Cambrian Evolutionary Radiation: Context, correlation, and chronostratigraphy—Overcoming deficiencies of the first appearance datum (FAD) concept Ed Landing a, ⁎, Gerd Geyer b, c , Martin D. Brasier d , Samuel A. Bowring e a New York State Museum, 222 Madison Avenue, Albany, NY 12230, USA b Institut für Geographie und Geologie, Lehrstuhl für Geodynamik und Geomaterialforschung, Bayerische Julius-Maximilians-Universität Würzburg, Am Hubland, 97074 Würzburg, Germany c Department of Earth Sciences (Palaeobiology), Uppsala University, Villavägen 16, 752 36 Uppsala, Sweden d Department of Earth Sciences, Oxford University, South Parks Road, Oxford OX1 3AN, UK e Department of Earth, Atmospheric, and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, MA 02139, USA abstract article info Article history: Received 17 February 2012 Accepted 31 March 2013 Available online 20 April 2013 Keywords: Lower Cambrian FAD concept Global correlation Laolinian Stage Lenaldanian Series Zhurinskyan Stage Use of the first appearance datum (FAD) of a fossil to define a global chronostratigraphic unit's base can lead to intractable correlation and stability problems. FADs are diachronous—they reflect species' evolutionary history, dispersal, biofacies, preservation, collection, and taxonomy. The Cambrian Evolutionary Radiation is characterised by diachronous FADs, biofacies controls, and provincialism of taxa and ecological communi- ties that confound a stable Lower Cambrian chronostratigraphy. Cambrian series and stage definitions require greater attention to assemblage zone successions and non-biostratigraphic, particularly carbon isotope, cor- relation techniques such as those that define the Ediacaran System base. A redefined, basal Cambrian Trichophycus pedum Assemblage Zone lies above the highest Ediacaran-type biotas (vendobionts, putative metazoans, and calcareous problematica such as Cloudina) and the basal Asteridium tornatum– Comasphaeridium velvetum Zone (acritarchs). This definition and the likely close correspondence of evolu- tionary origin and local FAD of T. pedum preserves the Fortune Head, Newfoundland, GSSP of the Cambrian base and allows the presence of sub-Cambrian, branched ichnofossils. The sub-Tommotian-equivalent base of Stage 2 (a suggested “Laolinian Stage”) should be defined by the I′/L4/ZHUCE δ 13 C positive peak, bracketed by the lower ranges of Watsonella crosbyi and Aldanella attleborensis (molluscs) and the Skiagia ornata– Fimbrioglomerella membranacea Zone (acritarchs). The W. crosbyi and A. attleborensis FADs cannot define a Stage 2 base as they are diachronous even in the Newfoundland “type” W. crosbyi Zone. The Series 2 base cannot be based on a species' FAD owing to the provincialism of skeletalised metazoans in the Terreneuvian–Series 2 boundary interval and global heterochrony of the oldest trilobites. A Series 2 and Stage 3 (a suggested “Lenaldanian Series” and “Zhurinskyan Stage,” new) GSSP base is proposed at the Siberian lower Atdabanian δ 13 C IV peak—which correlates into South China, Avalonia, and Morocco and assigns the oldest trilobites to the terminal Terreneuvian Series. © 2013 Elsevier B.V. All rights reserved. Contents 1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 134 2. The problem with FADs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 135 2.1. Definition and use of FADs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 135 2.2. FAD diachroneity: from confidence intervals to taxon history . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 135 2.3. FAD diachroneity: taxonomy, sampling, unconformities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 136 2.4. Improved lowest Cambrian correlations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137 3. Revised terminal Ediacaran–Early Cambrian palaeogeography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137 4. Chronostratigraphy and Stage 1 of the Cambrian Radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142 4.1. Historical development of chronostratigraphy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142 4.2. Trichophycus pedum Zone and basal Cambrian correlation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142 Earth-Science Reviews 123 (2013) 133–172 ⁎ Corresponding author at: New York State Museum, Albany, NY, USA. Tel.: +1 5184745816; fax: +1 5184340022. E-mail addresses: [email protected] (E. Landing), [email protected] (G. Geyer), [email protected] (M.D. Brasier), [email protected] (S.A. Bowring). 0012-8252/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.earscirev.2013.03.008 Contents lists available at SciVerse ScienceDirect Earth-Science Reviews journal homepage: www.elsevier.com/locate/earscirev

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Earth-Science Reviews 123 (2013) 133–172
Contents lists available at SciVerse ScienceDirect
Earth-Science Reviews
j ourna l homepage: www.e lsev ie r .com/ locate /earsc i rev
Cambrian Evolutionary Radiation: Context, correlation, andchronostratigraphy—Overcoming deficiencies of the firstappearance datum (FAD) concept
Ed Landing a,⁎, Gerd Geyer b,c, Martin D. Brasier d, Samuel A. Bowring e
a New York State Museum, 222 Madison Avenue, Albany, NY 12230, USAb Institut für Geographie und Geologie, Lehrstuhl für Geodynamik und Geomaterialforschung, Bayerische Julius-Maximilians-Universität Würzburg, Am Hubland,97074 Würzburg, Germanyc Department of Earth Sciences (Palaeobiology), Uppsala University, Villavägen 16, 752 36 Uppsala, Swedend Department of Earth Sciences, Oxford University, South Parks Road, Oxford OX1 3AN, UKe Department of Earth, Atmospheric, and Planetary Sciences, Massachusetts Institute of Technology, Cambridge, MA 02139, USA
⁎ Corresponding author at: New York State Museum,E-mail addresses: [email protected] (E. Landin
0012-8252/$ – see front matter © 2013 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.earscirev.2013.03.008
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 February 2012Accepted 31 March 2013Available online 20 April 2013
Keywords:Lower CambrianFAD conceptGlobal correlationLaolinian StageLenaldanian SeriesZhurinskyan Stage
Use of the first appearance datum (FAD) of a fossil to define a global chronostratigraphic unit's base can leadto intractable correlation and stability problems. FADs are diachronous—they reflect species' evolutionaryhistory, dispersal, biofacies, preservation, collection, and taxonomy. The Cambrian Evolutionary Radiationis characterised by diachronous FADs, biofacies controls, and provincialism of taxa and ecological communi-ties that confound a stable Lower Cambrian chronostratigraphy. Cambrian series and stage definitions requiregreater attention to assemblage zone successions and non-biostratigraphic, particularly carbon isotope, cor-relation techniques such as those that define the Ediacaran System base. A redefined, basal CambrianTrichophycus pedum Assemblage Zone lies above the highest Ediacaran-type biotas (vendobionts, putativemetazoans, and calcareous problematica such as Cloudina) and the basal Asteridium tornatum–
Comasphaeridium velvetum Zone (acritarchs). This definition and the likely close correspondence of evolu-tionary origin and local FAD of T. pedum preserves the Fortune Head, Newfoundland, GSSP of the Cambrianbase and allows the presence of sub-Cambrian, branched ichnofossils. The sub-Tommotian-equivalent baseof Stage 2 (a suggested “Laolinian Stage”) should be defined by the I′/L4/ZHUCE δ13C positive peak, bracketedby the lower ranges of Watsonella crosbyi and Aldanella attleborensis (molluscs) and the Skiagia ornata–Fimbrioglomerella membranacea Zone (acritarchs). The W. crosbyi and A. attleborensis FADs cannot definea Stage 2 base as they are diachronous even in the Newfoundland “type” W. crosbyi Zone. The Series 2base cannot be based on a species' FAD owing to the provincialism of skeletalised metazoans in theTerreneuvian–Series 2 boundary interval and global heterochrony of the oldest trilobites. A Series 2 andStage 3 (a suggested “Lenaldanian Series” and “Zhurinskyan Stage,” new) GSSP base is proposed at theSiberian lower Atdabanian δ13C IV peak—which correlates into South China, Avalonia, and Morocco andassigns the oldest trilobites to the terminal Terreneuvian Series.
© 2013 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1342. The problem with FADs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 135
2.1. Definition and use of FADs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1352.2. FAD diachroneity: from confidence intervals to taxon history . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1352.3. FAD diachroneity: taxonomy, sampling, unconformities . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1362.4. Improved lowest Cambrian correlations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 137
3. Revised terminal Ediacaran–Early Cambrian palaeogeography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1374. Chronostratigraphy and Stage 1 of the Cambrian Radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142
4.1. Historical development of chronostratigraphy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1424.2. Trichophycus pedum Zone and basal Cambrian correlation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142
Albany, NY, USA. Tel.: +1 5184745816; fax: +1 5184340022.g), [email protected] (G. Geyer), [email protected] (M.D. Brasier), [email protected] (S.A. Bowring).
rights reserved.

134 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
4.3. Eurytopic T. pedum organism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1424.4. Problems with the T. pedum FAD? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 142
5. Improved definition for the base of the Cambrian . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1435.1. Carbon isotope stratigraphy and geochronology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1435.2. Biotic successions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1435.3. Utility of the basal Cambrian GSSP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1455.4. Refined definition of the T. pedum Zone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 145
6. Stage 2 and the Cambrian Evolutionary Radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1456.1. Historical development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1456.2. Micromollusks and Stage 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1456.3. W. crosbyi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 146
6.3.1. Taxonomy and distribution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1466.3.2. Occurrence in South China . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1466.3.3. Range offset in West Gondwanan France . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1466.3.4. Range offsets in Mongolia and Australia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 147
6.4. A. attleborensis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1477. Stratigraphic ranges of W. crosbyi and A. attleborensis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 147
7.1. Avalonia and stage 2 of the Cambrian Radiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1477.2. Watsonella and Aldanella are not Tommotian-limited . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1477.3. Diachroneity of Avalonian W. crosbyi Zone base . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1487.4. “Falling FADs” of Siberian Watsonella and Aldanella . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1487.5. Biotic correlation of upper Watsonella–Aldanella associations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1497.6. Carbon isotopes and uppermost Watsonella–Aldanella association . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1497.7. S. imbricata Zone and carbon isotopes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 149
8. Correlation of the “type” W. crosbyi Zone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1508.1. Biostratigraphy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1508.2. Carbon isotope correlation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 150
9. Acritarchs and “type” W. crosbyi Zone correlation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1509.1. Sub-W. crosbyi Zone correlation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1509.2. Acritarchs from W. crosbyi Zone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1519.3. Correlation of S.–F. Zone acritarchs from W. crosbyi Zone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1519.4. Acritarch correlation of Avalonian lowest W. crosbyi Zone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 151
10. Significance of 528 Ma volcanic ash in Cambrian Stage 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15110.1. 528 Ma ash and pre-Tommotian skeletalised metazoans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15110.2. Acritarch correlation and the 528 Ma ash . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 152
11. “Laolinian Stage” of the upper Terreneuvian Series . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15211.1. Definition of concept and GSSP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15211.2. Metazoan fossils and a basal GSSP of the “Laolinian Stage” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15311.3. Acritarch-based definition for the base of Cambrian Stage 2? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15311.4. Carbon isotopes and chronostratigraphy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15411.5. Biostratigraphic and carbon isotope brackets on the “Laolinian Stage” base . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154
12. Third stage of the Cambrian Evolutionary Radiation—appearance of trilobites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15412.1. Terreneuvian trilobites? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15412.2. Taxonomic and community evolution, lithofacies, and trilobite FADs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154
13. Earliest trilobites . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15513.1. Baltica . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15513.2. West Gondwana—southern Morocco . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15513.3. Iberia and Normandy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15613.4. Avalonia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15613.5. Laurentia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15713.6. Siberian Platform . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15813.7. South China Platform . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15813.8. Australian East Gondwana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 159
14. Defining a Series 2 base . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16014.1. Earliest trilobites and their diachroneity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16014.2. Non-trilobite FADs for the Series 2 base . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
15. Suggested “Lenaldanian Series” and “Zhurinskyan Stage” (new) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16215.1. Definition, concepts, location, etymology, and preservation of GSSP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16215.2. Chronostratigraphic significance of “Lenaldanian Series” and “Zhurinskyan Stage” GSSP . . . . . . . . . . . . . . . . . . . . . . . 163
16. Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 163Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 164References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 165
1. Introduction
The International Commission on Stratigraphy (ICS) has workedsince 1974 to establish a standardardised geological time scale withformally defined global chronostratigraphic units (i.e., eonothems,systems, series, and stages). Global boundary stratotype section and
points (GSSPs) have been established for almost all of the chrono-stratigraphic divisions of the Phanerozoic. The Cambrian remainsthe only Phanerozoic system for which all of its component seriesand stages have not been formally named and defined.
One difficulty that has slowed completion of a unified Cambrianchronostratigraphy is the strong biotic provincialism that developed

135E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
after fragmentation of the late Precambrian supercontinent Rodinia(e.g., Torsvik et al., 1996). Most skeletalised metazoans having poten-tial for biostratigraphy at this level (e.g., Brasier, 1989) are thought tohave been benthic. Unfortunately, some of the pelagic/nekticskeletalised metazoans used for global correlation later in the Phaner-ozoic (e.g., euconodonts, cephalopods, graptoloids) only appeared inthe latest Cambrian (e.g., Landing et al., 2010). Part of the problemmay be that pelagic metazoan planktivores and macropredatorswere essentially unmineralised and are rare in the Cambrian fossil re-cord (e.g., Butterfield, 1995, 2001). A possible exception includesagnostoid arthropods, commonly thought to be pelagic and amongthe best aids to Cambrian correlation, but these also may have beenbenthic and associated with dysoxic water masses (e.g., Fatka andSzabad, 2011).
Despite these difficulties, Cambrian chronostratigraphy hasprogressed with the adoption of four global, series-level divisions(Landing, 1998a,b; Geyer and Shergold, 2000) divided into ten stages(Geyer and Shergold, 2000; Peng and Babcock, 2005b; Babcock et al.,2005). The lowest and upper series (Terreneuvian and Furongian)have been defined. However, with exception of the lowest CambrianFortunian Stage (Landing et al., 2007a), most formally named Cambri-an stages are Middle and Upper Cambrian (Peng and Babcock, 2011).
A stage-level division of the lowest Cambrian Terreneuvian Series andthe overlying, formally unnamed, upper Lower Cambrian Series 2, whichcommonly has the oldest trilobites (e.g., Zhu et al., 2006), has remainedelusive. This condition reflects Early Cambrian biotic provincialism, andthe problems of correlating and constructing a chronostratigraphy forthe facies-specific developments of the Cambrian Radiation as firstoutlined by Brasier (1979). The three successive stages of the CambrianRadiation—the diversification of coelomates that produced deep andcomplex burrows at ca. 541 Ma, the diversification of metazoans withmineralised skeletons at ca. 532 Ma, and appearance of trilobites and di-verse lingulate brachiopods at ca. 520.5 Ma (Brasier, 1989; Landing et al.,1989; Landing and Westrop, 2004; Maloof et al., 2010a; Erwin et al.,2011)—correspond broadly to the lowest three stages of the Cambrian(e.g., Zhu et al., 2008). The Cambrian Evolutionary Radiation and aLower Cambrian global chronostratigraphy based on it encompass thegrowing complexity of benthic communities (e.g., Sheehan, 1996;Landing and Westrop, 2004) and the radiation of microphytoplanktonand mesozooplankton (Moczydłowska, 1981; Buttterfield, 2001).
The diversification of coelomate burrowers in shallow, sub-littoral fa-cies provided a basis for defining the base of the Cambrian (Narbonne etal., 1987; Brasier et al., 1994a; Landing, 1994). However, the diversifica-tion of skeletalised metazoans that characterise Stage 2 seems to havetaken place in very shallow marine facies, with the subsequent offshorespread of temporally long-ranged taxa taking a considerable amount oftime—about equivalent to that of a geological age. Similarly, the onset ofbiomineralisation of early trilobites seems to have begun in offshore fa-cies, with trilobites being exceptionally rare in coeval peritidal environ-ments (Landing and Westrop, 2004).
Diachroneity in the lowest occurrences of diverse skeletalisedmetazoan communities and the likely offshore origin of trilobites(Landing and Westrop, 2004) suggest the limitations of a fossil-basedchronostratigraphy of the later part of the Early Cambrian. This leadsus to question the utility of defining the base of Stage 2 of theLower Cambrian at a horizon defined by the FADs of a small shelledmollusk (e.g., Parkhaev et al., 2011; Steiner et al, 2011), or the use ofthe lowest occurrence of trilobites to define the base of CambrianSeries 2 and Stage 3 (Zhu et al., 2006; Babcock and Peng, 2007;Yuan et al., 2011).
Palmer's (1998b) report titled “Why is intercontinental correlationin the Lower Cambrian so difficult?” summarises the consequences offaunal provincialism and facies control on Lower Cambrian biostrati-graphic correlation. As a belated reply to Palmer (1998b), this report re-views the “state of the art” in Lower Cambrian chronostratigraphy andproposes improved and stable global chronostratigraphic units. The
chronostratigraphic units proposed herein are based on multiplecriteria, and we consider them more rigorously defined than existingproposals of chronostratigraphic units largely based on singlespecies-based correlations. [In this report, “Lower”/“Early,” “Middle”/“Middle,” and “Upper”/“Late” Cambrian are informal subsystems andsubperiods that equal the Terreneuvian + Series/Epoch 2, Series/Epoch 3, and Furongian Series/Epoch, respectively (Landing, 2007). Inaccordance with stratigraphic recommendations, the two or threemajor divisions of a system or period are capitalised (North AmericanCommission on Stratigraphic Nomenclature, 1983; Salvador, 1994)].
2. The problem with FADs
2.1. Definition and use of FADs
Regional and global chronostratigraphic units can be defined, subdi-vided, and correlated by various biostratigraphic and non-conventionaltechniques (North American Stratigraphic Commission, 1983; Salvador,1994). However, the ICS and its subcommissions have tended to empha-sise the “first [actually “lowest”] appearance datum” (FAD) of selectedfossil organisms to define and correlate the bases of global chrono-stratigraphic units. A selected species' FAD is then used to define a globalboundary stratotype section and point (GSSP)—being the precise place ina selected sedimentary rock section that defines the base of a globalchronostratigraphic unit. The top of this unit is then defined by the baseof the overlying chronostratigraphic unit (Cowie, 1986; Cowie et al.,1986).
Fossils that provide globally useful, interregionally correlatable FADsare rare, but are identified by consensus by an ICS subcommission.These FADs should be placed in the context of all available informa-tion, such as geochronometric and chemo- and magnetostratigraphicbrackets. A test that a given FAD is approximately coeval between local-ities with stratigraphically unbroken, monofacial successions can alsobe based on the fossil's uniformity of lowest occurrence in a verticalsuccession of biotas (Van Couvering and Berggren, 1977). A cautioususe of FADs has led to the definition of primarily fossil-defined chrono-stratigraphic units through much of the Phanerozoic.
2.2. FAD diachroneity: from confidence intervals to taxon history
Attempts have been made to use this FAD concept for global sub-division of the Lower Cambrian (e.g., Brasier, 1989). One obviousproblem with this approach is the presumption that the FAD of achronostratigraphically important fossil is essentially simultaneousin geographically separated successions. However, the FAD and thelast appearance datum (LAD) of a fossil even in thoroughly collected,stratigraphically continuous, monofacial successions always underes-timate the true range of a fossil species in every stratigraphic section.The simple arithmetic of confidence intervals in the stratigraphicranges of fossils (Marshall, 1990) emphasises that even with thebest supplementary brackets, the seemingly coeval FADs of a fossilbetween sections are not time-equivalent; they are just temporallyindistinguishable. Indeed, the stratigraphic interval not representedin every section between the known FAD of a fossil organism andthe organisms evolutionary origin is its range offset (Patzkowskyand Holland, 2012).
Diachroneity in a FAD is possible even between stratigraphicallycontinuous, similar successions. Interregional diachroneity in FADs(Fig. 1) can occur even with low global provincialism. This patternis shown, for example, by the apparently slow dispersal rate ofsome fusulinids that appear a geological stage higher in northern Car-boniferous localities than in the low latitude Tethyan realm (Mamet,1977) (Fig. 1). Even more dramatic is the 35 million year intervalthat separates the Late Cretaceous extermination of belemnites andorigin of advanced coleoloids in the North Pacific region, long before

Fig. 1. Reasons for diachroneity (and range offsets) in the local first appearance datum (FAD) and local last appearance datum (LAD) of fossil organisms.
136 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
these events took place globally in the Cretaceous–Paleogene bound-ary interval (Iba et al., 2011).
The dramatic global heterochroneity in the FAD of modern cepha-lopods recalls the documentation by Landing and Westrop (2004) ofthe peritidal origin of benthic communities dominated by major,modern mineralised metazoan groups in the middle Terreneuvian(middle Early Cambrian). This benthic community did not spread off-shore until the late Terreneuvian (‘late Early Cambrian’ see Landing etal., 2007b, for Terreneuvian Series definition).
Strong lithofacies–biofacies linkages often cause the range of a fos-sil within a section to be controlled by vertical changes in deposition-al environments (Fig. 1). This obvious relationship of lithofacies to thefossil record of the Cambrian Radiation has long been known (Brasier,1979; Landing, 1992; Mount and Signor, 1992), but its implicationsfor Cambrian chronostratigraphy deserve much more acknowledg-ment For example, the lowest occurrence of trilobites regarded aslowest Middle Cambrian in South China lies at a change from oxygen-ated to dysoxic shelf facies in western Laurentia (e.g., Webster et al.,2008; Sundberg et al., 2011). This relationship links the Lower–Middle Cambrian boundary interval trilobite turnoverwith transgressionof the west Laurentian shelf by dysoxic/anoxic slope water (Landing,2012). This turnover is less dramatic in Laurentian outer shelf depositswith less dramatic facies shifts (e.g., Blaker and Peel, 1998).
Another example of diachroneity potentially associated with dis-persal time is suggested by Terfelt and Ahlberg's (2010) conclusionthat the FAD of the agnostoid arthropod Pseudagnostus rugosusErgaliev, 1980, is a useful guide to upper Upper Cambrian correlationin Baltica, southern Kazakhstan, and east-central China. However,they note that P. rugosus seems to appear a geologic stage lower insouth-central China.
Use of the local FADs of a fossil for correlation between sectionswithout rigorous supplementary information will lead to errors incorrelation or poorly defined chronostratigraphic units because sig-nificant time intervals likely will separate the local FADs. Each FADmust mirror biological phenomena ranging from evolutionary origi-nation, to dispersal, successful local colonisation, and appearance offacies that allow a species' fossilisation (Fig. 1).
2.3. FAD diachroneity: taxonomy, sampling, unconformities
Problems with the FAD concept may also result from divergent tax-onomic practices, thoroughness of collecting and the presence of un-conformities (Fig. 1). Thus, Westrop et al. (2011) questioned the useof the agnostoid Lotagnostus americanus (Billings, 1860) FAD to definethe base of a terminal Cambrian stage (Peng and Babcock, 2005a) be-cause the seeming “global” distribution of the form results from overly
broad species concepts and synonymisation of related but geographi-cally isolated species. Unreliable species concepts compounded bymisidentification can lead to unstable definitions of chronostratigraphicunits. Thus, the GSSP for the Ordovician System base is defined by theFAD of the conodont Iapetognathus fluctivagus Nicoll et al., 1999, atGreen Point, western Newfoundland. However, restudy of the sectionsuggests a different species of Iapetognathus Landing (in Fortey et al.,1982) occurs at the basal OrdovicianGSSP and that I. fluctivagus appearshigher in the section (Terfelt et al., 2011a,b).
The liability of the FAD concept in chronostratigraphy also resultsfrom problems related to sampling. One example involved a proposedGSSP for the Cambrian base at Meishucun quarry, South China. The sec-tion near the Kunyang Phosphorite Mine near Meishucun had been thereference for the sub-trilobitic, lowest Cambrian of South China, orMeishucun Stage, with its base defined by the lowestmineralisedmeta-zoan fossils in the “China A” interval (e.g., Zhang et al., 1979; Luo et al.,1980, 1982, 1984; Xing et al., 1983; Xing and Luo, 1984; Qian et al.,1985). Destruction by quarrying of much of this section led to examina-tion of a nearby section that extends fromYiaowaitoushan to BadaowanGully. This latter section is now termed the “Meishucun section.” How-ever, no small shelly fossils (SSFs) were recovered from the lowestChina A interval in this section (Qian and Bengtson, 1989), which sug-gested inadequate early sampling and showed that the lowest SSFsappeared at a facies change from white to overlying grey dolostonesin the Baiyanshao Member of the Dengying Formation.
FAD-based correlations and chronostratigraphic units are also con-founded by cryptic or non-cryptic unconformities that cut out thelower ranges of key fossils (Fig. 1). This problem is seen in the lowestCambrian where the lowest diverse, sub-trilobitic SSFs appearabove a long-recognised unconformity at the base of the SiberianTommotian Stage (e.g., Khomentovsky and Karlova, 1986; Knollet al., 1995). This abrupt lowest occurrence of many SSFs has hadlong-lasting consequences for Cambrian correlations and chrono-stratigraphic concepts.
The significance of the Tommotian Stage arose from the exquisitepreservation of its dominantly secondarily phosphatised SSF assem-blage, archaeocyathan succession, long history of research among alllowest Cambrian successions, and proposal of the base of theTommotian stratotype as the global GSSP for the Cambrian's base(Rozanov, 1966, 1967; Rozanov et al., 1969; Rozanov and Sokolov,1984). The length of the pre-Tommotian hiatus was long regardedas brief in the Russian literature (Khomentovsky and Karlova, 1986),and the sudden appearance of diverse skeletalised metazoan fossilswith the lowest archaeocyathans, lingulid brachiopods, a number ofmollusk classes, and hyoliths, among other forms, was portrayed asa sudden evolutionary event in the earliest Tommotian (Rozanov et

137E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
al., 1969; Rozanov and Zhuravlev, 1996). However, the abrupt changefrom underlying restricted-marine dolostone to highly fossiliferousopen-shelf limestone of the Tommotian, a deeply eroded surfaceunder the Tommotian at its stratotype at Dvortsy, and appearanceof over 170 named taxa in the lowest Tommotian all suggested alengthy hiatus at its stratotype (e.g., Landing, 1994, 1995a) and/or afacies unsuitable for fossil remains in the sub-Tommotian dolostones.
The duration of the sub-Tommotian hiatus was briefer in northernSiberia where carbon isotope studies record somewhat lower rockswith Tommotian-aspect faunas, as well as sub-Tommotian rockswith carbon isotope signatures not recorded at and interpreted ashigher than those at Dvortsy (Knoll et al., 1995; Rowland et al.,1998; Kouchinsky et al., 2001, 2005, 2007). The newer evidenceshortens the pre-Tommotian hiatus, but an adequate fossil record ofthe evolutionary precursors of the diverse Tommotian biota is stilllacking in Siberia. The poor sub-Tommotian fossil record likely re-flects a more restricted marine facies and a lower potential for skele-tal fossil preservation in the more dolomitic, less phosphaticsub-Tommotian succession.
Fig. 2. Terminal Ediacaran–lowest Cambrian palaeogeographic maps. (A) Revised palaeogeotent of tropical carbonate platform and evaporite-rich successions (light grey areas), and disditional Early Cambrian palaeogeographic map with major continents at high south latitubiotas of major continents. Abbreviations EA and WA, east and west Avalonia, respectiveand Westrop, 1998a); Baltica (Nielsen and Schovsbo, 2011); Caborca and Chihuahua regio1969; Pillola, 1993); Iran (Hamdi, 1995); Germany (Geyer et al., 2008); Greenland (Cowiand Mambetov, 1981); Mongolia (Brasier et al., 1996); Morocco (Geyer and Landing, 199Zhuravlev, 1996); South China (Steiner et al., 2004, fig. 13); Uruguay and Brazil (Gaucher et atains, California–Nevada (Mount and Signor, 1992).Panel A is highly modified from Landing and Westrop (2004, fig. 1). Selected sources for panUchman, 1995a; Gámez Vintaned, 1996; Li et al., 2009; Gehling et al., 2001); and numerou
2.4. Improved lowest Cambrian correlations
Reliance on one fossil's FAD can derail chronostratigraphic conclu-sions and confuse the geological community. This report emphasisesour conviction that a FAD should be only part of a GSSP definition.Greater stability will result from definition of GSSPs and a LowerCambrian chronostratigraphy within a series of assemblage zonescombined with non-biostratigraphic correlation, in particular carbonisotope chemostratigraphy (Maloof et al., 2010b).
3. Revised terminal Ediacaran–Early Cambrian palaeogeography
Provincialism and onshore–offshore litho- and biofacies linkages thatconfound correlation and development of a stable chronostratigrapythrough the Cambrian Evolutionary Radiation reflect the latitudinal dis-tribution and separation of Cambrian palaeocontinents. Existingpalaeogeographic maps do not resolve the patterns of terminal Ediacar-an–Early Cambrian biotic distributions and lithofacies. Thus, a revisedpalaeogeographic map is proposed (Fig. 2A) that expands the coverage
graphic map shows dominantly tropical distribution of major continents, preserved ex-tribution of basal Cambrian Trichophycus pedum Zone ichnofossil assemblages. (B) Tra-des emphasises mismatches between interpreted palaeolatitudes and lithofacies andly. Summaries of geographic regions: Avalonia (Landing, 1996a, 2004, 2005; Landingns, northern Mexico (Sánchez-Zavala et al., 1999, and sources therein); France (Doré,e, 1971); Iberia (Liñán et al., 1993, 2006); Maly Karatau, Kazakhstan (Missarzhevsky5, 2006); Oman (Schröder et al., 2005); Siberia (Brasier et al., 1994b; Rozanov andl., 2003, 2007); Vermont (Landing et al., 2007b; Landing, 2012); andWhite-Inyo Moun-
el A: Trichophycus pedum Assemblage Zone localities (Narbonne et al., 1987; Geyer ands localities in South China (Weber et al., 2007) shown by two arrows.

138 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
of Landing andWestrop's (2004, fig. 1) map and incorporates biotic andlithostratigraphic information from Landing (2005) and Zhuravlev et al.(2012).
Many terminal Ediacaran–Lower Cambrian palaeogeographicmaps are “Antarctocentric” and show the South American–West Afri-can margin of Gondwana at high latitudes or at the South Pole. Simi-larly, Avalonia, the Early Palaeozoic terrane in the later Acadian–Caledonian orogen, often is shown as an appendage of or as severalfragments near West Gondwana (Fig. 2B). The tradition of a highsouth latitude-polar West Gondwana with an Avalonian appendagebegan with maps by Scotese et al. (1979) and Smith et al. (1981)who cited little data for their Cambrian–Early Ordovician reconstruc-tions. The Early Cambrian palaeogeographic reconstruction by(McKerrow et al., 1992) is similar and based largely on biotic andinterpreted palaeoclimatic data and available palaeomagnetic infor-mation (e.g., Smith, 2001, p. 12).
Repeated use of an “Antarctocentric” and “Avalonian appendage”Cambrian world map has given it the status of a palaeogeographicparadigm (Keppie et al., 1996; Torsvik et al., 1996; Dalziel, 1997;Keppie and Ramos, 1999; Brasier and Lindsay, 2001; Smith, 2001;Álvaro et al., 2003; Linnemann et al., 2008; Zhuravlev et al., 2012).But for Piper (1987), who interpreted palaeomagnetic data to recon-struct an Arctic West Gondwana, most Early Palaeozoic world mapsare “Antarctocentric.” Dalziel (1997; Fig. 1) and Steiner et al. (2007)seemingly followed Barr and Raeside (1989) in regarding Early Cam-brian Avalonia as a number of isolated fragments. Steiner et al.'s(2007, fig. 13) reconstruction shows “east” and “west” Avalonia asseparate “bumps” on opposite (west South American and west Afri-can) margins of Gondwana.
The inconsistencies between Cambrian palaeogeographic maps andthe distribution of climatically-controlled lithofacies and biotic prov-inces have arisen, in part, because of the often low quality ofpalaeomagnetic data (95% confidence limits commonly >20° and occa-sionally >40°) and “acceptable” age uncertainties of about 10% on indi-vidual poles (Smith, 2001). Thus, palaeomagnetic data may beinadequate to recognise misfits between calculated palaeolatitude andapparent polar wander paths and a palaeocontinent's actual lithofaciesand biotas.
Smith (2001) concern that Early Cambrian archaeocyathan reefsand carbonate platform deposition might belie a 70° S to South Poleposition of southern Morocco (e.g., Dalziel, 1997; Keppie andRamos, 1999) might be regarded as less significant than other unifor-mitarian arguments. Modern ahermatypic coral reefs occur, for exam-ple, in deep, cold water at high latitudes, and the temperature andlatitudinal limits of archaeocyathan reefs should not be presumed asalways tropical without other evidence. More importantly, a locally2.0 km-thick terminal Ediacaran–Early Cambrian carbonate platformwith lower evaporate minerals and higher oosparites is wholly con-sistent with an equatorial–tropical location of southern Moroccoand West Gondwana (Geyer and Landing, 1995, 2006; Landing,1996a; Landing and Westrop, 2004, fig. 1). A high latitude positionfor Early Cambrian West Africa is also precluded by the lack of indica-tors for floating or grounded ice (Fig. 2). Indeed, it is Avalonia thatshows evidence of floating ice consistent with a high latitude positionand significant separation from tropical West Gondwana in the earli-est Cambrian (Landing and MacGabhann, 2010).
The McKerrow et al. (1992) interpretation of Avalonia as an ap-pendage of Gondwana has been used in many reports (e.g., Murphyet al., 1999; Fletcher, 2003; Parkhaev and Karlova, 2011; Zhuravlevet al., 2012). However, the original interpretation was largely basedon the same type of data later used by Álvaro et al. (2003) to groupAvalonian and West Gondwanan trilobites by a Phi–Pearson similari-ty analysis. McKerrow et al. (1992) and Álvaro et al. (2003) comparedquite obviously similar Lower–Middle Cambrian boundary intervaltrilobite faunas. However, this faunal similarity did not apply to theca. 20 Ma of the earlier Cambrian (geochronology of Landing et al.,
1998). Older Cambrian faunas and lithofacies of the two regions aredissimilar—abundant archaeocyathans appear with fallotaspid andbigotinid trilobites in Moroccan carbonates, while archaeocyathansare absent and olenellids are the oldest trilobites in the coevalsiliciclastic-dominated successions of Avalonia (McKerrow et al.,1992; Landing, 1996a, 2005).
Provincial barriers between Avalonia and Morocco started tobreak down later in the Early–Middle Cambrian boundary intervalwhen trilobite genera are first shared—likely reflecting movement ofthe Moroccan margin of West Gondwana into the south temperatelatitude of Avalonia (Theokritoff, 1979; Burrett et al., 1991; Landingand Westrop, 2004) (Fig. 2). Southern Morocco also underwent arapid change from an earlier carbonate platform to an overlyingsiliciclastic-dominated, more “Avalonian-type” lithofacies in thistime interval (Geyer and Landing, 1995, 2006; Landing, 1996a).
Thus, a longer term biotic and lithofacies history contradictsÁlvaro et al.'s (2003) interpretation of Avalonia linked to the Moroc-can margin of West Gondwana in the Early Cambrian. Similarly, theiruse of an uppermost Lower Cambrian limestone in the Comley area ofAvalonian England (Fig. 3) as a way to buttress this “Avalonian ap-pendage” interpretation is irrelevant. The Comley limestones do notform a tropical carbonate platform succession comparable in anyway to that of southern Morocco. They comprise only about 2.0 mof successive limestone beds, each of which has a distinct trilobitefauna (e.g., Cobbold, 1921, 1931; Lake, 1936; Rushton, 1974). Thelimestones are best regarded as a very condensed succession of fossiltempestites that mark transgressive highstands in a cool-water faciesthat never developed the evaporites, massive carbonates, oosparites,and other features of tropical carbonate platforms (e.g., Landing,1988, 1992, 1996a; Myrow and Landing, 1992; Landing andWestrop, 1998a,b, 2004; Harvey et al., 2011).
In short, biotic and lithofacies differences are consistent with iso-lation of Avalonia from the West African margin of Gondwana duringthe latest Ediacaran, with biotic and lithofacies similarities developingin the Lower–Middle Cambrian boundary interval (Landing, 1996a,2005). This conclusion contrasts with the traditional interpretation,primarily based on the presence of shared trilobite taxa that the sep-aration only took place in the late Early Ordovician (Floian) (e.g.,Fortey and Cocks, 2003). The alternative interpretation is that a longterm (late Early Cambrian–Middle Ordovician) faunal similarityreflected the transport of Avalonia and West Gondwana, possiblyalong transform faults and their proximity in temperate latitudesuntil the Late Ordovician. At that latter time, West Gondwanamoved across the South Pole and Avalonia moved north into the tro-pics (Landing, 1996a, 2005; Kim et al., 2002; Landing et al., 2003;Landing and Westrop, 2004).
Similarity in Nd isotopes and detrital zircon ages has been used torelate Avalonia to the Amazonia margin of Gondwana. Thus, Keppieand Ramos (1999, fig. 5) figured isolated fragments of a non-unifiedAvalonia outboard of a West African–South American margin ofGondwana implausibly located at the South Pole in the Precambrian–Cambrian boundary interval (Fig. 2B). While Avalonia may have origi-natedwith Ediacaran rifting along the Amazoniamargin, there is no ev-idence to link Avalonia to South America or to allow it to be termed“peri-Gondwana” (e.g., Murphy et al., 1999) after the late Ediacaran.The late Ediacaran carbonate platform successions in the adjacentGondwanan areas of Uruguay and Brazil (Fig. 2A, sources in caption)provide evidence that the Amazonian region was likely tropical andlatitudinally distant from the coeval, cool-water siliciclastic successionsof Avalonia (Fig. 3).
A unified Avalonia palaeocontinent is indicated by the regionaluniformity of a terminal Ediacaran–Ordovician cover sequence thatnon-conformably overlies a late Cryogenian–middle Ediacaran arcvolcanic and volcaniclastic basement (Landing, 1996a, 2004, 2005;Landing et al., 2013a,b; McIlroy et al., 1998). This cover sequence ex-tends from Rhode Island through eastern Newfoundland to southern

