54 lobes "
Timanoceratina VU : D (V1V1)U : D Total of 6 lobes "
Tornoceratida VU : D VLU : D Total of 12 lobes Unstable
Praeglyphioceratida (V2V1V2)LU : ID Total of 8 lobes Marginal, ventral
Clymeniida VU VU VLUVU U
Total less than 12 Marginal, dorsal
Prolecanitida VU : D VLU : ID Formation of umbilical lobes Marginal, ventral
Prolecanitina VU : D VU : D VLU : ID 8 to 22 "
Medlicottiina VU : ID VLU U1 … : ID 14 to 50 "
Goniatitida VU : D (V1V1)LU : ID "
Ceratitida VU : DVL : ID(V1V1)LU1 : ID
(V1V1)L : ID Usually owing to the forma-tion of umbilical lobes
On the adult whorls,marginal ventral

1184
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
Paleozoic epochs. Boiko (2005) suggested that the sub-central position of the siphuncle could prolong theplanktonic stage in the development of the young mol-lusk thereby facilitating wider distribution in the basin.This phenomenon is also observed at the early ontogeneticstages of many Mesozoic ceratites and ammonites.
Tornoceratida in the Devonian and Carboniferousincluded 76 genera, in the Permian 14–16 genera,mainly of the suborder Pseudohaloritina.
Order Clymeniida Hyatt, 1884 (Late Devonian,Famennian)—the only ammonoid order with a dorsalposition of the siphuncle. This order existed only in thesecond half of the Famennian but reached an unprece-dented taxonomic and morphological diversity (70 gen-era). The suture developed along two pathways:VU VLU : D or VU U:D; the ventral lobe wasoften replaced by the ventral saddle. In some groups,the ventral saddle possesses an incipient ventral lobe,simple or bipartite. The dorsal lobe is simple, bipartite,
sometimes absent. The shell is variable in shape,mainly discoconic, from involute to evolute.
Boiko (2005) suggested that the dorsal position ofthe siphuncle precludes fast liquid removal from thephragmocone chambers, i.e., the dorsal siphuncle doesnot allow efficient buoyancy control. In most clymeni-ids the shells were compressed and evolute. Theseorganisms could only be planktonic and, given theirdorsal siphuncle, could hardly be adapted to verticalmigrations.
Order Praeglyphioceratida (Famennian–Tournai-sian). The main feature of organization is the presenceof a broad tripartite ventral lobe. Apart from that thereare also external lateral, umbilical, internal lateral, anddorsal lobes. The shell is involute and discoconic. Thisis a very small order (not more than 10 genera). Appar-ently, ammonoids of this group once again tried to solvethe problem of buoyancy control by developing auxil-
V1V2V2 L UU1 D1 D1
(a)
(b) (c)
Fig. 1.15. Sutural ontogeny in Prolecanitida: (a) Becanites africanus C1t (Korn et al., 2003, p. 1129); (b) Synartinskia principalisP1s (Leonova and Voronov, 1989, p. 115); (c) Epicanites loeblichi C1 (Spinosa et al., 1975, p. 259).

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1185
iary reservoirs on the ventral lobe. In general, this trenddid not achieve further progress.
Order Prolecanitida (Tournaisian–Scythian). Taxaassigned to this order (54 genera), throughout their his-tory, occupy a particular, distinct morphometric spaceand form a compact group with a well-defined trendtoward increased degree of whorl overlap and suturalcomplexity (Saunders and Work, 1997). For manyyears it was thought that the initial sutural type for allprolecanitids is trilobate, including a ventral lobe,umbilical lobe, and a dorsal lobe. Spinosa et al. (1975)showed that in the Early Carboniferous prolecanitidEpicanites the development of the lobe U is followedby the lobe L (VU : D VLU : ID) (Fig. 1.15). Ourstudy of a specimen of Epicanites, kindly donated byour American colleagues, supported their conclusion.Korn et al. (2003) studied the ontogeny of the Tournai-sian prolecanitid Becanites africanus and also con-cluded that the suture of the early prolecanitids andgoniatitids developed in a similar way: primary sutureVU : D third suture VLU : ID (Fig. 1.15). Inyounger members of the order (suborder Medlicottiina)the primary suture has an internal lateral lobe (fourth),as ontogenetic studies of the Permian prolecanitidgenus Synartinskia have shown (Leonova and Voronov,1989), the development follows the pathway VU :ID VLUU1U2..: ID (Fig. 1.15). Apparently, it istime to reconsider the well-established view on prole-canitids as on a group following the U-type ontogeny.
The main cluster of evolution of the most diversesuborder Medlicottiina was the progressively increas-ing complexity of the top of the external saddle by thedevelopment of many adventive lobes on the ventrolat-eral shoulder. The number of the external and internalinner lobes also increases in phylogeny.
Order Goniatitida (Tournaisian–Changhsingian)is the most taxonomically and morphologically diversegroup of Paleozoic ammonoids. The total number ofgenera in this order is 330. The initial type the sutureVLU : ID. The primary suture is VU : D. Members ofthe suborder Goniatitina, which included most Carbon-iferous and some Permian families, typically had“goniatitic” eight-lobed sutures composed of ventral,dorsal, and paired external and internal lateral andumbilical lobes. However, in some groups, e.g., in thePermian suborder Cyclolobina, the number of lobesreached 60, whereas the level of their dissection wascomparable to that of Mesozoic ammonoids. Goniatitesrepresented all size classes known in Paleozoicammonoids, from dwarves of less than 1 cm in diame-ter to giants, with a diameter of more than half a meter.Like prolecanitids, goniatitids had a marginal ventralsiphuncle. The shell shape varied from serpenticonicevolute (Rhymmoceras, Svetlanoceras) to oxyconic,completely evolute (Girtyoceras, Kazakhoceras) orspheroconic (Proshumardites, Neocrimites). The whorlsection varied from high and isometric to low and slit-like. Naturally, this diversity of the morphological
structures determined a wide range of adaptations.Goniatitids occupied all ecological niches in the LatePaleozoic basins and, unlike other Paleozoic orders(dominated by one or two life forms), their representa-tion in major types of life forms was reasonably uni-form, and throughout their evolutionary history, goniatit-ids inhabited the entire water column including bottomlayers, and near the surface. Morphometrically, they neveroverlapped the morphospace of prolecanitids.
Order Ceratitida (Roadian–Rhaetian). The firstceratitids appeared at the very end of the Paleozoic, atthe Lower–Middle Permian boundary. Their initialsutural formula is VL : ID, although in the most primi-tive taxa the primary suture could be trilobite, and inadvanced taxa it could be five-lobed. Despite the factthat ceratitids are related to prolecanitids in their origin,their sutural plans are different in the shape of the ven-tral lobes: in prolecanitids, the ventral lobe is tripartite,whereas in ceratitids it is bipartite. The earliest ceratit-ids had an evolute, medium-sized shell with a primitivesuture. The position of the siphuncle changed through-out ontogeny, and in many taxa the siphuncle was a sub-central at early stages. In adult ceratitids the siphuncleis always marginal, ventral. Taxa of higher rank (subor-ders) (total number 10, and two in the Permian) are dis-tinguished by the major types of the sutural ontogeny(Shevyrev, 2006b). In the systematics of ceratitids ofthe family- and genus-level, the suture does not have adecisive importance, as in the systematics of gonitatit-ids or prolecanitids. Shell ornamentation is consider-ably more widely used for Late Permian groups. Themost characteristic morphological feature of the Per-mian ceratitites is a complete absence of taxa with asubspheroconic and spheroconic shells, widespread ingoniatitids. Almost the entire diversity of Permian cer-atitids was represented by more or less evolute taxawith a low and moderately wide whorl cross section,with the exception of Araxoceratidae, the shell of whichhad an unusual shape with a flattened or keeled venterand strongly extended umbilical shoulders (Fig. 1.16);some of these taxa we assign to the benthopelagic lifeform. Ceratitids lived in the water column and judgingfrom the shell morphology were mainly inactive(planktonic life forms) or more active (nektobenthiclife forms).
We attempted to show that the functional explana-tion of differences in the body plans of the abovegroups may be used as a basis for recognition of sepa-rate orders. These groups, except bactritids andammonoids, control buoyancy similar to how it is doneby the modern Nautilus, i.e., by filling the phragmo-cone chambers with liquid. We could not find any fun-damental functional differences that could be used ascriteria supporting their assignment to different sub-classes. The original shell shape could be used as themost likely functional character for such an assign-ment: orthoconic or cyrtoconic, which indeed deter-mine fundamental differences in the orientation andstability control, but not in buoyancy control. Had this

1186
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
interpretation been accepted, it would have resulted inthe acceptance of the system proposed by Zhuravleva(1972): two subclasses Nautiloda and Orthoceroda,with ammonoids and coleoids assigned to Orthoceroda.This decision seems unacceptable because it does nottake into account fundamental differences in buoyancycontrol in ammonoids. Separation of Endocerida andActinocerida, and also Ellesmerocerida (Shevyrev,2005a) as subclasses does not have functional basis,more important than in other orders.
It is possible that further studies will allow the rec-ognition of fundamental differences in these groups(apart from just morphological), which will increasetheir taxonomic ranks. However, at present we do nothave such data. Therefore in this work we subdividegroups of Paleozoic cephalopods, without giving thema formal taxonomic rank: nonammonoid and ammonoid.This subdivision is quite well accepted in the descrip-tive work on post-Silurian cephalopods.
Ammonoids are taxonomically the more homoge-neous group, and it is unlikely that they can be regardedas a separate subclass.
ORDERS OF PALEOZOIC CEPHALOPODS
Nonammonoids1. Protactinocerida Chen et Qi, 1979–Cm2.Yanhecerida Chen et Qi, 1979–Cm3. Ellesmerocerida Flower, 1950 Cm–O34. Intejocerida Balashov, 1960 O15. Endocerida Teichert, 1933 O1–O36. Actinocerida Teichert, 1933 O1–C2
7. Orthocerida Kuhn, 1940 O1–T8. Pseudorthocerida Barskov, 1968 O1–T9. Tarphycerida Flower, 1950 O1–D310. Lituitida Starobogatov, 1983 O1–D211. Oncocerida Flower, 1950 O2–C112. Ascocerida Kuhn, 1940 O2–S13. Discosorida Flower, 1950 O2–D314. Barrandeocerida Flower, 1950 O2–D315. Nautilida Agassiz, 1847 D1–R16. Bactritida Shimansky, 1951 O1? D–TAmmonoidea17. Anarcestida Miller et Furnish, 1954 D18. Tornoceratida Wedekind, 1918 D–P319. Goniatitida Hyatt, 1884 C1–P320. Praeglyphioceratida Ruzhencev, 1957 D3–C121. Clymeniida Hyatt, 1884 D322. Prolecanitida Miller et Furnish, 1954 C1–T123. Ceratitida Hyatt, 1884 P2–T
CHAPTER 2. LIFE-FORMS OF CEPHALOPODS
2.1. Concept of Life-Forms
Alexander von Humboldt (1806) was the first tointroduce the concept of what we now call a “life-form.” He recognized 19 major forms of plants, whichshared “physiognomic” appearance. Although the ideaof “major forms” was widely used in botany as early asthe 19th century, the first formal definition of life-formsfor plants was formulated only a hundred years later byWarming (1908, p. 27), who wrote that a life-form is a
Fig. 1.16. Late Permian ceratitid Araxoceras with a rotoconic shell.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1187
form in which the vegetative body of an individualplant is in harmony with the environment throughout itsentire life, from seed to death. There are several sys-tems of life-forms used in botany, with a system pro-posed by Raunkiaer (1905) being the most popular. Inthis system the similarity of the external appearance ofthe plants united in one life-form is connected with theposition of new buds over the ground.
In zoology the idea of life-forms appeared later andwas formulated for the first time by Friederichs (1930)who wrote: “one and the same life-form includes thoseliving beings (species, generations, or stages of devel-opment) which live in similar environment and have asimilar lifestyle … a tadpole belongs to the same life-form as most fishes, whereas a frog belongs to another(cit. from Kashkarov, 1933, p. 123).
Attempts to explain morphological similarities as aresponse to similar requirements of the environmentresulted in the establishment of general systems of life-forms as hierarchical structures parallel to taxonomicsystems. Three such systems have been proposed(Gams, 1918; Friederichs, 1930; Aleev and Burdak,1984). Aleev (1986) used the term “ecomorph,” ratherthan the “life-form.” He argued the necessity of intro-ducing a new field of “ecomorphology” as one of thefundamental disciplines of general biology. Despite theapparent usefulness of building general systems of life-forms (ecomorphs) for discussion of general problemsof ontogeny, phylogeny and relationships between theorganism and environment, they have not been used tosolve problems of structure, initial stages or furtherevolution of taxa.
According to Gams (1918) the characterization of alife-form (although he did not formally define thisterm) should be based on “epharmonic” (adaptive, eco-logical) characters. His paper initiated a widespreadunderstanding of a life-form as a group of organisms,which, despite being taxonomically distant, have a sim-ilar shape which resulted from adaptation to the envi-ronment.
However, until now, a widely accepted definition of,or unified criteria for recognition of, life-forms have notbeen available in either botany or zoology. There are atleast five contradicting approaches to their recognitionand understanding (Aleev, 1986). For instance, there isan understanding that each taxon, each species, repre-sents a separate life-form (Severtsov, 1937). In contrast,Remane (1943) believed that each species cannot rep-resent only one life-form but is a member of differentsystems of life-forms. Kuhnelt (1970) suggested usingseven parallel systems of life-forms. From our point ofview, the differences in approach and methods of recog-nition and understanding of life-forms reflect the differ-ences in goals and tasks of different research. It is alsocertain that in different groups, the understanding andcriteria of recognition of life-forms may be different.
Below we cite several definitions of life-forms takenfrom the internet: (1) A life-form is a uniform biologi-
cal form characterized by common shape that is relatedto the development of this form and its inner structureof organs, the form appeared in certain ecological envi-ronment and reflecting the adaptations to this environ-ment (http://www.cladonia.ru/dict.html#); (2) a life-form is an ecological type of animals that includes rep-resentatives of different orders inhabiting similar envi-ronments and elaborating a similar lifestyle and a similarbody shape (http://www.glossary.ru/); (3) a life-form of aspecies is an external form reflecting a mode of interac-tions with the environment (http://shkola.lv/index.php);(4) a life-forms is group of individuals (of different spe-cies or within one species) with similar ecological andmorphological adaptations to living in a similar envi-ronment (http://enciclopaedia.ru/); (5) a life-form is ageneral ecological characterization of a species, genus,and any larger systematic category (http://www.aqua-worlds.com/aquaculture/). The Paleontologicheskii slo-var’ (Paleontological…, 1965, p. 117) defines it in thefollowing way: “a group of organisms recognizedbased on common features of their appearance, reflect-ing adaptations to a specific environment.”
Despite the differences, all the above definitionsshow a mutual interdependence of three groups of sim-ilarities on which the recognition of life-forms is based:(1) similar morphological appearance (shape); (2) sim-ilar lifestyle and consequently similar physiologicalcharacters; (3) living in a similar environment as thereason for the two above group of similarities. Differ-ences in definitions depend on which group of thesesimilarities is given priority. In the original definitionsof a life-form both in botany and zoology, priority isgiven to similarity in the lifestyle and biotope, and to alesser extent, to morphological similarity. Each taxonhas its own specific historically evolved system ofenvironmental adaptations. Organisms inhabiting thesame biotope and with a similar lifestyle are not nec-essarily morphologically similar. For instance, pelagicmicroplanktonic forms have different morphology (com-pare foraminifers, radiolarians, ostracodes, and dia-toms). The same applies to meso- and macroplanktonicorganisms. For instance, the morphology of pelagicpteropods that have a similar lifestyle may be quitediverse. Among modern cephalopods, planktonic octo-puses, squids, and spirulids are morphologically differ-ent. Also, similar morphology does not necessarilyindicate the same biotope, or a similar lifestyle. Forinstance, note the similarity of the shell in the pelagicJantina and benthic Natica and many other examples.Certainly, in all cases, morphological characters havecertain ecological adaptive significance. In the ecologi-cal sense, a life-form is a group of organisms recog-nized based on their affinity to certain ecotopes, and ingeneral to a particular space in the ecosphere.
Life-forms are recognized among the presently liv-ing organisms primarily based on direct observations oforganisms inhabiting certain environments, of their lif-estyle and liaising similarities in morphological expres-sion of adaptations with this ecotope and lifestyle. For

1188
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
fossil taxa, the determination of their affinity to a cer-tain adaptive zone and interpretation of their lifestyleare based on functional interpretation of morphology ofskeletal remains and also on their affinity to certainfacies and on taphonomic observations. Recognition oflife-forms within taxa of any rank is important and use-ful because firstly, it allows characterization of the eco-logical structure of a taxon and tracking of historicalchanges in the geological past, the understanding of theecological background of the phylogeny and thetaxon’s ecogenesis. Secondly, the evaluation of the tax-onomic and morphological diversity in a region orbasin allows identification of the morphological andtaxonomic “richness,” proportions and relative size ofadaptive zones in this region or basin. Thirdly, trackingchronological changes in the ecological structure oftaxonomic groups allows evaluation of changes in thespatial distribution and the size, structure, and propor-tions of ecotopes in the system of the Earth’s ecospherein the past.
Of numerous general definitions of the life-form, thedefinition proposed by Krivolutskii (1971) is the mostappropriate in this analysis of fossil cephalopods. Thelife-form is identified as an adaptive type formed“among representatives of a single taxonomic group(although of high rank), when characters of consider-able convergent similarity appear in different branchesof this group when these representatives inhabit a sim-ilar environment.” When talking about marine habitats“a similar ecological environment” is understood as thesame adaptive zone. The term “adaptive zone” identi-fies a certain space (pelagic or benthic zone) with itsphysical parameters in combination with a certain life-style in this space.
Hence, the first level of the hierarchy of life-forms istheir division into benthic (epifaunal, infaunal, psam-mon) and pelagic (nekton, plankton) groups. Furtherdivision may be based on the trophic type (suspensionfeeders, detrital feeders, scavengers, predators of vari-ous kind, etc.), on degree and type of mobility and anyother parameters, depending on the taxonomic groupand targets of study. In this paper, while studying life-forms of fossil cephalopods, we are naturally confinedto the two first hierarchic levels of life-forms.
Taxonomic genus is chosen as a major unit of life-forms, when analyzing ecological specialization ofcephalopod orders and its changes in the Paleozoicepochs, whereas the changes in the ecological structureof cephalopod communities inhabiting the UralianPaleobasin are studied at the species level.
2.2. Life-Forms of Cephalopods
Dollo (1912, 1922), a founder of ethological paleon-tology, was the first to recognize ecological groups offossil cephalopods corresponding to the term “life-forms” (Fig. 2.1). Dollo’s conclusions on ecology andlifestyle of fossil cephalopod groups are now only
interesting from a historical point of view, but his prin-ciple of ecological interpretation, in which genera areassigned to plankton, nekton, or benthos based on mor-phological analysis retains its full importance. Dollo’sideas correspond more to reality than a widespread mis-conception that is perpetuated in text-books and evensome scientific papers (e.g., Kröger, 2005) that allammonoids were active nektonic predators. The latteris not only incorrect and cannot be assumed taking intoaccount the design of the outer shell of cephalopodsfunctioning as a gas-liquid float, but simply contradictscommon sense.
The first systematization of extant cephalopods intoadaptive zones was proposed by Voos in 1967; he rec-ognized epi, meso-, bathy-, and abyssopelagic life-forms (cit. after Nesis, 1973). These groups were basedon the habitats (pelagic or benthic zones) and depth,i.e., immediately observed occurrence in certain adap-tive zones. Other ecological and morphological fea-tures were not analyzed. This approach is evidently notsuitable for fossil forms, for which affinity to a specificadaptive zone can be mainly based on the morphologi-cal and functional analysis.
Nesis (1973) proposed a detailed classification oflife-forms of extant cephalopods. Nesis’s classificationis based on the dichotomous hierarchical principle,with a taxonomic genus as a classification unit. At theuppermost hierarchical level all cephalopods aredivided into inhabitants of benthic zone, shelf, andpelagic zone. The lowermost level contains 23 genericecological groups. Apart from sharing the samebiotope, genera belonging to the same ecological groupshare similarities in the degree of mobility and motionmechanism, and feeding strategy. Morphological char-acters are included in the characterization of groups,but are not used as criteria of classification, althoughthis is possible. Nesis’s 23 ecological groups are notequally significant life-forms, and in the later publica-tions Nesis reduced their number to five (Nesis, 1975,1976) (benthic, benthopelagic, nektobenthic, nek-tonic, and planktonic).
General characteristics of five life-forms of cephalo-pods (after Nesis, 1976):
Benthic life-form. These animals are bottom-dwell-ing, mainly crawling, rarely using their hyponome. Sizefrom small to large (the latter is rare). Solitary. Scaven-gers. Eggs are large, development is direct, withoutpelagic stages. This group mostly includes octopuses.
Benthopelagic life-form. Bottom-dwelling animals,do not crawl on the bottom, but slowly move over thebottom using their hyponome for very short distances.Good at maneuvering. Scavengers and stealth preda-tors. Development is direct, without a pelagic stage.Deep-water octopuses. The only extant representativewith an outer shell is Nautilus.
Nektobenthic life-form. There are two types of thislife-form, the type represented by Sepia and the typerepresented by neritic squids.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1189
Sepia type. These are intermediate forms betweenbenthic and pelagic. They are closely connected withthe sea floor where they spend nights, ambush, and layeggs. Small and medium-sized. Feed on moving prey.Capable of darting forward. Ambush predators. Oftenlive in group. Eggs large, laid on the bottom.
Neritic squid type. Forms transitional to nektoniccephalopods. Live on the shelf. Connected with the bot-tom only to lay eggs. Fast, active predators, hunting ingroups. Eggs are small. The development has apelagic stage.
Nektonic life-form. Permanent inhabitants of thepelagic zone, not connected with the bottom.
Swim quickly using the hyponome. Active grouppredators. Eggs are small, in large numbers. The devel-opment includes a pelagic “larva” stage, which is verydifferent from the adult animal. In modern fauna, thisgroup is represented by oceanic squids.
Planktonic life-form. These are strictly pelagic ani-mals small, medium-sized, or sometimes large. Feed onplankton. Sometimes live in schools. Incapable of mov-ing for long periods using the hyponome. Eggs are usu-ally pelagic. Development is very prolonged. Larvae
are often completely different in appearance from theadults.
The above groups, excluding the nektonic form(actively swimming organisms capable of chasing theirprey), which cannot be reliably substantiated for ecto-cochliate cephalopods with a gaseous-fluid buoyancydevice, will be used in the discussion below. One andthe same life-form may be represented by animals withdifferent shell morphology (straight, curved, planispiralwith various combinations of parameters, or hetero-morphic), but with functional characteristics indicatinga certain life-form.
2.3. Ecological (Adaptive) Significance of Constructive Differences of the Outer Shell in Fossil Cephalopods,
Criteria and Methods of Their Assignment to Various Life-Forms
The characterization of life-forms of extant endoco-chliate cephalopods is based on direct observations oftheir habitats, breeding, and feeding strategies. Similar-ities of their morphology, including their body shape,morphology of their hyponome, head, and mantleappendages are recorded but are not recognized asdefinitive criteria. However, according to the above def-
Baculites
secondary planktonic
Ammonitessecondary benthic
Rhabdoceras
and primary nektonic
Ectocochlia
secondary planktonic
Ceratites benthic and
primary planktonic
Lituite
Bactritessecondary planktonic
Goniatitessecondary benthic and
primary planktonic
secondary planktonics
Nautilusessecondary benthic
Orthocerasprimary planktonic
Archimollusk primary benthic
Endocochlia
Opisthoteuthistertiary benthic
Pyrrhoteuthis secondary nektonic
Octopussecondary benthic
Doratopsidsecondary planktonic
Spirulasecondary planktonic
Decapodesprimary nektonic
Fig. 2.1. Major stages in ecogenesis of cephalopods (after Dollo, 1912, 1922).

1190
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
inition of the “life-form,” it is the convergent morpho-logical similarity that is definitive for the recognition oflife-forms and assignment of cephalopod genera to oneof these. The recognition of life-forms among fossilcephalopods is impossible by direct observation of theirlife and distribution in the adaptive zones of the sea. Inaddition, parameters of the adaptive zones of the seacould have been different in the geological past. There-fore the characteristics given below for life-forms offossil cephalopods are mainly based on functional anal-ysis of constructive parameters of the shell, mainly ofthose elements of the shell that have adaptive signifi-cance and are supplemented by some soft body fea-tures, which may be derived from the shell. The simi-larities to the morphology of extant cephalopods maybe of little help, because in the modern fauna, shelledcephalopods which are morphologically similar to fos-sil cephalopods and have a similar hydrostatic mecha-nism are represented by approximately ten genera ofsepiids and two genera of nautilids. Note that threemajor morphological types of modern shelled cephalo-pods (a few genera of Sepiida; Spirula; and Nautilusand Allonautilus belong to different life-forms: nekto-benthic, planktonic, and benthopelagic, respectively.Of these, only the nautilids have an external shell.
The recognition of life-forms when dealing withfossils is based on the shell and siphuncle morphology,which determine the main hydrostatic and hydrody-namic qualities of the animal allowing their assignmentto a particular life-form. This is supplemented by somegeneral biological assumptions, knowledge of the typeof individual development and taphonomy, throughwhich a possible environment may be suggested.
The main hydrodynamic and hydrostatic character-istics, from which the constructive features of the outershell may be inferred, are listed below.
(1) Degree of buoyancy. The degree of buoyancy,i.e., the size of the gas-filled phragmocone in compari-son to the size (weight) of the soft body may be esti-mated quantitatively based on the shell expansion rate.In straight and curved cephalopods, the degree of buoy-ancy depends on the function of the angle of the conicalshell, whereas in the coiled cephalopods this parameterdepends on the whorl expansion rate. The smallerexpansion angle and lesser whorl expansion rate sug-gest greater buoyancy, which in most cases has to becompensated for by chambers filled with fluid.
(2) Ways to support orientation in the water are oftenexpressed in the shell shape, in the presence or absenceof specialized mechanisms of orientation (cameral andendosiphuncular deposits, in the specific arrangementof septa, and in the ornamentation). For instance, thepresence in straight shelled taxa of a ventral siphunclewith deposits suggest that they lived in a horizontalposition (horizontal longitudinal axis) and, hence, wererelatively actively swimming. Sinusoid septa posi-tioned on the dorsal side of the shell, as in some asco-cerids and modern sepiids, also suggest a horizontal
position of the body. A tightly coiled shell, in which thegeneral center of gravity coincides with the center ofbuoyancy, is in indifferent balance with the water anddoes not need additional mechanisms of orientation.Extensive ornamentation (long spines, collars, or longi-tudinal ribs) is interpreted as a means of support of theshell orientation.
(3) Active swimming is possible because of thedevelopment of the propulsive mechanism, whichincludes a variously sized and shaped mantle cavity anda hyponome. Swimming ability can be evaluated basedon the proportions of the body chamber and on thedegree of the hyponomic sinus on the shell. Forinstance, it is evident that shells with a very long, orvery short, or strongly flattened body chamber couldnot have had a mantle cavity that would allow a power-ful propulsive thrust and could not therefore be efficientswimmers or predators.
(4) Streamlining of the shell may be one of the addi-tional characteristics supplemented to the ability toswim and depends both on the shell shape and orna-mentation.
Different life-forms have hydrostatic and hydrody-namic qualities variously developed and combined,resulting from different shell morphology.
Several fundamentally different types of shellgeometry may be recognized among ectocochliatecephalopods, resulting in different shell hydrostaticsand hydrodynamics.
Major morphological types of cephalopod shell:(1) Straight (orthoceraconic);(2) Curved (cyrtoceraconic);(2a) Shells endogastrically curved (ventral side, on
which the hyponome is positioned, is concave, whilethe dorsal side is convex);
(2b) Shells exogastrically curved (ventral side isconvex, while the dorsal side is concave);
(3) Coiled in one plane (planispiral) with non-con-tacting whorls (gyroceraconic), with contacting whorls(nautilicones), among which, depending on the whorlexpansion rate, degree of whorl overlap and the cross-sectional shape, special names are used (see below);
(4) Spirally coiled (trochoid), with possibly non-touching whorls, or in contact with varying degrees ofoverlap;
(5) Heteromorphic shells, changing their shape inontogeny; for instance, with a trochoid coiling replacedby planispiral coiling, tightly coiled whorls replaced byloosely coiled whorls, a planispiral shell replaced bycyrtospiral or orthoceraconic.
Depending on geometric parameters, within virtu-ally each of the above shell types, the hydrostatic andhydrodynamic properties may be different, and there-fore, the animals could belong to different life-forms.Below an attempt is made to assign cephalopods with astraight and curved shell, with a coiled planispiral, tro-choid, or heteromorphic shell to various life-forms.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1191
2.4. Life-Forms of Cephalopods with a Curved Shell
Two types of shell may be recognized according tothe way they are curved. In exogastric shells the ventralside of the shell is convex, and the dorsal side is con-cave. In endogastric shells, the ventral side of the shellis concave, and the dorsal is convex. The endogastricshell is a more primitive morphological type of theexternal cephalopod shell.
Endogastric and exogastric shells have differentpotentials for ecological adaptations. In the absence ofspecialized mechanisms for orientation and stabilitycontrol, in a floating endogastric shell, as it grows, thecenter of buoyancy shifts apically. In such cases, theanimal was orientated with its concave ventral(hyponomic) side facing up, and the dorsal side and
head facing down (Fig. 2.2). This situation facilitatedscavenging on the bottom using head tentacles, but didnot help active swimming. Thus, such shell construc-tion allowed only slow passive floating above the bot-tom. This allows the placement of ammonoids withsuch shells within the benthopelagic life-forms.
The construction of the exogastric shells allowsbroader opportunities for ecological adaptations. As theshell grew, the shell orientation changed to raise thedorsal side of the aperture upwards, whereas the ventral(hyponomic) side became horizontal (Fig. 2.2.). Thisorientation is fundamentally different from the onedescribed previously, because it facilitates the use ofthe hyponome for swimming. An animal with an exo-gastric shell is capable of more active swimming over
(a) (c)
(d)(b)
1 2
Fig. 2.2. Hypothetical orientation of exogastric (a, b) and endogastric (c, d) shells throughout growth; (a, c) juvenile shells; (b, d) adultshells (1) position of the center of buoyancy; (2) position of the center of gravity.
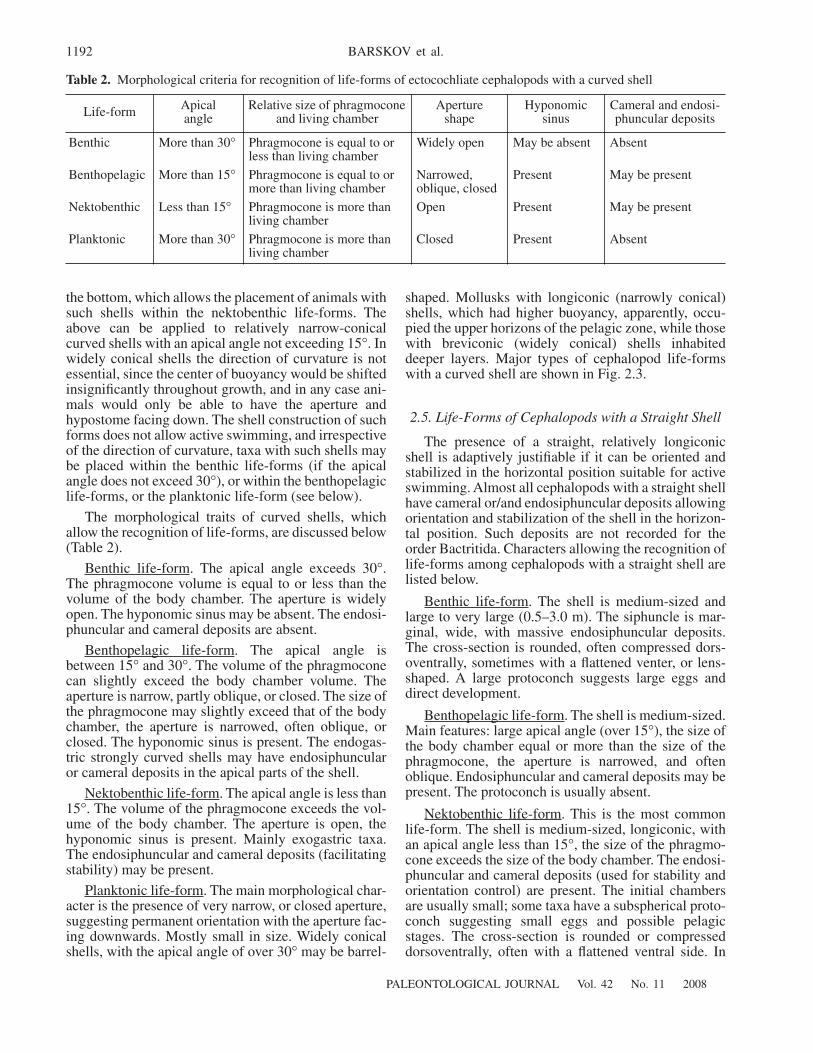
1192
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
the bottom, which allows the placement of animals withsuch shells within the nektobenthic life-forms. Theabove can be applied to relatively narrow-conicalcurved shells with an apical angle not exceeding 15°. Inwidely conical shells the direction of curvature is notessential, since the center of buoyancy would be shiftedinsignificantly throughout growth, and in any case ani-mals would only be able to have the aperture andhypostome facing down. The shell construction of suchforms does not allow active swimming, and irrespectiveof the direction of curvature, taxa with such shells maybe placed within the benthic life-forms (if the apicalangle does not exceed 30°), or within the benthopelagiclife-forms, or the planktonic life-form (see below).
The morphological traits of curved shells, whichallow the recognition of life-forms, are discussed below(Table 2).
Benthic life-form. The apical angle exceeds 30°.The phragmocone volume is equal to or less than thevolume of the body chamber. The aperture is widelyopen. The hyponomic sinus may be absent. The endosi-phuncular and cameral deposits are absent.
Benthopelagic life-form. The apical angle isbetween 15° and 30°. The volume of the phragmoconecan slightly exceed the body chamber volume. Theaperture is narrow, partly oblique, or closed. The size ofthe phragmocone may slightly exceed that of the bodychamber, the aperture is narrowed, often oblique, orclosed. The hyponomic sinus is present. The endogas-tric strongly curved shells may have endosiphuncularor cameral deposits in the apical parts of the shell.
Nektobenthic life-form. The apical angle is less than15°. The volume of the phragmocone exceeds the vol-ume of the body chamber. The aperture is open, thehyponomic sinus is present. Mainly exogastric taxa.The endosiphuncular and cameral deposits (facilitatingstability) may be present.
Planktonic life-form. The main morphological char-acter is the presence of very narrow, or closed aperture,suggesting permanent orientation with the aperture fac-ing downwards. Mostly small in size. Widely conicalshells, with the apical angle of over 30° may be barrel-
shaped. Mollusks with longiconic (narrowly conical)shells, which had higher buoyancy, apparently, occu-pied the upper horizons of the pelagic zone, while thosewith breviconic (widely conical) shells inhabiteddeeper layers. Major types of cephalopod life-formswith a curved shell are shown in Fig. 2.3.
2.5. Life-Forms of Cephalopods with a Straight Shell
The presence of a straight, relatively longiconicshell is adaptively justifiable if it can be oriented andstabilized in the horizontal position suitable for activeswimming. Almost all cephalopods with a straight shellhave cameral or/and endosiphuncular deposits allowingorientation and stabilization of the shell in the horizon-tal position. Such deposits are not recorded for theorder Bactritida. Characters allowing the recognition oflife-forms among cephalopods with a straight shell arelisted below.
Benthic life-form. The shell is medium-sized andlarge to very large (0.5–3.0 m). The siphuncle is mar-ginal, wide, with massive endosiphuncular deposits.The cross-section is rounded, often compressed dors-oventrally, sometimes with a flattened venter, or lens-shaped. A large protoconch suggests large eggs anddirect development.
Benthopelagic life-form. The shell is medium-sized.Main features: large apical angle (over 15°), the size ofthe body chamber equal or more than the size of thephragmocone, the aperture is narrowed, and oftenoblique. Endosiphuncular and cameral deposits may bepresent. The protoconch is usually absent.
Nektobenthic life-form. This is the most commonlife-form. The shell is medium-sized, longiconic, withan apical angle less than 15°, the size of the phragmo-cone exceeds the size of the body chamber. The endosi-phuncular and cameral deposits (used for stability andorientation control) are present. The initial chambersare usually small; some taxa have a subspherical proto-conch suggesting small eggs and possible pelagicstages. The cross-section is rounded or compresseddorsoventrally, often with a flattened ventral side. In
Table 2. Morphological criteria for recognition of life-forms of ectocochliate cephalopods with a curved shell
Life-form Apicalangle
Relative size of phragmocone and living chamber
Apertureshape
Hyponomic sinus
Cameral and endosi-phuncular deposits
Benthic More than 30° Phragmocone is equal to or less than living chamber
Widely open May be absent Absent
Benthopelagic More than 15° Phragmocone is equal to or more than living chamber
Narrowed, oblique, closed
Present May be present
Nektobenthic Less than 15° Phragmocone is more than living chamber
Open Present May be present
Planktonic More than 30° Phragmocone is more than living chamber
Closed Present Absent
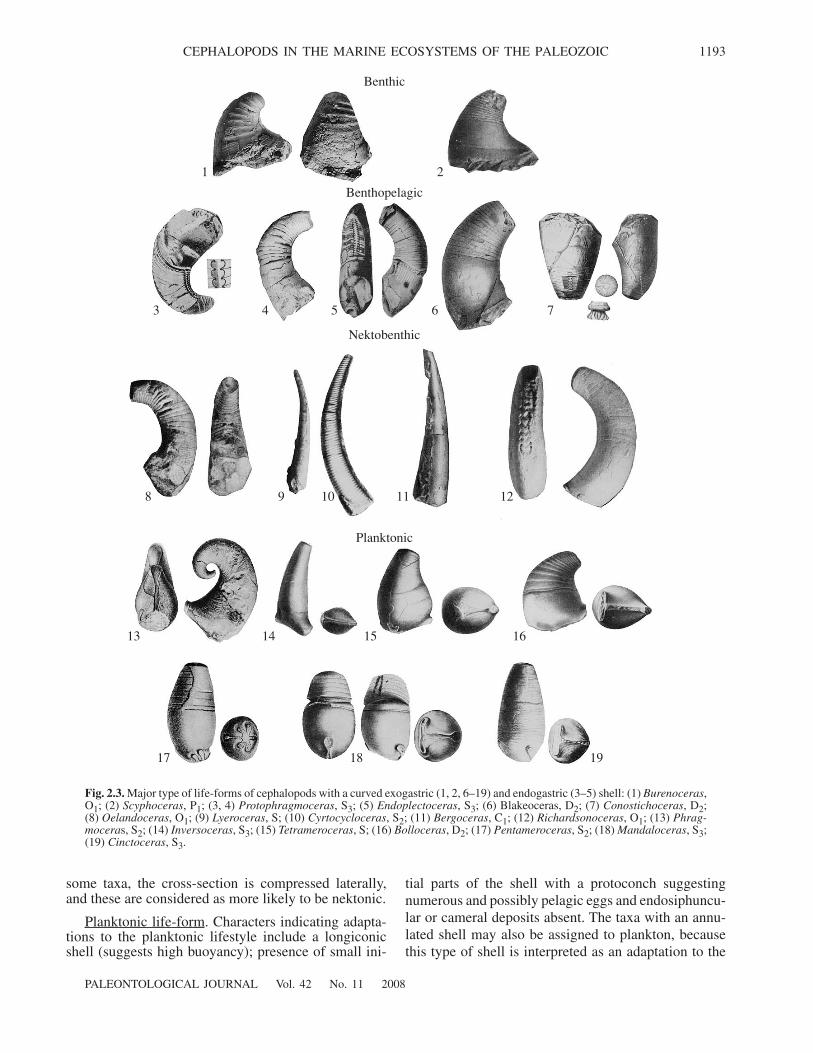
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1193
some taxa, the cross-section is compressed laterally,and these are considered as more likely to be nektonic.
Planktonic life-form. Characters indicating adapta-tions to the planktonic lifestyle include a longiconicshell (suggests high buoyancy); presence of small ini-
tial parts of the shell with a protoconch suggestingnumerous and possibly pelagic eggs and endosiphuncu-lar or cameral deposits absent. The taxa with an annu-lated shell may also be assigned to plankton, becausethis type of shell is interpreted as an adaptation to the
1 2
3 4 5 6 7
8 9 10 11 12
13 14 15 16
17 18 19
Benthic
Benthopelagic
Nektobenthic
Planktonic
Fig. 2.3. Major type of life-forms of cephalopods with a curved exogastric (1, 2, 6–19) and endogastric (3–5) shell: (1) Burenoceras,O1; (2) Scyphoceras, P1; (3, 4) Protophragmoceras, S3; (5) Endoplectoceras, S3; (6) Blakeoceras, D2; (7) Conostichoceras, D2;(8) Oelandoceras, O1; (9) Lyeroceras, S; (10) Cyrtocycloceras, S2; (11) Bergoceras, C1; (12) Richardsonoceras, O1; (13) Phrag-moceras, S2; (14) Inversoceras, S3; (15) Tetrameroceras, S; (16) Bolloceras, D2; (17) Pentameroceras, S2; (18) Mandaloceras, S3;(19) Cinctoceras, S3.
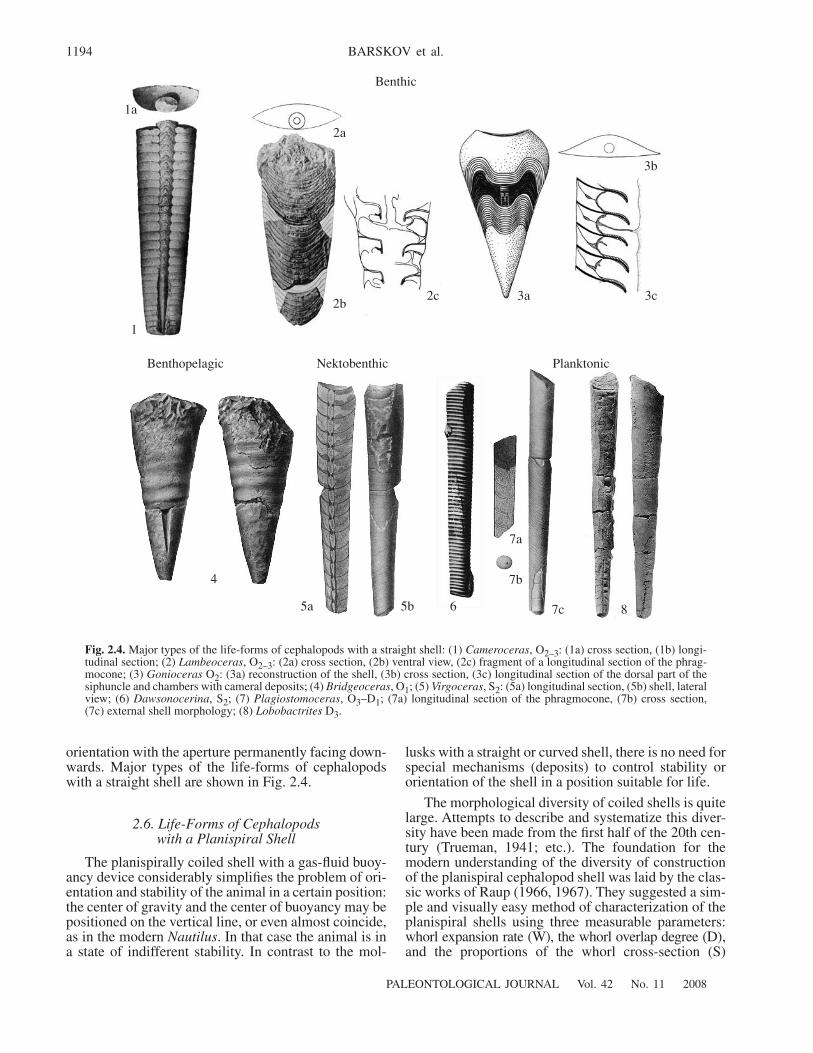
1194
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
orientation with the aperture permanently facing down-wards. Major types of the life-forms of cephalopodswith a straight shell are shown in Fig. 2.4.
2.6. Life-Forms of Cephalopods with a Planispiral Shell
The planispirally coiled shell with a gas-fluid buoy-ancy device considerably simplifies the problem of ori-entation and stability of the animal in a certain position:the center of gravity and the center of buoyancy may bepositioned on the vertical line, or even almost coincide,as in the modern Nautilus. In that case the animal is ina state of indifferent stability. In contrast to the mol-
lusks with a straight or curved shell, there is no need forspecial mechanisms (deposits) to control stability ororientation of the shell in a position suitable for life.
The morphological diversity of coiled shells is quitelarge. Attempts to describe and systematize this diver-sity have been made from the first half of the 20th cen-tury (Trueman, 1941; etc.). The foundation for themodern understanding of the diversity of constructionof the planispiral cephalopod shell was laid by the clas-sic works of Raup (1966, 1967). They suggested a sim-ple and visually easy method of characterization of theplanispiral shells using three measurable parameters:whorl expansion rate (W), the whorl overlap degree (D),and the proportions of the whorl cross-section (S)
Benthic
1‡
1
2b2c 3‡ 3c
3b
2‡
Benthopelagic Nektobenthic Planktonic
4
5‡ 5b 6
7‡
7b
7c 8
Fig. 2.4. Major types of the life-forms of cephalopods with a straight shell: (1) Cameroceras, O2–3: (1a) cross section, (1b) longi-tudinal section; (2) Lambeoceras, O2–3: (2a) cross section, (2b) ventral view, (2c) fragment of a longitudinal section of the phrag-mocone; (3) Gonioceras O2: (3a) reconstruction of the shell, (3b) cross section, (3c) longitudinal section of the dorsal part of thesiphuncle and chambers with cameral deposits; (4) Bridgeoceras, O1; (5) Virgoceras, S2: (5a) longitudinal section, (5b) shell, lateralview; (6) Dawsonocerina, S2; (7) Plagiostomoceras, O3–D1; (7a) longitudinal section of the phragmocone, (7b) cross section,(7c) external shell morphology; (8) Lobobactrites D3.
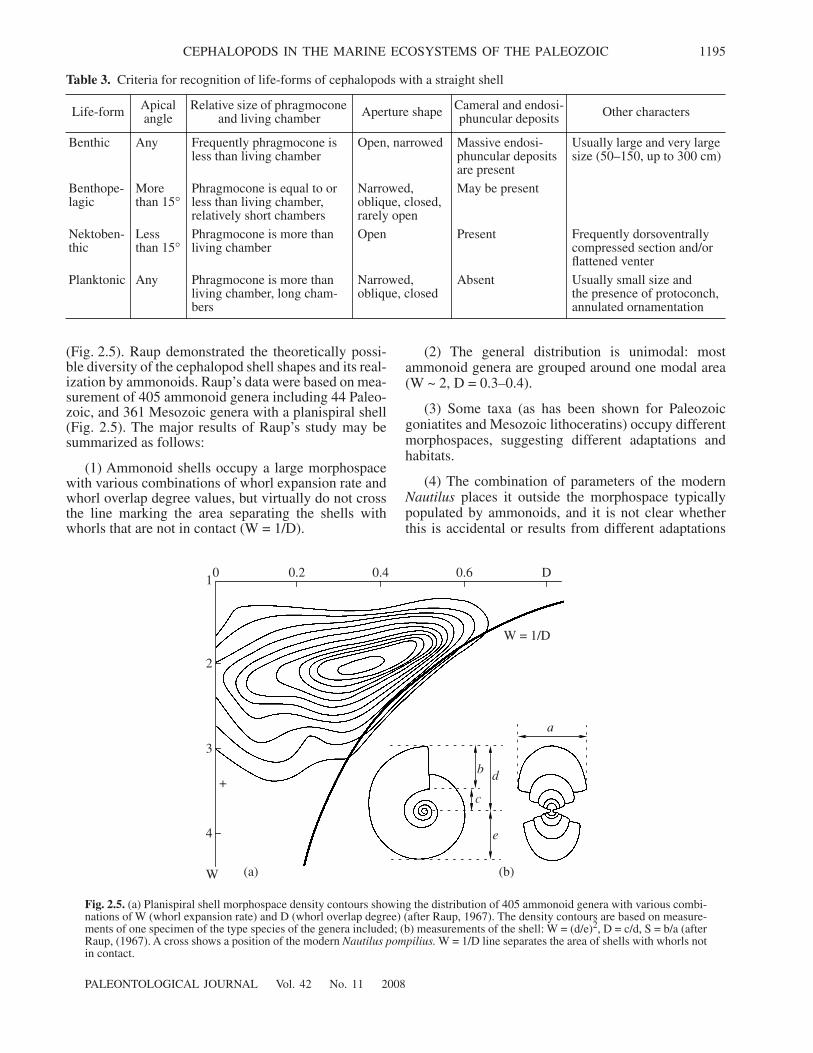
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1195
(Fig. 2.5). Raup demonstrated the theoretically possi-ble diversity of the cephalopod shell shapes and its real-ization by ammonoids. Raup’s data were based on mea-surement of 405 ammonoid genera including 44 Paleo-zoic, and 361 Mesozoic genera with a planispiral shell(Fig. 2.5). The major results of Raup’s study may besummarized as follows:
(1) Ammonoid shells occupy a large morphospacewith various combinations of whorl expansion rate andwhorl overlap degree values, but virtually do not crossthe line marking the area separating the shells withwhorls that are not in contact (W = 1/D).
(2) The general distribution is unimodal: mostammonoid genera are grouped around one modal area(W ~ 2, D = 0.3–0.4).
(3) Some taxa (as has been shown for Paleozoicgoniatites and Mesozoic lithoceratins) occupy differentmorphospaces, suggesting different adaptations andhabitats.
(4) The combination of parameters of the modernNautilus places it outside the morphospace typicallypopulated by ammonoids, and it is not clear whetherthis is accidental or results from different adaptations
Table 3. Criteria for recognition of life-forms of cephalopods with a straight shell
Life-form Apicalangle
Relative size of phragmocone and living chamber Aperture shape Cameral and endosi-
phuncular deposits Other characters
Benthic Any Frequently phragmocone is less than living chamber
Open, narrowed Massive endosi-phuncular deposits are present
Usually large and very large size (50–150, up to 300 cm)
Benthope-lagic
More than 15°
Phragmocone is equal to or less than living chamber, relatively short chambers
Narrowed, oblique, closed, rarely open
May be present
Nektoben-thic
Lessthan 15°
Phragmocone is more than living chamber
Open Present Frequently dorsoventrally compressed section and/or flattened venter
Planktonic Any Phragmocone is more than living chamber, long cham-bers
Narrowed, oblique, closed
Absent Usually small size andthe presence of protoconch, annulated ornamentation
2
0.20 0.4 0.6 D1
3
4
W
+
a
b
c
d
e
Fig. 2.5. (a) Planispiral shell morphospace density contours showing the distribution of 405 ammonoid genera with various combi-nations of W (whorl expansion rate) and D (whorl overlap degree) (after Raup, 1967). The density contours are based on measure-ments of one specimen of the type species of the genera included; (b) measurements of the shell: W = (d/e)2, D = c/d, S = b/a (afterRaup, (1967). A cross shows a position of the modern Nautilus pompilius. W = 1/D line separates the area of shells with whorls notin contact.
(a) (b)
W = 1/D
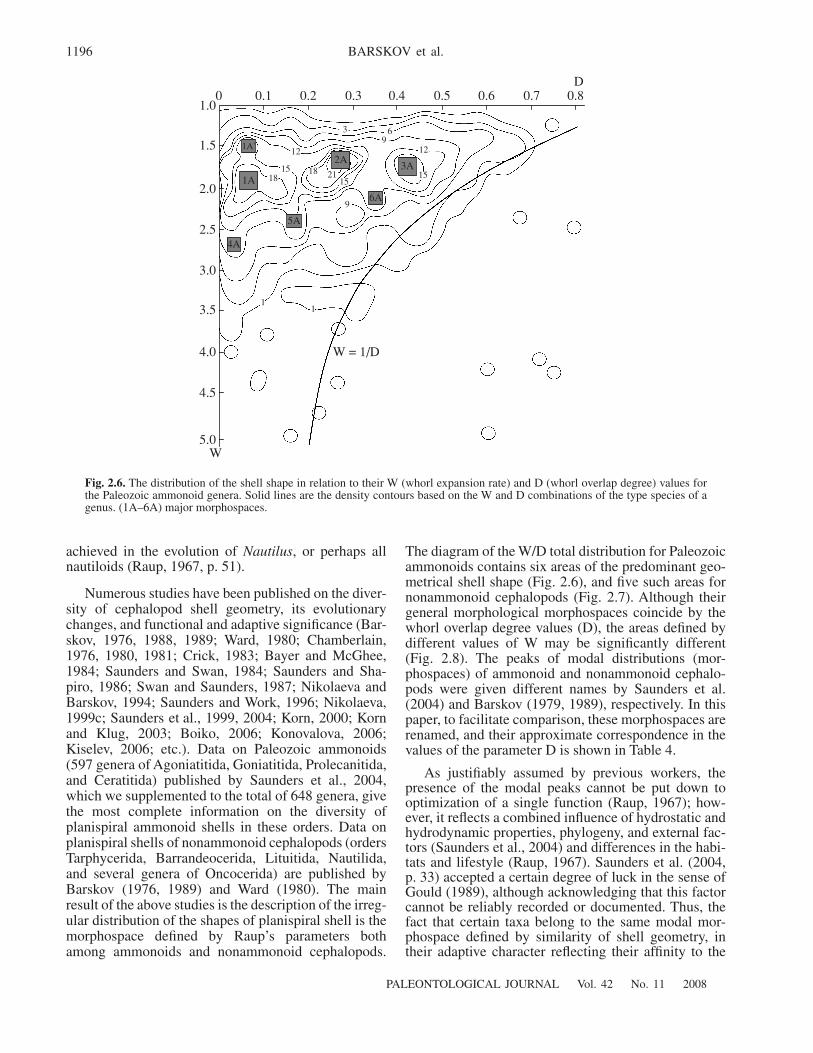
1196
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
achieved in the evolution of Nautilus, or perhaps allnautiloids (Raup, 1967, p. 51).
Numerous studies have been published on the diver-sity of cephalopod shell geometry, its evolutionarychanges, and functional and adaptive significance (Bar-skov, 1976, 1988, 1989; Ward, 1980; Chamberlain,1976, 1980, 1981; Crick, 1983; Bayer and McGhee,1984; Saunders and Swan, 1984; Saunders and Sha-piro, 1986; Swan and Saunders, 1987; Nikolaeva andBarskov, 1994; Saunders and Work, 1996; Nikolaeva,1999c; Saunders et al., 1999, 2004; Korn, 2000; Kornand Klug, 2003; Boiko, 2006; Konovalova, 2006;Kiselev, 2006; etc.). Data on Paleozoic ammonoids(597 genera of Agoniatitida, Goniatitida, Prolecanitida,and Ceratitida) published by Saunders et al., 2004,which we supplemented to the total of 648 genera, givethe most complete information on the diversity ofplanispiral ammonoid shells in these orders. Data onplanispiral shells of nonammonoid cephalopods (ordersTarphycerida, Barrandeocerida, Lituitida, Nautilida,and several genera of Oncocerida) are published byBarskov (1976, 1989) and Ward (1980). The mainresult of the above studies is the description of the irreg-ular distribution of the shapes of planispiral shell is themorphospace defined by Raup’s parameters bothamong ammonoids and nonammonoid cephalopods.
The diagram of the W/D total distribution for Paleozoicammonoids contains six areas of the predominant geo-metrical shell shape (Fig. 2.6), and five such areas fornonammonoid cephalopods (Fig. 2.7). Although theirgeneral morphological morphospaces coincide by thewhorl overlap degree values (D), the areas defined bydifferent values of W may be significantly different(Fig. 2.8). The peaks of modal distributions (mor-phospaces) of ammonoid and nonammonoid cephalo-pods were given different names by Saunders et al.(2004) and Barskov (1979, 1989), respectively. In thispaper, to facilitate comparison, these morphospaces arerenamed, and their approximate correspondence in thevalues of the parameter D is shown in Table 4.
As justifiably assumed by previous workers, thepresence of the modal peaks cannot be put down tooptimization of a single function (Raup, 1967); how-ever, it reflects a combined influence of hydrostatic andhydrodynamic properties, phylogeny, and external fac-tors (Saunders et al., 2004) and differences in the habi-tats and lifestyle (Raup, 1967). Saunders et al. (2004,p. 33) accepted a certain degree of luck in the sense ofGould (1989), although acknowledging that this factorcannot be reliably recorded or documented. Thus, thefact that certain taxa belong to the same modal mor-phospace defined by similarity of shell geometry, intheir adaptive character reflecting their affinity to the
1.5
0.10 0.2 0.3 0.4 0.5 0.6 0.7 0.8D
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
1.0
3
12
1518
18 2115
9
15
129
6
1A'
1A
4A
5A
6A
3A2A
W = 1/D
11
Fig. 2.6. The distribution of the shell shape in relation to their W (whorl expansion rate) and D (whorl overlap degree) values forthe Paleozoic ammonoid genera. Solid lines are the density contours based on the W and D combinations of the type species of agenus. (1A–6A) major morphospaces.
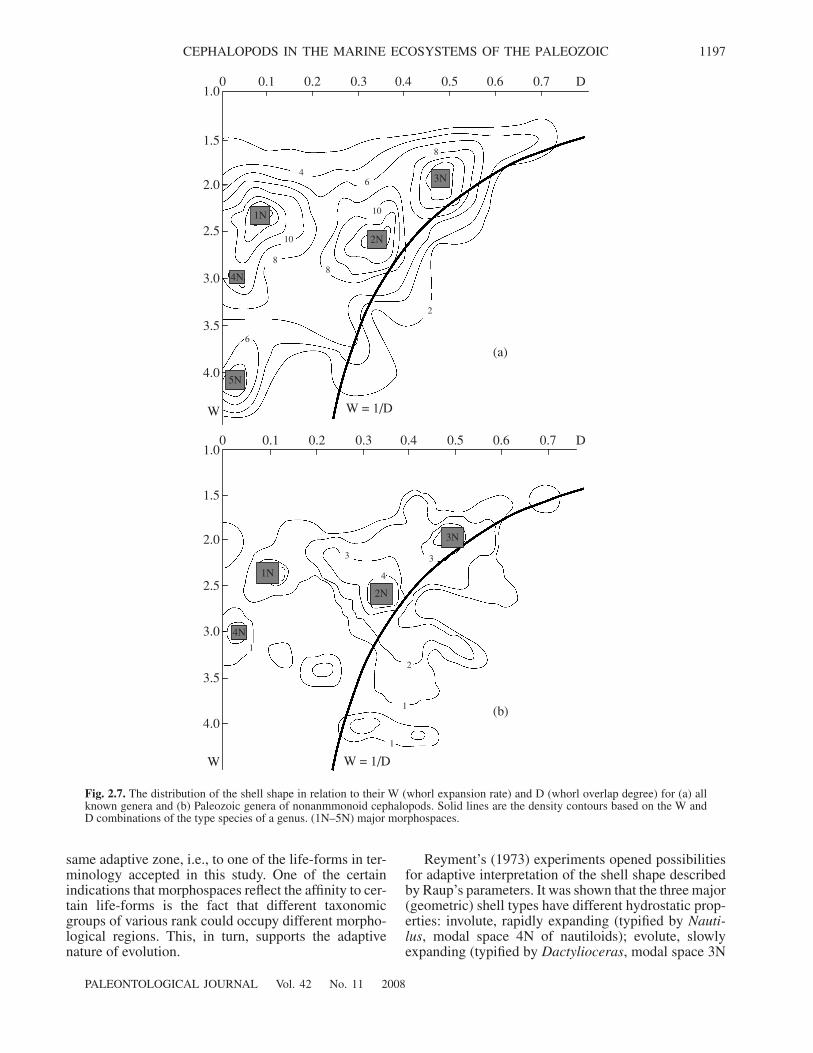
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1197
same adaptive zone, i.e., to one of the life-forms in ter-minology accepted in this study. One of the certainindications that morphospaces reflect the affinity to cer-tain life-forms is the fact that different taxonomicgroups of various rank could occupy different morpho-logical regions. This, in turn, supports the adaptivenature of evolution.
Reyment’s (1973) experiments opened possibilitiesfor adaptive interpretation of the shell shape describedby Raup’s parameters. It was shown that the three major(geometric) shell types have different hydrostatic prop-erties: involute, rapidly expanding (typified by Nauti-lus, modal space 4N of nautiloids); evolute, slowlyexpanding (typified by Dactylioceras, modal space 3N
1.5
0.10 0.2 0.3 0.4 0.5 0.6 0.7 D
2.0
2.5
3.0
3.5
4.0
W
1.0
43N
W = 1/D
2N
1N
4N
5N
10
8
6
10
88
6
2
1.5
0.10 0.2 0.3 0.4 0.5 0.6 0.7 D
2.0
2.5
3.0
3.5
4.0
W
1.0
3N
W = 1/D
2N
1N
4N
1
1
1
3 3
4
2
Fig. 2.7. The distribution of the shell shape in relation to their W (whorl expansion rate) and D (whorl overlap degree) for (a) allknown genera and (b) Paleozoic genera of nonanmmonoid cephalopods. Solid lines are the density contours based on the W andD combinations of the type species of a genus. (1N–5N) major morphospaces.
(a)
(b)
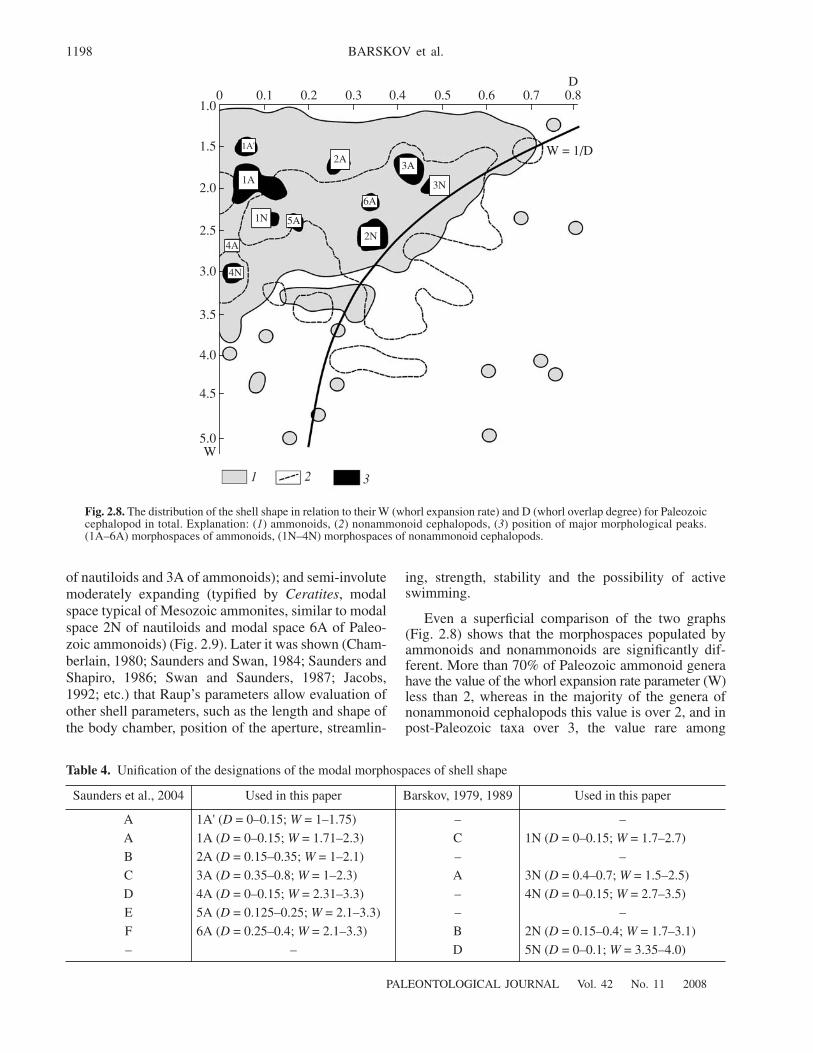
1198
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
of nautiloids and 3A of ammonoids); and semi-involutemoderately expanding (typified by Ceratites, modalspace typical of Mesozoic ammonites, similar to modalspace 2N of nautiloids and modal space 6A of Paleo-zoic ammonoids) (Fig. 2.9). Later it was shown (Cham-berlain, 1980; Saunders and Swan, 1984; Saunders andShapiro, 1986; Swan and Saunders, 1987; Jacobs,1992; etc.) that Raup’s parameters allow evaluation ofother shell parameters, such as the length and shape ofthe body chamber, position of the aperture, streamlin-
ing, strength, stability and the possibility of activeswimming.
Even a superficial comparison of the two graphs(Fig. 2.8) shows that the morphospaces populated byammonoids and nonammonoids are significantly dif-ferent. More than 70% of Paleozoic ammonoid generahave the value of the whorl expansion rate parameter (W)less than 2, whereas in the majority of the genera ofnonammonoid cephalopods this value is over 2, and inpost-Paleozoic taxa over 3, the value rare among
1.5
0.10 0.2 0.3 0.4 0.5 0.6 0.7 0.8D
1.0
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
3A
3N
2A
2N
1N
1A
1A'
5A
6A
4A
4N
W = 1/D
1 2 3
Fig. 2.8. The distribution of the shell shape in relation to their W (whorl expansion rate) and D (whorl overlap degree) for Paleozoiccephalopod in total. Explanation: (1) ammonoids, (2) nonammonoid cephalopods, (3) position of major morphological peaks.(1A–6A) morphospaces of ammonoids, (1N–4N) morphospaces of nonammonoid cephalopods.
Table 4. Unification of the designations of the modal morphospaces of shell shape
Saunders et al., 2004 Used in this paper Barskov, 1979, 1989 Used in this paper
A 1A' (D = 0–0.15; W = 1–1.75) – –
A 1A (D = 0–0.15; W = 1.71–2.3) C 1N (D = 0–0.15; W = 1.7–2.7)
B 2A (D = 0.15–0.35; W = 1–2.1) – –
C 3A (D = 0.35–0.8; W = 1–2.3) A 3N (D = 0.4–0.7; W = 1.5–2.5)
D 4A (D = 0–0.15; W = 2.31–3.3) – 4N (D = 0–0.15; W = 2.7–3.5)
E 5A (D = 0.125–0.25; W = 2.1–3.3) – –
F 6A (D = 0.25–0.4; W = 2.1–3.3) B 2N (D = 0.15–0.4; W = 1.7–3.1)
– – D 5N (D = 0–0.1; W = 3.35–4.0)

