This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Catalytic activity of soluble versus immobilized cauliflower(Brassica oleracea) bud peroxidase-concanavalin A complexand its application in dye color removal
Farrukh Jamal a,n, Sangram Singh a, Tabish Qidwai b, Dushyant Singh a, P.K. Pandey a,G.C. Pandey c, M.Y. Khan d
a Department of Biochemistry, Dr. Ram Manohar Lohia Avadh University, Faizabad 224001, U.P., Indiab Department of Biotechnology, Faculty of Engineering and Technology, R.B.S. College, Agra, U.P., Indiac Department of Environmental Science, Dr. Ram Manohar Lohia Avadh University, Faizabad 224001, U.P., Indiad Department of Biotechnology, Babasaheb Bhimrao, Ambedkar University, Lucknow, UP, India
a r t i c l e i n f o
Article history:Received 19 April 2013Received in revised form25 May 2013Accepted 28 May 2013Available online 12 June 2013
Keywords:Cauliflower bud peroxidaseImmobilizationDye decolorizationContinuous reactorDenaturantsKinetics
a b s t r a c t
Biocatalytic activity of soluble against immobilized cauliflower (Brassica oleracea) bud peroxidase (CBP)on calcium alginate gel beads have been assessed; internal conditions including temperature, pH andstability with denaturants optimized for application in removing color of textile industrial effluent. CBP–concanavalin A complex expressed �84% of original peroxidase whereas in entrapped state on calciumalginate pectin gel the activity reduced to �64%. Immobilized CBP–Con A complex retained �56.6%activity even at 70 1C which was achieved at pH 4.0. Prolonged urea treatment resulted in activity loss ofentrapped CBP-Con A complex by �19.8%. With dioxane at 50% (v/v), entrapped CBP-Con A complexexhibited an activity of over 58.7%; whereas an increasing concentration of dimethylformide causedreduction in peroxidase activity to 35.8% at 80% (v/v). With immobilized proteins there was considerablecolor removal upto �90.6% and �81.1% from disperse red 19 and dye mixture (disperse red 19+disperseblack 9) respectively, in continuous two reactor system and total organic carbon analysis was quitecomparable to color loss. Entrapped CBP-Con A complex were reusable upto ten cycles in dye colorremoval. This study suggests that immobilized and entrapped CBP catalytic system can be efficientlyexploited for dye color removal from industrial effluent due to its sustainability, durability and reusabilityover its soluble counterpart.
& 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Synthetic dyes are difficult to remove from effluents by con-ventional biological processes as they are highly stable andresistant to microbial attack (Pala and Toket, 2002). Althoughphysico-chemical methods are available for dye color removal butbeing cost effective finds limited application (Moreira et al., 2000).Another concern of such approaches are the generation of inter-mediates and end products that are carcinogenic and mostly moretoxic than the dyes per se. Approaches utilizing biodegradativeabilities of some white rot fungi and peroxidases from vegetablesources seems promising (Liu et al., 2004; Jamal et al., 2012).Owing to their extracellular nonspecific free radical-based
enzymatic system, they can completely eliminate a variety ofxenobiotics as well as synthetic dyes, giving rise to nontoxiccompounds (Bezalel et al., 1997; Pointing, 2001).
Peroxidases are monomeric heme-containing and hydrogenperoxide dependent oxidoreductase glycoproteins. The scope andapplications of such enzymes in detoxification, dye decolorizationand removal of various toxic organic pollutants especially phenoliccompounds which contaminate water and industrial effluents isgaining prominence (Azevedo et al., 2003; Husain, 2006; Lopezet al., 2002). As compared to microbial treatment enzymatictreatment are potentially better due to shorter treatment period;operation of high and low concentrations of substrates; absence ofdelays associated with the lag phase of biomass, reduction insludge volume and ease of controlling the process. Further onimmobilization such enzymes are advantageous over their solublecounterparts due to enhanced stability, easier product recoveryand purification, protection against denaturants, proteolysis, andreduced susceptibility to contamination over prolonged usage(Zille et al., 2003). The strategy adopted for immobilization of
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/bab
Biocatalysis and Agricultural Biotechnology
1878-8181/$ - see front matter & 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.bcab.2013.05.005
n Corresponding author at: Department of Biochemistry, Dr. Ram Manohar LohiaAvadh University, Faizabad 224001, U.P., India. Tel.: +91 9415075554;fax: +91 05278 246330.
E-mail addresses: [email protected],[email protected] (F. Jamal).
Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321

Author's personal copy
peroxidases is limited primarily due to the use of commerciallyavailable enzyme/expensive supports (Norouzian, 2003). Amongthe known techniques employed for immobilization bioaffinitybased physical adsorption using crude homogenate is useful andcost effective, with additional benefit, such as proper orientationof enzyme on the support (Mislovicova et al., 2000).
Novel research in the area of enzyme technology has providedsignificant clues and approaches that facilitate large scale opti-mum utilization of enzymes by entrapping and immobilizing(Gomez et al., 2006; Husain and Husain, 2008). Although, suchenzyme system has its own limitations of undergoing leakage andloss, such leakages can be minimized by controlling the poredimensions. Alternatively, entrapping cross-linked or pre-immobilized enzyme preparations could be better and sustainableapproaches (Betancor et al., 2005).
The present study demonstrates the biocatalytic activity ofsoluble and entrapped cauliflower bud peroxidase (CBP)–Con Acomplex among various internal conditions and the performanceof immobilized system in dye color removal of glycosylatedBrassica oleracea bud peroxidase. B.oleracea popularly knownas cauliflower is widely planted in tropical areas and consumed asvegetables and has earlier been shown by our group to besignificantly effective in decolorizing synthetic recalcitrant dye.Salt fractionated immobilized CBP with lectin Con A wasentrapped into calcium alginate-pectin beads. A comparativestudy on the biocatalytic activity of immobilized form of CBP(CBP–Con A complex and CBP–Con A–calcium alginate–pectincomplex) under different experimental conditions has been pre-sented for using such enzymes effectively in waste water treat-ment. Its ability to decolorize dyes in the batch processes andcontinuous two reactor system has also been examined. This is thefirst report on the effectivity and suitability of immobilized CBP indye color removal.
2. Material and methods
2.1. Materials
Sodium alginate, bovine serum albumin, Concanavalin A,O-dianisidine HCl, Disperse Red 19 (DR19), Disperse Black 9(DB9) was procured from Sigma Chemical Co. (St. Louis, MO,USA). Dioxane, dimethylformide (DMF), sodium dodecyl sulfate(SDS) and pectin were obtained from SRL Chemicals, Mumbai,India. All other chemicals were of analytical grade. The cauliflowerbuds were procured from Narendra Dev University of Agricultureand Technology, Faizabad, U.P., India. The samples were asepticallytransferred into sterilized plastic bags.
2.2. Partial purification, protein estimation and cauliflowerbud peroxidase (CBP) activity assay
Cauliflower buds (�350 g) was homogenized in 700 ml of100 mM sodium acetate buffer, pH 5.6; salt purification andmeasurement of peroxidase activity was done as described byJamal et al. (2012) (Jamal et al., 2010). This preparation of proteinwas aliquoted and stored for further use. Protein was estimatedusing BSA as a standard protein and following the procedure ofLowry et al. (1951).
2.3. Preparation of insoluble CBP–Con A complex and entrapmentin calcium alginate–pectin beads
The peroxidase proteins (1200 U) were mixed with an increas-ing concentration of Con A (0.1–1.0 ml) in a series of tubes and theprecipitate (CBP–Con A complex) exhibiting maximum activity
was obtained following the procedure of Jamal et al. (2012) (Jamalet al., 2010). Calcium alginate capsules were prepared by extrusionusing a simple one-step process as described by Nigma et al.(1988). CBP–Con A complex (1230 U) was mixed with sodiumalginate (2.5%) and pectin (2.5%) in 10 ml of 100 mM sodiumacetate buffer (pH 5.6) to obtain beads following the method ofJamal et al. (2012) (Jamal et al., 2010).
2.4. Peroxidase activity assay and effect of enzyme loading
Peroxidase activity was determined by the method of Jamalet al. (2012) (Jamal et al., 2010). One unit (1.0 U) of enzyme activityis the amount of enzyme protein that catalyzes oxidation of 1.0mmole of o-dianisidine HCl per min at 37 1C into colored product.An increasing concentration of enzyme (110–1000 U) was mixed tocalcium alginate-pectin gel in a series of tubes. Expression ofloaded enzyme was monitored by assaying the peroxidase activity.