Fig. 3. Lengthy bio- and lithostratigraphic range (650 m) ofWatsonella crosbyi and Aldanella attleborensis in Avalonian southeastern Newfoundland. Diagram shows uppermostEdiacaran–Lower Cambrian depositional sequences on marginal and inner platforms of the Avalonia palaeocontinent from North Wales to eastern Massachusetts.After Landing (1996a, 2004, fig. 4) with addition of volcanic Wades Lane Formation (Landing et al., 2008). Massachusetts and southeast Newfndland stratigraphy of Landing andBenus (1988), Landing (1992, 1996a), and Landing and Westrop (1998b) and southern New Brunswick of Landing and Westrop (1996), Landing (1996b, 2004), and Landing etal. (2008). Fletcher's (2003, 2006) lithostratigraphic terms in Newfoundland are junior synonyms of an established nomenclature and often rely on non-lithologic features (e.g.,“separation planes”) in their definition (see Westrop and Landing, 2011). Welsh stratigraphy (with asterisks) and English units (in italics) discussed in Landing et al. (2013b).
139E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Britain and underlies the Brabant region of Belgium and adjacentanticlinoria of westernmost Germany. Avalonia's regionally extensivedepositional sequence boundaries together with such distinctive de-velopments as a red limestone unit unconformably overlain by thelowest trilobite-bearing rocks from Massachusetts to England(Fig. 3) and a regional change to persistently dysoxic black mudstonedeposition in the middle Middle Cambrian (Landing, 1996a) showthat it is not credible to regard Avalonia as a collage of unrelated ter-ranes in the Early Palaeozoic (e.g., Steiner et al., 2007) is not credible.
The terminal Ediacaran–Early Cambrian map proposed herein(Fig. 2A) follows McKerrow et al. (1992) in emphasising a uniformi-tarian interpretation of lithofacies and biotic data. Thus, the lack ofevidence of grounded ice, permafrost, or widespread dropstone ordiamictite deposits argues against a large, high latitude or polarcontinent or a polar sea isolated by land masses in the terminalEdiacaran–Middle Ordovician (Landing, 2012). Similarly, the locallyevaporitic, carbonate platform successions in Uruguay and Braziland from Namibia to southern Morocco (Fig. 2A, sources in caption)show that these West Gondwanan regions likely were in low lati-tudes (Landing, 1996a; Landing and Westrop, 2004, fig. 1).
Similarities in Early Cambrian archaeocyathans, trilobites, andlithofacies show a likely proximity between the Moroccan margin ofWest Gondwana and Cadomian Europe. A progressive northern shift(in terms of modern latitude) of carbonate platform development inthis belt arguably reflects the southern movement of the Moroccan–
Cadomian margin (Fig. 2A). Thus, the Atlas region, southern Morocco,has a terminal Ediacaran–Lower Cambrian carbonate platform thatwas replaced by a mixed carbonate-siliciclastic shelf shortly afterthe earliest appearance of trilobites in the early Atlasian Epoch(Fig. 4, middle Issendalenian) (Geyer and Landing, 1995, 2006). Con-versely, the oldest carbonate platform (Pedroche Formation) ofsouthern Iberia developed only later in the early Atlasian (earlyIssendalenian) with appearance of these oldest West Gondwanan tri-lobites (e.g., Brasier et al., 1980; Alvaro et al., 2008; Elicki and Geyer,2010).
Carbonate platform deposition began even later with this progres-sive southern movement in the more northerly areas of CadomianEurope. A thin carbonate platform developed in the MontagneNoire, southern France, in the late Atlasian (early Banian Age)and persisted into the Lower–Middle Cambrian boundary interval(early Agdzian Epoch) before it was replaced by likely cool-watersiliciclastic shelf deposits (Álvaro et al., 2002). A similar “late” endto carbonate platform deposition in the late Early Cambrian occurredin the Saxo-Thuringian sector of the Variscan Foldbelt in Germany,and was followed by rapid development of Moroccan-type, latestEarly Cambrian siliciclastic deposits and shelly fossil faunas with pro-gressively more Baltic aspect in the siliciclastic-dominated MiddleCambrian sequence (Geyer et al., 2008; Elicki and Geyer, 2010).
The revised palaeography (Fig. 2A) is supported by similar termi-nal Ediacaran–Early Cambrian mixed siliciclastic–carbonate platform
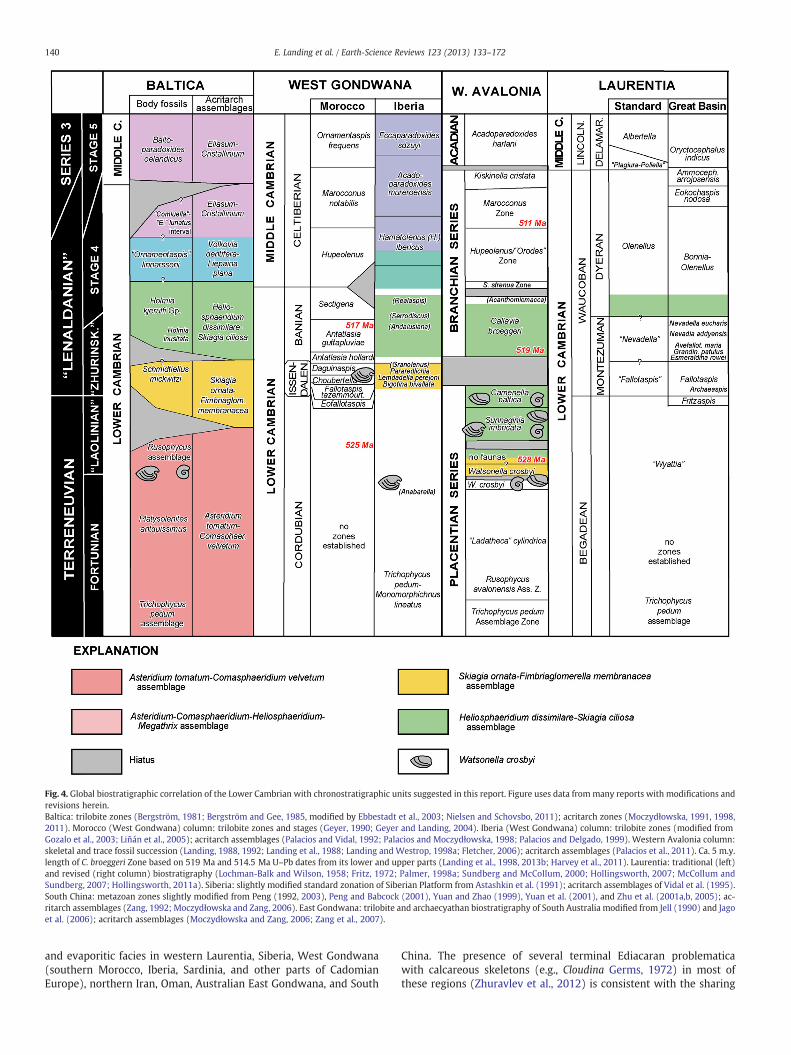
Fig. 4. Global biostratigraphic correlation of the Lower Cambrian with chronostratigraphic units suggested in this report. Figure uses data frommany reports with modifications andrevisions herein.Baltica: trilobite zones (Bergström, 1981; Bergström and Gee, 1985, modified by Ebbestadt et al., 2003; Nielsen and Schovsbo, 2011); acritarch zones (Moczydłowska, 1991, 1998,2011). Morocco (West Gondwana) column: trilobite zones and stages (Geyer, 1990; Geyer and Landing, 2004). Iberia (West Gondwana) column: trilobite zones (modified fromGozalo et al., 2003; Liñán et al., 2005); acritarch assemblages (Palacios and Vidal, 1992; Palacios and Moczydłowska, 1998; Palacios and Delgado, 1999). Western Avalonia column:skeletal and trace fossil succession (Landing, 1988, 1992; Landing et al., 1988; Landing and Westrop, 1998a; Fletcher, 2006); acritarch assemblages (Palacios et al., 2011). Ca. 5 m.y.length of C. broeggeri Zone based on 519 Ma and 514.5 Ma U–Pb dates from its lower and upper parts (Landing et al., 1998, 2013b; Harvey et al., 2011). Laurentia: traditional (left)and revised (right column) biostratigraphy (Lochman-Balk and Wilson, 1958; Fritz, 1972; Palmer, 1998a; Sundberg and McCollum, 2000; Hollingsworth, 2007; McCollum andSundberg, 2007; Hollingsworth, 2011a). Siberia: slightly modified standard zonation of Siberian Platform from Astashkin et al. (1991); acritarch assemblages of Vidal et al. (1995).South China: metazoan zones slightly modified from Peng (1992, 2003), Peng and Babcock (2001), Yuan and Zhao (1999), Yuan et al. (2001), and Zhu et al. (2001a,b, 2005); ac-ritarch assemblages (Zang, 1992; Moczydłowska and Zang, 2006). East Gondwana: trilobite and archaecyathan biostratigraphy of South Australia modified from Jell (1990) and Jagoet al. (2006); acritarch assemblages (Moczydłowska and Zang, 2006; Zang et al., 2007).
140 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
and evaporitic facies in western Laurentia, Siberia, West Gondwana(southern Morocco, Iberia, Sardinia, and other parts of CadomianEurope), northern Iran, Oman, Australian East Gondwana, and South
China. The presence of several terminal Ediacaran problematicawith calcareous skeletons (e.g., Cloudina Germs, 1972) in most ofthese regions (Zhuravlev et al., 2012) is consistent with the sharing
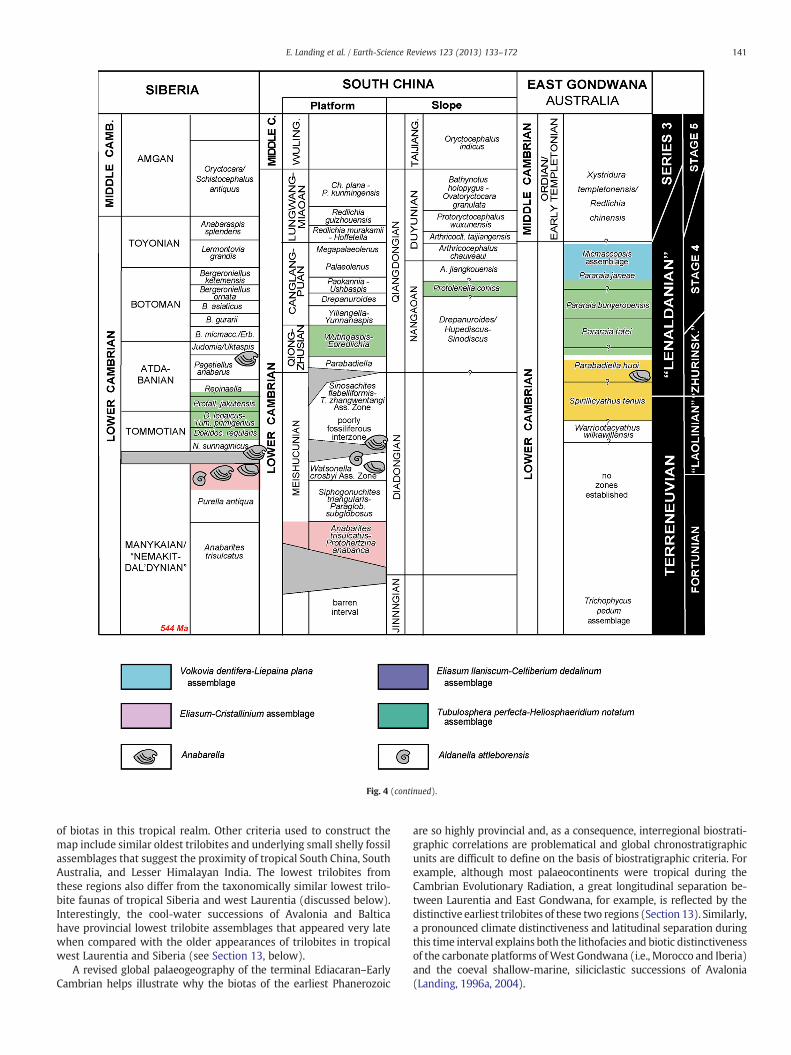
Fig. 4 (continued).
141E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
of biotas in this tropical realm. Other criteria used to construct themap include similar oldest trilobites and underlying small shelly fossilassemblages that suggest the proximity of tropical South China, SouthAustralia, and Lesser Himalayan India. The lowest trilobites fromthese regions also differ from the taxonomically similar lowest trilo-bite faunas of tropical Siberia and west Laurentia (discussed below).Interestingly, the cool-water successions of Avalonia and Balticahave provincial lowest trilobite assemblages that appeared very latewhen compared with the older appearances of trilobites in tropicalwest Laurentia and Siberia (see Section 13, below).
A revised global palaeogeography of the terminal Ediacaran–EarlyCambrian helps illustrate why the biotas of the earliest Phanerozoic
are so highly provincial and, as a consequence, interregional biostrati-graphic correlations are problematical and global chronostratigraphicunits are difficult to define on the basis of biostratigraphic criteria. Forexample, although most palaeocontinents were tropical during theCambrian Evolutionary Radiation, a great longitudinal separation be-tween Laurentia and East Gondwana, for example, is reflected by thedistinctive earliest trilobites of these two regions (Section 13). Similarly,a pronounced climate distinctiveness and latitudinal separation duringthis time interval explains both the lithofacies and biotic distinctivenessof the carbonate platforms ofWest Gondwana (i.e., Morocco and Iberia)and the coeval shallow-marine, siliciclastic successions of Avalonia(Landing, 1996a, 2004).

142 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
4. Chronostratigraphy and Stage 1 of the Cambrian Radiation
4.1. Historical development of chronostratigraphy
A relatively consistent vertical succession of increasingly complex,marine trace fossils was recognised in Precambrian–Cambrian bound-ary interval successions on a number of continents by the 1950s. As a re-sult, the lowest occurrences of particular ichnotaxa were suggested asways to define the base of the Cambrian (e.g., Seilacher, 1956; Albert,1977). These proposals received increased support as the earliest Cam-brian small shelly fossil assemblages of Siberia and South China provedto be provincial and often showed diachronous lowest occurrences thatcould mislead correlations (Bengtson, 1988; Brasier, 1989).
Proterozoic–Cambrian boundary sequences in the Burin Peninsula,southeastern Newfoundland (Figs. 2, 3), were found to show a verticalsuccession of ichnofaunas. The upper assemblages, with physicallyand behaviorally complex ichnofossils (vertical burrows, complex feed-ing burrows, graphoglyphids, arthropod traces), were regarded asmorecharacteristic of the coelomate burrowers of the Early Cambrian andPhanerozoic. Underlying strata had simpler, uppermost Precambrian-aspect assemblages (shallow burrows and surface trails) (Bengtsonand Fletcher, 1983; Crimes and Anderson, 1985).
The base of an ichnofossil assemblage with Phycodes pedum Seilacher,1955, later referred to Trichophycus or Treptichnus (e.g., Geyer, 1993;Geyer and Uchman, 1995; Jensen et al., 2000), was proposed as a GSSPfor the Cambrian base (Narbonne et al., 1987) (Fig. 3). The proposedGSSP at Fortune Head in the Burin Peninsula is low in the siliciclasticChapel Island Formation on the cool-water Avalon palaeocontinent(Myrow and Landing, 1992; Landing, 1996a) (Figs. 2, 3). This proposalwas accepted by the International Stratigraphic Commission in 1992 asthe standard for the base of the Cambrian System (Brasier et al., 1994a;Landing, 1994) (Fig. 4). Subsequently, this horizon was also designatedthe base of the lowest Cambrian Terreneuvian Series and FortunianStage (Landing et al., 2007a; Figs. 3, 4). Thus, the concept of the FortunianStage corresponds to the initial stage of the Cambrian Evolutionary Radi-ation and the diversification of coelomate burrowers that constructedmorphologically and behaviorally complex burrows (Landing et al.,1989; Landing and Westrop, 2004). This new infaunal community dra-matically changed sea floor biogeochemistry (Brasier et al., 2011).
4.2. Trichophycus pedum Zone and basal Cambrian correlation
Ratification of the Fortune Head GSSP placed the Cambrian base andT. pedum FAD well below the lowest occurrences of trilobites and otherskeletalisedmetazoans in southeast Newfoundland and a short distanceabove typical, terminal Proterozoic trace fossils. The basal CambrianGSSP is also above the highest occurrence of the problematical fossilsHarlaniella podolica Sokolov, 1972, and Palaeopascichnus delicatusPaliy, 1976. These geographically widespread, characteristic Ediacaranfossils are variously interpreted as trace fossils (Narbonne et al., 1987)or possible body fossils (Jensen, 2003). Palaeopascichnus is a puzzlingfossil interpreted as a giant xenophyophore foraminiferan (Shen et al.,2007) or as a “nonxenophyophore” agglutinated protozoan, becausexenophyophores are likely recently evolved foraminiferans (Antcliffeet al., 2011).
In addition to Avalonia, lowest Cambrian T. pedum Zone assem-blages occur in Baltica, East and West Gondwana, and west Laurentia(Narbonne et al., 1987; Gámez Vintaned, 1996) (Fig. 2A). Later workin Namibia showed that lowest Cambrian T. pedum Zone assemblagesoccur above the highest Ediacaran vendobionts and well above thecalcareous problematica Cloudina Germs and NamacalathusGrotzinger et al., 2000 (Geyer and Uchman, 1995; Grotzinger et al.,2000; Geyer, 2005a). However, a few Phanerozoic-aspect trace fossilsoccur below T. pedum and probably with the youngest Ediacaranvendobionts (Geyer, 2005a; Wilson et al., 2012).
Carbonate-dominated successions, as in Namibia, typically have limit-ed ichnofossils. However, lowdiversity assemblageswith T. pedum appearin carbonates with lowest Cambrian shelly fossils in the “Lower Phos-phate Bed” on the South China (Yangtze) Platform (Weber et al., 2007).As in southeastNewfoundland, the ichnogenusRusophycusHall, 1852, ap-pears above the T. pedum FAD in Baltica, Iberia, and the “Upper PhosphateBed” in South China (Fig. 2A). The successive appearances of lowdiversityplanolitid and helminthoidichnid traces in the terminal Precambrian,followed by lowest Cambrian-aspect trichophycids, and then cruzianidichnofossils record not only increased morphological and behaviouralcomplexity of metazoans, but also substrate community evolution withthe increased disparity in infaunal and epifaunal trace-producers. Signifi-cantly, Zhuravlev et al. (2012) have begun to develop an uppermost Edi-acaran biostratigraphy in carbonate platform successions of South Chinaand Iberia where remains of low diversity skeletalised conoidalproblematica and increasingly complex burrows occur below lowestCambrian assemblages with T. pedum.
4.3. Eurytopic T. pedum organism
Besides its wide palaeogeographic distribution (Fig. 2A), T. pedum isa useful trace fossil on which to base a GSSP because it represents a eu-rytopic pattern of behavior. Its range extends from desiccation-crackedmud flat to dysoxic subtidal facies (Geyer andUchman, 1995, table 1). T.pedum is not limited to the Cruziana ichnofacies as suggested bySeilacher (1970), and its broad lithofacies occurrence (Buatois et al.,2013) makes it one of the least habitat-specific Cambrian taxa. The util-ity of T. pedum in defining the base of the Cambrian or bracketing thePrecambrian–Cambrian boundary was emphasised by Gehling et al.(2001, p. 214, 215), who updated the list of regions where theichnospecies is found (Fig. 2A).
4.4. Problems with the T. pedum FAD?
A criticism about the use of the base of the T. pedum Zone to definethe base of the Cambrian boundary results from the current definitionof the T. pedum Zone—the zone is primarily defined by the FAD of itseponymous species (e.g., Jensen et al., 2000; Seilacher et al., 2005).The problem here is that traces resembling T. pedum also are foundwith latest Ediacaran-type organisms in Namibia (Geyer andUchman, 1995; Jensen et al., 2000) and South Australia (Gehlinget al., 2001). Thus, these T. pedum-type remains would have to beregarded as Ediacaran in age. What is now emerging is the recogni-tion of a gradual transition in ichnofossil communities across theEdiacaran–Cambrian boundary interval, notably in the thicker succes-sions of Namibia (Crimes and Germs, 1985; Geyer and Uchman, 1995;Geyer, 2005a), Spain (Gámez Vintaned, 1996), and Nevada (Jensenet al., 2000). In these successions, Conichnus Myannil, 1966 (possiblesea-anemone burrows), and Skolithos Haldemann, 1840 (verticaltubes), which Narbonne et al. (1987) considered more characteristicof the Cambrian, first appear below the lowest T. pedum datum and,thus, are of terminal Proterozoic age.
More problematical for a basal Cambrian GSSP defined primarilyupon the FAD of T. pedum was the recovery of T. pedum below thebasal Cambrian GSSP at Fortune Head—with Gehling et al. (2001)reporting T. pedum 3.11 m and 4.41 m below the GSSP with the char-acteristic terminal Ediacaran form H. podolica. The comments of Pengand Babcock (2011) on the presence of T. pedum below the basalCambrian GSSP are arguably less measured than those of Gehling etal. (2001). Peng and Babcock (2011) declared that the new range ofT. pedum at Fortune Head “casts doubt on the international correla-tion utility” of the GSSP and argued that no method can correlatethe GSSP beyond its site of definition.
These“problems”with a basal Cambrian GSSP primarily defined bythe FAD of T. pedum are far from intractable and easily resolved. Tobegin with, the trichophycids from the uppermost Ediacaran of

143E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Namibia, Spain, and Nevada are not conspecific with T. pedum. Theyrepresent, as might be expected, antecedent patterns of behaviorthat appear in a transitional interval and form part of a stepwisepattern of emergence of Phanerozoic-aspect behaviours in theEdiacaran–Cambrian boundary interval (Zhuravlev et al., 2012).
As the FAD of any fossil likely underestimates its true lowest oc-currence in any section, it can be suggested that Peng and Babcock(2011) placed too much reliance on the T. pedum FAD for definingthe base of the T. pedum Zone and the base of the Cambrian. In the ab-sence of unconformities or abrupt facies changes, it is expected thatany given FAD will be lowered with additional study. Thus, it is notsurprising that the T. pedum FAD was later discovered lower in theFortune Head section than Narbonne et al.'s (1987) original report.A 95% confidence interval calculation (Marshall, 1990), based on thestratigraphic range and number of horizons recorded for T. pedumby Narbonne et al. (1987), indicates that it could appear as much as40 m below the GSSP. The implication is that the GSSP of any chrono-stratigraphic unit that is based primarily upon the FAD of a single or-ganism, like that of T. pedum, is necessarily flawed. The challenge tothe basal Cambrian GSSP related to T. pedum (Peng and Babcock,2011) will apply to all FAD-based candidates in this stratigraphic in-terval. This challenge can be overcome if multiple criteria are usedto define and correlate the base of the Cambrian as argued byBrasier et al. (1990, 1992, 1996). For example, multiple methodscan be used to correlate the basal Cambrian GSSP beyond the FortuneHead stratotype. Perhaps even more heartening for the definition of aGSSP associated with T. pedum is the fact that the lowest occurrencerecorded for T. pedum by Narbonne et al. (1987) closely correspondsto Gehling et al.'s (2001) FAD for the ichnospecies in the shallow ma-rine succession at the Fortune Head succession. This near correspon-dence in the FADs reported by Narbonne et al. (1987) and Gehlinget al. (2001) reflects the fact that T. pedum is essentially eurytopic inshallow marine facies (see Section 4.3; Buatois et al., 2013). Inshort, the known lowest occurrence of T. pedum at Fortune Head like-ly approximates this burrower's evolutionary origin and relatively lit-tle range offset is likely present at Fortune Head.
5. Improved definition for the base of the Cambrian
5.1. Carbon isotope stratigraphy and geochronology
The utility of a basal Cambrian GSSP that features T. pedum isstrengthened by non-biostratigraphic correlation techniques. Astrong negative δ13C excursion occurs just below or at the lowest oc-currence of T. pedum in mixed siliciclastic–carbonate successions ofLaurentia (Northwest Territories, Canada; Death Valley, California)and northern Siberia. This negative excursion also occurs in non-fossiliferous successions in West Gondwana (Morocco and the sub-surface of Oman; see Li et al., 2009; Maloof et al., 2010a, p. 1736).This excursion is likely the “feature W” excursion recognised inMongolia by Brasier et al. (1996).
This δ13C nadir has been regarded as a proxy for the global Ediacaran–Cambrian boundary (e.g., Brasier et al., 1992, 1996; Strauss et al., 1992;Grotzinger et al., 1995) as it lies very high in the Ediacaran and some-what below biostratigraphically defined bases of the Cambrian. As anexample, at the Laolin section in Yunnan (Li et al., 2009), the lowestT. pedum specimen is reported variously as several to 15 m above thelowest small shelly fossils (Anabarites trisulcatus–Protohertzina anabaricaAssemblage Zone of Steiner et al., 2007) in the lowest ZhongyicunMember (Fig. 5). The phosphatic dolostones of the Zhongyicun are apoor medium for trace fossil preservation, and the biostratigraphic reso-lution provided by the lowest T. pedum is limited. In the absence of asatisfactory trace fossil succession, the SSFs at the base of the ZhongyicunMember lie within the Ediacaran–Cambrian boundary interval (e.g.,Zhuravlev et al., 2012). Use of the lowest specimens of T. pedum as a prac-tical lowest Cambrian indicator at Laolin and across the South China
Platform (e.g., Weber et al., 2007) is strengthened by the underlyingnegative δ13C excursion at Laolin which records global changes in theEdiacaran–Cambrian boundary interval (Li et al., 2009) and representsthe basal Cambrian excursion (BACE) of Zhu et al. (2004, 2006, 2008).
Rock successions with the strong terminal Ediacaran negativeδ13C values provide a practical bracket for the basal Cambrianboundary even in coeval successions without biostratigraphicallyuseful fossils in Morocco (Maloof et al., 2005) and Oman (Brasieret al., 2000; Grotzinger et al., 2000; Amthor et al., 2003). A secondnon-biostratigraphic correlation technique, IDTIMS dating of volca-nic zircons, provides statistically indistinguishable U–Pb dates ontwo ashes that closely under- and overlie the carbon isotopeexcursion in the unfossiliferous Oman succession (544 ± 3.3 Ma ofBrasier et al, 2000; 542.0 ± 0.4 Ma of Grotzinger et al., 2000;541.0 ± 0.2 Ma of Bowring et al., 2007) and on an ash just belowthe T. pedum FAD in Namibia (540.61 ± 0.88 Ma; recalculated byBowring et al., 2007, from Grotzinger et al., 1995).
An age on the Namibian successions comes from a 207Pb–206Pbvolcanic ash date in the upper Spitskop Member of the Urusis Forma-tion just below the lowest Trichophycus sp. Grotzinger et al. (1995)report 543 ± 1 Ma on this horizon, and an overlying ash in thelower Nomtsas Formation was dated at 539.4 ± 1 Ma. Grotzinger etal. (1995) placed the Proterozoic–Cambrian boundary at the base ofthe Nomtsas Formation. These dates are comparable to a U–Pb dateon a volcanic breccia from Cambrotubulus-bearing, earliest Cambrian(Manykaian) strata of the Olenek Uplift, northern Siberia, althoughthis horizon is also not very well constrained biostratigraphically(Bowring et al., 1993).
Carbon isotopic and U–Pb zircon data from the Ara Group of Omanbracket the highest appearance of the mineralised Ediacaran formsCloudina and Namacalathus. This highest occurrence appears to coin-cide with a strong negative carbon isotope excursion dated by U–Pbzircon methods at 542.0 ± 0.4 Ma (Grotzinger et al., 2000; Amthoret al., 2003). Since the Oman sections lack Cambrian fossils, the useby Gradstein et al. (2005) of this date as the best age for the Protero-zoic–Phanerozoic boundary is questionable. This conclusion assumes(1) that the highest occurrences of Cloudina and Namacalathus in theAra Group record the local extinction of these and other Ediacaran or-ganisms; (2) that the Oman extinctions are coeval with the disap-pearance of Cloudina and Namacalathus in Namibia; and, apparently,(3) that the higher, major positive carbon isotope shift in Oman coin-cides with the Siberian Lower Cambrian I′ excursion (discussedbelow).
5.2. Biotic successions
Many biostratigraphically distinctive features are related to a basalCambrian GSSP that is associated with the lower range of T. pedum,but not necessarily located at its local FAD. As discussed above, agradual increase in ichnofossil diversity and complexity, includingthe appearance of branched trichophycids, took place throughthe terminal Ediacaran–lowest Cambrian along with disappearanceof the characteristic terminal Proterozoic problematica H. podolicaand P. delicatus in cool-water successions in eastern Newfoundland(Narbonne et al., 1987) and Poland (Pacześna, 1985). Comparable faunaldisappearances took place on warm-water shelves of the terminalProterozoic with the disappearance of Cloudina and Namacalathus belowthe lowest occurrence of T. pedum in Namibia (Grotzinger et al., 2000).
The latest Ediacaran is also marked by the replacement of astratigraphically long-ranged, leiosphaeridid-dominated phytoplank-ton community by the more diverse acanthomorph acritarchs of theAsteridium tornatum–Comasphaeridium velvetum Zone assemblage(Figs. 3, 4). This diverse acritarch assemblage appears just below thelowest occurrence of T. pedum and just above the highest occurrencesof H. podolica and P. delicatus in Poland (Moczydłowska, 1991, 2011).
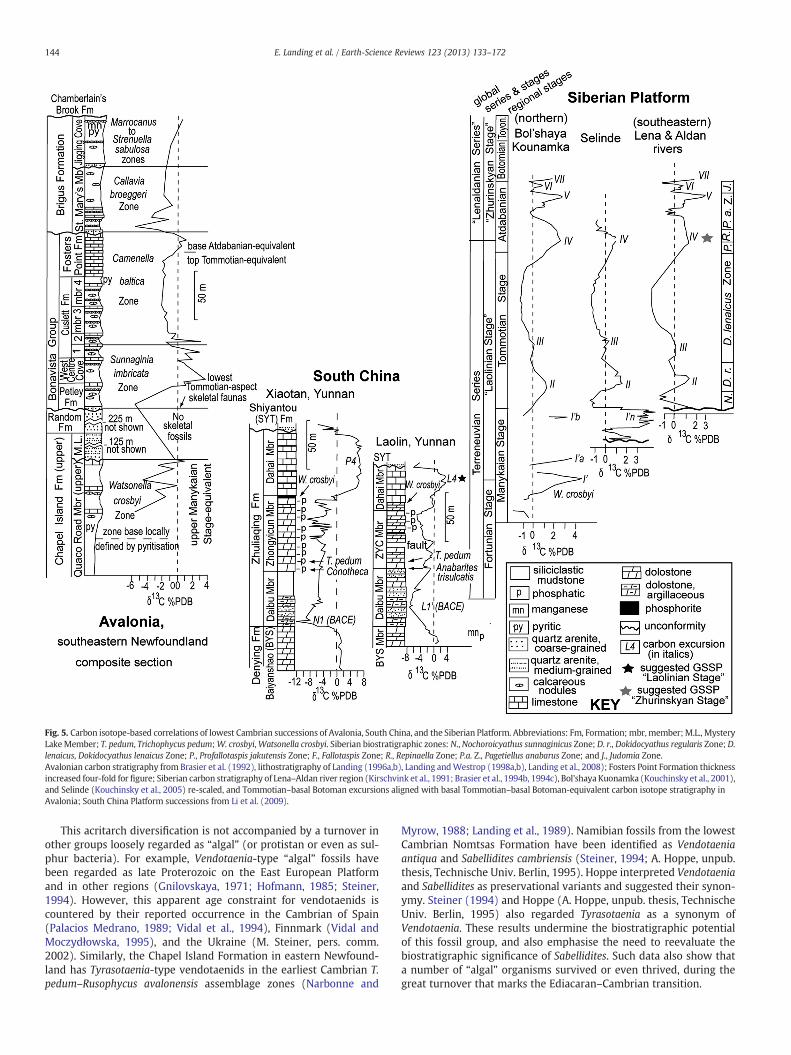
Fig. 5. Carbon isotope-based correlations of lowest Cambrian successions of Avalonia, South China, and the Siberian Platform. Abbreviations: Fm, Formation; mbr,member;M.L., MysteryLakeMember; T. pedum, Trichophycus pedum;W. crosbyi,Watsonella crosbyi. Siberian biostratigraphic zones: N.,Nochoroicyathus sunnaginicus Zone; D. r., Dokidocyathus regularis Zone; D.lenaicus, Dokidocyathus lenaicus Zone; P., Profallotaspis jakutensis Zone; F., Fallotaspis Zone; R., Repinaella Zone; P.a. Z., Pagetiellus anabarus Zone; and J., Judomia Zone.Avalonian carbon stratigraphy from Brasier et al. (1992), lithostratigraphy of Landing (1996a,b), Landing andWestrop (1998a,b), Landing et al., 2008); Fosters Point Formation thicknessincreased four-fold for figure; Siberian carbon stratigraphy of Lena–Aldan river region (Kirschvink et al., 1991; Brasier et al., 1994b, 1994c), Bol'shaya Kuonamka (Kouchinsky et al., 2001),and Selinde (Kouchinsky et al., 2005) re-scaled, and Tommotian–basal Botoman excursions aligned with basal Tommotian–basal Botoman-equivalent carbon isotope stratigraphy inAvalonia; South China Platform successions from Li et al. (2009).
144 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
This acritarch diversification is not accompanied by a turnover inother groups loosely regarded as “algal” (or protistan or even as sul-phur bacteria). For example, Vendotaenia-type “algal” fossils havebeen regarded as late Proterozoic on the East European Platformand in other regions (Gnilovskaya, 1971; Hofmann, 1985; Steiner,1994). However, this apparent age constraint for vendotaenids iscountered by their reported occurrence in the Cambrian of Spain(Palacios Medrano, 1989; Vidal et al., 1994), Finnmark (Vidal andMoczydłowska, 1995), and the Ukraine (M. Steiner, pers. comm.2002). Similarly, the Chapel Island Formation in eastern Newfound-land has Tyrasotaenia-type vendotaenids in the earliest Cambrian T.pedum–Rusophycus avalonensis assemblage zones (Narbonne and
Myrow, 1988; Landing et al., 1989). Namibian fossils from the lowestCambrian Nomtsas Formation have been identified as Vendotaeniaantiqua and Sabellidites cambriensis (Steiner, 1994; A. Hoppe, unpub.thesis, Technische Univ. Berlin, 1995). Hoppe interpreted Vendotaeniaand Sabellidites as preservational variants and suggested their synon-ymy. Steiner (1994) and Hoppe (A. Hoppe, unpub. thesis, TechnischeUniv. Berlin, 1995) also regarded Tyrasotaenia as a synonym ofVendotaenia. These results undermine the biostratigraphic potentialof this fossil group, and also emphasise the need to reevaluate thebiostratigraphic significance of Sabellidites. Such data also show thata number of “algal” organisms survived or even thrived, during thegreat turnover that marks the Ediacaran–Cambrian transition.