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1199
ammonoids. Because the expansion rate is functionallyconnected with the shell hydrostatics (Reyment, 1973;Saunders and Shapiro, 1986), these differences, in ouropinion, may be related to differences in the physiologyof these two groups mainly related to buoyancy control(Barskov, 1999).
Below we evaluate the adaptive significance of thefunctional possibilities of the shells from different mor-phospaces among ammonoids and nonammonoids. Thesequence of the evaluation is as follows: first we discuss
shells from morphospaces 3N, 4N, and 2N, becauseforms typifying these morphospaces were studied byReyment. These are followed by shells from region 1N.The sequence of consideration of the types of theammonoid shells follows the sequence of the mor-phospaces numbers: 1A to 6A.
Nonammonoid cephalopods. Note that for Paleo-zoic nonammonoid cephalopods morphospaces 1N and4N were represented by only a few genera, whereas forthe Mesozoic nautilid genera, including Triassic, thesemorphospaces became exclusive.
Morphospace 3N (Fig. 2.10). These are evoluteshells (D = 0.4–0.7) with a low whorl expansion rate(W = 1.5–2.5). As shown by Reyment (1973), when thebody chamber is less than one whorl (Fig. 2. 9) buoy-ancy is so high and stability is so low that the shell canonly float on the water surface in the subhorizontalposition. To go deeper in the water and turn into the ver-tical position such a shell should have a body chamberof more than one and a half whorls or a similar volumeof fluid in the phragmocone. In members of the orderTarphycerida, most of which populated this mor-phospace, in contrast to ammonoids from the samemorphospace (see below), the body chamber did notexceed one whorl. In addition, in these taxa the aper-tural part of the body chamber was turned outwards(Trochoceras, Shumardoceras, etc.) from the remain-ing part of the last whorl, while the aperture was oftennarrow and even almost closed (Ophioceras)
(a)
(b) (c) (d)
Fig. 2.9. Buoyancy and stability of spirally coiled shellswith different whorl expansion rate and their whorl overlapdegree; (a, b) evolute shells with a low whorl expansion rate(typified by Dactylioceras): (a) position of the shell in thewater, if the body chamber is less than one whorl, (b) posi-tion of the same shell, if the body chamber is one and a halfwhorls; (c) involute shell with rapidly expanding whorls(typified by Nautilus); (d) involute shell with moderatelyrapidly expanding whorls (typified by Ceratites) (after Rey-ment, 1973).
3N
2N
1N
4N
1
3
4
3
1
2
1
(a)
(b)
(c)
Morphospace 3NW = 1.5–2.5; D > 0.4Planktonic life-form
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
W W = 1/D
Fig. 2.10. Major life-forms of nonammonoid cephalopods with a planispiral shell. Morphospace 3N: (a) Ophioceras S3, (b) Disco-ceras O2–3, (c) Millkoninckioceras C1–P.
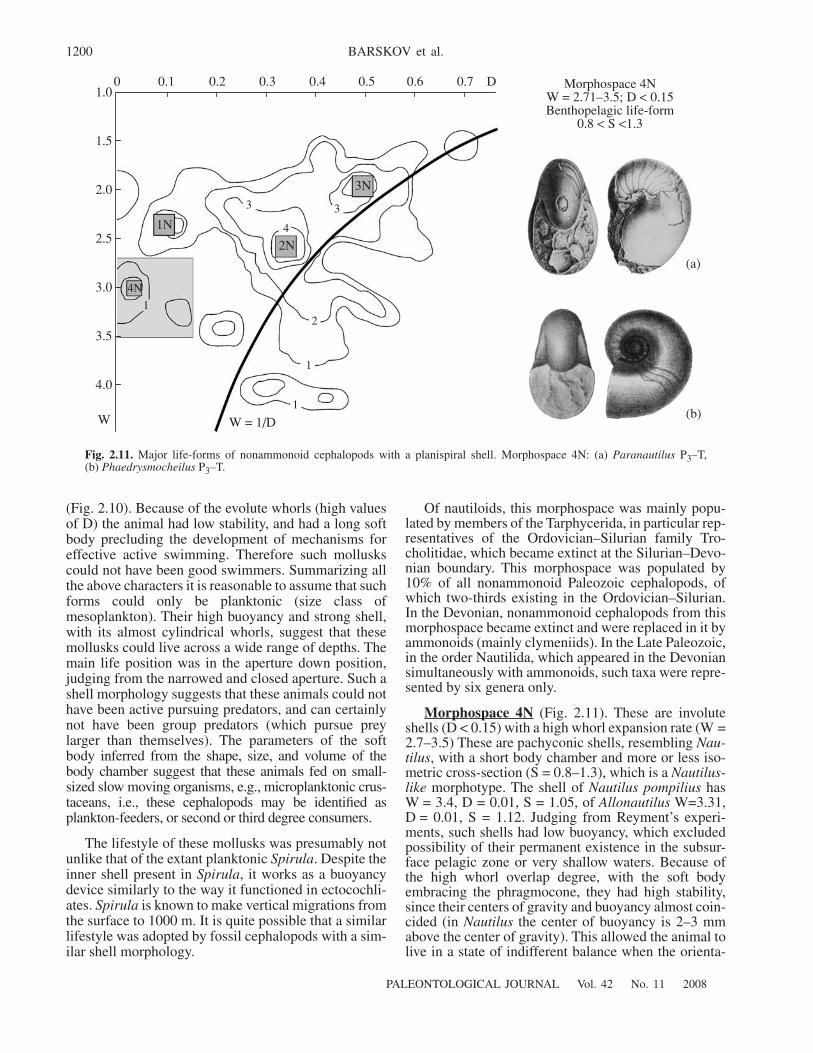
1200
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
(Fig. 2.10). Because of the evolute whorls (high valuesof D) the animal had low stability, and had a long softbody precluding the development of mechanisms foreffective active swimming. Therefore such molluskscould not have been good swimmers. Summarizing allthe above characters it is reasonable to assume that suchforms could only be planktonic (size class ofmesoplankton). Their high buoyancy and strong shell,with its almost cylindrical whorls, suggest that thesemollusks could live across a wide range of depths. Themain life position was in the aperture down position,judging from the narrowed and closed aperture. Such ashell morphology suggests that these animals could nothave been active pursuing predators, and can certainlynot have been group predators (which pursue preylarger than themselves). The parameters of the softbody inferred from the shape, size, and volume of thebody chamber suggest that these animals fed on small-sized slow moving organisms, e.g., microplanktonic crus-taceans, i.e., these cephalopods may be identified asplankton-feeders, or second or third degree consumers.
The lifestyle of these mollusks was presumably notunlike that of the extant planktonic Spirula. Despite theinner shell present in Spirula, it works as a buoyancydevice similarly to the way it functioned in ectocochli-ates. Spirula is known to make vertical migrations fromthe surface to 1000 m. It is quite possible that a similarlifestyle was adopted by fossil cephalopods with a sim-ilar shell morphology.
Of nautiloids, this morphospace was mainly popu-lated by members of the Tarphycerida, in particular rep-resentatives of the Ordovician–Silurian family Tro-cholitidae, which became extinct at the Silurian–Devo-nian boundary. This morphospace was populated by10% of all nonammonoid Paleozoic cephalopods, ofwhich two-thirds existing in the Ordovician–Silurian.In the Devonian, nonammonoid cephalopods from thismorphospace became extinct and were replaced in it byammonoids (mainly clymeniids). In the Late Paleozoic,in the order Nautilida, which appeared in the Devoniansimultaneously with ammonoids, such taxa were repre-sented by six genera only.
Morphospace 4N (Fig. 2.11). These are involuteshells (D < 0.15) with a high whorl expansion rate (W =2.7–3.5) These are pachyconic shells, resembling Nau-tilus, with a short body chamber and more or less iso-metric cross-section (S = 0.8–1.3), which is a Nautilus-like morphotype. The shell of Nautilus pompilius hasW = 3.4, D = 0.01, S = 1.05, of Allonautilus W=3.31,D = 0.01, S = 1.12. Judging from Reyment’s experi-ments, such shells had low buoyancy, which excludedpossibility of their permanent existence in the subsur-face pelagic zone or very shallow waters. Because ofthe high whorl overlap degree, with the soft bodyembracing the phragmocone, they had high stability,since their centers of gravity and buoyancy almost coin-cided (in Nautilus the center of buoyancy is 2–3 mmabove the center of gravity). This allowed the animal tolive in a state of indifferent balance when the orienta-
3N
2N
1N
4N
1
3
4
3
1
2
1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
W W = 1/D
(a)
(b)
Morphospace 4NW = 2.71–3.5; D < 0.15Benthopelagic life-form
0.8 < S <1.3
Fig. 2.11. Major life-forms of nonammonoid cephalopods with a planispiral shell. Morphospace 4N: (a) Paranautilus P3–T,(b) Phaedrysmocheilus P3–T.
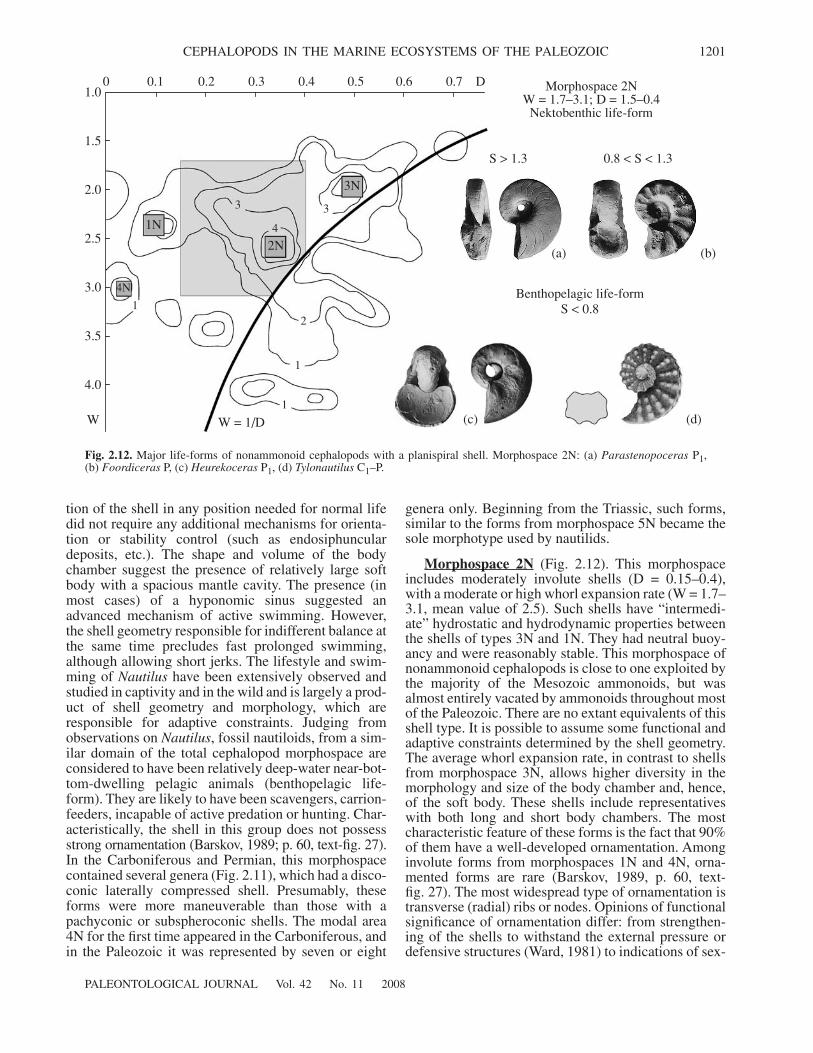
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1201
tion of the shell in any position needed for normal lifedid not require any additional mechanisms for orienta-tion or stability control (such as endosiphunculardeposits, etc.). The shape and volume of the bodychamber suggest the presence of relatively large softbody with a spacious mantle cavity. The presence (inmost cases) of a hyponomic sinus suggested anadvanced mechanism of active swimming. However,the shell geometry responsible for indifferent balance atthe same time precludes fast prolonged swimming,although allowing short jerks. The lifestyle and swim-ming of Nautilus have been extensively observed andstudied in captivity and in the wild and is largely a prod-uct of shell geometry and morphology, which areresponsible for adaptive constraints. Judging fromobservations on Nautilus, fossil nautiloids, from a sim-ilar domain of the total cephalopod morphospace areconsidered to have been relatively deep-water near-bot-tom-dwelling pelagic animals (benthopelagic life-form). They are likely to have been scavengers, carrion-feeders, incapable of active predation or hunting. Char-acteristically, the shell in this group does not possessstrong ornamentation (Barskov, 1989; p. 60, text-fig. 27).In the Carboniferous and Permian, this morphospacecontained several genera (Fig. 2.11), which had a disco-conic laterally compressed shell. Presumably, theseforms were more maneuverable than those with apachyconic or subspheroconic shells. The modal area4N for the first time appeared in the Carboniferous, andin the Paleozoic it was represented by seven or eight
genera only. Beginning from the Triassic, such forms,similar to the forms from morphospace 5N became thesole morphotype used by nautilids.
Morphospace 2N (Fig. 2.12). This morphospaceincludes moderately involute shells (D = 0.15–0.4),with a moderate or high whorl expansion rate (W = 1.7–3.1, mean value of 2.5). Such shells have “intermedi-ate” hydrostatic and hydrodynamic properties betweenthe shells of types 3N and 1N. They had neutral buoy-ancy and were reasonably stable. This morphospace ofnonammonoid cephalopods is close to one exploited bythe majority of the Mesozoic ammonoids, but wasalmost entirely vacated by ammonoids throughout mostof the Paleozoic. There are no extant equivalents of thisshell type. It is possible to assume some functional andadaptive constraints determined by the shell geometry.The average whorl expansion rate, in contrast to shellsfrom morphospace 3N, allows higher diversity in themorphology and size of the body chamber and, hence,of the soft body. These shells include representativeswith both long and short body chambers. The mostcharacteristic feature of these forms is the fact that 90%of them have a well-developed ornamentation. Amonginvolute forms from morphospaces 1N and 4N, orna-mented forms are rare (Barskov, 1989, p. 60, text-fig. 27). The most widespread type of ornamentation istransverse (radial) ribs or nodes. Opinions of functionalsignificance of ornamentation differ: from strengthen-ing of the shells to withstand the external pressure ordefensive structures (Ward, 1981) to indications of sex-
3N
2N
1N
4N
1
3
4
3
1
2
1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
W W = 1/D
(b)
S > 1.3 0.8 < S < 1.3
S < 0.8
Morphospace 2NW = 1.7–3.1; D = 1.5–0.4
Nektobenthic life-form
Benthopelagic life-form
Fig. 2.12. Major life-forms of nonammonoid cephalopods with a planispiral shell. Morphospace 2N: (a) Parastenopoceras P1,(b) Foordiceras P, (c) Heurekoceras P1, (d) Tylonautilus C1–P.
(a)
(c) (d)
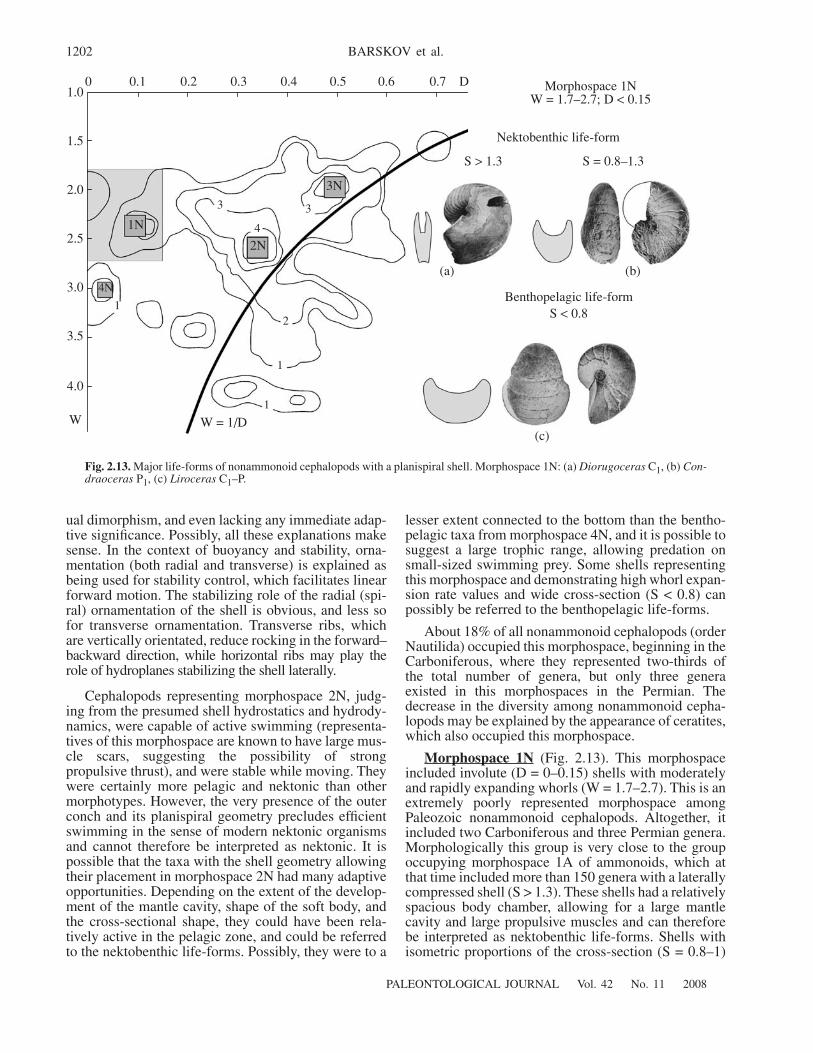
1202
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
ual dimorphism, and even lacking any immediate adap-tive significance. Possibly, all these explanations makesense. In the context of buoyancy and stability, orna-mentation (both radial and transverse) is explained asbeing used for stability control, which facilitates linearforward motion. The stabilizing role of the radial (spi-ral) ornamentation of the shell is obvious, and less sofor transverse ornamentation. Transverse ribs, whichare vertically orientated, reduce rocking in the forward–backward direction, while horizontal ribs may play therole of hydroplanes stabilizing the shell laterally.
Cephalopods representing morphospace 2N, judg-ing from the presumed shell hydrostatics and hydrody-namics, were capable of active swimming (representa-tives of this morphospace are known to have large mus-cle scars, suggesting the possibility of strongpropulsive thrust), and were stable while moving. Theywere certainly more pelagic and nektonic than othermorphotypes. However, the very presence of the outerconch and its planispiral geometry precludes efficientswimming in the sense of modern nektonic organismsand cannot therefore be interpreted as nektonic. It ispossible that the taxa with the shell geometry allowingtheir placement in morphospace 2N had many adaptiveopportunities. Depending on the extent of the develop-ment of the mantle cavity, shape of the soft body, andthe cross-sectional shape, they could have been rela-tively active in the pelagic zone, and could be referredto the nektobenthic life-forms. Possibly, they were to a
lesser extent connected to the bottom than the bentho-pelagic taxa from morphospace 4N, and it is possible tosuggest a large trophic range, allowing predation onsmall-sized swimming prey. Some shells representingthis morphospace and demonstrating high whorl expan-sion rate values and wide cross-section (S < 0.8) canpossibly be referred to the benthopelagic life-forms.
About 18% of all nonammonoid cephalopods (orderNautilida) occupied this morphospace, beginning in theCarboniferous, where they represented two-thirds ofthe total number of genera, but only three generaexisted in this morphospaces in the Permian. Thedecrease in the diversity among nonammonoid cepha-lopods may be explained by the appearance of ceratites,which also occupied this morphospace.
Morphospace 1N (Fig. 2.13). This morphospaceincluded involute (D = 0–0.15) shells with moderatelyand rapidly expanding whorls (W = 1.7–2.7). This is anextremely poorly represented morphospace amongPaleozoic nonammonoid cephalopods. Altogether, itincluded two Carboniferous and three Permian genera.Morphologically this group is very close to the groupoccupying morphospace 1A of ammonoids, which atthat time included more than 150 genera with a laterallycompressed shell (S > 1.3). These shells had a relativelyspacious body chamber, allowing for a large mantlecavity and large propulsive muscles and can thereforebe interpreted as nektobenthic life-forms. Shells withisometric proportions of the cross-section (S = 0.8–1)
3N
2N
1N
4N
1
3
4
3
1
2
1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
W W = 1/D
(a) (b)
(c)
Morphospace 1NW = 1.7–2.7; D < 0.15
Nektobenthic life-form
S > 1.3 S = 0.8–1.3
Benthopelagic life-formS < 0.8
Fig. 2.13. Major life-forms of nonammonoid cephalopods with a planispiral shell. Morphospace 1N: (a) Diorugoceras C1, (b) Con-draoceras P1, (c) Liroceras C1–P.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1203
possibly belonged to the benthopelagic life-forms, likethe modern Nautilus, but were more shifted toward thenektonic lifestyle. They were most likely efficient scav-engers and possibly predators of slow moving medium-sized prey.
Large subspheroconic (S < 0.7) shells with a dors-oventrally compressed body (Permian Permonautilus,Triassic Sibyllonautilus) had poorer hydrodynamics,but were stronger (Chamberlain, 1980; Kiselev, 2006)than Nautilus. They had a less efficient mechanism foractive swimming. They are interpreted as benthope-lagic life-forms, possibly less connected with the bot-tom than the modern Nautilus.
Barskov (1976, 1988, 1989) was the first to proposethe above typification of the life-forms of nonam-monoid cephalopods, which was later extended toammonoids. The interpretation was mainly based onthe parameters W and D. Differences in the parameter S,which determines the cross-sectional shape and there-fore largely the soft body and potential adaptations todifferent lifestyles, received less study. If this approachis to some extent justified for nonammonoid cephalo-pods, the total generic diversity of which was about100 genera and which are not much diverse in theircross-sectional shape: over 60% of all genera havealmost isometric cross-section (S = 0.8–1.3), about25% are compressed laterally (S over 1.3), and less than15% have the shell compressed dorsoventrally) (S lessthan 0.7). For ammonoids, which are considerablymore numerous (this analysis deals with more than600 genera) and have large quantitative and qualitativevariations of the cross-section, this approach is inade-quate. In addition, the above papers did not explain theconsiderable difference in the position of morphologi-cal peaks in ammonoids and nonammonoid cephalo-pods, while this, as shown below, allows the diversity ofthe ammonoid shells within the same morphospacedefined by parameter W and D only, which allows theirassignment to different life-forms.
AmmonoidsThe morphospaces occupied by the coiled shells of
Paleozoic ammonoids and nonammonoid cephalopodsare largely overlapped. This overlap indicates that theacquisition of similar shell geometry by completely dif-ferent and phylogenetically unrelated higher taxa (over10 orders) is to a large extent adaptive. At the same time,preferred (dominant) morphospaces of ammonoids andnonammonoid cephalopods for the same time spanshave different position in the total all-cephalopod mor-phospace. As noted above all preferred morphospacesoccupied by ammonoids, while coinciding with theareas occupied by nonammonoid cephalopods in thevalues of the parameter D (whorl overlap degree), havelower values of W. According to Saunders et al. (2004)and our own data, 72% of ammonoid genera have avalue for parameter W of less than 1.75, whereas innonammonoid cephalopods such genera are less than
10%. In other words, in general “typical” ammonoidswith the same values of the parameter D (which proba-bly characterizes stability) have a lower whorl expan-sion rate. From the point of view of hydrostatics, thissuggests that ammonoids had higher buoyancy,although some of them had shells that are morphologi-cally completely identical to those of nonammonoidcephalopods. Therefore, ammonoids had more possi-bilities of buoyancy control and, hence, better mecha-nisms of filling and emptying the phragmocone cham-bers. For instance, it is suggested that in this process, inaddition to the mechanism of the partial osmosis as inNautilus (Denton et al., 1961), ammonoids used capil-lary absorption (Barskov, 1999). In addition,ammonoids in general have a higher diversity of pro-portions of the body chamber and of the whorl cross-section. For instance, among nonammonoid cephalo-pods, only six genera have narrow whorls with anarrow-shaped cross-section with a pointed ventral keel,and no cadiconic or spheroconic shells with a value ofparameter S less than 0.5. This suggests a possibility ofmore diverse adaptations and, hence, higher morpho-logical diversity of life-forms of ammonoids belonging,according to their Raup’s parameters, to the same peakregion.
Morphospace 1A (Fig. 2.14): involute (D = 0–1.5)slowly expanding forms (W = 1–2.3). This region ispopulated by 240 genera (37% of the total number ofgenera of Paleozoic ammonoids).
A detailed analysis of the distribution of parametersW and D shows that this region can be split into twomore or less separate morphological groups. The first(morphological group 1A') embraces the region withthe lowest values of W: W < 1.7. The second (morpho-logical group 1A), which includes the majority offorms from this region is characterized by the values ofW around 1.7–2.3 with a mean value of W = 2.0. Below,each of these morphological groups are discussedin detail.
Morphogroup 1A' (Fig. 2.14). These are involute,narrowly umbilicate shells (D ~ 0.01–0.15) with thelowest whorl expansion rate (W < 1.7) among coiledforms. Very low whorls and their almost incompleteoverlap strongly change the shape of the body chamberand, hence, the soft body, which becomes compressedU- or V-like. An animal with this type of shell cannot beassumed to be capable of active swimming, or of con-siderable vertical migrations, while high stability couldallow the existence above the fair-weather wave base.The upper pelagic zone was their most likely, if not theonly possible, adaptive zone. This is supported by theappearance in this morphogroup of taxa with inflated,spheroconic chambers (Epiwocklumeria, Parawock-lumeria), with specific constrictions (Prolobites, Reni-tes) and, as usual, small-sized shells. This involute mor-photype we interpret as a planktonic life-form (in clas-sification accepted in this work, it is named “plankton 2”).Morphogroup 1A' contains forms with various propor-
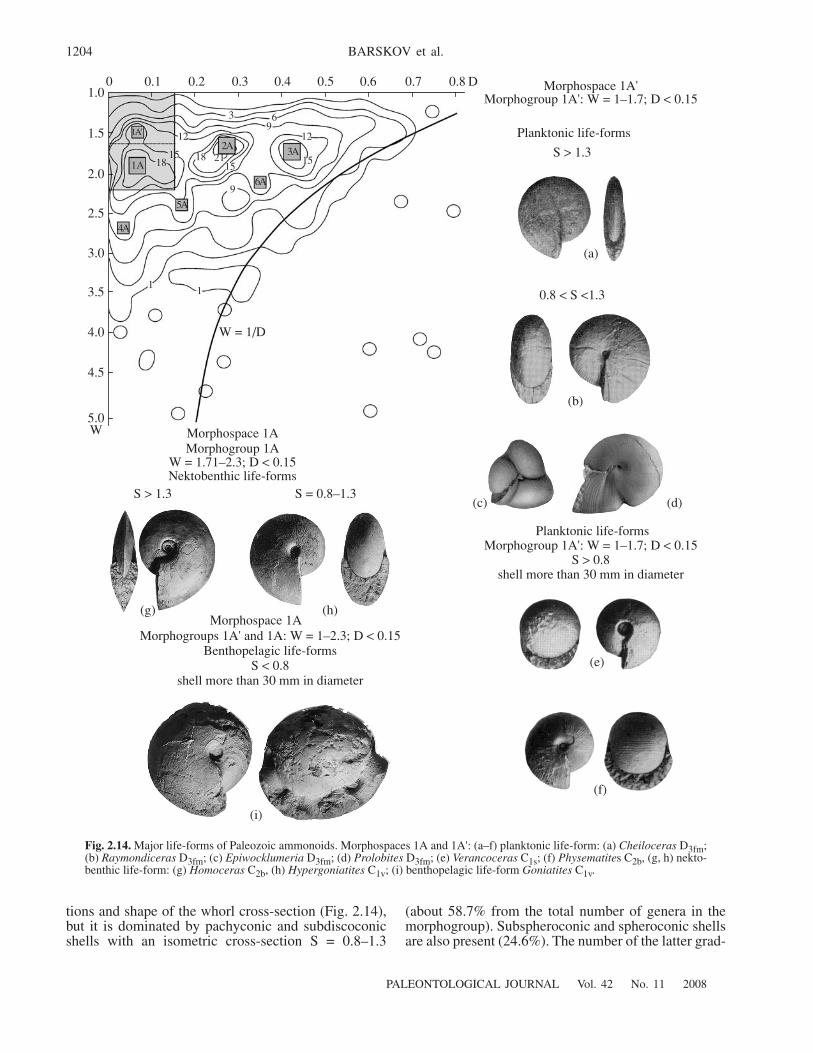
1204
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
tions and shape of the whorl cross-section (Fig. 2.14),but it is dominated by pachyconic and subdiscoconicshells with an isometric cross-section S = 0.8–1.3
(about 58.7% from the total number of genera in themorphogroup). Subspheroconic and spheroconic shellsare also present (24.6%). The number of the latter grad-
12
3 69
2118151518
11
12
15
1A'
1A
4A
5A
6A9
3A2A
W = 1/D
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
(a)
(b)
(c) (d)
(g) (h)
(e)
(f)
(i)
Morphospace 1A'Morphogroup 1A': W = 1–1.7; D < 0.15
Planktonic life-forms
S > 1.3
0.8 < S <1.3
S > 1.3 S = 0.8–1.3
Morphogroups 1A' and 1A: W = 1–2.3; D < 0.15Benthopelagic life-forms
S < 0.8shell more than 30 mm in diameter
Morphogroup 1A': W = 1–1.7; D < 0.15 Planktonic life-forms
S > 0.8shell more than 30 mm in diameter
Morphogroup 1AW = 1.71–2.3; D < 0.15Nektobenthic life-forms
Fig. 2.14. Major life-forms of Paleozoic ammonoids. Morphospaces 1A and 1A': (a–f) planktonic life-form: (a) Cheiloceras D3fm;(b) Raymondiceras D3fm; (c) Epiwocklumeria D3fm; (d) Prolobites D3fm; (e) Verancoceras C1s; (f) Physematites C2b, (g, h) nekto-benthic life-form: (g) Homoceras C2b, (h) Hypergoniatites C1v; (i) benthopelagic life-form Goniatites C1v.
Morphospace 1A
Morphospace 1A
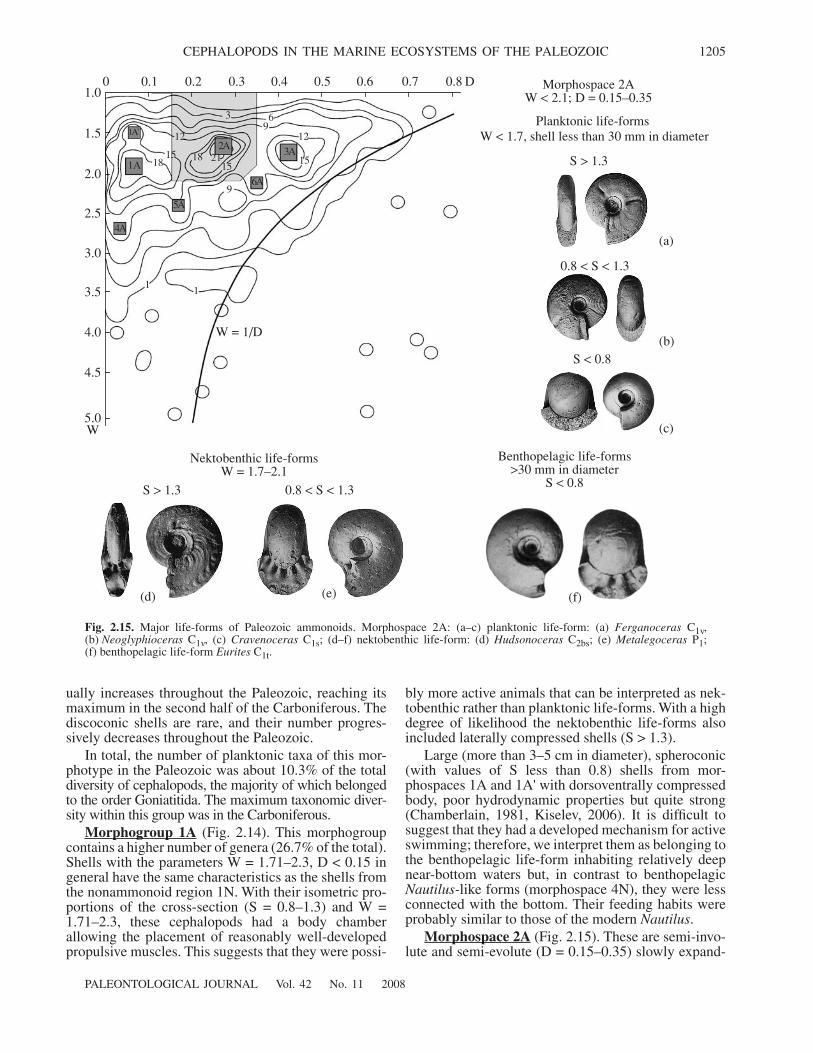
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1205
ually increases throughout the Paleozoic, reaching itsmaximum in the second half of the Carboniferous. Thediscoconic shells are rare, and their number progres-sively decreases throughout the Paleozoic.
In total, the number of planktonic taxa of this mor-photype in the Paleozoic was about 10.3% of the totaldiversity of cephalopods, the majority of which belongedto the order Goniatitida. The maximum taxonomic diver-sity within this group was in the Carboniferous.
Morphogroup 1A (Fig. 2.14). This morphogroupcontains a higher number of genera (26.7% of the total).Shells with the parameters W = 1.71–2.3, D < 0.15 ingeneral have the same characteristics as the shells fromthe nonammonoid region 1N. With their isometric pro-portions of the cross-section (S = 0.8–1.3) and W =1.71–2.3, these cephalopods had a body chamberallowing the placement of reasonably well-developedpropulsive muscles. This suggests that they were possi-
bly more active animals that can be interpreted as nek-tobenthic rather than planktonic life-forms. With a highdegree of likelihood the nektobenthic life-forms alsoincluded laterally compressed shells (S > 1.3).
Large (more than 3–5 cm in diameter), spheroconic(with values of S less than 0.8) shells from mor-phospaces 1A and 1A' with dorsoventrally compressedbody, poor hydrodynamic properties but quite strong(Chamberlain, 1981, Kiselev, 2006). It is difficult tosuggest that they had a developed mechanism for activeswimming; therefore, we interpret them as belonging tothe benthopelagic life-form inhabiting relatively deepnear-bottom waters but, in contrast to benthopelagicNautilus-like forms (morphospace 4N), they were lessconnected with the bottom. Their feeding habits wereprobably similar to those of the modern Nautilus.
Morphospace 2A (Fig. 2.15). These are semi-invo-lute and semi-evolute (D = 0.15–0.35) slowly expand-
12
3 69
2118151518
11
12
15
1A'
1A
4A
5A
6A9
3A2A
W = 1/D
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
(a)
(b)
(c)
(d) (e) (f)
Nektobenthic life-forms
S > 1.3
W = 1.7–2.1
Morphospace 2AW < 2.1; D = 0.15–0.35
Planktonic life-formsW < 1.7, shell less than 30 mm in diameter
S > 1.3
0.8 < S < 1.3
S < 0.8
Benthopelagic life-forms
0.8 < S < 1.3
>30 mm in diameterS < 0.8
Fig. 2.15. Major life-forms of Paleozoic ammonoids. Morphospace 2A: (a–c) planktonic life-form: (a) Ferganoceras C1v,(b) Neoglyphioceras C1v, (c) Cravenoceras C1s; (d–f) nektobenthic life-form: (d) Hudsonoceras C2bs; (e) Metalegoceras P1;(f) benthopelagic life-form Eurites C1t.
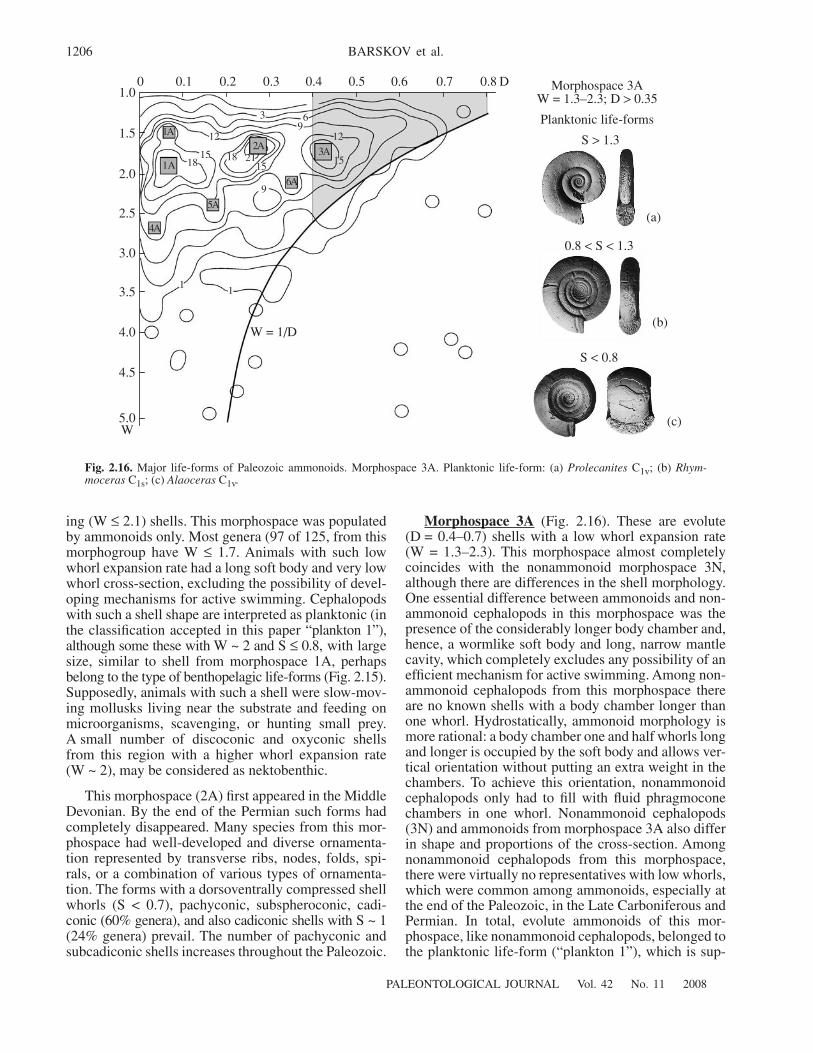
1206
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
ing (W ≤ 2.1) shells. This morphospace was populatedby ammonoids only. Most genera (97 of 125, from thismorphogroup have W ≤ 1.7. Animals with such lowwhorl expansion rate had a long soft body and very lowwhorl cross-section, excluding the possibility of devel-oping mechanisms for active swimming. Cephalopodswith such a shell shape are interpreted as planktonic (inthe classification accepted in this paper “plankton 1”),although some these with W ~ 2 and S ≤ 0.8, with largesize, similar to shell from morphospace 1A, perhapsbelong to the type of benthopelagic life-forms (Fig. 2.15).Supposedly, animals with such a shell were slow-mov-ing mollusks living near the substrate and feeding onmicroorganisms, scavenging, or hunting small prey.A small number of discoconic and oxyconic shellsfrom this region with a higher whorl expansion rate(W ~ 2), may be considered as nektobenthic.
This morphospace (2A) first appeared in the MiddleDevonian. By the end of the Permian such forms hadcompletely disappeared. Many species from this mor-phospace had well-developed and diverse ornamenta-tion represented by transverse ribs, nodes, folds, spi-rals, or a combination of various types of ornamenta-tion. The forms with a dorsoventrally compressed shellwhorls (S < 0.7), pachyconic, subspheroconic, cadi-conic (60% genera), and also cadiconic shells with S ~ 1(24% genera) prevail. The number of pachyconic andsubcadiconic shells increases throughout the Paleozoic.
Morphospace 3A (Fig. 2.16). These are evolute(D = 0.4–0.7) shells with a low whorl expansion rate(W = 1.3–2.3). This morphospace almost completelycoincides with the nonammonoid morphospace 3N,although there are differences in the shell morphology.One essential difference between ammonoids and non-ammonoid cephalopods in this morphospace was thepresence of the considerably longer body chamber and,hence, a wormlike soft body and long, narrow mantlecavity, which completely excludes any possibility of anefficient mechanism for active swimming. Among non-ammonoid cephalopods from this morphospace thereare no known shells with a body chamber longer thanone whorl. Hydrostatically, ammonoid morphology ismore rational: a body chamber one and half whorls longand longer is occupied by the soft body and allows ver-tical orientation without putting an extra weight in thechambers. To achieve this orientation, nonammonoidcephalopods only had to fill with fluid phragmoconechambers in one whorl. Nonammonoid cephalopods(3N) and ammonoids from morphospace 3A also differin shape and proportions of the cross-section. Amongnonammonoid cephalopods from this morphospace,there were virtually no representatives with low whorls,which were common among ammonoids, especially atthe end of the Paleozoic, in the Late Carboniferous andPermian. In total, evolute ammonoids of this mor-phospace, like nonammonoid cephalopods, belonged tothe planktonic life-form (“plankton 1”), which is sup-
12
3 69
2118151518
11
12
15
1A'
1A
4A
5A
6A9
3A2A
W = 1/D
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
(a)
(b)
(c)
Morphospace 3AW = 1.3–2.3; D > 0.35
Planktonic life-forms
S > 1.3
0.8 < S < 1.3
S < 0.8
Fig. 2.16. Major life-forms of Paleozoic ammonoids. Morphospace 3A. Planktonic life-form: (a) Prolecanites C1v; (b) Rhym-moceras C1s; (c) Alaoceras C1v.
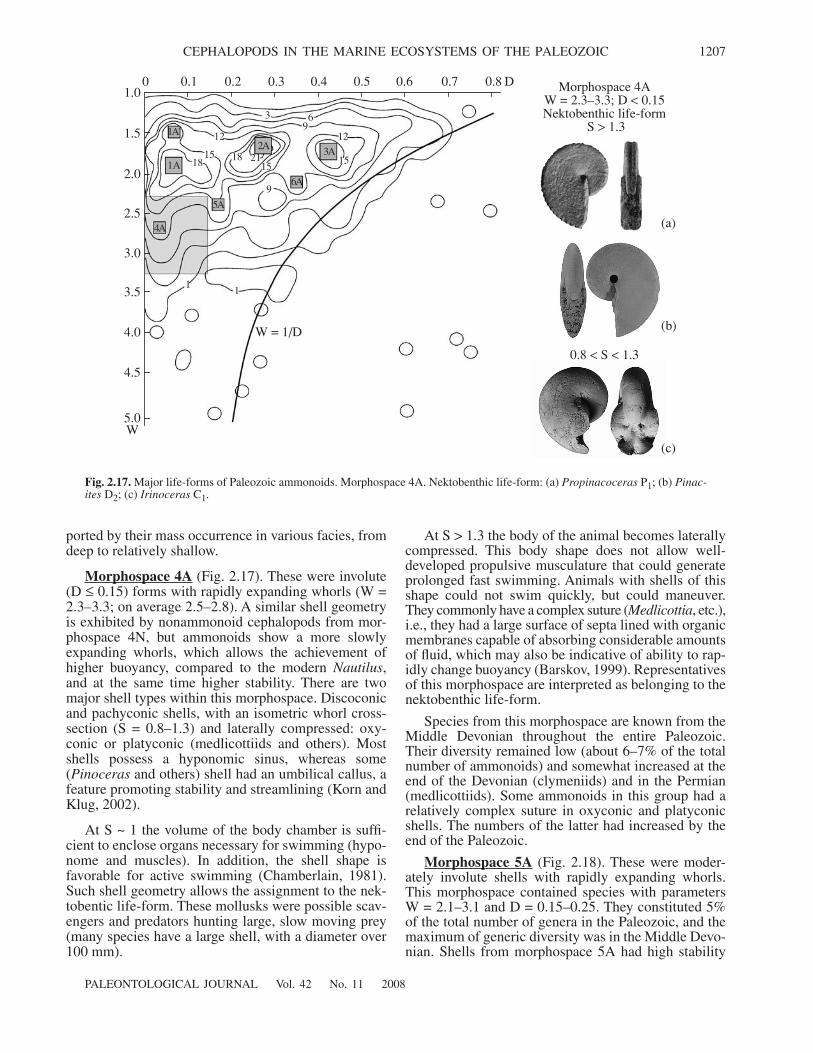
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1207
ported by their mass occurrence in various facies, fromdeep to relatively shallow.
Morphospace 4A (Fig. 2.17). These were involute(D ≤ 0.15) forms with rapidly expanding whorls (W =2.3–3.3; on average 2.5–2.8). A similar shell geometryis exhibited by nonammonoid cephalopods from mor-phospace 4N, but ammonoids show a more slowlyexpanding whorls, which allows the achievement ofhigher buoyancy, compared to the modern Nautilus,and at the same time higher stability. There are twomajor shell types within this morphospace. Discoconicand pachyconic shells, with an isometric whorl cross-section (S = 0.8–1.3) and laterally compressed: oxy-conic or platyconic (medlicottiids and others). Mostshells possess a hyponomic sinus, whereas some(Pinoceras and others) shell had an umbilical callus, afeature promoting stability and streamlining (Korn andKlug, 2002).
At S ~ 1 the volume of the body chamber is suffi-cient to enclose organs necessary for swimming (hypo-nome and muscles). In addition, the shell shape isfavorable for active swimming (Chamberlain, 1981).Such shell geometry allows the assignment to the nek-tobentic life-form. These mollusks were possible scav-engers and predators hunting large, slow moving prey(many species have a large shell, with a diameter over100 mm).
At S > 1.3 the body of the animal becomes laterallycompressed. This body shape does not allow well-developed propulsive musculature that could generateprolonged fast swimming. Animals with shells of thisshape could not swim quickly, but could maneuver.They commonly have a complex suture (Medlicottia, etc.),i.e., they had a large surface of septa lined with organicmembranes capable of absorbing considerable amountsof fluid, which may also be indicative of ability to rap-idly change buoyancy (Barskov, 1999). Representativesof this morphospace are interpreted as belonging to thenektobenthic life-form.
Species from this morphospace are known from theMiddle Devonian throughout the entire Paleozoic.Their diversity remained low (about 6–7% of the totalnumber of ammonoids) and somewhat increased at theend of the Devonian (clymeniids) and in the Permian(medlicottiids). Some ammonoids in this group had arelatively complex suture in oxyconic and platyconicshells. The numbers of the latter had increased by theend of the Paleozoic.
Morphospace 5A (Fig. 2.18). These were moder-ately involute shells with rapidly expanding whorls.This morphospace contained species with parametersW = 2.1–3.1 and D = 0.15–0.25. They constituted 5%of the total number of genera in the Paleozoic, and themaximum of generic diversity was in the Middle Devo-nian. Shells from morphospace 5A had high stability
12
3 69
2118151518
11
12
15
1A'
1A
4A
5A
6A9
3A2A
W = 1/D
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
(a)
(b)
(c)
Morphospace 4AW = 2.3–3.3; D < 0.15Nektobenthic life-form
S > 1.3
0.8 < S < 1.3
Fig. 2.17. Major life-forms of Paleozoic ammonoids. Morphospace 4A. Nektobenthic life-form: (a) Propinacoceras P1; (b) Pinac-ites D2; (c) Irinoceras C1.
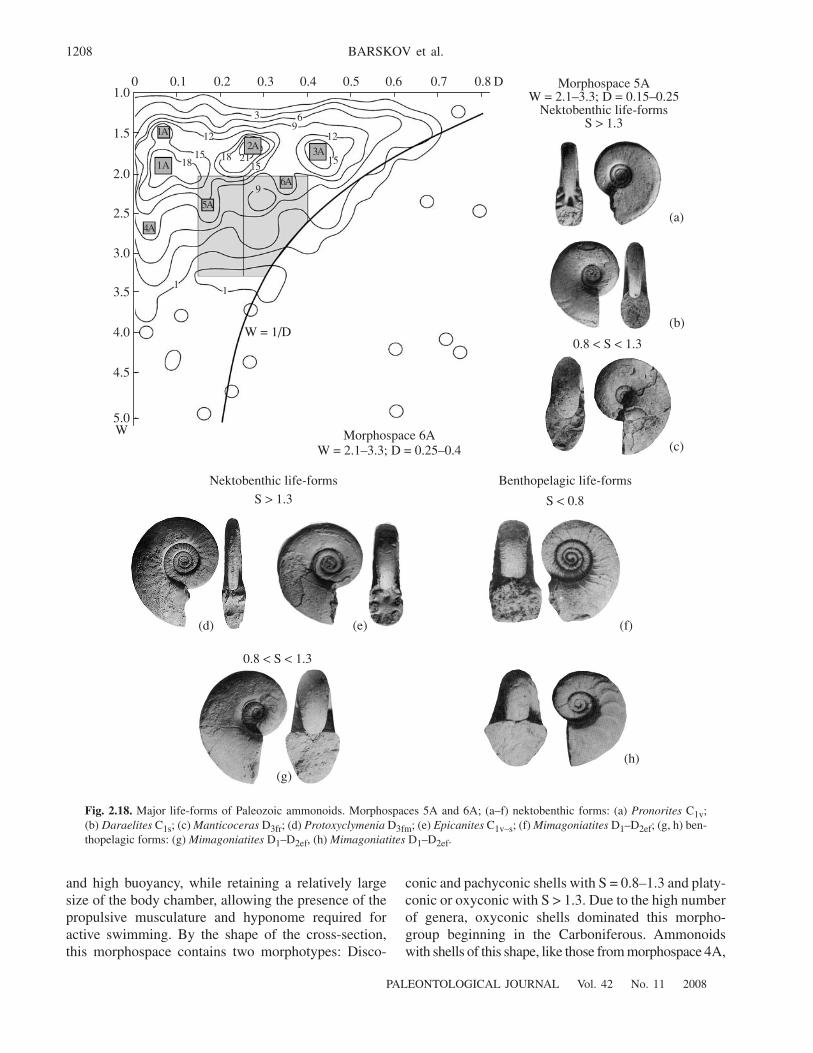
1208
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
and high buoyancy, while retaining a relatively largesize of the body chamber, allowing the presence of thepropulsive musculature and hyponome required foractive swimming. By the shape of the cross-section,this morphospace contains two morphotypes: Disco-
conic and pachyconic shells with S = 0.8–1.3 and platy-conic or oxyconic with S > 1.3. Due to the high numberof genera, oxyconic shells dominated this morpho-group beginning in the Carboniferous. Ammonoidswith shells of this shape, like those from morphospace 4A,
12
3 69
2118151518
11
12
15
1A'
1A
4A
5A
6A9
3A2A
W = 1/D
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
(a)
(b)
(c)
Morphospace 5AW = 2.1–3.3; D = 0.15–0.25
Nektobenthic life-formsS > 1.3
0.8 < S < 1.3
Morphospace 6AW = 2.1–3.3; D = 0.25–0.4
Nektobenthic life-forms Benthopelagic life-forms
S > 1.3 S < 0.8
0.8 < S < 1.3
(d) (e) (f)
(g)(h)
Fig. 2.18. Major life-forms of Paleozoic ammonoids. Morphospaces 5A and 6A; (a–f) nektobenthic forms: (a) Pronorites C1v;(b) Daraelites C1s; (c) Manticoceras D3fr; (d) Protoxyclymenia D3fm; (e) Epicanites C1v–s; (f) Mimagoniatites D1–D2ef; (g, h) ben-thopelagic forms: (g) Mimagoniatites D1–D2ef, (h) Mimagoniatites D1–D2ef.
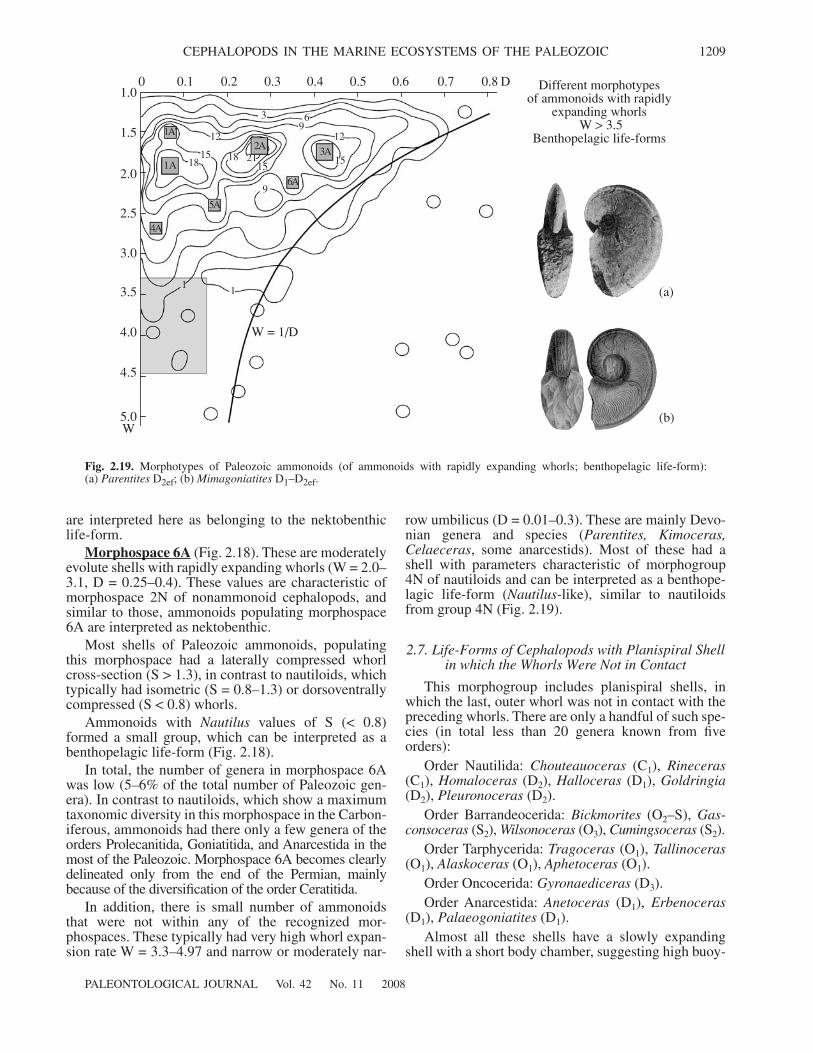
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1209
are interpreted here as belonging to the nektobenthiclife-form.
Morphospace 6A (Fig. 2.18). These are moderatelyevolute shells with rapidly expanding whorls (W = 2.0–3.1, D = 0.25–0.4). These values are characteristic ofmorphospace 2N of nonammonoid cephalopods, andsimilar to those, ammonoids populating morphospace6A are interpreted as nektobenthic.
Most shells of Paleozoic ammonoids, populatingthis morphospace had a laterally compressed whorlcross-section (S > 1.3), in contrast to nautiloids, whichtypically had isometric (S = 0.8–1.3) or dorsoventrallycompressed (S < 0.8) whorls.
Ammonoids with Nautilus values of S (< 0.8)formed a small group, which can be interpreted as abenthopelagic life-form (Fig. 2.18).
In total, the number of genera in morphospace 6Awas low (5–6% of the total number of Paleozoic gen-era). In contrast to nautiloids, which show a maximumtaxonomic diversity in this morphospace in the Carbon-iferous, ammonoids had there only a few genera of theorders Prolecanitida, Goniatitida, and Anarcestida in themost of the Paleozoic. Morphospace 6A becomes clearlydelineated only from the end of the Permian, mainlybecause of the diversification of the order Ceratitida.
In addition, there is small number of ammonoidsthat were not within any of the recognized mor-phospaces. These typically had very high whorl expan-sion rate W = 3.3–4.97 and narrow or moderately nar-
row umbilicus (D = 0.01–0.3). These are mainly Devo-nian genera and species (Parentites, Kimoceras,Celaeceras, some anarcestids). Most of these had ashell with parameters characteristic of morphogroup4N of nautiloids and can be interpreted as a benthope-lagic life-form (Nautilus-like), similar to nautiloidsfrom group 4N (Fig. 2.19).
2.7. Life-Forms of Cephalopods with Planispiral Shellin which the Whorls Were Not in Contact
This morphogroup includes planispiral shells, inwhich the last, outer whorl was not in contact with thepreceding whorls. There are only a handful of such spe-cies (in total less than 20 genera known from fiveorders):
Order Nautilida: Chouteauoceras (C1), Rineceras(C1), Homaloceras (D2), Halloceras (D1), Goldringia(D2), Pleuronoceras (D2).
Order Barrandeocerida: Bickmorites (O2–S), Gas-consoceras (S2), Wilsonoceras (O3), Cumingsoceras (S2).
Order Tarphycerida: Tragoceras (O1), Tallinoceras(O1), Alaskoceras (O1), Aphetoceras (O1).
Order Oncocerida: Gyronaediceras (D3).Order Anarcestida: Anetoceras (D1), Erbenoceras
(D1), Palaeogoniatites (D1).Almost all these shells have a slowly expanding
shell with a short body chamber, suggesting high buoy-
12
3 69
2118151518
11
12
15
1A'
1A
4A
5A
6A9
3A2A
W = 1/D
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 D1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0W
(a)
(b)
Different morphotypesof ammonoids with rapidly
expanding whorls W > 3.5
Benthopelagic life-forms
Fig. 2.19. Morphotypes of Paleozoic ammonoids (of ammonoids with rapidly expanding whorls; benthopelagic life-form):(a) Parentites D2ef; (b) Mimagoniatites D1–D2ef.
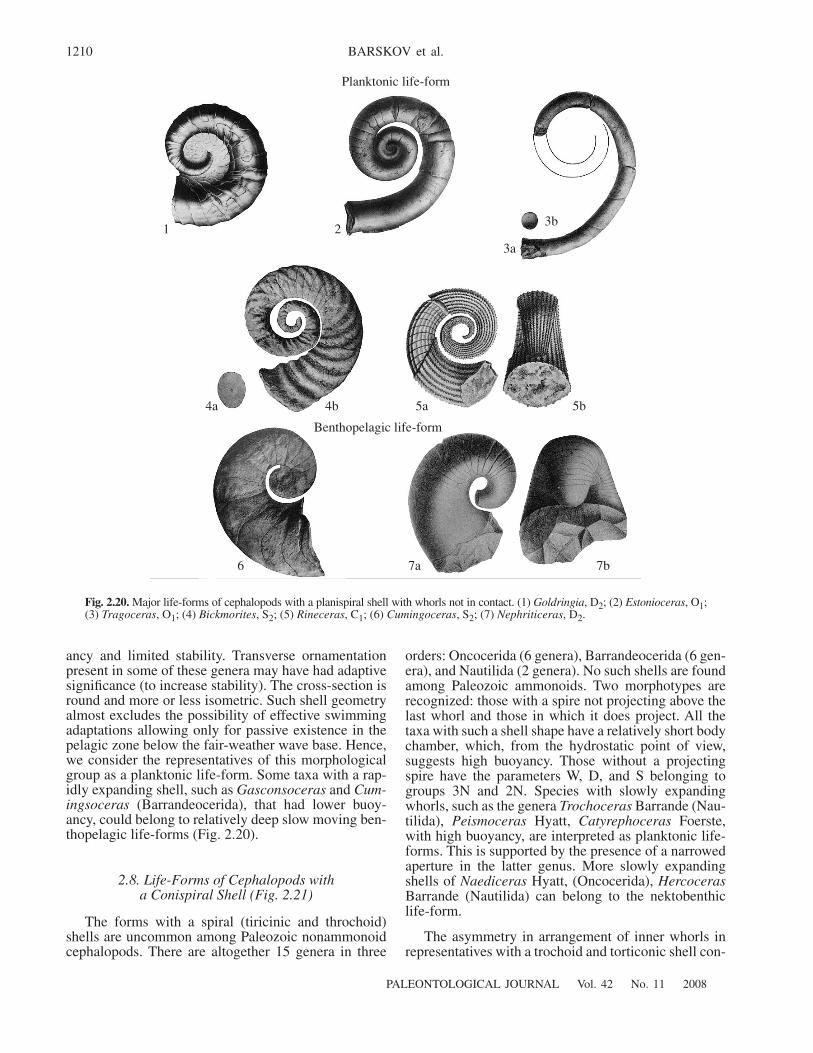
1210
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
ancy and limited stability. Transverse ornamentationpresent in some of these genera may have had adaptivesignificance (to increase stability). The cross-section isround and more or less isometric. Such shell geometryalmost excludes the possibility of effective swimmingadaptations allowing only for passive existence in thepelagic zone below the fair-weather wave base. Hence,we consider the representatives of this morphologicalgroup as a planktonic life-form. Some taxa with a rap-idly expanding shell, such as Gasconsoceras and Cum-ingsoceras (Barrandeocerida), that had lower buoy-ancy, could belong to relatively deep slow moving ben-thopelagic life-forms (Fig. 2.20).
2.8. Life-Forms of Cephalopods witha Conispiral Shell (Fig. 2.21)
The forms with a spiral (tiricinic and throchoid)shells are uncommon among Paleozoic nonammonoidcephalopods. There are altogether 15 genera in three
orders: Oncocerida (6 genera), Barrandeocerida (6 gen-era), and Nautilida (2 genera). No such shells are foundamong Paleozoic ammonoids. Two morphotypes arerecognized: those with a spire not projecting above thelast whorl and those in which it does project. All thetaxa with such a shell shape have a relatively short bodychamber, which, from the hydrostatic point of view,suggests high buoyancy. Those without a projectingspire have the parameters W, D, and S belonging togroups 3N and 2N. Species with slowly expandingwhorls, such as the genera Trochoceras Barrande (Nau-tilida), Peismoceras Hyatt, Catyrephoceras Foerste,with high buoyancy, are interpreted as planktonic life-forms. This is supported by the presence of a narrowedaperture in the latter genus. More slowly expandingshells of Naediceras Hyatt, (Oncocerida), HercocerasBarrande (Nautilida) can belong to the nektobenthiclife-form.
The asymmetry in arrangement of inner whorls inrepresentatives with a trochoid and torticonic shell con-
1 2
3a
3b
4a 4b 5a 5b
6 7a 7b
Planktonic life-form
Benthopelagic life-form
Fig. 2.20. Major life-forms of cephalopods with a planispiral shell with whorls not in contact. (1) Goldringia, D2; (2) Estonioceras, O1;(3) Tragoceras, O1; (4) Bickmorites, S2; (5) Rineceras, C1; (6) Cumingoceras, S2; (7) Nephriticeras, D2.
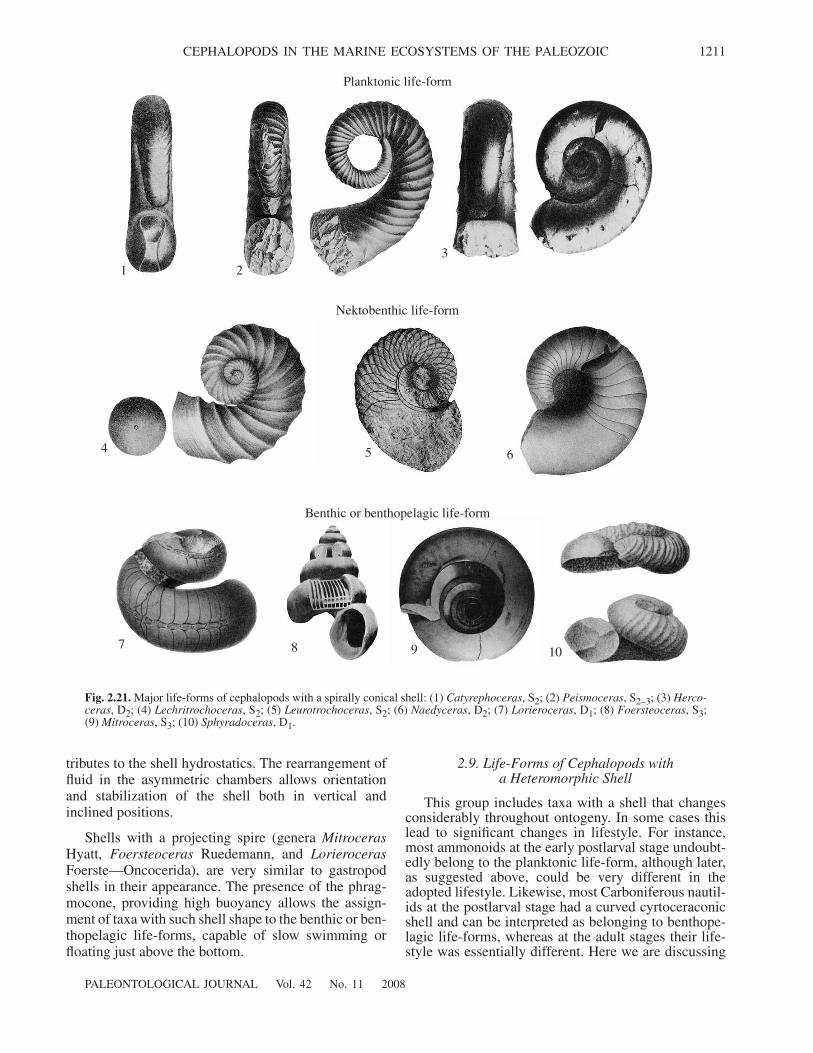
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1211
tributes to the shell hydrostatics. The rearrangement offluid in the asymmetric chambers allows orientationand stabilization of the shell both in vertical andinclined positions.
Shells with a projecting spire (genera MitrocerasHyatt, Foersteoceras Ruedemann, and LorierocerasFoerste—Oncocerida), are very similar to gastropodshells in their appearance. The presence of the phrag-mocone, providing high buoyancy allows the assign-ment of taxa with such shell shape to the benthic or ben-thopelagic life-forms, capable of slow swimming orfloating just above the bottom.
2.9. Life-Forms of Cephalopods witha Heteromorphic Shell
This group includes taxa with a shell that changesconsiderably throughout ontogeny. In some cases thislead to significant changes in lifestyle. For instance,most ammonoids at the early postlarval stage undoubt-edly belong to the planktonic life-form, although later,as suggested above, could be very different in theadopted lifestyle. Likewise, most Carboniferous nautil-ids at the postlarval stage had a curved cyrtoceraconicshell and can be interpreted as belonging to benthope-lagic life-forms, whereas at the adult stages their life-style was essentially different. Here we are discussing
1 23
4 5 6
7 8 9 10
Planktonic life-form
Nektobenthic life-form
Benthic or benthopelagic life-form
Fig. 2.21. Major life-forms of cephalopods with a spirally conical shell: (1) Catyrephoceras, S2; (2) Peismoceras, S2–3; (3) Herco-ceras, D2; (4) Lechritrochoceras, S2; (5) Leurotrochoceras, S2; (6) Naedyceras, D2; (7) Lorieroceras, D1; (8) Foersteoceras, S3;(9) Mitroceras, S3; (10) Sphyradoceras, D1.
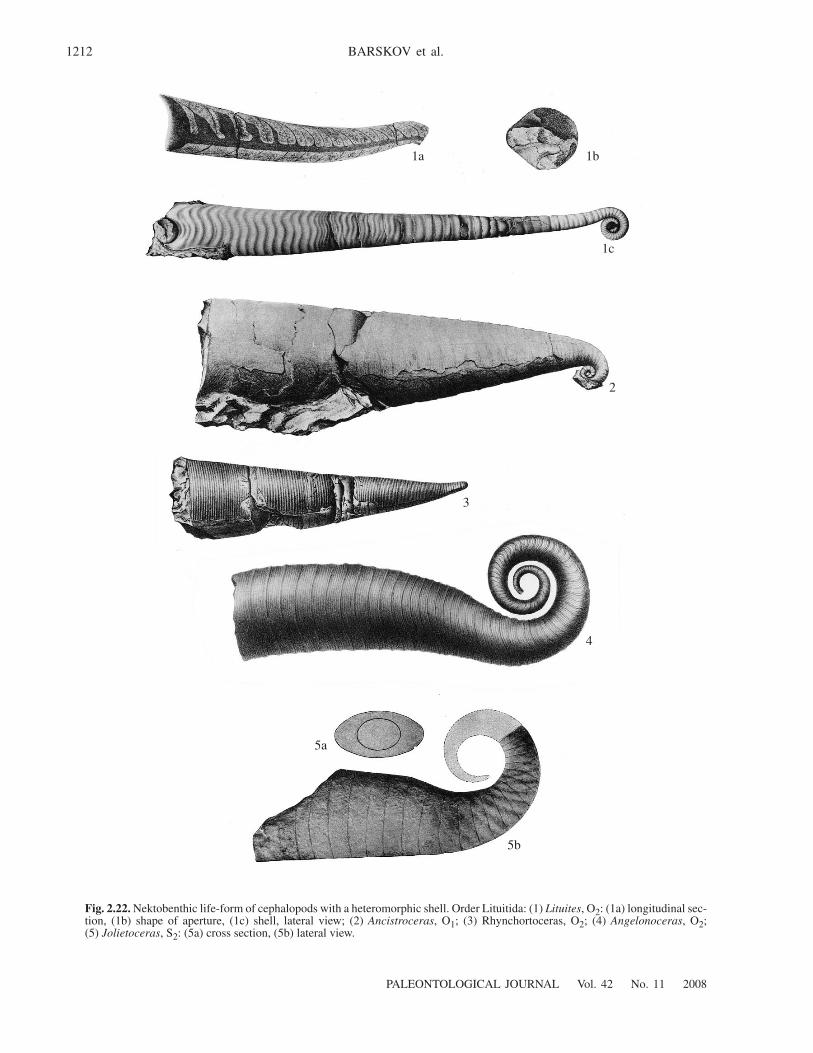
1212
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
1a 1b
1c
2
3
4
5a
5b
Fig. 2.22. Nektobenthic life-form of cephalopods with a heteromorphic shell. Order Lituitida: (1) Lituites, O2: (1a) longitudinal sec-tion, (1b) shape of aperture, (1c) shell, lateral view; (2) Ancistroceras, O1; (3) Rhynchortoceras, O2; (4) Angelonoceras, O2;(5) Jolietoceras, S2: (5a) cross section, (5b) lateral view.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1213
taxa in which shell morphology changed during theadult stage. In the Paleozoic such shell shape was char-acteristic of representatives of the orders Lituitida andAscocerida, and only of a few genera of other orders.
Order Lituitida (Fig. 2.22). In the members of thisorder the initial part of the shell is planispiral with con-
tacting or not contacting whorls, and later the shell isstraight. In one genus (Rhynchorthoceras Remele,1881) the initial part is cyrtoconic. A very short livingchamber and very long phragmocone are typical fea-tures of the body plan in this order. This morphologysuggests high buoyancy, which allows the interpreta-
1a
1b
1c
2a
2b
2c
2e3
4a 4b 5a 5b
6a 6b 6c
Nektobenthic life-form
Planktonic life-form
Fig. 2.23. Life-forms of cephalopods with a heteromorphic shell. Order Ascocerida: (1) Aphragmites, S3: (1a) dorsal side, (1b) sep-tal view, (1c) lateral view; (2) Glossoceras, S2–3 (2a) longitudinal section, (2b) lateral view, (2c) dorsal side, (2e) reconstruction ofthe mollusk; (3) Ascoceras, S2–3; (4) Billingsites, O3: (4a) lateral side, (4b) dorsal side; (5) Probillingsites, O2–3: (5a) ventral side,(5b) lateral side; (6) Schuchertoceras, O3: (6a) lateral view, (6b) ventral view, (6c) dorsal view.
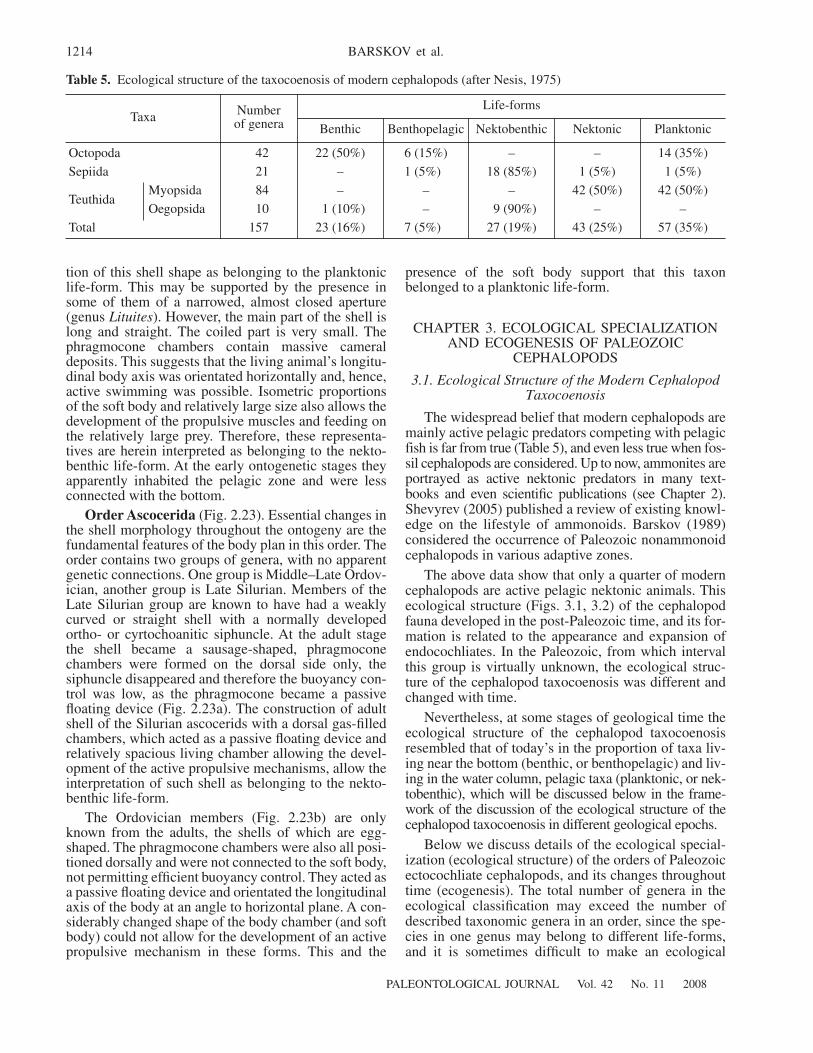
1214
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
tion of this shell shape as belonging to the planktoniclife-form. This may be supported by the presence insome of them of a narrowed, almost closed aperture(genus Lituites). However, the main part of the shell islong and straight. The coiled part is very small. Thephragmocone chambers contain massive cameraldeposits. This suggests that the living animal’s longitu-dinal body axis was orientated horizontally and, hence,active swimming was possible. Isometric proportionsof the soft body and relatively large size also allows thedevelopment of the propulsive muscles and feeding onthe relatively large prey. Therefore, these representa-tives are herein interpreted as belonging to the nekto-benthic life-form. At the early ontogenetic stages theyapparently inhabited the pelagic zone and were lessconnected with the bottom.
Order Ascocerida (Fig. 2.23). Essential changes inthe shell morphology throughout the ontogeny are thefundamental features of the body plan in this order. Theorder contains two groups of genera, with no apparentgenetic connections. One group is Middle–Late Ordov-ician, another group is Late Silurian. Members of theLate Silurian group are known to have had a weaklycurved or straight shell with a normally developedortho- or cyrtochoanitic siphuncle. At the adult stagethe shell became a sausage-shaped, phragmoconechambers were formed on the dorsal side only, thesiphuncle disappeared and therefore the buoyancy con-trol was low, as the phragmocone became a passivefloating device (Fig. 2.23a). The construction of adultshell of the Silurian ascocerids with a dorsal gas-filledchambers, which acted as a passive floating device andrelatively spacious living chamber allowing the devel-opment of the active propulsive mechanisms, allow theinterpretation of such shell as belonging to the nekto-benthic life-form.
The Ordovician members (Fig. 2.23b) are onlyknown from the adults, the shells of which are egg-shaped. The phragmocone chambers were also all posi-tioned dorsally and were not connected to the soft body,not permitting efficient buoyancy control. They acted asa passive floating device and orientated the longitudinalaxis of the body at an angle to horizontal plane. A con-siderably changed shape of the body chamber (and softbody) could not allow for the development of an activepropulsive mechanism in these forms. This and the
presence of the soft body support that this taxonbelonged to a planktonic life-form.
CHAPTER 3. ECOLOGICAL SPECIALIZATION AND ECOGENESIS OF PALEOZOIC
CEPHALOPODS
3.1. Ecological Structure of the Modern Cephalopod Taxocoenosis
The widespread belief that modern cephalopods aremainly active pelagic predators competing with pelagicfish is far from true (Table 5), and even less true when fos-sil cephalopods are considered. Up to now, ammonites areportrayed as active nektonic predators in many text-books and even scientific publications (see Chapter 2).Shevyrev (2005) published a review of existing knowl-edge on the lifestyle of ammonoids. Barskov (1989)considered the occurrence of Paleozoic nonammonoidcephalopods in various adaptive zones.
The above data show that only a quarter of moderncephalopods are active pelagic nektonic animals. Thisecological structure (Figs. 3.1, 3.2) of the cephalopodfauna developed in the post-Paleozoic time, and its for-mation is related to the appearance and expansion ofendocochliates. In the Paleozoic, from which intervalthis group is virtually unknown, the ecological struc-ture of the cephalopod taxocoenosis was different andchanged with time.
Nevertheless, at some stages of geological time theecological structure of the cephalopod taxocoenosisresembled that of today’s in the proportion of taxa liv-ing near the bottom (benthic, or benthopelagic) and liv-ing in the water column, pelagic taxa (planktonic, or nek-tobenthic), which will be discussed below in the frame-work of the discussion of the ecological structure of thecephalopod taxocoenosis in different geological epochs.
Below we discuss details of the ecological special-ization (ecological structure) of the orders of Paleozoicectocochliate cephalopods, and its changes throughouttime (ecogenesis). The total number of genera in theecological classification may exceed the number ofdescribed taxonomic genera in an order, since the spe-cies in one genus may belong to different life-forms,and it is sometimes difficult to make an ecological
Table 5. Ecological structure of the taxocoenosis of modern cephalopods (after Nesis, 1975)
Taxa Numberof genera
Life-forms
Benthic Benthopelagic Nektobenthic Nektonic Planktonic
Octopoda 42 22 (50%) 6 (15%) – – 14 (35%)
Sepiida 21 – 1 (5%) 18 (85%) 1 (5%) 1 (5%)
TeuthidaMyopsida 84 – – – 42 (50%) 42 (50%)
Oegopsida 10 1 (10%) – 9 (90%) – –
Total 157 23 (16%) 7 (5%) 27 (19%) 43 (25%) 57 (35%)
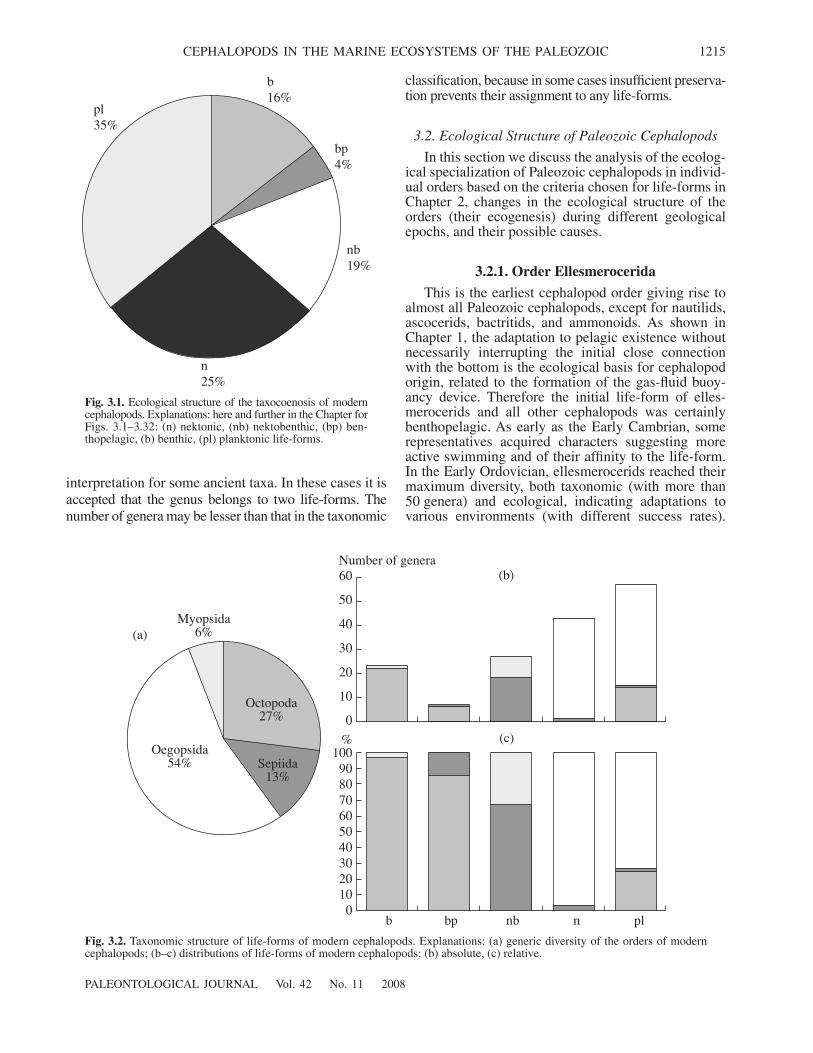
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1215
interpretation for some ancient taxa. In these cases it isaccepted that the genus belongs to two life-forms. Thenumber of genera may be lesser than that in the taxonomic
classification, because in some cases insufficient preserva-tion prevents their assignment to any life-forms.
3.2. Ecological Structure of Paleozoic Cephalopods
In this section we discuss the analysis of the ecolog-ical specialization of Paleozoic cephalopods in individ-ual orders based on the criteria chosen for life-forms inChapter 2, changes in the ecological structure of theorders (their ecogenesis) during different geologicalepochs, and their possible causes.
3.2.1. Order Ellesmerocerida
This is the earliest cephalopod order giving rise toalmost all Paleozoic cephalopods, except for nautilids,ascocerids, bactritids, and ammonoids. As shown inChapter 1, the adaptation to pelagic existence withoutnecessarily interrupting the initial close connectionwith the bottom is the ecological basis for cephalopodorigin, related to the formation of the gas-fluid buoy-ancy device. Therefore the initial life-form of elles-merocerids and all other cephalopods was certainlybenthopelagic. As early as the Early Cambrian, somerepresentatives acquired characters suggesting moreactive swimming and of their affinity to the life-form.In the Early Ordovician, ellesmerocerids reached theirmaximum diversity, both taxonomic (with more than50 genera) and ecological, indicating adaptations tovarious environments (with different success rates).
16%b
4%bp
19%nb
25%n
35%pl
Fig. 3.1. Ecological structure of the taxocoenosis of moderncephalopods. Explanations: here and further in the Chapter forFigs. 3.1–3.32: (n) nektonic, (nb) nektobenthic, (bp) ben-thopelagic, (b) benthic, (pl) planktonic life-forms.
Myopsida6%
Octopoda27%
Sepiida13%
Oegopsida54%
60
50
40
30
20
10
0
100%
908070605040302010
0b bp nb n pl
(b)
(c)
Number of genera
(a)
Fig. 3.2. Taxonomic structure of life-forms of modern cephalopods. Explanations: (a) generic diversity of the orders of moderncephalopods; (b–c) distributions of life-forms of modern cephalopods: (b) absolute, (c) relative.
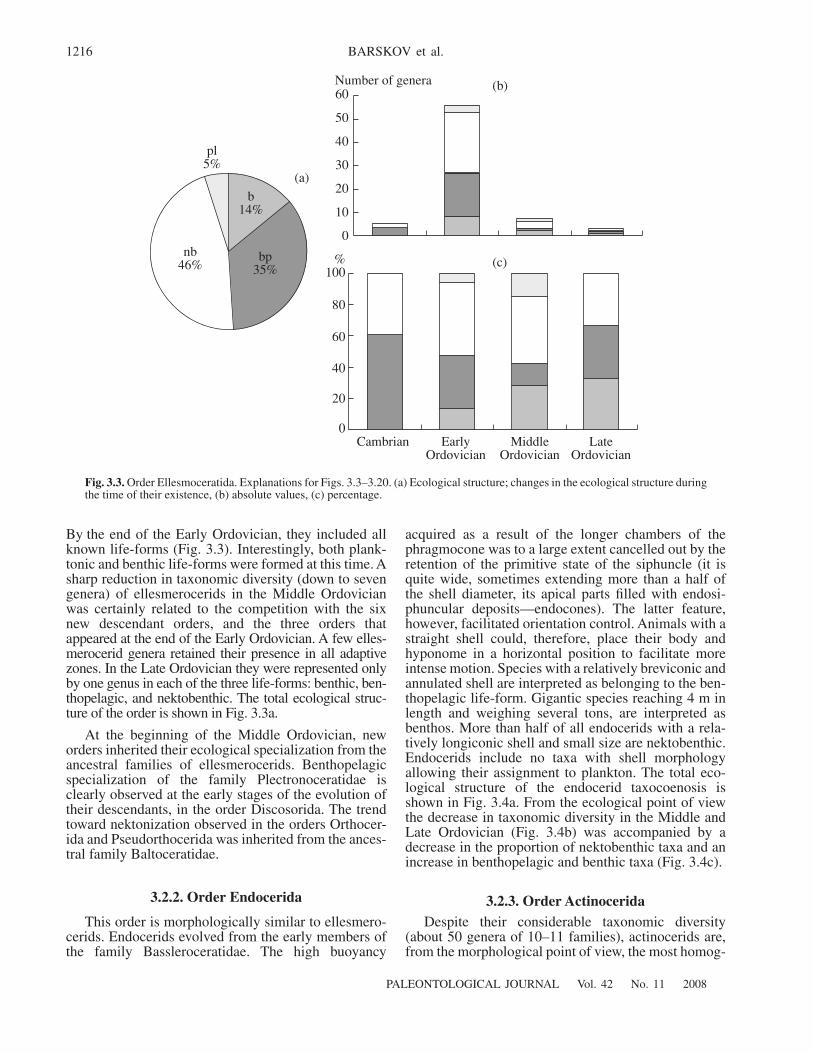
1216
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
By the end of the Early Ordovician, they included allknown life-forms (Fig. 3.3). Interestingly, both plank-tonic and benthic life-forms were formed at this time. Asharp reduction in taxonomic diversity (down to sevengenera) of ellesmerocerids in the Middle Ordovicianwas certainly related to the competition with the sixnew descendant orders, and the three orders thatappeared at the end of the Early Ordovician. A few elles-merocerid genera retained their presence in all adaptivezones. In the Late Ordovician they were represented onlyby one genus in each of the three life-forms: benthic, ben-thopelagic, and nektobenthic. The total ecological struc-ture of the order is shown in Fig. 3.3a.
At the beginning of the Middle Ordovician, neworders inherited their ecological specialization from theancestral families of ellesmerocerids. Benthopelagicspecialization of the family Plectronoceratidae isclearly observed at the early stages of the evolution oftheir descendants, in the order Discosorida. The trendtoward nektonization observed in the orders Orthocer-ida and Pseudorthocerida was inherited from the ances-tral family Baltoceratidae.
3.2.2. Order Endocerida
This order is morphologically similar to ellesmero-cerids. Endocerids evolved from the early members ofthe family Bassleroceratidae. The high buoyancy
acquired as a result of the longer chambers of thephragmocone was to a large extent cancelled out by theretention of the primitive state of the siphuncle (it isquite wide, sometimes extending more than a half ofthe shell diameter, its apical parts filled with endosi-phuncular deposits—endocones). The latter feature,however, facilitated orientation control. Animals with astraight shell could, therefore, place their body andhyponome in a horizontal position to facilitate moreintense motion. Species with a relatively breviconic andannulated shell are interpreted as belonging to the ben-thopelagic life-form. Gigantic species reaching 4 m inlength and weighing several tons, are interpreted asbenthos. More than half of all endocerids with a rela-tively longiconic shell and small size are nektobenthic.Endocerids include no taxa with shell morphologyallowing their assignment to plankton. The total eco-logical structure of the endocerid taxocoenosis isshown in Fig. 3.4a. From the ecological point of viewthe decrease in taxonomic diversity in the Middle andLate Ordovician (Fig. 3.4b) was accompanied by adecrease in the proportion of nektobenthic taxa and anincrease in benthopelagic and benthic taxa (Fig. 3.4c).
3.2.3. Order ActinoceridaDespite their considerable taxonomic diversity
(about 50 genera of 10–11 families), actinocerids are,from the morphological point of view, the most homog-
Cambrian Early Middle LateOrdovicianOrdovicianOrdovician
b14%
bp35%
nb46%
pl5%
60
50
40
30
20
10
0
%100
80
60
40
20
0
(a)
(b)
(c)
Fig. 3.3. Order Ellesmoceratida. Explanations for Figs. 3.3–3.20. (a) Ecological structure; changes in the ecological structure duringthe time of their existence, (b) absolute values, (c) percentage.
Number of genera
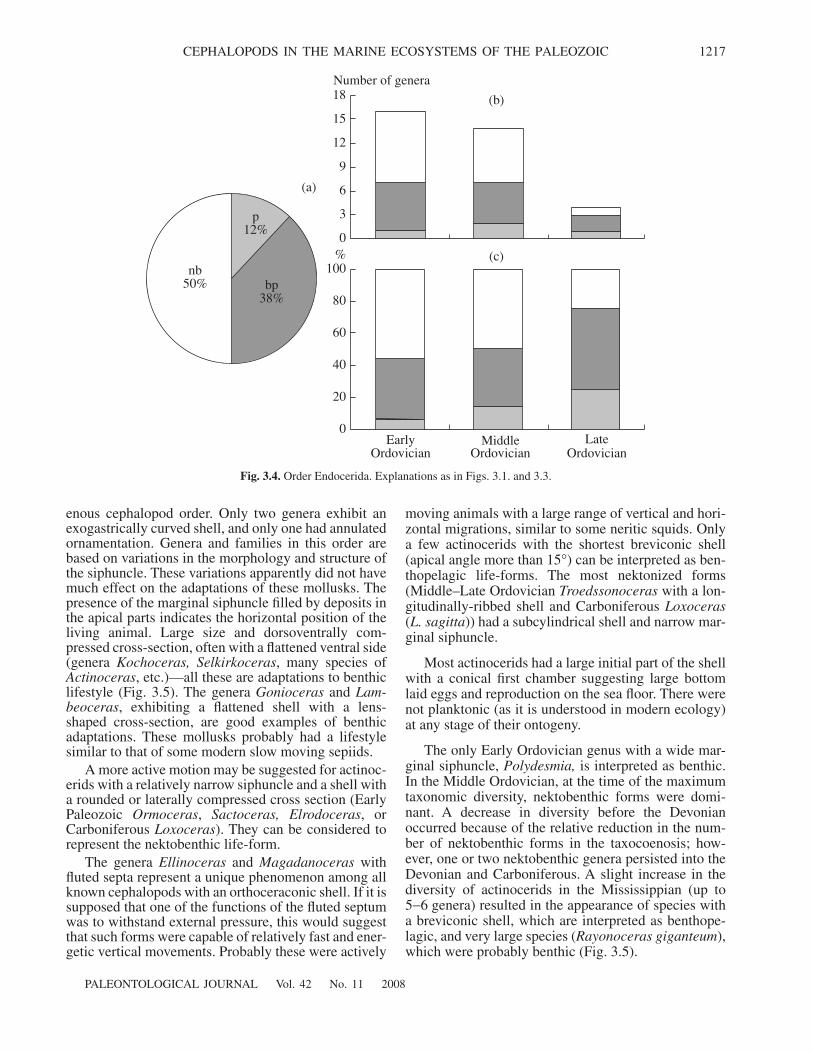
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1217
enous cephalopod order. Only two genera exhibit anexogastrically curved shell, and only one had annulatedornamentation. Genera and families in this order arebased on variations in the morphology and structure ofthe siphuncle. These variations apparently did not havemuch effect on the adaptations of these mollusks. Thepresence of the marginal siphuncle filled by deposits inthe apical parts indicates the horizontal position of theliving animal. Large size and dorsoventrally com-pressed cross-section, often with a flattened ventral side(genera
Kochoceras, Selkirkoceras
, many species of
Actinoceras
, etc.)—all these are adaptations to benthiclifestyle (Fig. 3.5). The genera
Gonioceras
and
Lam-beoceras
, exhibiting a flattened shell with a lens-shaped cross-section, are good examples of benthicadaptations. These mollusks probably had a lifestylesimilar to that of some modern slow moving sepiids.
A more active motion may be suggested for actinoc-erids with a relatively narrow siphuncle and a shell witha rounded or laterally compressed cross section (EarlyPaleozoic
Ormoceras
,
Sactoceras, Elrodoceras
, orCarboniferous
Loxoceras
). They can be considered torepresent the nektobenthic life-form.
The genera
Ellinoceras
and
Magadanoceras
withfluted septa represent a unique phenomenon among allknown cephalopods with an orthoceraconic shell. If it issupposed that one of the functions of the fluted septumwas to withstand external pressure, this would suggestthat such forms were capable of relatively fast and ener-getic vertical movements. Probably these were actively
moving animals with a large range of vertical and hori-zontal migrations, similar to some neritic squids. Onlya few actinocerids with the shortest breviconic shell(apical angle more than 15
°
) can be interpreted as ben-thopelagic life-forms. The most nektonized forms(Middle–Late Ordovician
Troedssonoceras
with a lon-gitudinally-ribbed shell and Carboniferous
Loxoceras
(
L. sagitta
)) had a subcylindrical shell and narrow mar-ginal siphuncle.
Most actinocerids had a large initial part of the shellwith a conical first chamber suggesting large bottomlaid eggs and reproduction on the sea floor. There werenot planktonic (as it is understood in modern ecology)at any stage of their ontogeny.
The only Early Ordovician genus with a wide mar-ginal siphuncle,
Polydesmia,
is interpreted as benthic.In the Middle Ordovician, at the time of the maximumtaxonomic diversity, nektobenthic forms were domi-nant. A decrease in diversity before the Devonianoccurred because of the relative reduction in the num-ber of nektobenthic forms in the taxocoenosis; how-ever, one or two nektobenthic genera persisted into theDevonian and Carboniferous. A slight increase in thediversity of actinocerids in the Mississippian (up to5
−
6 genera) resulted in the appearance of species witha breviconic shell, which are interpreted as benthope-lagic, and very large species (
Rayonoceras giganteum
),which were probably benthic (Fig. 3.5).
Early
18
15
12
9
6
3
0
100
80
60
40
20
0Late
OrdovicianMiddle
OrdovicianOrdovician
%
(b)
(c)
(a)
nb50% bp
38%
p12%
Fig. 3.4.
Order Endocerida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
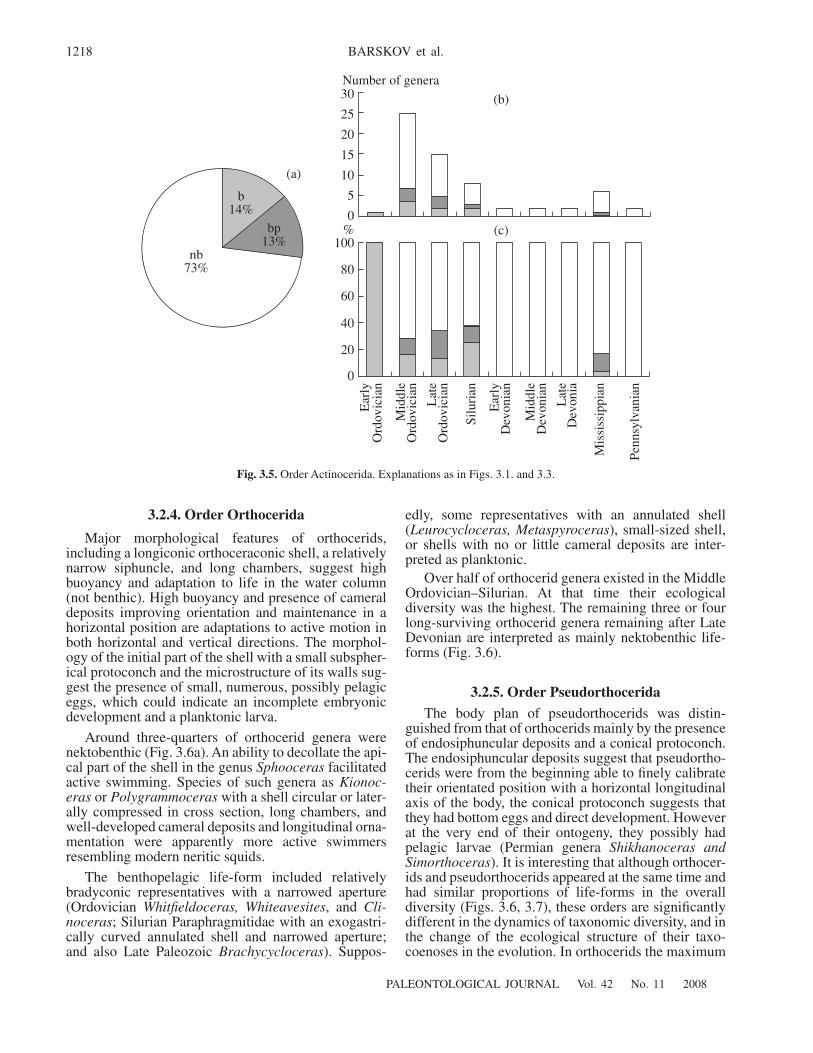
1218
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
3.2.4. Order Orthocerida
Major morphological features of orthocerids,including a longiconic orthoceraconic shell, a relativelynarrow siphuncle, and long chambers, suggest highbuoyancy and adaptation to life in the water column(not benthic). High buoyancy and presence of cameraldeposits improving orientation and maintenance in ahorizontal position are adaptations to active motion inboth horizontal and vertical directions. The morphol-ogy of the initial part of the shell with a small subspher-ical protoconch and the microstructure of its walls sug-gest the presence of small, numerous, possibly pelagiceggs, which could indicate an incomplete embryonicdevelopment and a planktonic larva.
Around three-quarters of orthocerid genera werenektobenthic (Fig. 3.6a). An ability to decollate the api-cal part of the shell in the genus
Sphooceras
facilitatedactive swimming. Species of such genera as
Kionoc-eras
or
Polygrammoceras
with a shell circular or later-ally compressed in cross section, long chambers, andwell-developed cameral deposits and longitudinal orna-mentation were apparently more active swimmersresembling modern neritic squids.
The benthopelagic life-form included relativelybradyconic representatives with a narrowed aperture(Ordovician
Whitfieldoceras, Whiteavesites
, and
Cli-noceras
; Silurian Paraphragmitidae with an exogastri-cally curved annulated shell and narrowed aperture;and also Late Paleozoic
Brachycycloceras
). Suppos-
edly, some representatives with an annulated shell(
Leurocycloceras, Metaspyroceras
), small-sized shell,or shells with no or little cameral deposits are inter-preted as planktonic.
Over half of orthocerid genera existed in the MiddleOrdovician–Silurian. At that time their ecologicaldiversity was the highest. The remaining three or fourlong-surviving orthocerid genera remaining after LateDevonian are interpreted as mainly nektobenthic life-forms (Fig. 3.6).
3.2.5. Order Pseudorthocerida
The body plan of pseudorthocerids was distin-guished from that of orthocerids mainly by the presenceof endosiphuncular deposits and a conical protoconch.The endosiphuncular deposits suggest that pseudortho-cerids were from the beginning able to finely calibratetheir orientated position with a horizontal longitudinalaxis of the body, the conical protoconch suggests thatthey had bottom eggs and direct development. Howeverat the very end of their ontogeny, they possibly hadpelagic larvae (Permian genera
Shikhanoceras andSimorthoceras
). It is interesting that although orthocer-ids and pseudorthocerids appeared at the same time andhad similar proportions of life-forms in the overalldiversity (Figs. 3.6, 3.7), these orders are significantlydifferent in the dynamics of taxonomic diversity, and inthe change of the ecological structure of their taxo-coenoses in the evolution. In orthocerids the maximum
b14%
bp13%
nb73%
(a)
(b)
(c)
30
25
20
15
10
5
0
100%
80
60
40
20
0 E
arly
Mid
dle
Penn
sylv
ania
n
Mid
dle
Lat
e
Silu
rian
Ear
ly
Lat
e
Mis
siss
ippi
an
Dev
onia
n
Dev
onia
Dev
onia
n
Ord
ovic
ian
Ord
ovic
ian
Ord
ovic
ian
Fig. 3.5.
Order Actinocerida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
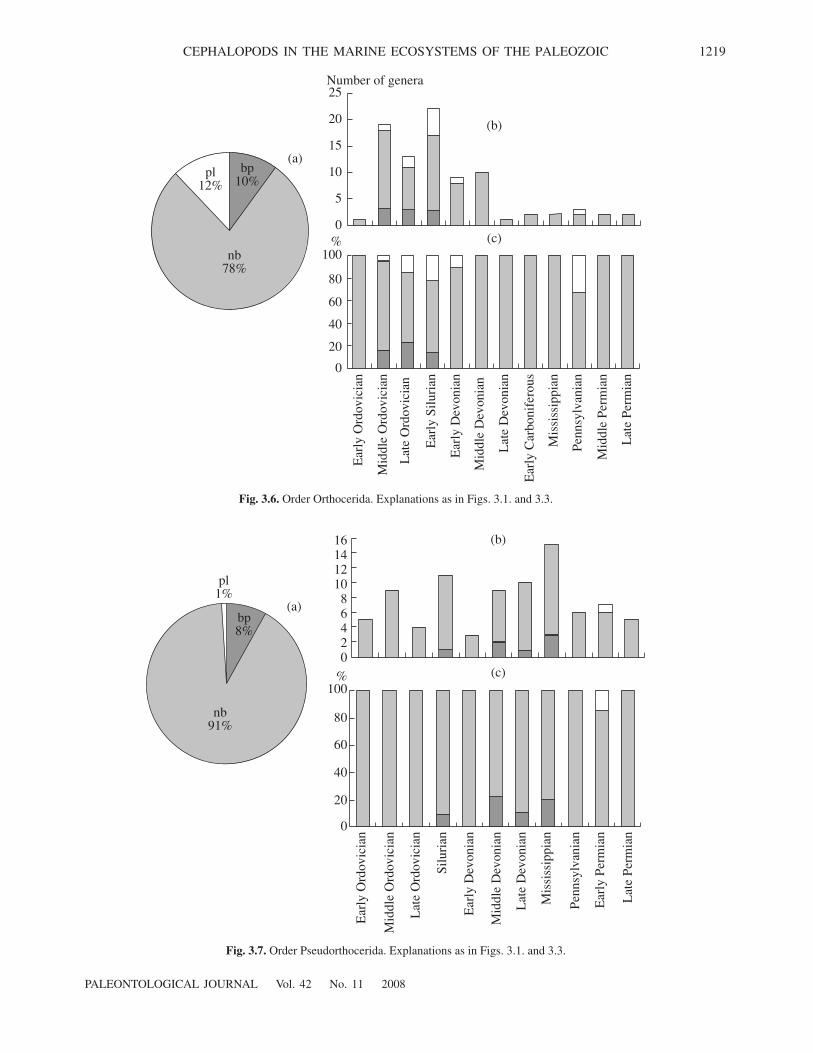
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1219
Ear
ly O
rdov
icia
n
Ear
ly S
iluri
an
Lat
e D
evon
ian
Penn
sylv
ania
n
25
20
15
10
5
0%
100
80
60
40
20
0M
iddl
e O
rdov
icia
n
Lat
e O
rdov
icia
n
Ear
ly D
evon
ian
Mid
dle
Dev
onia
n
Ear
ly C
arbo
nife
rous
Mis
siss
ippi
an
Mid
dle
Perm
ian
Lat
e Pe
rmia
n
(a)
(b)
(c)
bp10%
nb78%
pl12%
Fig. 3.6.
Order Orthocerida. Explanations as in Figs. 3.1. and 3.3.
Ear
ly O
rdov
icia
n
Mid
dle
Dev
onia
n
1614121086420
100
80
60
40
20
0
Mid
dle
Ord
ovic
ian
Lat
e O
rdov
icia
n
Silu
rian
Ear
ly D
evon
ian
Lat
e D
evon
ian
Mis
siss
ippi
an
Penn
sylv
ania
n
Ear
ly P
erm
ian
Lat
e Pe
rmia
n
(a)
(b)
(c)
pl1%
bp8%
nb91%
%
Fig. 3.7.
Order Pseudorthocerida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
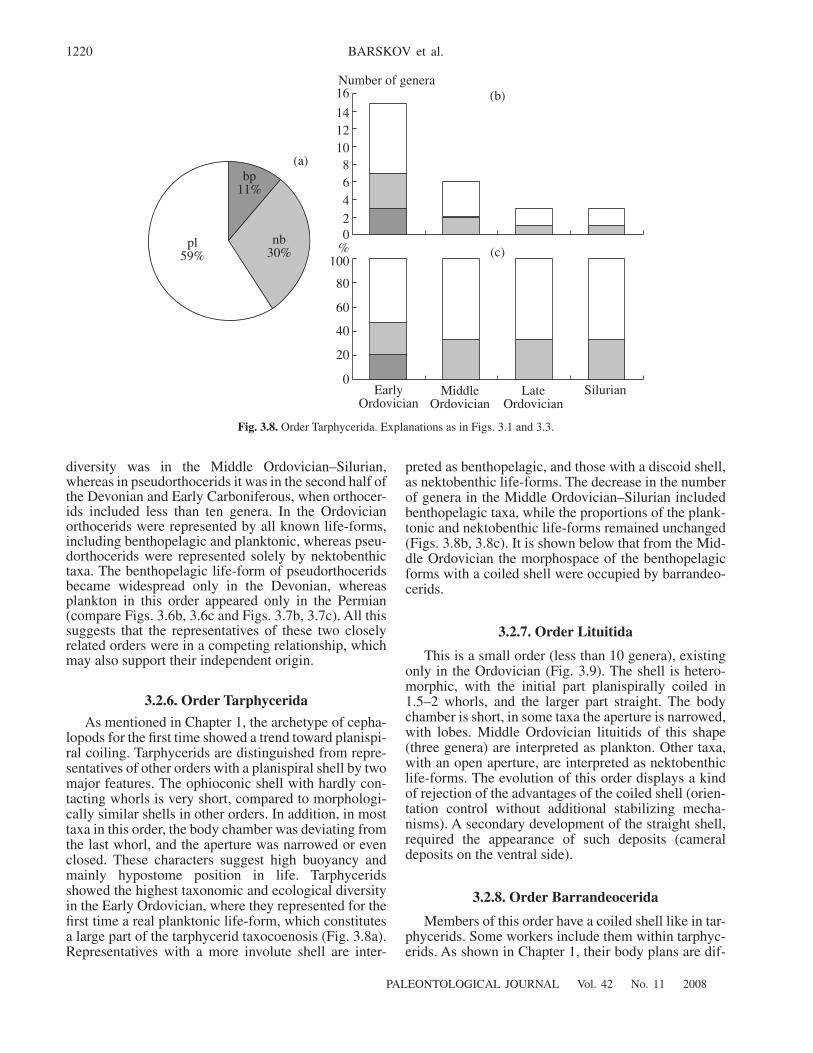
1220
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
diversity was in the Middle Ordovician–Silurian,whereas in pseudorthocerids it was in the second half ofthe Devonian and Early Carboniferous, when orthocer-ids included less than ten genera. In the Ordovicianorthocerids were represented by all known life-forms,including benthopelagic and planktonic, whereas pseu-dorthocerids were represented solely by nektobenthictaxa. The benthopelagic life-form of pseudorthoceridsbecame widespread only in the Devonian, whereasplankton in this order appeared only in the Permian(compare Figs. 3.6b, 3.6c and Figs. 3.7b, 3.7c). All thissuggests that the representatives of these two closelyrelated orders were in a competing relationship, whichmay also support their independent origin.
3.2.6. Order Tarphycerida
As mentioned in Chapter 1, the archetype of cepha-lopods for the first time showed a trend toward planispi-ral coiling. Tarphycerids are distinguished from repre-sentatives of other orders with a planispiral shell by twomajor features. The ophioconic shell with hardly con-tacting whorls is very short, compared to morphologi-cally similar shells in other orders. In addition, in mosttaxa in this order, the body chamber was deviating fromthe last whorl, and the aperture was narrowed or evenclosed. These characters suggest high buoyancy andmainly hypostome position in life. Tarphyceridsshowed the highest taxonomic and ecological diversityin the Early Ordovician, where they represented for thefirst time a real planktonic life-form, which constitutesa large part of the tarphycerid taxocoenosis (Fig. 3.8a).Representatives with a more involute shell are inter-
preted as benthopelagic, and those with a discoid shell,as nektobenthic life-forms. The decrease in the numberof genera in the Middle Ordovician–Silurian includedbenthopelagic taxa, while the proportions of the plank-tonic and nektobenthic life-forms remained unchanged(Figs. 3.8b, 3.8c). It is shown below that from the Mid-dle Ordovician the morphospace of the benthopelagicforms with a coiled shell were occupied by barrandeo-cerids.
3.2.7. Order Lituitida
This is a small order (less than 10 genera), existingonly in the Ordovician (Fig. 3.9). The shell is hetero-morphic, with the initial part planispirally coiled in1.5–2 whorls, and the larger part straight. The bodychamber is short, in some taxa the aperture is narrowed,with lobes. Middle Ordovician lituitids of this shape(three genera) are interpreted as plankton. Other taxa,with an open aperture, are interpreted as nektobenthiclife-forms. The evolution of this order displays a kindof rejection of the advantages of the coiled shell (orien-tation control without additional stabilizing mecha-nisms). A secondary development of the straight shell,required the appearance of such deposits (cameraldeposits on the ventral side).
3.2.8. Order Barrandeocerida
Members of this order have a coiled shell like in tar-phycerids. Some workers include them within tarphyc-erids. As shown in Chapter 1, their body plans are dif-
Early Middle Late SilurianOrdovicianOrdovicianOrdovician
16
14121086420
%100
80
60
40
20
0
(a)
(b)
(c)
bp11%
nb30%
pl59%
Fig. 3.8.
Order Tarphycerida. Explanations as in Figs. 3.1 and 3.3.
Number of genera
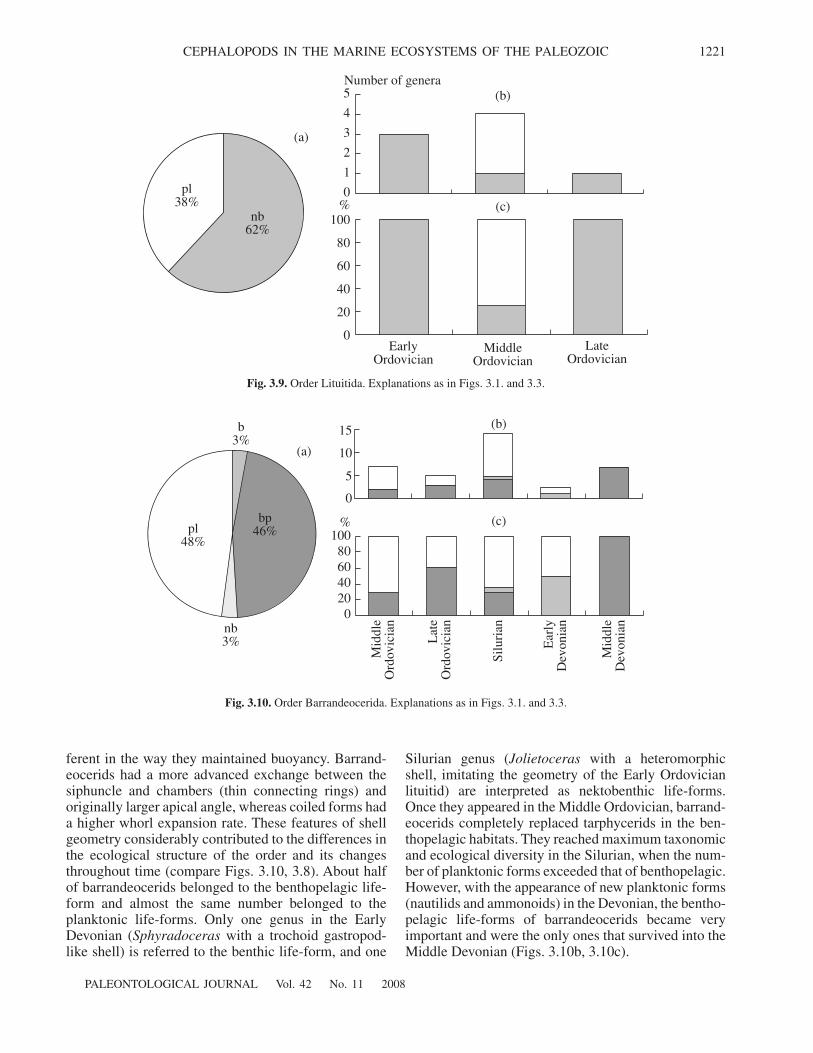
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1221
ferent in the way they maintained buoyancy. Barrand-eocerids had a more advanced exchange between thesiphuncle and chambers (thin connecting rings) andoriginally larger apical angle, whereas coiled forms hada higher whorl expansion rate. These features of shellgeometry considerably contributed to the differences inthe ecological structure of the order and its changesthroughout time (compare Figs. 3.10, 3.8). About halfof barrandeocerids belonged to the benthopelagic life-form and almost the same number belonged to theplanktonic life-forms. Only one genus in the EarlyDevonian (
Sphyradoceras
with a trochoid gastropod-like shell) is referred to the benthic life-form, and one
Silurian genus (
Jolietoceras
with a heteromorphicshell, imitating the geometry of the Early Ordovicianlituitid) are interpreted as nektobenthic life-forms.Once they appeared in the Middle Ordovician, barrand-eocerids completely replaced tarphycerids in the ben-thopelagic habitats. They reached maximum taxonomicand ecological diversity in the Silurian, when the num-ber of planktonic forms exceeded that of benthopelagic.However, with the appearance of new planktonic forms(nautilids and ammonoids) in the Devonian, the bentho-pelagic life-forms of barrandeocerids became veryimportant and were the only ones that survived into theMiddle Devonian (Figs. 3.10b, 3.10c).
Early Middle LateOrdovicianOrdovicianOrdovician
5
4
3
2
1
0
100%
80
60
40
20
0
(b)
(c)
(a)
pl38%
nb62%
Fig. 3.9.
Order Lituitida. Explanations as in Figs. 3.1. and 3.3.
Mid
dle
Lat
e
Silu
rian
Ear
ly
Mid
dle
Dev
onia
n
Dev
onia
n
Ord
ovic
ian
Ord
ovic
ian
15
10
5
0
%100806040200
b3%
bp46%
nb3%
pl48%
(a)
(b)
(c)
Fig. 3.10.
Order Barrandeocerida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
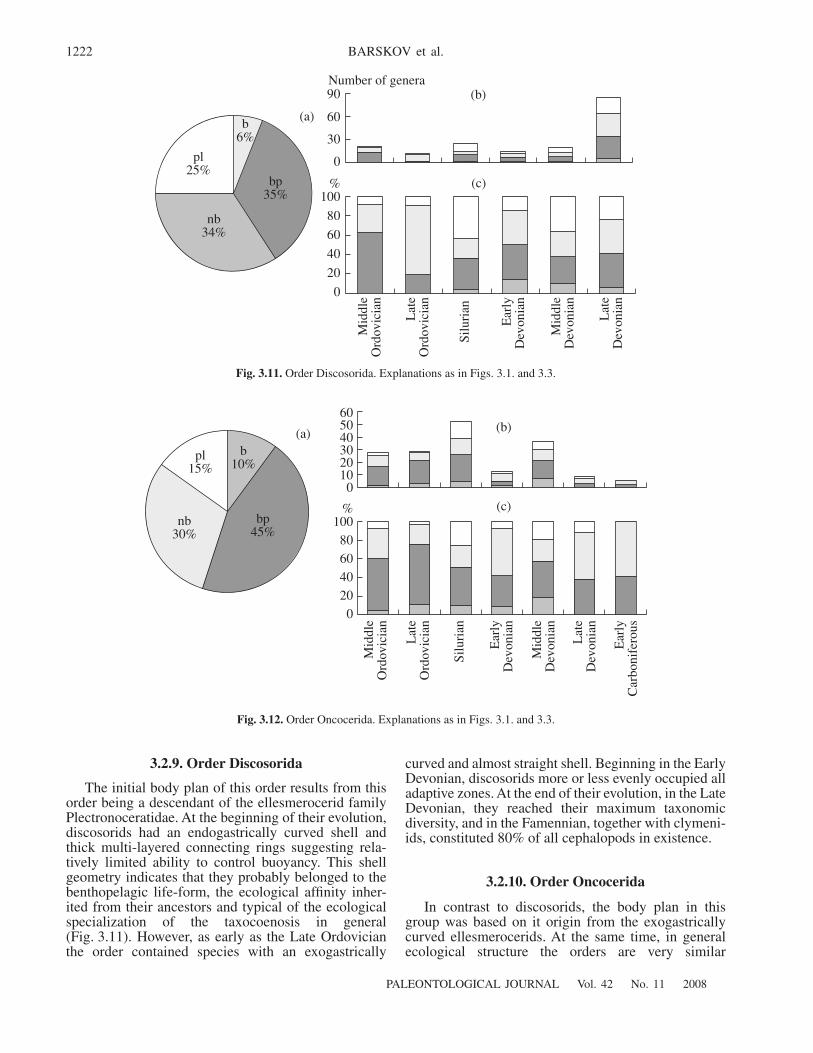
1222
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
3.2.9. Order Discosorida
The initial body plan of this order results from thisorder being a descendant of the ellesmerocerid familyPlectronoceratidae. At the beginning of their evolution,discosorids had an endogastrically curved shell andthick multi-layered connecting rings suggesting rela-tively limited ability to control buoyancy. This shellgeometry indicates that they probably belonged to thebenthopelagic life-form, the ecological affinity inher-ited from their ancestors and typical of the ecologicalspecialization of the taxocoenosis in general(Fig. 3.11). However, as early as the Late Ordovicianthe order contained species with an exogastrically
curved and almost straight shell. Beginning in the EarlyDevonian, discosorids more or less evenly occupied alladaptive zones. At the end of their evolution, in the LateDevonian, they reached their maximum taxonomicdiversity, and in the Famennian, together with clymeni-ids, constituted 80% of all cephalopods in existence.
3.2.10. Order Oncocerida
In contrast to discosorids, the body plan in thisgroup was based on it origin from the exogastricallycurved ellesmerocerids. At the same time, in generalecological structure the orders are very similar
Mid
dle
Ear
ly
90
60
30
0
%100
80
60
40
20
0
(b)
(c)
Lat
e
Silu
rian
Mid
dle
Lat
eD
evon
ian
Dev
onia
n
Dev
onia
n
Ord
ovic
ian
Ord
ovic
ian
(a)b
6%
bp35%
nb34%
pl25%
Fig. 3.11.
Order Discosorida. Explanations as in Figs. 3.1. and 3.3.
b10%
bp45%
nb30%
pl15%
Mid
dle
6050403020100
%100806040200
Lat
e
Silu
rian
Ear
ly
Mid
dle
Lat
e
Ear
lyC
arbo
nife
rous
Dev
onia
n
Dev
onia
n
Dev
onia
n
Ord
ovic
ian
Ord
ovic
ian
(a) (b)
(c)
Fig. 3.12.
Order Oncocerida. Explanations as in Figs. 3.1. and 3.3.
Number of genera

PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1223
(Figs. 3.11, 3.12) and supplemented each other by theproportions of life-forms throughout their co-existence.Until the Late Devonian, the generic diversity of onco-cerids was more than that of discosorids. In the secondhalf of the Silurian forms appeared among both onco-cerids and discosorids exhibiting a new, peculiar plank-tonic morphotype with a barrel-shaped shell and closedaperture (oncocerid families Hemiphragmoceratidaeand Trimeroceratidae, and discosorid families Phrag-moceratidae, Mandaloceratidae, and Mesoceratidae).Another example of such parallel emergence of formswith similar shell geometry in both orders was theappearance of nektobenthic life-forms with an almoststraight shell in the Late Devonian
3.2.11. Order Ascocerida
This is a small order, represented by one and a halfdozen genera, the main feature of the body plan ofwhich was a natural truncation of the posterior end ofthe orthoconic phragmocone and development at thefinal orthogenetic stages of an ellipsoidal, sausage-shaped, or barrel-shaped shell with chambers on theposterior and dorsal sides. In these forms, the phragmo-cone acted as a passive floating device. The membersrepresented two life-forms: (1) nektobenthic, whichcharacteristically displayed an open aperture and hori-zontal position of the longitudinal body axis (forinstance, the Ordovician
Probillingsites,
or Silurian
Ascoceras
) and (2) planktonic, shell of which had a nar-rowed aperture and was inclined or of hypostome ori-entation (the Ordovician
Billingsites, Schuchertoceras
,or Silurian
Glossoceras
and
Aphragmites
). The propor-tions of these life-forms in the community remained virtu-ally the same throughout their existence (Fig. 3.13).
3.2.12. Order Nautilida
The basic geometry of nautilids is a planispiralshell, but in the modern system the order includes sev-eral genera with a trochoid, curved shell (Middle Devo-nian
Rutoceras
,
Centrolitoceras,
and others and theEarly Permian
Sphooceras
) and even with a straightshell (
Casteroceras
).
Nautilids originally appeared in the Early Devonianas benthopelagic and planktonic forms. Later the groupcontained members of the nektobenthic life-form andeven benthic representatives. In general the ecologicalstructure of the order contains planktonic, benthope-lagic, and nektobenthic life-forms, with their propor-tions considerably changing throughout ecogenesis. Inthe evolution, the proportion of the planktonic life-formdecreases, whereas the proportions of nektobenthic andbenthopelagic life-forms increased (Fig. 3.14). To alarge extent, the order of development was determinedby competition with ammonoids, which appeared in theEarly Devonian and were a large group with a planispi-ral shell. In the pre-Permian epochs, especially in theCarboniferous, most nautilids were represented by nek-tobenthic and benthopelagic forms with a morphotypeof semi-evolute moderately or rapidly expanding shells(morphogroup 2N) (Fig. 3.14c). Among ammonoidssuch forms appear and predominate in the second halfof the Permian. From that time onward, the proportionof benthopelagic forms with a morphotype of the mod-ern
Nautilus
began to increase among nautilids, appar-ently because of the replacement by ammonoids,whereas ammonoids in this adaptive zone were repre-sented by another morphotype: spheroconic and cadi-conic shells with slowly expanding whorls.
nb54%
pl46%
Middle
8
6
4
2
0
%100
80
60
40
20
0Late Silurian
OrdovicianOrdovician
(a)
(b)
(c)
Fig. 3.13.
Order Ascocerida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
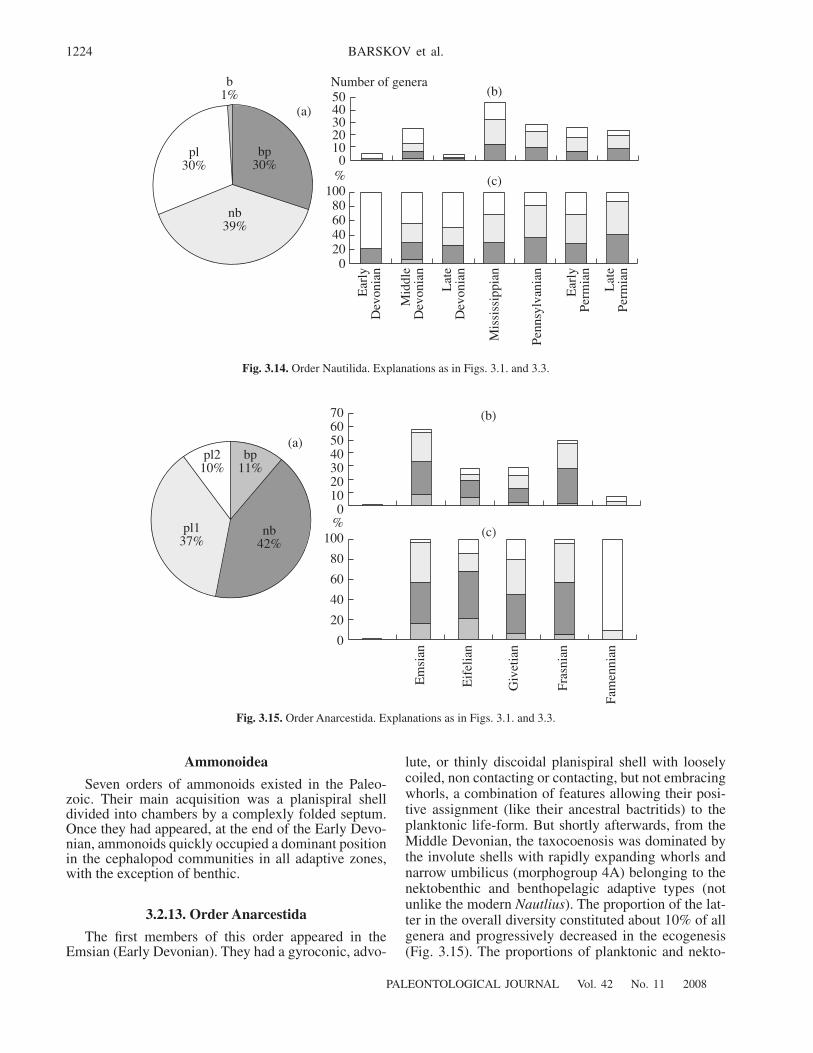
1224
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
BARSKOV et al.
Ammonoidea
Seven orders of ammonoids existed in the Paleo-zoic. Their main acquisition was a planispiral shelldivided into chambers by a complexly folded septum.Once they had appeared, at the end of the Early Devo-nian, ammonoids quickly occupied a dominant positionin the cephalopod communities in all adaptive zones,with the exception of benthic.
3.2.13. Order Anarcestida
The first members of this order appeared in theEmsian (Early Devonian). They had a gyroconic, advo-
lute, or thinly discoidal planispiral shell with looselycoiled, non contacting or contacting, but not embracingwhorls, a combination of features allowing their posi-tive assignment (like their ancestral bactritids) to theplanktonic life-form. But shortly afterwards, from theMiddle Devonian, the taxocoenosis was dominated bythe involute shells with rapidly expanding whorls andnarrow umbilicus (morphogroup 4A) belonging to thenektobenthic and benthopelagic adaptive types (notunlike the modern
Nautlius
). The proportion of the lat-ter in the overall diversity constituted about 10% of allgenera and progressively decreased in the ecogenesis(Fig. 3.15). The proportions of planktonic and nekto-
Ear
ly
Lat
e
50403020100
100806040200
%
Lat
e
Mid
dle
Mis
siss
ippi
an
Penn
sylv
ania
n
Ear
ly
Perm
ian
Perm
ian
Dev
onia
n
Dev
onia
n
Dev
onia
n
(a)
(b)
(c)
b1%
bp30%
nb39%
pl30%
Fig. 3.14.
Order Nautilida. Explanations as in Figs. 3.1. and 3.3.
pl210%
bp11%
nb42%
pl137%
706050403020100
%100
80
60
40
20
0
Em
sian
Eif
elia
n
Giv
etia
n
Fras
nian
Fam
enni
an
(a)
(b)
(c)
Fig. 3.15.
Order Anarcestida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
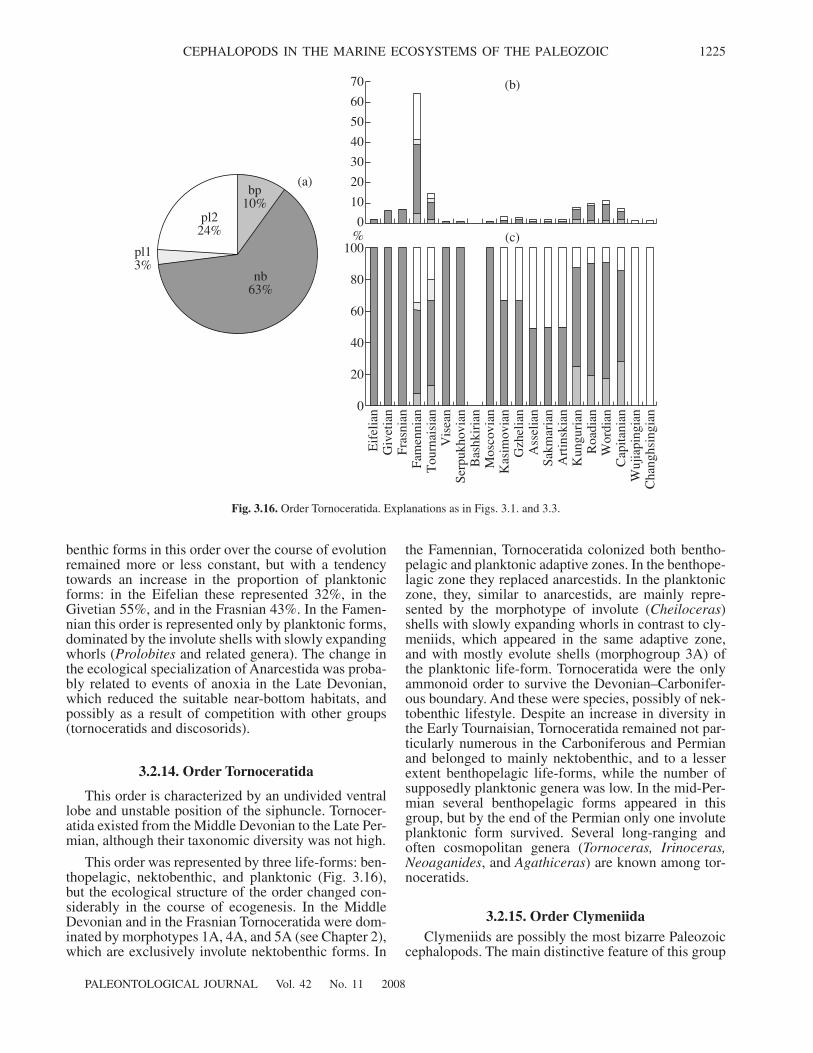
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 11
2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1225
benthic forms in this order over the course of evolutionremained more or less constant, but with a tendencytowards an increase in the proportion of planktonicforms: in the Eifelian these represented 32%, in theGivetian 55%, and in the Frasnian 43%. In the Famen-nian this order is represented only by planktonic forms,dominated by the involute shells with slowly expandingwhorls (
Prolobites
and related genera). The change inthe ecological specialization of Anarcestida was proba-bly related to events of anoxia in the Late Devonian,which reduced the suitable near-bottom habitats, andpossibly as a result of competition with other groups(tornoceratids and discosorids).
3.2.14. Order Tornoceratida
This order is characterized by an undivided ventrallobe and unstable position of the siphuncle. Tornocer-atida existed from the Middle Devonian to the Late Per-mian, although their taxonomic diversity was not high.
This order was represented by three life-forms: ben-thopelagic, nektobenthic, and planktonic (Fig. 3.16),but the ecological structure of the order changed con-siderably in the course of ecogenesis. In the MiddleDevonian and in the Frasnian Tornoceratida were dom-inated by morphotypes 1A, 4A, and 5A (see Chapter 2),which are exclusively involute nektobenthic forms. In
the Famennian, Tornoceratida colonized both bentho-pelagic and planktonic adaptive zones. In the benthope-lagic zone they replaced anarcestids. In the planktoniczone, they, similar to anarcestids, are mainly repre-sented by the morphotype of involute (
Cheiloceras
)shells with slowly expanding whorls in contrast to cly-meniids, which appeared in the same adaptive zone,and with mostly evolute shells (morphogroup 3A) ofthe planktonic life-form. Tornoceratida were the onlyammonoid order to survive the Devonian–Carbonifer-ous boundary. And these were species, possibly of nek-tobenthic lifestyle. Despite an increase in diversity inthe Early Tournaisian, Tornoceratida remained not par-ticularly numerous in the Carboniferous and Permianand belonged to mainly nektobenthic, and to a lesserextent benthopelagic life-forms, while the number ofsupposedly planktonic genera was low. In the mid-Per-mian several benthopelagic forms appeared in thisgroup, but by the end of the Permian only one involuteplanktonic form survived. Several long-ranging andoften cosmopolitan genera (
Tornoceras, Irinoceras,Neoaganides
, and
Agathiceras
) are known among tor-noceratids.
3.2.15. Order ClymeniidaClymeniids are possibly the most bizarre Paleozoic
cephalopods. The main distinctive feature of this group
Eif
elia
n
Ass
elia
n
70
60
50
40
30
20
10
0
100%
80
60
40
20
0
bp10%
nb63%
pl224%
pl13%
Giv
etia
nFr
asni
anFa
men
nian
Tou
rnai
sian
Vis
ean
Serp
ukho
vian
Bas
hkir
ian
Mos
covi
anK
asim
ovia
nG
zhel
ian
Sakm
aria
nA
rtin
skia
nK
ungu
rian
Roa
dian
Wor
dian
Cap
itani
anW
ujia
ping
ian
Cha
nghs
ingi
an
(a)
(b)
(c)
Fig. 3.16. Order Tornoceratida. Explanations as in Figs. 3.1. and 3.3.