2.5. Measurement of encapsulation efficiency and leakage of CBP
CBP concentration was measured both in calcium chloridesolution and capsule to assess the encapsulation efficiency. Theencapsulated enzyme concentration was estimated by cutting thecapsules in half and placing it in 5 mL phosphate buffer (pH¼7.4)solution. In order to obtain encapsulated protein concentration ofprotein in buffer was measured according to the Lowry's assayafter 2 h (Nigma et al., 1988). The percentage of encapsulatedenzyme was obtained from the difference between initial proteinintroduced to the calcium chloride hexahydrate solution andencapsulated protein measured as mentioned above.
Enzyme leakage measurement was carried out by placingcapsules in a test tube filled with Tris buffer (pH¼8.0) for 18 h.The capsules were cut in half and placed in phosphate buffer(pH¼7.4) solution. The protein concentration was measuredaccording to Lowry's assay and the leakage percentage wascalculated from the differences between encapsulated protein atthe beginning of time interval and the value found according tothe above procedure (Nigma et al., 1988).
2.6. Optimization of internal conditions in the presenceof contaminants
2.6.1. Effect of time, temperature and pH on immobilized CBPCBP–Con A complex and calcium alginate–pectin entrapped
CBP–Con A complex (1.20 U) were incubated at 60 1C in 100 mMsodium acetate buffer (pH 5.6) for varying time interval to obtaintime activity plot following the method of Jamal et al. (2012)(Jamal et al., 2010). The temperature activity (20–90 1C) and pHactivity plots (pH 2–9) were obtained following similar methods.
2.6.2. Effect of urea, organic solvents, detergents, sodium azideand mercuric chloride on immobilized CBP
CBP–Con A complex and calcium alginate–pectin entrappedCBP–Con A preparations (1.20 U) were incubated separately in4.0 M urea; 10–80% (v/v) of water-miscible organic solvents;dioxane and DMF and increasing concentrations of sodium dode-cyl sulfate (SDS) and non-ionic detergents; Triton X-100 andTween-20 (0.5–6.0%, v/v); with sodium azide, mercuric chloride(HgCl2) (0.05–6.0 mM) prepared in 100 mM sodium acetate buffer(pH 5.6) at 37 1C for 1 h. Catalytic activity was monitored at all theindicated detergent concentrations (Jamal et al., 2010). Activity ofenzyme without exposures to contaminants was taken as control(100%).
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321312

Author's personal copy
2.6.3. Determination of Km and Vmax of immobilized B. oleraceaperoxidase with respect to o- dianisidine HCl
As described previously, the initial enzymatic activity was mea-sured at various concentrations of o-dianisidine HCl. The solutionshaving different concentrations of o-dianisidine HCl ranging from0.002 to 1.2 mM were treated with CBP (1.20 U) in the presence of18.0 mM H2O2 in 100 mM sodium acetate buffer, pH 5.6, at 37 1C for15 min.
3. Preparation of synthetic dye solutions and calculationof dye color removal
The synthetic solutions of disperse dyes (25–50 mg/ml) wereprepared in distilled water to examine their decolorization bysoluble and immobilized CBP. A mixture of disperse dyes consist-ing of DR19 and DB9 was prepared by mixing each dye in equalproportion of color intensity (Jamal et al., 2011). Dye decoloriza-tion was monitored by measuring the difference at the maximumabsorbance for each dye as compared with control experimentswithout enzyme on UV-visible spectrophotometer (JASCO V-550,Japan). Untreated dye solution (inclusive of all reagents except theenzymes) was used as control for calculation of percent decolor-ization. The dye decolourization was calculated as the ratio of thedifference of absorbance of treated and untreated dye to that oftreated dye and converted in terms of percentage. Five indepen-dent experiments were carried out in duplicate and the mean wascalculated.
4. Decolorization of dye solution by immobilized peroxidasein batch processes
The dye solution (200 mL) was treated with immobilized CBP(27.6U) in 100 mM sodium acetate buffer, pH 5.6 in the presence of0.2 mM riboflavin as redox mediator and 0.8 mM H2O2 for 1 h at37 1C. The treated samples were centrifuged at 3000g for 15 min.The residual dye concentration was measured spectrophotometri-cally at specific wavelength maxima of the dye. Untreated dyesolution was considered as control (100%) for the calculation ofpercent decolorization.
5. Continuous dye decolorization using a two-reactor system
For the continuous removal of dyes from solutions two-reactorsystemwas developed. A column (15�2.0 cm) was filled with 10 gentrapped Con A–CBP complex (1256U) connected to secondcolumn (15�2.0 cm) containing activated silica. Silica was acti-vated by heating in an oven (120 1C) for overnight and thenwashed thrice with 30 ml distilled water. The flow rate wasmaintained at 6.4 ml/h and the feed solution contained eitherDR19 or mixture of DR19 and DB9 in two independent reactorsystems. Dye solutions were run under the same experimentalconditions. Reactors were operated at room temperature (37 1C)for a period of 4 months. Immobilized CBP treated and activatedsilica gel adsorbed samples were collected at an interval of 20 daysand the absorbance of each sample was recorded.
Procedure for the dye decolorization was followed by centrifu-gation and clear diluted solution was considered for TOC (totalorganic carbon) determination. TOC was evaluated by using a TOCanalyzer. Each control and treated DR19 or mixture of dyes (DR19+DB9) was diluted 10-fold before determining their TOC content.
6. Results and discussion
Peroxidases in conjunction with hydrogen peroxide can act onspecific recalcitrant pollutants to remove them either by precipita-tion or transformation to often innocuous products. They canchange the characteristics of a given waste rendering it moreamenable for treatment (Kasam and Niceu, 1997). Their catalyticaction is extremely efficient and selective as opposed to chemicalcatalysts due to higher reaction rates, milder reaction conditions(relatively low temperature and in the entire aqueous phase pHrange) and greater stereospecificity (Mohan et al., 2002). Thoughmuch attention has been paid in the utilization of biocatalysts inseveral fields, their involvement has been felt very recently insolving the environmental problems (Mohan et al., 2005; Nicell1994). The use of free enzymes poses inherent limitations as thestability and catalytic ability of free enzymes decreases with thecomplexity of the effluents (Zille et al., 2003). Some of theselimitations are overcome by the use of enzymes in immobilizedform which can be used as catalysts with long lifetime (Zille et al.,2003; Rogalski et al., 1995). Immobilization with different poly-meric materials is studied for enzyme encapsulation along withtheir application in treatment of various pollutants. However,appropriate selection of encapsulation material specific to theenzyme and optimization of process conditions is still underintensive investigation.
Immobilizing enzymes directly from crude homogenate isrelatively a much cheaper approach (Gupta and Mattiason 1992).Although the immobilized form of bio-molecules holds commer-cial importance, protocols available for such preparations arelimited. Immobilization by adsorption is an effective procedurefor binding enzymes directly from partially purified preparationsor even from crude homogenates (Musthapa et al., 2004). Calciumalginate mediated entrapment is a simple, economical, effectiveand sustainable approach for using enzymes either from crudeextract or directly from partially purified preparations for detox-ification and degradation of phenolic compounds in waste water(Kulshrestha and Husain, 2006; Akhtar et al., 2005). In the presentstudy although commercial Con A has been used, the process canbe made cost effective by using simple techniques to isolate Con Afrom cheaper sources such as Jack Bean extract and using suchpreparations to improve immobilization yields of these glycosy-lated peroxidases.
Immobilization process is an articulated approach due toinvolvement of key amino acids that must be avoided to preventloss of enzymatic activity. In case of glycoenzymes, glycosylmoieties can safely be used in immobilizations as they do notparticipate in catalysis. Lectins are proteins which recognize andinteract with exposed carbohydrate moieties of glycoproteins andglycoenzymes. These proteins are useful in characterizing glyco-proteins and certain glycoenzymes have been immobilized onconcanavalin A (Con A) affinity matrices or as Con A–glycoenzymecomplexes (Mislovicova et al., 2000; Kulshrestha and Husain,2006). Peroxidase from cauliflower bud is a better choice thanother vegetable peroxidases as it is sufficiently thermostable,operates in a wide range of pH, economic and effective with lowconcentration of redox mediators in decolorizing recalcitrantsynthetic dyes (Jamal et al., 2011).
6.1. Purification of CBP and entrapment of CBP–Con A complexin calcium alginate–pectin beads
The specific activity of peroxidase preparation increased to3.5 fold over crude enzyme which exhibited an initial specificactivity of 98 U/mg of protein. This enzyme preparation was usedfor direct immobilization as enzyme–Con A complex. Peroxidasesin soluble states may leak out of beads over prolonged retention or
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321 313
Author's personal copy
repeated use. Therefore, in order to prevent the leaching ofenzymes from porous gel beads, these enzyme molecules werecomplexed with lectin concanavalin A. The insoluble CBP–Con Acomplex was subsequently entrapped into calcium alginate–pectingel. The best biocatalytic properties including lower enzymeleakage and higher enzyme encapsulation achieved when thecalcium chloride and sodium alginate solution were 2.5% w/v.By selecting these concentrations, encapsulation efficiency increases
to 93% and leakage decreases to lower than 6%. With 0.2 ml of Con A,the CBP–Con A complex expressed an activity of �84% which onentrapment into calcium alginate–pectin gel resulted in furtherdecrease (�64%) of peroxidase activity (Table 1). Further, the effectof enzyme loading on entrapped activity was evaluated by entrap-ping increasing concentration of enzyme. Optimum concentration(512 U/ml) was sufficient for maximum expression of peroxidaseactivity by entrapped preparation.