145E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
5.3. Utility of the basal Cambrian GSSP
The existing basal CambrianGSSP can be readily sustained. There areno strong reasons to revisit what had been an almost 30 year-long de-bate, as suggested by Peng and Babcock (2011). The comments byGehling et al. (2001, p. 214, 217) must be repeated—the Fortune HeadGSSP “has proved its utility in recognising the base of the Cambrian”globally (i.e., inNamibia, South and central Australia, California, Nevada,England, southeastern Newfoundland, northwest Canada, China).
Furthermore, a strong negative δ13C excursion and appearance ofA. tornatum–C. velvetum Zone acritarchs just below the lowest localT. pedum specimens can serve as proxies or lower brackets for theCambrian base. Carbonate nodules through some 300 m of theQuaco Road Member above the GSSP have marked negative signalsat Fortune Head (Brasier et al., 1992). However, comparative analysisshould be undertaken of the δ13C of the associated organic matter asit may distinguish local (diagenetic) methanogenic from global seawater signals. We are confident that this differentiation can beattempted. With regard to other proxies, we agree with Gehling etal. (2001, p. 217) that the Fortune Head GSSP “mark(s) a pointabove the last appearance of H. podolica [and, it might be added,Palaeopasichnus delicatus] and below the inception of a marked diver-sification of ichnotaxa.”
5.4. Refined definition of the T. pedum Zone
The long-tested utility of the Fortune Head GSSP should berecognised. It should be strengthened by de-emphasising the signifi-cance of the T. pedum FAD, and by refining the definition of theGSSP horizon as the base of an assemblage zone within a successionof biotas. This is how the situation currently stands with regard tothe lowest Cambrian Terreneuvian Series and Fortunian Stage(Landing et al., 2007a). The Cambrian GSSP simply needs a slight re-statement, as follows—“The position of the coterminent bases of theCambrian System, Terreneuvian Series, and Fortunian Stage at theFortune Head section coincides with the base of the T. pedumIchnozone Assemblage, which has its base defined immediatelyabove the highest occurrences of H. podolica and P. delicatus. This ho-rizon lies slightly above the base of the A. tornatum–C. velvetum Zone.The T. pedum FAD lies below the base of the T. pedum Ichnozone As-semblage, which features the appearance of such characteristic Phan-erozoic ichnogenera as Arenicolites, as well as taxa known elsewhereto appear in the terminal Ediacaran (e.g., Cochlichnus, Didymaulichnus,Monomorphichnus).”
Although no other correlation method currently is available forthe Fortune Head GSSP, available evidence shows that a basal Cambri-an GSSP based on a T. pedum Ichnozone Assemblage as redefinedabove (and not based on the FAD of T. pedum) is applicable globally.In the absence of trace fossils, the nadir of a strong negative carbonisotope excursion that occurs globally just below the T. pedumIchnozone (BACE excursion of Zhu et al., 2004) is a proxy for theEdiacaran–Cambrian boundary. The Fortune Head GSSP for the baseof the Cambrian is over 20 years old. It has reached “maturity” by virtueof its utility in hundreds of scientific syntheses andmany text books. It isbracketed by highly resolved U–Pb dates. There is no reason to confusethe wider audience of non-Cambrian specialists; nor should the Inter-national Cambrian Subcommission be encouraged to perpetuate itsexistence by revisiting and changing its own conclusions.
This redefinition of the base of the Cambrian does not mean a re-version to the use of FADs. A redefinition of the concept of the Cam-brian basal GSSP is proposed that more accurately records thegradual diversification of coelomate burrowers and the extinction ofa number of characteristic Ediacaran taxa. This redefinition utilisesall available bio- and chemostratigraphic data and geochronologic in-formation. As emphasised by Cowie et al. (1986) and Cowie (1986), aGSSP provides a standard for the base of a chronostratigraphic unit
and is a horizon into which other sections are correlated. A GSSP isas a physical point in a stratigraphic section—all correlations into abiotically defined GSSP reflect the limits to resolution of all biostrati-graphic correlations. What is evident is that the Fortune Head GSSPand coeval sections worldwide provide the best basis for biostrati-graphic and nonconventional global correlation of the Cambrian base.
6. Stage 2 and the Cambrian Evolutionary Radiation
6.1. Historical development
The second major phase of the Cambrian Evolutionary Radiation in-volved the origin and diversification of most extant metazoan phyla andclasses with mineralised (carbonate and phosphate) or agglutinatedskeletons, together with the evolution of more diverse Phanerozoiccommunities (e.g., Landing et al., 1989; Landing and Westrop, 2004;Kouchinshy et al., 2012). The onset of this radiation included appearanceof low diversity skeletal fossil assemblages with possible coelenterates(Anabarites Missarzhevsky in Voronova and Missarzhevsky, 1969) andpseudoconodonts (not “protoconodonts,” see Landing and Murphy,1991) such as Protohertzina Missarzhevsky, 1973. These latter taxa havebeen traditionally reported above the terminal Ediacaran, negative δ13Cexcursion on the northern Iran and South China carbonate platforms(Hamdi et al., 1989; Brasier et al., 1990; Hamdi, 1995; Li et al., 2009),Mongolia (Brasier et al., 1990), northwest Laurentia (Nowlan et al.,1985), and Siberia (e.g., Knoll et al., 1995; summary in Kouchinsky et al.,2007). However, these early skeletalised taxa now are known to appearlocally below the BACE excursion as terminal Ediacaran forms in Siberiaand Spain (Zhuravlev et al., 2012).
Appearing higher in a number of lowest Cambrian successionsworldwide is a more dramatic skeletalised metazoan diversificationmarked by the appearances of micromollusks, enigmatic taxa withcap-shaped sclerites, and orthothecid hyoliths in the middleTerreneuvian Epoch about 10 m.y. after the beginning of the Cambri-an (Landing et al., 1998; Maloof et al., 2010b). These diverseskeletalised metazoan assemblages are known in tropical lithofaciesthat include the Siberian Tommotian Stage (e.g., Rozanov et al.,1969; Rozanov and Zhuravlev, 1996), in strata termed Tommotianin western Mongolia (Brasier et al., 1996; Esakova and Zhegallo,1996) and northern Iran (Brasier et al., 1990; Hamdi, 1995), and inthe middle Meishucunian Stage (“Interval B”) of South China (Luoet al., 1984; Brasier et al., 1990; Qian, 1999; Steiner et al., 2007). Di-verse skeletalised metazoans also abruptly appear in cool-waterlithofacies of the middle–upper parts of the regional Placentian Seriesin Avalonian England and southeastern Newfoundland (Brasier, 1984,1986; Landing et al., 1989; Brasier et al., 1992) (Figs. 4, 6).
6.2. Micromollusks and Stage 2
The FADs of the early mollusks Watsonella crosbyi Grabau, 1900,and Aldanella attleborensis (Shaler and Foerste, 1888) have been sug-gested to define GSSPs for the base of Cambrian Stage 2 (Steiner et al.,2007, 2011; Rozanov et al., 2008; Li et al., 2011; Parkhaev andKarlova, 2011; Parkhaev et al., 2011). Peng and Babcock (2011, fig.1), as officers of the International Subcommission on Cambrian Stra-tigraphy, even seem to presage a decision by the Subcommission byfiguring the FAD of W. crosbyi as a “preferred base” of Stage 2.
Globally distributed and known in tropical to temperate shelf set-tings (Fig. 6), the fossils of W. crosbyi and A. attleborensis would seemto provide a superb basis for correlations between Early Cambrianpalaeocontinents. However, as discussed below, the FADs of bothtaxa are highly diachronous locally and globally, are strongly tied tothe near-shore lithofacies in which they originated, and the preserva-tion of these micromolluscs is limited to certain diagenetic facies. Inaddition, W. crosbyi and A. attleborensis are very long-ranged withintheir “topotype” Avalonian faunal province (Figs. 3, 6), and their

Fig. 6. Lower Cambrian distribution of the rostroconch Watsonella crosbyi (W) and gastropod Aldanella attleborensis (A); sources recording occurrences are listed in text.
146 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
ranges on other Cambrian palaeocontinents typically only representpart of their Avalonian range. Finally, as a good example of the diffi-culties associated with FADs (Fig. 1), differing taxonomic practicesmean that A. attleborensis is reported under many (14) speciesnames in Siberia—this produces the problem that A. attleborensis isinterpreted herein as occurring well below the Tommotian Stage, al-though the much higher FAD of A. attleborensis as the species is taxo-nomically defined by Parkhaev and Karlova (2011), among others,means that its FAD has been proposed as an index to the bases ofthe regional Tommotian Stage and a global Cambrian Stage 2.
6.3. W. crosbyi
6.3.1. Taxonomy and distributionW. crosbyi is the oldest known rostroconch (Landing, 1989). It does
not have “undetermined” or non-rostroconch affinities within the mol-lusks as concluded by Carter et al. (2000), Parkhaev (2001), and Li et al.(2011). The lack of a rostrum is not an argument against Watsonellabeing a rostroconch as early rostroconchs, such as ribeirioids, lack arostrum (Pojeta and Runnegar, 1976). In addition, W. crosbyi mouldscommonly, but not invariably (Parkhaev, 2001), have an incisioninterpreted as a rostroconch pegma (Landing, 1988, fig. 5.16, Kerber,1988, pl. 4, figs. 1b, 9). An adventitious pegma may be related to W.crosbyi's small size and its life as a shallow burrower in soft mud(Landing, 1989). Its autecology likely did not require a buttress-likepegma to keep the valves and aperture open during burrowing.
W. crosbyi has been synonymised withHeraultipegma varensalensis(Cobbold, 1935a), first described from Gondwanan south France, andwith H. yunnanense He and Yang, 1982, of South China by Landing(1988) and Landing et al. (1989). This species is also regarded as thesenior synonym of the Siberian “species” W. sibirica (e.g., Li et al.,2011). These synonymies mean that W. crosbyi is known from mostCambrian palaeocontinents (Fig. 6). The species was used to definethe base of a newly proposed W. crosbyi Zone in the middle ChapelIsland Formation in eastern Newfoundland (Fig. 3) and to correlatethis zone with the middle Meishucunian Stage of South China(Landing et al., 1989).
6.3.2. Occurrence in South ChinaW. crosbyi (as Heraultipegma yunnanensis) was recognised as an
important biostratigraphic aid across South China by Qian et al.
(1996) who figured but did not define a variously named H.yunnanense “Range Zone,” “Acme Zone,” and “Assemblage Zone.”The zone ranged from less than a metre thick to ca. 20 m in SouthChina (Qian, 1999). Steiner et al. (2004, 2007, 2011) reviewed thedistribution of a renamed “W. crosbyi Assemblage Zone” in SouthChina and defined the zone by the association of W. crosbyi, thesnail A. attleborensis (their A. yangjiaheensis Chen, 1984, discussedbelow), and the helcionelloid Oelandiella korobkovi (Vostokova,1962). The W. crosbyi Assemblage Zone is of such importance toSouth China correlation that it is recorded at Meishucun quarry(Parkhaev and Demidenko, 2010), where the zone is completely cutout at an unconformity (Li et al., 2009).
The Chinese W. crosbyi Assemblage Zone is less an “assemblagezone” than a simple “range zone” of W. crosbyi. Both A. attleborensis,which is uncommon in South China, and O. korobkovi occur infre-quently with W. crosbyi in South China sections (Qian, 1999). Indeed,A. attleborensis and O. korobkovi, despite their definition as associatesin theW. crosbyi Zone, appear below the range ofW. crosbyi (e.g., Xinget al., 1983; Steiner et al., 2007, fig. 12), and this observation rein-forces a conclusion that the Chinese W. crosbyi Assemblage Zone issimply the W. crosbyi range zone.
The FAD of W. crosbyi and duration of the W. crosbyi AssemblageZone in South China arguably have little geochronologic significance,even though the fossils of W. crosbyi can be regarded as rangingthrough “about the correct” biostratigraphic position. No confidenceinterval (cf. Marshall, 1990) could ever be calculated for W. crosbyi'sFADs and LADs at any South China section because they lie atlithostratigraphic breaks. Indeed, the tops of the Chinese W. crosbyiAssemblage Zone and the Dahai Member in which it occurs are eithertruncated by an unconformity or are followed by poorly fossiliferous,siliciclastic-dominated sedimentary rocks (Zhu et al., 2001a; Li et al.,2009) (Fig. 5). Furthermore, the W. crosbyi FAD lies at a lithologicchange from dolostones to limestones in the Dahai Member (Fig. 5)—likely indicating environmental or preservational definition of thisFAD (e.g., Fig. 1).
6.3.3. Range offset in West Gondwanan FranceW. crosbyi, long known as Heraultia varensalensis Cobbold, 1935a, or
Heraultipegma valensalensis, occurs in a small oucrop belt of the HeraultiaLimestone on the northeast margin of theMontage Noire inlier, southernFrance. Traditional stratigraphic correlations between the autochthonous

147E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
succession of which the Heraultia Limestone is a part with apparentlyidentical successions in overlying thrust sheets led to an upper LowerCambrian (Atdabanian- or Botoman-equivalency) assignment of theHeraultia Limestone. Thus, the Heraultia Limestone was regarded asyounger than the underlying Pardailhan Formation with itsAtdabanian–Botoman-boundary interval archaeocyathans and was cor-related with the “Série noire” with Botoman archaeocyathans (see re-views in Kerber, 1988; Álvaro and Clausen, 2007; Álvaro et al., 2011). Aseeming problem with this correlation with commonly trilobite-bearingrocks is the absence of trilobites in the Heraultia Limestone (Kerber,1988). Although Álvaro and Clausen (2007) and Álvaro et al. (2011)noted that “trilobites” and “crustaceans” occur in the unit, only smallshelly fossils have been recovered (Cobbold, 1935a; Kerber, 1988).Gubanov (2002) noted that Kerber (1988) illustrated a pre-Botomanand -Atdabanian-equivalent microfauna from the Heraultia Limestonewith W. crosbyi associated with a number of conoidal fossils that areknown in the lower and middle Meishucunian of South China. However,the presence of apparent paterinid brachiopods (Kerber, 1988) sugggeststhat the older part of the range of W. crosbyi as recorded in theMeishucunian of South China and Siberian Manykaian–“Nemakit-Daldynian” is not recorded in the Heraultia Limestone. New workshows that the Heraultia Limestone is better regarded as a lithologicmember of a previously unrecognised, thick, older (likely Terreneuvian)dolostone-dominatedunit that overlies rhyolites. This rhyolite–dolostonesuccession forms the oldest part of the allochthonous cover sequence onthe Montagne Noire's Proterozoic core (Álvaro et al., 2013). This newwork further emphasises that the Heraultia Limestone lithofacies andthe FAD of W. crosbyi are preservational artefacts that escapeddolomitisation. Thus, a range offset of unknown length separates the spe-cies' Montagne Noire FAD fromW. crosbyi's evolutionary origin.
6.3.4. Range offsets in Mongolia and AustraliaIn Mongolia (Brasier et al., 1996), W. crosbyi occurs in younger,
Atdabanian-equivalent intervals. Esakova and Zhegallo (1996)assigned the Mongolian occurrence of W. crosby, which likely is ac-companied by A. attleborensis (Parkhaev and Karlova, 2011), toTommotian-equivalent rocks. However, a carbon-isotope-based cor-relation favours an Atdabanian correlation (Brasier et al., 1996). Sim-ilarly, W. crosbyi from the Bemella communis and Stenothecadrepanoida assemblages (equivalent to the Parabadiella huoi Zone)of the Sellick Hill Formation of South Australia occurs within anAtdabanian-equivalent interval (Gravestock et al., 2001; Gubanov,2002) (Fig. 4).
6.4. A. attleborensis
In southeast Newfoundland, the FADs ofA. attleborensis andW. crosbyiare essentially identical (Landing, 1988; Landing et al., 1989) (Fig. 3). A.attleborensis is morphologically identical to A. yangjiaheensis in SouthChina, and A. attleborensis is regarded as the senior synomym (Landing,1988; Parkhaev and Demidenko, 2010, p. 891; Parkhaev and Karlova,2011). Although A. attleborensis is uncommon in South China, it has thesame range asW. crosbyi at a northern Shaanxi section (Qian, 1999, p. 89).
Aldanella is recorded as formingmulti-species associations on the Si-berian Platform, and supposedly shows rapid evolution in the terminalManykaian/“Nemakit-Daldynian”–early Tommotian (Rozanov et al.,1969; Parkhaev and Karlova, 2011; Parkhaev et al., 2011). Parkhaevand Karlova (2011) exhaustively illustrate named Siberian Aldanellaspecies. Their species synonymies emphasise just how fluid the defini-tion and identification of these “species” has been over several genera-tions of regional research. Parkhaev and Karlova's (2011) synonymy ofA. attleborensis, whose FAD is proposed to define the base of Stage 2(Parkhaev et al., 2011), includes six named “species,” in addition to A.attleborensis, that were taxonomically differentiated in earlier reportson the Siberian material.
The interpretation herein of the “valid species” distinguished byParkhaev and Karlova (2011) is that they are all synonymous withthe Avalonian genotype A. attleborensis if a practical concept of taxon-omy is applied (i.e. Landing, 1988; Landing et al., 1989). With the lim-ited morphological and anatomical data available from the conchs ofthese tiny, early snails, a robust taxonomy that reflects biological dis-tinctiveness is difficult to develop.
The Siberian “species” recognised by Parkhaev and Karlova (2011)include A. crassaMissarzhevsky in Rozanov et al. (1969) (with the un-common sinistral variant A. golubevi Parkhaev, 2007, and the gentlyribbed variant termed Pseudoyangtzespira selindeica Bokova, 1990);A. operosa Missarzhevsky in Rozanov and Missarzhevsky (1966); A.utchurica Missarzhevsky in Rozanov et al. (1969); and A. sibiricaParkhaev and Karlova, 2011. The supposedly diagnostic features ofthe Siberian species include conch height, whorl expansion rate,whorl number (1.5–3) and cross-section (flattened or inflated), incli-nation of whorl long axis to coiling axis, whorl surface smooth or withoblique low ribs that are moulded on steinkerns, and conch diameter(up to 5 mm). However, all of these features fall within the range ofvariability shown by the holotype and topotypes of A. attleborensisfrom the Sunnaginia imbricata Zone (Fig. 3) in eastern Massachusetts(Landing, 1988, and unpublished data).
7. Stratigraphic ranges of W. crosbyi and A. attleborensis
7.1. Avalonia and stage 2 of the Cambrian Radiation
A summary of the geological setting of the “type” W. crosbyi Zone inAvalonia (Figs. 2A, 3) is required because this zone has potential forinterregional correlation and for understanding the geochronology ofthe skeletalised metazoans that appear in stage 2 of the CambrianRadiation. The “type”W. crosbyi Zonewas defined in the Burin Peninsula,southeast Newfoundland, from a cool-water, carbonate-poor suc-cession (see Landing, 1988, 1992, 2004, 2005; Landing et al., 1989,2013b; Myrow and Landing, 1992; Landing and Westrop, 2004).The Burin Peninsula lies on the Avalon marginal platform, whichextends from North Wales to southern New Brunswick and forms thenorthwest edge of the fragmented Avalonia continent in the laterAcadian–Caledonian orogen (Landing, 1996a). The marginal platformhas a regionally uniform terminal Ediararan–middle Terreneuvian suc-cessions that reach 3.0 km-thick. The nearly identical FADs ofW. crosbyiand A. attleborensis lie well above a lower feldspathic, commonlysubaerial conglomeratic unit on the marginal platform (RencontreFormation, Fig. 3). These two species have FADs in the middle of asiliciclastic marine shelf formation (Chapel Island Formation) eitherjust below a sequence boundary (Newfoundland) or just above thisregional sequence boundary (New Brunswick) (Landing et al., 1989;Landing, 2004) (Fig. 3). This marine shelf unit is conformably overlainby a coarse-grained, quartzose tidalite (Random Formation, Fig. 3)that lacks shelly fossils.
The Avalonian inner platform is southeast of the marginal platform(Fig. 7) and extends from eastern Massachusetts to England. A quartz-ose tidalite is the lowest Cambrian unit on the inner platform and un-conformably overlies an Avalonian basement with a Cryogenian–lower Ediacaran arc succession (Landing, 1996a, 2004; Landing et al.,2013b). W. crosbyi and A. attleborensis appear as “Lazurus taxa” abovethe tidalite sandstone and persist to the top of the subtrilobitic LowerCambrian (Landing, 1992) (Figs. 1, 3).
7.2. Watsonella and Aldanella are not Tommotian-limited
In southeastern Newfoundland, the LADs of W. crosbyi and A.attleborensis occur at the top of the Fosters Point Formation, and justunder the unconformity with the lowest trilobite-bearing (Callaviabroeggeri Zone) beds of the Brigus Formation (Fig. 3). No other Cambri-an region shows such a great stratigraphic range (650 m) of these two

Fig. 7. Suggested GSSP for the base of Stage 2, or the “Laolinian Stage.” (A) Locations ofthe Laolin, Meishucun, and Xiaotan sections in northeastern Yunnan, southwesternChina. (B) Sketch map of the Laolin section with sampled outcrops in the road cutand lithostraphic units. Abbreviations: BYS, Baiyanshao Member; DB, Daibu Member;ZYC, Zhongyicun Member; DH, Dahai Member; and SYT, Shiyantou Formation.Modified from Li et al. (2009, fig. 1).
148 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
potential GSSP-defining taxa (Fig. 3). The ca. 20 m range of these spe-cies in South China and Siberia (e.g., Rozanov et al., 1969; Qian, 1999)is striking—particularly as their 650 m range in Avalon lies largelywith-in a shale-basin facies deposited at a relatively low rate of accumulationfor siliciclastic sedimentary rocks (see Landing and Benus, 1988;Myrowand Landing, 1992; Landing and Westrop, 2004; Landing et al., 2013b).
The short stratigraphic ranges of the two species in Siberia and SouthChina likely reflect a number of factors: truncation of ranges at thewide-spread sub-Tommotian unconformity in Siberia and at the top ofthe Meishucunian in China (e.g., Landing, 1994), unfavourablepalaeoenvironments for colonisation or preservation, and incompletecollecting. The longer (150 m) range of W. crosbyi, apparently with A.attleborensis in western Mongolia (Parkhaev and Karlova, 2011), is alsotruncated at an unconformitywith theAtdabanian-equivalent SalanygolFormation (Brasier et al., 1996; Esakova and Zhegallo, 1996). Since bothspecies have been proposed to define the base of a global Stage 2 of theCambrian, it is important to evaluatewhether or not the Asiatic ranges ofW. crosbyi and A. attleborensis correlate completely with or only into asmall part of their 650 m range in Avalonia.
A particular concern, which is critically examined below, is whetheror not the composite Avalonian range ofW. crosbyi and A. attleborensis iscoeval with the Siberian Tommotian Stage as claimed by Steiner et al.(2007) and Rozanov et al. (2008). A second concern is whether thesetwo potential GSSP-defining FADs are diachronous in Avalonia or else-where, as this would question their use as a GSSP. Finally, it must beevaluated whether the chronostratigraphic and correlation significanceof the two species can be evaluated by biostratigraphic data on othergroups and by carbon isotope and geochronologic correlations, whichcan then be tied to the ranges of W. crosbyi and A. attleborensis.
7.3. Diachroneity of Avalonian W. crosbyi Zone base
The inadequacy of a Cambrian Stage 2 base defined by theW. crosbyior A. attleborensis FADs was detailed in theW. crosbyi Zone “type area.”Landing et al. (1989, p. 748, 749, fig. 1) showed a 50 m difference in theessentially coterminous FADs of W. crosbyi and A. attleborensis in sec-tions separated by 26 km in southest Newfoundland. The diachroneityin the two species' FADs is an artefact of preservation—where diagene-sis in dark green-greymudstone led to pyritisation, enhanced preserva-tion, and a much lower FAD of the W. crosbyi Zone assemblage at LittleDanzig Cove. In contrast, these species' conchs were dissolved awayduring diagenesis of the lighter coloured mudstones at the FortuneNorth section, and are limited to thin limestone beds.
A lithofacies–biofacies linkage is shown by the lowest appearanceof W. crosbyi Zone taxa with the onset of shoaling in the upper QuacoRoad Member of the Chapel Island Formation in southeastern New-foundland (Fig. 3). The highest diversity W. crosbyi Zone assemblages(19 species) in southeastern Newfoundland occur higher in theshallowest facies (peritidal shell hash and calcareous mud moundlimestone with caliche) that bracket the intra-Chapel Island sequenceboundary (Landing et al., 1989; Landing and Westrop, 2004) (Fig. 3).
Preservational bias in the occurrence of W. crosbyi Zone faunas isseen in southern New Brunswick, where the intra-Chapel Island Forma-tion sequence boundary is underlain by unfossiliferous, coarse-grainedsandstone and overlain by a glacial diamictite with lonestones followedby greenishmudstones. Methanogenic calcareous nodules in the green-ishmudstones have pyritised (now limonitic) A. attleborensis and otheroriginally calcareous remains in a low diversityW. crosbyi Zone assem-blage (Landing, 2004; Landing and MacGabhann, 2010).
7.4. “Falling FADs” of Siberian Watsonella and Aldanella
Most early reports (e.g., Khomentovsky and Karlova, 1993) re-stricted W. crosbyi (traditionally called H. varensalensis or H. sibirica)and Aldanella to the Siberian lower Tommotian Stage. Longoverlooked in this literature were reports by Missarzhevsky (1983,1989) of Aldanella “crassa” and other Aldanella “species” (discussedabove) in the underlying Manykaian/“Nemakit-Daldynian” Stage innorthern Siberia (Fig. 5). Controversy allowed dismissal ofMissarzhevsky's work, and this has long influenced thinking that W.crosbyi–Aldanella associations, including those in Avalonia, shouldbe regarded as Tommotian-equivalents (Steiner et al., 2007;Rozanov et al., 2008; Li et al., 2011).
In the Siberian lowest Tommotian, the Watsonella and AldanellaFADs are commonly reported at about the same level, or withAldanella appearing slightly below Watsonella (Rozanov et al., 2008).Almost all of these lowest Tommotian occurrences lie just above awidespread facies change, unconformity, and sequence boundary atthe base of the Tommotian in many areas in Siberia (e.g., Rowlandet al., 1998). Both genera have peculiar distributions in easternSiberia—often being limited to 1–2 m of glauconitic, sandy limestonewith intraclasts (e.g., Khomentovsky and Karlova, 1993). This islikely the onlap facies of the Tommotian across the Manykaian/“Nemakit-Daldynian” Stage dolostones. This litho-biofacies linkageof theW. crosbyi and A. attleborensis FADs and their lowest occurrenceabove an unconformity strongly suggest that the oldest range of thesespecies is typically not preserved in Siberia. Reports of Aldanella justbelow the Tommotian and in the uppermost Ust'-Yudoma Formation(e.g., Varlamov et al., 2008) likely represent specimens fromTommotian-age fissure fills in the Ust'-Yudoma (e.g., Landing, 1994,fig. 2A; and confirmed by our unpublished carbon isotopic studies).
Moczydłowska and Vidal (1988) and Landing (1988, 1994) concurwith Khomentovsky and Karlova (1993) and Brasier et al. (1996) thata widespread unconformity lies at the base of the Tommotian.Landing (1992, 1994) argued that the sudden appearance of a diversefauna (170+ named species) with the lowest archaeocyathans,

149E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
lingulates, and tommotiids in the basal Tommotian likely indicates along, not short, hiatus. Thus, even the early literature suggested thatthe association of W. crosbyi and Aldanella, though essentially simul-taneous in most reports on the Tommotian Stage, does not corre-spond to the base of their complete ranges.
This lowest co-occurrence of the two species in Siberia has notchanged significantly in more recent work on successions where thesub-Tommotian hiatus is regarded as shorter. For example, severalmetres of strata at the top of the Sukharikha Formation in northwestSiberian (Rowland et al., 1998) are referable to the Tommotian. Thesestrata represent at least part of the lowest Cambrian record lost at theUst'-Yudoma (Manykaian/“Nemakit-Daldynian” Stage)–Pestrotsvet(Tommotian Stage) unconformity at Dvortsy, the reference standardfor the Tommotian base in southeast Siberia (Repina and Rozanov,1992; Sukhov, 1997).
Missarzhevsky's (1983, 1989) reports are important for revealingthe sub-Tommotian occurrence of Aldanella in the Olenek Uplift. Heplaced the northern Siberian FAD of A. attleborensis, using theLanding (1988) synonymy discussed above, in the lower part of theI′ negative carbon isotope peak of Kaufman et al. (1996) (Fig. 5).Thus, Missarzhevsky's (1983, 1989) “old” Aldanella's come from strataabsent at the the Ust'-Yudoma–Pestrotsvet unconformity in easternSiberia (Landing, 1994; Kouchinsky et al., 2005). More recently(June 2011), A. Kouchinsky reported to the International CambrianSubcommission that W. crosbyi also occurs in the lower part of the I′negative excursion in the Olenek Uplift (Fig. 5).
These sub-Tommotian occurrences ofW. crosbyi and A. attleborensisnow conform with earlier conclusions about the older stratigraphicranges of these species in Avalonia and China (Landing, 1988; Landinget al., 1989). These reports also undermine assumptions (e.g., Steineret al., 2007; Rozanov et al., 2008) concerning the Tommotian-equivalence of assemblages bearing W. crosbyi and A. attleborensis inSouth China and in the Avalonian “type”W. crosbyi Zone. The lengthen-ing stratigraphic ranges for both species emphasise that their rangeshave been incompletely established in Siberia. This, in turn, weakensproposals to use the FAD of either species to define a GSSP for thebase of Stage 2.
7.5. Biotic correlation of upper Watsonella–Aldanella associations
The long ranges of W. crosbyi and A. attleborensis in Avalonia argu-ably record much longer time intervals than those recorded by thetwo species in the thin/condensed sections in Siberia and South China.As discussed above, the Avalonian “type” W. crosbyi Zone correlatesfar below and into sub-Tommotian-equivalent strata. Landing et al.(1989, pp. 752, 753; also Landing, 1988, 1992) therefore concludedthat only the higher parts of the ranges ofW. crosbyi and A. attleborensisin Avalonia (Fig. 3) may be as young as the Tommotian Stage.
The assumption that aW. crosbyi–A. attleborensis association equateswith the Siberian Tommotian has become a form of circular logic. Awayout of this logic is provided by other key taxa and the carbon isotopestratigraphy of the Siberian Tommotian. Indeed, the Tommotian taxaConotheca mammilata Missarzhevsky in Rozanov et al. (1969), S.imbricata Missarzhevsky in Rozanov et al. (1969), and Torellellalaevigata (Linnarsson, 1871) have their local FADs well above the“type” W. crosbyi Zone, and only appear in the S. imbricata Zone inAvalonia (Fig. 3). The S. imbricata Zone also has the oldest Avalonianspecimens of Tiksitheca korobkovi Missarzhevsky in Rozanov andMissarzhevsky (1966), a taxon with a FAD in the older Manykaian/“Nemakit-Daldynian” and which persists into the Tommotian.
W. crosbyi and A. attleborensis range higher into the AvalonianCamenella baltica Zone (Figs. 3, 4). The contact of these faunal zonesis transitional—the C. baltica Zone base is merely the FAD of C. balticain an unbroken sequence of thin-bedded limestone and redsiliciclastic mudstone alternations in the middle Cuslett Formation(Fig. 3) (Landing, 1988; Landing et al., 1988, p. 48, 49; Brasier et al.,
1992). No evidence exists for an “unrecognised hiatus” at the S.imbricata–C. baltica zonal boundary, as indicated by Steiner et al.(2007, p. 90, fig. 12).
Most of the C. baltica Zone is a generalised equivalent of the mid-dle–upper Tommotian based on the gradual addition of typically“younger” SSFs upward through the zone. Correlation of the C. balticaZone is based on the following: occurrence of C. baltica in the Siberianupper Tommotian–Atdabanian; shared lowest occurrences of thepseudoconodont Rhombocorniculum insolutum Missarzhevsky inMissarzhevsky and Mambetov (1981); FADs of hyolithid hyoliths inthe upper Tommotian and middle C. baltica Zone; appearance oforthothecids with triangular cross sections (e.g., GracilithecaSysoiev, 1968) in the middle Tommotian and lower C. baltica Zone;and appearance of the bivalve Fordilla troyensis Barrande, 1881, andRhombocorniculum cancellatum (Cobbold, 1921) in the upper C.baltica Zone and Atdabanian Stage. The lowest specimens ofAnabaritellus tricarinatus Missarzhevsky in Rozanov et al. (1969)(= A. hexasulcatus Missarzhevsky, 1974; see Landing and Murphy,1991) lies in the lower Tommotian and in the middle C. balticaZone, which further strengthens a Tommotian correlation for mostof the C. baltica Zone (Landing, 1992).
7.6. Carbon isotopes and uppermost Watsonella–Aldanella association
Brasier et al. (1992) determined a carbon isotope stratigraphythrough the S. imbricata–C. baltica zones (and upper range of the W.crosbyi–A. attleborensis assemblage) in southeast Newfoundland andAvalonian England. The analytical values obtained and used to createthe Avalonian carbon isotope stratigraphy are independent ofchanges in oxygen isotopes, and the gradual changes in carbon iso-tope values through the section are likely primary, and not resetwith burial diagenesis and chlorite-grade metamorphism.
This δ13C stratigraphy of the uppermost Watsonella–Aldanella as-sociation in Avalonia is comparable to intervals II–IV of Kaufman etal. (1996) from Siberia (Fig. 5). Two positive carbon isotope peaks(III and IV) separated by a long negative interval in the Siberian mid-dle Tommotian–lower Atdabanian (Kaufman et al., 1996) are plausi-bly recorded in the upper 100 m of the ranges of W. crosbyi andAldanella attleborenis in southeast Newfoundland (Brasier et al.,1992) (Fig. 3). These carbon isotopic correlations of Brasier et al.(1992) support the Avalonia–Siberian correlations based on smallshelly fossils (Landing, 1988, 1992, 1994; Landing et al., 1989), butyield more highly resolved correlations. Correlation of the strong pos-itive peak in the lower Cuslett Formation of Newfoundland with themiddle Tommotian III peak of the upper Dokidocyathus regularisZone (archaeocyathans) in Siberia also enables re-evaluation of a pro-posed acritarch-based correlation of the Avalonian lowest Cambrianby Palacios et al. (2011) (as discussed below).
A negative excursion through the middle Cuslett Formation isfollowed by a strong positive peak in the thin (ca. 12 m) Fosters PointFormation just below the unconformity with the lowest C. broeggeriZone trilobites (Figs. 3–5). Brasier et al. (1992) recorded a comparablecarbon isotope and faunal succession in the Jees and Home Farmmem-bers in the English Midlands (Fig. 3). They correlated this negative tostrong positive δ13C peak with the middle Tommotian–lowerAtdabanian carbon isotope record that leads to the peak of excursionIV in the lower Atdabanian (Margaritz et al., 1986; Kirschvink et al.,1991; Brasier et al., 1994b; Kaufman et al., 1996).
7.7. S. imbricata Zone and carbon isotopes
Most of the ca. 100 m-thick S. imbricata Zone in southeasternNewfoundland (Fig. 3) records a progression from strongly negativevalues to a positive δ13C peak, followed by the suggestion of oscillato-ry values. Brasier et al. (1992, fig. 14) provisionally equated these δ13Cvalues to excursion 1 of the upper Ust'-Yudoma Formation in the

150 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Uchur–Maya region of the eastern Siberian Platform (Fig. 5). A bettersupported correlation, given the lowest Tommotian-aspect smallshelly fossils in the lower S. imbricata Zone (discussed above) is thatthis strongly negative to positive excursion represents lowerTommmotian excursion II of Kaufman et al. (1996). The subsequentstrong fall and oscillation in carbon isotope values in higher BonavistaGroup strata (Fig. 5) are similar to the isotope sequence between ex-cursions II and III in eastern Siberia (Kouchinsky et al., 2005)
8. Correlation of the “type” W. crosbyi Zone
8.1. Biostratigraphy
Biostratigraphic correlation of the top 200 m of the Watsonella–Aldanella association in southeastern Newfoundland is consistentwith a conclusion that the much lower “type” W. crosbyi Zone of themiddle Chapel Island Formation is older than the Siberian TommotianStage (Fig. 3). A sub-Tommotian correlation was first suggested onthe basis of taxa in the “type” W. crosbyi Zone (Landing, 1988, 1992,1994; Landing et al., 1989) and its lack of typical Tommotian–lowerAtdabanian fossils, which only appear higher in the S. imbricata–C.baltica zones (listed above).
A few species in the “type”W. crosbyi Zone that do not persist abovethe Random Formation (Fig. 3) suggest correlation with the middleMeishucunian Stage and W. crosbyi Assemblage Zone in South Chinaand with the Siberian middle Manykaian/“Nemakit-Daldynian” Stage(Landing, 1988, 1992, 1994). These include the pseudoconodontsMaldeotaia bandalica Singh and Shukla, 1981, known from the lowestCambrian of the Lesser Himalaya, India, and the longer ranging P.anabarica Missarzhevsky, 1973, which occurs in the lowest Cambrianof Himalayan India, west Laurentia, Siberia, and South China (e.g.,Brasier and Singh, 1987). Other sub-Tommotian taxa occur in the“type” W. crosbyi Zone (e.g., Anabarella plana Vostokova, 1962) orclosely resemble congeneric species (i.e., Bemella? vonbitteri andArchaeospira? avalonensis of Landing et al., 1989) that have lowestoccurrences in the middle Manykaian/“Nemakit-Daldynian” and mid-dle Meishucunian (Landing et al., 1989). A biotic correlation of the low-est W. crosbyi- and A. attleborensis-bearing faunas of the “type” W.crosbyi Zone with sub-Tommotian-equivalent strata is also consistentwith the biostratigraphic and carbon isotope-based Tommotian–earlyAtdabanian correlation of the higher S. imbricata–C. baltica zones(discussed above).
8.2. Carbon isotope correlation
This sub-Tommotian correlation of the “type”W. crosbyi Zone is con-sistent with its provisionally sampled δ13C stratigraphy (Brasier et al.,1992). This sampling included only nine samples through ca. 85 m ofthe middle Chapel Island Formation (Fig. 5). A convincing carbonisotope-based correlation of this interval is limited by the fact that thesiliciclastics of the overlyingMystery LakeMember and RandomForma-tion do not provide a carbon isotope stratigraphy (Fig. 3).
However, the carbon isotope record shows an oscillatory rise fromnegative to positive values through the “type” W. crosbyi Zone(Brasier et al., 1992) (Fig. 5) that compares with the δ13C patternthat leads up to carbon excursion I of the lower–middle Manykaian/“Nemakit-Daldynian” (Kaufman et al., 1996). Alternatively, it compareswith the stratigraphically higher oscillatory pattern that leads to the pos-itive I′ excursion in the upperManykaian/“Nemakit-Daldynian” in Siberia(Kaufman et al., 1996) and the L4 excursion in the upper Dahai Memberin South China (Li et al., 2009). The latter alternative is preferred as theupper Manykaian/Nemakit/“Daldynian”and upper Dahai Member haveW. crosbyi–A. attleborensis assemblages. Li et al. (2009) also noted thatthe L4 peak lies above the terminal Ediacaran positive excursion andabove strata with low-diversity, earliest Cambrian small shelly fossilsand T. pedum.
Correlation of the I′ and L4 excursions (Li et al., 2009) and theirsimilarity with the incompletely known carbon stratigraphy fromthe “type” W. crosbyi Zone, as well as the SSF similarities of the Sibe-rian, South China, and Avalonian “type” W. crosbyi Zone (Landing,1988; Landing et al., 1989), support a sub-Tommotian equivalencyof the oldest Watsonella–Aldanella associations on three Cambrianpalaeocontinents (Figs. 3–5). The most parsimonious correlation ofthe oldest AvalonianWatsonella–Aldanella association is into an inter-val absent at the Manykaian/“Nemakit-Daldynian”–Tommotian un-conformity on the Aldan River at Dvortsy, Ulakhan–Sulugur, andelsewhere in southeast Siberia. At least part of this interval is pre-served in the Olenek Uplift (Kessyuse Formation; e.g., Meshkova etal., 1973; Missarzhevsky, 1980) of northwest Siberia (Landing, 1988,1992).
9. Acritarchs and “type” W. crosbyi Zone correlation
The “standard” terminal Ediacaran–Lower Cambrian acritarch zo-nation was developed in Baltica (Moczydłowska, 1991, 1998, 2011). Thezonation features an upward succession through the A. tomatum–C.velvetum (A.–C.), Skiagia ornata–Fimbrioglomerella membranacea (S.–P.), Heliosphaeridium dissimilare–Skiagia ciliosa (H.–S.), Volkoviadentifera–Liepaina plana (V.–L.), and Eliasum–Cristallinium (E.–C.)zones (Figs. 3, 4). These zonal assemblages have also been recordedfrom East Gondwana (South Australia; Zang et al., 2007), SouthChina (Zang, 1992; Yin et al., 2011; Yuan et al., 2011), West Gondwa-na (Iberia), and Avalonia (Palacios et al., 2011).
9.1. Sub-W. crosbyi Zone correlation
Palacios et al. (2011) sampled the Chapel Island Formation foracritarchs in the Saint John, New Brunswick, area (Figs. 4, 7). Their“Ratcliffe Brook Formation” (Hayes and Howell, 1937; Alcock, 1938;Tanoli and Pickerill, 1988), a traditional but abandoned designation(Landing, 1996b, 2004; Landing and Westrop, 1996, 1998a), includessubaerial rift conglomerates of the Rencontre Formation and overly-ing shelf siliciclastics of the Chapel Island Formation (Fig. 3). In theSaint John area, as in the Burin Peninsula, southeast Newfoundland,and elsewhere on the Avalonian marginal platform, the Chapel IslandFormation is overlain by coarse-grained tidalite sandstone of the Ran-dom Formation (i.e., “Glen Falls Formation” of Palacios et al., 2011)(see Landing and Westrop, 1996, 1998a; Landing et al., 2008).
Ichnofossils low in the Chapel Island Formation are lowest Cam-brian at the Hanford Brook section (Matthew, 1890; Hayes andHowell, 1937; Tanoli and Pickerill, 1988; Hofmann and Patel, 1989)(Fig. 7, locality HBE). Acritarchs from the lowest 50 m of the ChapelIsland Formation on Hanford Brook were referred to the terminal Edi-acaran–lowest Cambrian A.–C. Zone of Baltica by Palacios et al. (2011,fig. 2) (Figs. 3, 4).
Only two taxa limited to the A.–C. Zone in Baltica, Pterospermella velataMoczydłowska, 1988, and C. velvetum Moczydłowska, 1988, werereported from the lower Chapel Island Formation. The figured P. velataspecimen (Palacios et al., 2011, fig. 3F) is poorly preserved and cannotbe identified confidently (M. Moczydłowska, 2011 personal communica-tion to E. Landing andG. Geyer). A.H. Knoll (personal communication to E.Landing, 2011) agreed with this evaluation and noted that identificationof the specimen is compounded by its distortion by mineral growths. Inaddition, the figured C. velvetum specimen (Palacios et al., 2011, fig. 3C)is broken, and the sparse, short processes make a species- and perhapsgenus-level identification questionable. The ranges of P. velata and C.velvetum are only well defined on the Lublin Slope of the East EuropeanPlatform, and their occurrence there does notmean that they are restrict-ed to the lowermost Cambrian acritarch zone (M.Moczydłowska, person-al comm., 2012). Despite these uncertainties, the lowest Chapel IslandFormation on Hanford Brook is likely referable to the lowest Cambrianpart of the A.–C. Zone.