1226
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
is their dorsal siphuncle. They lived in the second halfof the Famennian. This order is characterized by anextremely high evolutionary rate (over 70 generaappeared and became extinct in about 10 million years)and a clear-cut planktonic specialization (74% of cly-meniids genera likely to have been planktonic)(Fig. 3.17). It is possible that the first clymeniids wereplanktonic, while nektobenthic and some benthope-lagic forms appeared later. Most planktonic formsbelonged to morphogroup 3A, whereas others are rep-resented by involute, slowly expanding shells, amongwhich there are some bizarre shells with inflated whorlsand triangular coiling (Wocklumeria, Synwocklumeria).Nektobenthic forms, mainly semi-involute, rapidlyexpanding shells forms (morphogroups 5A, 6A) consti-tuted 24% of the generic diversity in the order, whereasbenthopelagic forms are represented by only 2 genera.Because of their ecological specificity clymeniids oftenoccupied the deep shelf. Their maximum diversity wasaround the levels marking global transgressions. Theproportion of planktonic forms increased in the courseof evolution of this order reaching 100% at the end ofthe Famennian.
As mentioned above, one of the possible reasons forthe increasing proportions of planktonic forms,observed in the Famennian in all ammonoid orders,could be anoxic events affecting the near-bottom zonesof marine basins. Repeated large-scale anoxic events(at least four) are documented in the Famennian sec-tions by deposition of black shale (Becker, 1993). Thelast (Hangenberg) event if was not the main reason,contributed greatly to the extinction of mostammonoids and many other organisms at the Devo-nian–Carboniferous boundary.
3.2.16. Order Praeglyphioceratida
A few representatives (about 10 genera) of this orderare known from the Famennian and Tournaisian. In theDevonian they were characterized by only nektobenthicforms, whereas in the Tournaisian, while most genera
were nektobenthic, some (e.g., Kazakhstania) had evo-lute shells, which may suggest that these belonged tothe planktonic life-form.
3.2.17. Order Goniatitida
Goniatitida is the most numerous and morphologi-cally diverse order of Paleozoic cephalopods with aplanispiral shell. Ecologically it contains nektobenthos(37%), plankton (35%), and benthopelagic forms(28%) (Fig. 3.18). From the time of their appearance inthe Middle Tournaisian and during the entire Carbonif-erous and most of the Permian goniatitids played adominant role in the cephalopod communities, deter-mining their morphological and ecological structure.The first representative of the order, genus Goniacy-clus, is interpreted as nektobenthic, but numerous anddiverse genera appeared almost at the same time, andare interpreted as belonging to the benthopelagic life-forms or to somewhat late appearing planktonic forms.The main feature of the ecological structure of the orderis high taxonomic diversity of benthopelagic forms.The structure of the order changed significantly in thecourse of its evolution (Fig. 3.18). The main trend wastowards a gradual decrease in the diversity of nekto-benthic genera (45% in the Early Carboniferous, 25%at the end of the Permian) due to the increase in the pro-portions of planktonic and benthopelagic forms. Themost significant decrease in number of nektobenthicgoniatitids is recorded for the Early and Late Permian.Partly, this could be related to the competition of otherammonoid groups: prolecanitids in the Early Permian(their number increased from 12 genera in the Late Car-boniferous to 24 genera in the Early Permian), and cer-atitids in the Late Permian.
3.2.18. Order Prolecanitida
Prolecanitids appeared in the Early Tournaisian,almost immediately after a major extinction at theDevonian–Carboniferous boundary. This ammonoidgroup is characterized by a unique type of increasedsutural complexity and a distinctive trend in the evolu-tion of shell morphology. The first prolecanitids had anevolute, widely umbilicate shell with flattened flanks,which in the course of evolution became more andmore involute and platyconic. At the same time thewhorl expansion rate increased, which indicates achange in the ecological specialization of the order(Fig. 3.19) from mainly planktonic in the Early Carbon-iferous to nektobenthic in the Late Carboniferous andEarly Permian, when they were the most diverse. Atthat time specific platyconic nektobenthic forms withventrolateral keels and a number of other charactersappeared. A complex, strongly fluted septum typical ofthis group suggests the ability of the animal to quicklychange its buoyancy, and, in addition, it served to increasethe strength of the shell allowing it to exist at various
bp2%
nb24%
pl164%
pl210%
Fig. 3.17. Order Clymeniida. Explanations as in Figs. 3.1.and 3.3.
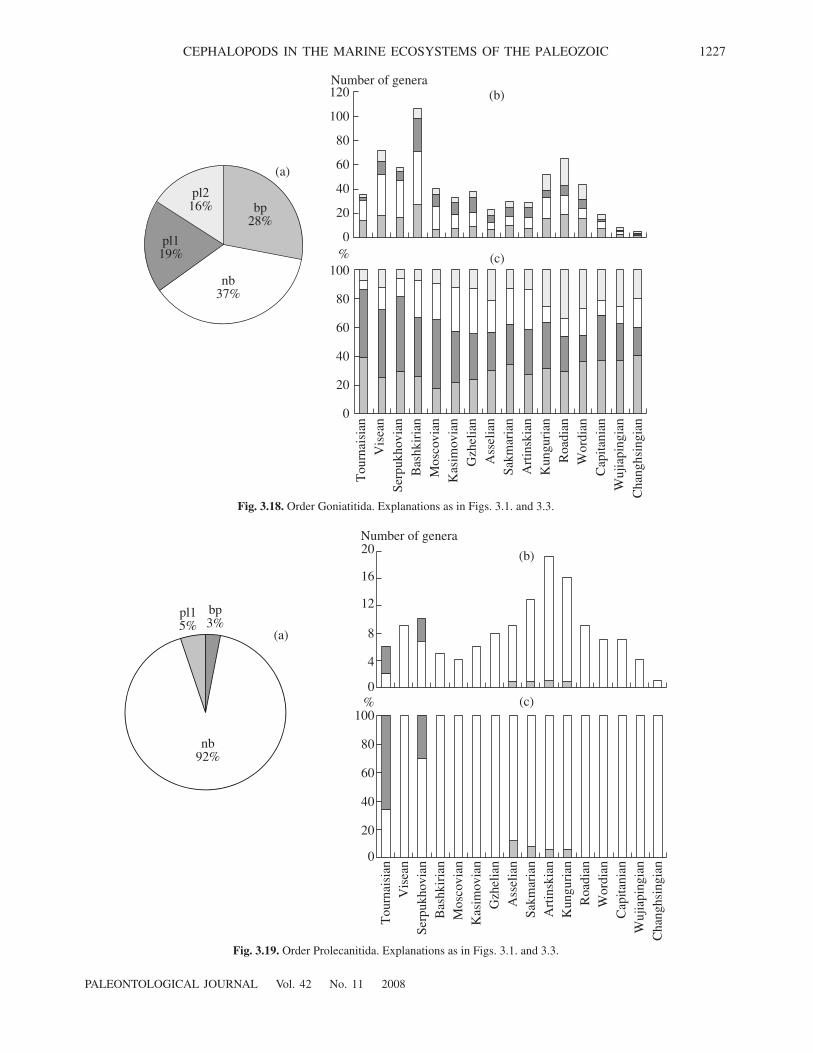
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1227
Tou
rnai
sian
Mos
covi
an
Sakm
aria
n
Wor
dian
bp28%
nb37%
pl216%
pl119%
120
100
80
60
40
20
0
100
80
60
40
20
0
%
Vis
ean
Serp
ukho
vian
Bas
hkir
ian
Kas
imov
ian
Gzh
elia
nA
ssel
ian
Art
insk
ian
Kun
guri
anR
oadi
an
Cap
itani
anW
ujia
ping
ian
Cha
nghs
ingi
an
(a)
(b)
(c)
Fig. 3.18. Order Goniatitida. Explanations as in Figs. 3.1. and 3.3.
Tou
rnai
sian
20
16
12
8
4
0%
100
80
60
40
20
0
Vis
ean
Serp
ukho
vian
Bas
hkir
ian
Mos
covi
anK
asim
ovia
nG
zhel
ian
Ass
elia
nSa
kmar
ian
Art
insk
ian
Kun
guri
anR
oadi
anW
ordi
anC
apita
nian
Wuj
iapi
ngia
nC
hang
hsin
gian
bp3%
nb92%
pl15%
(a)
(b)
(c)
Fig. 3.19. Order Prolecanitida. Explanations as in Figs. 3.1. and 3.3.
Number of genera
Number of genera
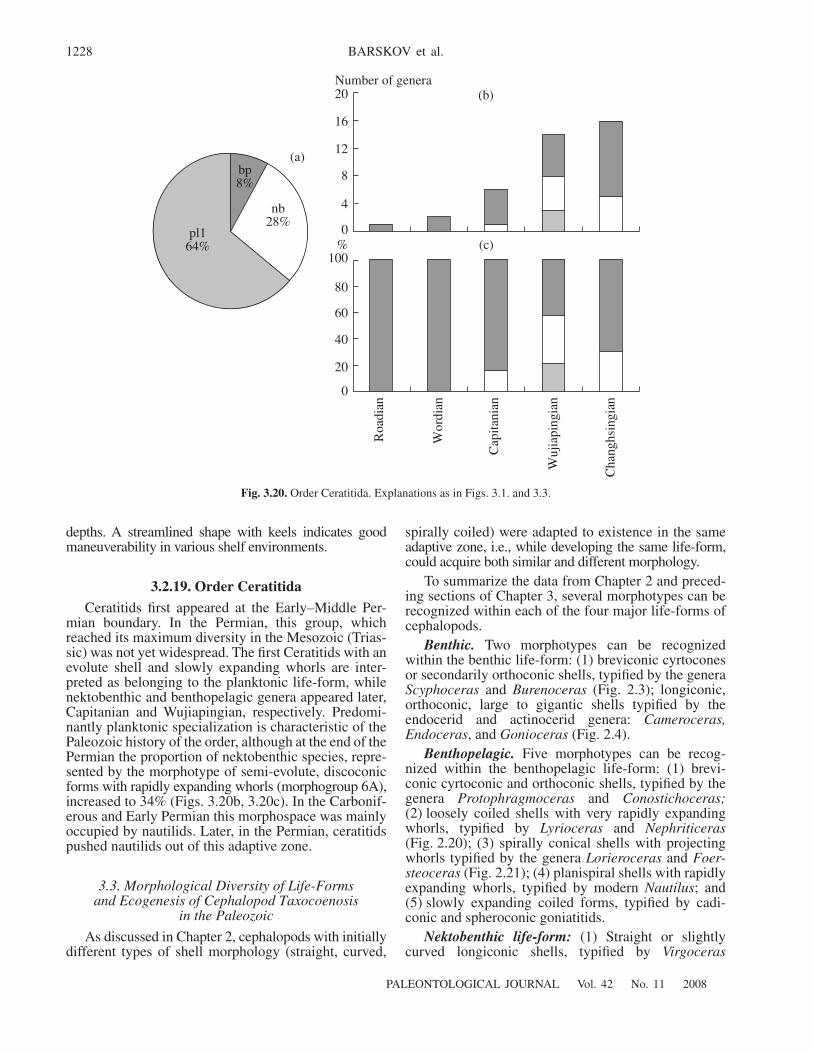
1228
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
depths. A streamlined shape with keels indicates goodmaneuverability in various shelf environments.
3.2.19. Order Ceratitida
Ceratitids first appeared at the Early–Middle Per-mian boundary. In the Permian, this group, whichreached its maximum diversity in the Mesozoic (Trias-sic) was not yet widespread. The first Ceratitids with anevolute shell and slowly expanding whorls are inter-preted as belonging to the planktonic life-form, whilenektobenthic and benthopelagic genera appeared later,Capitanian and Wujiapingian, respectively. Predomi-nantly planktonic specialization is characteristic of thePaleozoic history of the order, although at the end of thePermian the proportion of nektobenthic species, repre-sented by the morphotype of semi-evolute, discoconicforms with rapidly expanding whorls (morphogroup 6A),increased to 34% (Figs. 3.20b, 3.20c). In the Carbonif-erous and Early Permian this morphospace was mainlyoccupied by nautilids. Later, in the Permian, ceratitidspushed nautilids out of this adaptive zone.
3.3. Morphological Diversity of Life-Forms and Ecogenesis of Cephalopod Taxocoenosis
in the Paleozoic
As discussed in Chapter 2, cephalopods with initiallydifferent types of shell morphology (straight, curved,
spirally coiled) were adapted to existence in the sameadaptive zone, i.e., while developing the same life-form,could acquire both similar and different morphology.
To summarize the data from Chapter 2 and preced-ing sections of Chapter 3, several morphotypes can berecognized within each of the four major life-forms ofcephalopods.
Benthic. Two morphotypes can be recognizedwithin the benthic life-form: (1) breviconic cyrtoconesor secondarily orthoconic shells, typified by the generaScyphoceras and Burenoceras (Fig. 2.3); longiconic,orthoconic, large to gigantic shells typified by theendocerid and actinocerid genera: Cameroceras,Endoceras, and Gonioceras (Fig. 2.4).
Benthopelagic. Five morphotypes can be recog-nized within the benthopelagic life-form: (1) brevi-conic cyrtoconic and orthoconic shells, typified by thegenera Protophragmoceras and Conostichoceras;(2) loosely coiled shells with very rapidly expandingwhorls, typified by Lyrioceras and Nephriticeras(Fig. 2.20); (3) spirally conical shells with projectingwhorls typified by the genera Lorieroceras and Foer-steoceras (Fig. 2.21); (4) planispiral shells with rapidlyexpanding whorls, typified by modern Nautilus; and(5) slowly expanding coiled forms, typified by cadi-conic and spheroconic goniatitids.
Nektobenthic life-form: (1) Straight or slightlycurved longiconic shells, typified by Virgoceras
Roa
dian
bp8%
nb28%
pl164%
20
16
12
8
4
0%
100
80
60
40
20
0
Wor
dian
Cap
itani
an
Wuj
iapi
ngia
n
Cha
nghs
ingi
an
(a)
(b)
(c)
Fig. 3.20. Order Ceratitida. Explanations as in Figs. 3.1. and 3.3.
Number of genera

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1229
(Fig. 2.4); (2) shells with a dorsal gas-filled chambers,typified Ascoceras (Fig. 2.23); (3) shells with trochoid,non-projecting whorls, typified by Cumingoceras andLeurotrochoceras (Fig. 2.21); (4) Planispiral shellmoderately evolute with moderately rapidly expandingwhorls, usually with well-developed ornamentation—morphogroup 6A, 2N (Figs. 2.12, 2.18); and (5) planispi-ral involute discoconic and pachyconic shells, with rap-idly and moderately expanding whorls, smoothammonoids—morphogroups 1A, 4A (Fig. 2.14).
Planktonic life-form: (1) Cyrtoconic or secondarilyorthoconic breviconic shell with a narrowed or closedaperture, typified by Phragmoceras, Pentameroceras,or Cinctoceras (Fig. 2.3); (2) longiconic, straight orweakly curved shells without mechanism of orientationcontrol, typified by Bactrites (Fig. 2.4); (3) planispiralshell evolute, loosely coiled (typified by Estoniocerasand Bickmorites) or tightly coiled, slowly expandingwhorls—morphogroups 3A, 3N (Figs. 2.10, 2.16,2.20); and (4) planispiral involute shell with slowlyexpanding whorls, morphogroup 1A` (Fig. 2.14).
Below the ecological structure of the cephalopodcommunity is discussed for each stage of the Paleozoic.
About 100 cephalopod genera are known from theEarly Ordovician; half of these were ellesmerocerids,about 20% are tarphycerids, and about 15% endocerids.Another four orders: orthocerids, actinocerids, pseu-dorthocerids, and lituitids were represented by only afew genera. The ecological structure of the cephalopodcommunity (Fig. 3.21) was mainly composed of taxa ofthe former three orders (show in Fig. 3.21).
Nektobenthic forms constituted about half of thetotal taxocoenosis and were the most diverse taxonom-ically: this adaptive zone was populated by all thenexisting orders, with ellesmerocerids representing overhalf of the total number of taxa, while 20% belonged toendocerids, and the number of genera of the remainingfive orders constituted only a third of the total diversity.The morphotype of straight longiconic shells domi-nated, while only five genera were coiled tarphycerids.About 30% of the taxocoenosis was represented bybenthopelagic life-forms, which included three orders,
11%pl
31%bp
12%b
46%nb
Ellesme-
Tarphycerida
rocerida
Ellesmerocerida
Ellesmerocerida
Ellesmerocerida
Tarphycerida
Tarphy-cerida
Tarphy-cerida
Endo-cerida
Endo-cerida
Endo-cerida
Fig. 3.21. Ecological structure of cephalopods in the Early Ordovician. Small diagrams show the taxonomic structure of the life-form (for Figs. 3.21–3.32). Explanations: (nb) nektobenthic, (bp) benthopelagic, (b) benthic, (pl) planktonic life-forms.

1230
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
with three different morphotypes. Planktonic andbenthic forms each represented approximately one-tenth of the content of the taxocoenosis. Of ten plank-tonic genera, eight belonged to the orders Tarphyceridaand two to Ellesmerocerida. The benthic life-formsincluded ellesmerocerids (nine genera) and endocerids.
Middle Ordovician. About 140 genera are recordedfrom the Middle Ordovician. The taxonomic composi-tion of the assemblage is fundamentally different fromthat discussed above. Ellesmerocerids, endocerids, andtarphycerids, which dominated previously, weresharply reduced in diversity. The number of genera oforthocerids, pseudorthocerids, actinocerids, and lituit-ids increased significantly, while four new ordersemerged. At the same time, the proportions of the life-forms changed only insignificantly (Fig. 3.22).
The proportion of benthic and benthopelagic formsdecreased by 8% compared to the Early Ordovician.This may be a reflection of the increasing transgressionin the mid-Ordovician, which resulted in the pelagicadaptive zone increasing in size and becoming more
strongly differentiated. The adaptive vacancies thatemerged created an opportunity for new groups, with anew body plans and large adaptive potential, whichreplaced more primitive groups (ellesmerocerids,endocerids, and tarphycerids), and increased in diver-sity. The taxonomic composition of all life-forms in theMiddle Ordovician became more diverse. Despite theincrease in numbers and proportion of the total, newgenera of actinocerids and oncocerids appeared along-side the continuing Early Ordovician endocerids andellesmerocerids. Orthocerids and actinocerids becamethe dominant benthopelagic life-forms. Among plank-tonic forms, ellesmerocerids considerably reduced theirpresence, but six new orders appeared. To a varyingextent, the nektobenthic adaptive zone was occupied by11 of the Middle Ordovician orders.
In the Late Ordovician the general ecologicalstructure of the community (Fig. 3.23) changed onlyslightly: the proportion of the benthopelagic formsincreased, while that of the nektobenthic decreased by6%. This might have resulted from large scale reduction
12%pl
28%bp
7%b
53%nb
Ellesmerocerida
Tarphycerida
Ellesmerocerida
Endocerida
Actinocerida
Oncocerida
Oncocerida
Ascocerida
Discosorida
Orthocerida
Barrandeocerida
Lituitida
Ellesmerocerida
OncoceridaDiscosorida
Tarphycerida
Lituitida
Barrandeocerida
Pseudorthocerida
EllesmeroceridaAscocerida
Endocerida
Oncocerida
Discosorida Orthocerida
Actinocerida
Barrandeocerida
Pseudorthocerida
LituitidaTarphycerida
Fig. 3.22. Ecological structure of cephalopods in the Middle Ordovician. Explanations as in Fig. 3.21.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1231
of the shelf areas, which were the main habitats of nek-tobenthic organisms.
The taxonomic composition of all life-formschanged significantly. Although the same 11 orderscontinued from the Middle to the Late Ordovician, thenumber of representatives of different orders withineach of the life-forms decreased.
Among the representatives of the benthopelagiclife-form, pseudorthocerids were replaced by the two-fold increased number of oncocerids. Ellesmeroceridsdisappeared from the benthic communities, whileoncocerids became more prominent. Oncocerids took aleading position in the composition of the benthope-lagic life-form, and together with Discosorids consti-tuted 75% of the taxocoenosis. The number of ordersrepresenting the nektobenthic life-form (the mostdiverse life-form) decreased, while lituitids, ellesmero-cerids, and barrandeocerids disappeared. Representa-tives of the latter order constituted about 20% of theplanktonic life-form, which by then did not containlituitids. All this suggests the replacement of less
advanced order groups by more advanced ones, and theecological differentiation of the more advanced forms.
The ecological structure of the Silurian cephalopodcommunity (Fig. 3.24) is very generalized, since duringthe Silurian the taxonomic diversity and proportions ofthe life-forms were very changeable from province toprovince (Barskov and Kiselev, 1995). This structuremainly reflects the interval of the mid-Silurian–maxi-mum transgression, diversification of reef communitiesand graptolites, which were possibly the links in thesame trophic chain.
In the Silurian in the ecological structure of thecommunity, the proportion of pelagic forms (plank-tonic and nektobenthic) increased and now for the firsttime constituted 70%, and has not gone below this levelever since. Ecologically this may mean the beginningof the development of a balanced community, with thestructure changed only insignificantly, despite the fun-damental changes in the composition of life-forms andthe community in general.
14%pl
32%bp
7%b
47%nb
TarphyceridaActinocerida
Oncocerida
Ascocerida
Discosorida
Orthocerida
Barrandeo-
OncoceridaDiscosorida
Tarphycerida
Ascocerida
Oncocerida
Discosorida
Orthocerida
Actinocerida
Pseudorthocerida
Tarphycerida
cerida
Endocerida
Endocerida
Orthocerida
Endocerida
Fig. 3.23. Ecological structure of cephalopods in the Late Ordovician. Explanations as in Fig. 3.21.

1232
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
Another feature of the Silurian community was theappearance of a morphotype with a unconventionalshell morphology. Benthic communities were domi-nated by pseudorthocerids, which were represented byforms with a gastropod-like shell, and the newlyappeared orthocerids. 80% of the benthopelagic life-form was represented by discosorids and pseudortho-cerids with a curved shell and narrowed or closed aper-ture, which for the first time occupied this adaptivezone. The nektobenthic life-form was for more than ahalf represented by the morphotype of straight shellswith an advanced mechanisms of horizontal orientationcontrol (orthocerids, pseudorthocerids, actinocerids,although they included representatives with a weaklycurved, secondarily straight, or planispiral shell andabove-mentioned forms with a low trochoid and ascoc-eroid (apically decollating). A quarter of the planktoniclife-forms were represented by coiled barrandeoceridsand tarphycerids with slowly expanding, often looselycoiled shells, and among the latter by low trochoid
shells. Half of the planktonic life-forms were brevi-conic, often barrel-shaped shells with a narrowed orclosed aperture. It was the first time when the plank-tonic life-form included orthocerids with a small shellwithout mechanisms for orientation control (ancestorsof bactritids).
Early Devonian. A considerable decrease in thegeneric diversity and fundamental change in the domi-nant taxonomic group (appearance of first ammonoidsand or nautilids) did not affect the total ecologicalstructure of the cephalopod community. The proportionof pelagic forms reached 73% because of an increase inthe number of nektobenthic genera (Fig. 3.25).
A few benthic forms were represented by orthocer-ids, pseudorthocerids, and actinocerids with a brady-conic and trochoid shell. About half of benthopelagiclife-forms were discosorids. The other half comprisedorthocerids, oncocerids, and actinocerids with a brevi-conic-cyrtoconic or secondarily orthoconic shell. The
31%pl
26%bp
4%b
39%nb
ActinoceridaOncocerida
Discosorida
Tarphycerida
Ascocerida
Oncocerida
Discosorida
Orthocerida
Pseudortho-
Tarphycerida
Orthocerida
Discosorida
Barrandeocerida
Actinocerida
cerida
BarrandeoceridaTarphycerida
Ascocerida
Oncocerida
Discosorida
Pseudorthocerida
Orthocerida
Actinocerida
Barrandeocerida
Fig. 3.24. Ecological structure of cephalopods in the Silurian. Explanations as in Fig. 3.21.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1233
nektobenthic life-form was dominated for the first timeby the morphotype of coiled semi-involute shells, rep-resented by anarcestids. The remaining five orders,which belonged to this life-form, were represented bythe morphotype of primarily orthoconic (orthocerids,pseudorthocerids, actinocerids) and secondarily straightshells (oncocerids and discosorids). The compositionof the planktonic life-form changed fundamentally,both taxonomically and morphologically. Taxonomi-cally, the planktonic life-form in the Early Devoniancomprised almost 70% barrandeocerids, and 25%newly appearing groups of the order rank: ammonoidorder Anarcestida and nautilids. Two morphotypes ofcoiled forms dominated: evolute shells with slowlyexpanding whorls and spirally coiled shells withloosely coiled whorls, whereas involute, slowlyexpanding forms were represented by only a few gen-era. Among planktonic forms, a somewhat larger rolewas played compared to the Silurian by the morphotypeof orthoconic subcylindrical shells. The morphotype ofthe cyrtoconic shells with a closed aperture (oncocer-ids, discosorids), dominant in the Silurian, was repre-sented by a few genera.
Middle Devonian. The ecological structure retainedthe same proportions of life-forms as in the Early Devo-nian (Fig. 3.26). Benthic forms were dominated byoncocerids with a breviconic, cyrtoconic, and second-arily orthoconic shell, by representatives of barrandeo-cerids, discosorids, and nautilids, which were repre-sented by one genus each. The benthopelagic group hadthe morphotypes of coiled shells with rapidly expand-ing whorls equally represented, but taxonomically,apart from anarcestids and nautilids the communitycontained newly appeared barrandeocerids. The nekto-benthic life-form also became more diverse than in theMiddle Devonian: but while retaining equal propor-tions of the morphotypes of orthoconic and coiled invo-lute shells, the latter contained newly appeared tornoc-eratids, apart from the continuing anarcestids and nau-tilids. In the Middle Devonian planktonic living formsthe number of anarcestids and nautilids increased.Among the coiled forms the number of taxa with aninvolute shell, with slowly expanding whorls increased.The proportion of the morphotypes with a coiled shelland orthoconic shells (bactritids) remained the same.
28%pl
21%bp
6%b
45nb
Actinocerida
Barrandeocerida
Oncocerida
DiscosoridaActinocerida
Endocerida
Orthocerida
Orthocerida
Pseudorthocerida
Actinocerida
Actinocerida
Orthocerida
Pseudorthocerida
Discosorida
Oncocerida
Anarcestida
AnarcestidaNautilida
DiscosoridaOncocerida
Pseudorthocerida
Fig. 3.25. Ecological structure of cephalopods in the Early Devonian. Explanations as in Fig. 3.21.

1234
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
The number of breviconic forms with a closed aperturesomewhat increased (oncocerids, discosorids).
Late Devonian. In the late Devonian, the genericdiversity was the highest in the entire Paleozoic historyof the class. It increased due to the increase in the num-ber of planktonic forms, as a result of which the propor-tion of pelagic life-forms reached 85%. In the LateDevonian the taxonomic composition and ecologicalstructure of the cephalopod community was profoundlychanged (Fig. 3.27). The major transformations arerecorded in the Famennian, after the crisis of cephalo-pods at the Frasnian/Famennian boundary, whereas thestructure of the Frasnian community in general wasvery similar to that in the Middle Devonian.
The taxonomic identity of the Late Devonian ceph-alopods was determined by the appearance of theammonoid order Clymeniida, which existed only in theFamennian and by the increase in the generic diversityof tornoceratids (ammonoids), among which the plank-tonic and benthopelagic forms appeared for the firsttime, and of discosorids. Most clymeniids (two-thirdsof all existing genera), occupied the planktonic adap-
tive zone, representing the morphotype of evoluteshells (morphogroup 3A), and in the end of their evolu-tion also involute shells (morphogroup 1A) with slowlyexpanding whorls. Beginning in the Famennian, a dis-tinct planktonic specialization is also recorded in theorder Anarcestida. Among tornoceratids, planktonicforms also appear in the Famennian. Discosorids weremore flexible and adaptively diverse and displayed theentire spectrum of life-forms, although being mostlyconnected with benthic environments. Discosorids, aswell as clymeniids, completed their evolution at the endof the Famennian.
The taxonomic composition of life-forms. Benthicforms included only seven discosorid genera. Dis-cosorids also dominated the benthopelagic forms dis-playing a morphotype of breviconic cyrtocones with anarrowed aperture, whereas the morphotype of coiledforms was less than 20%, in contrast to the MiddleDevonian, when anarcestids, nautilids, and barrandeo-cerids constituted not less than half of all benthopelagicforms. Coiled ammonoids (tornoceratids, anarcestids,and newly appeared clymeniids) began playing a more
29%pl
21%bp
6%b
44%nb
Oncocerida
Oncocerida
Discosorida
Oncocerida
Discosorida
Barrandeocerida
Discosorida
Nautilida
Anarcestida
Barrandeocerida
Nautilida
Pseudorthocerida
AnarcestidaBarrandeocerida
Nautilida
Actinocerida
Orthocerida
Tornoceratida
PseudorthoceridaDiscosorida
Oncocerida
Barrandeocerida
Nautilida
Anarcestida
Bactritida
Fig. 3.26. Ecological structure of cephalopods in the Middle Devonian. Explanations as in Fig. 3.21.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1235
noticeable role in the composition of nektobenthic life-form. About a third of all nektobenthic ammonoidswere represented by the morphotype of the secondarilystraight shells of oncocerids and discosorids. In con-trast to the Middle Devonian, discosorids were domi-nant. Orthocerids, pseudorthocerids, and actinoceridswith an initially straight shell, comprising more than25% of the diversity in the Middle Devonian, in theLate Devonian, were represented by only a few genera.The taxonomic structure of the planktonic life-form forover 80% was dominated by ammonoids of the ordersTornoceratida, Anarcestida, and Clymeniida. By thenumber of genera, more than a half of these were cly-meniids, representing the morphotype of the evoluteshells with slowly expanding whorls. The morphotypeof involute shells with slowly expanding whorls wasrepresented by tornoceratids and, to a lesser extent, byanarcestids and late clymeniids. Less than 20% of theplanktonic cephalopods were represented by the mor-photype of straight (bactritids) and breviconic cyrto-cones with a narrowed or closed aperture. The maxi-mum in the Paleozoic generic diversity and a distinct
ecological structure of the cephalopod community ofthe Late Devonian reflects specific climatic and marineconditions of that time, and, to a lesser extent, may bean indicator of these conditions. A warm climate andabundant land vegetation, which, when washed into thewater, was not effectively utilized, resulted in an accu-mulation of non-oxidized organic matter on the sea bot-tom. The absence of the glacial cover and, hence, psy-chrosphere, precluded the convection of marine waterand resulted in the appearance of anoxic conditions inthe deep regions of the marine basins. Periodical spill-ing of anoxic waters onto shallow shelf areas (“blackshales events”) potentially causing mass extinctions(Kellwasser Event at the Frasnian–Famennian bound-ary, Hangenberg Event at the Famennian–Tournaisianboundary). The latter event was catastrophic in the evo-lution of cephalopods, and its consequences were asdramatic as the outcome of the biotic crisis at the Per-mian–Triassic boundary. Evidently, such global eventswere responsible for fundamental changes in the ceph-alopod communities, which represented the mostnumerous and important part of the Paleozoic marine
43%pl
14%bp
2%b
41%nb
Actinocerida
Discosorida
Oncocerida
Discosorida
PseudorthoceridaOrthocerida
Discosorida
Orthocerida
Anarcestida
TornoceratidaClymeniidaNautilida
Anarcestida Oncocerida
Tornoceratida
Clymeniida
NautilidaPraeglyphioceratida
Nautilida
Clymeniida
Discosorida
Änarcestida
Tornoceratida
Bactritida
Fig. 3.27. Ecological structure of cephalopods in the Late Devonian. Explanations as in Fig. 3.21.

1236
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
biota. Apparently, the reduction of the proportions ofthe coastal benthic forms in the total cephalopod diver-sity during the late Devonian was a direct result of thereduction of coastal habitats. The benthic life-form wasrepresented solely by discosorids. The benthopelagicadaptive zone was dominated by the morphotype ofbreviconic cyrtocones of discosorids. In the Late Devo-nian this adaptive zone was also occupied by theammonoid order tornoceratids. Benthopelagic tornoc-eratids were represented by relatively large forms withan involute, spheroconic or subspheroconic shell withslowly expanding whorls. Apparently this shell shapewas more successful in competition with anarcestids,some of which were benthopelagic forms with rapidlyexpanding whorls.
In the Mississippian (Early Carboniferous)changes in the total ecological structure were caused bythe relative reduction in the number of planktonic formsand by ectocochliates completely abandoning thebenthic adaptive zone. Slightly less than a half of thediversity was represented by nektobenthic forms, whileplanktonic and benthopelagic forms were in almostequal proportions. In addition, the composition of alllife-forms became dominated (up to 90%) by morpho-types of ammonoids, among which goniatitids, whichappeared around this time, constituted over half of thetotal diversity of cephalopods (Fig. 3.28).
Of the benthopelagic life-forms, goniatitids repre-sented around 70% and mainly had large, slowly
expanding involute and semi-involute shells with acompressed whorl cross section. The development ofthis morphotype, which in the Carboniferous and Permianbecame dominant among benthopelagic ammonoids, wasthe major change in the morphological structure of thislife-form at the Devonian–Carboniferous boundary.Together with nautilids and tornoceratids (morphotype ofinvolute shells with relatively rapidly expanding whorls),they almost completely replaced the morphotype of ortho-conic pseudorthocerids and the last oncocerids.
At the beginning of the Carboniferous a newammonoid order (Prolecanitids) took the place ofextinct anarcestids. The first prolecanitids (Tournai-sian) were represented by evolute shells with relativelyslowly expanding whorls interpreted as adaptive forms.Their further development resulted in the appearance ofinvolute, rapidly expanding nektobenthic forms.
The nektobenthic life-form was more diverse taxo-nomically and was represented by eight orders, but wasdominated by the coiled, involute shells, with moder-ately or rapidly expanding whorls (morphologicalregions 1A, 4A, 5A) (goniatitids, tornoceratids, andprolecanitids). The second nektobenthic morphotypewas represented by semi-evolute moderately expandingforms (mostly nautilids and a few genera of prolecanit-ids and goniatitids). Half of all nektobenthic generawere represented by goniatitids. The planktonic mor-photype of straight shells was represented by the last ofoncocerids genera and by bactritids (10%). The coiled
29%pl
25%bp
46%nb
Actinocerida
Oncocerida
Tornoceratida
Nautilida
Oncocerida
Tornoceratida
Nautilida
Bactritida Tornoceratida
Nautilida
Oncocerida
Prolecanitida
Goniatitida
Goniatitida
Prolecanitida
Orthocerida
Pseudorthocerida
GoniatitidaPseudorthocerida
Fig. 3.28. Ecological structure of cephalopods in the Mississippian. Explanations as in Fig. 3.21.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1237
planktonic forms were dominated by the morphotypeof evolute, slowly expanding shells, although the pro-portion of the involute and semi-involute slowlyexpanding shells increased (morphospace 2A).
Pennsylvanian assemblages (Middle and LateCarboniferous together). The general ecologicalstructure remained unchanged (Fig. 3.29). Ammonoidsdiversified, while nonammonoid cephalopods, exceptfor nautilids, were represented by only a few genera.Actinocerids and orthocerids were represented by nomore than five genera, and pseudorthocerids by lessthan ten genera. The benthopelagic adaptive zone wasnow populated only by goniatitids and nautilids, mainlyrepresented by involute and semi-involute, cadiconic,spheroconic, and subspheroconic shells with slowlyexpanding whorls. Involute shells with rapidly expand-ing whorls were less common.
Nektobenthic life-forms were more diverse taxo-nomically, although 80% of these were represented byammonoids and nautilids with a coiled semi-involuteand semi-evolute shells with moderately expandingwhorls. Thus, the cephalopod community became moremorphologically homogeneous. Taxa with orthoconic,cyrtoconic, and other shell shapes virtually disappearedfrom all adaptive zones.
Early Permian. The ecological structure of thecephalopod taxocoenosis changed by a slight increasein the number of nektobenthic genera and slight
decrease in the number of planktonic genera with anevolute slowly expanding shell (Fig. 3.30). The taxo-nomic composition of the life-forms changed as fol-lows. Among the benthopelagic life-forms the propor-tion of nautilids increased, while the number of goni-atitids genera decreased (number of goniatitids decreasedacross all adaptive zones); a few benthopelagic tornoc-eratid genera were present. Tornoceratids were repre-sented by one morphotype of involute shells with rap-idly expanding whorls. The number of goniatitids sig-nificantly decreased among nektobenthic forms, whereasprolecanitids reached an equal proportion with them.Nautilids and tornoceratids increased their proportionin the nektobenthos. Less than 20% were representedby the morphotype of orthoconic shells of orthoceridsand pseudorthocerids. Among planktonic forms, theproportion of goniatitids also decreased, due to the pro-portional increase of nautilids, and nautilids were rep-resented by the morphotype of evolute shells withslowly expanding whorls, whereas goniatitids weredominated by representatives of the second morpho-type (involute forms with slowly expanding whorls).
Middle Permian. The overall structure of the taxo-coenosis changed only slightly. The proportion of ben-thopelagic life-forms decreased to 24%, whereas theproportion of planktonic forms increased to 32%(Fig. 3.31). Among benthopelagic life-forms the pro-portions of the taxonomic groups somewhat changed:
32%pl
25%bp
43%nb
PseudorthoceridaOrthocerida
Goniatitida
Nautilida
Tornoceratida
Nautilida
NautilidaBactritida
Goniatitida
Actinocerida
ProlecanitidaGoniatitida
Fig. 3.29. Ecological structure of cephalopods in the Pennsylvanian. Explanations as in Fig. 3.21.

1238
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
nautilids decreased, while goniatitids prevailed(25 genera). Among nektobenthic forms, goniatitidsconstituted 45%, while prolecanitids, tornoceratids,and nautilids were less common. The proportion of themorphotype of orthoconic shells of orthocerids andpseudorthocerids constituted only about 5%. In plank-ton, the proportion of nautilids decreased, whereas cer-atitids appeared (5 genera). Both these groups wererepresented by evolute shells with slowly expandingwhorls. Among goniatitids, which contributed the larg-est proportion to plankton, were equally representedboth evolute and involute morphotypes. The proportionof straight shells (bactritids) decreased.
Late Permian. The Late Permian shows anextremely unusual ecological structure of the cephalo-pod community (Fig. 3.32), which somewhat resemblesthat of the Late Devonian. The taxonomic compositionof the life-forms fundamentally changed, primarilybecause of the Late Permian extinction of previouslydominant goniatitids and the early evolution of ceratit-ids. In the benthopelagic life-form nautilids becamedominant for the first time (more than 60%), whereas inthe Mesozoic and Cenozoic this adaptive zone becametheir only ecological domain. The number of goniatitidgenera, which had previously contributed more than80% to the benthopelagic life-form, experienced a four-
fold decrease. Ceratitids for the first time colonized thisadaptive zone.
Ceratitids, together with nautilids, in equal propor-tions constituted two-thirds of the generic compositionof the nektobenthic life-form. In plankton, the maincontributors were ceratitids, represented by an evolutemorphotype; whereas goniatitids, tornoceratids, andbactritids were represented by a few genera only.
The end-Permian extinction affected all of the thenexisting orders. Bactritids, pseudorthocerids, and goni-atitids became completely extinct. Only two genera ofceratitids, belonging to the planktonic life-form, con-tinued into the Triassic to give rise to the Triassic cer-atitid radiation. Among nautilids, the Permian–Triassiccrisis was survived by two or three benthopelagic gen-era, including one prolecanitid genus, which shortlyafterwards became extinct without descendants, and theonly orthocerid genus, which survived until the end ofthe Triassic.
The above discussion of the changes in the propor-tions and taxonomic composition of the life-forms inthe cephalopod taxocoenosis throughout geologicalepochs during the Paleozoic allows the following con-clusions:
There were several time spans of absolute durationof tens of million years, during which the ecologicalstructure of the cephalopod community changed insig-
27%pl
26%bp
47%nb
Pseudorthocerida
Orthocerida
Goniatitida
Nautilida
Tornoceratida
Nautilida
NautilidaBactritida
Goniatitida
Prolecanitida
Goniatitida
Prolecanitida
Tornoceratida
Fig. 3.30. Ecological structure of cephalopods in the Early Permian. Explanations as in Fig. 3.21.
Tornoceratida
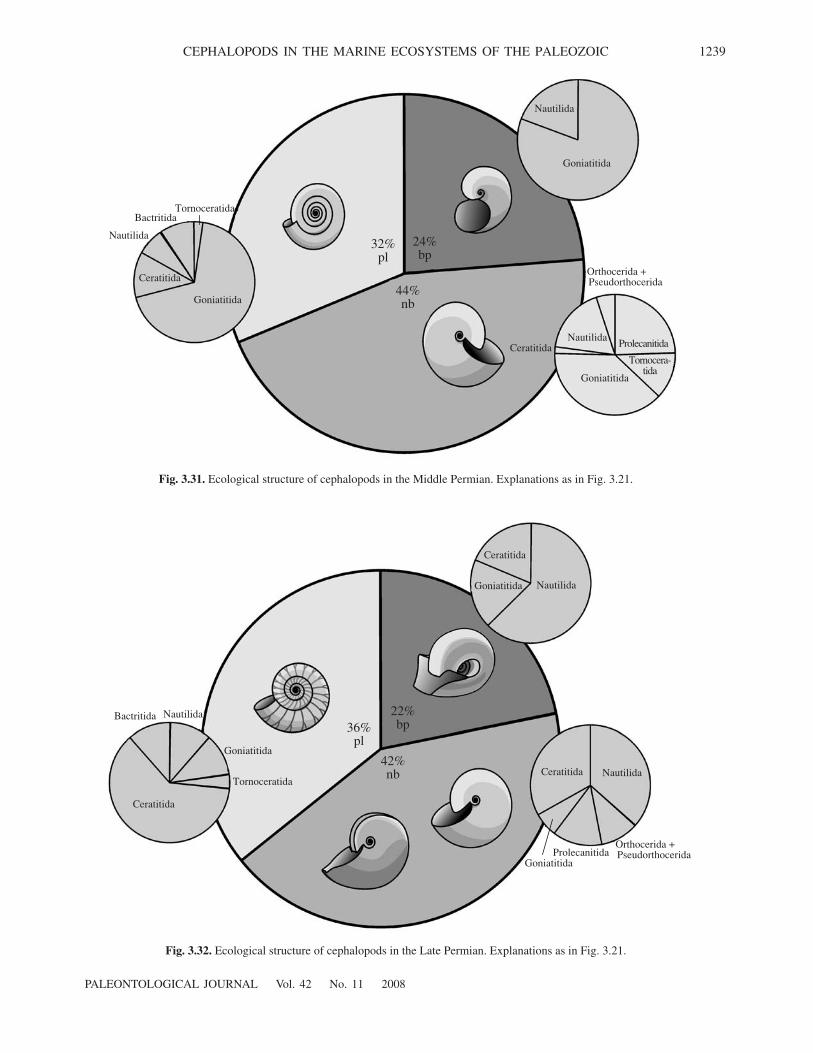
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1239
32%pl
24%bp
44%nb
PseudorthoceridaOrthocerida +
Goniatitida
Nautilida
Tornocera-
Nautilida
Nautilida
Bactritida
Goniatitida
Prolecanitida
Goniatitida
tida
Ceratitida
Ceratitida
Tornoceratida
Fig. 3.31. Ecological structure of cephalopods in the Middle Permian. Explanations as in Fig. 3.21.
36%pl
22%bp
42%nb
PseudorthoceridaOrthocerida +
Goniatitida Nautilida
Nautilida
NautilidaBactritida
GoniatitidaProlecanitida
Goniatitida
Ceratitida
Ceratitida
TornoceratidaCeratitida
Fig. 3.32. Ecological structure of cephalopods in the Late Permian. Explanations as in Fig. 3.21.