Table 1Activities of CBP immobilized with lectin Con A and calcium-alginate pectin gel and properties (Km and Vmax) of soluble and immobilized CBP.
Type of immobilization Original activity (%) Expressed activity (%) Enzyme property Enzyme preparation
Soluble CBP Immobilized CBP
CBP–Con A Complex 100 8472.02 pH optima 4.0 5.0CBP–Con A complex entrapped oncalcium-alginate pectin gel
100 6471.73 Temperature optima (1C) 40 40Km (mM) 0.076 0.089Vmax (mM) 22.4 16.6
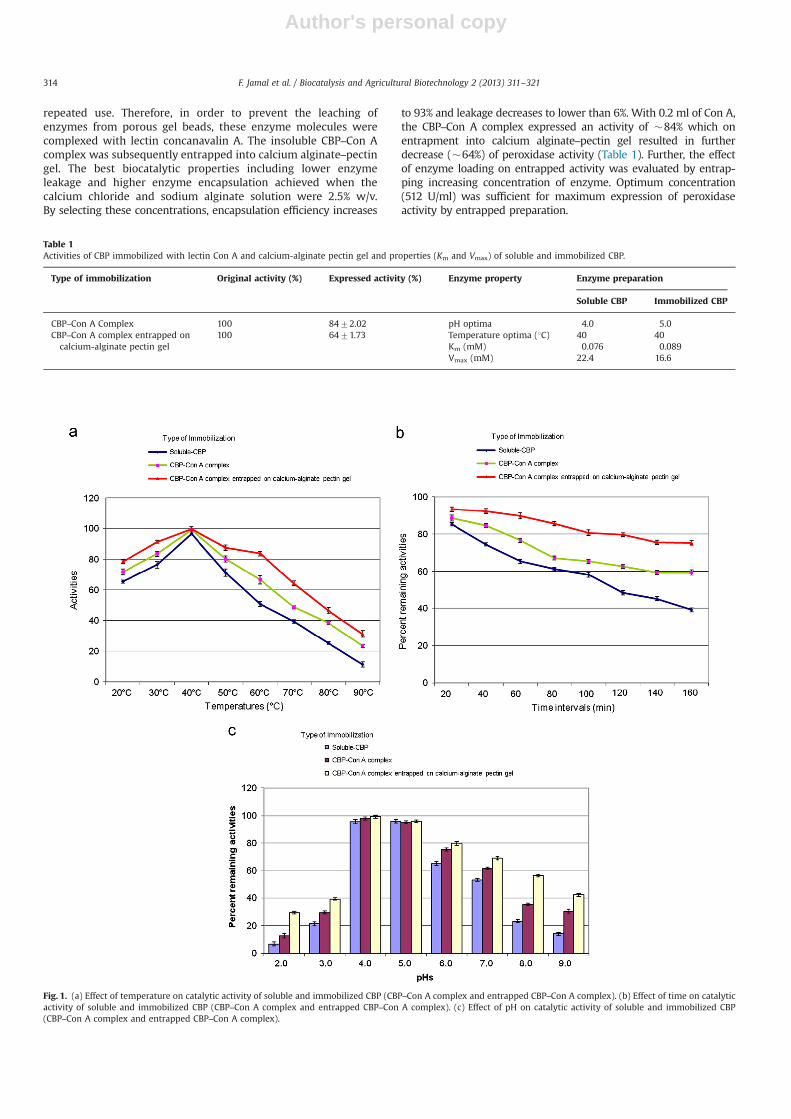
Fig. 1. (a) Effect of temperature on catalytic activity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex). (b) Effect of time on catalyticactivity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex). (c) Effect of pH on catalytic activity of soluble and immobilized CBP(CBP–Con A complex and entrapped CBP–Con A complex).
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321314
Author's personal copy
Pre-immobilized or cross-linked enzymes that remain insidepolymeric matrices for longer duration than their soluble counter-parts provide higher mechanical and operational stability(Betancor et al., 2005; Kulshrestha and Husain, 2006). It alsoindicated that enzymes with high molecular mass stay for longerduration inside polymeric matrix. Pre-immobilization increasesmolecular dimensioning of the enzyme and thereby prevents itsleaching from alginate beads.
6.2. Optimization of internal parameters with/without contaminantson soluble and immobilized CBP–Con A
6.2.1. Effect of time, temperature and pHImmobilized peroxidase preparations exhibited maximum per-
oxidase activity at 40 1C (Fig. 1a) which was true even for solublecounterpart of CBP. However, CBP–Con A and entrapped CBP–ConA complex retained greater fraction of catalytic activity at highertemperatures. CBP in the soluble state retained 48.4% of its initialactivity after 2 h incubation at 60 1C whereas CBP–Con A complexpreparation retained about 62.6% original activity under identical
incubation conditions. Interestingly, the entrapped CBP–Con Acomplex expressed 79.6% activity at 60 1C in an incubation periodof 2 h (Fig. 1b). The peroxidase activity was sufficiently high in thefirst 1 h for different immobilized forms (79.6% & 89.6% for CBP–Con A complex and entrapped states respectively) whereas arelatively progressive decline was observed with the soluble CBPon prolonging the time duration.
Immobilization enhanced the resistance/stability of enzyme tohigh temperatures. At reasonably high temperatures of 60 1C theenzyme expressed 66.6% and 83.7% peroxidase activity as CBP–ConA complex and calcium-alginate entrapped CBP–Con A complexrespectively (Fig. 1a). On increasing the temperature further to80 1C the entrapped immobilized form of CBP exhibited 46.5%peroxidase activity which declined to 31.2% at 90 1C. Calciumalginate–pectin entrapment of CBP–Con A complex contributedstrongly towards retention of its molecular structure, thus itsstability and consequently high activity was observed at elevatedtemperatures. Such enzyme preparation could certainly beexploited where operational temperatures are relatively high.Peroxidases being glycoprotein in nature, the lectin adsorptioncontributed towards improvement in thermal stability of calcium
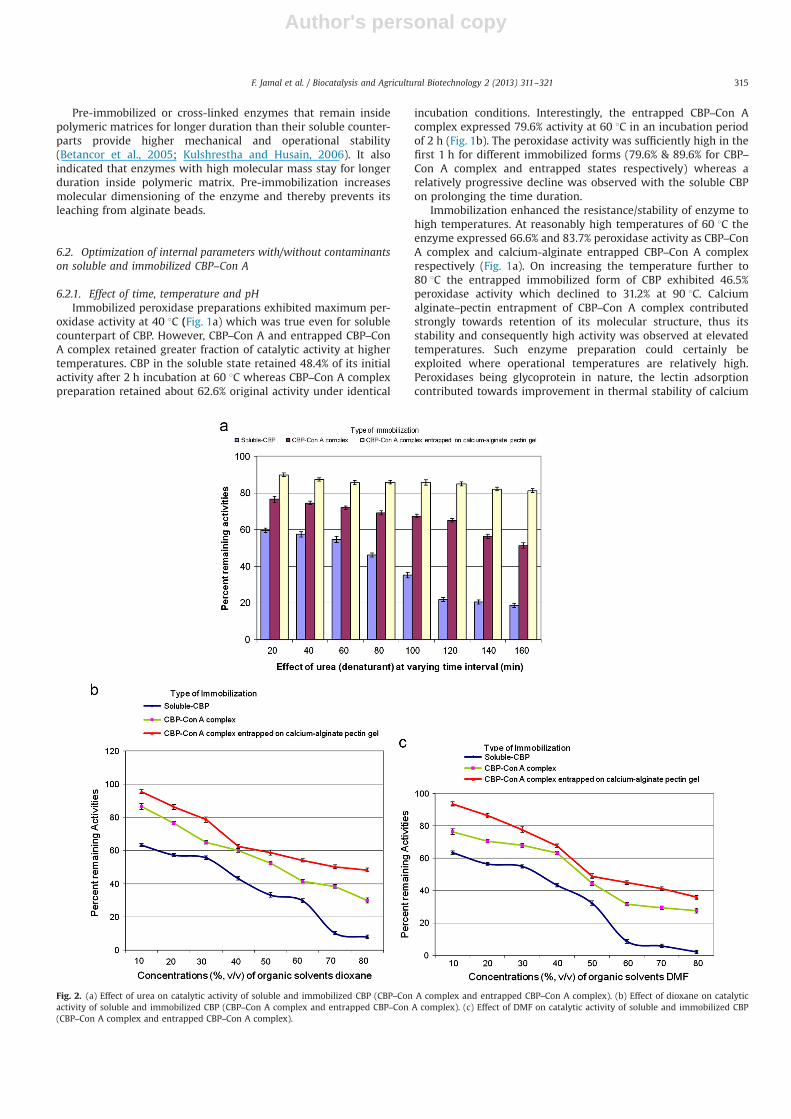
Fig. 2. (a) Effect of urea on catalytic activity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex). (b) Effect of dioxane on catalyticactivity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex). (c) Effect of DMF on catalytic activity of soluble and immobilized CBP(CBP–Con A complex and entrapped CBP–Con A complex).