151E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
9.2. Acritarchs from W. crosbyi Zone
A low diversity W. crosbyi Zone assemblage occurs just above thesequence boundary and in the Mystery Lake Member of the ChapelIsland Formation on Hanford Brook (Landing and Westrop, 1998a;Landing, 2004; Landing and MacGabhann, 2010). This assemblage isaccompanied by acritarchs assigned to the second Lower Cambrianacritarch assemblage of Baltica (Palacios et al., 2011) (Figs. 3, 4). Allof the S.–F. Zone acritarchs reported from this interval on HanfordBrook actually persist into the overlying H.–S. Zone in Baltica(Moczydłowska, 1991, 1998, 2011; Moczydłowska and Zang, 2006).Thus, an unambiguous correlation with the S.–F. Zone in Baltica isnot possible. The lowest range of this tentative S.–F. Zone assemblageis likely not recorded on Hanford Brook as its lowest occurrence isabove the intra-Chapel Island Formation sequence boundary, and anacritarch-free interval extends down to the A.–C. Zone.
9.3. Correlation of S.–F. Zone acritarchs from W. crosbyi Zone
Palacios et al. (2011, p. 54, fig. 7) mistakenly concluded that theW.crosbyi Zone and S.–F. Zone assemblage on Hanford Brook correlate withthe S. imbricata–C. baltica zones elsewhere in Avalonia (Fig. 3). However,biostratigrahic and carbon isotope correlations discussed above mandatethat the S. imbricata–C. baltica zones are Tommotian–lowest Atdabanianequivalents. The correlation by Palacios et al. (2011) followed a super-seded conclusion that S.–F. Zone acritarchs occur in the middleTommotian D. regularis Zone in eastern Siberia (Moczydłowska andVidal, 1988) (Figs. 4, 5). Indeed, the eponymous species H. dissimilareof the overlying H.–S. Zone was later described from the D. regularisZone through the lower Atdabanian by Moczydłowska (1991; seespecies synonymy). Vidal and Moczydłowska (1992) and Vidal et al.(1995) also documented H.–S. Zone acritarchs from the D. regularis Zone.
Palacios et al. (2011) used theMoczydłowska and Vidal (1988) reportto correlate the S.–F. Zone acritatchs from theW. crosbyi Zone on HanfordBrook into the upper part (middle Tommotian) of the range ofWatsonella–Aldanella assemblages. This miscorrelation of the S.–F. Zonewith the middle Tommotian–lower Atdabanian equivalent Avalonian S.sunnaginicus and C. baltica zones was repeated by Moczydłowska andYin (2012), although small shelly fossil and carbon isotope correlationsequate these two Avalonian zones with the Tommotian–lowerAtdabanian (Landing et al., 1989; Brasier et al., 1992).
The correlation of theW. crosbyi Zone at Hanford Brook by Palacioset al. (2011) runs counter to the probability that the New Brunswickand Newfoundland W. crosbyi Zone assemblages are of broadly simi-lar age. This conclusion is based on the fact that the lowest W. crosbyiZone faunas lie just above and below the same sequence boundarywithin the Chapel Island Formation and below the wave-dominatedupper Chapel Island Formation and higher Random Formationtidalites in southeastern Newfoundland and southern New Brunswick(Landing, 2004) (Fig. 3).
The age and distribution of lithofacies are well known in Avalonia.The Tommotian–lower Atdabanian-equivalent S. imbricata–C. balticazones invariably form a mudstone-dominated succession that uncon-formably overlies, not underlies, a massive tidalite sandstone (the“Glen Falls” of Palacios et al., 2011). The top of the C. baltica Zone istypically a peritidal limestone (Fig. 4) which occurs high above Ran-dom Formation quartzites in a sea cliff to the east of Saint John insouthern New Brunswick (Landing, 1996b; Fig. 7, locality CBr).There are no good reasons for making the Saint John areastratigraphically different from all other Avalonian areas, includingan adjacent New Brunswick area, by having S. imbricata–C. balticazone fossils below the Random Formation. The synthesis in Palacioset al. (2011) conflicts with the small shelly fossil-, acritarch-, and car-bon isotope-based correlations discussed above.
Palacios et al. (2011) favour a revised, much younger correlationof the Rencontre–Random formations (their “Ratcliffe Brook” and
“Glen Falls” formations) by listing unillustrated acritarchs from the“type” W. crosbyi Zone in southeast Newfoundland. Their list includesformswith a long Lower Cambrian range in Baltica (i.e., Granomarginatasquamacea Volkova, 1968; G. prima Naumova, 1960; Lophosphaeridiumtentativum Volkova, 1968). The unillustrated report of lowest CambrianP. velata from the upper Chapel Island (Mystery Lake Member) inNewfoundland is of a form misidentified in New Brunswick and whichdoes not have its range defined outside of Baltica (discussed above).An important result in Palacios et al. (2011) is that probable S.–F. Zoneacritarchs occur above the sequence boundary in the Chapel IslandFormation and with “type” W. crosbyi Zone fossils in New Brunswick
9.4. Acritarch correlation of Avalonian lowest W. crosbyi Zone
Correlation of the lowest “type” W. crosbyi Zone is not resolvedby Palacios et al. (2011). Moczydłowska (1991, p. 25, 26) andMoczydłowska and Yin, 2012) note that Aldanella and Anabarella,which first appear in the lowest W. crosbyi Zone (Landing et al., 1989),also occur in the Lontova Stage of Baltica with A.–C. Zone acritarchs.The presence of A. attleborensis, reported as A. kunda (Öpik, 1926) andA. polonica Łendzion, 1977 (synonymies in Landing, 1988; Parkhaevand Karlova, 2011), in the Baltic A.–C. Zone suggests that the Avalonianlower W. crosbyi Zone extends into the A.–C. Zone (Figs. 3, 4).
10. Significance of 528 Ma volcanic ash in Cambrian Stage 2
Isachsen et al. (1994) reported a 530.7 ± 0.9 Ma volcanic ashfrom the Chapel Island Formation in the Somerset Street section inSaint John, New Brunswick (Fig. 7, locality SoS). This date wasrecalculated as 528.1 ± 0.9 Ma (Compston et al., 2008), but reportedas 530 Ma in Palacios et al. (2011). This is one of the few dated ashesabove the appearance of diverse skeletalised metazoans and belowthe lowest trilobites (Bowring and Martin, 1999; Maloof et al.,2010b).
Calibration by Maloof et al. (2005) of Lower Cambrian carbon iso-tope stratigraphy with dated ashes in Morocco indicates that thisChapel Island Formation ash is older than a calculated 525.4 Mabase of the Tommotian. In South China, a 526.5 ± 1.1 Ma date fromBed 9 (Compston et al., 2008) just above the unconformity that cutsout the W. crosbyi Assemblage Zone at Meishucun quarry (Li et al.,2009) is temporally indistinguishable from the New Brunswick age.This ash sets an upper age limit on the W. crosbyi Assemblage Zonein South China.
10.1. 528 Ma ash and pre-Tommotian skeletalised metazoans
The ca. 528 Ma ash provides an upper bracket on the underlyingW. crosbyi Zone and S.–F. Zone acritarchs on Hanford Brook. The528 Ma ash lies a short distance (ca. 8 m) beneath the faulted contactof the upper Chapel Island and Random formations at SomersetStreet. Although Palacios et al. (2011) proposed that much of theChapel Island Formation could be cut out at the fault, publisheddata show that little of the Chapel Island and Random is cut out. Infact, wave-deposited sandstones of the upper Chapel Island areinterbedded with coarse-grained, tidalite sandstones of the Randomat Somerset Street—an interbedding seen at unfaulted contacts ofthe formations in southern New Brunswick from Ratcliffe Brook toBeaver Harbour (Landing, 2004; Landing et al, 2008; Fig. 7, localitiesRBr and Bhr).
Palacios et al. (2011, p. 54) provide two interpretations of the ash.By the first, they re-interpret previous reports on the ash (Isachsen etal., 1994; Landing and Westrop, 1998a; Landing, 2004) and concludethat it lies not at the top, but low in the Chapel Island Formation (inthe Quaco Road Member), and could represent any ash within the for-mation. Thus, a 528 Ma date could be assigned to the lowest acritarchson Hanford Brook (Palacios et al., 2011), an interpretation followed by

152 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Moczydłowska and Yin (2012). Alternatively, Palacios et al. (2011,p. 54) reported upper Lower Cambrian H.–S. Zone acritarchs (Figs. 3,4) just across Somerset Street and 12 m above the ash, and refer thison-strike, lithologically identical section to the upper Chapel Island(Mystery Lake Member).
Resolution of Palacios et al.'s (2011) conflicting interpretationssolves a minor problem—the correct stratigraphic location of the528 Ma ash within the Chapel Island Formation. This correct strati-graphic position of the 528 Ma ash contributes to the geochronologyand biostratigraphy of Cambrian Stage 2 and Stage 2 of the CambrianRadiation.
The 528 Ma ash at 24.4 m on Somerset Street is a marker bed inthe New Brunswick Cambrian (Landing, 2004). It is the only annealed,purple ash with mm-sized white glass fragments in the Cambrian ofthe region—other Chapel Island Formation ashes are fine-grained,greenish, and chalky or cherty (E. Landing, unpub. data). Another dis-tinctive ash is a thick (15 cm) ash at 16 m on Somerset Street. Besidesthe adjacent Somerset and Gooderich Street localities (Fig. 4, locali-ties SoS and GoS), the 24.4 m and 16.0 m ashes occur near the topof the Mystery Lake Member type section in east Saint John, at145.4–145.6 m and 116 m, respectively (Fig. 7, locality MyL). At Mys-tery Lake, the ashes appear above the intra-Chapel Island sequenceboundary (Landing, 2004, p. 423). The purple ash lies 85 m abovethe sequence boundary in the Chapel Island Formation and 20 mbelow the Random Formation on Ratcliffe Brook 40 km east of SaintJohn (Fig. 7, locality RBr) (Landing, 2004). The ash likely occurs inthe covered interval between the Chapel Island and Random forma-tions on Hanford Brook.
Geochronological calibration of the earliest Cambrian (Maloof etal., 2005, 2010a,b) should have led Palacios et al. (2011) andMoczydłowska and Yin (2012) to conclude that a ca. 528 Ma ash ispre-Tommotian and, thus, not regard the upper Chapel Island Forma-tion as Tommotian or younger. Because the ash lies just below theRandom Formation, it occurs well above the W. crosbyi Zone and S.–F. Zone acritarchs just above the intra-Chapel Island sequence bound-ary on Hanford Brook.
The ca. 528 Ma ash provides an upper date for the early diversifi-cation of skeletalised metazoans, in particular micromollusks, and forthe base of Cambrian Stage 2. Indeed, the W. crosbyi Zone and tenta-tive S.–F. Zone acritarchs lie 160 m below the Random Formation atHanford Brook and well below the the 528 Ma ash. Since most ofthis interval consists of rapidly deposited, wave-dominated sand-stones on Hanford Brook (Landing and Westrop, 1998a; Landing,2005), the W. crosbyi and S.–F. zones may only be several millionyears older than the 528 Ma ash.
10.2. Acritarch correlation and the 528 Ma ash
No acritarchs that allow a highly resolved correlation are reportedbelow the ca. 528 Ma ash at the Somerset Street (NE and SW) sectionsby Palacios et al. (2011). Microfossils low in the Somerset Streetsection, and below the ash include fragmentary debris (nanoscalefilaments, cyanobacterial sheaths) and long-ranging acritarch genera(Comasphaeridium Staplin et al., 1965; Leiosphaeridia Eisenack, 1958),but species-level identifications are not possible. Two long-ranging spe-cies are present—A. tornatum (Volkova, 1968), which ranges throughthe Lower Cambrian in Baltica, and Ceratophyton vernicosum Kiryanovin Volkova et al. (1979), a probable microscopic metazoan (e.g.,Vanguestaine and Léonard, 2005) that ranges from the Baltic upper Edia-caran through the S.–F.Zone (Moczydłowska, 1991) and almost to the topof the Lower Cambrian by appearing in the Gislöv Formation in Sweden(Vidal, 1981) (Figs. 3, 4). P. velata is listed below the 528 Ma ash at Som-erset Street, and is the basis for Palacios et al. (2011) referring the ash tothe lowest Cambrian. However, their identification of this species isquestioned, and its biostratigraphic significance as a lowest Cambrianform is uncertain (discussed above). The lowest phytoplankton fossils
at Somerset Street can be interpreted as a lithofacies-defined assemblagecomparable to the lowest remains on Hanford Brook—dominantly poorlypreserved filaments and sheaths in relatively organic-rich, somewhatpyritiferous sedimentary rock (E. Landing, unpub. data) that forms theonlap facies of the Chapel Island Formation.
The 528 Ma ash lies about 12 m below H.–S. Zone acritarchs whichwere recovered just under the Random Formation (Palacios et al.,2011, fig. 2) (Figs. 3, 4). This relationship suggests that the 528 Maash lies within the H.–S. Zone. S. ciliosa (Volkova in Rozanov et al.,1969) from this upper Chapel Island Formation assemblage has anupper Lower Cambrian FAD (Moczydłowska, 1991; Moczydłowskaand Zang, 2006; S.G. Molyneux in Rushton et al., 2011). The reportof C. vernicosum from the lowest Chapel Island at Somerset Streetmay be consistent with correlation into the probable S.–F. Zone inthe lower Mystery Lake Member (upper Chapel Island Formation)on Hanford Brook, but C. vernicosum ranges almost to the top of theLower Cambrian in Sweden (Vidal, 1981). Thus, the ca. 528 Ma ashmay provide an age for the base of the H.–S. Zone.
H.–S. Zone acritarchs just below the Random Formation on SomersetStreet indicate the lower Randommay be assigned to this upper, but notuppermost, Lower Cambrian acritarch zone. Palacios et al. (2011) reportacritarchs froma sample “attributed” to the top of the Random in south-east Newfoundland. These acritarchs are assigned the S.–F. Zone, al-though all of the taxa listed (F. minuta, F. membranea, S. ornata, S.orbiculare, Comasphaeridium strigosum, C. brachyspinosum) persistthrough the H.–S. Zone in Baltica (Moczydłowska, 1991) and are notlimited to the S.–F. Zone.
The available evidence is consistent with correlation of the Ran-dom Formation, as well as the overlying Bonavista Group (discussedabove), into the H.–S. Zone (Figs. 3, 4). This correlation supports a uni-form formational succession (lower rift facies of the Rencontre–openmarine shelf of the Chapel Island with middle sequence boundary andW. crosbyi Zone faunas–upper tidalites of the Random) that extendsfor 650 km from southeastern Newfoundland to southwest NewBrunswick (Landing et al., 2008).
Palacios et al. (2011) show that the upper Mystery Lake Memberat Somerset Street has H.–S. Zone acritarchs and is younger than theW. crosbyi Zone with its S.–F. Zone acritarchs near the base of theMystery Lake Member at Hanford Brook. The 32 m thickness of theMystery Lake at Somerset Street NE (Landing and Westrop, 1998b)is less than the ca. 41 m assigned by Palacios et al. (2011) to their“Ratcliffe Brook Formation.” Palacios et al. (2011) apparently included10.1 m of volcaniclastic sandstone, arkose, and rhyolite pebble con-glomerate at the base of their section that are best refered to themuch older Rencontre Formation. The Quaco Road Member of theChapel Island Formation (Fig. 3) is missing on Somerset Street(Landing and Westrop, 1998a, fig. 21; Landing, 2004). Its absence ismarked by a yellowish horizon on the Rencontre Formation thatPatel (1977) recognised as a weathering horizon.
11. “Laolinian Stage” of the upper Terreneuvian Series
11.1. Definition of concept and GSSP
Landing and Geyer (2012) suggested that a candidate for the Cam-brian Stage 2 basal GSSP is in the road cut near Laolin village, Yunnan,South China. They further suggested that a “Laolinian Stage” with itsbase defined by the L4 carbon excursion peak in the Laolin road cutand within the local range of W. crosbyi could replace Stage 2 of theCambrian. Of course, it is the role of the International CambrianSubcommission to determine the type of marker horizon that willbe used to define the base of chronostratigraphic units and to reachagreement on the actual GSSP locality. However, the informal Cam-brian chronostratigraphic divisions and their definition suggestedherein are comparable to Peng and Babcock's (2011, fig. 1) proposaland publication, as officers of the Cambrian Subcommission, that

153E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
the FAD of W. crosbyi is a “preferred base” for Stage 2. Though nevervoted on, their proposal, as ours, provides a basis for discussion ofCambrian chronostratigraphy.
Li et al. (2009) closely examined the well known Laolin road cut forits chemo- and biostratigraphy and correlated the peak of its L4 carbonexcursion with what they designated the as the P4 peak at the Xiaotanquarry and with coeval carbon isotope peaks across South China. Fur-thermore, Li et al. (2009) correlated both the L4 and P4 peaks with theSiberian I′ excursion. As noted above, the report of characteristic W.crosbyi Zone taxa low in the I′ excursion support a precise correlation be-tween these two palaeocontinents. The significance for intra- and inter-continental correlation of a strong positive peak within the lower rangeofW. crosbyi-bearing faunas was earlier emphasised by Zhu et al. (2004),who designated this as the ZHUCE excursion. In short, the positive excur-sion peaks designated as “L4” and “P4” and the peak of the ZHUCE excur-sion are all synonymous chemostratigraphic events in South China. TheChinese and Siberian faunas associated with L4/P4/peak ZHUCE/I′ werediscussed above as comparable taxonomically and approximately coevalwith the “type” W. crosbyi Zone of southeast Newfoundland. Furtherwork on the carbon isotope stratigraphy of the Newfoundland sectionwill likely strengthen the correlation of these sub-TommotianW. crosbyiZone faunas between three Cambrian palaeocontinents.
The suggested “Laolinian Stage” (previously, informal CambrianStage 2) is a globally extensive chronostratigraphic unit that com-prises the second and upper stage of the Terreneuvian Series. TheGSSP for the “Laolinian Stage” base is herein defined in China by thepeak of the L4 carbon isotope excursion within the W. crosbyi Assem-blage Zone and within the upper Dahai Member at a roadside sectionca. 800 m SSE of Laolin village, Huize County, eastern Yunnan Prov-ince, China (Figs. 5, 7). This peak value of ca. +3.47 δ13C (‰) lies at174.6 m in the measured roadside section. The Laolinian Stage GSSPlies some 9.4 m below the top of the Dahai Member (Li et al., 2009,table 1). The term “Laolinian Stage” comes from the location of theroadcut near Laolin village, and utilises a place name that has not pre-viously been used for any stratigraphic unit.
The Laolin road cut lies about 200 kmnorth of the classicMeishucunquarry section, which is near Jinning village and also in Huize County.The location and stratigraphy of the Laolin section is well knownas it was proposed as a parastratotype section for the Precambrian–Cambrian boundary. Indeed, Qian et al. (2002) strongly emphasisedthat this section is superb as a parastratotype as it is continuous, wellexposed, stratigraphically complete, structurally uncomplicated, andlisted its faunas. Subsequently, Li et al. (2009) completed a detailedrestudy of its terminal Edicaran–lowest Cambrian carbon isotope stra-tigraphy in the context of its biotic succession and pointed out a “crack-led” fault zone well below the base of the W. crosbyi Assemblage Zoneand even further below the L4 peak, but did not indicate any strati-graphic throw. Over time, the Laolin roadcut has become a standardfor carbon isotope stratigraphy in China and for global correlation(Shen and Schidlowski, 2000; Li et al., 2009, 2012). The South ChinaCambrian sections are very condensed by comparison with the muchthicker (but slowly deposited) Avalonian successions (Landing et al.,2013b). However, the presence of the globally recogniseable basal“Laolinian Stage” carbon excursion peak (L4/P4/ZHUCE) as a suggestedbase for Stage 2, and the location of this carbon isotope positive excur-sion peak within the lower W. crosbyi Assemblage Zone, makes thisSouth China section appropriate as a candidate GSSP for the base ofCambrian Stage 2. The similar, thicker section at Xiaotan, 200 kmnorth of Laolin, does not allow clear distinction of the A. trisulcatus–P.anabarica and Siphogonuchites triangularis–Paragloborilus subglobosuszones below the W. crosbyi Assemblage Zone and higher L4 excursionbut could serve as a parastratotype GSSP section (Li et al., 2009,2012). The traditional Meishucun section 200 km south of Laolin is in-appropriate as a stratotype for the base of Stage 2 as an unconformitycuts out the W. crosbyi Assemblage Zone at Meishucun (Li et al., 2009,2012).
11.2. Metazoan fossils and a basal GSSP of the “Laolinian Stage”
As detailed above, the proposed use of the W. crosbyi or A.attleborensis FADs to define the base of informal Cambrian Stage 2(Parkhaev and Karlova, 2011; Parkhaev et al., 2011; Peng andBabcock, 2011; Steiner et al., 2011) draws, in part, on the worldwidedistribution of both species (Fig. 6). A caveat to the use of either theW. crosbyi or A. attleborensis FAD to define a Stage 2 base reflectsthe fact that both species have a great stratigraphic range inAvalonia—which brackets an interval coeval with the Siberian upperManykaian/“Nemakit-Daldynian”–lower Atdabanian stages. Perhapsas a reflection of slow dispersal, preservational bias, and/or unconfor-mities, the FAD and range of both W. crosbyi and A. attleborensis arediachronous between Cambrian regions and may locally representonly a fraction of their Avalonian composite range.
Thus, the two species appear relatively early in Avalonia and SouthChina and apparently the Montagne Noire; higher in Atdabanian-equivalent rocks in South Australia and Mongolia; and only recentlyhave been found to co-occur at levels apparently comparable to theirlower Avalonian range in northern Siberia. A second caveat to the use ofthe FADs of W. crosbyi and A. attleborensis to define the GSSP for thebase of Stage 2 is that there is no evidence that precludes the possibilitythat the FADs of both species, which appear with shoaling at the LittleDanzig Cove section (Landing et al., 1989), might actually be significantlyolder than the species FADs in South China and northern Siberia. Withinsmall areas, preservational bias can cause the essentially coterminousFADs of the two species to differ—by 50 m along depositional strike insoutheasternNewfoundland. A combinationof preservational bias and in-hospitable faciesmeans theW. crosbyiZonebase is above the intra-ChapelIsland Formation sequence boundary and in the S.–F. Zone (acritarchs) insouthern New Brunswick, but about 50 m below this sequence boundaryand possibly in the A.–C. Zone in southeastern Newfoundland (Landing etal., 1989; Landing, 2004).
The reliability of these micromollusks for correlation is questionableeven in South China where only a few sections have the W. crosbyi–A.attleborensis–O. korobkovi association that defines the W. crosbyi Assem-blage Zone. This Chinese “assemblage zone” is actually the W. crosbyirange zone, but with the W. crosbyi FAD coincident with a lithofacieschange from dolostone to limestone in the Dahai Member (Fig. 5). Thus,the South China and Montagne Noire (discussed above) successionsshow that aW. crosbyi FAD is also unreliable for use in chronostratigraphicdefinition. In conclusion, all of the problems that confound use of a singlespecies FAD for correlation and definition of chronostratigraphic units areassociated withW. crosbyi and A. attleborensis (Fig. 1).
11.3. Acritarch-based definition for the base of Cambrian Stage 2?
The potential of acritarchs for global correlation is shown by theoccurrence of such forms as S. ciliosa on a number of Cambrianpalaeocontinents (Moczydłowska and Zang, 2006). As the W. crosbyiand A. attleborensis FADs are diachronous, an acritarch-based defini-tion for the base of Stage 2 might provide an alternative.
The local base of the S. ornata–F. membranacea (S.–F.) Zone lieswell below the lowest trilobites and with a W. crosbyi Zone microfau-na in a cool-water succession in Avalonian New Brunswick (Palacioset al., 2011). The S.–F. Zone base is also below the lowest trilobitesin tropical East Gondwana in the Wilkawillina Limestone of SouthAustralia (Moczydłowska and Zang, 2006).
S.–F. Zone acritarchs appear in Baltica in the SwedishHardeberga For-mation and correlative units. The conundrum for an S.–F. Zone-basedglobal correlation is that the Hardeberga has poorly preserved holmiidtrilobites (Ahlberg and Bergström, 1998) (Fig. 4), whereas the S.–F.and H.–S. zones in New Brunswick are definitely sub-trilobitic. A ca.528 Ma date on the lowest H.–S. Zone (or S.–F. to H.–S. zone boundary)in New Brunswick is definitely pre-trilobitic in terms of Cambrian

154 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
geochronology, with the calculated base of the lowest trilobite-bearingAtdabanian Stage at 521.5 Ma (Maloof et al., 2005, 2010a,b).
That S.–F. Zone acritarchs persist higher into the range of morpho-logically derived Schmidtiellus Zone trilobites in Baltica and withlower P. huoi Zone trilobites in South Australia is not presently explain-able (see Moczydłowska, 1991, 1998; Ahlberg and Bergström, 1998;Moczydłowska and Zang, 2006; Moczydłowska and Yin, 2012). Asdiscussed below, the latter Baltic and Australian trilobites are probablyAtdabanian or younger in age (Figs. 3, 4). It is consequently confusingthat “younger” H.–S. Zone acritarchs occur in the middle TommotianD. regularis Zone–lower Atdabanian (Moczydłowska, 1991; Vidalet al., 1995). Thus, a Cambrian Stage 2 base defined by S.–F. or H.–C.zone acritarchs would be problematical.
11.4. Carbon isotopes and chronostratigraphy
Uncertainties about the true time ranges of micromollusks andacritarchs emphasise that a simple biostratigraphic (FAD) approachshould not be considered for definition of the Stage 2 base. The bestdefinition for a globally correlatable GSSP is a combination of asub-trilobitic carbon isotope excursion within a biotic assemblagezone. The utility of Early Cambrian carbon isotope excursions forglobal correlation has long been appreciated with the peak excursionsof carbon excursions being used for highly resolved between the low-est Cambrian successions of Morocco, Siberia, Mongolia, South China,and Avalonia (e.g., Brasier et al., 1992, 1994a–c; Maloof et al., 2005,2010a,b; Li et al., 2009, 2012).
A combined carbon isotope and faunal definition of the GSSP for thebase of Stage 2 uses the correlation potential of the Chinese L4/P4/peakZuche excursion and Siberian I′ excursions within the W. crosbyi Zone.This definition will allow the base of the suggested “Laolinian Stage” tobe correlated by carbon isotope stratigraphy into unfossiliferous strata.
Acritarchs show that the Avalonian upperW. crosbyi Zone tentative-ly lies in the lower S.–F. Zone, while keyW. crosbyi Zone taxa appear inthe underlying A.–C. Zone in Baltica. Thus, the lower S.–F. Zone mightserve as a proxy for the base of the Laolinian Stage in successionswhere its micromollusks and distinctive carbon isotope excursion arenot present. As the W. crosbyi Zone and lower S.–F. Zone are olderthan 528 Ma, this suggests that the base of the “Laolinian Stage” liesat the strong positive carbon isotope excursion at ca. 531 Ma associatedwith the Cambrian mollusk diversification (Maloof et al., 2010a,b).
Use of the L4/P4/peak ZHUCE and I′ carbon isotope peaks forinterregional correlation has a long history. Brasier et al. (1990,1996) concluded that a strong positive carbon isotope peak at aboutthe lowest occurrence of diverse micromollusk assemblages can betraced across South China (“Marker B”), and had potential for correla-tion with positive carbon isotope peaks in the lowest Cambrian ofnorthern Iran, Siberia, Mongolia, Lesser Himalaya, India, Morocco,and South Australia. Brasier et al.'s (1996) correlations also supportedconclusion that the oldest diverse small shelly assemblages aresub-Tommotian (Landing, 1988, 1992, 1996a; Landing et al., 1989),and not, as regularly repeated in the literature, Tommotian orTommotian-equivalent (e.g., Steiner et al., 2007; Rozanov et al.,2008; Li et al., 2011; Moczydłowska and Yin, 2012).
A GSSP based primarily on a carbon isotope excursion has a strongprecedent—a strong negative δ13C excursion is the datum that definesthe GSSP of the base of the Ediacaran System in South Australia andallows its worldwide correlation (Knoll et al., 2004). The suggested“Laolinian Stage” builds firmly on this established procedure.
11.5. Biostratigraphic and carbon isotope brackets on the “LaolinianStage” base
The strength of the proposed definition of the “Laolinian Stage”GSSP lies in the multiple brackets that can be used to recognise thesuggested stage in successions worldwide. These include the much
older BACE excursion and lowest A.–C. Zone acritarchs in the terminalEdiacaran, the refined base of the T. pedum Assemblage Zone at thebase of the Cambrian, and the somewhat older appearance of diversemineralised metazoans, including micromollusks of the W. crosbyiZone, of stage 2 of the Cambrian Evolutionary Radiation.
Sub-“Laolinian Stage” developments include the appearance oflow diversity mineralised metazoan fossils with a potential for zonalbiostratigraphy in the terminal Ediacaran and lowest Cambrian(Zhuravlev et al., 2012), and the higher appearance of diverseskeletalised metazoans, including those of the lower W. crosbyiZone. The lower “Laolinian Stage” may be close to the base of theS.–F. Zone. Higher “Laolinian Stage” strata have Tommotian orTommotian-equivalent microfaunas and the lowest archaeocyathans.As Tommotian and Tommotian-equivalent rocks lie below the lowestoccurrence of trilobites in Siberia and the trilobite-bearing upperLower Cambrian rocks typically assigned to Series 2 (e.g., Zhu et al.,2005, 2006), the Tommotian correlates into the upper “LaolinianStage” of the upper Terreneuvian Series.
12. Third stage of the Cambrian Evolutionary Radiation—appearanceof trilobites
12.1. Terreneuvian trilobites?
The top of the Terreneuvian is defined by the base of informalupper Lower Cambrian Series 2 and lowest Stage 3. Much enthusiasmhas been expressed for defining the base of Series 2 at the lowest oc-currence of trilobites, which seemingly would preclude their occur-rence in Stage 2 of the upper Terreneuvian (e.g., Zhu et al., 2006;Babcock and Peng, 2007). A seeming problem here is that acritarch-based correlations could be used to argue, incorrectly as discussedabove, that trilobite fossils actually have their lowest occurrence, per-haps in the lower “Laolinian Stage” of the Terreneuvian, on severalpalaeocontinents (Moczydłowska and Vidal, 1988; Moczydłowskaand Yin, 2012) (Fig. 3, caption).
A sub-Series 2 first appearance of mineralised trilobites cannot beruled out. Indeed, the transition between Lower Cambrian chrono-stratigraphic units, such as the gradual increase in diversity in earlymineralised fossils and the greater complexity of trace fossils throughthe Ediacaran–Terreneuvian boundary (e.g., Jensen et al., 2000;Zhuravlev et al., 2012), is not necessarily accompanied by abruptfirst-appearances of higher-level taxa, such as that represented by tri-lobites. An obvious exception is seen at the base of the TommotianStage in southeast Siberia, and involves truncation of the lowerranges of high-level taxa at an unconformity. Without further evi-dence and without the definition of a formalised replacement for in-formal Series 2, the possibility exists that trilobites appeared in theterminal Terreneuvian. As proposed below, the best definition forthe GSSP of a formally defined replacement for informal Series 2 liessomewhat above the oldest Siberian trilobites—which means thatmineralised trilobites appeared, by the definition proposed herein,in the terminal Terreneuvian.
12.2. Taxonomic and community evolution, lithofacies, and trilobite FADs
The use of a trilobitic FAD for the Series 2 base provides a classicexample of the problems integral to the FAD concept (Fig. 1). Generalagreement has long existed that trilobites do not have a uniform FAD,even in stratigraphically unbroken sections (e.g., Brasier, 1989, fig7.16–7.19). The appearance of mineralised trilobites marks themost dramatic event in the modernisation of ecologic communitiesin stage 3 of the Cambrian Radiation (Landing and Westrop, 2004).Disagreement exists whether the appearance and earliest radiationof trilobites with mineralised carapaces took place in more off-shore shelf environments and that coeval peritidal environmentsinitially had very sparse trilobites (Landing et al., 1989; Landing

155E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
and Westrop, 2004) or whether nearshore shelf habitats featuredrapid diversification of the earliest trilobites accompanied by anunusual morphological plasticity in the earliest trilobite assem-blages (e.g., Geyer, 1996).
This “disagreement” may revolve around the fact that theperitidal, nearly trilobite-free Avalonian facies described by Landingand Westrop (2004) were actually intertidal and very close toshore, whereas the Moroccan facies described by Geyer (1996) werelittoral and more comparable to the more offshore locus of trilobitediversification proposed by Landing and Westrop (2004). Regardlessof the evolutionary history, strong lithofacies–biofacies controlsexisted on the distribution of the lowest local trilobites, and thesecontrols on their lowest occurrence are compounded by the high pro-vincialism of the oldest trilobite faunas.
Palmer and Repina (1993) outlined a relatively nuanced, traditional(non-cladistic) interpretation of ancestor–descendent relationships ofEarly Cambrian trilobite families. They suggested diachronous lowesttrilobite occurrences onCambrian palaeocontinents—with lowdiversityfallotapidids with temporally staggered, first appearances appearingearly on the tropical Siberian Platform and Moroccan margin ofGondwana. So-called “fallotaspidids” appeared later in tropical westernLaurentia. Even later-appearing assemblages with descendent holmiidscomprise the oldest trilobite faunas in the cool-water successions ofAvalonia and Baltica by this synthesis (Fig. 2). Palmer and Repina(1993) suggested that some Cambrian palaeocontinents or regionscould have remained sub- and pre-trilobitic and in “Series 1” whileother regions could have had mineralised trilobites and been in “Series2” at the same time. This clearly suggests that the global “trilobite FAD”is amyth. Provincialism in the earliest trilobite faunas contributes to un-certainty in interregional correlation, as do facies controls. Indeed, thelowest trilobites often appear at facies changes and above unconfor-mities (Figs. 4, 9).
A different approach was taken by Yuan et al. (2011) who essen-tially lined up the oldest trilobite horizons in South China, Siberia,Gondwana (south Morocco and Australia), and western Laurentiaand made them into a single trilobite FAD. Such an approach takesno account of independent biostratigraphic evidence (small shellyfossils, acritarchs), carbon isotope stratigraphy and geochronology,alternative models of trilobite evolution, or of stratigraphic breaksmarked by lithofacies changes or unconformities of the type neededto evaluate the temporal significance of the lowest local trilobites.
13. Earliest trilobites
13.1. Baltica
The Lower Cambrian biostratigraphy of Baltica is essentially a com-bination of acritarch and trilobite zones (Fig. 4). The trilobite zones re-flect local or regional occurrences of trilobites and have undergonegreat modifications in successive reports (Bergström, 1981; Ahlberg,1984; Bergström and Gee, 1985; Ahlberg et al., 1986; Moczydłowska,1991; Ebbestadt et al., 2003; Nielsen and Schovsbo, 2011). The instabil-ity in trilobite zonation reflects the facies dependence of the trilobiteassemblages, while the predominantly coarse-grained siliciclastic fa-cies make fossils rare in most Baltic sections. The trilobite zones ofBaltica are largely based on the Scandinavian succession; the acritarchzones are defined in Polish and southeast Baltic sections (e.g.,Moczydłowska and Vidal, 1986; Hagenfeldt, 1989; Moczydłowska,1991, 1998). Based on a new synthesis of depositional environmentsand sequence stratigraphy of the Scandinavian Lower Cambrian,Nielsen and Schovsbo (2011) modified the trilobite zonation into thesuccessive Schmidtiellus mickwitzi, Holmia inusitata, H. kjerulfi, and‘Ornamentaspis’ linnarssoni zones (Fig. 4).
The oldest identifiable trilobites of Baltica are in the S.mickwitziZone.The zonehas a lowdiversity faunawith S.mickwitzi (Schmidt, 1888) (in-cluding two subspecies); S. reetaeBergström, 1973;Wanneria? lundgreni
(Moberg, 1892); and Holmia mobergi Bergström, 1973. This fauna isknown from the Norretorp Sandstone (Norretorp Member of the LaesaFormation) in Scania, southern Sweden (Moberg, 1892, 1899;Bergström, 1973), and the Lükati Formation of Estonia (Ahlberg et al.,1986; Mens and Pirrus, 1997).
All of the species and clearly distinguishable genera of the S.mickwitzi Zone are limited to Baltica, and correlation betweenpalaeocontinents based on these trilobites is impossible. Associatedshelly fossils are of little help. Proposed correlations have been basedon S. ornata–F. membranacea Zone acritarchs from the S. mickwitziZone (Moczydłowska and Vidal, 1986). However, this acritarch-basedcorrelation equates the S. mickwitzi assemblage with strata so fardown in the subtrilobitic Cambrian on other palaeocontinents (dis-cussed above, Fig. 3) that these trilobites would be the oldest knownon Earth, even though Schmidtiellus shows many derived charactersand is not an “early” trilobite (Palmer and Repina, 1993). Thus, thisacritarch-based correlation seems highly questionable.
The S. mickwitzi Zone is underlain by an informal Rusophycus Zonecharacterised by the eponymous ichnofossil (Figs. 4, 9). Whether thistrace was made by a trilobite is unclear, but a poorly preserved trilo-bite thorax is known from this zone (Ahlberg et al., 1986).
A Homia inusitata Zone above the S. mickwitzi Zone was proposedby Ahlberg et al. (1986). However, H. inusitata Ahlberg and Bergströmin Ahlberg et al. (1986) is known from only one section in the MjøsaDistrict, southern Norway. Nielsen and Schovsbo (2011) show thatsequence stratigraphic correlations indicate these strata are notolder than the younger H. kjerulfi-bearing strata at Mjøsa (Fig. 4).Thus, they suggested abandoning the H. inusitata Zone. Their expand-ed H. kjerulfi Zone corresponds to the lower Vergalian–Rausvian Stageof the Baltic region. The trilobites of this zone were primarily recov-ered from Scania and the Mjøsa District and have taxonomic affinitieswhich place them into the later part of the trilobite-bearing LowerCambrian. Nielsen and Schovsbo's (2011) revisions, which combinethe H. kjerulfi and ‘O.’ linnarssoni zones, put very late Early Cambrianellipsocephalids known from isolated occurrences into H. kjerulfi-bearing strata. Thus, it is puzzling that Holmia could range nearlythrough the entire trilobite-bearing Early Cambrian of Baltica.
13.2. West Gondwana—southern Morocco
The key area for the earliest trilobites ofWest Gondwana is the Atlasranges ofMorocco (Sdzuy, 1978; Geyer and Landing, 1995, 2006; Geyer,1996). The revised zonation for the trilobite-bearing Lower–lowerMid-dle Cambrian in this region is useful across the Mediterranean region,and is the basis for a biostratigraphic standard for the Cambrian ofWest Gondwana (Geyer and Landing, 2004). The early trilobite recorddefines the Issendalenian Stage (Geyer, 1990). This stage consists of avertical sequence of the Eofallotaspis, Fallotaspis tazemmourtensis,Choubertella, and Daguinaspis zones (Fig. 4).
The lowest described trilobites of the Eofallotaspis Zone are knownfrom a section near Tiout, western Anti-Atlas, but a similar pattern ofappearance is known in such nearby sections as Tazemmourt (K.Sdzuy, unpub. data; Geyer and Landing, 1995, 2006). The oldest deter-minable trilobites at Tiout are in the upper Igoudine Formation, and ap-pear just above a facies change into less restricted marine habitats(Fig. 9). Lower, light and dark coloured, thick-bedded, micritic and do-lomitic limestones with large chert nodules and stromatolites areoverlain by trilobite-bearing, dark, oolitic, oncoid- or intraclast-and archaeocyathan-bearing limestones with variably developedolive-coloured, quartzose interbeds. This FADdoes not reflect the evolu-tionary origin of trilobites as trilobite fragments are found in the lowermember of the Igoudine Formation (Sdzuy and Geyer, 1988; Geyer andLanding, 1995). Indeed, fragments of probable trilobite sclerites occureven lower in carbonates near the diachronous top of the Lie de vinFormation (Geyer and Landing, 1995, 2006). These fragments arenot generically determinable, but may represent poorly preserved