1240
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
nificantly. The proportions of the benthic (benthic andbenthopelagic life-forms) and pelagic forms (nekto-benthic and planktonic) fluctuated insignificantly. Peri-ods when the ecological structure was stable (homeo-static) included the Ordovician, Early and MiddleDevonian, and Frasnian (Late Devonian), and Carbon-iferous to Middle Permian. When the proportions of thebenthic (benthic and benthopelagic) and pelagic (nek-tobenthic and planktonic) life-forms, are compared foreach of these periods, a distinct trend towards a gradualincrease of the proportion of the pelagic sector may beobserved. As early as the Early Ordovician, despite thepredominance of the most primitive groups, the result-ing ecological structure contained all life-forms. In theMiddle Ordovician, when five new orders appeared andanother three orders that were previously representedby a few genera, became widespread, a general ecolog-ical structure of this new community, which was twiceas numerous and more diverse taxonomically, changedless than might have been expected. The pelagic part ofthe community increased to 60% due to the appearanceof new groups originally adapted to the pelagic adap-tive zones (orthocerids, pseudorthocerids, actinocer-ids—all nektobenthos; barrandeocerids—plankton).However in the Late Ordovician among these groups,there appeared benthic and benthopelagic forms. Thus,the relative stability of the ecological structure of thecephalopod community in the Ordovician, despite theappearance of new groups and increase in taxonomicdiversity, was determined by the stability in the marineadaptive zones. The Ordovician community character-istically shows a greater taxonomic diversity for each ofthe existing life-forms, compared to any other Paleo-zoic epoch. All then existed cephalopod orders hadtheir representatives in all adaptive zones.
In the Ordovician communities, the proportion ofbenthic (benthic and benthopelagic) forms constitutes43 and 39% in the Early and Late Ordovician, respec-tively, and 35% in the Middle Ordovician. The reduc-tion of benthic forms in the Middle Ordovician corre-lates with the maximum transgression. At that time astructure of the marine ecosystem similar to that oftoday began developing. Beginning in the Devonian,the proportion of the benthic communities became sta-bilized at a level of 25%. (In the Recent community,this proportion is about 20%.) In the Devonian, thereremained some benthic forms, but beginning in the Car-boniferous no benthic forms among ectocochliates everappeared. Periods of stability in the ecological structurewere interrupted by shorter periods when the ecologicalstructure of the community was significantly different(epoch of disrupted adaptive homeostasis). Theseincluded Silurian, Famennian (Late Devonian), andLate Permian. All these periods show an increase in theproportion of planktonic forms.
Theoretically there are three groups of reasonsresponsible for upheaval in the ecological structure ofthe community. The internal reasons: appearance ofnew body plans, of new taxonomic groups, the physiol-
ogy of which (reflected by the shell geometry and con-struction) was adapted to a certain lifestyle. To someextent, this may explain the increase in the proportionof planktonic forms in the second half of the Permian,when the first ceratitids appeared.
Among the external reasons it is possible to assumeincreased pressure of competition with other, more suc-cessful bottom-dwelling organisms. For instance,changes in the taxonomic structure of the cephalopodcommunity in the Silurian due to more than twofoldincrease in the proportion of planktonic forms mainlyresulted from the reduction in the nektobenthos, theproportion of which in the community was the lowest inthe entire Paleozoic. It is the most likely that this, likethe reduction in the number of bottom-dwelling cepha-lopods, could be related to the appearance of newgroups of bottom-dwelling fauna. For some of the newcolonists, cephalopods could become prey (e.g., forlarge arthropods). Other new inhabitants (fish) couldhave been better hunters. Other external reasons couldinclude changes in the oceanic environment leading tothe reduction in the number of suitable niches in someadaptive zone. Possibly, this may explain the largestchange in the ecological structure of the community inthe second half of the Late Devonian. Anoxic events inthe Late Devonian, leading to the extinction of thegroups of benthic biota and groups connected with thebenthos, affected cephalopod communities by almosttwofold reduction of the proportion of benthic and ben-thopelagic forms and almost threefold increase in thenumber of the inhabitants of the planktonic life-form.Half of the planktonic life-form was represented by theorder Clymeniida (Ammonoidea), more precisely by amorphotype of evolute shells, which are interpreted asinhabitants of the open pelagic zone.
Probably, all of these reasons played their roles, tovarying extents, at times of upheaval of the ecologicalstructure of the cephalopod community.
CHAPTER 4. ECOLOGICAL STRUCTUREOF PALEOZOIC AMMONOID COMMUNITIES
IN THE URALIAN PALEOBASIN
The study of the evolution of ammonoid assem-blages over a prolonged time interval within one largepaleobasin allows detailed tracking of the nature andmechanism of the morphological and ecologicalchange in communities and their relationship with abi-otic events.
4.1. General Background
The Uralian Late Paleozoic basin is one of the mostfavorable objects for such studies. This ancient oceanexisted continuously during the second half of the Pale-ozoic. In the southeast it was connected, through theAral Region, with the marine basins of the Tien Shanand Pamir, in the east with central and eastern Kazakh-stan, in the west with the epicontinental seas of the Rus-

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1241
sian Platform, and in the north with the basins ofNovaya Zemlya and Timan-Pechora Province. Almostthe entire Middle–Upper Paleozoic sequence of theUrals, from the Emsian (Lower Devonian) to the Artin-skian Stage of the Lower Permian contains numerousammonoids. Some assemblages (Famennian, LateTournaisian, Late Visean, Serpukhovian, Early Bash-kirian, Gzhelian, Asselian, Sakmarian, Early and LateArtinskian) are unique both in taxonomic diversity andin the number of individuals.
During the time span under consideration this basinwas gradually closing and shallowing as the Siberianand Kazakhstan continents collided with the Russiancraton. Beginning in the Late Devonian, the UralianPaleo-ocean, which separated Baltica from Siberia andKazakhstania, was gradually closing. This process wasaccompanied by orogeny and volcanism on the easterncoast of the basin, and by extensive seismic eventsresulting from the sideways sliding of plates on both theeastern and western coasts.
By the end of the Devonian, in the Frasnian andFamennian a series of north-south structures wasformed in the territory of the modern Urals, includingmicrocontinents and island arcs separating marinebasins with different sedimentary settings (Puchkov,2000). In the west, between the margin of Baltica andthe microcontinent Uraltau, there was a deep epiconti-nental basin, which formed in the Early Paleozoic(Mizens, 1997, 2000; etc.). The submerged north-southtectonic structures of the Uralian paleocean were themain source of terrigenous material. In its northwesternpart there existed deep troughs, separated by uplifts. Inthe Frasnian and Famennian Domanik-like sedimentsand thick series of mainly terrigenous and terrigenous-carbonate beds accumulated in the troughs. In the LateDevonian, in the eastern regions there remained a deepoceanic basin, Magnitogorsk island arc, and a subduc-tion zone.
At the Devonian and Carboniferous boundary, aconsiderable change in geodynamic settings occurredin the Urals, resulting from the interruption of the sub-duction in the Magnitogorsk Megazone (Puchkov,2000). Thus, the Uralian Basin was gradually loosingits oceanic characteristics, as Laurasia became moreconsolidated by the beginning of the Carboniferous.
In the Early Carboniferous and at the beginning ofthe Middle Carboniferous, a deep marine basinremained in the north, which was bordered on the westby a shallow Carboniferous shelf of the Timan-PechoraProvince. Deep marine basins remained in the territorypresently occupied by the Middle and South Urals.These basins had the appearance of narrow, asymmetri-cal troughs in the west and east, and were freely con-nected in the south. In the deepest zones of the southernregions of the basin (Zilair Megasynclinorium) sedi-mentation, as in the Late Devonian, was mainly per-formed by gravity flows. The rising Urals Mountainswere the main source of the terrigenous material, and in
the southeast of the basin there were also volcanicislands of the Magnitogorsk arc. In the south regions ofthe basin, the depth increased from north to south fromBashkortostan to the Aktyubinsk Region. In the secondhalf of the Early Carboniferous and at the beginning ofthe Middle Carboniferous a shallow shelf sea waspresent on the eastern slope of the Urals, with accumu-lation of mainly carbonate series, and only in its westernpart had relatively deep settings (Mizens, 2002; etc.).
In the Bashkirian, the Uralian Fore-Deep begandeveloping, which led to a considerable change in thesedimentary and paleogeographic settings in the basin.The amount of polymictic material washed from theslopes of the Mountainous Urals increased resulting inthe accumulation of thick flyschoid series on the east-ern slope of the depression. At the same time the area ofthe Carboniferous shelf in the west decreased.
In the Moscovian, the Uralian Basin had a numberof submeridional facial zones. In the center, there was achain of islands, symmetrically separating the basininto the western and eastern areas. These areas wereisolated in the Moscovian in the North, Middle, andmost of the South Urals and were connected at the per-iclinal region, which is supported by several faunalgroups in common (Ivanova, 2002). In the eastern slopeof the South Urals the marine basin represented a gulfwith no exit in the north and a connection in the southwith the European part of the Basin and, via the AralRegion, with seas of Tien Shan, Pamir, central and east-ern Kazakhstan (Chuvashov et al., 1984; Ivanova andChuvashov, 1990). The European part of the Urals wasa more open deep epicontinental basin, which in thewest was separated by a chain of islands and shoalsfrom the Moscovian Sea of the Russian Platform.
In the Late Carboniferous, orogeny began andmarine conditions remained mainly on the westernslope of the Urals. At that time and during the EarlyPermian (Asselian, Sakmarian), the Uralian Basin wasa meridional asymmetrical strait, surrounding the EastEuropean inner sea in the east and connecting thebasins of Paleotethys and northern Panthalassa. Thestrait was separated from the platform regions of theEast European sea by a chain of shoals, islands, andreefs. The latter were interpreted as barrier reefs(Antoshkina, 2003; etc.). The Permian reef bodiesextended as a submeridional belt along the entire west-ern slope of the Urals from the south to the north, witha branch to the east of the Timan. The hydrological,geographical, and geological conditions were very dif-ferent on either side of the barrier reef. A that time alarge carbonate-evaporite platform existed on the EastEuropean plain. Thick molasse and flyschoid series,which accumulated in the fore-deep, were built of ter-rigenous material from the Hercynian Mountain chainsin the east and at the same time carbonate material fromthe slope of the platform in the west. During the entireLate Carboniferous and Early Permian the fore-deephad a distinct transverse asymmetry. From the east to
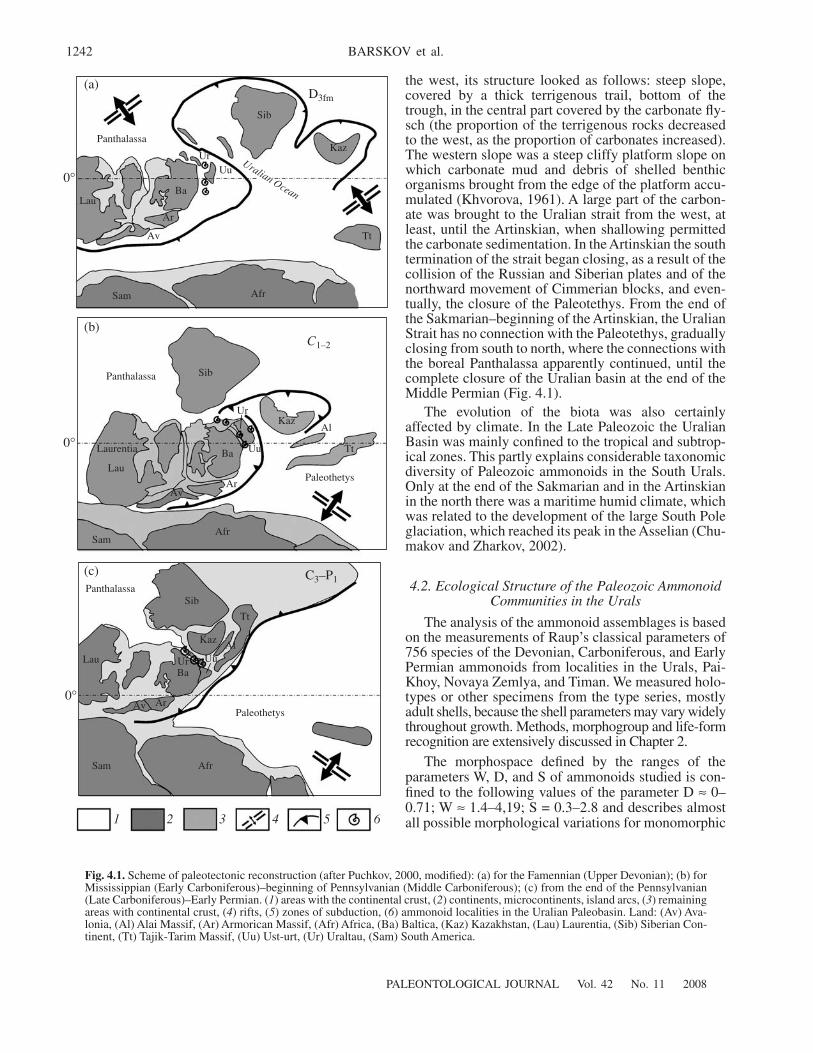
1242
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
Panthalassa
Panthalassa
Panthalassa
Uralian Ocean
Paleothetys
Paleothetys
D3fm
Lau
Av
AfrSam
Ar
Sib
Kaz
Tt
UrUu
Ba
C1–2
0°
0°
Sib
KazAl
TtBa
ArAv
Ur
Uu
AfrSam
Lau
Laurentia
C3–P1
Sib
Tt
Kaz Al
BaUuUrLau
Av Ar0°
Sam Afr
1 2 3 4 5 6
(a)
(b)
(c)
Fig. 4.1. Scheme of paleotectonic reconstruction (after Puchkov, 2000, modified): (a) for the Famennian (Upper Devonian); (b) forMississippian (Early Carboniferous)–beginning of Pennsylvanian (Middle Carboniferous); (c) from the end of the Pennsylvanian(Late Carboniferous)–Early Permian. (1) areas with the continental crust, (2) continents, microcontinents, island arcs, (3) remainingareas with continental crust, (4) rifts, (5) zones of subduction, (6) ammonoid localities in the Uralian Paleobasin. Land: (Av) Ava-lonia, (Al) Alai Massif, (Ar) Armorican Massif, (Afr) Africa, (Ba) Baltica, (Kaz) Kazakhstan, (Lau) Laurentia, (Sib) Siberian Con-tinent, (Tt) Tajik-Tarim Massif, (Uu) Ust-urt, (Ur) Uraltau, (Sam) South America.
the west, its structure looked as follows: steep slope,covered by a thick terrigenous trail, bottom of thetrough, in the central part covered by the carbonate fly-sch (the proportion of the terrigenous rocks decreasedto the west, as the proportion of carbonates increased).The western slope was a steep cliffy platform slope onwhich carbonate mud and debris of shelled benthicorganisms brought from the edge of the platform accu-mulated (Khvorova, 1961). A large part of the carbon-ate was brought to the Uralian strait from the west, atleast, until the Artinskian, when shallowing permittedthe carbonate sedimentation. In the Artinskian the southtermination of the strait began closing, as a result of thecollision of the Russian and Siberian plates and of thenorthward movement of Cimmerian blocks, and even-tually, the closure of the Paleotethys. From the end ofthe Sakmarian–beginning of the Artinskian, the UralianStrait has no connection with the Paleotethys, graduallyclosing from south to north, where the connections withthe boreal Panthalassa apparently continued, until thecomplete closure of the Uralian basin at the end of theMiddle Permian (Fig. 4.1).
The evolution of the biota was also certainlyaffected by climate. In the Late Paleozoic the UralianBasin was mainly confined to the tropical and subtrop-ical zones. This partly explains considerable taxonomicdiversity of Paleozoic ammonoids in the South Urals.Only at the end of the Sakmarian and in the Artinskianin the north there was a maritime humid climate, whichwas related to the development of the large South Poleglaciation, which reached its peak in the Asselian (Chu-makov and Zharkov, 2002).
4.2. Ecological Structure of the Paleozoic Ammonoid Communities in the Urals
The analysis of the ammonoid assemblages is basedon the measurements of Raup’s classical parameters of756 species of the Devonian, Carboniferous, and EarlyPermian ammonoids from localities in the Urals, Pai-Khoy, Novaya Zemlya, and Timan. We measured holo-types or other specimens from the type series, mostlyadult shells, because the shell parameters may vary widelythroughout growth. Methods, morphogroup and life-formrecognition are extensively discussed in Chapter 2.
The morphospace defined by the ranges of theparameters W, D, and S of ammonoids studied is con-fined to the following values of the parameter D ≈ 0–0.71; W ≈ 1.4–4,19; S = 0.3–2.8 and describes almostall possible morphological variations for monomorphic

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1243
shells (Fig. 4.2). The most widespread (over 40% of thetotal species diversity) were the nektobenthic life-forms from morphospace 1A (morphogroup 1A). Thismorphogroup is one of the most stable and is present inassemblages throughout the time span studied. Typicalrepresentatives include Tornoceras, Dzhaprakoceras,Goniatites, Dombarites, Proshumardites, Delepinoceras,Bilinguites, Agathiceras, Gonioloboceras, Thalassoceras,etc. Species with a discoconic, platyconic, or oxyconicshell with moderately or rapidly expanding whorls(W ~ 2.0–3.5)—morphogroup 4A, for instance Timan-ites, Kazakhoceras, Girtyoceras, Carinoceras, Medli-cottia, Artinskia, etc. also belonged to the nektobenthiclife-form, but, in contrast to group 1A, they were scarcethroughout the Paleozoic. Their species diversity isabout 10% of the total number of forms. A usual shellsize is Dm = 40–60 mm, whereas in some species the shellreached size considered large for the Paleozoicammonoids: Dm = 100–200 mm and over.
Planktonic evolute, widely umbilicate forms with avarying cross-sectional shape of the whorl from mor-phogroup 3A (plankton-1) constitute about 22% of thetotal taxonomic diversity. Typical representatives ofthis morph include Hexaclymenia, Trigonoclymenia,Clymenia, Eonomismoceras, Rhymmoceras, Alao-ceras, Dombarigloria, Cancelloceras, Eoasianites,Svetlanoceras, Paragastrioceras, etc. Small involuteand moderately involute shells with slowly expandingwhorls (Prolobites, Epiwocklumeria, Quasicravenoc-eras, Mirilentia, Lyrogoniatites, Ferganoceras, Emi-lites, Protopopanoceras, Crimites, etc.), included inmorphogroup 1A' and 2A and characterizing the sec-ond type of the planktonic life-form (P-2), are lessdiverse—9% of the total number of species.
Benthopelagic forms were not numerous. Theyincluded subspheroconic, spheroconic, and cadiconicshells, which were relatively large—with a diameter of35 mm and more, and with a slowly or moderatelyslowly expanding shell: the genera Eurites, Glaphy-rites, some species of the genera Goniatites, Dom-barites, Gastrioceras, Cravenoceras, etc. We refer rel-atively large (Dm = 35 mm and over) Pachyconic andsubdiscoconic shells with a moderately narrow ormedium-sized umbilicus from morphogroup 2A to thebenthopelagic forms. In total, the diversity of benthope-lagic forms was low—17% of the total number of species.
The proportions of the life-forms, taxonomic andmorphological diversity of the ammonoids from theUralian Basin changed considerably throughout thecourse of their evolution (Figs. 4.3a, 4.3b). It is possibleto recognize several stages of intense morphogenesisseparated by episodes of crises or stagnations, largelyrelated to global or regional abiotic events (transgres-sions and regressions, change in sedimentary settings,tectonic disruption, climatic changes, etc.).
Below the evolution of ammonoids is discussed bystratigraphic stages from the Devonian to Permian.
4.3.1. Early Devonian
Emsian
The evolution of ammonoids in the Uralian basinbegan from the Emsian (Early Devonian). The oldestammonoid assemblages are known from the easternslope of the North and Middle Urals (SverdlovskRegion), and also Novaya Zemlya. Ammonoids werestudied by B.I. Bogoslovsky (1962a, 1963, 1969,1972). Ammonoid collections are medium sized,including about 200 specimens housed in the Paleonto-logical Institute of the Russian Academy of Sciences(PIN). Ammonoids come from several localities.
In the north Urals they were found in the region ofthe Krasnotur’insk (right bank of the ZabolotnayaRiver, tributary of the Bolshya Volchanka). From dark-gray to black, bedded cherty limestone with thin lami-nations of calcareous sandstone and marl Bogoslovsky(1969) identified Erbenoceras advolvens, Mimosphinc-tes tenuicostatus, Teicherticeras lissovi, and Convolu-ticeras erbeni.
Ammonoids from the Upper Emsian beds are foundin two localities, situated on the left bank of the PyshmaRiver (eastern slope of the Middle Urals). The lime-stone inclusions from tuff interbeds and xenolithesfrom porphyrites yielded Teicherticeras pyshmense.
In the number of specimens the Uralian assem-blages are dominated by representatives of the genus
1.5
0.11.0
0.2 0.3 0.4 0.5 0.6 0.7 D0
2.0
2.5
3.0
3.5
4.0
4.5
W
W = 1/D
3A
6A
2A
5A
4A
1A
1A'
Fig. 4.2. Distribution of the shell shape of 756 species ofPaleozoic ammonoids from the localities in the Urals, Pai-Khoy, and Novaya Zemlya in relation to the values of thewhorl expansion rate (W) and whorl overlap degree (D).Morphospaces recognized for Paleozoic ammonoids areindexed (see Chapter 3).
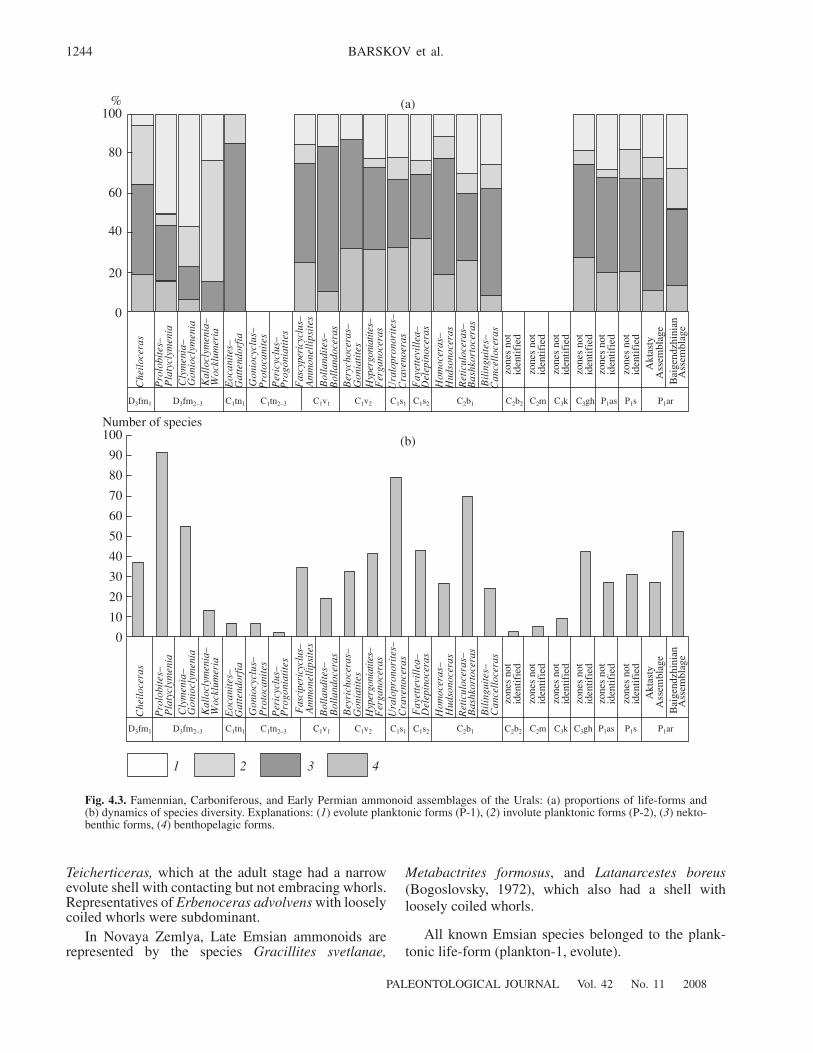
1244
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
Teicherticeras, which at the adult stage had a narrowevolute shell with contacting but not embracing whorls.Representatives of Erbenoceras advolvens with looselycoiled whorls were subdominant.
In Novaya Zemlya, Late Emsian ammonoids arerepresented by the species Gracillites svetlanae,
Metabactrites formosus, and Latanarcestes boreus(Bogoslovsky, 1972), which also had a shell withloosely coiled whorls.
All known Emsian species belonged to the plank-tonic life-form (plankton-1, evolute).
100%
80
60
40
20
0
(a)
Number of species100
90
80
70
60
50
40
30
10
0
(b)
Che
iloce
ras
Pro
lobi
tes–
Pla
tycl
ymen
iaC
lym
enia
–G
onio
clym
enia
Kal
locl
ymen
ia–
Woc
klum
eria
Eoc
anite
s–G
atte
ndor
fiaG
onio
cycl
us–
Pro
toca
nite
sP
eric
yclu
s–P
rogo
niat
ites
Fas
cype
ricy
clus
–A
mm
onel
lipsi
tes
Bol
land
ites–
Bol
land
ocer
asB
eryc
hoce
ras–
Gon
iatit
esH
yper
goni
atite
s–F
erga
noce
ras
Ura
lopr
onor
ites–
Cra
veno
eras
Fay
ette
ville
a–D
elep
inoc
eras
Hom
ocer
as–
Hud
sono
cera
sR
etic
uloc
eras
–B
ashk
orto
cera
sB
iling
uite
s–C
ance
lloce
ras
zone
s no
t
Akt
asty
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
d
Bai
gend
zhin
ian
Ass
embl
age
Ass
embl
age
Che
iloce
ras
Pro
lobi
tes–
Pla
tycl
ymen
iaC
lym
enia
–G
onio
clym
enia
Kal
locl
ymen
ia–
Woc
klum
eria
Eoc
anite
s–G
atte
ndor
fiaG
onio
cycl
us–
Pro
toca
nite
sP
eric
yclu
s–P
rogo
niat
ites
Fas
cipe
ricy
clus
–A
mm
onel
lipsi
tes
Bol
land
ites–
Bol
land
ocer
asB
eyri
choc
eras
–G
onia
tites
Hyp
ergo
niat
ites–
Fer
gano
cera
sU
ralo
pron
orite
s–C
rave
noce
ras
Fay
ette
ville
a–D
elep
inoc
eras
Hom
ocer
as–
Hud
sono
cera
sR
etic
uloc
eras
–B
ashk
orto
cera
sB
iling
uite
s–C
ance
lloce
ras
zone
s no
t
Akt
asty
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
dzo
nes
not
iden
tifie
d
Bai
gend
zhin
ian
Ass
embl
age
Ass
embl
age
D3fm1 D3fm2–3 C1tn1 C1tn2–3 C1v1 C1v2 C1s1 C1s2 C2b1 C2b2 C2m C3k C3gh P1as P1s P1ar
D3fm1 D3fm2–3 C1tn1 C1tn2–3 C1v1 C1v2 C1s1 C1s2 C2b1 C2b2 C2m C3k C3gh P1as P1s P1ar
1 2 3 4
20
Fig. 4.3. Famennian, Carboniferous, and Early Permian ammonoid assemblages of the Urals: (a) proportions of life-forms and(b) dynamics of species diversity. Explanations: (1) evolute planktonic forms (P-1), (2) involute planktonic forms (P-2), (3) nekto-benthic forms, (4) benthopelagic forms.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1245
4.3.2. Middle Devonian
Eifelian
Scarce ammonoids are known from the MiddleDevonian of the Uralian Basin (23 species of 13 generaare known from the Eifelian). Ammonoids were studiedby Bogoslovsky (1958a, 1958b, 1961, 1969). They arefound in several localities, situated on the eastern slopeof the North and Middle Urals. The collections (about150 specimens) are housed in the Paleontological Insti-tute, Russian Academy of Sciences.
Early Eifelian ammonoids have been describedfrom the light-gray, fine-grained micritic limestones onthe eastern slope of the North Urals: Gyroceratitesglaber, Laganites tenuis, Parentites praecursor, Augu-rites mirandus, and Latanarcestes kakvensis (Bog-oslovsky, 1961). Together with ammonoids there arefound occasional brachiopods, gastropods, tetracorals,crinoids, and nautiloids. Ammonoid assemblages aredominated by oxyconic and discoconic shells of mor-phogroups 4A and 6A, nektobenthic forms—3 species,planktonic life-form is represented by one evolute spe-cies Gyroceratites glaber. The species Parentites prae-cursor has a discoconic shell with very rapidly expand-ing whorls (W = 3.8) and is provisionally interpreted asa benthopelagic form.
A diverse ammonoid assemblage is described fromthe Lower Eifelian on the left bank of the BobrovkaRiver (eastern slope of the Middle Urals, ArtemovskiiDistrict). The following species were identified fromthe cherry-red, thickly bedded cherty limestones, over-lain by calcareous sandstones: Gyroceratites gracilis,Fasciculoceras uralense, Mimagoniatites obesus,M. angulostriatus, Agoniatites uralensis, Latanarces-tes pronini, L. ventroplanus, Subanarcestes macro-cephalus, S. bisulcatus, Mimanarcestes nalivkini, andWerneroceras bobrovkense (Bogoslovsky, 1969). Apartfrom ammonoids, the limestones include bivalves andnautiloids, and diverse benthic fauna: tetracorals, bra-chiopods, gastropods, crinoids, trilobites, and tentacu-lites. Ammonoids are scanty (only 132 specimens inour collections). The assemblages (both in number ofspecimens and in specific diversity) are dominated byoxyconic, discoconic, and subdisconic nektobenthicspecies (7 of 11 species). Planktonic forms are repre-sented by evolute shells with varying whorl height(three species). The benthopelagic life-form is repre-sented by one species only.
Givetian
Givetian ammonoids are known from one locality(left bank of the Bolshoi Elets River, upstream of thevillage of Eletskaya, near Vorkuta) on the western slopeof the Polar Urals. A few ammonoids have beendescribed from the thickly layered dark-gray lime-stones with foraminifers, brachiopods, and trilobites:Werneroceras uralicum, Pseudofoordites hyperboreus,and Wedekindella psittacina (Bogoslovsky, 1959). The
host rocks are dated Late Givetian. The species Werner-oceras uralicum and Pseudofoordites hyperboreus maybe referred to the nektobenthic life-form, because theyhave a discoconic medium-shelled shell (Dm = 30–50 mm) with a narrow umbilicus and relatively rapidlyexpanding whorls. Wedekindella psittacina have a dis-coconic, narrowly umbilicate small shell (less than20 mm in diameter), with slowly expanding whorls,characteristic of the planktonic (plankton-2) life-form.
The development of the Early and Middle Devonianammonoids from the Uralian Paleocean reflects thegeneral features of the evolution of this group at thisstage. Only planktonic forms are characteristic of theEmsian. In the Eifelian, the diversity of life-forms andmorphotypes increased, and nektobenthic and bentho-pelagic forms appeared. Ammonoids predominantlybelong to the nektobenthic life-form and are found inlimestones including diverse benthic fauna.
An extremely impoverished ammonoid assemblagefrom the Givetian of the Subpolar Urals is also mainlyrepresented by nektobenthic forms. Givetian ammonoids,similar to Eifelian, come from the relatively shallowcarbonate beds, including diverse benthic fauna.
4.3.3. Late Devonian
Frasnian
In the Late Devonian, the diversity of ammonoids inthe Uralian Paleocean increased considerably. Themain peak of morphogenesis was in the Famennian andwas to a large extent related to the appearance and evo-lution of the order Clymeniida. The taxonomic andmorphological diversity of Frasnian ammonoids wererelatively low. They are mainly known from the locali-ties on the western slope of the Middle and SubpolarUrals, in Novaya Zemlya, Pai-Khoy, and South Timan.In the second half of the Frasnian, the ammonoidsbegan their evolution in the South Urals.
The Lower, Middle, and (possibly) Upper Frasnian(Manticoceras Zone) contain numerous ammonoids.However, fine correlations with the Western Europeanzones is often complicated, because the majority of theUralian species are endemic or have a wide strati-graphic distribution. The assemblage includes 27 spe-cies, 11 genera, and four families. Collections includeabout 2900 specimens housed in the PaleontologicalInstitute of the Russian Academy of Sciences, Cherny-shev TsNIGR-Museum, St. Petersburg Mining Insti-tute, and in All-Russia Scientific Research GeologicalOil Institute (the taxonomic composition of Frasnianammonoids was studied by many workers (Bog-oslovsky, 1957, 1958b, 1969, 1971; Bogoslovsky et al.,1982; Yanischewsky, 1926; Lyashenko, 1956, 1957; etc.).
The morphospace defined by the parameters W, D,and S of Frasnian ammonoids is restricted by the valuesof W = 1.54–3.3; D = 0.01–0.57; and S = 0.67–1.9. Thefauna was dominated by the nektobenthic ammonoidswith discoconic and oxyconic shells, with a narrow or

1246
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
moderately narrow umbilicus (23 species, 85%). Evo-lute ophioconic or platyconic shells are also present(4 species, 15%). Ammonoids are known from variousparts of the basin, characterized by different sedimen-tary settings and geology. In total, the assemblages areimpoverished both taxonomically and morphologically.Frasnian ammonoids of the Urals show a high degree ofendemism at the species level.
In the north of the Uralian Paleobasin, ammonoidsare found in several localities, on the western slope ofthe Subpolar Urals and in the Chernyshev Range (basinof the Usa River). Ammonoids were found in gray anddark-gray, almost black, limestones (Kynov-SargaevoBeds). The assemblage included Hoeninghausia ucht-ensis, Koenetites uralensis, Timanites keyserlingi, Tor-noceras typum, Komioceras stuckenbergi, Ponticerassp., and Manticoceras sinuosum. This assemblage istypical of the Lower Frasnian (Manticoceras Zone,possibly an equivalent of Zone C, based on Timaniteskeyserlingi). All species have a discoconic, narrowlyumbilicate shell, are often large in size and representthe nektobenthic life-form. An equivalent ammonoidassemblage is known from the localities in the basin of theKara River (eastern slope of the Pai-Khoy Range). Timan-ites keyserlingi and Tornoceras typum are recorded fromblack pyritized limestone (Bogoslovsky, 1969).
On the South Timan, the Early Frasnianammonoids (Zones B–E) are found in the localities inthe Ukhta River basin. Ammonoids come from the car-bonate interbeds in the series of the greenish-gray clayswith rare layers of light-gray and greenish-gray marl ofthe Ust-Yarega Formation, and also from the carbonateinterbeds of the Domanik series. The Ust-Yarega For-mation is composed of clay, siltstone with marl inter-beds, and of sandy, clayey, and detrital limestone. Thedeposits of the Domanik Formation represent organic-rich carbonate-cherty shale with clayey interbeds.Together with ammonoids, there were found rare bac-tritoides, nautiloids, and tentaculites, and in addition,numerous remains of brachiopods and ostracodes. Theassemblage includes Tornoceras typum, Timanites key-serlingi, Ponticeras uralicum, P. uchtense, P. tsherny-schewi, P. bisulcatum, P. lebedeffi, P. auritum, Probelo-ceras keyserlingi, P. domanicense, Komioceras stucken-bergi, Aulotornoceras keyserlingi, Uchtites syrjanicus,Manticoceras ammon, etc. The assemblages are domi-nated by nektobenthic discoconic and oxyconic forms(11 species), and also narrow evolute planktonic speciesof the genera Ponticeras and Probeloceras (4 species).
The Late Frasnian ammonoids come from the local-ities in the Sed’yu River basin, along the rivers Lyaioland Vezha-Vozh from the Lyaiol Formation (Bog-oslovsky, 1969). This formation is represented by thesediments of the slope of the fore-reef depression,including turbidites: interbedding of organic-rich shaleand limestone. Ammonoids come from the limestone,which also contains numerous remains of brachiopods,ostracodes, and conodonts, whereas clay contains
miospores. Predominantly algal reef was located in thesouth and southeast of the region (Yudina and Mosk-alenko, 1997). The assemblages are dominated bynumerous species of the genus Manticoceras, with sub-dominant Carinoceras ljaschenkovae, C. menneri, Tor-noceras typum, and Timanoceras ellipsoidale. Mostspecies have a discoconic or oxyconic shell with a mod-erately narrow or narrow umbilicus and a moderate orhigh whorl expansion rate, characteristic of the nekto-benthic life-form. Shells frequently reach large size (upto 200 mm in diameter and over), while the averagediameter of the shells of the genus Manticoceras is 60–70 mm).
In the Middle Urals, ammonoids are found in thedeposits of the Lower and Upper Frasnian (Mantico-ceras). A few (53 specimens) of the Early Frasnianammonoids come from the Kynov-Sargaev Beds(locality on the left bank of the Khoroshevka River,near the town of Gubakha, Perm Region). Bogoslovsky(1969) recorded Koenetites uralensis, Hoeninghausiakoswensis, and Timanites keyserlingi from the compactlight gray, ferruginous limestone. These species have anarrow, discoconic or oxyconic, large (Dm = 50–140 mm) involute shell, characterized by the nekto-benthic life-form.
The Late Frasnian ammonoids are recorded fromthe localities on the Kos’va and Vil’va Rivers (westernslope of the Middle Urals, Perm Region). Manticocerasintumescens, M. cordatum, M. sinuosum, M. sp., Torno-ceras typum, and Aulotornoceras sp. have beenrecorded from the light gray, grained limestone beds(Bogoslovsky, 1969). All species have a discoconicinvolute shell with a narrow or moderately narrowumbilicus, high whorl expansion rate, and belong to thenektobenthic life-form.
In the south of the Uralian Paleobasin the evolu-tion of ammonoids began somewhat later. The earliestoccurrences are found in the Upper Frasnian. TheUpper Frasnian ammonoids come from the localities onthe western slope of the South Urals (Bashkortostan,basins of the Zilim and Zigan Rivers). This territory, sit-uated near the margin of Baltica and separated from theopen ocean by the Uraltau microcontinent, at the end ofthe Devonian represented an epicontinental basin. Atthe end of Frasnian, various facies accumulated in thisbasin, including those characteristic of the shallowshelf: limestones, calcareous-clayey shales, siltstones,clayey shales, etc., containing few ammonoids. The fol-lowing species have been recorded from these beds:Manticoceras sinuosum, M. cordatum, M. intumescens,and Tornoceras typum (Bogoslovsky, 1969). All thesespecies belonged to the nektobenthic life-form. Animpoverished taxonomic composition and homogenousecological structure of the communities apparently sug-gest unfavorable conditions in the usual ammonoidhabitats.
Thus, the assemblages of the Frasnian ammonoidsare characterized by the predominance of large involute

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1247
discoconic nektobenthic shells both in the number ofindividuals and taxonomic composition. Planktonicevolute, platyconic shells recorded from the Early Fras-nian of the South Timan were distributed to a muchlesser extent. Ammonoids inhabited the region of theCarboniferous shelf and upper part of the slope. Theabsence of benthopelagic forms, which became wide-spread later, is very characteristic of the Late Frasnian.
Famennian
In the mid-Frasnian–Tournaisian, the Timan-Pechora system of troughs was developing in the west-ern part of the North Uralian Basin (Malyshev, 2000).As a result, carbonate shoals were formed on the localuplifts, where the accumulative subaqueous structurese.g., carbonated banks, or small carbonate buildups sur-rounded by the zones of uncompensated sedimentationcarbonate banks and small isolated organic buildups(Antoshkina, 2003). Mainly clayey-carbonate andclayey-cherty beds of the Domanik type accumulated indeep troughs. Marine water surrounding the carbonateplateau, characterized by quiet hydrodynamic condi-tions, abundant decayed organic matter or presence ofvolcanic material, which affected the chemical compo-sition and facilitated the appearance of the anoxic con-ditions and hydrogen sulfide accumulation (Maksi-mova, 1970; Antoshkina, 2003). These conditions wereapparently unfavorable for ammonoids. During themost of the Famennian, in the territory of the North andSubpolar Urals they are almost completely absent. Theonly discovery of the Early Famennian ammonoids inthe North Urals (Yaiva River) is Cheiloceras verneuililisted by Krotov (1888).
A few Late Famennian (Kalloclymenia–Wocklume-ria Genozone) ammonoids are known from the Kozhimsection (western slope of the Subpolar Urals). Theammonoid assemblage contains Kalloclymenia kozhi-mensis and Rectimitoceras obsoletum and R. angustilo-batum collected from the overlying beds (Bogoslovskyand Kusina, 1980). Ammonoids come from the seriesof black clayey-cherty limestones with lenses and mas-sive cherty beds. A character of faunal preservationindicates a possible post-mortem transfer. The assem-blage is dominated by evolute planktonic shells of Kal-loclymenia kozhimensis occasionally forming layers,whereas involute, narrowly umbilicate species of thegenus Rectimitoceras, characteristic of the nektobenthiclife-form, are represented by only a few specimens.
The taxonomically richer Late Famennian ammonoidlocalities, containing Posttornoceras contiguum,Maeneceras sulciferum, Uraloclymenia volkovi, Fal-ciclymenia uralica, Protxyclymenia dubia, Cymacly-menia costata, and Kalloclymenia glabra are knownfrom the eastern slope of the Polar Urals (Man’ya andLoz’va rivers). This assemblage is dominated by evo-lute planktonic forms.
A regression over the territory of the South Urals atthe Frasnian–Famennian boundary reached its peak atthe beginning of the Famennian (triangularis conodontzone), and led to a crisis in the ammonoid communities.No ammonoids are found from the basal Early Famen-nian of the South Urals. The regression was followedby a transgression resulting in the increased outer shelf,which was the main ammonoid habitat in the Famen-nian (Becker, 1993; etc.); and this was probably themain reason for the rapid growth of their taxonomic andmorphological diversity.
The Famennian was a unique time in ammonoidevolution. At this time ammonoids displayed the high-est taxonomic diversity in their Paleozoic history.Famennian ammonoids included in this analysis comefrom the interval corresponding to the total of the fourammonoid genozones: Cheiloceras, Prolobites–Platy-clymenia, Clymenia–Gonioclymenia, and Kalloclyme-nia–Wocklumeria. A collection of more than 25 thou-sand shells is housed at the Paleontological Institute,Russian Academy of Sciences (coll. nos. 1263, 1266,1447, 2688, 2755, 3754). The systematic compositionof the Famennian ammonoid fauna of the South Ura-lian-Kazakhstan Region has been studied by manyauthors (Tokarenko, 1903; Perna, 1914; Kolotukhina,1938; Kind, 1944; Nalivkina, 1953; Bogoslovsky, 1969,1971, 1981; Nikolaeva and Bogoslovsky, 2005; etc.).
The Famennian beds of the South Urals yielded193 ammonoids species of 60 genera and 21 families.The morphospace for this interval is restricted by thevalues of W = 1.4–2.7; D = 0–0.65, and S = 0.33–2.0.
The assemblages of Famennian ammonoids weredominated by planktonic forms—49% of species, mostof which have evolute ophioconic or platyconicshells—116 species (Fig. 4.4). Most members of thismorphogroup belong to the order Clymeniida, includ-ing the families Cymaclymeniidae, Gonioclymeniidae,Hexaclymeniidae, Cyrtoclymeniidae, Rectoclymeni-idae, Clymeniidae, Carinoclymeniidae, Pachyclymeni-idae, Gonioclymeniidae, etc. of tornoceratids, this mor-phogroup includes species of the families Pseudocly-meniidae and Tornoceratidae (genus Posttornoceras).Other planktonic forms, 24 species, were representedby involute, slowly expanding shells, characteristic ofthe representatives of the orders Anarcestida and Torno-ceratida (species of the family Prionoceratidae, Spora-doceratidae, Dimeroceratidae, Cheiloceratidae, Post-tornoceratidae, and Prolobitidae). Among clymeniids,some representatives of the family Parawocklumeridaewith a triangular coiling in early whorls belonged tothis group.
Nektobenthic forms were also widespread (33% ofthe total number of species). Two-thirds of these(52 species) were represented by pachyconic and dis-coconic involute and semi-involute shells. Representa-tives of the nektobenthic life-form were present in allorders, with the majority in the order Tornoceratida(families Cheiloceratidae, Tornoceratidae, Posttornoc-

1248
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
eratidae, etc.), Praeglyphioceratida, and Anarcestida.To a lesser extent (26 species) this group is representedby clymeniids, by species with a discoconic semi-invo-lute shell. The nektobenthic oxyconic forms with rap-idly expanding whorls were uncommon.
Species, belonging to the benthopelagic life-formwere uncommon (8% of the total taxonomic diversity).
Characteristically Famennian ammonoid communi-ties displayed a trend towards gradual increase in theproportion of planktonic forms and reduction of ben-thopelagic, and later nektobenthic forms.
Morphological and taxonomic diversity and ecolog-ical structure of the communities of Famennianammonoids varied greatly in time and different facialenvironments (Figs. 4.3, 4.5).
Famennian ammonoid localities in the South Ura-lian region are subdivided into three major groupsbased on their type of preservation, diversity, and thenumber of shells, on the one hand, and on a facial typeof host rocks, on the other. The latter facial types arerecognized in: (1) Bashkortostan, (2) western Kazakh-stan, (3) eastern slope of the South Urals. These typesof localities belong to different tectonic zones and dif-ferent types of basins. The Famennian ammonoid faciesin Bashkortostan were accumulated in an epicontinen-tal basin of varying depth, with mostly carbonate sedi-mentation. Localities of Famennian ammonoids inwestern Kazakhstan (Aktyubinsk and Orenburgregions) belong to the deep regions of the western shelfof the Uralian Ocean. Famennian carbonates found inthe Chelyabinsk Region, with mass accumulations ofammonoid shells, were apparently deposited in theback-arc basin in the Magnitogorsk tectonic zone.Despite the geographic isolation of the above localities
and considerable differences in the lithology of the hostrocks assemblages of Famennian ammonoids are simi-lar in taxonomic composition and are considerably dif-ferent from the synchronous assemblages of the Rheno-Hercynian zone (Nikolaeva and Bogoslovsky, 2005). Atthe same time each type of locality displays an individ-ual ecological structure of ammonoid communities.
The Famennian section in Bashkortostan containsclymeniids and goniatitids of all four genozones.Deposits of the Cheiloceras genozone are well repre-sented on the Basu, Inzer, Ryauzyak, Zilim, Takata,Mendym, Terekly, and Ishikai rivers. They are com-posed of gray and light gray bedded limestone withnumerous ammonoids, often in association with brachi-opods, trilobites, and corals. In the Inzer and Basu riv-ers, and in some other localities, the basal beds of thegenozone are composed of a series of shale and claywith interbeds of cherty rocks and limestone, with atotal thickness of 3 m (Tyazheva, 1961). In the middleand upper parts of the series, thinly bedded gray lime-stones contain shells of straight nautiloids andammonoids. The middle part of the genozone is repre-sented by a series of gray thinly bedded micritic lime-stones, whereas the overlying beds are composed oflight gray medium-bedded limestones with accumula-tions of ammonoid shell of the genus Cheiloceras. Inmany neighboring regions of Bashkortostan the terrig-enous member at the base of the genozone is absent,whereas light gray and gray limestones compose theentire genozone and contain a rich fauna of ammonoidsand brachiopods. In total, these deposits may be inter-preted as having been accumulated in a shallow basin.The assemblages is dominated by numerous ammonoidsof the genus Cheiloceras (Cheiloceras circumflexum,C. subpartitum, C. inversum, C. globosum dorsatum,C. sacculum, C. amblylobum, Sporadoceras latiloba-tum, etc.). The communities are dominated by involutepachyconic and discoconic nektobenthic forms (55% ofspecies), whereas benthopelagic species to a lesserextent (27%). Planktonic forms constitute 18% of thetotal species diversity (Figs. 4.5a, 4.5b). They aremainly represented by small involute shells. Disco-conic forms, with moderately narrow umbilicus andslowly expanding whorls (Pseudoclymenia) are lesscommon.
The overlying gray and light gray bedded limestone,with interbeds of black calcareous-cherty shales andwith chert lenses contain numerous ammonoids of theProlobites–Platyclymenia Genozone, often togetherwith brachiopods. In general beds with ammonoids canbe interpreted as medium-deep thin carbonates. Thetotal thickness of the genozone is 2 to 9.5 m. Numerousammonoids: Prolobites delphinus, Sporadoceras rotun-dum, Platyclymenia tschernyschewi, Rectoclymenia cf.subflexuosa, and Protactoclymenia krasnopolski fromthe localities on the Terekly, Ishikai, and Sikashty riversare found in the carbonate interlayers (Karpinsky,1869; Chernyschev, 1887; Bogoslovsky, 1981). In gen-eral, the assemblage is uniform, noticeably dominated
49% 116 species
8%20 species
33%78 species
10%24 species
1 2 3 4
Fig. 4.4. Proportions of life-forms in the assemblage ofFamennian ammonoids of the Uralian basin. Explanationsas in Fig. 4.3.
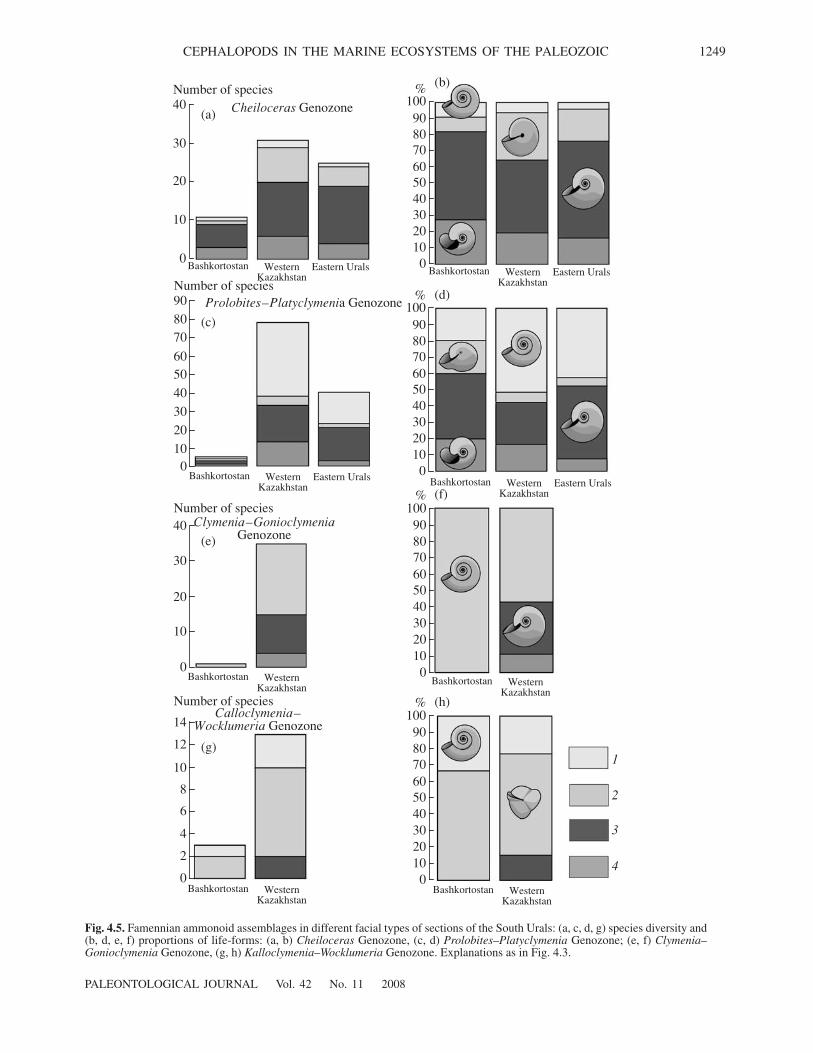
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1249
10
0
20
30
40Number of species
100
50
70
90Number of species
20
30
40
60
80
40Number of species
30
20
10
0
14
Number of species
10
6
2
0
4
8
12
100
50
70
90
203040
60
80
100%
100
50
70
90
203040
60
80
100%
100
50
70
90
203040
60
80
100%
100
50
70
90
203040
60
80
100%
Cheiloceras Genozone
Prolobites–Platyclymenia Genozone
Clymenia–Gonioclymenia
Calloclymenia–
Genozone
Wocklumeria Genozone
Bashkortostan WesternKazakhstan
Eastern Urals
Bashkortostan WesternKazakhstan
Eastern Urals
Bashkortostan WesternKazakhstan
Bashkortostan WesternKazakhstan
Bashkortostan WesternKazakhstan
Eastern Urals
Bashkortostan WesternKazakhstan
Eastern Urals
Bashkortostan WesternKazakhstan
Bashkortostan WesternKazakhstan
1
2
3
4
(a)
(c)
(e)
(g)
(b)
(d)
(f)
(h)
Fig. 4.5. Famennian ammonoid assemblages in different facial types of sections of the South Urals: (a, c, d, g) species diversity and(b, d, e, f) proportions of life-forms: (a, b) Cheiloceras Genozone, (c, d) Prolobites–Platyclymenia Genozone; (e, f) Clymenia–Gonioclymenia Genozone, (g, h) Kalloclymenia–Wocklumeria Genozone. Explanations as in Fig. 4.3.

1250
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
by the compressed shells of Platyclymenia tscherny-schewi. The average size of shells is about 25 mm. Theremains are unsorted. The shells are moderately wellpreserved, occasionally with remains of fine ornamen-tation, but in the majority there is no terminal aperturepreserved, whereas some shells are rounded, suggest-ing short-distance transportation.
In this period the species diversity was reduced con-siderably (5 species instead of 11 in the precedinggenozone) and the ecological structure of the commu-nity changed. The proportion of planktonic formsincreased, with evolute compressed clymeniid shellsprevailing in number of individuals. The diversity ofnektobenthic forms considerably decreased, and thenumber of benthopelagic forms was also somewhatreduced (Figs. 4.5c, 4.5d). These changes happenedwhile the basin was gradually becoming deeper due toa transgression, with reached its maximum floodinginterval in the annulata phase. The impoverished taxo-nomic diversity suggests that the environment in thebasin was probably unfavorable for ammonoids. Thetaxonomic diversity progressively decreased (Fig. 4.5e)in the later epochs. The assemblage of the Clymenia–Gonioclymenia genozone contains almost exclusivelyevolute shells of Clymenia laevigata. Beds of this ageare represented by gray and light gray, in places chertylimestones with ammonoids, often in association withbrachiopods, trilobites, bivalves, corals, and ostracodes(Sultanaev, 1973; Kochetkova et al., 1986). In the ZiganRiver basin (see Nalivkin, 1926, 1937, 1945) beds ofthis age contain interbeds of algal limestones, suggest-ing that the basin was relatively shallow.
The youngest assemblage (Kalloclymenia–Wock-lumeria Genozone) is exclusively represented byplanktonic evolute and involute shells, some of whichhad triangular coiling of early whorls or at all stages:Parawocklumeria paradoxa, Cymaclymenia evoluta,and Synwocklumeria baschkirica (Nalivkin, 1945; Sul-tanaev, 1973; Popov, 1975).
The evolution of ammonoids in Bashkortostan wasmainly restricted to a shallow carbonate shelf. Numer-ous remains of benthic organisms suggest the nearbypresence of a carbonate platform or several such plat-forms. In some localities the rocks of this age containterrigenous beds, indicating periodic influx of clastics.In the early Famennian this part of the basin becamedeeper, while the shallow marine habitats of the earliestFamennian were gradually diminished. Ammonoids areconsiderably less diverse than in western Kazakhstan,with communities dominated by one or two endemictaxa. A short-distance post-mortem transportation ofshells is possible.
Famennian ammonoids of western Kazakhstan arediverse and numerous. They mainly come from carbon-ates containing almost exclusively conodonts and ceph-alopods and no or very little benthic fauna. In additionto ammonoids, numerous remains of straight nautiloidsare present in the rocks. Ammonoid shells form huge
accumulations, often forming cephalopod shellstone.The shells are unsorted and not oriented. Many shellshave fine structures preserved (terminal apertures,spines, and keels). The shell matrix is often present.Body chambers of large shells often contain smallershells, indicating post-mortem transportation at shortdistances. This ammonoid fauna was studied by Kind(1944), Nalivkina (1953), Bogoslovsky (1955; 1960a,1960b; 1962a, 1962b; 1965; 1975; 1976; 1977; 1979a,1979b; 1981; 1982; 1983), Nikolaeva and Bogoslovsky(2005). Ammonoids come from the Kiinskaya Forma-tion (localities–Shiyli-Sai, Ornektotas-Sai, Aral-Tyube-Bakai, Kiya 1, Kiya 2, etc.), which represents a cherty-clayey terrigenous serious interbedded with gray ceph-alopod limestones (Bogoslovsky, 1969; Nikolaeva andBogoslovsky, 2005). Limestones of the Kiya Formationare irregularly bedded, often clayey, with scanty crinoiddebris, containing small thin-shelled planktonic ostra-codes, single-chambered foraminifers (Parathuram-mina spp.), few Septaglomospiranella sp., and numer-ous conodonts (Akhmetshina et al., 2004). The totalthickness does not exceed 15–20 m. The reduced thick-ness, specific microfacies and virtually completeabsence of benthic organisms suggest sedimentary set-tings of a deep shelf. In most localities the most diverseand numerous ammonoid assemblages are found in theCheiloceras and Prolobites–Platyclymenia genozones.The number of shells and diversity of ammonoidsnoticeably decrease upward in the Famennian section(Fig. 4.5).
The ammonoid assemblage of the Cheiloceras Zoneis represented by numerous species of Cheiloceras,Sporadoceras, Dimeroceras, etc. (section on the Aral–Tyube–Bakai River). Upward in the section, the rockscontain mass accumulations of Sporadoceras clarkei,S. muensteri, S. equale, Pseudoclymenia pseudogonia-tites, etc. The communities were dominated by nekto-benthic species—47 %, in most cases with a discoconicor pachyconic shell, with moderately and even slowlyexpanding whorls (W ~ 1.7). Benthopelagic forms arerepresented to a lesser extent (19% of species). Plank-tonic forms are considerably more diverse taxonomi-cally compared to those from localities in Bashkor-tostan, in total they constitute 34% of the species diver-sity (Figs. 4.5a, 4.5b). At the base of the genozone theyare represented by involute tornoceratid species, withcompressed evolute Pseudoclymenia pseudogoniatitesappearing upward in the section.
Beginning from the Prolobites–PlatyclymeniaGenozone, ammonoid assemblages of western Kazakh-stan are noticeably dominated by clymeniids (about80% of the total number of species). The appearance ofclymeniids was apparently not related to changes in thesedimentary setting because neither lithologicalchanges, nor discontinuities are observed in the sec-tions at the level with the earliest clymeniids. The firstclymeniids (genera Cyrtoclymenia, Platyclymenia, Genu-clymenia, Pleuroclymenia) appear after the completedisappearance of the tornoceratid genus Pseudoclymenia,

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1251
which were similar to clymeniids in their shell (com-pressed evolute shell). Simultaneously with clymeni-ids, assemblages contain many shells of Prolobiteswhich may be interpreted as planktonic. The shells ofProlobites are small, spheroconic, with very slowlyexpanding whorls and with deep constrictions, and withthe last (terminal) constriction almost completely clos-ing the aperture.
The highest ammonoid diversity coincides with themaximum of the large-scale transgression (zones del-phinus and annulata). Beds at this level contain largeaccumulations of Prolobites delphinus, P. nanus, Reni-tes striatus, Sporadoceras cf. inequale, Sporadocerascf. clarkei, S. pisum, numerous representatives ofMaeneceras, Falcitornoceras, Araneites, Nothospora-doceras, Praeglyphioceras, compressed ribbed andsmooth clymeniids Pricella stuckenbergi, Platyclyme-nia pompeckii, P. cf. subnautilina, Rectoclymenia subpli-cata, Rectoclymenia roemeri, Genuclymenia aktubensis,G. frechi, Posttornoceras contiguum, Platyclymeniasubnautilina, P. valida, Genuclymenia frechi, Trigono-clymenia protacta, T. spinosa, Protoxyclymenia dubia,Genuclymenia sp., etc. Juvenile and adult shells arepresent, many with body chambers. Average size of theshells is 2.5–3 cm, whereas the maximum size is almost25 cm. In the overlying beds similar limestone con-tained Uraloclymenia nodosa, Trigonoclymenia tigra,Pleuroclymenia costata, Protactoclymenia sp., Protox-yclymenia carinata, and Sporadoceras discoidale.
In the Prolobites–Platyclymenia Genozone the eco-logical structure of ammonoid communities changedfundamentally. The communities (58% of species) arecomposed of planktonic, mainly evolute forms. Theproportion of nektobenthic forms (25% of species) andbenthopelagic forms (17% of species) considerablydecreased (Figs. 4.5c, 4.5d). The latter are representedmainly by pachyconic shells with a narrow or moder-ately narrow umbilicus and slowly expanding whorls.These changes and the appearance and diversificationof clymeniids apparently could not be solely related tothe deepening of the basin during the local transgres-sion, but rather are the product of the evolution of theammonoid taxocoenoses of the deep shelf, on the glo-bal scale.
In the Clymenia–Gonioclymenia Genozone thenumber and diversity of ammonoids decreased consid-erably. The assemblages are dominated by large shellsof the genus Clymenia, which represent over a half ofthe total number of specimens. The assemblageincludes Rectimitoceras pompeckji, Sporadoceraskiense, Mimimitoceras liratum, Rectimitoceras kiense,R. cf. substriatum, Progonioclymenia aff. acuticostata,Cymaclymenia barbarae, Protoxyclymenia rotundata,Cyrtoclymenia angustiseptata, Clymenia laevigata,Discoclymenia cucullata, Renites kiensis, Costaclyme-nia binodosa, Gonioclymenia levis, Sphenoclymeniasp., Biloclymenia aktubensis, Maeneceras inflexum,Gonioclymenia hoevelensis, Ornatoclymenia ornata,
Gonioclymenia hoevelensis Wedekind, Sphenoclyme-nia maxima, Protoxyclymenia pseudoserpentina, Bilo-clymenia dubia, Kiaclymenia semiplicata, etc.
Despite the considerable change in the taxonomiccomposition, the overall ecological structure of thecommunities did not change (Fig. 4.5). The number ofnektobenthic forms somewhat increased (30% of spe-cies instead of 25% in the preceding genophase) due tothe reduction in the diversity of benthopelagic forms.The proportion of planktonic forms remainedunchanged. This may be explained by stable conditionsin the ammonoid habitats through the genophase.
At the end of the Famennian, in the Kalloclymenia–Wocklumeria Genophase the diversity decreased evenmore. The number of shells also considerablydecreased. The shells are scattered in the rocks (clayeylimestones), and are poorly preserved. The communi-ties are mainly composed of planktonic forms (84% ofspecies) (Figs. 4.5g, 4.5h). The newly appeared wock-lumeriids are most abundant, and they had small (1.2–2 cm in diameter) inflated and spheroconic, or triangu-larly coiled shells. The diversity and abundance of kos-moclymeniids, cyrtoclymeniids, and cymaclymeniidsvery much decreased. This is an unusual situationbecause in the underlying beds with a similar lithology(Clymenia–Gonioclymenia Genozone) kosmoclymeni-ids, cyrtoclymeniids, and cymaclymeniids are verydiverse (Nikolaeva and Bogoslovsky, 2005a; Nikolaevaand Bogoslovsky, 2005b). The absence of representa-tives of these families in the assemblages of thesphaeroides Zone in western Kazakhstan distinguishesit from synchronous communities of Great Britain,Germany, Morocco, Poland, and Northern Caucasus,where the evolution of kosmoclymeniids, cyrtoclyme-niids, and cymaclymeniids continued, and wocklumeri-ids existed together with evolute cymaclymeniids untilthe very end of the Famennian.
In the Late Famennian, only a few genera wereendemic to western Kazakhstan, but there were manyendemic species. In addition, in comparison with otherregions of the Uralian-Kazakhstanian region, the basinof western Kazakhstan retained a high diversity ofammonoids during the entire Famennian. Planktonicevolute shells of clymeniids were the key players in theammonoid communities until the end of the Famen-nian. In the Clymenia–Gonioclymenia Genophase,pachyconic, involute, narrowly umbilicate clymeniids,belonging the nektobenthic life-form, became consid-erably more widespread. At the very end of the Famen-nian, among the planktonic forms, narrow evoluteshells of clymeniids almost disappeared, beingreplaced by small, inflated, triangularly coiled wock-lumeriids.
A decrease in the ammonoid communities in west-ern Kazakhstan in the Late Famennian could be con-nected with the total reduction of the shelf areas in thecourse of progressive collision of plates and microcon-tinents in the area of the future Ural mountains. The