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321 315
Author's personal copy
alginate-pectin entrapped CBP–Con A preparation possibly due tomultipoint attachment of peroxidases with Con A. This enhance-ment in thermal stability is possibly due to formation of severallinkages between the enzyme molecules and support matrix (Janand Husain, 2004).
pH activity profile of soluble CBP, CBP–Con A and entrappedCBP–Con A was evaluated by incubating these preparations in thebuffers of varying pH values (2.0–9.0). The pH range 4.0–6.0 wassufficient for optimum enzyme activity (Fig. 1c). Maximum activity(96%) at pH 5.0 was observed for soluble CBP whereas progressivedecline in activity was recorded in alkaline medium. On thecontrary the CBP–Con A complex and entrapped CBP–Con Aperformed optimally at pH 4.0 (98.1% and 99.4% respectively).Interestingly, these immobilized states of CBP retained remarkableactivity in alkaline medium as well as compared to soluble CBP.Moreover, the entrapped CBP–Con A retained more peroxidaseactivity (69.2%–42.2%) than CBP–Con A complex (61.6%–30.4%) inalkaline range from pH 7 to pH 9.
The immobilized CBP preparations exhibited significant broad-ening in pH activity profiles reflecting a marked increase instability (Fig. 1c). This broadening in pH-activity remarkablyincreased in case of entrapped CBP–Con A followed by CBP–ConA complex, suggesting that entrapment of enzymes in gel beadsprovides a microenvironment for enzyme, which may play an
important role in the state of protonation/deprotonation of proteinmolecules (Musthapa et al., 2004). Formation of CBP–Con Acomplex promotes retention of molecular structure and conse-quently confers additional resistance to enzyme against extremeconditions of pH (Akhtar et al., 2005; Lopez et al., 2004).
6.2.2. Effect of urea (denaturant)Soluble and immobilized CBP preparations were treated with
4.0 M urea for varying times. Soluble CBP lost nearly 78% of initialactivity on 2 h treatment whereas activity of Con A–CBP complexdiminished by �35%. Although, with time there was progressivedecline in the peroxidase activity, but the CBP–Con A complexretained much higher activity as against soluble CBP (Fig. 2a). Incase of entrapped CBP–Con A complex the activity retention wasprofound with the progressive increase in incubation time.The loss was in the range of �10%–18% in duration of 20–160 min.
Entrapped CBP–Con A preparation was remarkably resistant tourea induced inactivation and retained nearly 85% of its initialactivity even after 2 h incubation (Fig. 2a). On the contrary solubleCBP lost �90% of its activity under similar conditions. Urea (4.0 M)is a strong denaturant of some proteins and it irreversiblydenatures soluble CBP (Makhatadze and Privalor, 1992). Although,action mechanism of urea on protein structure is not yet
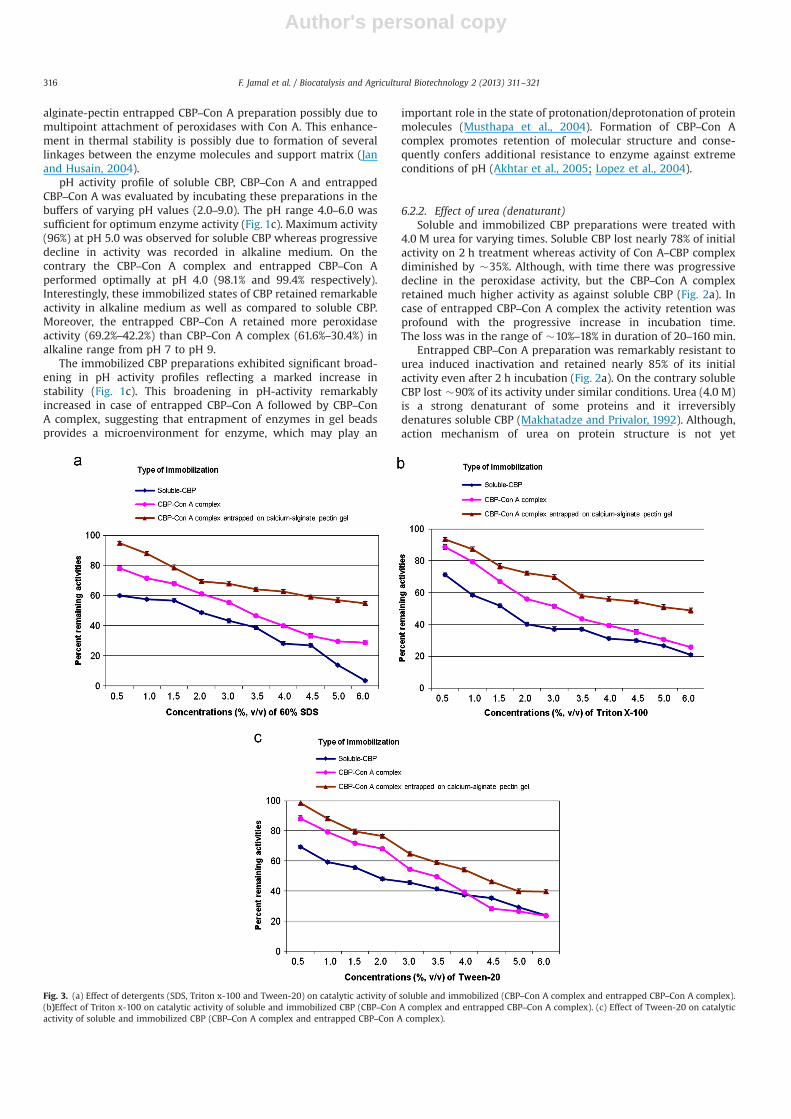
Fig. 3. (a) Effect of detergents (SDS, Triton x-100 and Tween-20) on catalytic activity of soluble and immobilized (CBP–Con A complex and entrapped CBP–Con A complex).(b)Effect of Triton x-100 on catalytic activity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex). (c) Effect of Tween-20 on catalyticactivity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex).
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321316
Author's personal copy
completely understood, several findings have indicated that pro-tein unfolding by direct interaction of urea molecule with apeptide backbone via non-covalent interactions, which contributesto maintenance of protein conformation (Musthapa et al., 2004;Akhtar et al., 2005). Complexing of glycoenzymes with Con A hasbeen reported in an enhancement of their resistance to denatura-tion mediated by urea (Fatima and Husain, 2007). Thus, theseobservations indicate that entrapment protected CBP–Con A com-plex from urea induced inactivation.
6.2.3. Effect of organic solventsAn increasing concentration of dioxane (10–80%, v/v) was used
to assess the activity of soluble and immobilized CBP. With lowerconcentration of dioxane (10–30%, v/v) the soluble and immobi-lized CBP exhibited over 60% of peroxidase activity, neverthelessactivities of immobilized counterparts was sufficiently high (86.6–64.9% for CBP–Con A complex and 95.4–78.6% for entrapped CBP–Con A complex, respectively) on treatment for 2 h. On exposure todioxane (60%, v/v), soluble enzyme retained only 29.8% of itsoriginal activity while CBP–Con A complex and entrapped CBP–Con A retained nearly 41.6% and 54.1% of their initial activity,respectively. Interestingly, at 80% v/v dioxane, soluble CBP lost�92% of peroxidase activity, whereas immobilized CBP exhibited29.9% and 48.2% activity (Fig. 2b).
A decrease in enzyme activity was observed of soluble andimmobilized CBP with increasing concentrations of DMF (10–80%,v/v). At lower concentration of DMF (10–30%) the soluble andimmobilized CBP exhibited sufficiently high activity (460%) andat 30% v/v concentration of DMF, activities expressed was 54.9% forsoluble, 67.9% and 77.6% for CBP–Con A and entrapped CBP–Con Arespectively. On exposure to DMF (60%, v/v) for 2 h, soluble CBPlost nearly 91.4% of its initial activity while Con A–CBP complexand entrapped Con A–CBP retained nearly 31.6% and 44.9% oforiginal activity, respectively (Fig. 2c). On increasing the concen-tration of DMF further, progressive decline in activities wereobserved and at 80% v/v DMF the soluble CBP lost �98% of activitywhereas immobilized CBP exhibited 27.6% (CBP–Con A complex)and 35.8% (entrapped CBP–Con A) activities.