156 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
remains of species of the Eofallotaspis assemblage of the upper IgoudineFormation.
Most of the lowest Moroccan trilobites are primitive bigotinoids(Sdzuy, 1978, 1981; Geyer and Landing, 2006). Only one species ofthis taxonomic group has been described—Hupetina antiqua Sdzuy,1978, from the lowest horizon of the Igoudine Formation at Tioutwith determinable trilobite remains. Bigotina sp. is known from aslightly higher horizon. Small fallotaspidids occur and include severalspecies of Eofallotaspis Sdzuy, 1978. Material from higher horizonswas collected by K. Sdzuy, but only E. tioutensis Sdzuy, 1978, and E.prima Sdzuy, 1978, have been described.
The Eofallotaspis Zone faunas with Eofallotaspis and Hupetina comefrom carbonate facies. The overlying F. tazemmourtensis throughDaguinaspis zones (Hupé, 1953) are largely based on faunas fromshales (Geyer, 1990; Geyer and Landing, 1995, 2006) (Fig. 4). Fossils,including archaeocyathans (Debrenne and Debrenne, 1978, 1995),from associated limestone beds expand the biostratigraphically sig-nificant faunal elements.
The base of the F. tazemmourtensis Zone, and its oldest Fallotaspisspecies, is in the uppermost Igoudine Formation in the Tiout section.This zone is usually thin and has a low diversity faunawith Pararedlichiapulchella Hupé, 1953; P. rochi Hupé, 1953; P. subtransversa Hupé, 1953;and others. Eofallotaspis overlaps the range of F. tazemmourtensis.Lemdadella tioutensis Sdzuy in Liñán and Sdzuy (1978) is assigned tothe zone and occurs in thin limestone beds just above the base of theoverlying, shaly Amouslek Formation.
The F. tazemmourtensis Zone is overlain by the Choubertella Zone,which is characterised by Choubertella ambroggii Hupé, 1953; C.spinosa Hupé, 1953; and C. latifrons Hupé, 1953. Associated trilobitesinclude Fallotaspis bondoni (Neltner and Poctey, 1950); F. planaHupé, 1953; Bigotinops dangeardi Hupé, 1953; and other species.
The overlying Daguinaspis Zone has locally abundant fossils withsuch trilobites as Daguinaspis ambroggii Hupé and Abadie, 1950; D.abadiei Hupé, 1953; D. subabadiei Hupé, 1953; and D. crassioculataHupé, 1953. Other trilobites include Fallotaspis planospinosa Hupé,1953; F. bondoni (Neltner and Poctey, 1950); Resserops falloti Hupé,1953; R. brevilimbatus Hupé, 1953; R. uncioculatus Hupé, 1953; R.bourgini Hupé, 1953; Marsaisia robauxi Hupé, 1953; M. parvifronsHupé, 1953; and many others.
Fallotaspis is not limited to the F. tazemmourtensis–Daguinaspiszones as Hupé (1953) claimed. It is common in the higher Antatlasiahollardi Zone (Fig. 4) and also occurs above that zone (Geyer, 1996;Geyer and Landing, 2006). The genotype F. typica Hupé, 1953, camefrom the A. hollardi Zone (Hupé, 1953; Geyer, 1996). Thus, theso-called “Fallotaspis zone” or “Fallotaspis stage” (Fritz, 1972;Repina, 1986) is imprecisely defined and cannot be applied on an in-tercontinental scale.
13.3. Iberia and Normandy
The oldest determinable trilobites from Iberia occur in the PedrocheFormation in the Sierra de Córdoba, southern Spain (e.g., Liñán Guijarro,1978). They include, in ascending stratigraphic order, the redlichioidtrilobites Bigotina bivallata Cobbold, 1935b; Lemdadella linaresae Liñánand Sdzuy, 1978; L. perejoni Liñán et al., 2005; and Pararedlichia cf.ovetensis Sdzuy in Liñán and Sdzuy (1978) (recently termed Eoredlichiacf. ovetensis). These species define the B. bivallata, L. linaresae, and L.perejoni zones plus “levels with Eoredlichia” (Liñán et al., 2005, 2006)(Fig. 4). It should be noted that Liñán et al. (2005) synonymisedPararedlichia with Eoredlichia but kept the genus in a subfamilyPararedlichiinae.
Serrania verae Liñán, 1978, from the Pedroche Formation can bereassigned, based on data in its original description, to the L. linaresaeZone (Liñán et al., 2005). However, discrepancies exist in the avail-able information, and S. veraei likely was collected from Perejón's(1986, 1994) archaeocyathan zone I and close to the base of the
Ovetian Stage of Iberia (= Issendalenian Stage of Fig. 4) as indicatedby data in Liñán et al. (2006). The B. bivallata Zone, by contrast, coin-cides more or less with Perejón's (1986, 1994) archaeocyathan zoneIII. Bigotinidae resembling Bigotinella were reported from older strataand even from the upper part of the supposedly sub-trilobiticCorduban Stage (Liñán et al., 2005).
These data indicate that the top of the traditional IberianCorduban Stage does not correspond to the top of the Corduban Seriesof the West Gondwanan standard. The top of the Iberian Cordubiancorrelates into the lowest part of the trilobite-bearing IssendalenianStage. Furthermore, the traditional Ovetian Stage of Iberia corre-sponds to the middle–upper Issendalenian of the West Gondwananstandard (Fig. 4; Geyer and Landing, 2004, fig. 2). The earliest trilo-bites from southern Iberia are probably approximately coeval withthe oldest Morocco trilobites. However, the known occurrences andpreservation of the Iberian trilobites are considered inadequate forreliable taxonomy and correlation (Geyer and Landing, 2004).
B. bivallata is based on a small collection from the shore outcrop inthe Saint-Jean-de-la-Rivière Formation at Carteret, Normandy. Thespecies' range at Carteret (Pillola et al., 1994) appears to be longerthan in southern Spain. However, other trilobites of comparable ageare not known from Normandy.
13.4. Avalonia
The earliest determinable trilobites in Avalonia include theolenelloid C. broeggeri (Walcott, 1890) from southeast Newfoundland,Massachusetts, and England. The species appears across Avalonia insiliciclastics or condensed, thin carbonates above a major unconformitythat overlies a lower Atdabanian-equivalent limestone that lacks trilo-bites (Brasier et al., 1978; Landing, 1988; Landing et al., 1989, 2013b;Brasier et al., 1994a; McIlroy et al., 1998; Landing and Westrop, 2004;Fletcher, 2006) (Figs. 3–5, 9). In southeast Newfoundland, the C.broeggeri Zone has C. broeggeri and Acanthomicmacca walcotti(Matthew, 1899) occurring through the zone. Fletcher (2006) nameda lower A. walcotti Subzone with C. broeggeri and Comluella sp. fromthe lowest limestone beds of the St. Mary's Member of the Brigus For-mation (Fig. 3) [= “Broad Cove Member” of Fletcher's (2006) nomen-clatural scheme for the Brigus Formation, and which is a juniorsynonyn of the basal limestone of Landing and Westrop's (1998b) St.Mary's Member, see Westrop and Landing, 2011)].
A “Dipharus attleborensis Subzone” is named from southeasternNewfoundland (Fletcher, 2006; Harvey et al., 2011). Several problemsexist with this supposed upper part of the C. broeggeri Zone. Thefirst is that D. attleborensis (Shaler and Foerste, 1888) is a nomendubium described from eastern Massachusetts, and which is notrecogniseable in Avalonian Newfoundland or Britain (Westrop andLanding, 2011). Secondly, forms comparable to those which Fletcher(2006) and Fletcher and Theokritoff (2008) called D. attleborensis insoutheast Newfoundland occur within 3 m of the base of the BrigusFormation, and are not an element of an upper subzone of the C.broeggeri Zone (Westrop and Landing, 2011).
More diverse taxa occur in the higher C. broeggeri Zone. They includethe eodiscoids “D. attleborensis,” Delgadella plana (Hutchinson, 1962),Calodiscus lobatus (Hall, 1847), C. meeki (Ford, 1876), C. schucherti(Matthew, 1896), Weymouthia nobilis (Ford, 1872), Acimetopus helena(Walcott, 1889), Mallagnostus? llarenai (Richter and Richter, 1941),Serrodiscus bellimarginatus (Shaler and Foerste, 1888), Triangulaspismeglitzkii (von Toll, 1899), and T. vigilans (Matthew, 1889). Non-eodiscoids include “Strenuaeva” cf. spinosa (Ahlberg and Bergström,1978), Strenuella strenua (Billings, 1874), Comluella pustulosa (Cobbold,1910), Pseudatops reticulatus (Walcott, 1890) (Hutchinson, 1962;Fletcher, 2006; Westrop and Landing, 2011).
The earliest English Avalonian trilobite zone is the traditionalCallavia Zone with C. broeggeri (Walcott, 1890) and “D. attleborensis”in the very thin Green and Red Callavia limestones of the Comley

157E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
area (Fig. 3). This English zone has been correlated with the upper C.broeggeri Zone in southeast Newfoundland (Fletcher, 2006; Harvey etal., 2011), but, as discussed above, this correlation is based on the incor-rect assumption that “D. attleborensis” is an upper C. broeggeri Zone ele-ment in North American Avalon. Thus, the English Callavia Zone andNorth American lower C. broeggeri Zone are biostratigraphically indistin-guishable at present. Apparent Callavia sclerites occur with the lowesttrilobites ca. 1.5 m above the base of the Purley Formation in Warwick-shire (Brasier, 1984). Rushton (1966) described faunas with S.bellimarginatus and M.? llarenai higher in the Purley Formation (Fig. 3).
A cranidium from the middle of the Lower Comley Sandstone(unit Ac3) below the Green and Red Callavia sandstones is oftenregarded as the oldest trilobite fossil in British Avalonia (Fig. 3).Harvey et al. (2011) used the fragment to define a “Fallotaspis Zone”in Avalonia, although the fragment is not identifiable even at thegenus level (e.g., Bergström, 1973). It cannot be used to define a“Fallotaspis” Zone, let alone indicate that unit Ac3 is older than thelower C. broeggeri Zone (Landing et al., 2013b).
Callavia faunas are relatively late among early trilobites, and arecorrelated with the traditional Nevadella Zone in Laurentia (e.g.,Palmer and Repina, 1993), although no direct evidence for correlationcan be based on the paraendemic Avalonian trilobites. The “D.attleborensis” Subzone of Newfoundland and the Purley Formationof England is dominated by such eodiscoids as “D. attleborensis,”Calodiscus spp., M.? llarenai, S. bellimarginatus, and Triangulaspis spp.This assemblage characterises a nearly pandemic, relatively lateEarly Cambrian fauna termed the Serrodiscus–Triangulaspis–Hebediscus (“STH”) fauna or “band” (Robison et al., 1977; Geyer,1988, 1990, 2005b).
13.5. Laurentia
The oldest Laurentian trilobites have long been assigned to a sup-posed Fallotaspis assemblage. This assemblage, first described fromthe White-Inyo Mountains in California (Nelson and Hupé, 1964)(Fig. 2A), has long been assumed to include not precisely identifiablespecies of the fallotaspidids Fallotaspis and Daguinaspis (Nelson andHupé, 1964; Nelson, 1971, 1978; Fritz, 1972), and has been the sub-ject of repeated taxonomic reevaluations. Later work in adjacentwestern Nevada, particularly in the Esmeralda Mountains, EsmeraldaCounty, revealed a more detailed sequence of the earliest Laurentiantrilobites and brachiopods, and has led to a major taxonomic revision(Hollingsworth, 1999, 2000, 2005, 2007, 2011a,b).
The “Fallotaspis” assemblage in the Esmeralda Mountains occurs inthe upper Campito Formation. A report of F. tazemmourtensis Hupé,1953, in the quartzitic, middle Andrews Mountain Member nearWaucoba Springs (Nelson and Hupé, 1964; Alpert, 1976; Nelson,1978), which was used to define the base of the Fallotaspis Zone inLaurentia (Fritz, 1972), was not later confirmed (Hollingsworth,2005).
Trilobites first appear in the upper part of the traditional AndrewsMountain Member—in dark dysoxic sandstone and quartzite withsimple trace fossils now termed the Gold Coin Member(Hollingsworth, 2011a, 2011c). Trilobites and brachiopods are fairlycommon in light-coloured quartzitic sandstones with calcareouslenses that coincide with a lowstand systems tract near the top ofthe member. Hollingsworth (2007) proposed a Fritzaspis Zone forthis lowest trilobite assemblage (earlier the Repinaella? fauna ofHollingsworth, 2005). The Fritzaspis Zone (Fig. 4) is a 50–72 m inter-val in several sections. The assemblage includes Fritzaspis generalisHollingsworth, 2007; F. ovalis Hollingsworth, 2007; Profallotaspis?sp.; Amplifallotaspis keni Hollingsworth, 2007; and Repinaella sp.(Hollingsworth, 2007, 2011c).
The appearance of trilobites in these west Laurentian sections isinterpreted to reflect an upward facies change to shallow, well oxy-genated water (Fig. 9). Brachiopods (mainly obolellids) are common
in the quartzitic sandstones and constitute about two-thirds of thebody fossils. They occur with various trace fossils. The base of this in-terval has been suggested as the FAD of trilobites for Laurentia, andpossibly globally (Hollingsworth, 2005).
Above the Fritzaspis Zone is a 40–62 m interval without body fos-sils and with few trace fossils across the the Gold Coin–Montenegromember boundary. Archaeaspis trilobites appear abruptly and arecommon in a 30 cm interval, earlier termed the Archaeaspis Zone(Hollingsworth, 2005) and later used to mark the base of theFallotaspis Zone of Hollingsworth (2011a,b). This interval, 25–36 mabove the base of the Montenegro Member, is now regarded as asubzone within a continuous succession of greenish sandstone andsiltstone that forms the upper part of a transgressive systems tract.The index fossil is an undescribed, new species of Archaeaspis thatclosely resembles the Siberian Archaeaspis hupei Repina inKhomentovsky and Repina (1965). This interval has brachiopods, farless common than trilobite fragments, with bradoriids, hyoliths,tube-like conchs tentatively identified as Ladatheca and Sabellidites.
This horizon is overlain in Esmeralda County by a thick interval(ca. 130 m) with species originally assigned to Fallotaspis that forma virtually continuous fossil record. This true Fallotaspis assemblagebegins 5–15 m above the Archaeaspis Zone and has Fallotaspismacropleuron (Lieberman, 2002), F. nelsoni (Lieberman, 2002), andforms reported as F. cf. plana (Hupé, 1953) and F. aff. plana (Hupé,1953). Trilobites in the lower–middle parts of this Fallotaspis Zone in-clude “Daguinaspis” sp. and Cambroinyoella wallacei Lieberman, 2001.
Lieberman (2002) complementedW.H. Fritz and J.S. Hollingsworth'sfield studies by describing trilobites from classic localities in theWhite-Inyo Mountains. He proposed Archaeaspis macropleuronLieberman, 2002, and A. nelsoni Lieberman, 2002, based on material ear-lier assigned to Fallotaspis and Parafallotaspis. Lieberman's (2002) assign-ments were disputed by Hollingsworth (2005, 2007) because theyconflict with well preserved features of the glabella and palpebrallobes that show them to be members of the Fallotaspididae and notthe Archaeaspididae. Apart from the similarity of some of the Laurentianspecimens early assigned to Fallotaspiswith Fallotaspis species from theirtopotype area in the Moroccan Anti-Atlas, all of the Moroccan speciesdiffer in having an ocular line (“midinterocular ridge” of Lieberman,2002) that is absent in specimens from the White-Inyo Mountains, asdiscussed by Geyer (1996).
A “mixed Fallotaspis assemblage” (Hollingsworth, 2005) in the upperthird of the revised Fallotaspis Zone has trilobites, small shelly fossils,and brachiopods. The trilobites include Parafallotaspis, “Montezumaspis,”“Selindella,” Paranevadella, Nevadia, Judomia, Bradyfallotaspis?, two newspecies of Fallotaspis, and F. nelsoni (faunal list modified fromHollingsworth, 2011a). Several forms in this interval are very similar totrilobites of the Siberian middle Atdabanian Pagetiellus anabarus Zone(Figs. 4, 5).
Above this interval in Esmeralda County, holmiids and nevadiidsappear in the maximum flooding facies and mark the base of anEsmeraldina rowei Zone, which corresponds to the lowest part of thetraditional Laurentian Nevadella Zone (Fig. 4). Its base is the FAD ofMontezumaspis parallela (Fritz, 1995). Other trilobites includeArchaeaspis? sp., Nevadia n. sp., and Montezumaspis cometes (Fritz,1995) (Hollingsworth, 2011a)—an assemblage apparently unknownin the White-Inyo region but which resembles lower faunas of theLaurentian traditional Nevadella Zone. E. rowei (Walcott, 1910), abun-dant higher in the zone, is accompanied by E. elliptica Hollingsworth,2006; Nevadia weeksi Walcott, 1910; Palmettaspis consorta Fritz,1995; and P. lidensis (Fritz, 1995), among others (Hollingsworth,2011a). The traditional Nevadella Zone (Fritz, 1972) is nowsubdivided, in ascending order, into the E. rowei, Grandinasus patulus,Avefallotaspis maria, Nevadia addyensis, and N. eucharis zones(Hollingsworth, 2011a; Hollingsworth and Babcock, 2011) (Fig. 4).
Other Fallotaspis Zone trilobites in western Laurentia are found inthe Cassiar Range of northern British Columbia, where thin limestone

158 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
beds yield a sparse fauna with Parafallotaspis? sp. and Fallotaspis (or“Cirquella”) nelsoni overlain by Nevadella Zone trilobites (Fritz,1978). Calcareous rocks with Parafallotaspis represent the upperFallotaspis Zone in the Mackenzie Mountains (Fritz, 1976), wherethe lowest Sekwi Formation has a fauna similar to Hollingsworth's(2005) “mixed Fallotaspis assemblage” and an overlying Esmeraldinafauna.
Reports of Fallotaspis-type trilobites from the Death Valley regionof California (Hunt, 1990) and the Carborca area of Sonora, Mexico(McMenamin, 1987), are based on problematical identifications ofpoorly preserved specimens and await confirmation. The oldest de-terminable trilobite from the Puerto Blanco Formation at Caborcacan be assigned to Esmeraldina.
13.6. Siberian Platform
Trilobites appear on the Siberian Platform near or somewhat abovethe base of the Atdabanian Stage, which is defined by the base of theRetecoscinus zegebarti Zone (archaeocyathans) (Zhuravleva et al., 1969;Sokolov and Zhuravleva, 1983). Key sections for the earliest trilobitesare located in cliffs along the middle Lena River in the Lena–Aldanfacies region and in the Bulkur Anticline of the Khara–Ulakh ranges(e.g., Astashkin et al., 1990, 1991). The lowest trilobites in theLena River sections are found in the middle Pestrotsvet Formation,with Profallotaspis jakutensis Repina in Khomentovsky and Repina(1965) the index fossil of the lowest Atdabanian P. jakutensis Zone(Figs. 4, 5).
The FAD of Siberian trilobites accompanies a distinct lithofacieschange from interbedded reddish, fine-grained limestones and thinmudstones to carbonates with “algal limestones” (i.e., thin planarthrombolites in Varlamov et al., 2008, fig. 30) deposited on the openshelf (Repina, 1981; Pegel, 2000) (Fig. 9). At Zhurinsky Mys on theLena River, the base of the Atdabanian corresponds to the appearanceof thrombolite beds and the abrupt re-appearance of archaeocyathanswithin the Pestrotsvet Formation above a 35 m-thick interval withoutarchaeocyathans (despite their absence, this 35 m interval is assignedto the Dokidocyathus lenaicus Zone). The oldest trilobites of theProfallotasis jakutensis Zone occur 2.6 m above this linked appearanceof thrombolites and R. zegebarti Zone archaeocyathans at ZhurinskyMys (Astashkin et al., 1991; Varlamov et al., 2008, fig. 34).
The lowest Atdabanian P. jakutensis Zone trilobites are overlain bylow diversity Repinaella Zone assemblages with Repinaella explicata(Repina in Khomentovsky and Repina, 1965) and R. sibirica (Repinain Khomentovsky and Repina, 1965). Both species were first assignedto Fallotaspis, which lead to a genus-based correlation with the Mo-roccan and Laurentian fallotaspidid faunas, but both species wereshown to be archaeaspidids (Geyer, 1996). These Repinaella speciesshow only a superficial similarity with Repinaella? sp. describedfrom western Laurentia (see above).
A dramatic development in the Repinaella Zone is a very strongpositive δ13C perturbation—the peak of the Siberian carbon isotopeexcursion IV (Kaufman et al., 1996). First reported near the top ofthe P. jakutensis Zone at Isit' and Zhurinsky Mys (Kirschvink et al.,1991; Brasier et al., 1994b,c), re-study of the Zhurinsky Mys sectionplaces the excursion IV peak in the lower Repinaella Zone (Varlamovet al., 2008, fig. 27) (Fig. 5).
The third Atdabanian trilobite zone is the P. anabarus Zone, orDelgadella anabarus Zone of Varlamov et al. (2008). At Oy-Muran, thediverse trilobites include P. anabarus Lazarenko, 1962; Triangullinaparvula Repina, 1964; A. hupei Repina in Khomentovsky and Repina(1965); Nevadella subgroenlandica (Repina); Pseudoresserops oculatusRepina, 1964; Bigotinella rara Repina, 1981; and B. malykanica(Suvorova, 1960). The carbon isotope profile shows a strong positiveexcursion (V) in the upper P. anabarus Zone across Siberia (Brasier etal., 1994b,c; Kaufman et al., 1996) (Figs. 4, 5).
The Judomia–Prouktaspis Zone forms the upper Atdabanian andhas trilobites such as Pagetiellus lenaicus (Toll and von, 1899);Triangulaspis lermontovae Lazarenko, 1957; Bonnia arguta Repina,1960; and Metadoxides patria Repina. These forms are distinctly de-rived morphologically and do not represent very early trilobites.
A section along the Lena River near Ulakhan–Ald'yarkhay Creek inthe Bulkur Anticline exposes the Tommotian–Atdabanian Tyuser For-mation. The lowest trilobites include undetermined, poorly preservedfallotaspidids and archaeaspidids in the lowest Atdabanian R.zegebarti Zone 33–40 m above the formation's base, and overlie richsmall shelly fossil assemblages in the upper Tommotian D. lenaicusZone (Figs. 4, 5). The middle Atdabanian P. anabarus Zone has P.anabarus; P. sakhaicus Nikiforov, 1974; Nevadella effusa Repina,1974; and N. subgroenlandica (Demokidov and Lazarenko, 1959,1964; Repina et al., 1974; Lazarenko and Repina, 1983; Astashkin etal., 1991).
13.7. South China Platform
On the South China Platfom, the lowest trilobites define the basesof the Qiandongian Series and Nangaoan Stage (e.g., Peng, 2000, 2003,2009), which more or less coincide with the traditional ChiungchussiStage as defined by the All-China Committee on Stratigraphy in 1959(e.g., Zhang, 1980). Distinct lateral lithofacies changes accompanychanges in the earliest trilobite assemblages.
On the Yangtze Platform, in its strict sense, the lowest trilobitesoccur above a distinct unconformity on shelf carbonates and appearin overlying shelf black shale, calcareous shale, siltstone, and marl-stone (e.g., Niutitang Formation) (Fig. 8). The trilobites form a diverseassemblage with bradoriids, brachiopods, and fossils of other groups.In the Jiangnan Slope Belt, the lower Qiandongian Series is dominatedby black siliceous and phosphatic rocks of the Niutitang Formation.The trilobite faunas are mostly composed of species of the eodiscoidTsunyidiscus. The Jiangnan Basin facies is characterised in the lowerQiangdongian by siliceous and calcareous siliciclastics (XiaoyanxiFormation) with in situ, commonly articulated trilobites.
The trilobites traditionally believed to be the oldest in China occurin the lower, but not lowest, Yu'anshan Formation (previously amember of the so-called Chiungchussu or Qiangzhusi Formation) inYunnan on the Yangtze Platform (e.g., Steiner et al., 2001). The trilo-bites again appear above an unconformity on shelf carbonates whichis mantled by dark mudstone with a phosphatised small shelly fauna.The lowest trilobites from the overlying fine-grained siliciclastic rocksare characterised by Parabadiella and define the Parabadiella Zone.The Parabadiella assemblage in eastern Yunnan includes P. conicaLuo, 1981; P. yunnanensis Luo, 1981; Mianxiandiscus badaowanensisLuo, 1981; M. jinningensis Luo, 1981; Wutingaspis tingi Kobayashi,1944; W. kuanyangensis Luo, 1981; W. planifrons Luo, 1981;Yunnanocephalus planifrons Luo, 1981; and Y. subparallelus Luo, 1981.
Parabadiella specimens from the Meishucun section have been con-sidered to be Earth's oldest trilobites (Luo, 1981; Zhang, 1987). However,Xiang and Zhou (1987) incorrectly claimed that Tsunyidiscus species rep-resent even older trilobites in the eastern Yangtze Gorges in HubeiProvince. Nevertheless, these forms were reported near the base ofthe Shujingtuo Formation in that region, where they were describedto be associated with small shelly fossil assemblage III (Sinosachitesflabelliformis–Tannuolina zhangwentangi Assemblage) of the Meishucunsection (Fig. 4). Following this interpretation, the Hubei trilobites shouldbe older than the Parabadiella assemblage in Yunnan (see discussion inPeng and Babcock, 2001). However, Tsunyidiscus is younger in all sec-tions in which Parabadiella is also found.
This problem of which Chinese trilobite fauna is older is presentlyunresolved, but is a relatively unimportant question in global correla-tion of the oldest trilobites. The presence of the “oldest” Chinese trilo-bites in small shelly fossil assemblage III shows that the Chinesetrilobites are relatively high in the trilobite-bearing Lower Cambrian.

Fig. 8. Locations of the 528 Ma ash in southern New Brunswick marked by abbreviated locality names with asterisks (e.g., RBr*). Abbreviations: BHr, Beaver Harbour; CBr, CradleBrook; GoS, Gooderich Street; HBE, Hanford Brook east section; IS., Island; McC, McCleod Brook; MyL, Mystery Lake; pl., platform; RBr, Ratcliffe Brook; SoS, Somerset Street; and113, roadcut at kilometre 113 of Trans-Canada highway.
159E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Small shelly fossil assemblage III includes forms best regarded asupper Atdabanian- or Botoman Stage-indicators in terms of the Sibe-rian Platform (Landing et al., 1989; Landing, 1992, 1994; comparewith Brasier et al., 1990).
The middle–upper Yu'anshan Formation in Yunnan belongs tothe Eoredlichia–Wutingaspis Zone (Zhang et al., 2001; Fig. 4),which is sometimes divided into a lower Tsunyidiscus and anupper Yunnanocephalus subzone. The well known Chengjianglagerstätte is in the Yunnanocephalus Subzone and has suchbiostratigraphically significant trilobites as Eoredlichia intermedia(Lu, 1940), Yunnanocephalus yunnanensis (Mansuy, 1912), andKuanyangia pustulosa (Lu, 1941).
Eoredlichia shensiensis Zhang, 1966, and the genotype species P.huoi Zhang, 1966, were first described from Shaanxi Province, withE. shensiensis from the upper part of the traditional Chiungchussu For-mation and P. huoi from the lower–middle part, which is now theYu'anshan Formation (Zhang, 1966; Chen, 1985). Both are criticalspecies for correlation into South Australia.
The trilobites of the lower Nangaoan Stage occur in deeper water de-posits and include such eodiscoids as Tsunyidiscus (particularly T.niutitangensis Zhang in Lu et al., 1974), Neocobboldia, Sinodiscus, andHupeidiscus, aswell as such redlichioids as Paraichangia andMetaredlichia.These taxa characterise the lower Hupeidiscus–Sinodiscus Zone of theNangaoan. AHupeidiscus fengdongensis–Zhuxiella fangxianensisZone is de-fined in southeast Guizhou and Hubei and is assigned to the lowerCanglanpuan (Fig. 4). Zhuxiella has been interpreted as a subgenus ofEstaingia (Yuan et al., 2011). This suggests a correlation with the Pararaiajaneae Zone of South Australia, but this correlation does not seem to bevery plausible if based on other criteria. The Hupeidiscus–SinodiscusZone is overlain by the Sichuanolenus–Chengkouia Zone with trilobitesthat indicate a high interval in the trilobite-bearing Lower Cambrian(Fig. 4).
13.8. Australian East Gondwana
The Lower Cambrian trilobite sequence proposed by Jell (1990) isbased on South Australian successions (Fig. 2A), particularly in theFlinders Ranges (e.g., Jago et al., 2006). Four trilobite zones are distin-guished (Fig. 4). The lowest Abadiella huoi Zone has only five species—A. huoi (Zhang, 1966); Elicicola calva Jell, 1990; Yorkella australis
(Woodward, 1884); Eoredlichia sp.; and a form identified as Alanisiaguillermoi (Richter and Richter, 1940) in the upper part.
A nomenclatorial problem exists with A. huoi—first described as P.huoi from South China, it suggests a direct correlation with theParabadiella Zone of South China (Fig. 4). This species has beenassigned in Australia to Abadiella Hupé, 1953, which is based on a Mo-roccan Lower Cambrian species.
The genotype and only known specimen of Abadiella bourginiHupé, 1953, is an incomplete cranidium (examined by G. Geyer)that is slightly obliquely compacted laterally. Hupé's (1953, fig.45.2) original drawing is a poor representation. The A. bourgini holo-type is much smaller than the Parabadiella species from South Chinaand has a distinctly longer genal spine, but the length of the palpebrallobes is difficult to determine. The holotype was collected at a poorlydescribed horizon from a discontinuous section in the westernAnti-Atlas. Although its provenance is poorly known, the data inHupé's (1953) description allow it to be referred to the DaguinaspisZone (Fig. 4). The preservation and provenance of A. bourgini suggestthat this genus' and species' name should be restricted to the holo-type, and that the best name for the index species from South Chinaand Australia is P. huoi.
The presumed A. guillermoi from the lowest Australian trilobitezone was first described from Richter and Richter's (1940) Saukiandafauna of the Sierra Morena, Spain. The Saukianda fauna has Resseropsresserianus Richter and Richter, 1940; Saukianda andalusiae Richterand Richter, 1940; Perrector perrectus Richter and Richter, 1940; andStrenuaeva sampelayoi Richter and Richter, 1940. Most Sierra Morenaspecimens are distorted tectonically. Thus, the genus and species con-cepts of A. guillermoi are poorly defined (Henningsmoen, 1958;Sdzuy, 1961, 1962). The species is known from internal or compositemoulds in deformed shale, and a comparison with well preserved ma-terial is problematical. The Saukianda fauna is in the traditionalIberian middle Marianian Stage, and clearly occurs in the WestGondwanan Banian Stage (Fig. 4). Equivalent strata in southernMorocco are part of the Antatlasia guttapluviae Zone (e.g., Geyer andLanding, 2004, 2006), and are younger than those from which A.bourgini was reported by Hupé (1953).
The overlying Australian Pararaia tatei Zone has at least ten types oftrilobites. Four species persist from the A. huoi Zone. These include P.huoi and A. guillermoi. Newly appearing species include P. tatei(Woodward, 1884); E. shensiensis Zhang, 1966; and Egyngolia willochra

160 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Jell, 1990. E. shensiensis allows correlation with the Eoredlichia–Wutingaspis Zone of South China (Fig. 4).
14. Defining a Series 2 base
14.1. Earliest trilobites and their diachroneity
As discussed above, the Avalonian Callavia faunas are undoubtedlyrelatively late among the early trilobites, and correlate with the tradi-tional Nevadella Zone in Laurentia. The “D. attleborensis” fauna ofsoutheastern Newfoundland and central England is dominated byeodiscoids that include elements of the nearly pandemic, late EarlyCambrian Serrodiscus–Triangulaspis–Hebediscus (“STH”) fauna or“band” (Robison et al., 1977; Geyer, 1988, 2005b). Faunas withsome of these trilobites are found in Taconic Laurentia (e.g., Rasetti,1967), West Gondwana (upper A. guttapluviae Zone of Morocco,Spain, and Sardinia; Richter and Richter, 1941; Hupé, 1953; Geyer,1990; Álvaro and Liñán, 1997; Gozalo et al., 2003; Geyer andLanding, 2004), the Altay–Sayan Fold Belt (Repina, 1964; Repinaand Romanenko, 1978), the Siberian Platform (Demokidov andLazarenko, 1959; Yegorova and Savitsky, 1969; Repina et al., 1974),and the Russian Far East (Okuneva and Repina, 1973). The STHfauna is older than the global Hawke Bay regression (see Geyer,2005b), and its sequence stratigraphic position and biota suggest itcould define a Cambrian Stage 4 base quite high in the upper LowerCambrian.
The oldest Avalonian Callavia-dominated assemblages areinterpreted to have a ca. 519 Ma lower bracket in Wales (Landing etal., 1998) and occur above a regional unconformity on a lowerAtdabanian-equivalent limestone in eastern Newfoundland andEngland that lacks trilobites (Landing, 1988, 1991, 1992, 1995b;Brasier et al., 1992; Landing and Westrop, 2004; Landing et al.,2013b) (Figs. 3–5). Its geochronologic age and the domination ofthe Callavia fauna by derived holmiid trilobites (Palmer and Repina,1993) suggest that it is much younger than the earliest Atdabaniantrilobites in Siberia (ca. 521.4 Ma as suggested by Maloof et al.,2010b).
Fig. 9. Diachroneity in appearance of earliest Cambrian trilobites (curved grey shapes) and reGondwana, Laurentia, and Siberia are strongly influenced by available data on the strong poin the Moroccan Atlas ranges (Maloof et al., 2005, 2010a).
Carbon isotope stratigraphic work by Brasier et al. (1992) on thisuppermost, “sub-trilobitic” limestone in Avalonia reveals that thepeak of Siberian excursion IV in the lower, but not lowest, AtdabanianAge (lower Repinaella Zone; Figs. 4, 5) is recogniseable in the FostersPoint Formation in southeast Newfoundland and the lithologicallysimilar, coeval upper Home Farm Member in England (Fig. 3). Thisdata on the late age of the C. broeggeri Zone assemblage and correla-tion of the Siberian lowest trilobites (lower Repinaella Zone andlower) with subtrilobitic strata in Avalonia (Figs. 5, 9) demonstratethe extremely diachronous, lowest local occurrence (FAD) of trilo-bites in the Early Cambrian.
Correlation between South Australia and the South China isallowed by P. huoi—an index fossil in the oldest trilobite-bearingfaunas on both continents (Jell in Bengtson et al., 1990; Geyer andShergold, 2000; Steiner et al., 2001; Yuan et al., 2011). That theParabadiella assemblage of Yunnan has the Earth's oldest trilobites(e.g., Luo, 1981; Zhang, 1987) has been disproven (discussedabove). This assemblage and the overlying faunas of South China arelargely endemic. However, their likely correlation into Siberia iswith the latest Atdabanian–early Botoman.
Parabadiella occurs in Yunnan in calcareous siltstones above phospho-rites. These siltstones have been examined with non-conventionalmethods and correlated with the Tommotian Stage (Kirschvink et al.,1991; Zhang et al., 2001). However, small shelly fossils from the basalYu'anshan Formation phosphorite directly under the Parabadiella-bearing faunas are considered an upper Atdabanian–Botoman-equivalent(Landing et al., 1989; Landing, 1992, 1994). It should be noted, however,that correlations based on non-trilobite fossils are highly divergent (Qianand Bengtson, 1989; Yuan et al., 2011, table 1).
A correlation of the P. tatei Zone of South Australia with theEoredlichia–Wutingaspis Zone of South China uses the presence of E.shensiensis in both zones. In accordance with the correlation of theunderlying zones, these two zones should be about coeval with thelower Botoman Bergeroniellus micmacciformis–Erbiella Zone (Fig. 4).
The Siberian Platform trilobites were long regarded by far as theEarth's oldest, which was subsequently questioned by a number ofauthors (e.g., Geyer, 1996; Hollingsworth, 2005, 2007, 2011a). Thelowest Siberian trilobites comprise fairly uniform assemblages,
lationship to unconformities (vertical lines) and lithofacies. Correlations betweenWestsitive carbon isotope excursion at the Lie de vin–Igoudine formation boundary interval