1252
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
reduction of the epicontinental part of the basin, cer-tainly led to structural changes in the communities andaffected the food resources of cephalopods. Communi-ties, almost entirely composed of highly specializedforms, probably could not adapt to environmentalchanges and new habitats.
Several localities of Famennian ammonoids are sit-uated on the eastern slope of the Urals, in the Verkh-neuralsk Region to the southwest of the town Verkh-neuralsk (Karpinsky, 1884; Tokarenko, 1903; Perna,1914; Bogoslovsky, 1969, 1971. 1981; Nikolaeva andBogoslovsky, 2005a). The lower part of the section iscomposed of reddish limestone with greenish-grayinterbeds, with numerous shells of ammonoids, nauti-loids, and small corals, remains of trilobites and gastro-pods. The overlying beds are represented by light grayand brownish-gray compact, medium-grained lime-stone, containing a rich fauna of brachiopods, bivalves,and to a lesser extent gastropods and ammonoids. Themain source of the carbonate could be local subaqueousuplifts inhabited by small-sized shelled benthos. Theseare relatively shallow water deposits probably repre-senting the back-arc facies. Well-preserved ammonoidshells do not suggest significant post-mortem transpor-tation. Therefore, it is possible that the simultaneouspresence of remains of relatively deep-sea organism(ammonoids) and shallow-sea benthos suggest that thematerial was transported by a flow of debris from theedge of a shelf or the boundary of a carbonate platformto the deep shelf. Ammonoid assemblages in theselocalities belong to the Cheiloceras and Prolobites–Platyclymenia genozones.
The ammonoid assemblage of the CheilocerasGenozone is diverse. It includes goniatitids and tornoc-eratids: Sporadoceras muensteri, Sporadoceras rotun-dum, Dimeroceras mamilliferum, Dyscheiloceras latil-obus, a variety of Cheiloceras and Tornoceras species.Similar to other regions the communities are mainlycomposed of nektobenthic forms—61% of species.Benthopelagic (16%) and planktonic (23%) forms arerepresented to a lesser extent (Figs. 4.5a, 4.5b). Thisecological structure of the communities, apparentlyresulted from a relatively shallow ammonoid habitats.
In the Prolobites–Platyclymenia Genophase thediversity of planktonic forms (47% of species)increased, mostly due to the evolution of evolute cly-meniids (Figs. 4.5c, 4.5d). The number of nektobenthicand benthopelagic forms decreased (45 and 8%, repsec-tively). The assemblage is composed of various species ofthe genera Prolobites, Maeneceras, Sporadoceras, Perno-ceras, Protornoceras, Kirsoceras, Armatites, Platyclyme-nia, Cyrtoclymenia, Pricella, Genuclymenia, Rectocly-menia, and Protoxyclymenia, typical of the lower andupper parts of the Prolobites–Platyclymenia Genozone(delphinus–annulata and dunkeri zones). The taxo-nomic diversity, increased at the beginning of the geno-zone later rapidly decreased, which, apparently, was
related to the overall decrease of the shelf areas. Theoverlying beds do not contain ammonoids.
Two large stages with different ecological structureof the ammonoid communities are recognized in theevolution of the fauna of the Upper Devonianammonoids of the Uralian Paleobasin. The first (Fras-nian–Middle Famennian) shows low taxonomic diver-sity, predominance of nektobenthic forms (85%), whileplanktonic forms were uncommon and benthopelagicforms are almost entirely absent. At the second stage(Late Famennian) that taxonomic diversity was high.All major life-forms are represented in the communi-ties. The main trend may be identified as a focused evo-lution of planktonic forms in all three orders, whichrapidly became dominant in the communities.
Despite different habitats of Famennian ammonoids(shallow shelf near a carbonate platform, open deepshelf, etc.) leading to the formation of communities dif-ferent ecologically, it is possible to recognize somegeneral patterns in the development of ammonoid faunain this territory.
(1) The richest assemblages come from the deep-seasediment suggesting that Famennian ammonoids, likemodern Nautilus, preferred deep sea. This is confirmedby the fact that the peaks of ammonoid diversity in eachof the regions studied coincided with the annulataphase, a time of a global-scale highstand (Becker,1993; etc.). After a flooding interval the diversity andabundance of ammonoids in all studied localities of theUralian region gradually decreased.
(2) The major trend in the evolution of theammonoid communities in the Famennian was anincrease in the diversity of planktonic life-forms andreduction of the diversity of the benthopelagic life-form, followed by the decrease in the diversity of nek-tobenthic forms (Fig. 4.3). In the second half of Famen-nian, with the appearance of the order Clymeniida, theproportion of planktonic species increased in all fourtypes of localities and represented more than 50% ofspecies diversity.
(3) The increase in the number of planktonic formswas happening at the time of the maximum floodinginterval with a peak in the annulata phase. At the sametime, it is apparent from the analysis of the westernKazakhstan ammonoids that this reason cannot aloneaccount for the increased abundance of the planktonicforms, nor for the explosive radiation of clymeniids.
(4) The decrease in the ammonoid diversity in theUralian region by the end of the Famennian was grad-ual. It is unlikely that it was related solely to globaleustatic fluctuations. It is most likely the decrease wascontributed to considerably by local tectonic eventsresulting in a decrease of the shelf, which was a tradi-tional ammonoid habitat and possibly by a decrease infood resources. The extinction primarily affected theplanktonic evolute forms, and their species diversitysharply decreased. At the end of the Famennian thecommunity was dominated by planktonic narrowly

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1253
umbilicate, slowly expanding forms. Representativesof this morphogroup, were apparently the best adaptedto the unfavorable conditions established from the endof the Famennian.
The Devonian–Carboniferous boundary is markedby a deep crisis in the ammonoid communities, whichis often explained by a series of large-scale transgres-sion-regression pulses recorded in many places on theglobe and accompanied by the development of short-term anoxic events (so-called Hangenberg Event) fol-lowed by sea level drops. As a result of the mass extinc-tion at the Devonian–Carboniferous boundary, only afew prionoceratid species survived to cross the bound-ary (with an involute, narrowly umbilicate forms, withslowly and moderately expanding whorls, representativesof the nektobenthic and planktonic (P-2) life-forms.
4.3.4. Mississippian (Early Carboniferous)
Tournaisian
In the Urals and adjacent territories of Pai-Khoy,ammonoids are apparently present in all genozones(here we used the European zonation) of the Tournai-sian: Eocanites–Gattendorfia, Goniocyclus–Protocanites,Pericyclus–Progoniatites, Fascipericyclus–Ammonellip-sites. Because the Tournaisian–Visean boundary isbased on foraminifers, it divided the Fascipericyclus–Ammonellipsites ammonoid genozone into two parts.However, in the opinion of most ammonoid workers,the Fascipericyclus–Ammonellipsites Phase representsa single definitive stage in the evolution of the groupand is not divided into the Tournaisian and Visean sub-stages, and therefore this phase is analyzed here as asingle unit. The degree of knowledge of the assem-blages is different. Ammonoid collections include morethan 750 specimens, housed in the PaleontologicalInstitute, Russian Academy of Sciences, in ChernyshevTsNIGR Museum, and Department of Paleontology ofSt. Petersburg State University. The systematic compo-sition of ammonoids was studied by many authors(Librovitch, 1941; Balashova, 1953; Kusina, 1971,1973, 1974, 1980, 1983, 2000; Popov, 1975; Popov andKusina, 1997; Kusina and Konovalova, 2004; etc.).
The Tournaisian ammonoid assemblage includes54 species of 31 genera and 7 families. The mor-phospace of the used combination of the parameters W,D, and S is delineated by the values D = 0.02–0.4; W =1.54–2.7, and S = 0.35–2.4. In general, communities ofTournaisian ammonoids have involute pachyconic andsubdiscoconic nektobenthic forms (49% of species).Spheroconic and pachyconic involute forms with a nar-row or medium-sized umbilicus are less common—benthopelagic (27%). Evolute and involute planktonicforms constituted 24% of species (Fig. 4.6). Theammonoid diversity remained low throughout the Tour-naisian and only slightly increased at the very end ofthe Tournaisian. (Fig. 4.3b).
Early and Middle Tournaisian were not epochs ofhigh diversity of ammonoids in the Uralian Basin. Theearliest Tournaisian ammonoids are known from local-ities in western Kazakhstan (Berchogur) (Kusina,1985). The assemblage includes a few weakly orna-mented, involute, narrowly umbilicate species: Acutim-itoceras subbilobatum, A. carinatum, A. mugodzarense,A. yatskovi, A. pulchrum, Rectimitoceras aff. substria-tum, and R. bertchogurense. Ammonoids are found insmall lens-shaped accumulations in yellowish-gray,clayey, bedded limestone, deposited in a shallow shelf(Kusina, 1985, Barskov et al., 1984). The remains aremoderately well preserved, and the shell matrix is pre-served fragmentarily. The average shell size is 12–25 mm. The taphonomy and preservation of theseammonoids suggest considerable post-mortem trans-portation. Together with ammonoids, conodonts andsometimes brachiopods are found. Ammonoid assem-blages contain a few species, characterizing nekto-benthic and, to a lesser extent, planktonic (1 species)life-forms. Such ecological structure is apparentlycharacteristic of the post-crisis or crisis phase of com-munity evolution.
Occasional finds of Middle Tournaisian ammonoids,so called Goniocyclus fauna, are known from the west-ern slope of the South Urals (Orenburg Region) andSubpolar Urals (Popov and Kusina, 1997). In the SouthUrals ammonoids are found in terrigenous-carbonatebeds on the Kamsak River. The assemblage containsthe following species: Goniocyclus subtilis, G. dom-barovensis, G. vodoresovi, Muensteroceras modestum,Aquilonites uralensis, and Gattendorfia uralica, char-acterizing mainly nektobenthic and, to a lesser extent,benthopelagic life-forms.
In the Subpolar Urals, ammonoids of this age arefound in the clayey-carbonate beds (Vangyr River),
7%2 species
17%5 species
49%15 species
27%8 species
1 2 3 4
Fig. 4.6. Proportions of life-forms in the Tournaisianammonoid assemblages in the Urals. Explanations as inFig. 4.3.
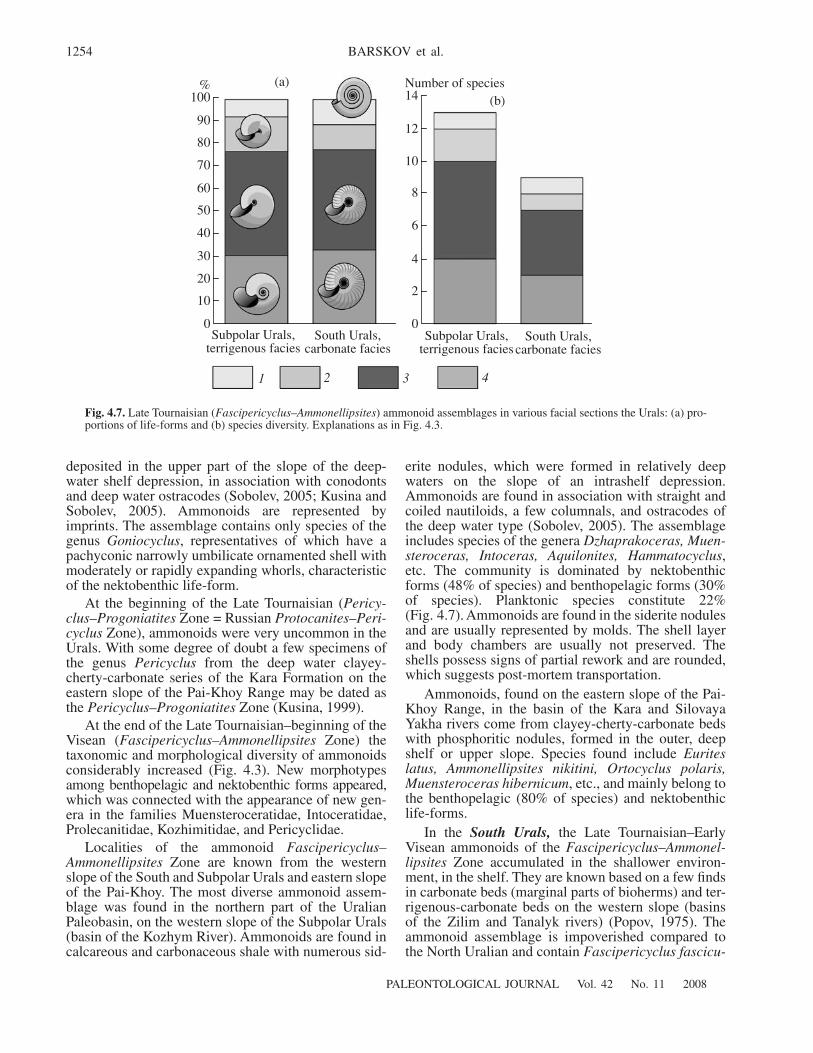
1254
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
deposited in the upper part of the slope of the deep-water shelf depression, in association with conodontsand deep water ostracodes (Sobolev, 2005; Kusina andSobolev, 2005). Ammonoids are represented byimprints. The assemblage contains only species of thegenus Goniocyclus, representatives of which have apachyconic narrowly umbilicate ornamented shell withmoderately or rapidly expanding whorls, characteristicof the nektobenthic life-form.
At the beginning of the Late Tournaisian (Pericy-clus–Progoniatites Zone = Russian Protocanites–Peri-cyclus Zone), ammonoids were very uncommon in theUrals. With some degree of doubt a few specimens ofthe genus Pericyclus from the deep water clayey-cherty-carbonate series of the Kara Formation on theeastern slope of the Pai-Khoy Range may be dated asthe Pericyclus–Progoniatites Zone (Kusina, 1999).
At the end of the Late Tournaisian–beginning of theVisean (Fascipericyclus–Ammonellipsites Zone) thetaxonomic and morphological diversity of ammonoidsconsiderably increased (Fig. 4.3). New morphotypesamong benthopelagic and nektobenthic forms appeared,which was connected with the appearance of new gen-era in the families Muensteroceratidae, Intoceratidae,Prolecanitidae, Kozhimitidae, and Pericyclidae.
Localities of the ammonoid Fascipericyclus–Ammonellipsites Zone are known from the westernslope of the South and Subpolar Urals and eastern slopeof the Pai-Khoy. The most diverse ammonoid assem-blage was found in the northern part of the UralianPaleobasin, on the western slope of the Subpolar Urals(basin of the Kozhym River). Ammonoids are found incalcareous and carbonaceous shale with numerous sid-
erite nodules, which were formed in relatively deepwaters on the slope of an intrashelf depression.Ammonoids are found in association with straight andcoiled nautiloids, a few columnals, and ostracodes ofthe deep water type (Sobolev, 2005). The assemblageincludes species of the genera Dzhaprakoceras, Muen-steroceras, Intoceras, Aquilonites, Hammatocyclus,etc. The community is dominated by nektobenthicforms (48% of species) and benthopelagic forms (30%of species). Planktonic species constitute 22%(Fig. 4.7). Ammonoids are found in the siderite nodulesand are usually represented by molds. The shell layerand body chambers are usually not preserved. Theshells possess signs of partial rework and are rounded,which suggests post-mortem transportation.
Ammonoids, found on the eastern slope of the Pai-Khoy Range, in the basin of the Kara and SilovayaYakha rivers come from clayey-cherty-carbonate bedswith phosphoritic nodules, formed in the outer, deepshelf or upper slope. Species found include Euriteslatus, Ammonellipsites nikitini, Ortocyclus polaris,Muensteroceras hibernicum, etc., and mainly belong tothe benthopelagic (80% of species) and nektobenthiclife-forms.
In the South Urals, the Late Tournaisian–EarlyVisean ammonoids of the Fascipericyclus–Ammonel-lipsites Zone accumulated in the shallower environ-ment, in the shelf. They are known based on a few findsin carbonate beds (marginal parts of bioherms) and ter-rigenous-carbonate beds on the western slope (basinsof the Zilim and Tanalyk rivers) (Popov, 1975). Theammonoid assemblage is impoverished compared tothe North Uralian and contain Fascipericyclus fascicu-
0
10
20
40
50
30
60
70
80
90
100% (a)
0
14Number of species
(b)
12
10
8
6
4
2
Subpolar Urals,terrigenous facies
South Urals,carbonate facies
Subpolar Urals,terrigenous facies
South Urals,carbonate facies
1 2 43
Fig. 4.7. Late Tournaisian (Fascipericyclus–Ammonellipsites) ammonoid assemblages in various facial sections the Urals: (a) pro-portions of life-forms and (b) species diversity. Explanations as in Fig. 4.3.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1255
latus, Pericyclus princeps, Muensteroceras sp., Orto-cyclus worki, and Polaricyclus rileyi, mainly belongingto the benthopelagic (32% of species) and nektobenthic(47%) life-forms. A few evolute and involute plank-tonic species are also present (Fig. 4.7).
A few general conclusions may be made based onthe analysis of the ammonoid evolution in the Tournai-sian in the Urals.
(1) As in the Late Devonian, Tournaisian ammonoidspreferred deep-water regions of the outer shelf: themost abundant (in the number of specimens) and taxo-nomically diverse ammonoid assemblages are found indeep-water terrigenous beds of the North Urals.
(2) A distinctive feature of the Tournaisian commu-nities is the predominance of nektobenthic (49%),while benthopelagic species are less diverse (27%), andplanktonic forms are uncommon.
(3) Two large stages may be recognized in the evo-lution of ammonoid communities of the Uralian basin,like in the Famennian: Early Tournaisian (Eocanites–Gattendorfia Zone) and a later interval from the MiddleTournaisian to Early Visean. Each of these stages, sep-arated by a crisis, had its own morphological and eco-logical community structure. At the first stage, morpho-logical diversity impoverished (only involute forms arepresent) with predominance of nektobenthic forms andslight presence of the planktonic species. Imitoceratidswith an undivided ventral lobe prevailed. In the Uralian,communities of this stage do not contain evolute formsfrom the order Prolecanitidae (genus Eocanites),widely distributed in the deeper water deposits of theRheno-Hercynian Basin, North Africa, and Pamir.Characteristically most Early Tournaisian ammonoidcommunities show a little presence of benthic forms,which, is apparently explained by the reduction of theseecotypes as a result of the crisis at Devonian–Carbonif-erous boundary.
At the second stage, goniatitids with a bipartite ven-tral lobe appeared and began their evolution. At thattime communities with an increased diversity of nekto-benthic and benthopelagic forms and a few planktonictaxa, appear. The increase in the morphological diver-sity was to a large extent connected with a rapid evolu-tion of ornamented forms of the family Pericyclidae,and with the appearance oxyconic, narrowly umbilicateforms in the family Intoceratidae. Compared to otherfamilies, Pericyclidae were not only the most taxonom-ically diverse (16 species, 8 genera) but are representedby various morphotypes and life-forms. This indicatesa high degree of differentiation of Uralian Pericyclidae,which agrees with the general trend in the evolution ofthe group, which reached its maximum diversificationat the end of the Tournaisian.
Visean
Ammonoids are found in all Visean genozones.They come from various localities in Novaya Zemlya,
Subpolar, and South Urals. This fauna was studied byLibrovitch (1938), Ruzhencev (1949a, 1949b, 1958,1966; Ruzhencev and Bogoslovskaya, 1971), Bog-oslovskaya (1966), Kusina (1974, 1980, 1983), Kusinaand Konovalova (2004), Kusina and Yatskov (1990,1999), Konovalova (2004), etc. Collections includeover 20000 specimens, from the Paleontological Insti-tute, Chernyshev TsNIGR Museum, and in the Depart-ment of Paleontology at the St. Petersburg State Uni-versity. The assemblage is considerably richer than theTournaisian one: 86 species, 38 genera, and 17 families.The morphospace considerably increased compared tothe Tournaisian and is delineated by the values of D =0.01–0.6; W = 1.31–2.8, and S = 0.35–3.1. In general,the main constituents of the communities of Viseanammonoids are pachyconic, discoconic or oxyconic,narrowly umbilicate nektobenthic (47% of species) andbenthopelagic forms (about 30% of species) (Fig. 4.8).The quantity of planktonic forms was low during mostof the Visean. Only in the terminal Visean their propor-tion in the communities increased up to 27% of species.The species diversity among planktonic forms gradu-ally increased by the end of the Visean, and at the sametime new planktonic morphotypes—species with lati-conic and aperticonic shells, of the new superfamilyNeoglyphiocerataceae appeared and became wide-spread.
The end of the Tournaisian–beginning of the Viseanin the Uralian basin were marked by a large regression,which reached its peak at the end of the Early Visean(Alekseev et al., 1996; Puchkov, 2000; Sobolev, 2005;etc.). The increased regression, which caused a reduc-tion in the outer shelf areas, led to almost complete dis-appearance of ammonoids. The extinctions primarilyaffected highly specialized planktonic and benthope-lagic forms, while nektobenthic forms decreased later.
The ammonoid diversity and structure of their com-munities changed considerably throughout the Visean(Fig. 4.3).
Fig. 4.8. Proportions of life-forms in Visean ammonoidassemblages in the Urals. Explanations as in Fig. 4.3.
29%22 species
47%36 species
21%16 species
1 2 3 4
3%2 species
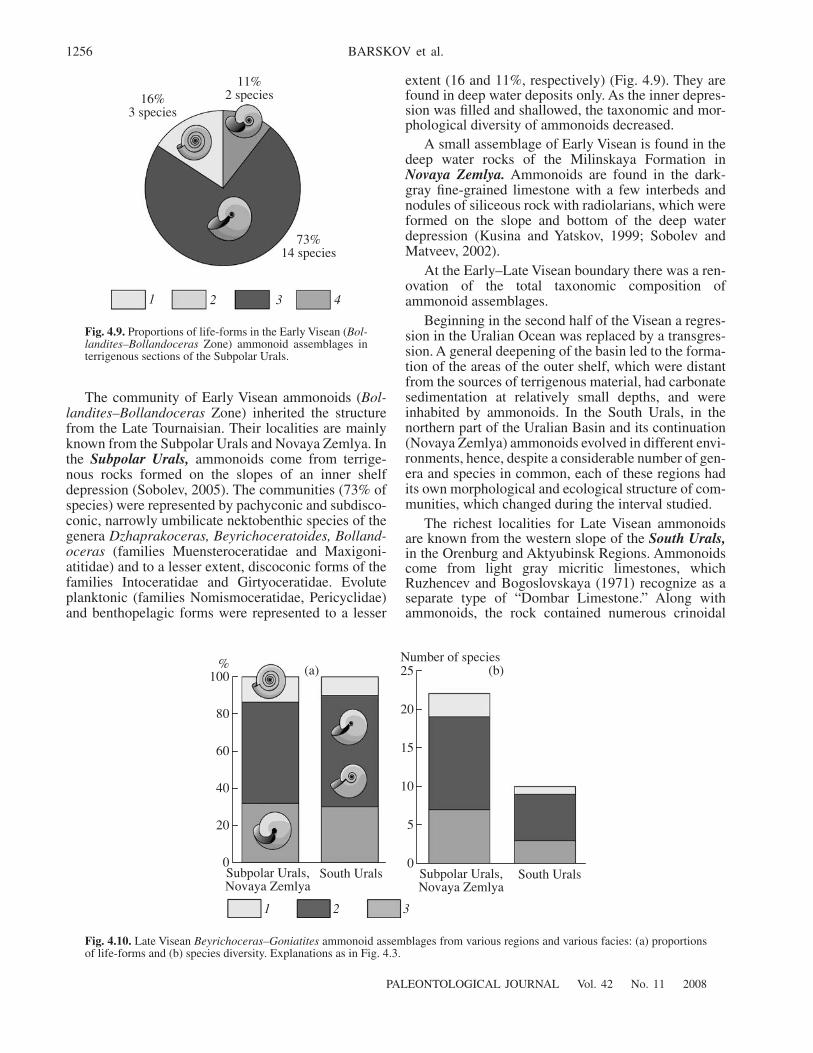
1256
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
The community of Early Visean ammonoids (Bol-landites–Bollandoceras Zone) inherited the structurefrom the Late Tournaisian. Their localities are mainlyknown from the Subpolar Urals and Novaya Zemlya. Inthe Subpolar Urals, ammonoids come from terrige-nous rocks formed on the slopes of an inner shelfdepression (Sobolev, 2005). The communities (73% ofspecies) were represented by pachyconic and subdisco-conic, narrowly umbilicate nektobenthic species of thegenera Dzhaprakoceras, Beyrichoceratoides, Bolland-oceras (families Muensteroceratidae and Maxigoni-atitidae) and to a lesser extent, discoconic forms of thefamilies Intoceratidae and Girtyoceratidae. Evoluteplanktonic (families Nomismoceratidae, Pericyclidae)and benthopelagic forms were represented to a lesser
extent (16 and 11%, respectively) (Fig. 4.9). They arefound in deep water deposits only. As the inner depres-sion was filled and shallowed, the taxonomic and mor-phological diversity of ammonoids decreased.
A small assemblage of Early Visean is found in thedeep water rocks of the Milinskaya Formation inNovaya Zemlya. Ammonoids are found in the dark-gray fine-grained limestone with a few interbeds andnodules of siliceous rock with radiolarians, which wereformed on the slope and bottom of the deep waterdepression (Kusina and Yatskov, 1999; Sobolev andMatveev, 2002).
At the Early–Late Visean boundary there was a ren-ovation of the total taxonomic composition ofammonoid assemblages.
Beginning in the second half of the Visean a regres-sion in the Uralian Ocean was replaced by a transgres-sion. A general deepening of the basin led to the forma-tion of the areas of the outer shelf, which were distantfrom the sources of terrigenous material, had carbonatesedimentation at relatively small depths, and wereinhabited by ammonoids. In the South Urals, in thenorthern part of the Uralian Basin and its continuation(Novaya Zemlya) ammonoids evolved in different envi-ronments, hence, despite a considerable number of gen-era and species in common, each of these regions hadits own morphological and ecological structure of com-munities, which changed during the interval studied.
The richest localities for Late Visean ammonoidsare known from the western slope of the South Urals,in the Orenburg and Aktyubinsk Regions. Ammonoidscome from light gray micritic limestones, whichRuzhencev and Bogoslovskaya (1971) recognize as aseparate type of “Dombar Limestone.” Along withammonoids, the rock contained numerous crinoidal
Fig. 4.9. Proportions of life-forms in the Early Visean (Bol-landites–Bollandoceras Zone) ammonoid assemblages interrigenous sections of the Subpolar Urals.
20
0
40
60
80
100%
5
0
10
15
20
25Number of species
Subpolar Urals,Novaya Zemlya
Subpolar Urals,Novaya Zemlya
South Urals South Urals
1 2 3
(a) (b)
Fig. 4.10. Late Visean Beyrichoceras–Goniatites ammonoid assemblages from various regions and various facies: (a) proportionsof life-forms and (b) species diversity. Explanations as in Fig. 4.3.
11%2 species
73%14 species
16%3 species
1 2 3 4

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1257
ossicles and benthic fauna of bivalves, gastropods, tri-lobites, tetracorals, brachiopods, and also deep-waterostracodes and conodonts. The development of themorphological and taxonomic diversity of ammonoidswas gradual.
In the Beyrichoceras–Goniatites genophase theassemblages were dominated by nektobenthic formswith pachyconic and discoconic involute and semi-involute shells (Figs. 4.3, 4.10). Species of the generaBeyrichoceras, Lusitanoceras, Goniatites, Girtyo-ceras, and Pronorites are typical representatives ofthese morphogroups. Species with a spheroconic orsubspheroconic involute shell, interpreted as benthope-lagic life-form were also widespread. Their massappearance was connected with the genus Goniatites(Goniatites sphaeroides, G. shimanskyi). The evolutecompressed shells of the families Prolecanitidae,Nomismoceratidae, interpreted as planktonic were lesswidespread. This structure of the assemblage appar-ently reflects the initial stages of restoration of thediversity in the community.
In the Hypergoniatites–Ferganoceras Genophasemorphological and taxonomic diversity of ammonoidsrapidly increased. Vast accumulations of their shells arefound together with numerous crinoids, conodonts, anda few gastropods, bivalves, trilobites, etc. (Ruzhencevand Bogoslovskaya, 1971). Shells are represented byvarious growth stages, are well preserved, not rounded,and many with fine ornamentation. Ammonoids comefrom crinoid or micritic limestones. Rock lithology,impoverished diversity of typical shallow-water organ-isms (corals, brachiopods, algae, and foraminifers),suggest that sedimentation took place in the outer deepwater part of the shelf, which is supported by the pres-ence of numerous crinoids. For crinoids, parts of the seafloor with stenohaline environment and good circula-tion (the conditions on the shelf) are the most suited(Krammer and Ausich, 2006). Nevertheless, in this casethe ideal model of a carbonate shelf (Ahr, 1973, 1998)was probably distorted and modified by a strong differ-entiation of the sea floor resulting from a continuingtectonic evolution of the South Urals (Puchkov, 2000).Territories inhabited by crinoids and ammonoids wereapparently semi-isolated from the neighboring shallowwater zones inhabited by corals and brachiopods.Apparently, these were quiet areas of the marine basin,near or on the margins of the subaqueous uplifts(Nikolaeva, 2006).
The assemblages are dominated by ammonoids withsubspheroconic and pachyconic, weakly ornamentedshells with slowly expanding whorls, not suitable foractive swimming. The assemblages, taxonomically andin number of individuals, contain benthopelagic (32%of species) and planktonic forms (27% of species) ofthe families Goniatitidae, Neoglyphioceratidae, Craveno-ceratidae, Delepinoceratidae, Ferganoceratidae, Rhym-moceratidae, etc. (Fig. 4.11). Nektobenthic forms arealso diverse (41 % of species), but the majority of spe-
cies that belonged to this life-form are represented inthe assemblages by a small number of individuals. Thenektobenthic life-form is represented by species of thegenera Kazakhoceras, Arcanoceras, Megapronorites,families Girtyoceratidae, Dimorphoceratidae, Goni-atitidae, Agathiceratidae, Prolecanitidae, etc.
An increase in the number of benthopelagic forms ismainly related to the rapidly evolving family Goniatiti-dae and superfamily Neoglyphiocerataceae. Theappearance of the latter is one of the most importantevolutionary events at the end of the Visean. Its devel-opment is also connected with an increase in the mor-phological diversity of planktonic forms. Representa-tives of this superfamily (families Cravenoceratidaeand Neoglyphioceratidae) form two new planktonicmorphotypes—laticones and aperticones, widely umbili-cate shells. These morphotypes were the most success-ful, which was responsible for the morphologicalappearance of the Late Carboniferous and Early Per-mian communities.
The evolution of ammonoids in the Beyrichoceras–Goniatites and Hypergoniatites–Ferganoceras geno-phases on the western slope of the South Urals occurredin similar environments. Transformations in ammonoidcommunities apparently were not determined by largeabiotic changes in the environment (change in the sed-imentary settings, sea level fluctuations, etc.) and mayreflect successive stages in the evolution of the commu-nity: (a) early evolution of the morphological diversity;(b) radiation and exploration of new niches.
On the eastern slope of the Urals (Verkhnyaya Kar-dailovka section), the evolution of Visean ammonoidsoccurred in the deeper water settings, which is indi-cated by radiolarians, deep water ostracodes, and by thespecific lithofacies (Pazukhin and Gorozhanina, 2002;Pazukhin et al., 2002). The ammonoid assemblages inthis region are very impoverished being represented bybenthopelagic and planktonic forms.
32%13 species
41%17 species
22%9 species
1 2 3 4
5%2 species
Fig. 4.11. Proportions of life-forms in the Late Visean(Hypergoniatites–Ferganoceras Genozone) ammonoidassemblages from the carbonate sections of the South Urals.Explanations as in Fig. 4.3.

1258
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
In the North Urals and Novaya Zemlya, the LateVisean ammonoids also existed on the outer margin ofthe carbonate shelf, but they lived in significantly shal-lower environment. In the Subpolar Urals they arefound in the bioherm structures similar to Walsourtianmounds on the quiet slope (Antoshkina, 2003; Kono-valova, 2004; Skompski et al., 2001). Ammonoids arefound in carbonate deposits together with abundantbenthic fauna brachiopods and bivalves, sometimesforming lens-shaped formations, with gastropods, bry-ozoans, corals, and trilobites (Konovalova and Sobolev,2005). Numerous remains of foraminifers and algae arealso present. In places, ammonoids form lens-like accu-mulations (cephalopod shellstones) and are representedby complete shells of varying size. The assemblages aredominated by spheroconic and pachyconic benthope-lagic and discoconic nektobenthic forms: Goniatitesolysya, Lusitanoceras kusinae, L. nadotense, Kazakho-ceras hawkinsi, Girtyoceras kazakhorum, etc.
Upper Visean ammonoids of Novaya Zemlya arefound in the deposits of the Milinskaya Formation andin the Gorbovsky Reefoid Massif, which formed in adifferent environments. The Milinskaya Formation iscomposed of mainly carbonate, fine-grained, micritic,cherty limestones with infrequent interbeds of clayey-cherty limestone and dolomites, scanty ammonoids,conodonts, and foraminifers. The accumulation of theUpper Visean beds apparently occurred on the slopeand on the bottom of a deep-sea depression (Platonovand Chernyak, 1982; Schecoldin et al., 1994, Sobolevand Matveev, 2002).
The ammonoid assemblage includes benthopelagic(56% of species), nektobenthic (31%) and a few plank-tonic forms. Ammonoids are also found in biohermlimestones of the Gorbovsky Reefoid Massif (BerkhIsland). The frame of the reef is composed of algal andcoral-algal limestones, occasionally overfilled withfauna. Ammonoids are found in association with typi-cally shallow-water benthic fauna: gastropods, brachi-opods, gigantic tetracorals, fusulinids, etc. (Kusina andYatskov, 1999). The specific composition of theammonoid assemblage from the bioherm deposits isimpoverished, mainly including spheroconic, pachy-conic, and oxyconic benthopelagic and nektobenthicforms of the families Goniatitidae, Berkhoceratidae,and Girtyoceratidae. Ophioconic planktonic forms(Nomismoceras) are uncommon and occur in deeperwater deposits of the Milinskaya Formation.
In general, the taxonomic diversity of the ammonoidassemblages from the Upper Visean of the northernUralian basin is low. The communities are dominatedby species, characterizing the nektobenthic life-form(54% of species). Benthopelagic forms are also rela-tively diverse (28%), whereas species of the planktonicadaptive type are few (18%) (Fig. 4.10).
Two stages can be recognized in the evolution of theVisean ammonoid communities of the Urals. Thesestages were separated by a crisis caused by a large
regression, which happened across the entire Uralianbasin and adjacent regions of the Russian Platform andPeri-Caspian at the Early–Late Visean boundary (Alek-seev et al., 1996; etc.).
The Early Visean ammonoid communities devel-oped in the same environments as the Late Tournaisianand had a similar morphological and ecological struc-ture (Figs. 4.7, 4.9). They mainly occurred in the north-ern region of the basin. During the Early Visean themorphological diversity of ammonoids decreased andmany taxa became extinct.
In the second half of the Visean ammonoid assem-blages were completely renewed while their communi-ties were significantly restructured, as they now devel-oped in shallower environments compared to the Tour-naisian and Famennian. At that time, poorly adapted toactive swimming benthopelagic and planktonic formsincreased in diversity and rapidly diversified. Aperti-cones and laticones appeared and became widespread,which is related to the appearance of the superfamilyNeoglyphiocerataceae. At the same time, the diversityamong nektobenthic forms, better adapted for swim-ming, decreased. At that time, large taxa (families oreven superfamilies) were characterized by a singlemorphotype or morphogroup, in contrast to the Tour-naisian, when species of several families (Pericyclidae,Gattendorfiidae) were represented by different morpho-groups. This feature may suggest that the groups werehighly specialized, i.e., communities were moremature.
The above analysis of the ammonoid assemblages ofthe western slope of the South Urals shows two succes-sive stages in the evolution of communities, unrelatedto fundamental changes in the environments: (a) earlyevolution of the diversity, (b) radiation and explorationof new niches. At this stage new taxa appeared, and newmorphotypes were developed, and all that determined thestructure of the Serpukhovian ammonoid communities.
Serpukhovian
In the Urals, ammonoids characterize two geno-zones of the Serpukhovian stage: Uralopronorites–Cravenoceras and Fayettevillea–Delepinoceras. Therichest localities of Serpukhovian ammonoids areknown from the South Urals (western slope). On theeastern slope of the South Urals ammonoids arerecorded from the Chelyabinsk Region (VerkhnyayaKardailovka section). In addition, ammonoids arefound on Berkh Island (Novaya Zemlya). Ammonoidcollections contain over 32000 specimens, housed inthe Paleontological Institute, Russian Academy of Sci-ences, Chernyshev TsNIGR Museum, St. Petersburg,and in the Paleontology Department of St. PetersburgState University. The systematic composition of theSerpukhovian ammonoid faunas was studied byRuzhencev (1947a, 1947b, 1949a, 1949b, 1956a, 1958,1965, etc.), Librovitch (1941, 1957), Ruzhencev and

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1259
Bogoslovskaya (1971), Kusina and Yatskov (1990,1999), Nikolaeva and Konovalova in Pazukhin et al.(2002), etc.
In the Serpukhovian time, the taxonomic diversityof ammonoids considerably increased compared to theVisean. The Serpukhovian assemblage includes121 species, 53 genera, and 19 families.
The morphological diversity of ammonoids at thattime is also relatively high. The morphospace definedby the parameters W, S, and D is delimited by the val-ues of D = 0.01–0.6; W = 1.26–3.27; S = 0.34–1.81.The benthopelagic spheroconic and cadiconic formswith a narrow and moderately narrow umbilicus wererepresented by 32% of the overall species diversity.Nektobenthic (30% of species) were mainly repre-sented by pachyconic and discoconic shells, whereasoxyconic and platyconic forms with rapidly expandingwhorls were less common. Planktonic forms were quitediverse, both taxonomically and morphologically andwere represented by 38% of species in the communi-ties. In general, the ecological structure of communitiesof Serpukhovian ammonoids is similar to those of LateVisean communities (Fig. 4.12). The diversity amongplanktonic forms increased due to the radiation of thesuperfamily Neoglyphiocerataceae, primarily of thefamilies Cravenoceratidae and Neoglyphioceratidae.
One of the trends in the evolution of the planktonicforms in the communities was connected to an increasein the number of species with a wide platyconic or aper-ticonic widely umbilicate shell, and also the appearanceof dwarf (not more than 20 mm in diameter) involutespecies. The appearance among the benthopelagicforms of various cadiconic and subcadiconic shells inthe second half of the Serpukhovian, connected withthe appearance of the families Glaphyritidae andStenoglaphyritidae, was also important.
At the Visean–Serpukhovian boundary no new high-rank taxa or new morphotypes appeared. In addition,the Uralopronorites–Cravenoceras Genophase charac-teristically shows a higher species and generic diver-sity, in particular among Neoglyphioceratida (nine newgenera), which became widespread.
The morphological and taxonomic diversity ofammonoids remained high during the entire Ser-pukhovian, reaching its maximum in the Early Ser-pukhovian (Uralopronorites–Cravenoceras Geno-zone). The maximum species diversity coincides withthe transgression at the beginning of the Serpukhovian,across the territory of the Urals. At the end of the Ser-pukhovian the taxonomic diversity decreased some-what. The latter was, apparently, caused by a decreasein the area of the shelf in the course of a regression, themaximum of which was at the end of the Ser-pukhovian–beginning of the Bashkirian (Pazukhinet al., 2002).
Ammonoid localities can be divided into threegroups, differing in taxonomic composition and lithol-ogy of the host rock. These are (1) western slope of the
South Urals, (2) eastern slope of the South Urals, and(3) Novaya Zemlya.
On the western slope of the South Urals(Aktyubinsk and Orenburg Regions) communities ofSerpukhovian ammonoids existed in the environmentsof the outer deep shelf. Their finds are known from thesame “Dombar limestone,” as the earlier assemblage ofthe Hypergoniatites–Ferganoceras Genozone. In gen-eral, they retained major evolutionary trends that wereoutlined in the Late Visean. The communities aremainly composed of benthopelagic (29% of species)and planktonic (33% of species) forms (Figs. 4.13a,4.13b). The diversity of planktonic forms somewhatincreased, mainly as a result of the appearance of newgenera and species in the families Rhymmoceratidae,Neoglyphioceratidae, and, especially, in the familyCravenoceratidae. The representatives of this familyare dominant among planktonic forms both in terms ofspecies diversity and in the number of individuals. Inthe family Girtyoceratidae, representatives of whichmainly belonged to the nektobenthic life-form, a newgenus Tumulites had a subspheroconic shell character-istic of benthopelagic forms. The diversity of nekto-benthic forms, most of which were represented by spe-cies continued from the Visean, continued to slowlydecrease.
The changes in the Early Serpukhovian ammonoidcommunities of the western slope of the South Uralswere likely to have been caused by internal factors,because no serious lithological changes have beenrecorded in the section around that time. They can beconsidered as the next stage in the evolution of the com-munities—the phase of maximum diversification withthe maximum number of species and morphologicaldiversity.
Beginning in the second half of the Serpukhoviantime, the sea level on the western margin of the Uralianbasin dropped, a process caused by the intensified orog-eny of the Urals (Puchkov, 2000). A simultaneous cool-
32%36 species
30%35 species
28%32 species
1 2 3 4
10%12 species
Fig. 4.12. Proportions of life-forms in the Serpukhovianammonoid assemblages of the Urals. Explanations as inFig. 4.3.
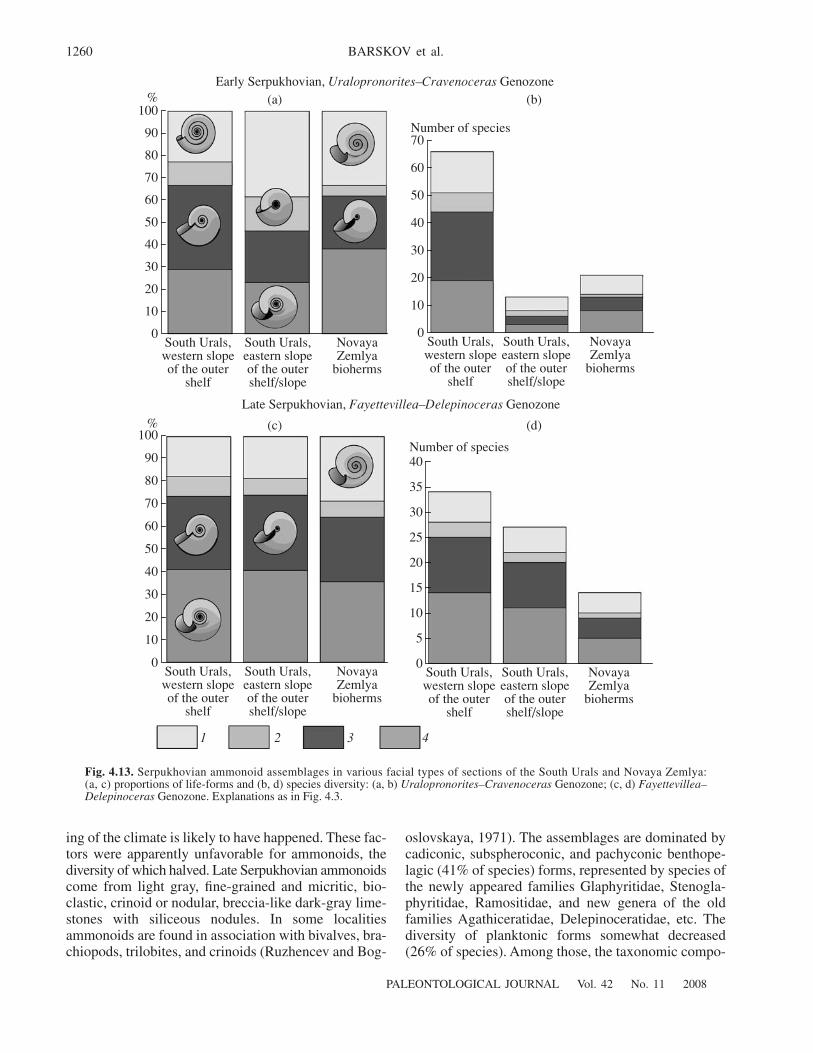
1260
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
ing of the climate is likely to have happened. These fac-tors were apparently unfavorable for ammonoids, thediversity of which halved. Late Serpukhovian ammonoidscome from light gray, fine-grained and micritic, bio-clastic, crinoid or nodular, breccia-like dark-gray lime-stones with siliceous nodules. In some localitiesammonoids are found in association with bivalves, bra-chiopods, trilobites, and crinoids (Ruzhencev and Bog-
oslovskaya, 1971). The assemblages are dominated bycadiconic, subspheroconic, and pachyconic benthope-lagic (41% of species) forms, represented by species ofthe newly appeared families Glaphyritidae, Stenogla-phyritidae, Ramositidae, and new genera of the oldfamilies Agathiceratidae, Delepinoceratidae, etc. Thediversity of planktonic forms somewhat decreased(26% of species). Among those, the taxonomic compo-
100
South Urals,0
Late Serpukhovian, Fayettevillea–Delepinoceras Genozone
70
80
90
60
50
40
30
20
10
%Early Serpukhovian, Uralopronorites–Cravenoceras Genozone
western slopeof the outer
shelf
South Urals,eastern slopeof the outershelf/slope
NovayaZemlya
bioherms
South Urals,0
70
60
50
40
30
20
10
western slopeof the outer
shelf
South Urals,eastern slopeof the outershelf/slope
NovayaZemlya
bioherms
Number of species
South Urals,0
40
35
25
20
15
10
5
western slopeof the outer
shelf
South Urals,eastern slopeof the outershelf/slope
NovayaZemlya
bioherms
Number of species
30
100
South Urals,0
70
80
90
60
50
40
30
20
10
%
western slopeof the outer
shelf
South Urals,eastern slopeof the outershelf/slope
NovayaZemlya
bioherms
1 2 3 4
(a) (b)
(d)(c)
Fig. 4.13. Serpukhovian ammonoid assemblages in various facial types of sections of the South Urals and Novaya Zemlya:(a, c) proportions of life-forms and (b, d) species diversity: (a, b) Uralopronorites–Cravenoceras Genozone; (c, d) Fayettevillea–Delepinoceras Genozone. Explanations as in Fig. 4.3.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1261
sition was considerably renewed. Laticonic widelyumbilicate forms disappeared. The number of nekto-benthic forms decreased (32%) (Figs. 4.13c, 4.13d).Despite being relatively diverse in species composition,the communities are usually represented by small num-ber of specimens.
On the eastern slope of the Urals (VerkhnyayaKardailovka) the evolution of ammonoids occurred inthe deeper environments of the outer slope of a carbon-ate platform. In the Early Serpukhovian, this areabecame somewhat shallower, and a few small bryo-zoan-serpulid bioherm buildups appeared (Pazukhinet al., 2002). The assemblage of Early Serpukhovianammonoids is less diverse compared to that of theDombar Limestone in western Kazakhstan, but is morediverse than that from the Upper Visean in the samesection. The communities are mainly composed ofplanktonic species (54% of species) of the families Cra-venoceratidae, Rhymmoceratidae, Neoglyphioceratidae,and Prolecanitidae; whereas the remaining part of thecommunity is represented by benthopelagic and nekto-benthic forms (Figs. 4.13a, 4.13b). Despite a high pro-portion of planktonic forms in the total diversity, thenumber of their shells in the assemblages is very low,and the communities were dominated by benthopelagicforms.
In the second half of the Serpukhovian the basinexisted in this territory deepened (Kulagina et al.,2001). The ammonoid assemblage is far more diverse,compared to the Early Serpukhovian (27 speciesinstead of 11, respectively). It was dominated by ben-thopelagic (40% of species) and planktonic (26% ofspecies) species of the families Agathiceratidae, Dele-pinoceratidae, Cravenoceratidae, Fayettevilleidae, Gla-phyritidae, Stenoglaphyritidae, etc. Nektobenthic forms(families Girtyoceratidae, Pronoritidae, Agathicer-atidae, etc.) constituted 34% of species of the diversity(Figs. 4.13c, 4.13d).
Novaya ZemlyaAmmonoid communities of Novaya Zemlya devel-
oped throughout the Serpukhovian in the environmentof a shallow-water bioherm that existed from the LateVisean to the Bashkirian. In the Early Serpukhovian thediversity of ammonoids increased considerably.Despite a noticeable renovation of the taxonomic com-position and reduction in diversity (23 species and 14,respectively), the ecological structure of ammonoidcommunities was not significantly changed at the tran-sition from the Early to Late Serpukhovian (Fig. 4.13).The assemblages were equally composed of pachy-conic, subspheroconic, and cadiconic narrowly umbili-cate benthopelagic and widely umbilicate aperticonicand laticonic planktonic forms (37 and 38% of species,respectively). They are represented by species of thefamilies Cravenoceratidae, Goniatitidae, Glaphyriti-dae, Fayettevilleidae, Delepinoceratidae, etc. Nekto-benthic forms were also relatively diverse (27% of spe-
cies diversity) and are represented by oxyconic, disco-conic, and, to a lesser extent, pachyconic narrowlyumbilicate forms of the families Berkhoceratidae,Dimorphoceratidae, Girtyoceratidae, etc. (Fig. 4.13).
The assemblages of Serpukhovian ammonoids fromNovaya Zemlya show a high degree of endemism at thespecies level (32 species of 37, over 80%), impover-ished taxonomic (at the species and especially genericlevel) and morphological diversity, and a high numberof taxa representing the nektobenthic life-form. Plank-tonic forms are dominated by aperticones, whereasophiocones and laticones are uncommon, whereasplanktonic small involute forms, widespread in thecommunities of the South Urals, are absent.
Despite the fact that the evolution of the Ser-pukhovian ammonoids occurred in the distant parts ofthe basin, several general patterns may be recognized.
(1) The main trend in the evolution of the communi-ties was the increase in the taxonomic and morpholog-ical diversity of planktonic and benthopelagic forms,which were dominant in the communities. In the sec-ond half of the Serpukhovian the number of benthope-lagic species with a cadiconic shell increased, mainlydue to the appearance of the new families, Glaphyriti-dae and Stenoglaphyritidae. Thus, in the Serpukhovianspecialization in the planktonic and benthopelagic seg-ments of community increased. At the same time, theproportion of the nektobenthic forms slightly decreased.As in the Late Visean, most families contain a singlemorphotype or morphogroup; i.e., each family repre-sents a single life-form.
(2) The evolution of the ammonoid communities ofthe Serpukhovian in the Urals retained trends that wererooted in the Late Visean, while the ammonoid habitatsdid not change much. Neither new high-rank taxa nornew morphotypes appeared at the Visean–Ser-pukhovian boundary. Changes in their ecological struc-ture in the Early Serpukhovian probably reflected thenext stage in the evolution of the communities—themaximum diversification phase with the highest num-ber of species and the highest morphological diversity.
(3) Serpukhovian ammonoids inhabited the outerpart of the open carbonate shelf. Their maximum diver-sity is registered in the southwestern regions of thebasin, where at that time communities of crinoids andammonoids developed on the outer, deeper shelf inquiet environments. In the north of the Uralian basin,ammonoids inhabited shallow-water settings near thebioherms. Their communities were less diverse bothtaxonomically, and morphologically.
(4) The ammonoid diversity at the beginning of theSerpukhovian and its reduction in the second half of theSerpukhovian coincides with the maximum highstandand subsequent sea level drop, which led to the reduc-tion in the area of the deeper outer shelf, primarily inthe western part of the basin.
At the end of the Early Carboniferous a large sealevel drop is recorded for the Urals. This regression