Enzymes exploited for the treatment of wastewaters containingaromatic pollutants would be affected by the presence of water-miscible organic solvents. Therefore, we have investigated thestability of CBP preparations against some water miscible organicsolvents. It was also observed that entrapped CBP–Con A complexretained remarkably high stabilization against inactivation caused
by dioxane and DMF as compared to soluble CBP and CBP–Con Acomplex (Fig. 2c). Earlier reports also suggested that bioaffinitybound enzymes were significantly more stable perhaps due todecrease in flexibility and increased molecular rigidity againstexposure to water miscible organic solvents (Akhtar et al., 2005).
6.2.4. Effect of detergentsSeveral types of denaturants which contaminate wastewater
from various elimination sites include detergents that can stronglydenature enzymes used for treatment of polluted wastewater.Soluble and immobilized CBP were treated with increasing con-centrations of SDS, Triton X-100 and Tween-20 for 1 h at 37 1C.A progressive decline in the activities was observed for soluble andimmobilized CBPs with increasing concentration of both the non-ionic and ionic detergent. As compared to Triton X-100 andTween-20 the soluble CBP was adversely affected at 6.0% (v/v) ofSDS. Soluble CBP exhibited only 3.5% with 6% SDS whereas20.9% and 23.9% activity was retained on treatment with TritonX-100 and Tween-20 respectively. Entrapped CBP–Con A complexretained 54.7%, 48.8% and 39.5% of its initial activity in thepresence of 6.0% (v/v) SDS, Triton X-100 and Tween-20, respec-tively. However, CBP–Con A complex exhibited only 28.7%, 25.8%and 23.5% of initial activity in the presence of 6.0% SDS, TritonX-100 and Tween-20, respectively (Fig. 3a–c).
Our study also revealed that entrapped CBP–Con A complexwas markedly more stable to inactivation induced by detergents(SDS, Triton X-100 and Tween-20) (Fig. 3a–c). Entrapped CBP–ConA could work quite efficiently in the presence of contaminants likesoaps and detergents. Immobilized peroxidases are reported to besignificantly stabilized against denaturation induced by somecommonly used detergents (Musthapa et al., 2004). The immobi-lization of enzymes by multipoint attachment accords protectionfrom denaturation mediated by organic solvents. Further, thestabilization of immobilized enzymes against various forms ofwater-miscible organic solvents could possibly be due to lowwater requirement or enhanced rigidity of the enzyme structure.Enzymatic catalysis in organic solvents is possible if the organicsolvent does not substantially disturb the active site conformation(Ryu and Dorick, 1992).
6.2.5. Effect of enzyme inhibitor, metal ion chelator and HgCl2The effect of sodium azide on the activity of soluble and
immobilized CBP is shown in Fig. 4a. Soluble CBP lost 58.2% of
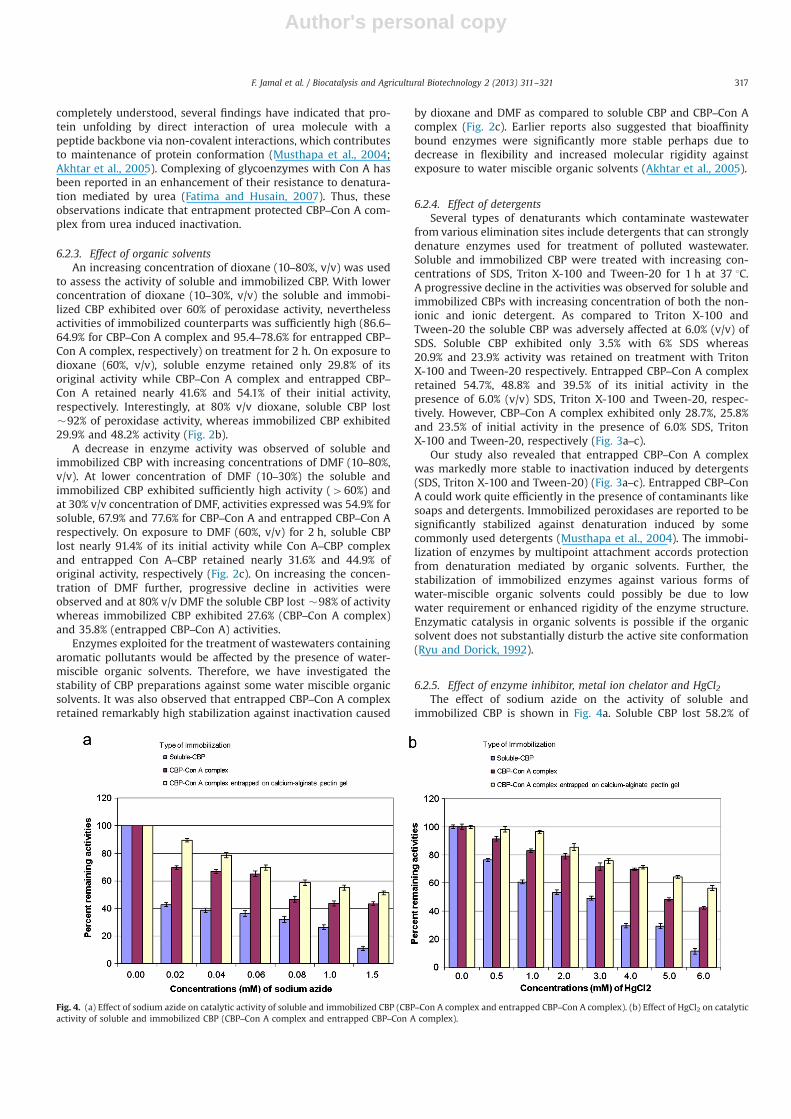
Fig. 4. (a) Effect of sodium azide on catalytic activity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex). (b) Effect of HgCl2 on catalyticactivity of soluble and immobilized CBP (CBP–Con A complex and entrapped CBP–Con A complex).
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321 317

Author's personal copy
its initial activity while entrapped CBP–Con A complex retained89.3% of its original activity after 1 h exposure to 0.02 mM sodiumazide. However, CBP–Con A complex retained an activity of 69.5%which was better than soluble CBP nevertheless lower thanentrapped CBP–Con A complex. With progressive increase inconcentration of sodium azide to 1.5 mM concentration therewas decrease in enzymatic activities of soluble and immobilizedCBP. However, the decrease in percent activity was highest in caseof soluble CBP (89.9% at 1.5 mM of sodium azide) whereas theCBP–Con A complex and entrapped CBP–Con A retained 43.4% and51.3% peroxidase activities, respectively. The effect on soluble andimmobilized CBP was also studied with metal ion chelator EDTAand at 30 mM concentration and there was no remarkable effecton their activities.
The peroxidase activity was concentration dependent in thepresence of HgCl2. At low concentrations of 1.0 mM immobilizedCBP exhibited peroxidase activities of 96.5% (entrapped CBP-Con Acomplex) and 82.9% (CBP-Con A complex) which was considerablyhigher than soluble CBP (60.7%). Upon increasing the concentra-tion of HgCl2 further to 4.0 mM soluble CBP lost 70.6% of initialactivity whereas immobilized states of CBP still retained over 69%of activities. However, at 6.0 mM HgCl2 soluble CBP lost 89.7%whereas immobilized states of CBP still exhibited above 40% ofperoxidase activity (Fig. 4b).
Sodium azide, an enzyme inhibitor was found to inhibit CBPstrongly (Fig. 4a). On the contrary, ethylenediamine tetra-aceticacid showed no significant effect on the activity of soluble andimmobilized CBP even when its concentration was raised 30 mM(data not given). A number of studies have already been performedon the inhibitory effect of such compounds such as horseradishperoxidase where sodium azide has been shown to be a potentinhibitor of many heme protein-catalyzed reactions (Kvaratskheliaand Winkel, 1997). Peroxidase in the presence of sodium azide andH2O2 mediates one electron oxidation of azide ions forming azidylfree radicals which bind covalently to the heme moiety ofperoxidase, thus inhibiting the enzyme activity (Tatarko andBumpus ,1997). EDTA did not have any significant effect on theactivity of CBP and such an observation on enzyme activity hasalready been reported (Lorenzo et al., 2005).