161E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
which probably reflect the fact that a relatively uniform calcareous fa-cies has been sampled at comparatively few sections. Correlations ofthe Atdabanian trilobite zones are difficult because of their ende-mism, and interregional correlation based on shared genera is prob-lematic as the Siberian trilobites are defined by broad genus-levelconcepts.
The Botoman Stage likely represents only a short time interval asspecies from its zones occur in the STH band (see above). Thus, theunderlying Judomia–Prouktaspis Zone at the top of the Atdabanianprobably correlates with the higher part of the traditional NevadellaZone or even lower Olenellus Zone of Laurentia (Fig. 4).
Suggested correlations of the earliest Laurentian trilobites focuson apparent similarities with the archaeaspidid and fallotaspidid tri-lobites of Siberia and Morocco. Hollingsworth (2005, 2011a) noted aresemblance of Fritzaspis interval trilobites with those from theSiberian P. jakutensis and R. explicata zones. He also noted the resem-blance of A. keni to forms from the Moroccan Eofallotaspis Zone andemphasised a similarity of Eofallotaspis prima to Profallotaspis. Finally,Archaeaspis sp. from the Archaeaspis assemblage of the basal FallotaspisZone (sensu stricto) of the Esmeralda Basin was compared with A.hupei from the P. anabarus Zone of Siberia and Eofallotaspis tioutensisfrom Morocco, which Hollingsworth (2005, 2011a) interpreted as evi-dence of a second, and lower, band of intercontinentally correlativefaunas.
These correlations are only based on morphological similarities inearly trilobites from several Cambrian regions. Thus, the trilobitesmerely suggest faunal similarities, but do not necessarily indicatethat they are coeval. Hollingsworth (2011a) proposed an evolution-ary history in which bigotinids in the earliest trilobite-bearing strataof southern Morocco and Spain appear slightly later in Siberia, where-as the earliest trilobites with facial sutures appeared in westernLaurentia as late as the A. maria Zone (Fig. 4). This history, in fact, re-flects facies differences and shows that environmental conditionsplayed an important role in the composition of trilobite faunas(Fig. 9)—bigotinids are confined to shallow marine limestones inWest Gondwana and Siberia but apparently coeval siliciclastic stratain Laurentia lack bigotinids.
Several forms from the Laurentian Fallotaspis Zone (sensuHollingsworth) resemble Moroccan species, and one has been identi-fied as Fallotaspis cf. plana (and listed as F. plana by Hollingsworth,2011a, fig. 8). Another correlation has been suggested from theLaurentian lower Fallotaspis Zone to the Moroccan ChoubertellaZone based on a similarity of “Cirquella” nelsoni with Choubertella(Fritz, 1993). This similarity is superficial and mainly based on theabsence of genal spines in both taxa. The similarity of “C.” nelsoniactually seems to be with Daguinaspis from Morocco, which wouldshift the correlations notably (Fig. 4). In addition, the Laurentian“Daguinaspis” from the upper Fallotaspis Zone is difficult to accom-modate in the generic concept of Fallotaspis from Morocco. Neverthe-less, Hollingsworth (2005) correctly noted that small olenelloidswithout genal spines occur in a short stratigraphic interval in westernLaurentia, Siberia, and Morocco—these include “C.” nelsoni in Laurentia,Lenallina in Siberia, and Choubertella and Daguinaspis in Morocco.
A stronger similarity is shown between trilobites of the Laurentianupper Fallotaspis Zone (or“mixed Fallotaspis Zone”) and the SiberianP. anabarus Zone. The latter has Nevadella spp. and Selindella, whilea form similar to Selindella occurs in Nevada. In addition, a trilobitetentatively assigned to Judomia occurs high in the Fallotaspis Zone,which suggests a correlation with part or all of the Siberian JudomiaZone (Fig. 4). It needs to be emphasised, however, that Judomiaranges up to the middle Botoman Bergeroniellus gurarii Zone. Thus, re-markable affinities exist between west Laurentian forms and Siberianupper P. anabarus Zone faunas.
The lowest identifiable Moroccan trilobites are incompletelyknown and occur in a monofacial carbonate sequence of the upperIgoudine Formation in an assemblage dominated by archaeocyathans.
Only a few trilobite species have been described from the lowest ho-rizons. Unpublished Eofallotaspis material indicates several specieswhich are morphologically linked by rare intermediate forms. Sucha pattern is known in metazoans early in their evolutionary prolifer-ation (e.g., Pfennig et al., 2010), and this pattern in the Moroccan ma-terial may record rapid evolutionary developments in Eofallotaspis.Maloof et al. (2005, 2010a,b) recognise the IV carbon isotope peakat the boundary interval of the uppermost Lie-de-vin and lowermostIgoudine Formation, which appears to be well below the level ofSdzuy's (1978) Eofallotaspis and Hupetina horizons. This recognitionof the IV excursion peak suggests that the known Eofallotaspis andHupetina specimens are well above the base of Series 2 and Stage 3.However, the carbon isotope curve was determined from sections inwhich fossils have not yet been found, and related isotope studieshave not been carried out in the Tiout-Aguerd Trough of the westernAnti-Atlaswith its pronounced facies-shifts and resultingly diachronouslithostratigraphic boundaries. A puzzling consequence of the peakpositive excursion at the top Lie-de-vin–lowermost Igoudine inter-val and well below the recognised base of the Eofallotaspis Zoneis that it squeezes the earliest Moroccan trilobite zones into a veryshort period.
As noted above, Fallotaspis is not limited to the named fallotaspididzones in Morocco and ranges up into the A. guttapluviae Zone. Thus, the“Fallotaspis zone” or “Fallotaspis stage” (e.g., Fritz, 1972; Repina, 1986)lacks a clear definition, intercontinental correlation would extend this“zone” at least into the Laurentian upper Nevadella Zone and SiberianBotoman Stage (Geyer, 1990).
West Gondwana–Siberia–East Gondwana correlations are difficultand can yield conflicting results that reflect the taxa used. B. bivallatain Normandy and southern Spain, as well as a tentative Bigotinella,have been used for correlation into the Siberian lower Atdabanian(Liñán et al., 2005) although bigotinids are strongly facies dependentand restricted to calcareous strata. This means that their local occur-rences and FADs do not record their complete ranges. Other correla-tions between these three Early Cambrian regions often rely ontrilobite genera with problematical taxonomic definitions. Smallshelly fossils in the Pedroche Formation of the Sierra de Córdoba,southern Spain (Fernández Remolar, 1996, 2001), suggest correlationof the lowest trilobites in Iberia and Normandy into the lowerIssendalenian Stage in West Gondwana (traditional Ovetian of Iberia)and with small shelly fossil assemblage III of South China (Fig. 4).
The problem of dating the oldest identifiable trilobites of Balticais discussed above. The endemic S. mickwitzi Zone fauna does notallow a reliable trilobite-based correlation. Correlations based on as-sociated S. ornata–F. membranacea Zone acritarchs have been usedto suggest that the S. mickwitzi Zone has the Earth's oldest trilobites(Moczydłowska and Vidal, 1988; Moczydłowska and Yin, 2012).However, S. mickwitzi and associated trilobites do not have a primi-tive morphology and are likely late Early Cambrian (Palmer andRepina, 1993). In addition, the overlying Holmia-dominated assem-blages have late Early Cambrian ellipsocephaloids. As Holmia evenoccurs in the S. mickwitzi Zone, Holmia seemingly ranges throughnearly the entire trilobite-bearing Lower Cambrian in Baltica.Holmia's range would be astonishingly long if the S. mickwitziZone is greatly older than the lower Atdabanian as suggested bythe acritarch reports.
14.2. Non-trilobite FADs for the Series 2 base
Because of diachronous local first appearances of trilobites andproblems in trilobite-based interregional correlations, the FADs ofsmall shelly fossils have been proposed to define a GSSP for thebase of Cambrian Series 2. Rozanov et al. (2011) suggested the FADof Mobergella radiolata Bengtson, 1968, a problematicum knownfrom phosphatic, operculum-like microfossils, to define the Series 2base. They also used this FAD to redefine and lower the base of the

Fig. 10. Lower Cambrian stage and series designations proposed in this report.
162 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Siberian Atdabanian Stage. Their proposal is problematical becauseM.radiolata was long considered characteristic of the sub-trilobiticLower Cambrian of Siberia.
Missarzhevsky (1989, fig. 14, table 5) reported M. radiolata fromupper Tommotian strata (upper D. regularis and D. lenaicus zones)and used it to define an upper Tommotian Mobergella Zone.Rozanov et al. (2011, fig. 2) seemingly truncated the lower range ofM. radiolata by not recording it in the D. lenaicus and D. regulariszones, and extended its range upward halfway through theAtdabanian. These reports suggest that M. radiolata's range is not re-liably established in Siberia, and it is premature to use its FAD to de-fine the GSSP of a Series 2 base. Rozanov et al.'s (2011) redefinition ofthe Atdabanian base also creates a new, much thicker stage that mustreceive a new name, otherwise the expanded “Atdabanian” is a juniorhomonym of the traditional Atdabanian (e.g., Salvador, 1994).
M. radiolata is known in Siberia and Baltica, but its occurrence inBaltica is a poor correlation datum as it is facies controlled(Moczydłowska, 1991). Its presence in the S.–F. Zone (acritarchs) inBaltica would indicate a much older FAD than reported in Siberia.
Steiner et al. (2011) suggested the lowest occurrences of severalsmall shelly taxa as potential bases for a Series 2 base. While notingthe provincial restriction or relative rarity of most of them, they sug-gested the FAD of the gastropod Pelagiella subangulata Tate, 1892, as away to define a GSSP. Indeed, P. subangulata is found on a number ofCambrian palaeocontinents, including tropical Gondwanan Australia(Bengtson et al., 1990) and South China (Steiner et al., 2007), and itis rare in the cool-water platform succession of the upper Cuslett For-mation of Avalonian southeast Newfoundland (Landing et al., 1989).
The problem of the FAD concept (Fig. 1) applies to P. subangulata—the confidence level in its stratigraphic range is limited and itsinterregional correlation is controversial. Diachroneity in the P.subangulata FAD is shown by its rare appearance in upperTommotian–lower Atdabanian-equivalent strata in Newfoundland(Landing, 1992). It appears in the lowest Yu'anshan Formation onthe South China Platform in a younger interval that is variouslyregarded as an Atdabanian-equivalent (Qian and Bengtson, 1989;Steiner et al., 2007), a Botoman-equivalent (Landing et al., 1989), oran unresolved Atdabanian–Botoman-equivalent (Jago et al., 2006)(Fig. 4). More importantly, its FAD is unknown as the species appearsabove a sequence boundary in China. The likelihood of an extremelylow range in South Australia is shown by the fact that the base of aP. subangulata Zone lies below the S. ornata–Fimbriaglomerelladissimilare Zone (acritarchs) and apparently within the A. tornatum–
C. velvetum Zone (Moczydłowska and Zang, 2006)—this would placethe lowest P. subangulata specimens in the Laolinian Stage (CambrianStage 2) and at the level of the lower W. crosbyi Zone.
15. Suggested “Lenaldanian Series” and “Zhurinskyan Stage”(new)
15.1. Definition, concepts, location, etymology, and preservation of GSSP
The base of Cambrian Series 2 and Stage 3 form the top of theTerreneuvian Series (Landing et al., 2007a) and its upper “LaolinianStage” (this report) (Fig. 10). The “Lenaldanian Series” (new) and“Zhurinskyan Stage” (new) are suggested as appropriate designationsto replace informal Cambrian Series 2 and Stage 3. It is the role of theInternational Subcommission on Cambrian Stratigraphy to evaluateand accept the names of chronostratigraphic divisions, and“Lenaldanian” and “Zhurinskyan” are suggested in this report as ap-propriate, defensible, and completely defined chronostratigraphicunits, and should be seen as informal units. The GSSP for both the“Lenaldanian Series” and “Zhurinskyan Stage” occurs in the ZhurinskyMys (Zhurinsky's Cape) section on the Lena River in Yakutia, easternSiberia. The GSSP is a horizon and point marked by the peak of the IVcarbon isotope excursion in the lower Repinaella Zone and above the
base of the Atdadanian Stage. This point is 75 m above the base of theZhurinsky Mys section opposite the mouth of Negyurchyune Creek, aleft tributary of the Lena River. The coordinates of the Zhurinsky Myssection are 60° 55′ 0″ N and 125° 53′ 26″ E.
The GSSP for the base of the suggested “Lenaldanian” and“Zhurinshyan” lies at the positive carbon isotope peak earlier de-scribed as 70 m above the base of Zhurinsky Mys section and withinthe upper part of the lowest Siberian trilobite assemblage (P.jakutensis Zone) by Kirschvink et al. (1991), as based on stratigraphicdata from Kirschvink and Rozanov (1984). Reevaluation and restudyof the Zhurinsky Mys section places the peak of the IV carbon isotopeexcursion and GSSP for the base of the “Lenaldanian Series” and“Zhurinskyan Stage” at ca. 75 m above the base of the ZhurinskyMys section (data in Astashkin et al., 1990, 1991; Brasier et al.,1994b; Varlamov et al., 2008). The GSSP horizon is ca. 1 m abovethe base of the 80 m-thick, dolomitic limestone of the NokhoroyMember of the Pestrotsvet Formation and near the base of bed 8 ofthe Zhurinsky Mys section (Varlamov et al., 2008, figs. 27, 31). Thishorizon can be further specified as located in the middle R. zegebartiZone (archaeocyathans) and in the lower Repinaella Zone (trilobites).The GSSP for the “Lenaldanian Series” and “Zhurinskyan Stage” is lo-cated ca. 33 m above the regional stratotype horizon for theTommotian–Atdabanian boundary. It is also ca. 50 m above the low-est reported occurrence of M. radiolata (data from Astashkin et al.,1990; Varlamov et al., 2008).
The “Lenaldanian Series” features the compound word“Lenaldanian,” which would be recogniseable by many EarlyPalaeozoic workers because the Lena–Aldan river area in theYakutianregion of eastern Siberia is well known for its terminal Ediacaran–Cambrian successions (e.g., Varlamov et al., 2008). The GSSP of the“Lenaldanian Series” and “Zhurinskyan Stage” is in a section on the

163E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Lena River, and “Lenaldanian” emphasises its location. The names ofalmost all suitable local geographic and cultural features have beenused for litho- and chronostratigraphic terminology in the region(e.g., “Lenian Series,” “Yakutian horizon”), while “Lenaldanian” is anon-preoccupied name and not a homonym of an existing term.“Zhurinskyan Stage” is based on a topographic name that has notbeen used as part of any stratigraphic term. It is named for theZhurinsky Mys (“Zhurinsky's Cape”) section on the Lena Riverwhere the GSSP for the coterminous bases of Series 2 and Stage 3 ofthe Cambrian is herein defined. One name that is precluded fromuse for either Series 2 or Stage 3 is “Atdabanian,” because the GSSPfor the “Lenaldanian Series” and “Zhurinskyan Stage” lies above theformally defined base (base of archaeocyathan Retecoscincus zegebartiZone) of the regional Siberian Atdabanian Stage at the Zhurinsky Myssection (Interdepartmental Stratigraphic Committee, 1984).
The Zhurinsky Mys section is a shallow shelf succession withmixed thin carbonate beds and calcareous mudstone beds that hasno apparent stratigraphic breaks through and above the GSSP inter-val. The completely exposed section is abundantly fossiliferous withwell described archaeocyathans, small shelly fossils, and trilobites ofthe middle Tommotian–lower Atdabanian stages. The section's fossilsallow for precise correlations across the Siberian Platformwhile smallshelly fossils allow interregional correlations that help bracket thebase of the “Lenaldanian Series” (e.g., Landing et al., 1989; Brasier etal., 1994b; Landing, 1994). The Zhurinsky Mys section has been thesubject of thorough litho-, bio-, chemo-, and magnetostratigraphicstudies. Features of the Zhurinsky Mys section that are appropriateto a global GSSP locality include its relative accessibility, tectonicallyundeformed character, and location in a sparsely populated areathat will allow long-term preservation.
A lower bracket for correlation of the GSSP horizon is the appear-ance of the lowest fallotaspidid and archaeaspidid trilobites belowthe GSSP that allow correlation across Siberia and into westernLaurentia. In addition, preliminary palaeomagnetic results show asuccession of normal and reversed polarity intervals through theTerreneuvian–“Lenaldanian” boundary interval at Zhurinsky Mys(e.g., Kirschvink et al., 1991; Varlamov et al., 2008, fig. 27) that willlikely offer another non-conventional global correlation method.
15.2. Chronostratigraphic significance of “Lenaldanian Series” and“Zhurinskyan Stage” GSSP
The “FAD” of trilobites and the FADs of several small shelly fossilsare inappropriate for defining the base of Series 2 and its lowest Stage3 because of the high provincialism and facies controls of late EarlyCambrian faunas. The best standard for definition and global correla-tion of the Terreneuvian Series–“Lenaldanian Series” boundary in-volves carbon isotope stratigraphy. The strongest positive δ13C peakof excursion IV in the lower Repinaella Zone (trilobites) of the lowerAtdabanian Stage provides a readily correlated horizon in Siberia(Kirschvink et al., 1991; Brasier et al., 1994b,c; Kaufman et al., 1996).
The peak of the IV excursion is likely known from the top Lie devin–lower Igoudine Formation in southern Morocco (Maloof et al.,2005, 2010a,b). The oldest described Moroccan trilobites of theEofallotaspis Zone (Sdzuy, 1978; Sdzuy and Geyer, 1988) occur imme-diately above the facies change to less restricted, more energetic ma-rine environments in the middle Igoudine Formation and distinctlyabove the IV positive peak suggested by Maloof et al. (2005, 2010a).Thus, the oldest described Moroccan trilobites are referable to the“Lenaldanian Series” (Fig. 9). This carbon isotope-based correlationnot only further corroborates the traditional correlation of the oldestMoroccan archaeocyathans at about the base of the Eofallotaspis Zonewith the Siberian Atdabanian Stage (Geyer and Landing, 1995, 2006),but also appears to demonstrate that the oldest described trilobites ofMorocco (Eofallotaspis Zone) are not particularly ancient by compar-ison with the early trilobites of Siberia. However, undescribed,
fragmentary trilobite remains lower in the Igoudine Formation(Sdzuy and Geyer, 1988) show the presence of mineralised trilobitespossibly in the upper Terreneuvian Series of Morocco, as well asbelow the “Lenaldanian Series” in the series' “type region” in Siberia(Fig. 9).
Brasier et al. (1992) recognised the IV peak in Avalonia near the topof a distinctive, relatively thick carbonate unit (up to 12 m) and justbelow the unconformity with the earliest Callavia-bearing trilobitefaunas in southeast Newfoundland (upper Fosters Point Formation)and England (upper Home Farm Member). This carbonate unit, whichlacks any trilobites and straddles the Terreneuvian–“Lenaldanian”boundary, shows that the stratigraphically lowest trilobites are highlydiachronous in the Early Cambrian and that there is no trilobite “FAD”(Figs. 3, 5).
Zhu et al. (2004) documented and named the CARE (Cambrian Ar-thropod Radiation Excursion) carbon excursion in South China and cor-related it with excursion IV (Fig. 5). However, Zhu et al. (2006)suggested that the CARE excursion is associated with the appearanceof a “wide variety of arthropods, particularly in major CambrianLagerstätten deposits” and listed the Chengjiang and Balang (China),Poleta (western Laurentia), Sirius Passet (North Greenland), and EmuBay (South Australia) Lagerstätten. It must be emphasised that thislist includes localities of distinctly different ages so that the suggestedcorrelation and the naming of the CARE excursion are inappropriate(Fig. 5).
Use of the peak of the positive δ13C excursion of the Siberian IV ex-cursion, its recognition in Mongolia and Morocco, as the most highlypositive excursion in the uppermost C. baltica Zone in Avalonia, andas the peak of a coeval, redefined CARE-like excursion in South Chinaallows interregional correlation that cannot be confounded by the se-vere provincialism and dissimilar trilobite and non-trilobite fossils ondifferent palaeocontinents (Kirschvink et al., 1991; Brasier et al.,1992, 1994a; Maloof et al., 2005, 2010a,b). The great potential of aGSSP defined by the peak of carbon isotope excursion IV at ZhurinskyMys is that this excursion allows correlation into sparsely ornon-fossiliferous strata, as those in the lower Igoudine Formation (orLie de vin–Igoudine boundary interval; discussed above) of Morocco.
The 519 Ma U–Pb zircon date on a volcanic ash low in the range ofAvalonian trilobites (Landing et al., 1998, 2013b) provides a lower ageon the C. broeggeri Zone with its late-appearing olenelloid trilobites. Acalculated 520.4 Ma age for the Atdabanian Stage base (Maloof et al.,2010a,b) is probably closer to the age of the basal “Lenaldanian.” Thus,a ca. 520 myr date is shown for the base of the “Lenaldanian” as this ho-rizon is somewhat younger than that of the Atdabanian Stage (Fig. 4).This age is greater than the estimated 530–534 Ma bracket for theonset of stage 2 of the Cambrian radiation—which included the diversi-fication of molluscs (Maloof et al., 2005, 2010a) and the appearance ofWatsonella and Aldanella. The base of the “Laolinian Stage” is withinthe lower ranges of W. crosbyi and A. attleborensis and is figured as ca.530 Ma (Fig. 10). This estimated age for the base of the “Laolinian” ap-proximately divides the (generally) pre-trilobitic Early Cambrian intotwo roughly equal parts.
16. Discussion
The Cambrian has been the focus of research for over 50 years sincethe International Subcommission on Cambrian Stratigraphy wasestablished in 1961—with its role to develop international standardsfor the base and internal divisions (series and stages) of the period(Peng and Babcock, 2011). Traditional techniques that proposed corre-lations between highly provincial biotas and defined Cambrian chrono-stratigraphic divisions have been largely limited to FADs of individualtaxa. Evaluations and critiques of the FADs of proposed GSSP-definingtaxa for the bases of Cambrian Stage 2 and Series 2 are required This re-port critiques a number of earlier proposed FADs and finds them tobe both locally and globally diachronous, while often lying at

164 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
unconformities or facies changes. More robust methods to define theGSSPs of the bases of Stage 2 and Series 2 must be utilised. This reporturges that the International Commission on Stratigraphy no longer pro-mote a wholly FAD-based chronostratigraphy—which presumes thatthe lowest occurrence of single species can be geochronologically iden-tical worldwide.
The GSSP of a global chronostratigraphic unit should in the future bedefined on the basis of all available evidence that helps bracket its def-inition and correlation. Thus, GSSPs for the bases of Stage 2 and Series 2of the Cambrian Sysytem are suggested that are associatedwith, but notdefined by, a biotic zone, which itself lies within a biotic zonal succes-sion. In addition, GSSP definitions must be accompanied byreproduceable, high resolution, non-conventional correlation tech-niques, particularly carbon isotope techniques in the Lower Cambrian(Brasier et al., 1990, 1992, 1994a; Maloof et al., 2005, 2010a; Landinget al., 2011; Landing and Geyer, 2012). Intercontinental and regionalcorrelations based on biostratigraphic and non-conventional correla-tion should eventually be supported by detailed, high-frequency se-quence stratigraphic analyses. GSSPs only provide a standard tocompare with and correlate into, and it must be conceded that long dis-tance correlation based on a single taxon become less precise at greaterdistances from a GSSP (Cowie, 1986; Cowie et al., 1986) and may evenbe essentially impossible across biofacies and provincial boundaries.There are limits to the resolution allowed by biostratigraphy, and thisincludes uncertainties associated even with establishing the completestratigraphic range of a fossil even in a stratigraphically continuous,monofacial section.
The problemwith use of a single species' FAD to define the base of achronostratigraphic unit is discussed above by the use of the conodont I.fluctivagus FAD to define the base of the Ordovician—possiblemisidentificationmeans that the base of the Ordovician at its stratotypemay lie a few beds higher than its GSSP. Limitation of the FAD concept isshown by sole use of the T. pedum FAD to define the base of the Cambri-an at its GSSP. The supposedly “established” FAD later proved to be atleast 4 m higher than later discovery of the taxon at Fortune Head.However, this lower occurrence does not reduce in any way the utilityof involving T. pedum as part of the definition of the base of the Cambri-an. Additional biotic and nonconventional techniques can be used to as-sist correlation of this well established datum.
The concept of using T. pedum in definition of the GSSP for the baseof the Cambrian has been accompanied by a too strict reliance on useof its FAD alone to define the base of a system, as well as its lowest se-ries, and stage-level divisions. Criticism by Peng and Babcock (2011)of the Fortune Head GSSP is based on the mistaken concept that aFAD can provide a precise geochronometric guide. In truth, the Cam-brian (and Phanerozoic) base was actually defined by the base of theT. pedum Assemblage Zone (Narbonne et al., 1987; Landing et al.,2007a), characterised by the broadly coeval appearance of an arrayof different and comparatively complex trace fossils that occurabove the highest Edicaran fossils. The assemblage zone marks theonset of stage one of the Cambrian Evolutionary Radiation and signalsthe rapid appearance of metazoans with complex behaviour, as iswell recorded in the Fortune Head section. However, the relativelyeurytopic distribution of the producer of T. pedum in shallow marinefacies suggests that its lowest occurrence at Fortune Head likely ap-proximates the evolutionary origin of the producer of the trace, andthat a relatively minor range offset is present for the FAD of thisichnospecies at the basal Cambrian GSSP.
All that is required to conserve a proven useful and durable horizonin the Cambrian Radiation is a minor redefinition of the T. pedum Zone.The Ediacaran–Cambrian boundary interval is now understood to in-clude a record of ever-more Phanerozoic-aspect trace fossil communi-ties, plus an initially slow diversification of skeletalised metazoans andthe disappearance of characteristic Ediacaran vendobionts andskeletalised forms over an estimated five million year-long interval(Zhuravlev et al., 2012). The appearance of trichophycids, including T.
pedum, forms part of these boundary-inverval biotic developments.Thorough studies over the last generation have repeatedly shown thatthe appearance of T. pedumwith such ichnofossils asMonomorphichnus,forms part of a distinct bio- and chemostratigraphic succession that isyounger than characteristic terminal Ediacaran taxa and developments.The latter include the problematica Harlaniella and Palaeopascichnus,the skeletalised taxa Cloudina and Namacalathus, the BACE carbon iso-tope excursion, and an acritarch diversification marked by the base ofthe A. tornatum–C. velvetum Zone (e.g., Zhuravlev et al., 2012). Inshort, the best definition of the basal Cambrian GSSP lies within therange of the producer of T. pedum and lies at the base of the hereinredefined T. pedum Assemblage Zone at Fortune Head, southeasternNewfoundland.
A GSSP for the base of Cambrian Stage 2 must avoid the problems as-sociatedwith a biotic FAD, and should be definedwithin the ranges of thepalaeogeographically widespread, but intra- and interregionallydiachronous appearance of the micromollusks W. crosbyi and A.attleborensis. Early occurrences of both species in Avalonia, South China,and Siberia are sub-Tommotian on the basis of small shelly fossil—pluscarbon isotope-based correlation. The lowest occurrence of this two-species association must no longer be regarded as Tommotian. A pro-posed definition of the Fortunian Stage–Stage 2 boundary lies at thepeak of the I′/L4/ZHUCE carbon isotope excursion within the earlier partof the range of W. crosbyi and A. attleborensis in the road cut near Laolinvillage, Yunnan, China. This suggested GSSP for the base of the suggested“Laolinian Stage” (i.e., a reproposed informal designation Cambrian Stage2) of theupper Terreneuvian Series is significantly older than528 Ma. The“Laolinian Stage” approximately embraces the appearance and diversifi-cation of skeletalised metazoans and the age of its base may be close to531 Ma (e.g., Maloof et al., 2005, 2010a,b) (Fig. 9).
Correlations based on small shelly fossils, acritarchs, and early trilo-bites and examination of lithofacies and stratigraphic continuity clearlyshow that trilobites had a diachronous lowest local occurrence. Hence,the base of Series 2 cannot be based on the FAD of trilobites in generalor on any one of the highly provincial trilobites of this time interval.All of the suggested FADs of small shelly fossils proposed to define thebase of Series 2 suffer from the problems inherent to the FAD concept,while the candidate species are limited to several palaeocontinents.
Themost robust definition for the base of Series 2 cannot be definedby the FAD of any trilobite or small shelly fossil, and is best defined to lieat the peak of the strong IV carbon isotope excursion that isrecogniseable in Avalonia, South China, Siberia, Mongolia, and WestGondwana (Morocco). In these areas, the IV peak and a necessarilyredefined CARE carbon isotope excursion occur in fossiliferous stratawith good potential for biostratigraphic correlation. This IV/refinedCARE excursion is clearly recogniseable in the Fosters Point Formationand Home Farm Member in southeast Newfoundland and England,and this emphasises the long held conclusion that Series 2 includes lo-cally sub-trilobitic strata. The GSSP for the base of Series 2 is suggestedto lie at the peak of carbon isotope excursion IV, and somewhat aboveboth the lowest occurrence of trilobites and the base of the Atdabanianat the ZhurinskyMys section on the Lena River. This suggested GSSP de-fines the top of the Terreneuvian Series at the coterminous bases of the“Lenaldanian Series” (new) and “Zhurinskyan Stage” (new), and in-cludes occurrences of the oldest mineralised trilobited in Siberian andprobably western Laurentia in the terminal Terreneuvian. The 519 Madate based on the oldest trilobite bearing unit in Wales and 521.5 Macalculated age on the base of the Atdabanian suggest a 520 Ma age onthe Terrenauvian–“Lenaldanian” Series boundary (Fig. 9).
Acknowledgements
M. Moczydłowska, A. H. Knoll, and P. Wignall are thanked for use-ful comments. Much of EL's work was done under National ScienceFoundation grant support while at the New York State Museum.GG's work was supported by research grant GE 549/21-1 of the