1262
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
resulted in the considerable reduction of the deeperouter shelf. At the same time the orogeny increasedleading to the appearance in the Bashkirian of a deeptrough in the Fore-Urals (western part of the basin). Thedevelopment of the fore-deep and increased processedof uplifting resulted in considerable changes in the sed-imentary settings and sea floor paleogeography(Khvorova, 1961; Puchkov, 2000; Kulagina et al.,2000; etc.) and caused a crisis in ammonoid communi-ties. At the Serpukhovian–Bashkirian boundary over60% of genera and families become extinct, while thearea of ammonoid distribution decreased considerablyand fundamental changes in the structure of the com-munities took place.
4.3.5. Pennsylvanian (Middle and Late Carboniferous)
Bashkirian
In the Urals, numerous ammonoids are found in thelower Bashkirian (Homoceras–Hudsonoceras, Reticu-loceras–Bashkortoceras, and Bilinguites–Cancello-ceras genozones). The riches localities of Bashkirianammonoids are known from the western and easternslopes of the South Urals, in the Aktyubinsk, Orenburg,and Chelyabinsk regions, and Bashkortostan. Theammonoid fauna was studied by Librovitch (1939a,1939b, etc.) Ruzhencev (1947a, 1947b, 1955, etc.),Ruzhencev and Bogoslovskaya (1978), Nikolaeva(1999; Nikolaeva in Kulagina et al., 2000, 2001, etc.).A few ammonoids were also described by Kusina andYatskov (1999) from the Bashkirian of Novaya Zemlya.Collections of Bashkirian ammonoids include over11600 specimens and are housed at the PaleontologicalInstitute, Russian Academy of Sciences, in ChernyshevTsNIGR Museum, and Department of Paleontology ofSt. Petersburg State University.
The Bashkirian assemblage is somewhat moreimpoverished than the Serpukhovian—106 species,
39 genera, and 15 families. Because of a decrease indiversity the morphological space of the assemblages issomewhat narrower: D = 0.02–0.55; W = 1.28–3.38;and S = 0.32–2.39.
The main trend in the morphological evolution ofammonoids at this stage was an increase in the numberof species with involute, rapidly expanding whorls anda narrow or moderately narrow umbilicus. These spe-cies, characterizing the nektobenthic life-form, consti-tute 48% of the total taxonomic diversity. Spheroconicand cadiconic benthopelagic forms were not abundant(19% of the total number of species). Evolute widelyumbilicate shells and small narrowly umbilicatespheroconic and pachyconic forms, constituting theplanktonic part of the community, show lower diversitycompared to that of the Serpukhovian (33% of speciesinstead of 38%), but were still numerous. Planktonicforms were now dominated by laticones and aperti-cones, whereas the number of ophioconic forms con-siderably decreased compared to that of the Ser-pukhovian. The species diversity among involuteplanktonic forms also decreased (Fig. 4.14).
The evolution of Bashkirian ammonoids occurred inconsiderably different environments compared to thatof the Serpukhovian, which was considerably reflectedby the structure of communities. At the Serpukhovian–Bashkirian boundary an extensive regression occurredin the South Urals. A large portion of the sedimentationarea in the South Urals in the Bashkirian represented arelatively shallow shelf, and only the Upper Bashkiriandeposits in the Zilair megasynclinorium are interpretedas deposits accumulated in the submerged margin of theshelf and continental slope (Kulagina et al., 2001).From this territory the earliest Lower Bashkirianammonoid assemblages are described by Nikolaeva(1999).
Localities of Bashkirian ammonoids occur both onthe western, and the eastern slope of the Urals. Each ofthese regions has a particular type of sedimentary set-tings, reflected in the structure of the communities.Ammonoid localities on the western slope of the Uralsbelong to the Central-Uralian facial zone. This zonewas a fore-mountain trough with carbonate-terrige-neous and terrigenous sediments, which began itsdevelopment at the beginning of the Bashkirianbetween the East European platform in the west and thearea of the basin presently covered by the Ural Moun-tains in the east. The deposits of the Lower Bashkirianyielding ammonoids belong to the Kuruil Formation(Khvorova, 1961), which is represented by thick seriesof carbonate rocks, interbedded with series of thinlyand medium-bedded dark limestone-cherty-clayeydeposits with rare interbeds of carbonate breccias.
The western slope of the South Urals contains themost diverse ammonoid assemblage. Three LowerBashkirian ammonoid genozones are recognized. In thelowermost Homoceras–Hudsonoceras Genozone, theassemblages mainly contain discoconic and pachy-
19%20 species
48%50 species
25%27 species
1 2 3 4
8%9 species
Fig. 4.14. Proportions of life-forms in the Bashkirianammonoid assemblages of the Urals. Explanations as inFig. 4.3.
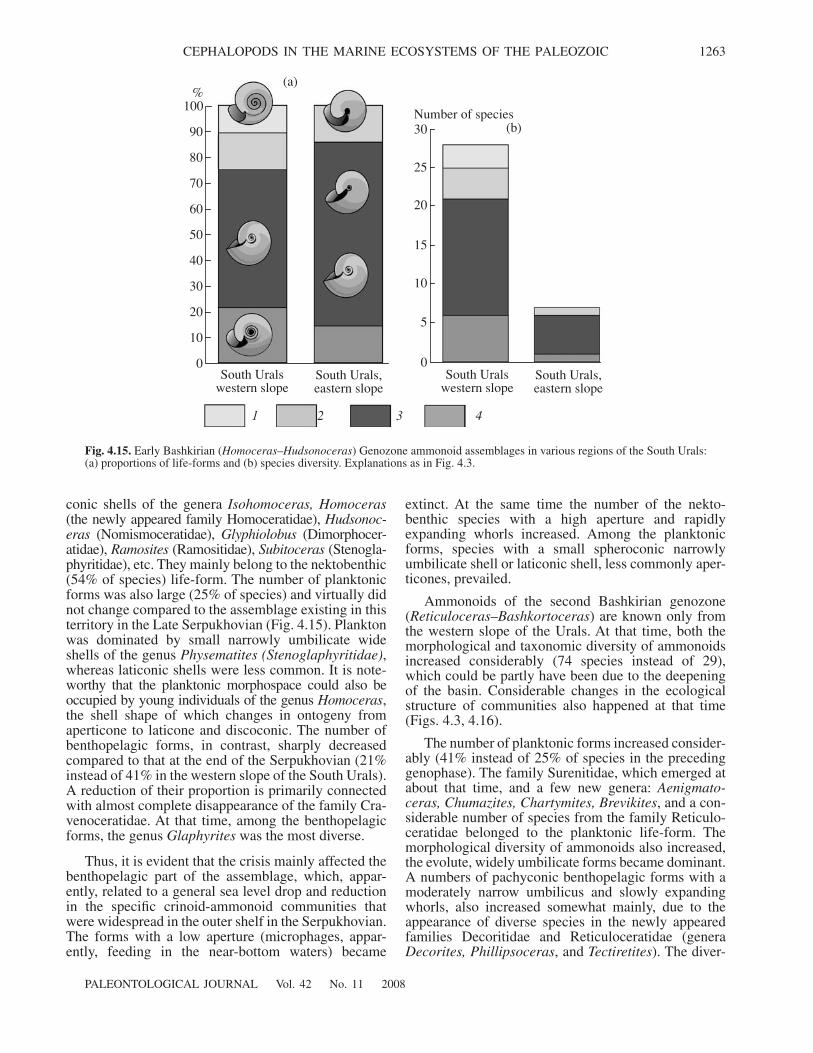
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1263
conic shells of the genera Isohomoceras, Homoceras(the newly appeared family Homoceratidae), Hudsonoc-eras (Nomismoceratidae), Glyphiolobus (Dimorphocer-atidae), Ramosites (Ramositidae), Subitoceras (Stenogla-phyritidae), etc. They mainly belong to the nektobenthic(54% of species) life-form. The number of planktonicforms was also large (25% of species) and virtually didnot change compared to the assemblage existing in thisterritory in the Late Serpukhovian (Fig. 4.15). Planktonwas dominated by small narrowly umbilicate wideshells of the genus Physematites (Stenoglaphyritidae),whereas laticonic shells were less common. It is note-worthy that the planktonic morphospace could also beoccupied by young individuals of the genus Homoceras,the shell shape of which changes in ontogeny fromaperticone to laticone and discoconic. The number ofbenthopelagic forms, in contrast, sharply decreasedcompared to that at the end of the Serpukhovian (21%instead of 41% in the western slope of the South Urals).A reduction of their proportion is primarily connectedwith almost complete disappearance of the family Cra-venoceratidae. At that time, among the benthopelagicforms, the genus Glaphyrites was the most diverse.
Thus, it is evident that the crisis mainly affected thebenthopelagic part of the assemblage, which, appar-ently, related to a general sea level drop and reductionin the specific crinoid-ammonoid communities thatwere widespread in the outer shelf in the Serpukhovian.The forms with a low aperture (microphages, appar-ently, feeding in the near-bottom waters) became
extinct. At the same time the number of the nekto-benthic species with a high aperture and rapidlyexpanding whorls increased. Among the planktonicforms, species with a small spheroconic narrowlyumbilicate shell or laticonic shell, less commonly aper-ticones, prevailed.
Ammonoids of the second Bashkirian genozone(Reticuloceras–Bashkortoceras) are known only fromthe western slope of the Urals. At that time, both themorphological and taxonomic diversity of ammonoidsincreased considerably (74 species instead of 29),which could be partly have been due to the deepeningof the basin. Considerable changes in the ecologicalstructure of communities also happened at that time(Figs. 4.3, 4.16).
The number of planktonic forms increased consider-ably (41% instead of 25% of species in the precedinggenophase). The family Surenitidae, which emerged atabout that time, and a few new genera: Aenigmato-ceras, Chumazites, Chartymites, Brevikites, and a con-siderable number of species from the family Reticulo-ceratidae belonged to the planktonic life-form. Themorphological diversity of ammonoids also increased,the evolute, widely umbilicate forms became dominant.A numbers of pachyconic benthopelagic forms with amoderately narrow umbilicus and slowly expandingwhorls, also increased somewhat mainly, due to theappearance of diverse species in the newly appearedfamilies Decoritidae and Reticuloceratidae (generaDecorites, Phillipsoceras, and Tectiretites). The diver-
0
10
20
40
50
30
60
70
80
90
100%
(a)
0
30Number of species
(b)
25
20
15
10
5
South Uralswestern slope
South Urals,eastern slope
1 2 43
South Urals,eastern slope
South Uralswestern slope
Fig. 4.15. Early Bashkirian (Homoceras–Hudsonoceras) Genozone ammonoid assemblages in various regions of the South Urals:(a) proportions of life-forms and (b) species diversity. Explanations as in Fig. 4.3.
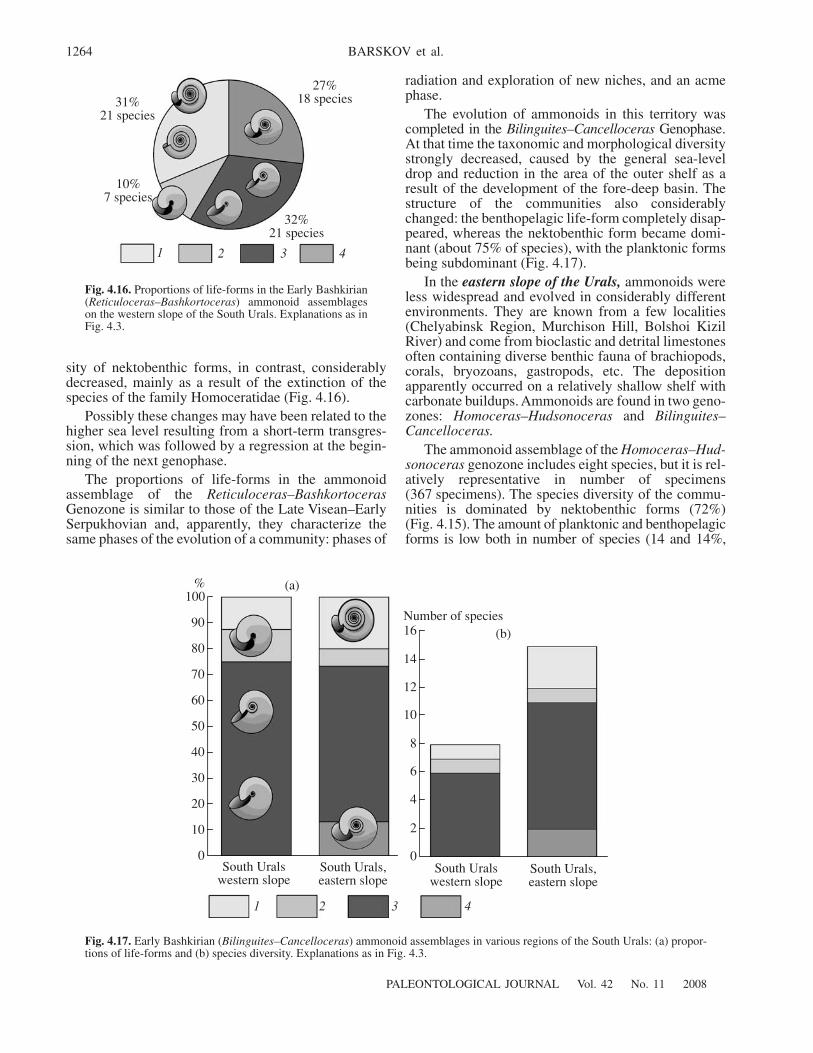
1264
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
sity of nektobenthic forms, in contrast, considerablydecreased, mainly as a result of the extinction of thespecies of the family Homoceratidae (Fig. 4.16).
Possibly these changes may have been related to thehigher sea level resulting from a short-term transgres-sion, which was followed by a regression at the begin-ning of the next genophase.
The proportions of life-forms in the ammonoidassemblage of the Reticuloceras–BashkortocerasGenozone is similar to those of the Late Visean–EarlySerpukhovian and, apparently, they characterize thesame phases of the evolution of a community: phases of
radiation and exploration of new niches, and an acmephase.
The evolution of ammonoids in this territory wascompleted in the Bilinguites–Cancelloceras Genophase.At that time the taxonomic and morphological diversitystrongly decreased, caused by the general sea-leveldrop and reduction in the area of the outer shelf as aresult of the development of the fore-deep basin. Thestructure of the communities also considerablychanged: the benthopelagic life-form completely disap-peared, whereas the nektobenthic form became domi-nant (about 75% of species), with the planktonic formsbeing subdominant (Fig. 4.17).
In the eastern slope of the Urals, ammonoids wereless widespread and evolved in considerably differentenvironments. They are known from a few localities(Chelyabinsk Region, Murchison Hill, Bolshoi KizilRiver) and come from bioclastic and detrital limestonesoften containing diverse benthic fauna of brachiopods,corals, bryozoans, gastropods, etc. The depositionapparently occurred on a relatively shallow shelf withcarbonate buildups. Ammonoids are found in two geno-zones: Homoceras–Hudsonoceras and Bilinguites–Cancelloceras.
The ammonoid assemblage of the Homoceras–Hud-sonoceras genozone includes eight species, but it is rel-atively representative in number of specimens(367 specimens). The species diversity of the commu-nities is dominated by nektobenthic forms (72%)(Fig. 4.15). The amount of planktonic and benthopelagicforms is low both in number of species (14 and 14%,
27%18 species
32%21 species
31%21 species
1 2 3 4
10%7 species
Fig. 4.16. Proportions of life-forms in the Early Bashkirian(Reticuloceras–Bashkortoceras) ammonoid assemblageson the western slope of the South Urals. Explanations as inFig. 4.3.
0
10
20
40
50
30
60
70
80
90
100% (a)
0
16Number of species
(b)
14
12
10
8
6
South Uralswestern slope
South Urals,eastern slope
1 2 43
South Urals,eastern slope
South Uralswestern slope
4
2
Fig. 4.17. Early Bashkirian (Bilinguites–Cancelloceras) ammonoid assemblages in various regions of the South Urals: (a) propor-tions of life-forms and (b) species diversity. Explanations as in Fig. 4.3.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1265
respectively) and in the number of specimens. In generalsuch ecological structure is characteristic of the crisisphase or of the initial stage of radiation in a community.
The evolution of ammonoids of the Bilinguites–Cancelloceras Genozone occurred in a similar environ-ment, which was reflected in the structure of the com-munities. In general, the proportions of the life-formsrepeat the structure of the communities of theHomoceras–Hudsonoceras Genozone. The proportionof planktonic forms somewhat increased (27% of spe-cies) (Fig. 4.17).
In general, ammonoid communities of the Bashkir-ian were unstable, rapidly changing, with high rate ofevolutionary processes, which can reflect the responseof the group to sharp changes in the environment(Figs. 4.3; 4.18). The second half of the Bashkirian wasmarked by a local (confined to the Urals) extinction ofammonoids, which was caused by sharp changes in theoverall structure of the basin, in connection with thedevelopment of the fore-deep basin.
Moscovian
During the Moscovian the ammonoid communitiesof the Urals entered a deep crisis, apparently related tothe total change in the settings in the whole of the Ura-lian basin following the development of the fore-deepbasin. Thick coarse-clastic and flysch formations accu-mulated in the eastern part of the fore-deep basin,whereas the western part was delimited by the steepslope of the carbonate platform. The Moscovian in thewestern slope of the South Urals is mainly character-
ized by foraminifers, which are used in the recent strati-graphic schemes (see Ivanova, 2000, 2002) and bybenthic fauna. Ammonoids in the Moscovian Stage arevery poorly represented, and are found only in the west-ern slope of the Urals and in Novaya Zemlya. The Mos-covian ammonoid assemblages were studied byRuzhencev (1951a, 1952a, 1955); Librovitch (1957),Kusina and Yatskov (1999), Popov (1975), etc. The col-lections are housed in the Paleontological Institute,Russian Academy of Sciences. Despite low speciesdiversity, ammonoids are represented by several mor-photypes, characterizing the nektobenthic, planktonic,and benthopelagic life-forms.
In the South Urals, it is difficult to reliably identifythe composition of the ammonoid assemblage of theMoscovian Stage, due to the uncertainty in the strati-graphic positions of the few finds. Various authors indi-cate the presence of the following species: Diabolo-ceras uralicum, Syngastrioceras orientale, Steno-pronorites karpinskii, and Pseudoparalegocerastzwetaevi. Tentatively, this assemblage may be supple-mented by Wellerites russiensis and Aktubites trifidus.A limestone boulder in the Ziyanchura breccia (Kasimov-ian) contained Eoasianites kajraklensis (Ruzhencev,1950); however, the dating of this boulder as the Mos-covian is doubtful because other ammonoids from thislocality are not referred to the Moscovian.
In Novaya Zemlya, Moscovian ammonoids (Diab-oloceras–Winslowoceras) Genozone are found inBerkh Island in the bedded carbonate series with aslight admixture of terrigenous material (KrestovskyHorizon). Ammonoids are represented by a few shells of
0
10
20
40
50
30
60
70
80
90
100% (a)
0
80Number of species
(b)
10
Homoceras–Hudsonoceras
Reticuloceras–Bashkortoceras
1 2 43
Billinguites–Cancelloceras
70
60
50
40
30
20
Fig. 4.18. Changes in the proportions of the life-forms (a) and species diversity (b) in the Early Bashkirian ammonoids from thelocalities on the western slope of the South Urals. Explanations as in Fig. 4.3.

1266
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
the species Pseudopronorites aquilonalis, Pseudobisa-toceras gorbovense, Glaphyrites vulgaris, and Diab-oloceras sp. To the north of this region in the RussianHaven there was a record of Winslowoceras sp. (Kusinaand Yatskov, 1999, identified by M.F. Bogoslovskaya).The deposits containing ammonoids also contain fora-minifers and Moscovian conodonts (Sobolev andMatveev, 2002). Despite a very impoverished taxo-nomic composition of ammonoids, the assemblagecontains four different morphotypes, representing twolife-forms. Pachyconic or subdiscoconic shells with avery narrow umbilicus and a subdiscoconic shell with amoderately narrow umbilicus and with flattened flanks(Prolecanites-like) are characteristic of the nekto-benthic life-form. The planktonic life-form (P-1) alsocontains two morphotypes: aperticonic widely umbili-cate and ophioconic, with triangularly coiled whorls.
Kasimovian
In the South Urals the Kasimovian is mainly repre-sented by clayey deposits with subdominant interbedsof clayey and sandy limestone. In the east of the region,the sections contain clayey breccias and pebbled lime-stones.
Kasimovian ammonoids of the Urals were studiedmostly by Ruzhencev (1950). The material on this stageincludes about 200 shells housed in the collection of thePaleontological Institute of the Russian Academy ofSciences (coll. nos. 319, 320).
The Kasimovian assemblage is one of the mostimpoverished (9 species, 9 genera, 9 families). Althoughthe taxonomic diversity is somewhat higher than in theMoscovian, the assemblage had still not recovered fromthe crisis. During the Kasimovian, a very slow recoveryof diversity is observed, mostly due to the nektobenthicpachyconic forms.
The total morphospace occupied by Kasimovianammonoids is small: D = 0.1–0.59; W = 1.5–2.5; S =0.4–2.4. In addition, the actual coverage of this mor-phological space is very small. Despite the extremelylow taxonomic diversity, Kasimovian ammonoids rep-resent a considerable number of morphotypes becauseof their scattered distribution in the morphospace.
Except the ancient Pronoritidae and Glaphyritidae,all the other families are newly appeared (Neoicocer-atidae, Agathiceratidae, Thalassoceratidae, Uddeniti-dae, Schistoceratidae, Shumarditidae, and Marathoniti-dae). It is essential that only one widely umbilicatemorphotype was present, represented by aperticonicEoasianites kajraklensis (Neoicoceratidae), which weinterpret as a benthopelagic form (its stratigraphic posi-tion was discussed above). The planktonic life-form isalso represented by only one spheroconic form, Kar-galites (Subkargalites) neoparkeri.
It is noteworthy that all Kasimovian ammonoidscome from a single limestone boulder in the lower partof the Ziyanchura breccia (Lower Gzhelian). This boul-
der was dated Kasimovian based on certain lithologicaland tectonic evidence (Ruzhencev, 1950, p. 16). Thus,if Ruzhencev’s view is accepted it has to be remem-bered that the distribution of the life-forms for the Kasi-movian is based on a single ammonoid associationfrom a single locality.
Gzhelian
In the Gzhelian, the geodynamic structure of theUralian Fore-Deep can be considered to be completelyformed. This is responsible for the transverse lithologi-cal and facial zonation typical of such structures.
Three major types of sections corresponding to dif-ferent sedimentary settings are recognized for theGzhelian (and for the overlying Permian stages). Thesedimentary settings in the Uralian basin were mostlycontrolled by the influx of clastic material from theeastern and western margins. From the east to the westthe following zones are recognized: (1) zone of verythick terrigenous series; (2) zone of considerably thin-ner clayey-carbonate deposits; and (3) zone of moder-ately thick carbonate formations. These zones corre-spond, respectively, to eastern orogenic margin, centralpart of the depression, and western platform margin.
In the South Urals, in the environment of active tec-tonics accompanying the collision of plates and eleva-tion of the Magnitogorsk meganticlinorium, accumula-tion of terrigenous sediments considerably exceededcarbonate deposition. On the eastern margin of thesouthern regions of the strait, bullion and boulder-richbreccias, conglomerates, and gravelites accumulated.The slope facies contain numerous traces of turbiditeflows and slides. In the central zone of the basin clayey-sandy rocks contain large boulders, brought along theslope from the east. In the west of the strait, carbonateswith frequent interbeds of coarse limestone brecciaswere accumulated. This indicates that the Gzhelian,especially its first half, was marked by a series of earth-quakes. The earthquakes were caused by sliding alongthe contact zone of the plates. The active tectonic move-ments were accompanied by noticeable volcanic activ-ity. The lower horizons of the Gzhelian contain manyash layers originated from the eastern slope (Khvorova,1961).
Gzhelian ammonoids are relatively numerous inthin layers of marls and sandy limestones in coarse-grained flyschoid series on the eastern slope of thedepression and are virtually unknown from the areas ofdistribution of cherty-carbonate-clayey and carbonaterocks of the central and western zones. The affinity ofammonoids to the regions of flyschoid sedimentationapparently explains the almost complete absence ofammonoids to the north of the Belaya River basin, anarea where, at that time, carbonate mud was mostlydeposited.
On the other hand, the remains of the benthic faunamostly come from carbonates of the western, platform

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1267
facial zone. Apart from ammonoids, fusulinids are theonly fossils that are found in considerable quantities inflyschoid rocks. However, ammonoids and fusulinidsdo not usually occur together in the same bed. Majorfinds of ammonoids come from carbonate members offlyschoid cycles, whereas fusulinids come from terrig-enous members. This suggests different time and placeof their habitats. A similar pattern of the facial affinityof fossil groups was observed to the end of the basin’sexistence.
Gzhelian ammonites were studied by Karpinsky(1874), Ruzhencev (1950), Bogoslovskaya and Popov(1986a, 1986b), and Borisenkov (2002, 2003, 2004a,2004b). Material on Gzhelian ammonoids includesover 5000 specimens, housed in the collection of thePaleontological Institute nos. 319, 320; TsNIGRMuseum, coll. no. 12257, etc.
The Gzhelian assemblage includes 44 species of24 genera of 16 families and has a complex ecologicalstructure. The total morphospace increased consider-ably: D = 0.1–0.62; W = 1.4–3,7; S = 0.3–3.29, whichis somewhat larger than in the Serpukhovian, althoughthe taxonomic diversity is much lower. However, it canbe positively suggested that the ecological structure ofthe community recovered almost completely. The mor-phological distribution is more or less bimodal, withpeaks separated at approximately D = 0.18. Ammonoidswith lesser values of D belonged to the nektobenthiclife-form, while the group with high values of D is rep-resented by planktonic and benthopelagic forms.
The first morphogroup includes the families Pronor-itidae, Medlicottiidae, Uddenitidae (all Prolecanitida),Thalassoceratidae, Neodimorphoceratidae, Agathicer-atidae, Pseudohaloritidae, Adrianitidae, Vidrioceratidae,and Marathonitidae. This group comprises narrowlyumbilicate morphotypes with moderately and rapidlyexpanding whorls, which we interpret as nektobenthicforms.
The second group comprises species of the familiesShumarditidae, Glaphyritidae, Daraelitidae, Schisto-ceratidae, Somoholitidae, and Neoicoceratidae. Thesemorphotypes can be interpreted as planktonic and ben-thopelagic adaptive types.
Despite the relatively high morphological diversityof Gzhelian ammonoids, it has to be said that theassemblage contains very few compressed morpho-types with a medium-sized or wide umbilicus—platy-cones and ophiocones.
The ecological distribution of the Gzhelian assem-blage is unusual. The nektobenthic and benthopelagicgroups are represented as follows: 48% of species—nektobenthic and 27%—benthopelagic. The remainingportion of the assemblage is interpreted as planktonic:18% of species—evolute forms, 7%—involute forms(Fig. 4.19).
Ophioconic widely umbilicate forms are repre-sented by a single form only (Eoasianites vodorezovi),with the whorl width considerably exceeding typical
representatives of this morphotype in the youngerassemblages. E. vodorezovi is apparently an ancestor ofthe genus Svetlanoceras (Borisenkov, 2003), whichwas initial in the family Paragastrioceratidae, represen-tatives of which belong to the morphotype of widelyumbilicate shells with slowly expanding whorls.
One of the most ammonoid-rich sections is locatednear the village of Nikol’skoe on the watershedbetween the Sakmara and Urals rivers. This sectionbelongs to the clayey type of sequences, in whichammonoids are usually rarely found, and therefore it isof particular interest. Numerous ammonoids are foundin brownish, compact clay with layers of marly nodules(Ruzhencev, 1950). The assemblage is represented by20 species, among which 57% represent nektobenthic,32%—benthopelagic, and 11%—planktonic ophio-conic forms (Fig. 4.20). This distribution of life-formssuggest an unusual nature of this locality. A consider-able proportion of nektobenthic and benthopelagic spe-cies is usually characteristic of communities inhabitingrelatively shallow waters. This is contradicted by thedeep-water nature of this sequence largely composed ofclayey rocks. The absence of the involute planktonicforms (such as Vidrioceras borissiaki, Marathonitesuralensis, and Emilites plummeri) also remains unex-plained.
Flyschoid sequences on Aidaralash Creek are rich inammonoids. A large ammonoid material was collectedfrom several levels of sandy limestones and marl nod-ules. Of 22 species described from this section(Ruzhencev, 1950) 45% of species belong to the nekto-benthic life-form and 32% of species to the benthope-lagic life-form (Fig. 4.20). Planktonic forms are morenumerous, compared to assemblages from clayeysequences, and constitute 23% of species diversity.They are represented by both evolute (18%) and invo-lute (5%) morphotypes. In general, the ecologicalstructure is similar to that of the entire Gzhelianammonoid assemblage.
18%8 species 27%
12 species
48%21 species
7%3 species
1 2 3 4
Fig. 4.19. Proportions of life-forms in the Gzhelian ammonoidassemblage of the Urals. Explanations as in Fig. 4.3.
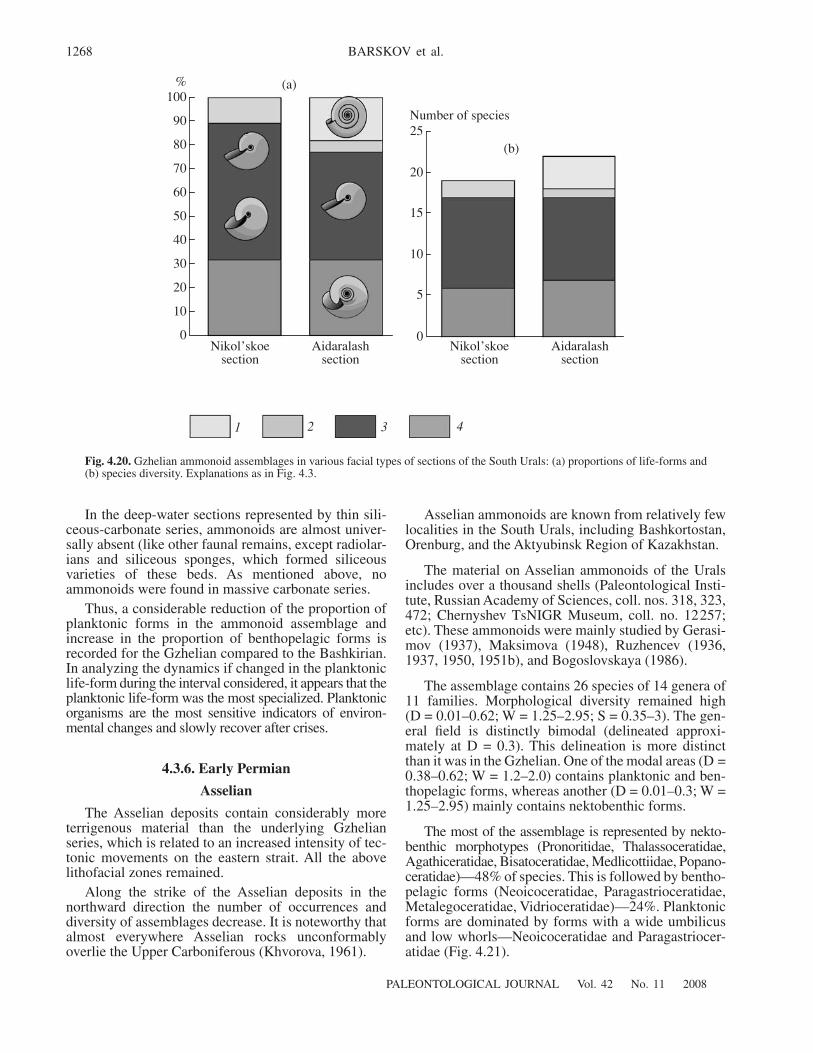
1268
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
In the deep-water sections represented by thin sili-ceous-carbonate series, ammonoids are almost univer-sally absent (like other faunal remains, except radiolar-ians and siliceous sponges, which formed siliceousvarieties of these beds. As mentioned above, noammonoids were found in massive carbonate series.
Thus, a considerable reduction of the proportion ofplanktonic forms in the ammonoid assemblage andincrease in the proportion of benthopelagic forms isrecorded for the Gzhelian compared to the Bashkirian.In analyzing the dynamics if changed in the planktoniclife-form during the interval considered, it appears that theplanktonic life-form was the most specialized. Planktonicorganisms are the most sensitive indicators of environ-mental changes and slowly recover after crises.
4.3.6. Early Permian
Asselian
The Asselian deposits contain considerably moreterrigenous material than the underlying Gzhelianseries, which is related to an increased intensity of tec-tonic movements on the eastern strait. All the abovelithofacial zones remained.
Along the strike of the Asselian deposits in thenorthward direction the number of occurrences anddiversity of assemblages decrease. It is noteworthy thatalmost everywhere Asselian rocks unconformablyoverlie the Upper Carboniferous (Khvorova, 1961).
Asselian ammonoids are known from relatively fewlocalities in the South Urals, including Bashkortostan,Orenburg, and the Aktyubinsk Region of Kazakhstan.
The material on Asselian ammonoids of the Uralsincludes over a thousand shells (Paleontological Insti-tute, Russian Academy of Sciences, coll. nos. 318, 323,472; Chernyshev TsNIGR Museum, coll. no. 12257;etc). These ammonoids were mainly studied by Gerasi-mov (1937), Maksimova (1948), Ruzhencev (1936,1937, 1950, 1951b), and Bogoslovskaya (1986).
The assemblage contains 26 species of 14 genera of11 families. Morphological diversity remained high(D = 0.01–0.62; W = 1.25–2.95; S = 0.35–3). The gen-eral field is distinctly bimodal (delineated approxi-mately at D = 0.3). This delineation is more distinctthan it was in the Gzhelian. One of the modal areas (D =0.38–0.62; W = 1.2–2.0) contains planktonic and ben-thopelagic forms, whereas another (D = 0.01–0.3; W =1.25–2.95) mainly contains nektobenthic forms.
The most of the assemblage is represented by nekto-benthic morphotypes (Pronoritidae, Thalassoceratidae,Agathiceratidae, Bisatoceratidae, Medlicottiidae, Popano-ceratidae)—48% of species. This is followed by bentho-pelagic forms (Neoicoceratidae, Paragastrioceratidae,Metalegoceratidae, Vidrioceratidae)—24%. Planktonicforms are dominated by forms with a wide umbilicusand low whorls—Neoicoceratidae and Paragastriocer-atidae (Fig. 4.21).
0
10
20
40
50
30
60
70
80
90
100% (a)
0
25Number of species
(b)
5
Nikol’skoe
1 2 43
10
15
20
Aidaralash Nikol’skoe Aidaralash
Fig. 4.20. Gzhelian ammonoid assemblages in various facial types of sections of the South Urals: (a) proportions of life-forms and(b) species diversity. Explanations as in Fig. 4.3.
section section section section
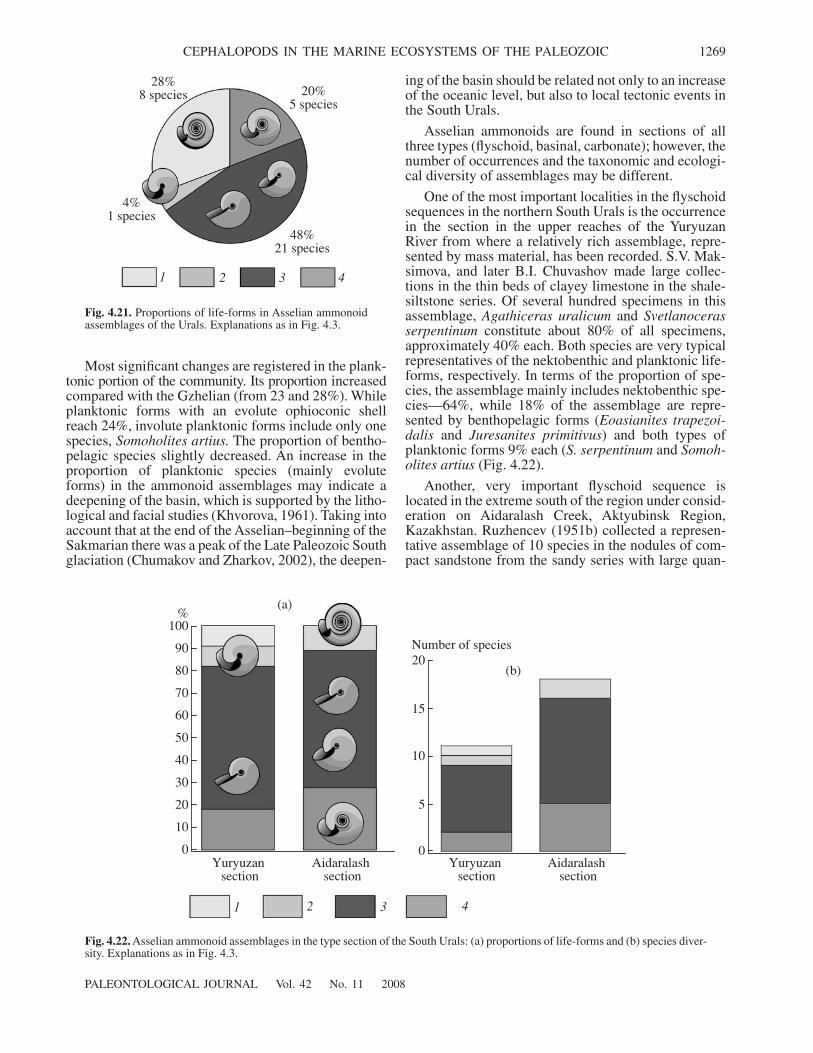
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1269
Most significant changes are registered in the plank-tonic portion of the community. Its proportion increasedcompared with the Gzhelian (from 23 and 28%). Whileplanktonic forms with an evolute ophioconic shellreach 24%, involute planktonic forms include only onespecies, Somoholites artius. The proportion of bentho-pelagic species slightly decreased. An increase in theproportion of planktonic species (mainly evoluteforms) in the ammonoid assemblages may indicate adeepening of the basin, which is supported by the litho-logical and facial studies (Khvorova, 1961). Taking intoaccount that at the end of the Asselian–beginning of theSakmarian there was a peak of the Late Paleozoic Southglaciation (Chumakov and Zharkov, 2002), the deepen-
ing of the basin should be related not only to an increaseof the oceanic level, but also to local tectonic events inthe South Urals.
Asselian ammonoids are found in sections of allthree types (flyschoid, basinal, carbonate); however, thenumber of occurrences and the taxonomic and ecologi-cal diversity of assemblages may be different.
One of the most important localities in the flyschoidsequences in the northern South Urals is the occurrencein the section in the upper reaches of the YuryuzanRiver from where a relatively rich assemblage, repre-sented by mass material, has been recorded. S.V. Mak-simova, and later B.I. Chuvashov made large collec-tions in the thin beds of clayey limestone in the shale-siltstone series. Of several hundred specimens in thisassemblage, Agathiceras uralicum and Svetlanocerasserpentinum constitute about 80% of all specimens,approximately 40% each. Both species are very typicalrepresentatives of the nektobenthic and planktonic life-forms, respectively. In terms of the proportion of spe-cies, the assemblage mainly includes nektobenthic spe-cies—64%, while 18% of the assemblage are repre-sented by benthopelagic forms (Eoasianites trapezoi-dalis and Juresanites primitivus) and both types ofplanktonic forms 9% each (S. serpentinum and Somoh-olites artius (Fig. 4.22).
Another, very important flyschoid sequence islocated in the extreme south of the region under consid-eration on Aidaralash Creek, Aktyubinsk Region,Kazakhstan. Ruzhencev (1951b) collected a represen-tative assemblage of 10 species in the nodules of com-pact sandstone from the sandy series with large quan-
28%8 species 20%
5 species
48%21 species
4%1 species
1 2 3 4
Fig. 4.21. Proportions of life-forms in Asselian ammonoidassemblages of the Urals. Explanations as in Fig. 4.3.
10
20
40
50
30
60
70
80
90
100%
(a)
Number of species
(b)
5
Yuryuzan
1 2 43
10
15
20
Aidaralash section section
0 0Yuryuzan
sectionAidaralash
section
Fig. 4.22. Asselian ammonoid assemblages in the type section of the South Urals: (a) proportions of life-forms and (b) species diver-sity. Explanations as in Fig. 4.3.

1270
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
tity of fusulinids. Bogoslovskaya and Popov (1986a,1986b) supplemented this record with another eightspecies. Unfortunately, Bogoslovskaya and Popov didnot indicate the actual number of shells. The analysis ofRuzhencev’s data gives the following distributions:specimens of nektobenthic taxa ≈ 60% (Boesites,Neopronorites, Artinskia, Agathiceras, Prothalasso-ceras, Aristoceras), benthopelagic ≈ 25% (Prost-ascheoceras, Glaphyrites), planktonic ≈ 15% (Eoasi-anites). It is important that the proportions of the life-forms based on quantity of species gives similar results:nektobenthic, 61%; benthopelagic, 28%; and plank-tonic, 11% (Fig. 4.22).
The sections discussed represent a typical exampleof flyschoid series and indicate relatively deep-waterenvironments at the foot of the eastern slope of thestrait. In the clayey-carbonate basinal sections typicalof the central regions of the strait (basins of the Sim,Assel, and Usolka rivers), ammonoids are uncommon,and their diversity is minimal (usually two or three spe-cies are represented by a few specimens). All knownsamples are to some extent accidental. Apparentlyammonoids did not live permanently in the mainstreamof the strait, where strong meridional currents couldhave existed, but entered this zone only occasionally.
In the reefoid sections, ammonoids are known onlyfrom Sterlitamak Shikhans (large fossil reefs). In gen-eral, it is known that ammonoids are rarely found in theextremely shallow-water facies, although sometimes afew specimens can be found in reef deposits.Ruzhencev (1951b) recorded an assemblage of 10 spe-cies from the Tra-Tau Shikhan. Except Neopronoritesrotundus (≈25%) and Agathiceras uralicum (≈50%)other species are represented by a few specimens. Thelife-forms are distributed as follows: nektobenthic—70%, benthopelagic—10%, and planktonic—20%. It isnoteworthy that Agathiceras uralicum almost alwayswhen found in a sample noticeably prevail over otherspecies in the number of specimens, sometimes consti-tuting over 50%. This applies to almost all stages andall localities. It is possible that specific adaptations ofthis species allow it to inhabit various environments,retaining considerable abundance everywhere.
Asselian assemblage is almost half of the Gzhelianand displays a somewhat different distribution of life-forms. Nektobenthic ammonoids dominate, represent-ing about half of the total diversity, and at the same timethe diversity of planktonic forms increases. The plank-ton shows a predominance of ophioconic and similarforms, whereas morphogroup 1A' (involute shells) isrepresented by a single species. The appearance of theophioconic morphotype in the Gzhelian and its furtherevolution in the Asselian is exclusively connected to thesuperfamily Neoicocerataceae. Furthermore, in thesubsequent epochs this life-form will be representedsolely by neoicoceratids.
Sakmarian
In the Sakmarian, flyschoid series continued accu-mulating in the Uralian Strait. By the end of the Sak-marian, the thickness and abundance of the terrigenousseries considerably increased. In the eastern coastalzone, thick underwater fans were formed. These fanscontinued the underwater flow systems, which existedfrom the Asselian. This suggests the existence of long-existing water flows running across the PaleouralianMountain Range. The underwater fans formed a goodsubstrate for coral reefs, being populated by abundantbenthos. Ammonoids preferred inhabiting deeper partsof the basin with flyschoid sedimentary settings.
General trends in the evolution of Sakmarianammonoids continued from the Asselian. No new fam-ilies appeared at this time. Evolution was restricted tothe generic and species levels, while Carboniferoustaxa continued to become extinct. As in the Asselian(and even to a greater extent), Sakmarian ammonoidswere more abundant and diverse in the south of theregion. Localities for Sakmarian ammonoids are notknown north of the Ufa River basin.
Material on Sakmarian ammonoids includes severalthousands shells, occurring mainly from the sections inBashkortostan, Chelyabinsk Region, and the Orenburgand Aktyubinsk regions of Kazakhstan (collection ofthe Paleontological Institute of the Russian Academy ofSciences, nos. 318, 323, 472, 590, etc.).
Sakmarian ammonoids of the Urals were mainlystudied by Ruzhencev (1938, 1951b, 1952b) and Mak-simova (1935, 1938).
The assemblage includes 34 species of 16 genera of10 families, i.e., taxonomically somewhat richer thanthe Asselian ones. The morphospace of the Sakmarianammonoids: D = 0.01–0.62; W = 1.22–3, S = 0.3–2.8.Although the total number of species is higher than inthe Asselian, the boundaries of the morphospaceremained the same. The distinctness of the groupsincreased. The group with D ≥ 0.24 (morphotypes witha wide and medium-sized umbilicus) is almost exclu-sively Neoicocerataceae—species of the genera Svetl-anoceras, Paragastrioceras, Uraloceras (Paragastrio-ceratidae), and Metalegoceras (Metalegoceratidae).
These assemblages are dominated by nektobenthicand planktonic forms. The proportion of benthopelagicspecies changed considerably. A large portion of theassemblage is represented by nektobenthic morpho-types (Pronoritidae, Medlicottiidae, Daraelitidae, Thalas-soceratidae, Agathiceratidae, Popanoceratidae, someParagastrioceratidae)—46%. The Sakmarian commu-nities show more even distribution of planktonic life-forms, plankton-1 (evolute forms with low whorls) con-stitutes 18% (Paragastrioceratidae), plankton-2 (invo-lute forms with wide whorls)—15% (Adrianitidae)(Fig. 4.23). We also refer to this life-form the species ofthe genus Sakmarites, because they are always repre-sented by small shells. The smallest proportion is rep-

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1271
resented by benthopelagic forms—21% (Metalegocer-atidae, Popanoceratidae, Vidrioceratidae).
The proportion of nektobenthic and benthopelagicforms is similar to that of the Asselian assemblage. Atthe same time, despite the Sakmarian regression, theproportion of planktonic forms increased, mainly dueto the involute forms. The shallowing of the basin wasapparently not sufficient to seriously affect planktonicforms.
Ammonoid occurrences are mainly found in thincarbonate interbeds in flyschoid series, carbonate nod-ules in clayey series and limestones and sandy lime-stones, frequently in limestone lenses. Ammonoids arerarely found in association with other fossils. Forinstance, the interbeds of compact micritic limestonewith ammonoid shells contain layers and lenses ofdetrital limestone with fusulinids and bryozoans. Thelatter possess traces of transportation. In other casesammonoids are found in black marl nodules in shalewith fusulinids. Judging from the facial preferences ofammonoids, at that time they mostly inhabited relativelydeep zones with carbonate-clayey substrates at the foot ofthe underwater fans and on the local uplifts with clayey-sandy bottom in the central zone of the strait.
One of the richest Sakmarian ammonoid assem-blages is described by Ruzhencev (1951b) (the samplecontains almost 500 specimens of 12 species) fromsandy limestones in the Ultugan-Sai Gully, in the basinof the Aktasty River, Aktyubinsk Region of Kazakh-stan. The sample is dominated by specimens of Agath-iceras uralicum, Neopronorites tenuis (36 and 26%, intotal 62%), i.e., nektobenthic forms prevail in thetaphocoenosis. At the same time, planktonic species arerepresented by evolute paragastrioceratids, constitutingonly slightly more than 2% of all specimens. When theabundances of life-forms are compared by the numberspecies they include, nektobenthic forms prevail (50%species), whereas the proportion of planktonic forms(35% species) is considerably different from the pro-portion based on the number of specimens (Fig. 4.24).The number of benthopelagic forms is almost half ofthat of planktonic, and a third of that of nektobenthic.Apparently, in this case, the conditions in a relativelyshallow marine basin with considerable carbonate pre-cipitation and moderate influx of clastics were favor-able for a nektobenthic ammonoid assemblage.
The sections from the Sakmarian type region—rightbank of the Sakmara River near the village of Kon-durovka (Mt. Kurmaya). Ammonoids are found in bedsof gray micritic, in places cherty limestone in the UpperSakmarian limestone-clayey series (Ruzhencev, 1951b).The assemblage, composed of 16 species, contains37% of nektobenthic forms (less that average for thestage, 46%), 19% of benthopelagic forms (close toaverage), 19% of planktonic subspherical forms, and25% of planktonic evolute forms (Fig. 4.24). In thisassemblage the proportion of all planktonic species
(44%) is higher than the average for the stage (33%).Apparently, the ammonoid-bearing beds accumulatedin the more basinal parts of the basin.
The terminal part of the Sakmarian contains numer-ous ammonoids in the Verkhnii Ozernyi Section 10–15 km to the south of the previously described locality.In the topmost part of the stage, the series of sandstonesinterbedding with platy limestones contains bullionsnear the basal part of the bed, overfilled by ammonoidshells. The assemblage contains eight species (Levenet al., 2002). Apparently, the same bed was the sourceof the sample described by Ruzhencev (1951b), whichsupplements this list with three more species names. Inthe assemblage, the proportion of benthopelagic spe-cies (Metalegoceras, Andrianovia) is higher than theaverage for the stage—27% instead of 21%, and interms of the number of specimens, benthopelagic formsalso constitute about 30%. At the same time, the pro-portion of involute planktonic forms (plankton-2) wasreduced to a total of 9%. The proportion of evoluteplanktonic forms (Paragastrioceras, Uraloceras) is18% and the proportion of nektobenthic (46%) corre-sponds to the average percentage (Fig. 4.24).
This assemblage is the closest to the assemblageaverage for the whole of the Sakmarian. Apparently, theVerkhnii Ozernyi Section can be considered to repre-sent a typical ammonoid habitat for this time. The sec-tion consists of a series of thick terrigenous members(sandstones and siltstones—tens of meters) and thincarbonates (rarely more than 0.5 m).
It is noteworthy that in the stratotype region, theupper and the lower parts of the Sakmarian are faciallydifferent. The Lower Sakmarian is represented by car-bonate-clayey beds interlayered by calcareous breccia.A similar type of sedimentation is more likely charac-teristic of the foot of the western margin. The uppersubstage is represented by flyschoids dominated by
18%6 species 21%
7 species
46%16 species
15%5 species
1 2 3 4
Fig. 4.23. Proportions of life-forms in the Sakmarianammonoid assemblage of the Urals. Explanations as inFig. 4.3.
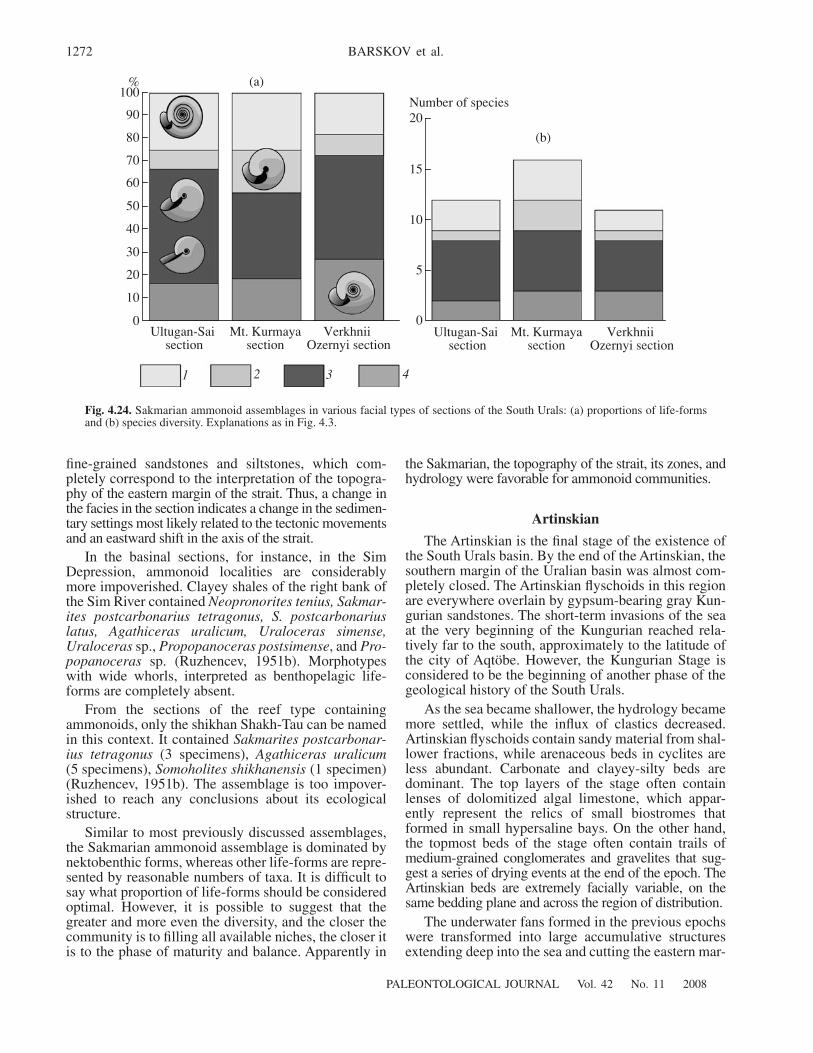
1272
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
fine-grained sandstones and siltstones, which com-pletely correspond to the interpretation of the topogra-phy of the eastern margin of the strait. Thus, a change inthe facies in the section indicates a change in the sedimen-tary settings most likely related to the tectonic movementsand an eastward shift in the axis of the strait.
In the basinal sections, for instance, in the SimDepression, ammonoid localities are considerablymore impoverished. Clayey shales of the right bank ofthe Sim River contained Neopronorites tenius, Sakmar-ites postcarbonarius tetragonus, S. postcarbonariuslatus, Agathiceras uralicum, Uraloceras simense,Uraloceras sp., Propopanoceras postsimense, and Pro-popanoceras sp. (Ruzhencev, 1951b). Morphotypeswith wide whorls, interpreted as benthopelagic life-forms are completely absent.
From the sections of the reef type containingammonoids, only the shikhan Shakh-Tau can be namedin this context. It contained Sakmarites postcarbonar-ius tetragonus (3 specimens), Agathiceras uralicum(5 specimens), Somoholites shikhanensis (1 specimen)(Ruzhencev, 1951b). The assemblage is too impover-ished to reach any conclusions about its ecologicalstructure.
Similar to most previously discussed assemblages,the Sakmarian ammonoid assemblage is dominated bynektobenthic forms, whereas other life-forms are repre-sented by reasonable numbers of taxa. It is difficult tosay what proportion of life-forms should be consideredoptimal. However, it is possible to suggest that thegreater and more even the diversity, and the closer thecommunity is to filling all available niches, the closer itis to the phase of maturity and balance. Apparently in
the Sakmarian, the topography of the strait, its zones, andhydrology were favorable for ammonoid communities.
Artinskian
The Artinskian is the final stage of the existence ofthe South Urals basin. By the end of the Artinskian, thesouthern margin of the Uralian basin was almost com-pletely closed. The Artinskian flyschoids in this regionare everywhere overlain by gypsum-bearing gray Kun-gurian sandstones. The short-term invasions of the seaat the very beginning of the Kungurian reached rela-tively far to the south, approximately to the latitude ofthe city of Aqtöbe. However, the Kungurian Stage isconsidered to be the beginning of another phase of thegeological history of the South Urals.
As the sea became shallower, the hydrology becamemore settled, while the influx of clastics decreased.Artinskian flyschoids contain sandy material from shal-lower fractions, while arenaceous beds in cyclites areless abundant. Carbonate and clayey-silty beds aredominant. The top layers of the stage often containlenses of dolomitized algal limestone, which appar-ently represent the relics of small biostromes thatformed in small hypersaline bays. On the other hand,the topmost beds of the stage often contain trails ofmedium-grained conglomerates and gravelites that sug-gest a series of drying events at the end of the epoch. TheArtinskian beds are extremely facially variable, on thesame bedding plane and across the region of distribution.
The underwater fans formed in the previous epochswere transformed into large accumulative structuresextending deep into the sea and cutting the eastern mar-
10
20
40
50
30
60
70
80
90
100% (a)
Number of species
(b)
5
Ultugan-Sai
1 2 43
10
15
20
section
0 0Mt. Kurmaya
sectionVerkhnii
Ozernyi sectionUltugan-Sai
sectionMt. Kurmaya
sectionVerkhnii
Ozernyi section
Fig. 4.24. Sakmarian ammonoid assemblages in various facial types of sections of the South Urals: (a) proportions of life-formsand (b) species diversity. Explanations as in Fig. 4.3.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1273
gin into a series of bays. These bays were places of dep-osition of carbonate mud with ammonoid shells. Lime-stones overfilled with ammonoid shells form frequentnarrow lenses in sandy-clayey series. It may be sug-gested that semi-liquid mud containing ammonoidshells was sliding down the underwater slopes andfilled in the depressions on the sea floor next to the feetof the fans. The absence of the remains of benthicorganisms in these ammonoid limestones suggests thatinitially ammonoids inhabited the bays with their softmud grounds and avoided unstable coastal environ-ments with high turbulence of water and sediment. Insome cases, another sedimentary model explaining theaccumulation of ammonoid shellstones may be pro-posed, which interprets it as concentration of shells inthe tidal waters of semi-isolated lagoons.
In contrast to the beginning of the Permian, Artin-skian ammonoids are known almost throughout theUrals, from the Mugodzhary Hills in the south to Pai-Khoy and Vaigach Island in the north (Bogoslovskaya,1997). The further north, the more impoverished are theassemblages. While on the extreme south of the region,the communities are extremely rich, richer than allother synchronous associations in the world, in centralBashkortostan the assemblages are noticeably moreimpoverished, not more than two to four genera.
In the South Urals, the Artinskian Stage has a two-fold division (Ruzhencev, 1956b). The existing modelof subdivision of the Artinskian Stage into four hori-zons (Rauzer-Chenousova, 1949) cannot be used in theextreme south of the regions, from where the mainammonoid occurrences are recorded. Attempts to use ascheme based mainly on the sections of the Ufa Plateauto stratigraphic subdivision of the southern section havenot been successful. In addition, in recent decades, thisscheme is updated (the upper horizon is assigned to theKungurian). The controversy of this approach has been
repeatedly discussed (see Chuvashov et al., 2002). In ouranalysis we accept the twofold subdivision of the stage.
The numerous and diverse Artinskian ammonoids ofthe Urals were studied by Karpinsky (1890), Krotov(1885), Tchernov (1907), Maksimova (1935, 1945),Voinova (1934), Ruzhencev (1936, 1956b), and Bo-goslovskaya (1962). Material on Artinskian ammonoidsincludes over 20 thousand shells collected in the sec-tions all over the western Fore-Urals. Of these, the col-lections made by Ruzhencev, coll. PIN, no. 317, repre-sent three-quarters.
The Early Artinskian Aktasty assemblage includes28 species of 17 genera of 12 families, i.e., somewhatless diverse than that of Sakmarian. The total mor-phospace did not change considerably D = 0.1–0.53;W = 1.3–2.8; S = 0.35–3.25. The bimodal distributionremained, while the peaks are separated by an intervalwith D between 0.2 and 0.25. The morphospace with thelower values of D contains narrowly umbilicate morpho-types of the nektobenthic and benthopelagic types.
The group of the higher values of D includes repre-sentatives of all three life-forms.
The nektobenthic life-form (Pronoritidae, Medlicot-tiidae, Daraelitidae, Thalassoceratidae, Agathiceratidae,Popanoceratidae) constitutes 57%. The proportion ofthe types of planktonic life-forms is as follows: plank-ton-1, six species (21%) (Paragastrioceratidae, Eothin-itidae) and plankton-2, three species (11%) (Adrianiti-dae, Marathonitidae). Benthopelagic forms (Metalego-ceratidae, Somoholitidae) constitute 11% (Fig. 4.25a).The proportion of the benthopelagic species noticeablydecreased due to an increase in the proportion of thenektobenthic life-form. The proportion of planktoniclife-forms remained virtually the same as that in theSakmarian, but the distribution of its subtypes wasshifted towards a decrease in the proportion of the invo-lute species.
21%6 species
11%3 species
57%16 species
11%3 species
1 2 3 4
12%3 species
52%13 species
12%species
24%6 species
(a)(b)
Fig. 4.25. Proportions of life-forms in the Artinskian (Aktastynian) ammonoid assemblages: (a) of the South Urals; (b) Aktasty sec-tion. Explanations as in Fig. 4.3.

1274
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
In the Aktasty time, the South Uralian basin becameshallower (Khvorova, 1961). The role of shallowing inthe reduction of the diversity of benthopelagicammonoids is relatively easy to explain. Throughouttheir history the ammonoid communities in the basinshow an increase in the proportion of the nektobenthosas the basin became shallower. Among the sections ofthe Aktasty Substage, the section Aktasty in theAktyubinsk Region of Kazakhstan is one of the mostimportant. The dolomitized limestones of the upperpart of the substage contain an assemblage of 26 spe-cies. In a sample of this assemblage over half the spec-imens are Kargalites typicus, whereas Agathicerasuralicum, which normally dominates other samples, ishere represented by only a few specimens. The nekto-benthic life-form constitutes 52%. The proportion ofthe subtype of the planktonic life-forms is as follows:6 species (24%) evolute and 3 species (12%) involute(Fig. 4.25b). Benthopelagic forms constitute 12%. Thisis different from the total assemblage in the absence ofthree nektobenthic species, whereas all other speciesare present. This is reasonable because almost the entireAktasty assemblage comes from a locality on theAktasty River. In addition, there are a few more locali-ties, with much poorer content.
The Late Artinskian (Baigendzhinian) assemblageshows an increased taxonomic diversity, which was thehighest since the Late Carboniferous (Fig. 4.3).
The assemblage includes 52 species of 19 genera of13 families. The total morphospace is delineated in thefollowing way: D = 0.02–0.6; W = 1.4–2.8; S = 0.3–3.4.Nektobenthic species constitute 39% of the total diver-sity. Plankton-1 constitutes 21%, and plankton-2, 27%.Benthopelagic species constitute 13% (Fig. 4.26a).
In the Baigendzhinian, the community experiencedexplosive speciation, primarily among representativesof the superfamilies Neoicocerataceae (Paragastrio-ceras, Uraloceras, Metalegoceras, and Eothinites).
Almost all new species of this group are, according toour interpretation, planktonic or benthopelagic. Theyare responsible for the very rapid growth of the plank-tonic portion of the assemblages. Within the planktonicgroup, the number of species with a subspherical shellincreased, increasing the proportion of the involute por-tion of this community. Thus, the total proportion ofplankton reached 48%. In this case, the reduction in theproportion of nektobenthic species is not related to adecrease in their absolute numbers (they became 5 spe-cies more) but to a sharp growth of species, whichbelonged to other life-forms.
The section of Mt. Zhil-Tau in the AktyubinskRegion contained a uniquely rich ammonoid assem-blage of Baigendzhinian age.
In the lower part of the substage the basal layers ofthe member of dolomitized limestone contain a lens-shaped bed of cephalopod shellstones. Various workerscollected more than 2000 shells of 41 species from thelenses (Ruzhencev, 1956b). Nektobenthic species con-stitute 40% of the total diversity. Plankton constitutes41%, of which 14% are involute. Benthopelagic speciesconstitute 19% (Fig. 4.26b). Although the total numberof specimens of various species may fluctuate from sev-eral specimens to three thousand specimens, while thedistribution of the shells across the life-forms wasapproximately the same. In addition, no single speciescan be identified as dominant in this sample, as the pro-portion of the most numerous one does not exceed12%. The composition of this assemblage differs fromthat of the assemblages of the entire stage (substage) inthe reduced proportion of involute planktonic forms.Compared to the above-mentioned sections, this assem-blage is most similar in the proportion of the life-formsto that of the Sakmarian assemblage of Mt. Kurmaya.Apparently the rootstocks of this similarity are quitedifferent in different sections. Ammonoids from thesection of Mt. Kurmaya come from several levels, and
27%14 species
13%7 species
39%20 species
21%11 species
1 2 3 4
19%9 species
40%15 species
14%5 species
27%10 species
(a)(b)
Fig. 4.26. Proportions of life-forms in the Artinskian (Baigendzhinian Substage) ammonoid assemblages: (a) of the South Urals;(b) from the Zhil-Tau section. Explanations as in Fig. 4.3.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1275
hence reflect a sequence of communities. All ammonoidsfrom Zhil-Tau were collected from the same thin layer.It seems that this was a result of accumulation of float-ing shells from a large territory, which explains averageand similar proportions of various life-forms. It is sup-ported by the virtually complete absence of benthos.
Thus, the Baigendzhinian at the same time showsthe closure of the Uralian Strait and a peak of diversityof the ammonoid community. This suggests that thisbasin ceased to exist very quickly when the declinebegan. There are not any features of degradation of theammonoid assemblage, whereas the richest collectionscome from the topmost beds of the stage. In addition,Kungurian ammonoids, which are known from the sec-tions of the Middle Urals are taxonomically impover-ished, i.e., the continuity of the assemblages is verysmall. Hence, the drying event in the south part of thebasin had a catastrophic effect on ammonoid communi-ties, which could not adapt to new conditions.
Changes in the ecological structure of Paleozoicammonoids communities of the Uralian Paleobasin areto a large extent connected with general changes in theabiotic environment, including the depth of the basin,sedimentary settings, and spatial distribution of adap-tive zones. Several stages during which the ecologicalstructure remained relatively stable can be recognized.
(1) Early Devonian (Emsian)—ammonoids arerepresented by exclusively planktonic forms, evolute orloosely coiled whorls, which indicate the period of earlyevolution of ammonoids, whereas their occurrences in theopen shelf deposits suggest that this was the adaptive zoneof their origin. The species diversity is low.
(2) Eifelian–Frasnian. The ammonoid communitybecame considerably more diverse ecologically, althoughthe species diversity remains the same. Nektobenthicinvolute forms dominate (over 80%), plankton is repre-sented by the evolute morphotype; the benthopelagiclife-form is represented by only one species. They arefound in various types of deposits, which suggests thatammonoids inhabited various marine environments.
(3) Famennian. In the Famennian, the ammonoidcommunities changed fundamentally: the taxonomicand morphological diversity sharply increased, the pro-portion of planktonic form increased (the second typeof planktonic morphotype represented by involuteshells with slowly expanding whorls appeared) and theproportion of benthopelagic forms became larger. Themaximum diversity of ammonoids is recorded from themost basinal sections with condensed sedimentation.The catastrophic reduction of the biological diversity atthe end of the Famennian primarily affected benthope-lagic and evolute planktonic forms, which can be con-nected with shallowing and reduction in the area of theouter (deep) shelf. Only involute nektobenthic andplanktonic forms continued into the Carboniferous.
(4) Early Carboniferous. In the Early Carbonifer-ous, the Uralian basin can be interpreted as a large epi-
continental marine basin, which is reflected in the eco-logical structure of ammonoid communities. In theTournaisian and Early Visean–beginning of the LateVisean the assemblages are dominated by nektobenthiclife-forms and contained benthopelagic life-forms,while the proportion of planktonic life-forms was min-imal. At the end of the Visean and in the Serpukhovian,a specific ammonoid community was formed, the mainfeature of which was the prevalence of planktonic andbenthopelagic forms, which reflected the extension ofthe adaptive zone of the relatively deep outer shelf.
(5) Early Bashkirian. New ammonoid communi-ties in the Early Bashkirian after a large-scale regres-sion at the end of the Serpukhovian in its structure issimilar to the Serpukhovian, but at the end of the EarlyBashkirian the diversity decreased as the number ofplanktonic and benthopelagic life-forms became lower.Very few or no Late Bashkirian and Moscovianammonoids are found in the Urals.
(6) Kasimovian–Baigendzhinian (Permian). Thestructure of the communities in this long period was asubject of fluctuations, but in general it differed consid-erably from the Early Carboniferous and Early Bash-kirian structures. At this stage the assemblages weredominated by nektobenthic forms with gradual increasein the proportion of planktonic forms and decrease inthe proportion of benthopelagic forms. Interestingly, at theend of this stage, in the Baigendzhinian, the ecologicalstructure of ammonoids does not show traces of sharpchanges indicating its degradation and shows features of amature community with proportional quantities of life-forms. This apparently suggests that the Uralian Basinclosed very quickly, and fast change in the abiotic condi-tions did not lead to changes in the community.
CONCLUSIONS
The main feature of the cephalopod archetype is thedevelopment of the phragmocone, a new functionalbuoyancy device, allowing cephalopods to colonize anew adaptive zone (pelagic zone), which was at thattime outside the reach of other mollusks. The develop-ment of the main body plans within the archetype,which correspond to the higher taxonomic divisions,can functionally be explained by different approachesto buoyancy and orientation control and facilitation ofactive swimming. In the course of discussion of funda-mentally different approaches to these problems,23 orders of Paleozoic cephalopods are considered andsubstantiated. Seven ammonoid orders represented afunctionally and phylogenetically connected group,which can be considered as a single taxon of a higherrank. The combination of other cephalopod orders, mostof which did not survive beyond the Paleozoic in supraor-dinal taxa, cannot be justified from the functional point ofview. They are considered as a group of nonammonoidcephalopods with uncertain taxonomic status.