The chemical contamination of water by a wide range of toxicderivatives, particularly, heavy metals are a serious environmentalproblem owing to their potential human toxicity. In view of theirpresence in wastewater, it became necessary to evaluate the effectof some heavy metals on the activity of CBP. Our results revealedthat CBP exhibited more resistance to heavy metal inducedinhibition; a concentration-dependent gradual inhibition of CBPactivity by HgCl2 was observed (Fig. 4b). Some recent reportsindicated that horseradish peroxidase was remarkably inhibited byheavy metal ions (Keyhani et al., 2003; Einollahi et al., 2006).However, the inhibition of immobilized CBP by HgCl2 was quitelow as compared to the soluble enzyme. Although, metals induceconformational changes in enzymes, however peroxidases remainactive even in the presence of a number of metal ions, as a part oftheir detoxifying role. The effect of different metal ions on theactivity of different enzymes is related to their affinity to differentfunctional groups present in the enzymatic structure (Bagirovaet al., 2006; vel Krawczyk et al., 2000). The stability of immobilizedCBP against several metal compounds showed that such enzymepreparations could be exploited for the treatment of aromaticpollutants even in the presence of heavy metals.
7. Km and Vmax of soluble and immobilized CBP
The kinetic parameters of soluble and immobilized CBP weredetermined using different concentrations of o-dianisidine HCl.
The plot of initial enzyme activity versus different concentrationsof o-dianisidine HCl for both the enzyme preparations followed ahyperbolic pattern as expected according to the Michaelis–Mentonkinetics (data not given). Lineweaver–Burk plots of soluble andimmobilized CBP were also found to be linear and Michaelis–Menton constant, Km, for soluble and entrapped CBP–ConA complex was 0.076 and 0.089 mM, respectively (Table 1).The Vmax values for soluble and immobilized CBP were found tobe 22.4 and 16.6 mM/min, respectively.
Vmax and Michaelis–Menten constant (Km) for free and immo-bilized CBP were found from the Lineweaver–Burk plot. AlthoughKm values of soluble and immobilized CBP were close to eachother, Vmax of soluble enzyme was found to be more thanimmobilized CBP (Table 1).This observation suggested that immo-bilization of CBP by entrapment using calcium alginate pectinslightly altered the accessibility for the substrate whereas theconformational change in the enzyme was almost unaffected.Glutaraldehyde crosslinking enhances the structural rigidity ofprotein and also maintains the native structure (Wang et al., 2007).Immobilization by entrapment serves similar functions due towhich the CBP becomes more resistant to environment.
Since the entrapment of CBP–Con A complex is non-specificand masking of certain amino acids at or near the active sitecontributed towards lesser formation of enzyme-substrate com-plex a decrease in Vmax was observed in the case of immobilizedenzyme preparation. It is well documented that the Km values ofseveral immobilized enzymes were either unaltered or exhibited‘minor alteration as compared to those of their respective solublecounterparts (Jan et al., 2006).
8. Dye decolorization in batch process and a two-reactorsystem
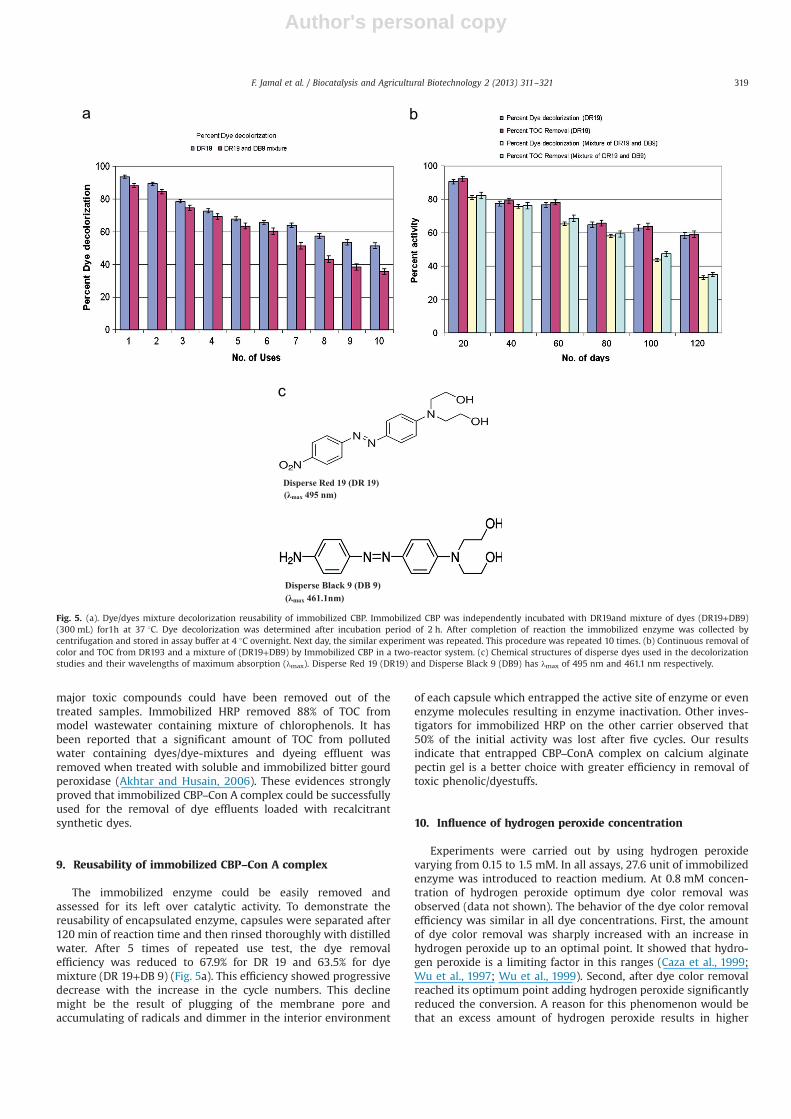
The decolorization of dyes by CBP has been illustrated in Fig. 5a.Entrapped immobilized CBP decolorized 93.7% and 88.2% of DR 19and mixture of dyes (DR19+DB9) after 2 h of incubation, respec-tively. Immobilized CBP was more effective as compared to itssoluble counterpart in the decolorization of both DR 19 andmixture of dyes. On the contrary the performance of the two-reactor system in terms of dye decolorization is shown in Fig. 5b.Entrapped Con A–CBP complex decolorized 90.6% and 81.1% of theinitial color from DR19 and dye mixture (DR19+DB9) , respectivelyafter 20 days. Considerable color removal from DR19 (64.8%) andmixture of disperse dyes (56.8%) was found even after 120 d and80 d respectively of operation of the two-reactor system. Thechemical structures dyes and its absorption maxima are shownin Fig. 5c.
The treatment of DR 19 and mixture of dyes (DR19+DB9) bypassing through a double reactor system provided almost thewater free from dyes. The dyes treated by immobilized CBPpresent in the first column, got adsorbed in the second column,which contained activated silica. Both the reactors worked formore than 120 days approximately, thus explaining their efficiencytowards dye decolorization. A significant loss of color appearedwhen DR19 or mixture of dyes was treated with entrapped CBP–Con A complex in the presence of redox mediator, riboflavin in acontinuous reactor system. It has earlier been reported that thedisappearance of peak in visible region was either due to thebreakdown of chromophoric groups present in dyes or theremoval of pollutants in the form of insoluble products (Moreiraet al., 2001).
In case of continuous reactors the level of TOC was significantlydecreased in the presence of immobilized CBP treated pollutedwater. However, the CBP treated dye solutions exhibited great lossof TOC from the wastewater (Fig. 5b), which suggested that the
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321318
Author's personal copy
major toxic compounds could have been removed out of thetreated samples. Immobilized HRP removed 88% of TOC frommodel wastewater containing mixture of chlorophenols. It hasbeen reported that a significant amount of TOC from pollutedwater containing dyes/dye-mixtures and dyeing effluent wasremoved when treated with soluble and immobilized bitter gourdperoxidase (Akhtar and Husain, 2006). These evidences stronglyproved that immobilized CBP–Con A complex could be successfullyused for the removal of dye effluents loaded with recalcitrantsynthetic dyes.
9. Reusability of immobilized CBP–Con A complex
The immobilized enzyme could be easily removed andassessed for its left over catalytic activity. To demonstrate thereusability of encapsulated enzyme, capsules were separated after120 min of reaction time and then rinsed thoroughly with distilledwater. After 5 times of repeated use test, the dye removalefficiency was reduced to 67.9% for DR 19 and 63.5% for dyemixture (DR 19+DB 9) (Fig. 5a). This efficiency showed progressivedecrease with the increase in the cycle numbers. This declinemight be the result of plugging of the membrane pore andaccumulating of radicals and dimmer in the interior environment
of each capsule which entrapped the active site of enzyme or evenenzyme molecules resulting in enzyme inactivation. Other inves-tigators for immobilized HRP on the other carrier observed that50% of the initial activity was lost after five cycles. Our resultsindicate that entrapped CBP–ConA complex on calcium alginatepectin gel is a better choice with greater efficiency in removal oftoxic phenolic/dyestuffs.