165E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Deutsche Forschungsgemeinschaft (DFG). A.C. Maloof provided usefulcomments and editorial help. This report is dedicated to the contribu-tions to Cambrian palaeobiology and correlation of Jan Bergström.
References
Ahlberg, P., 1984. Lower Cambrian trilobites and biostratigraphy of Scandinavia. LundsPublications in Geology 22, 1–37.
Ahlberg, P., Bergström, J., 1978. Lower Cambrian ptychopariid trilobites from Scandina-via. Sveriges Geologica Undersökning, Avhandlingar och Uppsatser, I 4:o, Ser. Ca,49 (40 pp.).
Ahlberg, P., Bergström, J., 1998. The Cambrian of Scania. In: Ahlberg, P. (Ed.), Guide toExcursions in Scania and Västergötland, Southern Sweden: Lunds Publications inGeology, 14, pp. 20–23.
Ahlberg, P., Bergström, J., Johansson, J., 1986. Lower Cambrian olenellid trilobites from theBaltic Faunal Province. Geologiska Föreningens i Stockholm Förhandlingar 108, 39–56.
Albert, S.P., 1977. Trace fossils and the basal Cambrian boundary. In: Crimes, T.P.,Harper, J.C. (Eds.), Trace FossilsGeological Journal Special Issue 9, 1–8.
Alcock, F.J., 1938. Geology of Saint John region, New Brunswick. Geological Society ofCanada Memoir 216 (65 pp.).
Alpert, S.P., 1976. Trilobites and star-like trace fossils from the White-Inyo Mountains,California. Journal of Paleontology 50, 226–240.
Álvaro, J.J., Clausen, S., 2007. Lower Cambrian shelled phosphorites from the northernMontagne Noire, France. In: Álvaro, J.J., Aretz, M., Boulvain, F., Munnecke, A.,Vachard, D., Vennin, E. (Eds.), Palaeozoic Reefs and Bioaccumulations: Climatic andEvolutionary ControlsGeological Society, London, Special Publication 275, 17–28.
Álvaro, J.J., Liñán, E., 1997. Nuevos datos acerca del Bilbiliense (Cámbrico inferior ter-minal) en las Cadenas Ibéricas y su correlación con otras áreas. Revista Españolade Paleontología 12, 277–280 (in Spanish).
Álvaro, J.J., Elicki, O., Debrenne, F., Vizcaïno, D., 2002. Small shelly fossils from theLower Cambrian Lastours Formation, southern Montagne Noire, France. Geobios35, 397–409.
Álvaro, J.J., Elicki, O., Geyer, G., Rushton, A.W.A., Shergold, J.H., 2003.Palaeogeographical controls on the Cambrian trilobite immigration and evolution-ary patterns reported in the western Gondwana margin. Palaeogeographica,Palaeoclimatology, Palaeoecology 195, 5–35.
Álvaro, J.J., Monceret, E., Monceret, S., Verraes, G., Vizcaïno, D., 2011. Stratigraphic re-cord and palaeogeographic context of the Cambrian Epoch 2 subtropial carbonateplatforms and their basinal counterparts in SW Europe, West Gondwana. Bulletinof Geosciences 85, 573–584.
Álvaro, J.J., Zamora, S., Clausen, S., Vizcaïno, D., Smith, A.B., 2013. The role of abiotic fac-tors in the Cambrian Substrate Revolution: a review from the benthic communityreplacements of West Gondwana. Earth-Science Reviews 118, 69–82.
Amthor, J.E., Grotzinger, J.P., Schröder, A., Bowring, S.A., Ramezani, J., Martin, M.W.,Matter, A., 2003. Extinction of Cloudina and Namacalathus at the Precambrian–Cambrian boundary in Oman. Geology 31, 431–434.
Antcliffe, J.B., Gooday, A.J., Brasier, M.D., 2011. Testing the protozoan hypothesis forEdiacaran fossils: a developmental analysis of Palaeopascichnus. Palaeontology54, 1157–1175.
Astashkin, V.A., Varlamov, A.I., Esakova, N.V., Zhuravlev, A.Yu, Repina, L.N., Rozanov,A.Yu, Fedorov, A.B., Shabanov, Yu.Ya, 1990. Putevoditel'ekskursii po rekam Aldanui Lene. Sibirskaya platforma. Tretiy Mezhdunarodnyy simpozium po kembriyskoysisteme. Akademiya Nauk SSSR, Sibirskoe otdelenie, Institut geologii i geofiziki,Paleontologicheskiy institut. SNIIGiMS, Novosibirsk (115 pp. (in Russian)).
Astashkin, V.A., Pegel, T.V., Shabanov, Yu.Ya, Sukhov, S.S., Sundukov, V.M., Repina, L.N.,Rozanov, A.Yu, Zhuravlev, A.Yu, 1991. The Cambrian system on the Siberian Plat-form. Correlation Chart and Explanatory Notes, 27. International Union GeologicalSciences, Pub. (133 pp.).
Babcock, L.E., Peng, S.C., 2007. Cambrian chronostratigraphy: current state and futureplans. Palaeogeography, Palaeoclimatology, Palaeoecology 254, 62–66.
Babcock, L.E., Peng, S.C., Geyer, G., Shergold, J.H., 2005. Changing perspectives on Cam-brian chronostratigraphy and progress toward subdivision of the Cambrian Sys-tem. Geosciences Journal 9, 101–106.
Barr, S.M., Raeside, R.P., 1989. Tectono-stratigraphic terranes in Cape Breton Island,Nova Scotia: implications for the configuration of the northern Appalachianorogen. Geology 17, 822–825.
Barrande, J., 1881. Système Silurien du centre de la Bohême. Acephales. : , 6 (Publisherunnamed, Paris and Prague (1881), 361 pp. (in French)).
Bengtson, S., 1968. The problematic genus Mobergella from the Lower Cambrian of theBaltic area. Lethaia 1, 325–351.
Bengtson, S., 1988. Skeletal fossils and the Precambrian–Cambrian transition. In:Landing, E., Narbonne, G.M., Myrow, P. (Eds.), Trace Fossils, Small Shelly Fossilsand the Precambrian–Cambrian Boundary: New York State Museum Bulletin,464, p. 8.
Bengtson, S., Fletcher, T.P., 1983. The oldest sequence of skeletal fossils in the Lower Cambri-an of southeastern Newfoundland. Canadian Journal of Earth Sciences 20, 526–536.
Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A., Runnegar, B.N., 1990. Early Cam-brian fossils from South Australia. Memoirs of the Association of AustralasianPalaeontologists 9 (364 pp.).
Bergström, J., 1973. Classification of olenellid trilobites and some Balto-Scandian spe-cies. Norsk Geologisk Tidsskrift 53, 283–314.
Bergström, J., 1981. Lower Cambrian shelly faunas and biostratigraphy in Scandinavia. In:Taylor, M.E. (Ed.), Short Papers for the Second International Symposium on the Cam-brian System 1981: U. S. Geological Survey Open-File Reptort, 81-743, pp. 22–25.
Bergström, J., Gee, B.G., 1985. The Cambrian in Scandinavia. In: Gee, D.G., Sturt, B.A.(Eds.), The Caledonian Orogen — Scandinavia and Related Areas. John Wiley andSons, Ltd., Chichester, pp. 247–271.
Billings, E., 1860. On some new species of fossils from limestone at Point Levis oppositeQuebec. Canadian Naturalist 5, 301–324.
Billings, E., 1874. On some new species of fossils from the Primordial rocks of New-foundland. Paleozoic Fossils. : Part 1. Geological Survey of Canada, vol. 2. DawsonBrothers, Montréal, pp. 64–78.
Blaker, M.R., Peel, J.S., 1998. Lower Cambrian trilobites from North Greenland.Meddeleser om Grønland. Geosciences 35 (145 pp.).
Bokova, A.R., 1990. Novye nizhnekembriyskie gastropody Sibirskoy platformi.Paleontologichesky Zhurnal 1990, 123–126 (in Russian).
Bowring, S.A., Martin, H.W., 1999. High precision U–Pb geochronology, the tempo of evo-lution and the record from Gondwana. Journal of African Earth Sciences 28, 187–201.
Bowring, S.A., Grotzinger, J.P., Isachsen, C.E., Knoll, A.H., Pelechaty, S.M., Kolosov, P.,1993. Calibrating rates of Early Cambrian evolution. Science 161, 1293–1298.
Bowring, S.A., Grotzinger, J.P., Condon, J.P., Ramezani, J., Newall, M.J., Allen, P.A., 2007.Geochronologic constraints on the chronostratigraphic framework of theNeoproterozoic Huqf Supergroup, Sultanate of Oman. American Journal of Science307, 1097–1145.
Brasier, M.D., 1979. The Cambrian radiation event. In: House, M.R. (Ed.), The Origin ofMajor Invertebrate Groups: Systematics Association Special, vol. 12, pp. 103–159.
Brasier, M.D., 1984. Microfossils and small shelly fossils from the Lower Cambrian HyolithesLimestone at Nuneaton, English Midlands. Geological Magazine 121, 229–253.
Brasier, M.D., 1986. The succession of small shelly fossils (especially conoidal microfos-sils) from English Precambrian–Cambrian boundary beds. Geological Magazine 123,237–256.
Brasier, M.D., 1989. Towards a biostratigraphy of the earliest skeletal biotas. In: Cowie,J.W., Brasier, M.D. (Eds.), The Precambrian–Cambrian Boundary: Oxford Mono-graphs in Geology and Geophysics, 12, pp. 117–165.
Brasier, M.D., Lindsay, J.F., 2001. Did supercontinental amalgamation trigger the “Cam-brian Explosion?”. In: Zhuravlev, A.Yu., Riding, R. (Eds.), The Ecology of the Cam-brian Radiation. Columbia University Press, New York, pp. 69–106.
Brasier, M.D., Singh, P., 1987. Microfossils and Precambrian–Cambrian boundary stra-tigraphy at Maldeota, Lesser Himalaya. Geological Magazine 124, 323–345.
Brasier, M.D., Hewitt, R.A., Brasier, C.J., 1978. On the Late Precambrian–Early CambrianHartshill Formation of Warwickshire. Geological Magazine 115, 21–36.
Brasier, M.D., Perejón, A., de San José, M.A., 1980. Discovery of an important fossilifer-ous Precambrian–Cambrian sequence in Spain. Estudios Geologicas 35, 35–57.
Brasier, M.D., Margaritz, M., Corfield, R.M., Luo, H., Wu, X., Ouyang, L., Jiang, Z., Hamdi,B., He, T., Frasier, A.G., 1990. The carbon- and oxygen-isotope record of the Precam-brian–Cambrian boundary interval in China and Iran and their correlation. Geolog-ical Magazine 127, 319–332.
Brasier, M.D., Anderson, M.M., Corfield, R.M., 1992. Oxygen and carbon isotope stratig-raphy of Early Cambrian carbonates in southeastern Newfoundland and England.Geological Magazine 129, 265–279.
Brasier, M.D., Cowie, J., Taylor, M.E., 1994a. Decision on the Precambrian–Cambrianboundary. Episodes 17, 95–100.
Brasier, M.D., Rozanov, A.Yu., Zhuravlev, A.Yu., Corfield, R.M., Derry, L.A., 1994b. A car-bon isotope reference scale for the Lower Cambrian succession in Siberia: report ofIGCP Project 303. Geological Magazine 131, 767–783.
Brasier, M.D., Corfield, R.M., Derry, L.D., Rozavov, A.Yu., Zhuravlev, A.Yu., 1994c. Multi-ple δ13C excursions spanning the Cambrian explosion to the Botomian crisis in Si-beria. Geology 22, 455–458.
Brasier, M.D., Shields, G., Kuleshov, V.N., Zhegallo, E.A., 1996. Integrated chemo- andbiostratigraphic calibration of early animal evolution: Neoproterozoic–Early Cam-brian of southeast Mongolia. Geological Magazine 133, 445–489.
Brasier, M.D., McCarron, G., Tucker, R., Leather, J., Allen, P.A., Shields, G., 2000. New U–Pb zircon dates for the Neoproterozoic Gubrah glaciation and for the top of theHuqf Supergroup, Oman. Geology 28, 175–178.
Brasier, M.D., Antcliffe, J.B., Callow, R., 2011. Evolutionary trends in remarkable preser-vation across the Ediacaran–Cambrian transition and the impact of metazoanmixing. In: Allison, P., Bottger, D.J. (Eds.), Taphonomy and Bias through Time.Springer Verlag, Dordrecht, pp. 519–567.
Buatois, L.A., Almond, J., Germs, G.J.B., 2013. Environmental tolerance and range offsetof Treptichnus pedum: implications for the recognition of the Ediacaran–Cambrianboundary. Geology. http://dx.doi.org/10.1130/G33938.1 (2013).
Burrett, C., Long, J., Stait, B., 1991. Early–Middle Palaeozoic biogeography of Asianterranes derived from Gondwana. In: McKerrow, W.S., Scotese, C.R. (Eds.),Palaeozoic Palaeogeography and Biogeography: Geological Society, London,Memoir, 12, pp. 163–174.
Butterfield, N.J., 1995. Secular distribution of Burgess Shale-type preservation. Lethaia28, 1–13.
Buttterfield, N.J., 2001. Ecology and evolution of Cambrian plankton. In: Zhuravlev,A.Yu., Riding, R. (Eds.), The Ecology of the Cambrian Radiation. : Perspectives in Pa-leobiology & Earth History. Columbia University Press, New York, pp. 200–216.
Carter, J.G., Campbell, D.C., Campbell, M.R., 2000. Cladistic perspectives on early bivalveevolution. In: Harper, E.M., Taylor, J.D., Crane, J.A. (Eds.), The Evolutionary Biologyof the BivalviaGeological Society, London, Special Publication 17, 47–79.
Chen, P., 1984. Discovery of Lower Cambrian small shelly fossils from Jijapo, Yichang,west Hubei, and its significance. Professional Papers in Stratigraphy and Paleontol-ogy, 13, pp. 61–65.
Chen, R.Y., 1985. Lower Cambrian trilobites from Liangshan district of Nanzheng, Shaanxi.Acta Palaeontologica Sinica 24, 332–336 (in Chinese, with English abstract).
Cobbold, E.S., 1910. On some small trilobites from the Cambrian rocks of Comley(Shropshire). Quarterly Journal of the Geological Society of London 66, 19–50.

166 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Cobbold, E.S., 1921. The Cambrian horizons of Comley (Shropshire) and theirBrachiopoda, Pteropoda, Gasteropoda, etc. Quarterly Journal of the Geological Soci-ety of London 76, 325–386.
Cobbold, E.S., 1931. Additional fossils from the Cambrian rocks of Comley, Shropshire.Quarterly Journal of the Geological Society of London 87, 459–512.
Cobbold, E.S., 1935a. Lower Cambrian faunas from Hérault, France. Annual Magazine ofNatural History 16 (91), 25–48.
Cobbold, E.S., 1935b. A new genus of the Trilobita and a new species of theConchostraca from the Cambrian of the Carteret region, N.W. France. Annual Mag-azine of Natural History, Zoology, Botany, and Geology 15 (10), 381–392.
Compston, W., Zhang, Z., Cooper, D.A., Ma, G., Jenkins, R.J.F., 2008. Further SHRIMP geo-chronology on the Early Cambrian of South China. American Journal of Science 308,399–420.
Cowie, J.W., 1971. The Cambrian of the North American Arctic regions. In: Holland, C.H.(Ed.), Lower Palaeozoic Rocks of the World. : Cambrian of the New World, vol. 1.Wiley-Interscience, London and New York, pp. 325–383.
Cowie, J.W., 1986. Guidelines for boundary stratotypes. Episodes 9, 78–82.Cowie, J.W., Ziegler, W., Boucot, A.J., Bassett, M.G., Remane, J., 1986. Guidelines and
statutes of the International Commission on Stratigraphy (ICS). CourierForschungsinstitut Senckenberg, 83, pp. 1–14.
Crimes, T.P., Anderson, M.M., 1985. Trace fossils from late Precambrian–Early Cambrianstrata of southeastern Newfoundland (Canada): temporal and environmental im-plications. Journal of Paleontology 59, 310–343.
Crimes, T.P., Germs, G.J.B., 1985. Trace fossils from the Nama Group (Precambrian–Cambrian) of southwest Africa (Namibia). Journal of Paleontology 56, 890–907.
Dalziel, I.W.D., 1997. Overview: Neoproterozoic–Paleozoic geography and and tecton-ics: review, hypotheses, environmental speculations. Geological Society of AmericaBulletin 106, 16–42.
Debrenne, F., Debrenne, M., 1978. Archaeocyathid fauna of the lowest fossiliferouslevels of Tiout (Lower Cambrian, southern Morocco). Geological Magazine 115,1101–1109.
Debrenne, F., Debrenne, M., 1995. Limite Précambrien–Cambrien: évolution des con-cepts. Bulletin de la Société Géologique de France 166, 311–320.
Demokidov, K.K., Lazarenko, N.P., 1959. Novye dannye po stratigrafiya kembriyjskikhotlozheniy zapadnogo sklona severnogo Kharaulakha. Sbornik statey popaleontolologii i stratigrafii, 16, p. 11 (12).
Demokidov, K.K., Lazarenko, N.P., 1964. Stratigrafiya verkhnego dokembriya i kembriya inizhnekembriyskie trilobity severnoy chasti Sredney. Sibiri i ostrovov Sovetskoy Arktiki.Trudy nauchno-issled. Institut Geologii Arktiki (NIIGA), 137. Nedra, Moskow (286 pp.).
Doré, F., 1969. Les formations cambriennes de Normandie. (Ph.D. dissertation)Université de Caen (790 pp.).
Ebbestadt, J.O., Ahlberg, P., Høyberget, M., 2003. Redescription of Holmia inusitata(Trilobita) from the Lower Cambrian of Scandinavia. Palaeontology 46, 1039–1054.
Eisenack, A., 1958. Tasmanites Newton 1875 und Leiosphaeridia n. gen. aus Gattungender Hystrichosphaeridia. Palaeontographica, Abteilung A, 110, pp. 1–10.
Elicki, O., Geyer, G., 2010. The faunal province of the southern margin of the RheicOcean. Cambrian biostratigraphy. In: Linnemann, U., Romer, R.L. (Eds.), Pre-Meso-zoic Geology of Saxo-Thuringia — From the Cadomian Active Margin to theVariscan Orogen. Schweizerbart, Stuttgart, pp. 103–114.
Ergaliev, G.K., 1980. Trilobity srednego i verkhnego kembriya Malogo Karatau.Akademiya Nauk Kazakhskoy SSR, Ordena trudnogo krasnogo znameni InstitutGeologii. Nauk im. K. I. Satpaeva, Alma-Ata (211 pp. (in Russian)).
Erwin, D.H., LaFlamme, M., Tweed, S.M., Sperling, E.A., Pisani, D., Peterson, K.J., 2011.The Cambrian conundrum: earliest divergence and later ecological success in theearly history of animals. Science 334, 1091–1097.
Esakova, N.V., Zhegallo, E.A., 1996. Biostratigrafiya i fauna nizhnego kembriya Mongolii.Nauka, Moscow (214 pp. (in Russian)).
Fatka, O., Szabad, M., 2011. Agnostids entombed under exoskeletons ofparadoxidid trilobites. Neues Jahrbuch für Paläontologie, Abhandlungen 259,207–215.
Fernández Remolar, D.C., 1996. Los microfósiles problemáticos del Cámbrico Inferior dela Sierra de Córdoba: datos preliminares. Coloquios de Paleontología 48, 161–173.
Fernández Remolar, D.C., 2001. Chancelloriidae del Ovetiense inferior de la Sierra deCórdoba, España. Revista Española de Paleontología 16, 39–62.
Fletcher, T.P., 2003. Ovatoryctocara granulata, the key to a global Cambrian stageboundary and the correlation of the olenellid, redlichiid, and paradoxidid realms.Special Papers in Palaeontology 70, 73–102.
Fletcher, T.P., 2006. Bedrock geology of the Cape St. Mary's Peninsula, Southwest AvalonPeninsula (includes parts of 1M/1, 1N/4, 1L/16 and 1K/14 Map Areas). Geological Sur-vey, 06-02. Newfoundland and Labrador Department of Natural Resources (117 pp.).
Fletcher, T.P., Theokritoff, G., 2008. The Early Cambrian of eastern Massachusetts.Northeastern Geological and Environmental Sciences 30, 301–329.
Ford, S.W., 1872. Descriptions of some new species of primordial fossils. AmericanJournal of Science 7, 419–422 (3rd Series).
Ford, S.W., 1876. On additional species of fossils from the primordial of Troy andLansingburg, Rensselaer County, New York. American Journal of Science 11,369–371 (3rd Series).
Fortey, R.A., Cocks, L.R., 2003. Palaeontological evidence bearing on global Ordovician–Silurian continental reconstructions. Earth-Science Reviews 61, 245–307.
Fortey, R.A., Landing, E., Skevington, D., 1982. Cambrian–Ordovician boundary sectionsin the Cow Head Group, western Newfoundland. In: Bassett, M.G., Dean, W.T.(Eds.), The Cambrian–Ordovician Boundary: Sections, Fossil Distributions, and Cor-relations. : Geological Series, 3. National Museum of Wales, pp. 95–129.
Fritz, W.H., 1972. Lower Cambrian trilobites from the Sekwi Formation type section,Mackenzie Mountains, northwestern Canada. Bulletin of the Geological Survey ofCanada 212 (90 pp.).
Fritz, W.H., 1976. Ten stratigraphic sections from the Lower Cambrian Sekwi Forma-tion, Mackenzie Mountains, northwestern Canada. Geological Survey of CanadaPaper 76-22 (42 pp.).
Fritz, W.H., 1978. Upper (carbonate) part of Atan Group, Lower Cambrian, north-central British Columbia. Current Research, Part A, Geological Survey of CanadaPaper 78-1A, 7–16.
Fritz, W.H., 1993. New Lower Cambrian olenelloid trilobite genera Cirquella andGeraldinella from southwestern Canada. Journal of Paleontology 67, 856–868.
Fritz, W.H., 1995. Esmeraldina rowei and associated Lower Cambrian trilobites (1ffauna) at the base of Walcott's Waucoban Series, southern Great Basin, USA. Jour-nal of Paleontology 69, 708–723.
Gámez Vintaned, J.A., 1996. The Río Huso section. In: Liñán, E., Gámez Vintinaed, J.A.,Gozalo, R. (Eds.), Field Conference of the Cambrian Stage Subdivision WorkingGroups. International Subcommission on Cambrian Stratigraphy. Spain, September1996. : Field Trip Guide and Abstracts. Servicio de Publicaciones de la Universidadde Zaragoza, Zaragoza, pp. 21–31.
Gaucher, C., Boggiani, P.C., Sprechmann, P., Sial, A.N., Fairchild, T., 2003. Integrated cor-relation of the Vendian to Cambrian Arroyo del Sodado and Corumbá groups(Uruguay and Brazil): palaeogeographic, palaeoclimatic and palaeobiogeographicimplications. Precambrian Research 120, 241–278.
Gaucher, C., Sial, A.N., Ferreira, V.P., Pimental, M.M., Chiglino, L., Sprechmann, P., 2007.Chemostratigraphy of the Cerro Victoria Formation (Lower Cambrian, Uruguay):evidence for climate stabilization across the Precambrian–Cambrian boundary.Chemical Geology 237, 46–64.
Gehling, J., Jensen, S., Droser, M.L., Myrow, P.M., Narbonne, G.M., 2001. Burrowingbelow the basal Cambrian GSSP, Fortune Head, Newfoundland. Geological Maga-zine 132, 213–218.
Germs, G.J.B., 1972. New shelly fossils from Nama Group, South West Africa. AmericanJournal of Science 272, 752–761.
Geyer, G., 1988. Agnostida aus dem höheren Unterkambrium und dem Mittelkambriumvon Marokko. Teil 2: Eodiscina. Neues Jahrbuch der Geologie und Paläontologie,Abhandlungen 177, 93–133.
Geyer, G., 1990. Revised Lower to lower Middle Cambrian biostratigraphy of Morocco.Newsletters on Stratigraphy 22, 53–70.
Geyer, G., 1993. Untersuchungen zur Lithostratigraphie, Ablagerungsgeschichte undPaläontologie der Nama-Gruppe. Zentralblatt für Geologie und Paläontologie, TeilI. 241–243.
Geyer, G., 1996. The Moroccan fallotaspidid trilobites revisited. Beringeria 18, 89–199.Geyer, G., 2005a. The Fish River Subgroup in Namibia: stratigraphy, depositional envi-
ronments, and the Proterozoic–Cambrian boundary problem revisited. GeologicalMagazine 142, 465–498.
Geyer, G., 2005b. The base of a revised Middle Cambrian: are suitable concepts for a se-ries boundary in reach? Geosciences Journal 9, 81–99.
Geyer, G., Landing, E., 1995. The Cambrian of the Moroccan Atlas regions. In: Geyer, G.,Landing, E. (Eds.), Morocco '95—The Lower–Middle Cambrian Standard of WesternGondwana: Beringeria Special Issue, 2, pp. 7–46.
Geyer, G., Landing, E., 2004. A unified Lower–Middle Cambrian chronostratigraphy forWest Gondwana. Acta Geologica Polonica 54, 179–218.
Geyer, G., Landing, E., 2006. Latest Ediacaran and Cambrian of the Moroccan Atlas re-gions. In: Geyer, G., Landing, E. (Eds.), Morocco 2006. Ediacaran–Cambrian Deposi-tional Environments and Stratigraphy of the Western Atlas Regions. ExplanatoryDescription and Field Excursion Guide: Beringeria Special Issue, 6, pp. 9–75.
Geyer, G., Shergold, J.H., 2000. The quest for internationally recognized divisions ofCambrian time. Episodes 23, 188–195.
Geyer, G., Uchman, A., 1995. Ichnofossil assemblages from the Nama Group(Neoproterozoic–Lower Cambrian) in Namibia and the Proterozoic–Cambrianboundary problem revisited. Beringeria Special Issue 2, 175–202.
Geyer, G., Elicki, O., Fatka, O., Żylinska, A., 2008. Cambrian. In: McCann, T. (Ed.), The Ge-ology of Central Europe. : Volume 1: Precambrian and Palaeozoic. Geological Soci-ety, London, pp. 155–202.
Gnilovskaya, M.B., 1971. Drevsneyshiye rasteniya venda Russkoy platformy (pozdnyydokembriy). Paleontologisches Zhurural 1971, 101–107 (in Russian).
Gozalo, R., Liñán, E., Palacios, T., Gámez Vintaned, J.A., Mayoral, E., 2003. The Cambrianof the Iberian Peninsula: an overview. Geologica Acta 1, 103–112.
Grabau, A.W., 1900. Palaeontology of the Cambrian terranes of the Boston Basin. Occa-sional Papers of the Boston Society of Natural History, 4, pp. 601–694.
Gradstein, F.M., Ogg, J.G., Smith, A.G., 2005. A Geologic Timescale 2004.Cambridge Uni-versity Press, Cambridge (384 pp.).
Gravestock, D.I., Alexander, E.M., Demidenko, Yu.E., Esakova, N.V., Holmer, L.E., Jago,J.B., Lin, T.R., Melnikova, L.M., Parkhaev, P.Yu, Rozanov, A.Yu, Ushatinskaya, G.T.,Zang, W.I., Zhegallo, E.A., Zhurvalev, A.Yu, 2001. The Cambrian biostratigraphy ofthe Stansbury Basin, South Australia. Transactions of the Palaeontological Institute,282. Russian Academy of Science (344 pp.).
Grotzinger, J.P., Bowring, S.A., Saylor, B.Z., Kaufman, A.J., 1995. Biostratigraphic andgeochronologic constraints on early animal evolution. Science 270, 598–604.
Grotzinger, J.P., Watters, W.A., Knoll, A.H., 2000. Calcified metazoans in thrombolite–stromatolite reefs of the terminal Proterozoic Nama Group, Namibia. Paleobiology26, 334–359.
Gubanov, A.P., 2002. Early Cambrian palaeogeography and the probable Iberia–Siberiaconnection. Tectonophysics 352, 153–168.
Hagenfeldt, S.E., 1989. Lower Cambrian acritarchs from the Baltic Depression andsouth-central Sweden, taxonomy and biostratigraphy. Stockholm Contributionsin Geology 41, 1–176.
Haldemann, S.S., 1840. Supplement to number one of “A monograph of the limnniadesand other fresh-water univalve shells of North America, containing new descrip-tions of apparently new animals in different classes, and the names and character

167E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
of the subgenera in Paludina and Aculosa”. Philadelphia Academy of Sciences,pp. 1–3 (no volume number).
Hall, J., 1847. Palaeontology of New-York. Volume 1. Containing Descriptions of the Or-ganic Remains of the Lower Divisions of the New-York System (Equivalent of theLower Silurian Rocks of Europe.C. Van Benthuysen, Albany, NY (338 pp.).
Hall, J., 1852. The Palaeontology of New York, vol. II. C. Van Benthuysen, Albany, NY(362 pp.).
Hamdi, B., 1995. Precambrian–Cambrian deposits in Iran. Treatise on the Geology ofIran 20 (304 pp. in Pharsi, with English summary).
Hamdi, B., Brasier, M.D., Jiang, Z.W., 1989. Earliest skeletal fossils from Precambrian–Cambrian boundary strata, Elburz Mountains, Iran. Geological Magazine 125,283–289.
Harvey, T.H.P., Williams, M., Condon, D.J., Wilby, P.R., Siveter, D.J., Rushton, A.W.A.,Leng, M.J., Gabbott, S.H., 2011. A refined chronology of the Cambrian sucession ofsouthern Britain. Journal of the Geological Society of London 168, 705–716.
Hayes, A.O., Howell, B.F., 1937. Geology of Saint John, New Brunswick. Geological Soci-ety of America, Special Paper 5 (146 pp.).
He, T.G., Yang, X.H., 1982. Lower Cambrian Meishucun Stage of the western Yangtzestratigraphic region and its small shelly fossils. Bulletin of the Chendu Institute ofGeology and Mineral Resources, Chinese Academy of Geological Sciences 3,69–95 (in Chinese).
Henningsmoen, G., 1958. Los trilobites de las capas de Saukianda, Cámbrico inferior, enAndalucía. Con observaciones acerca de la deformación en los fósiles y sobre laclassificación de los Redlichiacea y los Elleipsocephalacea. Estudios Geológicos14, 251–271.
Hofmann, H.J., 1985. Precambrian carbonaceous megafossils. In: Toomey, D.F., Nitecki,M. (Eds.), Paleoalgology. Springer, Berlin, pp. 20–33.
Hofmann, H.J., Patel, I.M., 1989. Trace fossils from the type ‘Etcheminian’ Series' (LowerCambrian Ratcliffe Brook Formation), Saint John area, New Brunswick, Canada.Geological Magazine 126, 139–157.
Hollingsworth, J.S., 1999. Stop 10. A candidate position for the base of the MontezumanStage. In: Palmer, A.R. (Ed.), Laurentia 99, V Field Conference of the Cambrian StageSubdivision Working Group. : International Subcommission on Cambrian Stratig-raphy, September 12–22, 1999. Institute for Cambrian Studies, Boulder, Earth-Science Reviews 2013, pp. 34–37.
Hollingsworth, J.S., 2000. Environmental setting of the earliest trilobites in Laurentia(Cambrian, Begadean). Geological Society of America, Abstracts with Programs32 (7), A-301.
Hollingsworth, J.S., 2005. The earliest occurrence of trilobites and brachiopods in theCambrian of Laurentia. Palaeogeography, Palaeoclimatology, Palaeoecology 220,153–165.
Hollingsworth, J.S., 2006. Holmiidae (Trilobita: Olenelina) of the Montezuman Stage(Early Cambrian) in western Nevada. Journal of Paleontology 80, 309–332.
Hollingsworth, J.S., 2007. Fallotaspidoid trilobite assemblage from the Esmeralda Basin(western Nevada, U.S.A.). Memoirs of the Australasian Association ofPalaeontologists 32, 123–140.
Hollingsworth, J.S., 2011a. Lithostratigraphy and biostratigraphy of Cambrian Stage 3 inwestern Nevada and eastern California. Museum of Northern Arizona Bulletin 67,26–42.
Hollingsworth, J.S., 2011b. Stop 7D. The base of the Fallotaspis Zone (base of theMontezuman Stage, Cambrian Stage 3), Montezuma Range, Nevada. Museum ofNorthern Arizona Bulletin 67, 252–256.
Hollingsworth, J.S., 2011c. Stop 8B. Base of Cambrian Series 2, Stage 3 in the Gold CoinMine area, Esmeralda County, Nevada. Museum of Northern Arizona Bulletin 67,263–266.
Hollingsworth, J.S., Babcock, L.E., 2011. Stop 8A. Base of the Dyeran Stage (CambrianStage 4) in the middle Member of the Poleta Formation, Indian Springs Canyon,Montezuma Range, Nevada. Museum of Northern Arizona Bulletin 67, 256–262.
Hunt, D.L., 1990. Trilobite faunas and biostratigraphy of the Lower Cambrian WoodCanyon Formation, Death Valley Region, California. (unpublished M.Sc. thesis)Univ. California, Davis (103 pp.).
Hupé, P., 1953. Contribution à l'étude du Cambrien inférieur et du Précambrien III del'Anti-Atlas marocain. Notes et Mémoirs de la Service géologique du Maroc, 103(402 pp.).
Hupé, P., Abadie, J., 1950. Sur l’existence de trilobites du Cambrien inférieur marocain.Comptes Rendus des Séances de l’Academie des Sciences 230, 2112–2113.
Hutchinson, R.D., 1962. Cambrian stratigraphy and trilobite faunas of southeasternNewfoundland. Bulletin of the Geological Survey of Canada 88 (156 pp.).
Iba, Y., Mutterlose, J., Tanabe, K., Sano, S.-i, Misaki, A., Terabe, K., 2011. Belemnite extinctionand the origin of modern cephalopods 35 m.y. prior to the Cretaceous–Paleogeneevent. Geology 39, 483–486.
Interdepartmental Stratigraphic Committee, 1984. Resolutions of the InterdepartmentalStratigraphic Committee and Its Permanent Commissions.MSK 22, Leningrad (74pp.).
Isachsen, C.E., Bowring, S.A., Landing, E., Samson, S.D., 1994. New constraint on the di-vision of Cambrian time. Geology 22, 496–498.
Jago, J.B., Zang, W.L., Sun, X.W., Brock, G.A., Paterson, J.R., Skovsted, C.B., 2006. A re-view of the Cambrian biostratigraphy of South Australia. Palaeoworld 15,406–423.
Jell, P.A., 1990. Trilobita. In: Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A.,Runnegar, B.N. (Eds.), Early Cambrian Fossils from South Australia. : Memoir, 9. As-sociation of Australasian Paleontologists, pp. 257–322.
Jensen, S., 2003. The Proterozoic and earliest Cambrian trace fossil record; patterns,problems and perspectives. Integrative and Comparative Biology 43, 219–228.
Jensen, S., Saylor, B.Z., Gehling, J.G., Germs, G.J.B., 2000. Complex trace fossils from theterminal Proterozoic of Namibia. Geology 28, 143–146.
Kaufman, A.J., Knoll, A.H., Semikhatov, M.A., Grotzinger, J.P., Jacobsen, S.B., Adams, W.,1996. Integrated chronostratigraphy of Proterozoic–Cambrian boundary beds inthe western Anabar region, northern Siberia. Geological Magazine 133, 509–533.
Keppie, J.D., Ramos, V.A., 1999. Odessey of terranes in the Iapetus and Rheic oceansduring the Paleozoic. In: Ramos, V.A., Keppie, J.D. (Eds.), Laurentia–GondwanaConnections Before PangaeaGeological Society of America, Special Paper 336,267–276.
Keppie, J.D., Dostal, J., Murphy, J.B., Nance, R.D., 1996. Terrane transfer between easternLaurentia and western Gondwana in the Early Paleozoic: constraints on global re-construction. In: Nance, R.D., Thompson, M.D. (Eds.), Avalonian and Related Peri-Gondwana Terranes of the Circum-North AtlanticGeological Society of America,Special Paper 304, 369–380.
Kerber, M., 1988. Mikrofossilien aus unterkambrischen Gesteinen der Montagne Noire,Frankreich. Palaeontographica Abteilung A 202, 127–203.
Khomentovsky, V.V., Karlova, G.A., 1986. O nizhney granitse pestrotsvetnoy svity vbasseyne r. Aldan. In: Khomentovsky, V.V. (Ed.), Pozdniy dokembrii i rannipaleozoi Sibiri. Sibirskaya platforma i vneshnaya zona Altae-Sayanskoyskladchatoy oblasti. : Akademiya Nauk SSSR, Sibirskoe otdelenie, Institut geologiii geofizikii, Sbornik nauchnykh trudov. Nauka, Novosibirsk, pp. 3–22.
Khomentovsky, V.V., Karlova, G.A., 1993. Biostratigraphy of the Vendian–Cambrianboundary beds and the Lower Cambrian boundary in Siberia. Geological Magazine130, 29–45.
Khomentovsky, V.V., Repina, L.N., 1965. Nizhniy Kembriy stratotipicheskogo razrezaSibiri. Akademiya Nauk SSSR, Sibirskoe otdelenie, Institut geologii i geofizikii.Nauka, Moscow (198 pp.).
Kim, D.K., Westrop, S.R., Landing, E., 2002. Middle Cambrian (Acadian Series)conocorhyphid and paradoxidid trilobites from the upper Chamberlain's Brook For-mation, Newfoundland and New Brunswick. Journal of Paleontology 76, 822–842.
Kirschvink, J.L., Rozanov, A.Yu., 1984. Magnetostratigraphy of Lower Cambrian stratafrom the Siberian Platform: a palaeomagnetic pole and a preliminary polaritytime-scale. Geological Magazine 121, 189–203.
Kirschvink, J.L., Magaritz, M., Ripperdan, R.L., Zhuravlev, A.Yu, Rozanov, A.Yu, 1991. ThePrecambrian/Cambrian boundary: magnetostratigraphy and carbon isotopes re-solve correlation problems between Siberia, Morocco, and South China. GSAToday 1, 69–72 (87, 91).
Knoll, A.H., Grotzinger, J.P., Kaufman, A.J., Kolosov, P., 1995. Integrated approaches toterminal Proterozoic stratigraphy: an example from the Olenek Uplift, northeast-ern Siberia. Precambrian Research 73, 251–270.
Knoll, A.H., Walter, M.R., Narbonne, G.M., Christie-Blick, N., 2004. A new period for thegeologic time scale. Science 305 (621), 622.
Kobayashi, T., 1944. On the Cambrian formations in Yunnan and Haut-Tonkin and thetrilobites contained. Miscellaneous notes on the Cambro-Ordovician Geology andPalaeontology 16. Japanese Journal of Geology and Geography 19, 107–138.
Kouchinsky, A., Bengtson, S., Misssarzhevsky, V.V., Pelechaty, S., Torssander, P., Val'kov,A.K., 2001. Carbon isotope stratigraphy and the problem of a pre-Tommotian stagein Siberia. Geological Magazine 138, 387–396.
Kouchinsky, A., Bengtson, S., Pavlov, V.E., Pavlov, V., Runnegar, B.N., Val'kov, A., Young,E., 2005. Pre-Tommotian age of the lower Pestrotsvet Formation in the Selinde sec-tion on the Siberian platform: carbon isotopic evidence. Geological Magazine 142,319–325.
Kouchinsky, A., Bengtson, S., Pavlov, V.E., Runnegar, B.N., Torssander, P., Young, E., Ziegler,K., 2007. Carbon isotope stratigraphy of the Precambrian–Cambrian Sukharikha Riversection, northwestern Siberian platform. Geological Magazine 144, 1–10.
Kouchinsky, A., Bengtson, A., Runnegar, B.N., Skovsted, C., Steiner, M., Vendrasco, M.,2012. Chronology of Early Cambrian biomineralization. Geological Magazine 149,221–251.
Lake, P., 1936. A monograph of the British Cambrian trilobites. Palaeontographical So-ciety, London, Monograph 1936, 225–248.
Landing, E., 1988. Lower Cambrian of eastern Massachusetts: stratigraphy and smallshelly fossils. Journal of Paleontology 62, 661–695.
Landing, E., 1989. Paleoecology and distribution of the Early Cambrian rostroconchWatsonella crosbyi Grabau. Journal of Paleontology 63, 566–573.
Landing, E., 1991. Upper Precambrian through Lower Cambrian of Cape Breton Island:faunas, paleoenvironments, and stratigraphic revision. Journal of Paleontology 65,570–595.
Landing, E., 1992. Lower Cambrian of southeastern Newfoundland: epeirogeny and Lazarusfaunas, lithofacies–biofacies linkages, and the myth of a global chronostratigraphy. In:Lipps, J.H., Signor, P.W. (Eds.), Origins and Early Evolution of Metazoa. Plenum Press,New York, pp. 283–309.
Landing, E., 1994. Precambrian–Cambrian global stratotype ratified and a new perspec-tive of Cambrian time. Geology 22, 179–182.
Landing, E., 1995a. Precambrian–Cambrian global stratotype ratified and a new per-spective of Cambrian time—Reply. Geology 23, 286.
Landing, E., 1995b. Upper Placentian–Branchian Series of mainland Nova Scotia (mid-dle–upper Lower Cambrian: faunas, paleoenvironments, and stratigraphic revi-sion. Journal of Paleontology 69, 475–495.
Landing, E., 1996a. Reconstructing the Avalon continent: marginal-to-inner platformtransition in the Cambrian of Avalonian New Brunswick. Canadian Journal ofEarth Sciences 33, 623–632.
Landing, E., 1996b. Avalon—insular continent by the latest Precambrian. In: Nance, R.D.,Thompson, M. (Eds.), Avalonian and Related Peri-Gondwanan Terranes of theCircum-North AtlanticGeological Society of America, Special Paper 304, 27–64.
Landing, E., 1998a. Cambrian subdivisions and correlations: introduction. CanadianJournal of Earth Sciences 35 (321), 322.
Landing, E., 1998b. Avalon 1997—a pre-meeting viewpoint. In: Landing, E., Westrop,S.R. (Eds.), AVALON 1997—The Cambrian Standard. Third International Field