1276
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
A discussion of differences in the hydrostatic andhydrodynamic features of various shell morphotypesallows the evaluation of their potential colonization ofvarious adaptive zones in the sea. Based on this, in eachof 23 genera, the following life-forms are recognized:benthic, benthopelagic, nektobenthic, and planktonic,and changes in the ecological structure of each of thecephalopod orders are tracked throughout their evolu-tionary history. The predominant ecological specializa-tion of cephalopod orders is defined by the shell shapeinitial for the body plan of each order: cyrtoconic,orthoconic, or spirally coiled. Ellesmerocerids, dis-cosorids, and oncocerids initially with a cyrtoconicshell were the most ecologically flexible and capable ofliving in all adaptive zones. The orders with an initiallyorthoconic shell and well-developed mechanisms oforientation and stability control in the form of cameraland/or endosiphuncular deposits (endocerids, actinoc-erids, orthocerids, and pseudorthocerids) are mainlyrepresented by the nektobenthic life-form only. Bactrit-ids, which did not have specific mechanisms of orienta-tion control are considered as planktonic organisms.Orders with an initially planispiral shell (nautilids,ammonoids) were the most adapted to life in the pelagiczone and gave a broad spectrum of planktonic forms.
The recognition of the life-forms in cephalopodorders and tracking their ecogenesis created a basis forconsideration of the ecological evolution of the entiretaxocoenosis of cephalopods throughout the Paleozoic.The benthopelagic life-form was initial in the evolutionof cephalopods, and it was the life-form of the earliestLate Cambrian genus Plectronoceras. From the begin-ning of the Early Ordovician, the ecological structure ofthe cephalopod community included all known life-forms. A relatively constant proportion of the life-formsremained throughout the entire Ordovician, despite thechanges in the taxonomic composition of the commu-nity in the mid-Ordovician. This could be because thenewly appearing orders relatively evenly filled adaptivezones and niches, and due to a relatively stable structureof the adaptive sea zone throughout the Ordovician.The first significant change in the ecological structureof the cephalopod community occurred in the Silurian,when the number of bottom-dwelling (benthic and ben-thopelagic) life-forms increased, while the number ofplanktonic life-forms decreased. In the Devonian, theecological structure becomes more balanced, althoughthe relative number of pelagic forms is higher than inthe Ordovician. Similar to the Ordovician, the appear-ance of new groups (nautilids and ammonoids) in theDevonian did not change the proportions of life-formsin the community. This change, the largest in the Pale-ozoic history of cephalopods, was related to the globalevents at the end of the Frasnian. The number of benthicand benthopelagic forms halved, but the number ofplanktonic forms increased three times (mainly becauseof the appearance of clymeniids). The reduction in thenumber of benthic forms quite logically follows the dis-
tribution of anoxic conditions, which led to the reduc-tion of bottom habitats.
A catastrophic decrease in the taxonomic diversityof cephalopods at the Devonian–Carboniferous bound-ary was restored relatively quickly due to the radiationof a few nektobenthic ammonoids that survived the cat-astrophic event. From the Carboniferous onward, nobenthic forms appeared in the cephalopod community.The Carboniferous–Middle Permian ammonoid com-munity is characterized by a relatively stable ecologicalstructure, taxonomically represented by the orders ofammonoids and nautilids with a planispiral shell. Thetaxonomic structure changed at the end of the MiddlePermian, and like in the previous periods of instability,was related to an increase in the number of pelagic taxa.While the changes in the ecological structure in the Sil-urian and at the end of the Frasnian were related to theglobal extinction events, in this case, changes in theecological structure actually preceded the Permian–Tri-assic extinction event. Apparently, the ecologicalchange in the cephalopod community resulted from theappearance in the mid-Permian of the order Ceratitida,an initially pelagic group, which survived the crisis andbecame the rootstock of the Mesozoic radiation ofammonoids.
Thus, the analysis of the changes in the ecologicalstructure of the cephalopod community and changes inthe taxonomic composition of life-forms in the Paleo-zoic epochs shows that the appearance of new highertaxa in the cephalopod community has no certain corre-lation with abiotic events and is not caused by theseevents. The appearance of new groups usually did notchange the ecological structure of the cephalopod com-munity.
The major trend in the evolution of the ecologicalstructure of cephalopod community in the Paleozoicwas the reduction in the benthic life-forms and increasein abundance and morphological diversity of pelagiclife-forms, while the number of higher taxa decreased.Periods of “anomalous” ecological structure of thecommunity always resulted from changes in the envi-ronment. This is observed not only when the entirecephalopod community is analyzed from stage to stage,but also in the analysis of changes in ecological struc-ture in ammonoid communities based on the exampleof the Uralian Paleobasin. The ecological structure ofthe ammonoid community in this paleobasin is ana-lyzed for stages and substages from the Early Devonianto Early Permian. Several periods are recognized whenthe structure of the communities was relatively stable:Emsian, Eifelian–Frasnian, Famennian, Early Carbon-iferous (Tournaisian–Visean), Early Bashkirian, andKasimovian–Early Permian. The disruptions in the eco-logical structure partly reflect changes in the globalcephalopod taxocoenosis and partly result from thechanges in the geology and oceanography in the Ura-lian Paleobasin.

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1277
ACKNOWLEDGMENTSThe authors are grateful to Prof. L.A. Nevesskaja
and Dr. E.B. Naimark for valuable comments and sug-gestions, which were essential in preparation and pub-lication of this monograph.
The study is supported by the Program of the Presid-ium of the Russian Academy of Sciences “Origin andEvolution of the Biosphere” (Subprogram II, Part 5.Problem of Coevolution of Abiotic and Biotic Events).
REFERENCES1. W. M. Ahr, “The Carbonate Ramp; an Alternative to the
Shelf Model,” Today’s New Technology, Tomorrow’sNew Targets, Gulf Coast Assoc. Geol. Soc. Transact.23, 221–225 (1973).
2. W. M. Ahr, “Carbonate Ramps, 197–1996: A HistoricalReview,” Geol. Soc. London, Special Publ. 149, 7–14(1998).
3. L. Z. Akhmetshina, N. B. Gibshman, and S. V. Ni-kolaeva, “The Famennian Oil- and Gas-Bearing Forma-tion of Western Kazakhstan (The Caspian Sea Basin),”in Reports of Kazakhstan Geologists (Kazakh. Geol.Ob-vo “KazGeo,” Almaty, 2004), pp. 89–97 [in Rus-sian].
4. Yu. G. Aleev, Ecomorphology (Kiev, Naukova Dumka,1986) [in Russian].
5. Yu. G. Aleev and V. D. Burdak, “Ecological and Mor-phological Convergence and a Unified Ecomorphologi-cal System of Organisms,” Ekol. Morya, Issue 4, 3–17(1984).
6. A. S. Alekseev, L. I. Kononova, and A. M. Nikishin,“The Devonian and Carboniferous of the MoscowSineclise (Russian Platform): Stratigraphy and Sea-Level Changes,” Tectonophysics 268 (2), 149–168(1996).
7. A. I. Antoshkina, Reef Formation in the Paleozoic (TheNorthern Urals and Adjacent Areas) (Ural. Otd. Ross.Akad. Nauk, Yekaterinburg, 2003) [in Russian].
8. Z. G. Balashov, “Order Tarphycerida,” in Fundamentalsof Paleontology: Mollusks: Cephalopods 1 (Akad.Nauk SSSR, Moscow, 1962), pp. 77–82 [in Russian].
9. E. A. Balashova, “Goniatites from the Carboniferous ofBerchogur,” Ezhegodn. Vses. Paleontol. O–va, Issue 14,189–202 (1953).
10. I. S. Barskov, Late Ordovician and Silurian Cephalo-pod Mollusks of Kazakhstan and Central Asia (Mos-cow, 1972) [in Russian].
11. I. S. Barskov, “Geometry of Coiled Shells of Cephalo-pods and Its Biological Significance,” in Main Avenuesin the Application of Mathematical Methods in Paleon-tology (Kishinev, 1976), pp. 16–19 [in Russian].
12. I. S. Barskov, “Analysis of Functional Morphology ofSpirally Coiled Shells of Cephalopod Mollusks,” inModern Paleontology (Nedra, Moscow, 1988), Vol. 1,pp. 139–159 [in Russian].
13. I. S. Barskov, Morphogenesis and Ecogenesis of Paleo-zoic Cephalopods (Mosk. Gos. Univ., Moscow, 1989)[in Russian].
14. I. S. Barskov, “Why Ammonoids Have Complex Pat-terns of Septa and Sutures?,” in Fossil Cephalopods:
The Latest Advances in Their Study (Paleontol. Inst.Ross. Akad. Nauk, Moscow, 1999), pp. 53–61 [in Rus-sian].
15. I. S. Barskov and G. N. Kiselev, “Dynamics of theDiversity of Silurian Cephalopods in Various Bio-chores,” in Ecosystem Rearrangement and Evolution ofthe Biosphere (Paleontol. Ins. Ross. Akad. Nauk, 1995),Issue 2, pp. 41–44 [in Russian].
16. I. S. Barskov, K. V. Simakov, and A. S. Alekseev,“Devonian–Carboniferous Transitional Deposits of theBerchogur Section, Mugodzhary, USSR (PreliminaryReport),” Cour. Forsch. Inst. Senckenberg 67, 207–230(1984).
17. U. Bayer and G. R. McGhee Jr., “Iterative Evolution ofMiddle Jurassic Ammonite Faunas,” Lethaia 17, 1–16(1984).
18. R. T. Becker, “Anoxia, Eustatic Changes, and Upper Devo-nian to Lowermost Carboniferous Global AmmonoidDiversity,” in The Ammonoidea: Environment, Ecology,and Evolutionary Change: Syst. Assoc.; Spec. Pap.,Vol. 47, Ed. by M. R. House (Clarendon Press, Oxford,1993), pp. 115–163.
19. M. F. Bogoslovskaya, Artinskian Ammonoids of TheMiddle Urals: Proceedings of the Paleontological Insti-tute of the Academy of Sciences of the USSR, Vol. 87(Akad. Nauk SSSR, Moscow, 1962) [in Russian].
20. M. F. Bogoslovskaya, “The Genus Goniatites and ItsRepresentatives in the Southern Urals,” Paleontol. Zh.,No. 1, 38–46 (1966).
21. M. F. Bogoslovskaya, “Ammonoids,” in Atlas of Char-acteristic Assemblages of Permian Fauna and Flora ofthe Urals and Russian Platform (Leningrad, Nedra,1986), pp. 37–59 [in Russian].
22. M. F. Bogoslovskaya, “Permian Ammonoids from thePai-Khoy Range and Vaigach Island,” Paleontol. Zh.,No. 6, 23–28 (1997) [Paleontol. J. 31 (6), 588–594(1997)].
23. M. F. Bogoslovskaya and A. V. Popov, “Ammonoids atthe Carboniferous–Permian Boundary,” in Carbonifer-ous–Permian Boundary Beds in the Urals, Cis-Urals,and Central Asia: Biostratigraphy and Correlation(Nauka, Moscow, 1986a), pp. 56–63 [in Russian].
24. M. F. Bogoslovskaya and A. V. Popov, “New Species ofAmmonoids from the Carboniferous–Permian Bound-ary Beds of the Southern Urals,” in Carboniferous–Per-mian Boundary Beds in the Urals, Cis-Urals, and Cen-tral Asia: Biostratigraphy and Correlation (Nauka,Moscow, 1986b), pp. 125–129 [in Russian].
25. M. F. Bogoslovskaya, I. A. Mikhailova, and A. A. She-vyrev, “The System of Ammonoids,” in Systematics andPhylogeny of Invertebrates: Criteria for DistinguishingHigher Taxa (Nauka, Moscow, 1990), pp. 69–98 [inRussian].
26. M. F. Bogoslovskaya, L. F. Kusina, and T. B. Leonova,“Classification and Distribution of Late PaleozoicAmmonoids,” in Fossil Cephalopods: The LatestAdvances in Their Study (Palentol. Inst. Ross. Akad.Nauk, Moscow, 1999), pp. 89–124 [in Russian].
27. B. I. Bogoslovsky, “On the Family Biloclymeniidaefam. nov.,” Dokl. Akad. Nauk SSSR 104 (1), 134–137(1955).

1278
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
28. B. I. Bogoslovsky, “A Contribution to the Classificationof Agoniatitida,” Dokl. Akad. Nauk SSSR 116 (3), 489–492 (1957).
29. B. I. Bogoslovsky, “On the Origin of the Order Goni-atitida,” Dokl. Akad. Nauk SSSR 123 (5), 921–924(1958a).
30. B. I. Bogoslovsky, “A Contribution to the Evolution ofthe Family Gephuroceratidae Frech, 1901,” Izv. Akad.Nauk SSSR 64, 1–156 (1958b).
31. B. I. Bogoslovsky, “New Representatives of Ammonoidsfrom the Middle Devonian Deposits of the SubpolarUrals,” Paleontol. Zh., No. 3, 61–65 (1959).
32. B. I. Bogoslovsky, “New Records of Some Little-Known Representatives of Devonian Ammonoids,”Byull. Mosk. O–va Ispyt. Prir., Otd. Geol. 35 (4), 155–156 (1960a).
33. B. I. Bogoslovsky, “New Representatives of Some Lit-tle-Known Genera of Devonian Ammonoids from theFamennian Deposits of the Urals,” Paleontol. Zh.,No. 4, 69–73 (1960b).
34. B. I. Bogoslovsky, “Eifelian Ammonoids of the UralMountains and Problems of the Classification of Agoni-atitida,” Paleontol. Zh., No. 4, 60–70 (1961).
35. B. I. Bogoslovsky, Fundamentals of Paleontology:Mollusks: Cephalopods: 1. Devonian Ammonoids(Moscow, Akad. Nauk SSSR, 1962a), pp. 60–70 [inRussian].
36. B. I. Bogoslovsky, “An Uncommon Type of Ornamen-tation in Clymenids,” Paleontol. Zh., No. 1, 166–168(1962b).
37. B. I. Bogoslovsky, “The Earliest Devonian Ammonoidsof the Ural Mountains,” Paleontol. Zh., No. 2, 26–37(1963).
38. B. I. Bogoslovsky, “Carinoclymenia—a New Genus ofthe family Rectoclymeniidae,” Paleontol. Zh., No. 4,88–91 (1965).
39. B. I. Bogoslovsky, Devonian Ammonoids: 1. Agoni-atitida: Proceedings of the Paleontological Institute ofthe Academy of Sciences of the USSR, Vol. 124 (Akad.Nauk SSSR, Moscow, 1969) [in Russian].
40. B. I. Bogoslovsky, Devonian Ammonoids: 2. Goni-atitida: Proceedings of the Paleontological Institute ofthe Academy of Sciences of the USSR, Vol. 127 (Akad.Nauk SSSR, Moscow, 1971) [in Russian].
41. B. I. Bogoslovsky, “New Early Devonian Cephalopodsof Novaya Zemlya,” Paleontol. Zh., No. 4, 44–51(1972).
42. B. I. Bogoslovsky, “A New Family of Clymenids,” Pale-ontol. Zh., No. 3, 35–41 (1975).
43. B. I. Bogoslovsky, “Early Ontogeny and Origin of Cly-menids,” Paleontol. Zh., No. 2, 41–50 (1976).
44. B. I. Bogoslovsky, “On the Family MiroclymeniidaeSchindewolf, 1924,” Paleontol. Zh., No. 4, 47–58(1977).
45. B. I. Bogoslovsky, “Cteroclymenia—a New Genus ofthe Family Rectoclymeniidae,” Paleontol. Zh., No. 1,140–143 (1979a).
46. B. I. Bogoslovsky, “Systematics and Phylogeny of Cly-meniina,” Paleontol. Zh., No. 2, 32–47 (1979b).
47. B. I. Bogoslovsky, Devonian Ammonoids: 3. Clymenids(Suborder Gonioclymeniina): Proceedings of the Pale-
ontological Institute of the Russian Academy of Sci-ences, Vol. 191 (Akad. Nauk SSSR, Moscow, 1981) [inRussian].
48. B. I. Bogoslovsky, “An Interesting Form of AperturalStructures on the Shell of Clymenids,” Dokl. Akad.Nauk SSSR 264 (6), 1483–1486 (1982).
49. B. I. Bogoslovsky, “A New Genus of the Family Carin-oclymeniidae,” Paleontol. Zh., No. 3, 196–109 (1983).
50. B. I. Bogoslovsky and L. F. Kusina, “Late DevonianAmmonoids of the Kozhim River Basin in the SubpolarUrals,” Paleontol. Zh., No. 2, 67–73 (1980).
51. B. I. Bogoslovsky, I. A. Poslavskaya, and O. E. Belyaev,“Records of Frasnian Ammonoids in Central Kazakh-stan,” Paleontol. Zh., No. 3, 35–40 (1982).
52. M. S. Boiko, “Late Paleozoic Ammonoids with a Non-ventral Siphuncule,” Paleontol. J. 39 (Suppl. 5), 607–614 (2005).
53. M. S. Boiko, “Morphometric Methods in the Study ofAmmonoids,” in Modern Problems in the Study ofCephalopod Mollusks: Morphology, Systematics, Evo-lution, and Biostratigraphy: Proceedings of All-RussiaConference (Paleontol. Inst. Ross. Akad. Nauk, Mos-cow, 2006), pp. 85–88 [in Russian].
54. K. V. Borisenkov, “Evolution of Uralian Somoholitidsat the Carboniferous–Permian Boundary,” Vestn.S.-Peterb. Univ., Ser. 7, Issue 1, No. 7, 78–83 (2002).
55. K. V. Borisenkov, “Zonal Subdivisions of the GzhelianStage in the Southern Urals and Their InterregionalCorrelation,” Regional’naya Geol. i Metallogeniya,No. 18, 73–82 (2003).
56. K. V. Borisenkov, “On Some Permian Somoholitids(Ammonoidea) from the Ural Mountains,” Paleontol.Zh., No. 1, 19–25 (2004a) [Paleontol. J. 38 (1), 20–26(2004a)].
57. K. V. Borisenkov, “New Dates for the Upper Carbonif-erous (Pennsylvanian) Genus Neodimorphoceras(Ammonoidea),” in VI Intern. Symp. Cephalopods—Present and Past, Fayetteville, 2004 (2004b), pp. 27–28.
58. J. A. Chamberlain, Jr., “Flow Patterns and Drag Coeffi-cients of Cephalopod Shells,” Palaeontology 19, 593–563 (1976).
59. J. A. Chamberlain, Jr., “Motor Performance and Jet Pro-pulsion in Nautilus: Implications for Cephalopod Pale-obiology and Evolution,” Bulletin of the American Mal-acol. Union. 37–42 (1980).
60. J. A. Chamberlain, Jr., “Hydromechanical Design ofFossil Cephalopod,” in The Ammonoidea: Syst. Assoc.Spec. Vol. 18 (Academic Press, London, 1981),pp. 289–336.
61. N. M. Chumakov and M. A. Zharkov, “Climate duringPermian–Triassic Biosphere Reorganizations, Article 1:Climate of the Early Permian,” Stratigr. Geol. Korre-lyatsiya 10 (6), 62–81 (2002) [Stratigr. Geol. Correla-tion 10 (6), 586–602 (2002)].
62. B. I. Chuvashov, R. M. Ivanova, and A. N. Kolchina,Upper Paleozoic of the Eastern Slope of the Urals(Stratigraphy and Geological History) (Ural’sk.Nauchn. Tsentr Akad. Nauk SSSR, Sverdlovsk, 1984)[in Russian].
63. B. I. Chuvashov, V. V. Chernykh, and M. F. Bogos-lovskaya, “Biostratigraphic Characteristic of StageStratotypes of the Permian System,” Stratigr. Geol. Kor-

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1279
relyatsiya 10 (4), 3–19 (2002) [Stratigr. Geol. Correla-tion 10 (4), 317–333 (2002)].
64. R. E. Crick, “The Practicality of Vertical Cephalopod-Shells As Paleobathymetric Markers,” Bul. Geol. Soc.of America. 94, 1109–1116 (1983).
65. E. J. Denton and J. B. Gilpin-Brown, “The Buoyancy ofthe Cuttlefish Sepia officinalis (L.),” J. Mar. Biol. Assoc.U. K. 41, 319–342 (1961).
66. E. J. Denton and J. B. Gilpin-Brown, “Further Observa-tions on the Buoyancy of Spirula,” J. Mar. Biol. Assoc.U. K. 51, 363–373 (1971).
67. E. J. Denton, J. B. Gilpin-Brown, and J. V. Howart, “TheOsmotic Mechanisms of the Cuttlebone,” J. Mar. Biol.Assoc. U. K. 41, 351–364 (1961).
68. L. Dollo, “Les Céphalopodes adapté à la vie nectoniquesecondaire et à le vie benthonique tertiaire,” Zool. Jb.Suppl. 1, 105–140 (1912).
69. L. Dollo, “Les Céphalopodes déroulés et l’irréversibil-ité de l’évolution,” in Koninklijk Zoologisch Genootsc-hap Natura Artis Magistra te Amsterdam (N. V. Boekhan-del en Drukkerij voorheen E.J. Brill, Leiden, 1919–1922), pp. 215–226.
70. D. T. Donovan, “Cephalopod Phylogeny and Classifica-tion,” Biol. Rev. 39 (3), 259–287 (1964).
71. V. V. Drushchits and V. N. Shimansky, “The Main Prob-lems in the Systematics of Ectocochleate Cephalo-pods,” Byull. Mosk. O–va Ispyt. Prir., Otd. Geol. 57 (6),94–107 (1982).
72. V. V. Drushchits, M. F. Bogoslovskaya, and L. A. Dogu-zhaeva, “Evolution of Septal Tubes in Ammonoids,” Pale-ontol. Zh., No. 1, 41–56 (1976).
73. J. Dzik, “Origin of the Cephalopoda,” Acta Paleonto-logica Polonica. 26 (2), 161–191 (1981).
74. R. H. Flower, “Studies of the Actinoceratida,” Mem.Inst. Mine Technol. New Mex. 9, 1–68 (1957).
75. R. H. Flower, “The Nautiloid Order Ellesmeroceratida,”Mem. Inst. Mine and Technol. New Mex. 12, 1–234(1964).
76. R. H. Flower and B. Kümmel, “A Classification of theNautiloidea,” J. Paleontol. 24 (5), 604–616 (1950).
77. K. Friederichs, Die Grundfragen Gesetzmässigkeitender land- und forstwirtschaftlichen Zoologie, insbeson-dere der Entomologie (Parey, Berlin, 1930), Vols. 1 and 2.
78. Fundamentals of Paleontology: Mollusks: Cephalo-pods 1, Ed. by V. E. Ruzhencev (Akad. Nauk SSSR,Moscow, 1962) [in Russian].
79. W. M. Furnish and B. F. Glenister, “Tarphycerida,” inTreatise on Invertebrate Paleontology, Part K, Ed. byR. C. Moore (Univ. Kansas Press, Lawrence, 1964).pp. 343–368.
80. H. Gams, “Principienfragen der Vegetationforschung.Ein Beitrag zur Begriffsklarung und Methodik der Bio-cenologie,” Vierteljahrsschr. Naturf. Ges. Zurich. 63,293–493 (1918).
81. N. P. Gerasimov, “The Uralian Series of the PermianSystem: Geology, Books 3 and 4,” Uch. Zap. Kaz. Gos.Univ. 97 (8–9), 1–68 (1937).
82. Gould J. S., Wonderful Life, Ed. by W. W. Norton(New York, 1989).
83. C. Gregoire, “Experimental Alternation of the NautilusShell by Factors Involved in Diagenesis and Metamor-
phism: Part. 1. Thermal Changes in Conchiolin Matrixof Mother-of-Pearl,” Bull. Inst. R. Sci. Nat. Belg. 44(25), 1–69 (1968).
84. R. A. Hewitt, “On the Connecting Rings of the Pseudor-thocerataceae (Carboniferous Cephalopoda),” J. Mol-luscan Studies 48, 374–375 (1982).
85. M. R. House, “On the Origin, Classification, and Evo-lution of the Early Ammonoidea,” in The Ammonoidea(Acad. Press, New York, 1981), pp. 3–36.
86. A. Hyatt, “Cephalopoda in Zittel,” in Text-Book ofPalaeontology, 1st ed. (1900), Vol. 1, pp. 502–592.
87. R. M. Ivanova, “Zonal Stratigraphy and the Boundariesof the Middle Carboniferous in the Urals Defined Basedon Fusulinids,” in Zonal Subdivisions of the Carbonif-erous of the Unified Stratigraphic Scale of Russia: Pro-ceedings of All-Russia Conference, May 29–31, 2000(Gilem, Ufa, 2000), pp. 44–46 [in Russian].
88. R. M. Ivanova, “Fusulinid Zones of the MoscovianStage of the Ural Mountains,” in Stratigraphy andPaleogeography of the Carboniferous of Eurasia: Col-lection of Scientific Papers (Inst. Geol. Geokhim. Ural.Otd. Ross. Akad. Nauk, Yekaterinburg, 2002), pp. 127–138 [in Russian].
89. R. M. Ivanova and B. I. Chuvashov, “Biogeographicand Facies Features of the Zonal Assemblages ofFusulinids of the Middle Carboniferous of the UralMountains and Adjacent Areas,” in Boundaries of Bios-tratigraphic Units of the Carboniferous of the UralMountains (Ural. Otd. Ross. Akad. Nauk, Sverdlovsk,1990), pp. 130–142 [in Russian].
90. D. K. Jacobs, “Shape, Drag, and Power Consumption inAmmonoid Swimming,” Paleobiology 18 (2), 203–220(1992).
91. A. P. Karpinsky, “Fossil Coal Prospecting in the Ufa andOrenburg Provinces in 1868,” Gornyi Zh. 4 (11), 213–219 (1869).
92. A. P. Karpinsky, “Geological Investigations in the Oren-burg Region,” Zapiski Mineral. O–va, Ser. 2, Part 9,212–310 (1874).
93. A. P. Karpinsky, On Ammonoids of the Artinskian Stageand on Some Carboniferous Forms Resembling Them(St. Petersburg, 1890), pp. 1–192 [in Russian].
94. A. P. Karpinsky, “On the Remains of ClymenidsRecorded in the Ural Mountains,” Izv. Zap. Leningr.Univ., Ser. Geol.-Pochv. Nauk, Issue 11, 137–166(1984).
95. D. N. Kashkarov, Environment and Community(Medgiz, Moscow, 1933) [in Russian].
96. I. V. Khvorova, “Flysch and Lower Molasse Formationof the Southern Urals,” Tr. Geol. Inst. Akad. NaukSSSR, Issue 37, 1–352 (1961).
97. N. V. Kind, “Goniatites and Clymenids of the WesternSlope of the Mugodzhar Hills,” Uchen. Zap. Leningr.Univ., Issue 11, 137–166 (1944).
98. D. N. Kiselev, “A Quantitative Description of the Cross-Sectional Shape of the Shell of Ammonites Based onthe Morphofunctional Approach,” in Modern Problemsin the Study of Cephalopod Mollusks: Morphology, Sys-tematics, Evolution, and Biostratigraphy: Proceedingsof All-Russia Conference (Paleontol. Inst. Ross. Akad.Nauk, Moscow, 2006), pp. 92–96 [in Russian].

1280
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
99. G. N. Kiselev, “On the Paleoecology of Silurian Ceph-alopods,” Geosc. Mag. “Chigakukenkyi” 41 (3), 145–148 (1992).
100. I. Klebaba, “Gradual Opening of the Siphonal Tube inOrthoconic Cephalopod from Silurian of Central Bohe-mia (Czech Republic),” J. Czech Geol. Soc. 44 (1–2),131–136 (1999a).
101. I. Klebaba, “Sipho-Cameral Structures in Some Sil-urian Cephalopods from Barrandian Area (Bohemia),”Acta Musei Nationalis Pragae, Ser. B, Historia Natu-ralis 55 (1–2), 1–15 (1999b).
102. T. Kobayashi, “The Ancestory of the Cephalopoda.From Helcionella to Plectronoceras,” Proceeding JapanAcademy, Ser. B 63 (5), 135–138 (1987).
103. N. M. Kochetkova, E. A. Reitlinger, and V. N. Pazukhin,“The Devonian–Carboniferous Boundary in the South-ern Urals,” in Devonian–Carboniferous Boundary onthe Territory of the USSR: Proceedings of All-UnionConference, Minsk, April 1986 (BelNIGRI, Minsk,1986), pp. 173–176 [in Russian].
104. S. E. Kolotukhina, “On the Clymenid Facies of theUpper Devonian of Central Kazakhstan,” Izv. Akad.Nauk SSSR, Ser. Geol., No. 5/6, 671–686 (1938).
105. V. A. Konovalova, “The Distributional Pattern of ViseanAmmonoids in the Subpolar Ural Mountains,” in Eco-system Turnovers and Evolution of the Biosphere,Issue 6 (Paleontol. Inst. Ross. Akad. Nauk, Moscow,2004), pp. 119–124 [in Russian].
106. V. A. Konovalova, “Morphological and Ecological Evo-lution of Ammonoids in the Ural Paleobasin at theVisean–Serpukhovian Boundary,” in Modern Problemsin the Study of Cephalopod Mollusks: Morphology, Sys-tematics, Evolution, and Biostratigraphy: Proceedingsof All-Russia Conference (Paleontol. Inst. Ross. Akad.Nauk, Moscow, 2006), pp. 54–57 [in Russian].
107. V. A. Konovalova and D. B. Sobolev, “ViseanAmmonoids from the Deposits of the Bol’shenadotin-skii Bioherm Structure (Subpolar Urals),” in Proceed-ings of International Conference on Geology of Reefs(Inst. Geol. Komi Nauchn. Tsentr Ural. Otd. Ross.Akad. Nauk, Syktyvkar, 2005), pp. 81–84 [in Russian].
108. D. Korn, “Morphospace Occupation of Ammonoids atthe Devonian–Carboniferous Boundary,” Paläontol. Z.74 (3), 247–257 (2000).
109. D. Korn and C. Klug, “Occluded Umbilicus in thePinacitinae (Devonian) and Its Palaeoecological Impli-cations,” Palaeontology 45 (4), 917–931 (2002).
110. D. Korn and C. Klug, “Morphological Pathways in theEvolution of Early and Middle Devonian Ammonoids,”Paleobiology 29 (3), 329–348 (2003).
111. D. Korn, V. Ebbighausen, J. Bockwinkel, and C. Klug,“The A-Mode Sutural Ontogeny in ProlecanitidAmmonoids,” Palaeontology 46 (Part 6), 1123–1132(2003).
112. B. Kröger, “Adaptive Evolution in Paleozoic CoiledCephalopods,” Paleobiology 31 (2), 253–268 (2005).
113. T. W. Krammer and W. I. Ausich, The “Age ofCrinoids”: A Mississippian Biodiversity Spike Coinci-dent with Widespread Carbonate Ramps, Palaios 21,238–248 (2006).
114. D. A. Krivolutskii, “Modern Concepts of the LivingForms of Animals,” Ekologiya, No. 3, 19–25 (1971).
115. P. I. Krotov, “Artinskian Stage: A Monograph on theGeology and Paleontology of the Artinskian Sand-stone,” Tr. O–va Estestvoispyt. pri Kazansk. Univ. 13(5), 1–314 (1885).
116. P. I. Krotov, “Geological Investigations on the WesternSlope of the Solikamsk and Cherdym Regions of theUral Mountains,” Tr. Geol. Kom. 6 (1), 1–563 (1888).
117. W. Kuhnelt, Grundriss der Ökologie (VEB Fisher, Jena,1970).
118. E. I. Kulagina, V. N. Pazukhin, S. V. Nikolaeva, andN. N. Kochetova, “Biozonation of the Syuran Horizonof the Bashkirian Stage in the South Urals As Indicatedby Ammonoids, Conodonts, Foraminifers, and Ostra-codes,” Stratigr. Geol. Korrelyatsiya 8 (2), 38–56(2000) [Stratigr. Geol. Correlation 8 (2), 137–156(2000)].
119. E. I. Kulagina, V. N. Pazukhin, N. M. Kochetkova,Z. A. Sinitsina, and N. N. Kochetova, Stratotype andReference Sections of the Bashkirian Stage of the Car-boniferous of the Southern Ural Mountains (Gilem,Ufa, 2001) [in Russian].
120. B. Kümmel, “Nautiloidea—Nautilida,” in Treatise onInvertebrate Paleontology, Part K (Univ. Kansas Press,Lawrence, 1964), pp. 383–457.
121. L. F. Kusina, “New and Little-Known Lower Viséan(Saur) Ammonoids,” Paleontol. Zh., No. 1, 37–48(1971).
122. L. F. Kusina, “Contribution to the Revision of the GenusMuensteroceras,” Paleontol. Zh., No. 3, 14–25 (1973).
123. L. F. Kusina, “The Saur Assemblage of Early Carbonif-erous Ammonoids,” Paleontol. Zh., No. 4, 18–31(1974).
124. L. F. Kusina, “Saur Ammonoids,” Tr. Paleontol. Inst.Akad. Nauk SSSR 181, 1–108 (1980).
125. L. F. Kusina, “On Some New Early CarboniferousAmmonoids of the Ural Mountains,” Paleontol. Zh.,No. 4, 91–95 (1983).
126. L. F. Kusina, “Contribution to the Revision of the GenusImitoceras (Ammonoidea),” Paleontol. Zh., No. 3, 35–48(1985).
127. L. F. Kusina, “Biostratigraphic Distribution of theLower Carboniferous Ammonoids in Northern Russia,”in Advancing Research on Living and Fossil Cephalo-pods (New York–Kansas, 1999), pp. 505–519.
128. L. F. Kusina, “Ammonoids from the Tournaisian–Viséan Boundary Beds of Pai-Khoy and the SouthUrals,” Paleontol. Zh., No. 5, 16–24 (2000) [Paleontol. J.34 (5), 486–494 (2000)].
129. L. F. Kusina and S. V. Yatskov, “Representatives of theSuperfamily Neoglyphiocerataceae in the Dombar(Lower Carboniferous) Deposits of Novaya Zemlya,” inFossil Cephalopods: Evolutionary Paths and Systemat-ics of Selected Groups: Proceedings of the Paleontolog-ical Institute of the Academy of Sciences of the USSR,Vol. 243 (Nauka, Moscow, 1990), pp. 51–65 [in Rus-sian].
130. L. F. Kusina and S. V. Yatskov, “Lower- and MiddleCarboniferous Ammonoids of Novaya Zemlya,”Tr. Paleontol. Inst. Ross. Akad. Nauk 275, 1–144 (1999).
131. L. F. Kusina and V. A. Konovalova, “New Ammonoidsfrom the Tournaisian–Visean Boundary Beds on the

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1281
Kozhim River (Subpolar Ural Mountains),” Paleontol. Zh.,No. 1, 10–18 (2004) [Paleontol. J. 38 (1), 10–19 (2004)].
132. L. F. Kusina and D. B. Sobolev, “The Middle Tournai-sian Fauna with Goniocyclus,” in Syktyvkar Collectionof Paleontological Papers No. 6: Proceedings of theInstitute of Geology of the Komi Scientific Center of theUral Division of the Russian Academy of Sciences,Issue 117 (Inst. Geol. Komi Nauchn. Tsentr Ural. Otd.Ross. Akad. Nauk, Syktyvkar, 2005), pp. 79–85 [inRussian].
133. I. E. Leonova and A. V. Voronov, “Ontogenetic Devel-opment of the Suture Line in the Early PermianAmmonoid Genus Synartinskia,” Paleontol. Zh., No. 1,113–116 (1989).
134. T. B. Leonova, “Permian Ammonoids: Classificationand Phylogeny,” Paleontol. J. 36 (Suppl. l), S1–S114(2002).
135. E. Ya. Leven, M. S. Boiko, A. N. Reimers, T. B. Leo-nova, and M. F. Bogoslovskaya, “Lower Permian in theVicinity of the Verkhneozernoe Village (SouthernUrals),” Stratigr. Geol. Korrelyatsiya 10 (5), 44–58(2002) [Stratigr. Geol. Correlation 10 (5), 460–475(2002)].
136. L. S. Librovitch, “Carboniferous Ammonoids from theSouthern Island of Novaya Zemlya,” Tr. Arkt. Inst.Akad. Nauk SSSR 101, 47–107 (1938).
137. L. S. Librovitch, “Carboniferous Deposits in the Shar-tymka River Region and in the Upper Reaches of theUral River,” Tr. Tsentral. Nauchno-Issled. Geologorazv.Inst., Issue 114, 1–43 (1939a).
138. L. S. Librovitch, “Class Cephalopoda: Cephalopods,” inAtlas of Index Forms of Fossil Faunas of the USSR,Vol. 5 (Middle and Upper Series of the CarboniferousSystem) (Gosgeolizdat, Moscow-Leningrad, 1939b),pp. 130–141 [in Russian].
139. L. S. Librovitch, “Class Cephalopoda: Cephalopods,”in Atlas of Index Forms of Fossil Faunas of the USSR,Vol. 4 (Lower Series of the Carboniferous System) (Gos-geolizdat, Moscow–Leningrad, 1941), pp. 132–153 [inRussian].
140. L. S. Librovitch, “On Some New Groups of Goniatitesfrom the Carboniferous Deposits of the USSR,” Ezhe-godn. Vses. Paleontol. O–va 16, 246–272 (1957).
141. G. P. Lyashenko, “Goniatites at the Base of the FrasnianStage of Timan,” Izv. Akad. Nauk SSSR, Ser. Geol.,No. 5, 87–92 (1956).
142. G. P. Lyashenko, “New Species of Devonian Gonia-tites,” Tr. Vseros. Nauch. Issled. Geologorazv. Neft.Inst., Issue 8, 192–211 (1957).
143. S. V. Maksimova, “On Some New Species of ArtinskianAmmonites,” Byull. Mosk. O–va Ispyt. Prir., Otd. Geol.13 (2), 273–288 (1935).
144. S. V. Maksimova, On Some Representatives of the Fam-ily Pronoritidae: Proceedings of the PaleontologicalInstitute of the Academy of Sciences of the USSR, Vol. 9,Issue 1 (Akad. Nauk SSSR, Moscow, 1938), pp. 1–44[in Russian].
145. S. V. Maksimova, “Contribution to the Faunistic Char-acterization of the Artinskian Stage s. str.,” Dokl. Akad.Nauk SSSR 46 (2), 74–76 (1945).
146. S. V. Maksimova, Ammonites from the Lower Part ofSchwagerina Beds of the Yurezan’ River: Proceedings
of the Paleontological Institute of the Academy of Sci-ences of the USSR, Vol. 14, Issue 4 (Akad. Nauk SSSR,Moscow, 1948), pp. 1–42 [in Russian].
147. S. V. Maksimova, Ecological and Facies Features andthe Formation Conditions of the Domanik Formation(Moscow, Nauka, 1970) [in Russian].
148. N. Malyshev, “Tectonic Evolution of the PechoraBasin,” in Pan-Arctic Palaeozoic Tectonics: Evolutionof Basins and Faunas (Ichtyolith Issues Spec. Publ. 6)(Geoprint, Syktyvkar, 2000), pp. 56–58.
149. G. A. Mizens, Upper Paleozoic Flysch of the WesternUral Mountains (Ural. Otd. Ross. Akad. Nauk, Yekater-inburg, 1997) [in Russian].
150. G. A. Mizens, “Sedimentary Basins in the PaleozoicHistory of the Southern Urals,” in Sedimentary Basinsof the Ural Mountains and Adjacent Areas: Regularitiesin Structure and Mineral Formation (Inst. Geol.Geokhim. Ural. Otd. Ross. Akad. Nauk, Yekaterinburg,2000), pp. 53–67 [in Russian].
151. G. A. Mizens, “On the Sedimentary Basins of the Earlyand Middle Carboniferous in the Southern Urals,” inStratigraphy and Paleogeography of the Carboniferousof Eurasia: Collection of Scientific Papers (Inst. Geol.Geokhim. Ural. Otd. Ross. Akad. Nauk, Yekaterinburg,2002), pp. 213–219 [in Russian].
152. H. Mutvei, “Ultrastructural Studies on CephalopodaShells: Part 2. Orthoconic Cephalopods from the Penn-sylvanian Buckhorn Asphalt,” Bull. Geol. Inst. Univ.Uppsala, New Ser. 3 (9), 263–272 (1972).
153. H. Mutvei, “The Nacreous Layer in Molluscan Shell,”in The Mechanisms of Biomineralization in Animalsand Plants, Ed. by M. Omori and N. Watabe (TokaiUniversity Press, 1980), p. 8.
154. H. Mutvei, “Characterization of Actinoceratoid Cepha-lopods by Their Siphuncular Structure,” Lethaia 29 (4),339–348 (1997).
155. D. V. Nalivkin, “Report on the Fieldwork between theZilim and Zigan Rivers in 1925,” Izv. Geol. Kom. 45 (4)(1926).
156. D. V. Nalivkin, “The Sterlitamak Intersection of theSouthern Urals,” in XVII International Geological Con-gress: Permian Excursion: Southern Itinerary (1937),pp. 93–116 [in Russian].
157. D. V. Nalivkin, “The Tournaisian Stage of the Sterlita-mak Region,” Materialy Vses. Geol. Inst., Paleontol.Stratigr., Collection No. 4, 11–23 (1945).
158. A. K. Nalivkina, “Upper Devonian Goniatites and Cly-menids of the Mugodzhar Hills,” Tr. Vses. Neft.Nauchno-Issled. Geologorazv. Inst., N. S., Issue 72, 60–125 (1953).
159. K. N. Nesis, “Ecological Classification (Life-Forms) ofCephalopod Mollusks,” in The Results of Science: TheSeries “Zoology of Invertebrates,” Vol. 2: Mollusks ofCommercial Importance (Nauka, Moscow, 1973),pp. 8–59 [in Russian].
160. K. N. Nesis, “Evolution of Living Forms of CephalopodMollusks,” Tr. Inst. Okeanol. Akad. Nauk SSSR 101,124–142 (1975).
161. K. N. Nesis, “Ecological Evolution of Cephalopod Mol-lusks,” in Mollusks and Their System, Evolution, andRole in Nature: Collection of Papers No. 5 (Nauka,Leningrad, 1976), pp. 152–155 [in Russian].

1282
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
162. S. V. Nikolaeva, “Ammonoids from the BogdanovskyHorizon (Middle Carboniferous) of the South Urals,”Paleontol. Zh., No. 1, 12–19 (1999a) [Paleontol. J. 33(1), 10–18 (1999a)].
163. S. V. Nikolaeva, “Morphological Diversity of Ammonoidsfrom the Lower Namurian of Central Asia,” in Advanc-ing Research on Living and Fossil Cephalopods (NewYork, 1999b), pp. 295–313.
164. S. V. Nikolaeva, “An Assemblage of Ammonoids fromthe Dombar Limestone (The Southern Urals),” in Mod-ern Problems in the Study of Cephalopod Mollusks:Morphology, Systematics, Evolution, and Biostratigra-phy: Proceedings of All-Russia Conference (Paleontol.Inst. Ross. Akad. Nauk, Moscow, 2006), pp. 73–75[in Russian].
165. S. V. Nikolaeva and I. S. Barskov, “MorphogeneticTrends in the Evolution of Carboniferous Ammonoids,”N. Jb. Geol. Paläontol. Abh. 193 (Fasc. 3), 401–418(1994).
166. Nikolaeva S.V. and B. I. Bogoslovsky, “DevonianAmmonoids: IV. Clymeniines (Suborder Clymeniina),”Tr. Paleontol. Inst. Ross. Akad. Nauk 287, 1–220(2005a).
167. S. V. Nikolaeva and B. I. Bogoslovsky, “Late FamenianAmmonoids from the Upper Part of the Kiya Formationof the South Urals,” Paleontol. J. 39 (Suppl. 5), S527–S537 (2005b).
168. S. V. Nikolaeva, N. B. Gibshman, E. I. Kulagina,I. S. Barskov, and V. N. Pazukhin, “Correlation of theVisean–Serpukhovian Boundary in Its Type Region(Moscow Basin) and the South Urals and a Proposal ofBoundary Markers (Ammonoids, Foraminifers, Con-odonts),” Newsl. Carb. Stratigr. 20, 16–21 (2002).
169. Paleontological Dictionary, Ed. by G. A. Beznosovaand F. A. Zhuravleva (Nauka, Moscow, 1965), p. 117[in Russian].
170. V. N. Pazukhin and E. N. Gorozhanina, “Section‘Verkhnyaya Kardailovka’,” in Guidebook of Geologi-cal Excursions on the Carboniferous of the Ural Moun-tains: Part 1. Southern Ural Excursion (Inst. Geol.Geokhim. Ural. Otd. Ross. Akad. Nauk, Yekaterinburg,2002), pp. 59–67 [in Russian].
171. V. N. Pazukhin, E. I. Kulagina, S. V. Nikolaeva,K. N. Kochetova, and V. A. Konovalova, “Zonal Subdi-visions of the Upper Viséan and Serpukhovian Depositsin the Section Verkhnyaya Kardailovka (Eastern Slopeof the Southern Urals),” in Stratigraphy and Paleogeog-raphy of the Carboniferous of Eurasia: Collection ofScientific Papers (Inst. Geol. Geokhim. Ural. Otd. Ross.Akad. Nauk, Yekaterinburg, 2002), pp. 220–229 [inRussian].
172. E. Ya. Perna, “Ammonoids of the Upper Neodevonianof the Eastern Slope of the Ural Mountains,” Tr. Geol.Kom., N. Ser., Issue 99, 1–114 (1914).
173. E. G. Platonov and G. E. Chernyak, “CarboniferousDeposits of the Southern Island of Novaya Zemlya,” inGeology of the Southern Island of Novaya Zemlya (Sev-morgeologiya, Leningrad, 1982), pp. 25–36 [in Rus-sian].
174. A. V. Popov, “Ammonoids (Ammonoidea),” in Paleon-tological Atlas of the Carboniferous Deposits of theUral Mountains: Proceedings of the All-Union
Research Institute of Petroleum and Geological Explo-ration, Vol. 383 (Leningrad, 1975), pp. 111–130 [inRussian].
175. A. V. Popov and L. F. Kusina, “The Earliest Goniatitina(Ammonoidea) from the South Urals,” Paleontol. Zh.,No. 1, 30–36 (1997) [Paleontol. J. 31 (1), 28–34(1997)].
176. V. N. Puchkov, Paleogeodynamics of the Southern andMiddle Urals (Dauriya, Ufa, 2000) [in Russian].
177. C. Raunkiaer, “Types biologiques pour la géographiebotanique,” Dan. Vid. Selsk. Forhandl., No. 5, 345–437(1905).
178. D. M. Raup, “Geometric Analysis of Shell Coiling.General Problems,” J. Paleontol. 40, 1178–1190 (1966).
179. D. M. Raup, “Geometric Analysis of Shell Coiling:Coiling in Ammonoids,” J. Paleontol. 47 (1), 43–65(1967).
180. D. M. Rauser-Tschernoussowa, “Stratigraphy of theUpper Carboniferous and Artinskian Deposits of theBashkir Region of the Cis-Urals,” Tr. Inst. Geol. Nauk,Issue 105, Ser. Geol., No. 35, 3–21 (1949).
181. A. Remane, “Die Bedeutung der Lebenformtypen fürdie Ökologie,” Biol. Gen. 17 (1/2), 164–182 (1943).
182. R. A. Reyment, “Factors in the Distribution of FossilCephalopods: Part 3. Experiments with Exact Modelsof Certain Shell Types,” Bull. Geol. Inst. Univer. Upp-sala, N. S. 4, 7–41 (1973).
183. V. E. Ruzhencev, “Paleontological Notes of the Carbon-iferous and Permian Ammonoids,” Probl. Sov. Geol. 6(12), 1072–1088 (1936).
184. V. E. Ruzhencev, “Problem of the Carboniferous andPermian,” Byull. Mosk. O–va Ispyt. Prir., Otd. Geol. 15(5), 401–411 (1937).
185. V. E. Ruzhencev, “Ammonoids of the Sakmarian Stageand Their Stratigraphic Significance,” Probl. Paleontol.4, 187–285 (1938).
186. V. E. Ruzhencev, “Representatives of the FamilyDimorphoceratidae Hyatt in Carboniferous Deposits ofthe Ural Mountains,” Dokl. Akad. Nauk SSSR 56 (5),521–524 (1947a).
187. V. E. Ruzhencev, “A New Genus of the Family Cheilo-ceratidae in Namurian Deposits of the Ural Mountains,”Dokl. Akad. Nauk SSSR 57 (3), 281–284 (1947b).
188. V. E. Ruzhencev, “Some New Namurian Ammonitesfrom the Aktyubinsk Region,” Dokl. Akad. Nauk SSSR67 (4), 737–740 (1949a).
189. V. E. Ruzhencev, “Systematics and Evolution of theFamilies Pronoritidae Frech and Medlicottiidae Karpin-sky,” Tr. Paleontol. Inst. Akad. Nauk SSSR 19, 1–204(1949b).
190. V. E. Ruzhencev, “Upper Carboniferous Ammonites ofthe Ural Mountains,” Tr. Paleontol. Inst. Akad. NaukSSSR 29, 1–220 (1950).
191. V. E. Ruzhencev, “On the Record of the GenusPseudoparalegoceras in the Aktyubinsk Region of theKazakh SSR,” Dokl. Akad. Nauk SSSR 78 (4), 769–772(1951a).
192. V. E. Ruzhencev, “Lower Permian Ammonites of theSouthern Urals: 1. Ammonites of the Sakmarian Stage,”Tr. Paleontol. Inst. Akad. Nauk SSSR 33, 1–188(1951b).

PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
CEPHALOPODS IN THE MARINE ECOSYSTEMS OF THE PALEOZOIC 1283
193. V. E. Ruzhencev, “First Representatives of the FamilyWelleritidae from the Middle Carboniferous Depositsof the Ural Mountains,” Dokl. Akad. Nauk SSSR 84 (1),131–134 (1952a).
194. V. E. Ruzhencev, “Biostratigraphy of the SakmarianStage in the Aktyubinsk Region of the Kazakh SSR,”Tr. Paleontol. Inst. Akad. Nauk SSSR 42, 1–85 (1952b).
195. V. E. Ruzhencev, “On the Origin of the Family Shu-marditidae,” Dokl. Akad. Nauk SSSR 103 (6), 1107–1110 (1955).
196. V. E. Ruzhencev, “On Some New Genera ofAmmonoids,” Dokl. Akad. Nauk SSSR 107, 158–161(1956a).
197. V. E. Ruzhencev, “Lower Permian Ammonites of theUral Mountains: II. Ammonites of the ArtinskianStage,” Tr. Paleontol. Inst. Akad. Nauk SSSR 60, 1–271(1956b).
198. V. E. Ruzhencev, “Two New Genera of Goniatites fromthe Lower Namurian of the Southern Urals,” Dokl.Akad. Nauk SSSR 122 (2), 293–296 (1958).
199. V. E. Ruzhencev, Principles of Systematics, System, andPhylogeny of Paleozoic Ammonoids (Akad. NaukSSSR, Moscow, 1960) [in Russian].
200. V. E. Ruzhencev, “Main Ammonoid Assemblages of theCarboniferous Period,” Paleontol. Zh., No. 2, 3 (1965).
201. V. E. Ruzhencev, “Viséan Ammonoids in the SouthernUrals,” Paleontol. Zh., No. 1, 47–59 (1966).
202. V. E. Ruzhencev and M. F. Bogoslovskaya, “NamurianStage in the Evolution of Ammonoids: Early NamurianAmmonoids,” Tr. Paleontol. Inst. Akad. Nauk SSSR133, 1–382 (1971).
203. V. E. Ruzhencev and M. F. Bogoslovskaya, “NamurianStage in the Evolution of Ammonoids: Late NamurianAmmonoids,” Tr. Paleontol. Inst. Akad. Nauk SSSR167, 1–336 (1978).
204. R. A. Schecoldin, N. N. Sobolev, and V. P. Matveev,“Carboniferous Deposits of the Southern Area ofNovaya Zemlya,” Newslett. Carbon. Stratigr. 12, 17–20(1994).
205. O. H. Schindewolf, “Vergleichende Morphologie undPhylogenie der Anfangskammer Tetrabranchiates Ceph-alopoden,” Preuß. Geol. Landesanst. Abh. N. E.,Issue 148, 1–116 (1933).
206. S. A. Severtsov, “Darwinism and Ecology,” Zool. Zh. 16(4), 591–613 (1937).
207. A. A. Shevyrev, “Triassic Ammonoids,” Tr. Paleontol.Inst. Akad. Nauk SSSR 217, 1–184 (1986).
208. A. A. Shevyrev, “The Cephalopod Macrosystem: A His-torical Review, the Present State of Knowledge, andUnsolved Problems: 1. Major Features and OverallClassification of Cephalopod Mollusks,” Paleontol. Zh.,No. 6, 33–42 (2005a) [Paleontol. J. 39 (6), 606–614(2005a)].
209. A. A. Shevyrev, “Heteromorph Ammonoids of the Tri-assic: A Review,” Paleontol. J. 39 (Suppl. 5), 614–628(2005b).
210. A. A. Shevyrev, “The Cephalopod Macrosystem: A His-torical Review, the Present State of Knowledge, andUnsolved Problems: 2. Classification of NautiloidCephalopods,” Paleontol. Zh., No. 1, 43–52 (2006a)[Paleontol. J. 40 (1), 46–54 (2006a)].
211. A. A. Shevyrev, “The Cephalopod Macrosystem: A His-torical Review, the Present State of Knowledge, andUnsolved Problems: 3. Classification of Bactritoideaand Ammonoidea,” Paleontol. Zh., No. 2, 34–46(2006b) [Paleontol. J. 40 (2), 150–161 (2006b)].
212. V. N. Shimansky, “Superorder Nautiloidea: GeneralPart,” in Fundamentals of Paleontology: Mollusks:Cephalopods 1 (Akad. Nauk SSSR, Moscow, 1962),pp. 66–68 [in Russian].
213. V. N. Shimansky, Nautilids (Degree of Knowledge,Stratigraphic and Geographic Distribution, Stages ofDevelopment), Tr. Paleontol. Inst. Akad. Nauk SSSR170, 1–67 (1979).
214. V. N. Shimansky and F. A. Zhuravleva, Main Problemsin Systematics of Nautiloidea and Related Groups: Pro-ceedings of the Paleontological Institute, RussianAcademy of Sciences, Vol. 90 (Akad. Nauk SSSR, Mos-cow, 1961) [in Russian].
215. S. Skompski, M. Paszkowski, M. Krobicki, et al., “Dep-ositional Setting of the Devonian/Carboniferous Bio-hermal Bol’shaya Nadota Carbonate Complex. Subpo-lar Urals,” Acta Geol. Polon. 51 (3), 217–235 (2001).
216. D. B. Sobolev, “Ostracodes and Biostratigraphy of theTournaisian Stage in the Northern Urals (Ural. Otd.Ross. Akad. Nauk, Yekaterinburg, 2005) [in Russian].
217. N. N. Sobolev and V. P. Matveev, “BiostratigraphicRationale of the Stage Boundaries of the CarboniferousSystem in Novaya Zemlya,” in Stratigraphy and Paleo-geography of the Carboniferous of Eurasia (Inst. Geol.Geokhim. Ural. Otd. Ross. Akad. Nauk, Yekaterinburg,2002), pp. 288–292 [in Russian].
218. W. B. Sounders and A. R. H. Swan, “Morphology andMorphologic Diversity of Mid-Carboniferous (Namurian)Ammonoids in Time and Space,” Paleobiology 10 (2),195–228 (1984).
219. W. B. Sounders and E. A. Shapiro, “Calculation andSimulation of Ammonoid Hydrostatics,” Paleobiology12 (1), 64–79 (1986).
220. W. B. Sounders and D. M. Work, “Shell Morphologyand Suture Complexity in Upper CarboniferousAmmonoids,” Palebiology 22 (2), 189–218 (1996).
221. W. B. Sounders and D. M. Work, “Evolution of ShellMorphology and Suture Complexity in Paleozoic Prole-canitids, the Root-Stock of Mesozoic Ammonoids,”Paleobiology 23 (3), 301–325 (1997).
222. W. B. Sounders, D. M. Work, and S. V. Nikolaeva,“Evolution of Complexity in Paleozoic AmmonoidSutures,” Science 286, 760–763 (1999).
223. W. B. Sounders, D. M. Work, and S. V. Nikolaeva, “TheEvolutionary History of Shell Geometry in PaleozoicAmmonoids,” Palebiology 30 (1), 19–43 (2004).
224. C. Spinosa, W. M. Furnish, and B. F. Glenister, “TheXenodiscidae, Permian Ceratoid Ammonoids,” J. Pale-ontol. 49 (2), 239–283 (1975).
225. Ya. I. Starobogatov, “Xenoconchia and Their Signifi-cance for the Phylogeny and Systematics of CertainClasses of Mollusks,” Paleontol. Zh., No. 1, 3–18(1974).
226. Ya. I. Starobogatov, “The System of Cephalopod Mol-lusks,” in Systematics and Ecology of Cephalopod Mol-lusks (Zool. Inst. Akad. Nauk SSSR, Leningrad, 1983),pp. 4–7 [in Russian].

1284
PALEONTOLOGICAL JOURNAL Vol. 42 No. 11 2008
BARSKOV et al.
227. A. A. Sultanaev, “Reference Sections of the TournaisianStage of the Southern Urals: Usuili, Ryauzyak, andZigan,” in Reference Sections and Fauna of the Tour-naisian Stage in the Southern Urals: Proceedings of theAll-Union Research Institute of Petroleum and Geolog-ical Exploration, Issue 346 (Leningrad, 1973), pp. 8–67[in Russian].
228. A. R. H. Swan and W. B. Sounders, “Function andShape in Late Paleozoic (Mid-Carboniferous)Ammonoids,” Paleobiology 12 (2), 297–311 (1987).
229. W. C. Sweet, “Nautiloidea—Oncocerida,” in Treatiseon Invertebrate Paleontology, Part K (1964a), pp. 277–319.
230. W. C. Sweet, “Nautiloidea—Barrandeocerida,” in Trea-tise on Invertebrate Paleontology, Part K (1964b),pp. 368–382.
231. A. A. Tchernov, “Artinskian Stage: Ammonoids of theBasins of the Yaiva, Kos’va, and Chusovaya Rivers,”Byull. Mosk. O–va Ispyt. Prir., Nov. Ser. 20 (3–4), 270–401 (1907).
232. C. Teichert, “Der Bau der Actinoceroiden Ceph-aloloden,” Palaeontographica, Abt. A 78, 111–230(1933).
233. C. Teichert, “Nautiloid Cephalopods from the Devonianof Western Australia,” Royal Soc. W. Australia. J. 25,103–120 (1939).
234. C. Teichert, “Actinoceratoidea,” in Treatise on Inverte-brate Paleontology, Part K (1964), pp. 4–12.
235. C. Teichert, “Major Feature of Cephalopod Evolution.Essays in Paleontology and Stratigraphy, Raymond C.Moore Commemorative Volume,” Univ. Kansas Geol.Spec. Publ., No. 2, 162—210 (1967).
236. C. Teichert, “Main Features of Cephalopod Evolution,”in The Mollusca: Vol. 12. Paleontology and Neontologyof Cephalopods (Acad. Press, London, 1988), pp. 11–79.
237. L. Tokarenko, “Fauna of the Upper Devonian Lime-stones in the Vicinity of the Town of Verkhneural’sk inthe Orenburg Province,” Tr. O–va Estestvoisp. priKazansk. Univ. 37 (2), 1–40 (1903).
238. Treatise on Invertebrate Paleontology: Part L, Mol-lusca 4, Ammonoidea, Ed. by R. C. Moore (Univ. Kan-sas Press, Lawrence, 1957).
239. Treatise on Invertebrate Paleontology: Part K, Mol-lusca 3, Cephalopoda: General Features, Endocera-toidea, Actinoceratoidea, Nautiloidea, Bactritoidea,Ed. by R. C. Moore (Univ. Kansas Press, Lawrence,1964).
240. A. E. Trueman, “The Ammonite Body Chamber, withSpecial Reference to Buoyancy and Mode of Life of the
Living Ammonite,” Quart. J. Geol. Soc. London 96(384), 26–32 (1941).
241. F. N. Tschernyschew, “Fauna of the Middle and UpperDevonian of the Western Slope of the Ural Mountains,”Tr. Geol. Kom. 3 (3), I–XII 1–208 (1887).
242. A. P. Tyazheva, “Stratigraphy of the Devonian Depositson the Western Slope of the Southern Urals,” in Devo-nian Deposits of Bashkiria: Part 1: Stratigraphy (Akad.Nauk SSSR, Moscow, 1961), pp. 5–130 [in Russian].
243. E. V. Voinova, “Some Ammonoids from theBaigendzhinskii Horizon of the Artinskian Stage of theSouthern Urals,” Tr. Vses. Geol. Razved. Ob”ed., Issue352, 1–60 (1934).
244. P. Ward, “Comparative Shell Shape Distribution inJurassic–Cretaceous Ammonites and Jurassic–TertiaryNautiloids,” Paleontology 6 (1), 32–43 (1980).
245. P. Ward, “Shell Sculpture As a Defensive Adaptation inAmmonoids,” Paleobiology 7 (1), 96–100 (1981).
246. P. D. Ward and A. W. Martin, “On the Buoyancy of thePearly Nautilus,” J. Exp. Zool. 205, 5–12 (1978).
247. E. Warming, Om Planterigets lifsformer (Festsk. udg.Univ. Kjobenhavn, Kjobenhavn, 1908).
248. M. E. Yanishevskii, “Materials for Study of the Paleo-zoic Fauna of Novaya Zemlya,” Tr. Geol. Mineralog.Museum Akad. Nauk SSSR 5 (4), 73–116 (1926).
249. E. L. Yochelson, R. H. Flower, and G. F. Webers, “TheBearing of the New Late Middle Cambrian Monopla-cophoran Genus Knightoconus upon the Origin of theCephalopoda,” Lethaia 6, 275–310 (1973).
250. Yu. A. Yudina and M. N. Moskalenko, “Reference Sec-tions of the Frasnian Stage of Southern Timan,” in FieldExcursion Guidebook (Ukhta, 1997) [in Russian].
251. A. Zeiss, “Weichteile ectocochleater paläozoischerCephalopode in Röntgenaufnahmen und ihre paläontol-ogische Bedeuting,” Paläontol. Z. 43 (1–2), 13–27(1969).
252. F. A. Zhuravleva, “Devonian Nautiloids: Order Dis-cosorida,” Tr. Paleontol. Inst. Akad. Nauk SSSR 134,1−320 (1972).
253. F. A. Zhuravleva, “Septal Necks in Evolution of Cepha-lopods,” Paleontol. Zh., No. 3, 55–76 (1994).
254. F. A. Zhuravleva and L. A. Doguzhaeva, “CameralDeposits of Pseudorthocerids and Actinocerids throughthe Scanning Electron Microscope,” in Fossil Cephalo-pods: The Latest Advances in Their Study (Paleontol.Inst. Ross. Akad. Nauk, Moscow, 1999), pp. 213–269[in Russian].
Related Documents