10. Influence of hydrogen peroxide concentration
Experiments were carried out by using hydrogen peroxidevarying from 0.15 to 1.5 mM. In all assays, 27.6 unit of immobilizedenzyme was introduced to reaction medium. At 0.8 mM concen-tration of hydrogen peroxide optimum dye color removal wasobserved (data not shown). The behavior of the dye color removalefficiency was similar in all dye concentrations. First, the amountof dye color removal was sharply increased with an increase inhydrogen peroxide up to an optimal point. It showed that hydro-gen peroxide is a limiting factor in this ranges (Caza et al., 1999;Wu et al., 1997; Wu et al., 1999). Second, after dye color removalreached its optimum point adding hydrogen peroxide significantlyreduced the conversion. A reason for this phenomenon would bethat an excess amount of hydrogen peroxide results in higher
Disperse Red 19 (DR 19)(λmax 495 nm)
Disperse Black 9 (DB 9)(λmax 461.1nm)
Fig. 5. (a). Dye/dyes mixture decolorization reusability of immobilized CBP. Immobilized CBP was independently incubated with DR19and mixture of dyes (DR19+DB9)(300 mL) for1h at 37 1C. Dye decolorization was determined after incubation period of 2 h. After completion of reaction the immobilized enzyme was collected bycentrifugation and stored in assay buffer at 4 1C overnight. Next day, the similar experiment was repeated. This procedure was repeated 10 times. (b) Continuous removal ofcolor and TOC from DR193 and a mixture of (DR19+DB9) by Immobilized CBP in a two-reactor system. (c) Chemical structures of disperse dyes used in the decolorizationstudies and their wavelengths of maximum absorption (λmax). Disperse Red 19 (DR19) and Disperse Black 9 (DB9) has λmax of 495 nm and 461.1 nm respectively.
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321 319

Author's personal copy
concentrations of intermediate products which inhibit the activityof enzyme, and/or that enzyme is inactivated by an excess ofhydrogen peroxide. The deviation of the aforementioned rationmight be the result of polymer produced in the catalytic processlarger than dimmer (Nicell et al., 1993).
Aromatic pollutants are present in the wastewaters of numerousindustries. Compliance with increasingly stringent discharge limitsrequires the development of technologically and economically effec-tive methods for reducing the quantity of these contaminantsreleased into the environment. The immobilized enzyme catalyzedpolymerization and precipitation processes shows potential forremoving aromatic compounds from industrial wastewaters whichare not removed effectively through conventional chemical/physicalor biological wastewater treatment methods (Kilbanov et al., 1983).While inorganic compounds require acidic conditions to be effective,they may increase costs associated with initial pH of waste waters,corrosion of hardware during treatment and pH neutralization ofwastes prior to their release. Enzyme based catalysis reflects pre-ference over intact organisms (containing a multitude of enzymes)because the isolated enzymes act with greater specificity, therebyallowing specific group of pollutants to be targeted for treatment;their potency can be better standardized, they are easier to handleand store and enzyme concentration is not dependent on bacterialgrowth rates. Enzymes are naturally occurring chemical species andour source comprises of readily renewable resources. Moreover,in contrast to many other enzymes, CBP retains its catalyticactivity over wide ranges of temperature, pH, contaminant concen-tration, consequently making it particularly suitable for application inindustrial environment. In addition unlike other enzymes withsimilar functions CBP is relatively non-specific in terms of its organicsubstrates, making it applicable for treatment of large variety ofaromatic pollutants.
A critical aspect that governs and probably limits the industrialapplications of enzymes is inactivation through mechanical, che-mical and thermal processes that interact to influence enzymeactivity. In particular inactivation of CBP can result from oxidationof the enzyme to inactive forms, phenoxy radical inhibition andadsorption and/or entrapment of the enzyme in precipitatingpolymers. Therefore reducing the cost of the catalysts has beenthe focus of much attention. We have demonstrated that thecatalytic lifetimes of pointed gourd peroxidase as well as CBPimproves significantly when the reaction is conducted at theoptimal pH, temperature while maintaining a low instantaneousenzyme concentration (Jamal et al., 2011). The efficiency of thecrude enzyme preparation was independent of its purity as earlierreported by Alberti and Klibanov (1982)). By immobilization usingentrapment on calcium alginate pectin beads, the apparent rate ofenzyme inactivation was reduced which allowed a significantreduction in enzyme requirements for treatment. This increasein enzyme lifetime represents a very significant saving in terms oftreatment costs.
11. Conclusions
The preparation and application of immobilized cauliflowerbud peroxidase in Ca-alginate beads for dye color removal fromaqueous solution was investigated. The experimental resultsobtained in the present work revealed the effectiveness of theencapsulated peroxidase in sustainable dye color removal. Theimmobilization of CBP by Con A in Ca-alginate provides immobi-lized derivatives with adequate protein content and storagestability. Moreover, the high percentages of immobilized activitymake them good alternatives to the corresponding soluble forms.The immobilization has a protective effect on CBP against inactiva-tion and/or inhibition and so higher dye color removal can be
reached with the same concentration of immobilized CBP as freeCBP. The performance of dye color removal was found to be highlydependent on enzyme dose, hydrogen peroxidase and aqueous pH.The encapsulated enzyme activity shows higher relative activity inacidic solutions over a broader range which are the most commonconditions appeared in waste stream. Enzyme retention activity,encapsulation and leakage percentage of enzymes are influencedby gel preparation condition and finding a proper value for abovequantities totally depends on alginate species used. The reusabilityexperiment showed that these biocatalysts can be used up to ninecycles without serious deficiency in their catalytic performance. Atwo-reactor system with simple operational protocol for decolor-ization/degradation of disperse dyes has been focused for thepotential future use of immobilized peroxidases. Interestingly, thedescribed system is developed with a cheaper biocatalyst that isquite effective in treating dyes continuously in a small laboratoryreactor. Thus, immobilized CBP preparations could be exploited fordeveloping bioreactors for the treatment of phenolic and otheraromatic pollutants including synthetic dyes present in industrialeffluents.
Acknowledgment
We are thankful to the Department of Science and Technology(DST-FIST) under the Ministry of Science and Technology forproviding financial assistance towards infrastructure developmentfor carrying out this work. There is no conflict of interest whatso-ever regarding this manuscript.
References
Akhtar, S., Husain, Q., 2006. Potential of immobilized bitter gourd (Momordicacharantia) peroxidase in the removal of phenols from polluted water. Chemo 651228–5.
Akhtar, S., Khan, A.A., Husain, Q., 2005. Simultaneous purification and immobiliza-tion of bitter gourd (Momordica charantia) peroxidases on bioaffinity support.J. Chem. Technol. Biotechnol. 80 198-5.
Alberti, B.N., Klibanov, A.M., 1982. Peroxidase for removal of hazardous aromaticsfrom industrial wastewaters. In: Exemer, J.H. (Ed.), In Detoxification ofHazardousWastes. Ann Arbor Science Publishers, Ann Arbor, MI 349-256.
Azevedo, A.M., Martins, V.C., Prazers, D.M., Vojinovic, V., Cabral, J.M., Fonseca, L.P.,2003. Horseradish peroxidase: a valuable tool in biotechnology. Biotechnol.Ann. Rev. 9 199-7.
Bagirova, N.A., Muginova, S.V., Shekhovtsova, T.N., Gazaryan, I.G., Van Huystee, R.B.,2006. Effect of mercury (II) traces on catalytic activity of peanut and horse-radish peroxidases. Anal. Lett. 39 521-1.
Betancor, L., Lopez-Gallego, F., Hidalgo, A., Fuentes, M., Podrasky, O., Kuncova, G.,Guisan, J.M., Fernandez-Lafuente, R., 2005. Advantages of the preimmobiliza-tion of enzymes on porous supports for their entrapment in sol–gels. Bioma-cromolecules 6 1027-0.
Bezalel, L., Hadar, Y., Cerniglia, C.E., 1997. Enzymatic mechanisms involved inphenanthrene degradation by the white rot fungus Pleurotus ostreatus. Appl.Environ. Microbiol. 63 2495-1.
Caza, N., Bewtra, J.K., Biswas, N., Taylor, K.E., 1999. Removal of phenolic compoundfrom synthetic wastewater using soybean peroxidase. Water Res. 33, 3012–8.
Einollahi, N., Abbasi, S., Dashti, N., Vaezzadeh, F., 2006. Effect of mercuric chlorideon kinetic properties of horseradish peroxidase. Iran J. Public Health 35 49–6.
Gomez, J.L., Bodalo, A., Gomez, E., Bastida, J., Hidalgo, A.M., Gomez, M., 2006.Immobilization of peroxidases on glass beads: an improved alternative forphenol removal. Microb. Technol. 39 1016-2.
Gupta, M.N., Mattiason, B., 1992. Unique application of immobilized proteins inbioanalytical systems. Meth. Biochem. Anal. 36, 1–4.
Fatima, A., Husain, Q., 2007. A role of glycosyl moieties in the stabilization of bittergourd (Momordica charantia) peroxidase. Int. J. Biol. Macro. 41 56-3.
Husain, M., Husain, Q., 2008. Applications of redox mediators in the treatment oforganic pollutants by using oxidoreductive enzymes: a review. Crit. Rev.Environ. Sci. Technol. 38, 1–42.
Husain, Q., 2006. Potential applications of the oxidoreductive enzymes in thedecolorization and detoxification of textile and other synthetic dyes frompolluted water: a review. Crit. Rev. Biotechnol. 60 201-1.
Jamal, F., Pandey, P.K., Qidwai, T., 2010. Potential of peroxidase enzyme fromTrichosanthes dioica to mediate disperse dye decolorization in conjunctionwith redox mediators. J. Mol. Catal. B Enzy. 66 177–1.
Jamal, F., Qidwai, T., Singh, R., Pandey, P.K., Azo, Singh S., 2011. and Anthraquinonedye decolourization in relation to its molecular structure using soluble
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321320

Author's personal copy
Trichosanthes dioica peroxidase supplemented with redox mediator. Catal.Commun. 12 1218-3.
Jamal, F., Qidwai, T., Singh, D., Pandey, P.K., 2011. Catalytic potential of cauliflower(Brassica oleracea) bud peroxidase in decolorization of synthetic recalcitrantdyes using redox mediator. Catal. Commun. 15, 93–8.
Jamal, F., Qidwai, T., Singh, D., Pandey, P.K., 2012. Biocatalytic activity of immobi-lized pointed gourd (Trichosanthes dioica) peroxidase -concanavalin A complexon calcium alginate pectin gel. J. Mol. Catal. B Enzy. 74 125-1.
Jan, U., Husain, Q., 2004. Preparation of a highly stable, very active and high-yieldmultilayered assembly of glucose oxidase using carbohydrate-specific poly-clonal antibodies. Biotechnol. Appl. Biochem. 39, 233–9.
Jan, U., Khan, A.A., Husain, Q., 2006. A study on the comparative stability ofinsoluble complexes of glucose oxidase obtained with concanavalin A andspecific polyclonal antibodies. World J. Microb. Biotechnol. 22, 1033–9.
Kasam, J., Niceu, J.A., 1997. Potential application of enzymes in waste treatment.J. Chem. Tech. Biotechnol. 69, 141–3.
Keyhani, J., Keyhani, E., Einollahi, N., Minai-Tehrani, D., Zarchipour, S., 2003.Heterogeneous inhibition of horseradish peroxidase activity by cadmium.Biochim. Biophys. Acta 1621, 140–8.
Kilbanov, A.M., Tu, T.M., Scott, K.P., 1983. Peroxidase-catalyzed removal of phenolsfrom coal conversion wastewater. Science 221 259–1.
Kulshrestha, Y., Husain, Q., 2006. Adsorption of peroxidase on DEAE-cellulosedirectly from ammonium sulfate fractionated proteins of bitter gourd. Enzy.Microb. Technol. 38, 470–7.
Kvaratskhelia, M., Winkel, C., 1997. Thorneley RNF. Purification and characteriza-tion of a novel class III peroxidase isoenzyme from tea leaves. Plant Physiol. 1141237–5.
Liu, W., Chao, Y., Yang, X., Buo, H., Qian, S., 2004. Biodecolourization of azo,anthraquinonic and triphenylmethane dyes by white-rot fungi and a laccasesecreting engineered strain. J. Ind. Microbiol. Biotechnol. 31 127-2.
Lopez, C., Mielgo, I., Moreira, M.T., Feijoo, G., Lema, J.M., 2002. Enzymatic membranereactors for biodegradation of recalcitrant compounds: application to dyedecolourisation. J. Biotechnol. 29 249–7.
Lopez, C., Moreira, M.T., Feijoo, G., Lema, J.M., 2004. Dye decolorization bymanganese peroxidase in an enzymatic membrane bioreactor. Biotechnol. Prog.20 74-1.
Lorenzo, M., Moldes, D., Rodriguez, C.S., Sanroma, M.A., 2005. Inhibition of laccaseactivity from Trametes versicolor by heavy metals and organic compounds.Chemo 60, 1124–8.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith Folin-phenol reagent. J. Biol. Chem. 193 265-5.
Makhatadze, G.I., Privalor, P.I., 1992. Protein interactions with urea and guanidi-nium hydrochloride: a calorimetric study. J. Mol. Biol. 226, 491–5.
Mislovicova, D., Gemeiner, P., Sandula, J., Masarova, J., Vikartovska, A., Doloo-mansky, P., 2000. Examination of bioaffinity immobilization by precipitation ofmannan and mannan-containing enzymes with legume lectins. Biotechnol.Appl. Biochem. 31, 153–9.
Mohan, S.V., Prasad, K.K., Prakasham, R.S., Sarma, P.N., 2002. Enzymatic pretreat-ment to enhance the biodegradability of industrial wastewater. Chem. Wkly 23,163–8.
Mohan, S.V., Prasad, K.K., Roa, N.C., Sarma, P.N., 2005. Acid azo dye degradation byfree and immobilized horseradish peroxidase (HRP) catalyzed process. Chemo58 1097-5.
Moreira, M.T., Melgo, I., Feijoo, G., Lema, J.M., 2000. Evaluation of different fungalstrains in the decolorization of synthetic dye. Biotechnol. Lett. 22 1499-3.
Moreira, M.T., Palma, C., Mielgo, I., Feijoo, G., Lema, J.M., 2001. In vitro degradationof a polymeric dye (Poly R-478) by manganese peroxidase. Biotechnol. Bioeng.75, 362–8.
Musthapa, S.M., Akhtar, S., Khan, A.A., Husain, Q., 2004. An economical, simple andhigh yield procedure for the immobilization/stabilization of peroxidases fromturnip roots. J. Sci. Ind. Res. 63, 540–7.
Nicell, J.A., 1994. Kinetics of horseradish peroxidase-catalysed polymerization andprecipitation of aqueous 4-chlorophenol. J. Chem. Technol. Biotechnol. 60,203–5.
Nicell, J.A., Bewtra, J.K., Biswas, N., Pierre, C.S., 1993. Enzyme catalyzed polymeriza-tion and precipitation of aromatic compound from wastewater. Can. J. Civ. Eng.20 725–5.
Nigma, S.C., Tsao, I.-F., Sakoda, A., Wang, H.Y., 1988. Techniques for preparinghydrogel membrane capsule. Biotechnol. Technol. 2, 271–6.
Norouzian, D., 2003. The state of art in biotechnology. Iran J. Biotechnol. 1 197–6.Pala, A., Toket, E., 2002. Color removal from cotton textile industry wastewater by
an activated sludge system with various additives. Water Res. 36, 2922–5.Pointing, S.B., 2001. Feasibility of bioremediation by white-rot fungi. Appl. Micro-
biol. Biotechnol. 57, 20–3.Rogalski, J., Jozwik, E., Hatakka, A., Leonomicz, A., 1995. Immobilization of laccase
from Phlebia radiata on controlled porosity glass. J. Mol. Catal. B Enzy. 95 99-8.Ryu, K., Dorick, J., 1992. How do organic solvents affect peroxidase structure and
function? Biochemistry 31 2588–8.Tatarko, M., Bumpus, J.A., 1997. Further studies on the inactivation by sodium azide
of lignin peroxidase from Phanerochaete chrysosporium. Arch. Biochem.Biophys. 339, 200–9.
Wang, J., Liang, Z., Wang, L., Fan, C., Li, G., 2007. Electron transfer reactivity andcatalytic activity of structurally rigidized haemoglobin. Sens. Act. B 125 17–1.
Wu, Y., Taylor, K.E., Biswas, N., Bewtra, J.K., 1997. Comparison of additive in theremoval of phenolic compound by peroxidase-catalyzed polymerization. WaterRes. 31 2699– 4.
Wu, Y., Taylor, K.E., Biswas, N., Bewtra, J.K., 1999. Kinetic model for removal ofphenol by horseradish peroxidase with PEG. J. Environ. Eng. 125, 451–8.
Zille, A., Tzanov, T., Gubitz, G.M., Cavaco-Paulo, A., 2003. Immobilized laccase fordecolorization of Reactive Black 5 dyeing effluent. Biotechnol. Lett. 25, 1473–7.
vel Krawczyk, T.K., Moszczynska, M., Trojanowicz, M., 2000. Inhibitive determina-tion of mercury and other metal ions by potentiometric urea biosensor.Biosens. Bioelectron. 15 681–1.
F. Jamal et al. / Biocatalysis and Agricultural Biotechnology 2 (2013) 311–321 321
Related Documents