168 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Conference of the Cambrian Chronostratigraphy Working Group and I.G.C.P. Pro-ject 366 (Ecological Aspects of the Cambrian Radiation): New York State MuseumBulletin, 492, pp. 1–3.
Landing, E., 2004. Precambrian–Cambrian boundary interval deposition and themarginal platform of the Avalon microcontinent. Journal of Geodynamics 37,411–435.
Landing, E., 2005. Early Paleozoic Avalon–Gondwana unity: an obituary—response to‘Palaeontological evidence bearing on global Ordovician–Silurian continental re-constructions’ by R.A. Fortey and L.R.M. Cocks. Earth-Science Reviews 69, 169–175.
Landing, E., 2007. East Laurentia 2007―a pre-meeting statement. In: Landing, E. (Ed.),Ediacaran–Ordovician of East Laurentia. S.W. Ford Memorial Volume: New YorkState Museum Bulletin, 510, p. 3 (4).
Landing, E., 2012. Time-specific black mudstones and global hyperwarming on theCambrian–Ordovician slope and shelf of the Laurentia palaeocontinent.Palaeogeography, Palaeoclimatology, Palaeoecology 367–368, 256–272.
Landing, E., Benus, A.P., 1988. Stratigraphy of the Bonavista Group, southeastern New-foundland: growth faults and the distribution of the sub-trilobitic Lower Cambrian.In: Landing, E., Narbonne, G.M., Myrow, P. (Eds.), Trace Fossils, Small Shelly Fossils,and the Precambrian–Cambrian BoundaryNew York State Museum Bulletin 463,59–71.
Landing, E., Geyer, G., 2012. Misplaced faith—limitations of the first appearance datum(FAD) in chronostratigraphy and proposal of more robust Lower Cambrian correla-tion standards. In: Zhao, Y., Zhu, M.Y., Peng, J., Gaines, R.R., Parsley, R.L. (Eds.),Cryogenian–Ediacaran to Cambrian Stratigraphy and Paleontology of Guizhou,ChinaJournal of Guizhou University (Natural Sciences) 29 (Supplement 1),170–171.
Landing, E., MacGabhann, B.A., 2010. First evidence for Cambrian glaciation providedby sections in Avalonian New Brunswick and Ireland—additional data forAvalon–Gondwana separation by the earliest Palaeozoic. Palaeogeography,Palaeoclimatology, Palaeoecology 285, 174–185.
Landing, E., Murphy, J.B., 1991. Uppermost Precambrian(?)–Lower Cambrian of main-land Nova Scotia: faunas, depositional environment, and stratigraphic revision.Journal of Paleontology 65, 382–396.
Landing, E., Westrop, S.R., 1996. Upper Lower Cambrian depositional sequence inAvalonian New Brunswick. Canadian Journal of Earth Sciences 33, 404–417.
Landing, E., Westrop, S.R., 1998a. Cambrian faunal sequence and depositional history ofAvalonian Newfoundland and New Brunswick: field workshop. In: Landing, E.,Westrop, S.R. (Eds.), Avalon 1997—The Cambrian Standard. Third InternationalField Conference of the Cambrian Chronostratigraphy Working Group and I.G.C.P.Project 366 (Ecological Aspects of the Cambrian Radiation)New York State Muse-um Bulletin 492, 5–75.
Landing, E., Westrop, S.R., 1998b. Revisions in the stratigraphic nomenclature in theCambrian of Avalonian North America and comparisons with Avalonian Britain.In: Landing, E., Westrop, S.R. (Eds.), Avalon 1997—The Cambrian Standard. ThirdInternational Field Conference of the Cambrian Chronostratigraphy WorkingGroup and I.G.C.P. Project 366 (Ecological Aspects of the Cambrian Radiation)New York State Museum Bulletin 492, 76–87.
Landing, E., Westrop, S.R., 2004. Environmental patterns in the origin and evolutionand diversification loci of Early Cambrian skeletalised Metazoa: evidence fromthe Avalon microcontinent. In: Lipps, J.H., Wagoner, B. (Eds.), Neoproterozoic–Cambrian Biological Revolutions: Paleontological Society Paper, 10, pp. 93–105.
Landing, E., Narbonne, G.M., Myrow, P., Benus, A.P., Anderson, M.M., 1988. Faunas anddepositional environments of the upper Precambrian through Lower Cambrian,southeastern Newfoundland. In: Landing, E., Narbonne, G.M., Myrow, P. (Eds.),Trace Fossils, Small Shelly Fossils, and the Precambrian–Cambrian BoundaryNewYork State Museum Bulletin 463, 18–52.
Landing, E., Myrow, P., Benus, A.P., Narbonne, G.M., 1989. The Placentian Series: ap-pearance off the oldest skeletalized faunas in southeastern Newfoundland. Journalof Paleontology 63, 739–769.
Landing, E., Bowring, S.A., Davidek, K., Westrop, S.R., Geyer, G., Heldmaier, W., 1998.Duration of the Early Cambrian: U–Pb ages of volcanic ashes from Avalon andGondwana. Canadian Journal of Earth Sciences 35, 329–338.
Landing, E., Westrop, S.R., Kim, D.H., 2003. First Middle Ordovician biota from southernNew Brunswick: stratigraphic and tectonic implications for the evolution of theAvalon continent. Canadian Journal of Earth Sciences 40, 715–730.
Landing, E., Peng, S.C., Babcock, L.E., Geyer, G., Moczydłowska-Vidal, M., 2007a. Globalstandard names for the lowermost Cambrian series and stage. Episodes 30,283–289.
Landing, E., Franzi, D.A., Hagadorn, J.W., Westrop, S.R., Kröger, B., Dawson, J., 2007b.Cambrian of east Laurentia: field workshop in eastern New York and westernVermont. In: Landing, E. (Ed.), Ediacaran–Ordovicianp of East Laurentia―S.W.Ford Memorial VolumeNew York State Museum Bulletin 510, 25–80.
Landing, E., Johnson, S.C., Geyer, G., 2008. Faunas and Cambrian volcanism on theAvalonian marginal platform, southern New Brunswick. Journal of Paleontology82, 884–905.
Landing, E., English, A., Keppie, J.D., 2010. Cambrian origin of all skeletalized metazoanphyla—discovery of Earth's oldest bryozoans (Upper Cambrian, southern Mexico).Geology 38, 547–550.
Landing, E., Westrop, S.R., Adrain, J.M., 2011. The Lawsonian Stage—the Eoconodontusnotchpeakensis (Miller, 1969) FAD and HERB carbon isotope excursion define aglobally correlatable terminal Cambrian stage. Bulletin of Geoscience 86, 621–640.
Landing, E., Mohibullah, M., Williams, M., 2013a. First Middle Ordovician ostracodsfrom western Avalonia: paleogeographic and paleoenvironemntal significance.Journal of Paleontology 87, 269–276.
Landing, E., Westrop, S.R., Bowring, S.A., 2013b. Reconstructing the Avaloniapalaeocontinent in the Cambrian: A 519 Ma caliche in South Wales and
transcontinental middle Terreneuvian Epoch sandstones. Geological Magazine15 pp. http://dx.doi.org/1017/S001676813000228.
Lazarenko, N.P., 1957. Novye dannye o trilobitakh roda Triangulaspis. Sbornik statey popaleontologii i biostratigrafii, 3, pp. 3–17 (in Russian).
Lazarenko, N.P., 1962. Novye nizhnekembriyskie trilobity Sovetskoy Arktiki. Sbornikstatey po paleontologii i biostratigrafii, 29, pp. 29–77 (in Russian).
Lazarenko, N.P., Repina, L.I., 1983. Razrez pogranichnykh sloev nizhnego i srednegokembriya vostochnogo kryla Bulkurskoy antiklinali (khr. Tuora-Sis, SevernoeVerkhoyan'e). In: Repina, L.N., Suvorova, N.P. (Eds.), Biostratigrafiya i faunapogranichnykh otlozheniy nizhnego i srednego kembriya Sibiri. : Nauk SSSR,Trudy institut geologii i geofizikii, Sibirskoe otdelenie, 548. Nauka, Novosibirsk,pp. 22–35.
Łendzion, K., 1977. First gastropod fauna from the Klimontovian Stage (Lower Cambri-an) of south-eastern Poland. Kwartalnik Geologniczny 21, 235–243.
Li, D., Ling, H.F., Jiang, S.Y., Pan, J.Y., Chen, Y.Q., Cai, Y.F., Feng, H.Z., 2009. New carbonisotope stratigraphy of the Ediacaran–Cambrian boundary interval from SWChina: implications for global correlation. Geological Magazine 146, 465–484.
Li, G.X., Zhao, D., Gubanov, A., Zhu, M.Y., Na, L., 2011. Early Cambrian molluskWatsonella crosbyi: a potential GSSP index fossil for the base of Cambrian Stage2. Acta Geologica Sinica 85, 309–319.
Li, D., Ling, H.-F., Shields-Zhou, G.A., Chen, X., Cremonese, L., Och, L., Thirwall, M., Manning,C.J., 2012. Carbon and strontium isotope evolution of seawater across the Ediacaran–Cambrian transition: evidence from the Xiaotan section, NE Yunnan, South China. Pre-cambrian Research. http://dx.doi.org/10.1016/j.precamres.2012.01.002.
Lieberman, B.S., 2001. Phylogenetic analysis of the Olenellina Walcott, 1890 (Trilobita,Cambrian). Journal of Paleontology 75, 96–115.
Lieberman, B.S., 2002. Phylogenetic analysis of some basal early Cambrian trilobites,the biogeographic origins of the Eutrilobita, and the timing of the Cambrian radia-tion. Journal of Paleontology 76, 692–708.
Liñán Guijarro, E., 1978. Bioestratigrafía de la Sierra de Córdoba. (Ph.D. dissertation)Facultad de Ciencias: Departamento Paleontológico, Universidad de Granada(212 pp.).
Liñán, E., Sdzuy, K., 1978. A trilobite from the Lower Cambrian of Córdoba (Spain) andits stratigraphical significance. Senckenbergiana lethaea 59, 378–399.
Liñán, E., Perejón, A., Sdzuy, K., 1993. The Lower–Middle Cambrian stages and stratotypesfrom the Iberian Peninsula: a revision. Geological Magazine 130, 817–833.
Liñán, E., Dies, M.E., Gámez Vintaned, J.A., Gozalo, R., Mayoral, E., Muñiz, F., 2005. LowerOvetian (Lower Cambrian) trilobites and biostratigraphy of the Pedroche Forma-tion (Sierra de Córdoba, southern Spain). Geobios 38, 365–381.
Liñán, E., Gámez Vintaned, J.A., Gozalo, R., Dies, M.E., Mayoral, E., 2006. Events and bio-stratigraphy in the Lower Cambrian of Iberia. Zeitschrift der deutschen Gesellschaftfür Geowissenschaften 157, 597–609.
Linnarsson, J.G.O., 1871. Omnågra försteningar från Sveriges och Norges »Primordialzon«.Öfversigt af konglika Vetenskaps-Akademiens Förhandlingar (6), 789–796.
Linnemann, U., Romer, R.L., Pin, C., Aleksandrowski, P., Buła, Z., Geisler, T., Kachlik, V.,Krzemińska, E., Mazur, S., Motuza, G., Murphy, J.B., Nance, R.D., Pisarevsky, A.,Schulz, B., Ulrich, J., Wiszniewska, J., Żaba, J., Zeh, A., 2008. Precambrian. In:McCann, T. (Ed.), The Geology of Central Europe. Volume 1: Precambrian andPalaeozoicGeological Society, London 21–101.
Lochman-Balk, C., Wilson, J.L., 1958. Cambrian biostratigraphy in North America. Jour-nal of Paleontology 32, 312–350.
Lu, Y.H., 1940. On the ontogeny and phylogeny of Redlichia intermedia Lu (sp. nov.).Bulletin of the Geological Society of China 20, 333–341 (in Chinese and English).
Lu, Y.H., 1941. Lower Cambrian stratigraphy and trilobite fauna of Kunming, Yunnan.Bulletin of the Geological Society of China 21, 71–90 (in Chinese and English).
Lu, Y.H., Chang, W.T., Chien, Y.Y., Chu, C.L., Lin, H.L., Zhou, Z.Y., Qian, Y., Zhang, S.G., Yuan,J.L., 1974. Cambrian trilobites. Handbook of Stratigraphy and Palaeontology of South-west China. Science Press, Beijing, pp. 82–107 (in Chinese, with English summary).
Luo, H.L., 1981. Trilobites from the Chiungchussu Formation (Lower Cambrian) inMeishucun of Jinning, Yunnan Province. Acta Palaeontologica Sinica 20, 331–340(in Chinese, with English abstract).
Luo, H.L., Jiang, Z.W., Xu, Z.J., Song, X.L., Xue, X.F., Luo, H.L., Jiang, Z.W., Xu, Z.J., Song,X.L., Xue, X.F., 1980. On the Sinian–Cambrian boundary of Meishucun andWangjiawan, Jinning County, Yunnan. Acta Geologica Sinica, pp. 95–111 (in Chi-nese, with English summary).
Luo, H.L., Jiang, Z.W., Wu, X.C., Song, X.L., Ouyang, L., et al., 1982. The Sinian–CambrianBoundary in Eastern Yunnan, China.People's Publishing House, Kunming, Yunnan,China (265 pp. in Chinese, with English summary).
Luo, H.L., Jiang, Zh.W., Wu, X.C., Song, X.L., Ouyang, L., Xing, Y.Sh, Liu, G.Z., Zhang, S.S.,Tao, Y.H., 1984. Sinian–Cambrian Boundary Stratotype Section at Meishucun,Jinning, Yunnan, China.People's Publishing House, Kunming, Yunnan, China (154pp. in Chinese and English).
Maloof, A.C., Schrag, D.P., Crowley, J.L., Bowring, S.A., 2005. An expanded record of EarlyCambrian carbon recycling from the Anti-Atlas margin. Canadian Journal of EarthSciences 42, 2195–2216.
Maloof, A.C., Ramezani, J., Bowring, S.A., Fike, D.A., Porter, S.M., Mazouad, M., 2010a.Constraints on early carbon cycling from the duration of the Nemakit-Daldynian–Tommotian boundary δ13C shift, Morocco. Geology 38, 623–626.
Maloof, A.C., Porter, S.H., More, J.L., Dudás, F.Ö., Bowring, S.A., Higgins, J.A., Fike, D.A.,Eddy, M.P., 2010b. The earliest Cambrian record of animals and ocean geochemicalchange. Geological Society of America Bulletin 122, 1731–1774.
Mamet, B., 1977. In: Kaufman, E.G., Hazel, J.E. (Eds.), Foraminiferal Zonation of the LowerCarboniferous: Methods and Stratigraphic Implications. Dowden, Hutchinson & Ross,Stroudsburg, Pennsylvania, pp. 445–462.
Mansuy, H., 1912. Étude géologique du Yunnan oriental. Part 2: Paléontologie.Mémoires du Service Géologique de l'Indochine 1 (146 pp.).

169E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Margaritz, M., Holser, W.T., Kirschvink, J.L., 1986. Carbon-isotope events across thePrecambrian–Cambrian boundary on the Siberian Platform. Nature 320 (258), 259.
Marshall, C.R., 1990. Confidence intervals on stratigraphic ranges. Paleobiology 16,1–10.
Matthew, G.F., 1889. Sur le développement des prémiers trilobites. Société RoyaleMalacologique de Belgique, Annales, 23, pp. 351–362.
Matthew, G.F., 1890. On Cambrian organisms in Acadia. Transactions of the Royal Soci-ety of Canada 7, 135–162 (Section IV).
Matthew, G.F., 1896. Notes on the Cambrian faunas—the genus Microdiscus. AmericanGeologist 18, 28–31.
Matthew, G.F., 1899. A Palaeozoic terrane beneath the Cambrian. Annals of the NewYork Academy of Science 12, 41–56.
McCollum, L.B., Sundberg, F.A., 2007. Cambrian trilobite biozonation of the LaurentianDelamaran Stage in the southern Great Basin, U.S.A.: implications for global corre-lations and defining a Series 3 global boundary stratotype. Memoires of the Asso-ciation of Australasian Palaeontologists 34, 147–156.
McIlroy, D., Brasier, M.D., Moseley, J.B., 1998. The Proterozoic–Cambrian transitionwithin the ‘Charnian Supergroup’ of central England and the antiquity of theEdiacara fauna. Journal of the Geological Society of London 155, 401–411.
McKerrow, W.S., Scotese, C.R., Brasier, M.D., 1992. Early Cambrian continental recon-structions. Journal of the Geological Society of London 149, 599–606.
McMenamin, M.A.S., 1987. Lower Cambrian trilobites, zonation, and correlation ofthe Puerto Blanco Formation, Sonora, Mexico. Journal of Paleontology 61,738–749.
Mens, K., Pirrus, E., 1997. Cambrian. In: Raukas, A., Teedumäe, A. (Eds.), Geology andMineral Resources of Estonia. Estonian Academic Publishers, Tallinn, pp. 39–51.
Meshkova, N.P., Zhuravleva, I.T., Luchinina, V.A., 1973. Nizhniy kembriy i niznaya chast'srednego kembriya Olenekskogo podnyatiya. In: Zhuravleva, I.T. (Ed.), Problemypaleontologii i biostratigrafii nizhnego kembriya Sibiri i Dal'nego Vostok. : Trudyinstitut geologii i geofizikii, Sibirskoe otdelenie, Akademiya Nauk SSSR, 49.Nauka, Novosibirsk, pp. 194–216 (in Russian).
Missarzhevsky, V.V., 1973. Konodontoobraznie organizma iz pogranichnyk sloevkembriya i dokembriya Sibirskoy platformi i Kazakhstana. . Editor not listedProblemy paleontologii i biostratigrafiya nizhnego kembriya Sibiri i Dal'negoVostoka. Nauka, Novosibirsk, pp. 53–58 (in Russian).
Missarzhevsky, V.V., 1974. Noviye dannye o drevyneyskikh okamenelostyakh rannegokembriya Sibirskoy platformi. In: Zhuravleva, I.T., Rozanov, A.Yu. (Eds.),Biostratigrafiya i paleontologiya nizhnego kembriya Evropii i Severnoy Azii.Nauka, Novosibirsk, pp. 179–189 (in Russian).
Missarzhevsky, V.V., 1980. O progranichnykh sloyakh kembriya i dokembriyazapadnogo sklona Olenkskogo podnyatiya (r. Olenek). Byulletin MoskovskogoObshchstva Spytateley Priordy, Otdelenie geologii, 55, pp. 23–34 (in Russian).
Missarzhevsky, V.V., 1983. Stratigrafiya drevneyshich tolshch fanerozoya Anabaraskogomassiva. Sovetskaya Geologiya 9, 62–73 (in Russian).
Missarzhevsky, V.V., Mambetov, A.M., 1981. Stratigrafiya i fauna pogranichnykh sloevkembriya dokembriya Malogo Karatau. Akademiya Nauk SSSR. : Trudy InstitutGeologii, 326. Nauka, Moscow (90 pp. in Russian).
Missarzhevsky, V.V., 1989. Drevneyskie skeletnye okamenelosti i stratigrafiyapogranichnykh tolshch dokembriya i kembriya. Trudy Ordena Trudvogo KrasnogoZnameni Geologischeskogo Instituta Akademiya Nauk SSSR 443 238 pp. (inRussian).
Moberg, J.C., 1892. Om Olenellusledet i sydliga Skandinavien. Forhandlungen ved deSkandinaviske naturforskeres 14:de møde, København. 434–439 (in Swedish).
Moberg, J.C., 1899. Sveriges älsta kända trilobiter. Geologiska Föreningens i StockholmFörhandlingar 21, 309–348 (in Swedish).
Moczydłowska, M., 1981. Lower and Middle Cambrian acritarchs from northeasternPoland. Precambrian Research 15, 63–74.
Moczydłowska, M., 1988. New Lower Cambrian acritarchs from Poland. Reviews inPalaeobotany and Palynology 54, 1–10.
Moczydłowska, M., 1991. Acritarch biostratigraphy of the Lower Cambrian and thePrecambrian–Cambrian boundary in southeastern Poland. Fossils & Strata 29 (127 pp.).
Moczydłowska, M., 1998. Cambrian acritarchs from Upper Silesia, Poland: biochronologyand tectonic implications. Fossils & Strata 46 (121 pp.).
Moczydłowska, M., Vidal, G., 1986. Lower Cambrian acritarch zonation in southernScandinavia and southeastern Poland. Geologiska Föreningens i StockholmFörhandlingar 108, 201–223.
Moczydłowska, M., Vidal, G., 1988. How old is the Tommotian? Geology 16, 166–168.Moczydłowska, M., Yin, L.M., 2012. The lower Cambrian phytoplankton record in China
and global perspective for stage subdivision. In: Zhao, Y.L., Zhu, M.Y., Peng, J.,Gaines, R.R., Parsley, R.L. (Eds.), Cryogenian–Ediacaran to Cambrian Stratigraphyand Paleontology of Guizhou, ChinaJournal of Guizhou University (Natural Sci-ences) 29, 176–177.
Moczydłowska, M., Zang, W.L., 2006. The Early Cambrian acritarch Skiagia and its sig-nificance for global correlation. Palaeoworld 15, 328–347.
Moczydłowska, M., Landing, E., Zang, W., Palacios, T., 2011. Proterozoic phytoplanktonand timing of chlorophyte algae origins. Palaeontology 54, 721–733.
Mount, J.F., Signor, P.W., 1992. Faunas and facies—fact and artifact. Paleoenvironmentalcontrols on the distribution of Early Cambrian faunas. In: Lipps, J.H., Signor, P.W.(Eds.), Origins and Early Evolution of Metazoa. Plenum Press, New York, pp. 27–51.
Murphy, J.B., Keppie, J.D., Dostal, J., Nance, R.D., 1999. Neoproterozoic–Early Paleozoicevolution of Avalonia. In: Ramos, V.A., Keppie, J.D. (Eds.), Laurentia–GondwanaConnections Before PangaeaGeological Society of America, Special Paper 336,253–266.
Myannil, R.M., 1966. A small vertically excavated cavity in Baltic Ordovician limestone.. Editor not listed Organizm i sreda v geologischeskom proshlom. : AkademiyaNauk SSSR. Paleontologishe Institut, Moscow, pp. 200–207 (in Russian).
Myrow, P.M., Landing, E., 1992. Mixed siliciclastic–carbonate deposition in a LowerCambrian oxygen-stratified basin, Chapel Island Formation, southeastern New-foundland. Journal of Sedimentary Petrology 62, 455–473.
Narbonne, G.M., Myrow, P.M., 1988. Trace fossil biostratigraphy in the Precambrian–Cambrian boundary interval. In: Landing, E., Narbonne, G.M., Myrow, P. (Eds.),Trace Fossils, Small Shelly Fossils, and the Precambrian–Cambrian BoundaryNewYork State Museum Bulletin 463, 72–76.
Narbonne, G.M., Myrow, P., Landing, E., Anderson, M.M., 1987. A candidate stratotypefor the Precambrian–Cambrian boundary, Fortune Head, Burin Peninsula, south-eastern Newfoundland. Canadian Journal of Earth Sciences 24, 1277–1293.
Naumova, S.N., 1960. Sporovo-pyl'tsevye kompleksy rifeyskikh i nizhnekembriyskikhotlozheniy SSSR. In: Shatsky, N.S. (Ed.), Stratigrafiya pozdnego dokembriya ikembriya: Mezhdunarodnyj Geologicheskij Kongress, XXI Sessiya, DokladySovetskikh Geologov, Problemy, 8. Akademiya Nauk SSSR, Moscow, pp. 109–117(in Russian).
Nelson, C.A., 1971. Geologic map of the Waucoba Spring quadrangle, Inyo County,California. U.S. Geological Survey Map GQ 921..
Nelson, C.A., 1978. Late Precambrian–Early Cambrian stratigraphic and faunal succes-sion of eastern California and the Precambrian–Cambrian boundary. GeologicalMagazine 115, 121–126.
Nelson, C.A., Hupé, P., 1964. Sur l'éxistence de Fallotapsis et Daguinaspis, trilobites mar-ocains, dans le Cambrien inférieur de Californie, et sés conséquences. ComptesRendus de l'Académie des Sciences 258, 621–623 (Paris).
Neltner, L., Poctey, N., 1950. Quelques faunes géorgiennes du Maroc. Notes etMémoires du Service Géologique du Maroc, 2, pp. 53–83.
Nicoll, R.S., Miller, J.F., Nowlan, G.S., Repetski, J.E., Ethington, R.L., 1999. Iapetonudus(new genus) and Iapetognathus Landing, unusual earliest Ordovician multielementtaxa and their utility for biostratigraphy. Brigham Young University Geology Stud-ies, 44, pp. 27–101.
Nielsen, A.T., Schovsbo, N.H., 2011. The Lower Cambrian of Scandinavia: depositional en-vironment, sequence stratigraphy and palaeogeography. Earth-Science Reviews 107,207–310.
Nikiforov, N.I., 1974. Opisanie trilobitov. In: Repina, L.N., Lazarenko, N.P., Meshkova,N.P., Korshunov, V.I., Nikiforov, N.I., Aksarina, N.A. (Eds.), Biostragrafiya i faunanizhnego kembriya Karaulakha (kh. Tuora Sis): Akademiya Nauk SSSR, Sibiriskoyotdelenie, Trudy institut geologii i geofiziki, 235 (298 pp.).
North American Commission on Stratigraphic Nomenclature, 1983. North Americanstratigraphic code. American Association of Petroleum Geologists Bulletin 67,841–875.
Nowlan, G.S., Narbonne, G.M., Fritz, W.H., 1985. Small shellly fossils and trace fossilsnear the Precambrian–Cambrian boundary in the Yukon Territory, Canada. Lethaia18, 233–256.
Okuneva, O.G., Repina, L.N., 1973. Biostratigrafiya i fauna kembriya Primor'ya.Akademiya Nauk SSSR, Sibirskoe otdelenie, Trudy institut geolgii i geofiziki, 37.Nauka, Novosibirsk (284 pp. (in Russian)).
Öpik, A., 1926. Über estländischen Blauen Ton. Sitzungsberichte der naturforschendenGesellschaft der Universität Tartu, 33, pp. 1–57.
Pacześna, J., 1985. Skamienałości śladowe górnego wendu i dolnego kambrupołudniowej Lubelszczyzny. Kwartalnik Geologie 29, 255–270.
Palacios Medrano, T., 1989. Microfósiles de pared orgánica del Proterozóico superior(región central de la Península Ibérica). Memorias del Museo Paleontológico dela Universidad de Zaragoza, 3 (91 pp. (in Spanish)).
Palacios, T., Delgado, D., 1999. Acritarch assemblages at the Lower–Middle Cambrianboundary in the Iberian Peninsula and their utility in global correlation. Journalof Conference Abstracts 4 (3), 1017.
Palacios, T., Moczydłowska, M., 1998. Acritarch biostratigraphy of the Lower–MiddleCambrian boundary in the Iberian Chains, Province of Soria, northeastern Spain.Revista Española Paleontologicas, Número Extraordinario en Homage al ProfessorGonzalo Vidal 65–82.
Palacios, T., Vidal, G., 1992. Lower Cambrian acritarchs from northern Spain: thePrecambrian–Cambrian boundary and biostratigraphic implications. GeologicalMagazine 129, 421–436.
Palacios, T., Jensen, S., Barr, S.M., White, C.E., Miller, R.F., 2011. Newbiostratigraphical constraints on the Lower Cambrian Ratcliffe Brook Formation,southern New Brunswick, Canada, from organic-walled microfossils. Stratigra-phy 8, 45–60.
Paliy, V.M., 1976. Ostatki besskeletnoy fauny i sledy zhiznedeyatel'nosti iz nizhnegokembriya Podolii. In: Rybenko, V.A. (Ed.), Paleontologiya i stratigrafiya verhnegodokembriya i nizhnego paleozoya yuga-zapada Vostochno-Evropeyskoyplatformy. Nauk Dumka, Kiev, pp. 63–77 (in Russian).
Palmer, A.R., 1998a. A proposed nomenclature for stages and series for the Cambrian ofLaurentia. Canadian Journal of Earth Sciences 35, 323–333.
Palmer, A.R., 1998b. Why is international correlation in the Lower Cambrian so diffi-cult? Revista Española Paleontologica, Número Extraordinario en Homage al Pro-fessor Gonzalo Vidal. 17–21.
Palmer, A.R., Repina, L.N., 1993. Through a glass darkly: taxonomy, phylogeny, and bio-stratigraphy of the Olenellina. University of Kansas PaleontologicalContribributions, New Series, 3 (35 pp.).
Parkhaev, P.Yu, 2001. Mollusks and siphonoconchs. In: Alexander, E.M., Jago, J.B.,Rozanov, A.Yu., Zhuravlev, A.Yu. (Eds.), The Cambrian Biostratigraphy of theStansbury Basin, South Australia. Nauka, Moscow, pp. 133–210.
Parkhaev, P.Yu., 2007. Shell chirality in Cambrian gastropods and sinistral members ofthe genus Aldanella Vostokova, 1962. Paleontologicheskii Zhurnal 2007 (3), 3–9.
Parkhaev, P.Yu., Demidenko, Yu.E., 2010. Zooproblematica and Mollusca from theLower Cambrian Meishucun section (Yunnan, China) and taxonomy and systemat-ics of the Cambrian small shelly fossils. Palaeontological Journal 44, 883–1161.

170 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Parkhaev, P.Yu., Karlova, G.A., 2011. Taxonomic revision and evolution of Cambrianmollusks of the genus Aldanella Vostokova, 1962 (Gastropoda: Archaeobranchia).Paleontological Journal 45, 1145–1205.
Parkhaev, P.Yu., Karlova, G.A., Rozanov, A.Yu., 2011. Taxonomy, stratigraphy and bioge-ography of Aldanella attleborensis—a possible candidate for defining the base ofCambrian Stage 2. Museum of Northern Arizona Bulletin 67, 298–300.
Patel, I.M., 1977. Late Precambrian fossil soil horizon in southern New Brunswick, and itsstratigraphic significance. Geological Society of America, Abstracts with Programs 9,308.
Patzkowsky, M.E., Holland, S.M., 2012. Stratigraphic Paleobiology. Understanding theDistribution of Fossils in Time and Space. University of Chicago Press. 256 pp.
Pegel, T.V., 2000. Evolution of trilobite biofacies in Cambrian basins of the Siberian Plat-form. Journal of Paleontology 74, 1000–1019.
Peng, S.C., 2000. Cambrian of slope facies. In: Nanjing Institute of Geology and Palaeontology,the Chinese Academy of Sciences (Ed.), Stratigraphical Studies in China (1979–1999).China University of Technological Science, Hefei, pp. 23–38 (in Chinese).
Peng, S.C., 2003. Chronostratigraphic subdivision of the Cambrian of China. GeologicaActa 1, 135–144.
Peng, S.C., 2009. The newly-developed Cambrian biostratigraphic succession andchronostratigraphic scheme for South China. Chinese Science Bulletin 54,4161–4170.
Peng, S.C., Babcock, L.E., 2001. Cambrian of the Hunan–Guizhou region, South China. In:Peng, S.C., Babcock, L.E., Zhu, M.Y. (Eds.), Cambrian System of South China:Palaeoworld, 13, pp. 3–51.
Peng, S.C., Babcock, L.E., 2005a. Two Cambrian agnostoid trilobites, Agnostotes orientalis(Kobayashi, 1935) and Lotagnostus americanus (Billings, 1860): key species for de-fining global stages of the Cambrian System. Geoscience Journal 9, 105–119.
Peng, S.C., Babcock, L.E., 2005b. Towards a new global subdivision of the Cambrian Sys-tem. Journal of Stratigraphy 29 (171–178), 204.
Peng, S.C., Babcock, L.E., 2011. Continuing progress on chronostratigraphic subdivisionof the Cambrian System. Bulletin of Geosciences 86, 391–396.
Perejón, A., 1986. Bioestratigrafía de los arqueociatos en España. Cuadernos GeologicoaIbérica, 9, pp. 212–266.
Perejón, A., 1994. Palaeogeographic and biostratigraphic distribution of Archaeocyathain Spain. Courier Forschungsinstut Senckenberg, 172, pp. 341–354.
Pfennig, D.W., Wund, M.A., Snell-Rood, E.C., Cruickshank, T., Schlichting, C.D., Moczek,A.P., 2010. Phenotypic plasticity's impacts on diversification and speciation. Trendsin Ecological Evolution 25, 459–467.
Pillola, G.L., 1993. The Lower Cambrian trilobite Bigotina and allied genera.Palaeontology 36, 855–881.
Pillola, G.L., Gámez Vintaned, J.A., Dabard, M.P., Leone, F., Liñán, E., Chauvel, J.-J., 1994.The Lower Cambrian ichnospecies Astropolichnus hispanicus: palaeoenvironmentand palaeogeographical significance. Bolletino Societá Paleontologica Italiana, Spe-cial, volume 2, pp. 253–267.
Piper, J.D.A., 1987. Paleomagnetism and the Continental Crust. Open University Press,Milton Keynes, 372 pp.
Pojeta Jr., J., Runnegar, B., 1976. The paleontology of rostroconch mollusks and the earlyhistory of the PhylumMollusca. United States Geological Survey Professional Paper968 (88 pp.).
Qian, Y. (Ed.), 1999. Taxonomy and Biostratigraphy of Small Shelly Fossils in China. Sci-ence Press, Beijing (247 pp.).
Qian, Y., Bengtson, S., 1989. Palaeontology and biostratigraphy of the Early CambrianMeishucunian Stage in Yunnan Province, south China. Fossils & Strata 24 (156 pp.).
Qian, Y., Yu,W., Liu, D.Y., Wang, Z.Z., 1985. Restudy of the Precambrian–Cambrian boundarysection at Meishucun of Jinnin, Yunnan. Kexue Tongbao 15, 928–931 (in Chinese).
Qian, Y., Zhu, M.Y., He, T.G., Jiang, Z.W., 1996. New investigation of Precambrian–Cambrian boundary sections in eastern Yunnan. Acta Micropalaeontologica Sinica13, 225–240 (in Chinese with English summary).
Qian, Y., Zhu, M., Li, G., Jiang, Z., Van Iten, H., 2002. A supplemental Precambrian–Cambrian boundary stratotype section in SW China. Acta Palaeontologica Sinica41, 19–26 (in Chinese with English summary).
Rasetti, F., 1967. Lower and Middle Cambrian trilobite faunas from the Taconic se-quence of New York. Smithsonian Miscellaneous Collections 152 (4) (111 pp.).
Repina, L.N., 1960. Opisanie trilobitov. In: Khalfin, L.L. (Ed.), Biostratigrafiya paleozoyaSayano-Altayskoy gornoy oblasti. Tom 1, Nizhniy paleozoy. Trudy SibirskogoNauchno-Issledovatel’skogo Instituta Geologii, Geofoziki i Mineral’nogo Syr’ya(SNIIGGiMS), 19. Nauka, Novosibirsk, 152–253.
Repina, L.N., 1964. Paleontologicheskiy atlas. 2, Trilobity. In: Repina, L.N., Khomentovsky,V.V., Zhuravleva, I.T., Rozanov, A.Yu. (Eds.), Biostratigrafiya nizhnego kembriyaSayano–Altayskoy skladchatoy oblasti. : Akademiya Nauk SSSR, Institut geologii igeofizikii, Sibirskoe otdelenie. Nauka, Moscow, pp. 252–362 (in Russian).
Repina, L.N., 1981. Trilobite biostratigraphy of the Lower Cambrian stages ofSiberia. In: Taylor, M.E. (Ed.), Short Papers for the Second International Sympo-sium on the Cambrian System: U.S. Geological Survey, Open-File Report, 81-743, pp. 173–180.
Repina, L.N., 1986. Planetarnaia korreliatsiia iarusnikh podrazdeleniia nizhnegokembriya po trilobitam. In: Zhuravleva, I.T. (Ed.), Biostratigrafiia i paleontologiiakembriya svernoi Azii: Akademiya Nauk SSSR, Sibiriskoe otdelenie, Trudy Institutgeologii i geofizikii, 699 (240 pp. (in Russian)).
Repina, L.N., Romanenko, E.V., 1978. Trilobity i stratigrafiya nizhnego kembriya Altaya.Akademiya Nauk SSSR, Sibirskoe otdelenie, Trudy Institut geologii i geofizikii, 382.Nauka, Moscow (303 pp.).
Repina, L.N., Rozanov, A.Yu. (Eds.), 1992. Kembrii Sibiri. Nauka, Novosibirsk (135 pp.(in Russian)).
Repina, L.N., Lazarenko, N.P., Meshkova, N.P., Korshunov, V.I., Nikiforov, N.I., Aksarina,N.A., 1974. Biostratigrafiya i fauna nizhnego kembriya Kharaulakha (kh. Tuora-
Sis). Akademiya Nauk SSSR, Sibirskoe otdelenie, Trudy institut geologii i geofizikii,235 (298 pp. (in Russian)).
Richter, R., Richter, E., 1940. Die Saukianda-Stufe von Andalusien, eine fremde Fauna imeuropäischen Ober-Kambrium. Abhandlungen der Senckenbergische NaturforschendeGesellschaft 450 (88 pp.).
Richter, R., Richter, E., 1941. Die Fauna des Unter-Kambriums von Cala in Andalusien.Abhandlungen der Senckenbergiana Naturforschung Gesellschaft 455 (90 pp.).
Robison, R.A., Rosova, A.V., Rowell, A.J., Fletcher, T.P., 1977. Cambrian boundaries anddivisions. Lethaia 10, 257–262.
Rowland, S.M., Luchinina, V.A., Korovnikov, I.V., Sipin, D.P., Tarletskov, A.I., Fedoseev,A.V., 1998. Biostratigraphy of the Vendian–Cambrian Sukharikha River section,northwestern Siberia. Canadian Journal of Earth Sciences 35, 339–352.
Rozanov, A.Yu, 1966. Problema nizhney granitsy kembriya. In: Keller, B.M. (Ed.), Itoginauka. Obshchaya geologiya. Stratigrafiya. VINITI Press, Moscow, pp. 92–111 (inRussian).
Rozanov, A.Yu., 1967. The Cambrian lower boundary problem. Geological Magazine104, 415–434.
Rozanov, A.Yu, Missarzhevsky, V.V., 1966. Biostratigrafiya i fauna nizhnych gorizontovkembriya. Akademiya Nauk SSSR, Geologicheskiy Institut, Trudy, 148. Nauka,Moscow (126 pp. (in Russian)).
Yarusnoe raschlenenie nizhnego kembriya Sibiri. Stratigrafiya. In: Rozanov, A.Yu,Sokolov, B.S. (Eds.), Akademiya Nauk SSSR, Otdelenie geologii, geofiziki,geokhimiki. Nauka, Moscow (183 pp. (in Russian)).
Rozanov, A.Yu, Zhuravlev, A.Yu, 1996. The Lower Cambrian fossil record of the SovietUnion. In: Lipps, J.H., Signor, P.W. (Eds.), Origin and Early Evolution of the Metazoa.Topics in Geobiology, 10. Plenun Press, New York and London, pp. 206–282.
Rozanov, A.Yu, Missarzhevsky, V.V., Volkova, N.A., Voronova, L.G., Krylov, I.N., Keller,B.M., Korolyuk, I.K., Lendzion, K., Michniak, R., Pykhova, N.G., Sidorov, A.D., 1969.The Tommotian Stage and the Cambrian Lower Boundary Problem. Nauka,Moscow.Amerind Publishing Co., New Delhi, India (1981, in English, 359 pp.).
Rozanov, A.Yu, Khomentovsky, V.V., Shabanov, Ya.Yu, 2008. To the problem of stage sub-division of the Lower Cambrian. Stratigraphy and Geological Correlation 16, 1–19.
Rozanov, A.Yu., Parkhaev, P.Yu., Demidenko, Yu., Skorlotova, N.A., 2011. Mobergellaradiolata—a possible candidate for defining the base of Cambrian Series 2 andStage 3. Museum of Northern Arizona Bulletin 67, 304–306.
Rushton, A.W.A., 1966. The Cambrian trilobites from the Purley Shales of Warwick-shire. Palaeontographical Society Monographs 120 (511) (55 pp.).
Rushton, A.W.A., 1974. The Cambrian of Wales and England. In: Holland, C.H. (Ed.),Cambrian of the British Isles Norden, and Spitsbergen, Lower Palaeozoic Rocks ofthe World, vol. 2. John Wiley & Sons, New York, pp. 43–121.
Rushton, A.W.A., Bruck, P.M., Williams, M., Woodcock, N.H., 2011. Revised correlationof the Cambrian rocks in the British Isles. Geological Society, London, Special Re-port 25 (64 pp.).
Salvador, A. (Ed.), 1994. International Stratigraphic Guide. A Guide to StratigraphicClassification, Terminology, and Procedure, Second edition. International Unionof Geologists and the Geological. Society of America (214 pp.).
Sánchez-Zavala, J.L., Centenero-García, E., Ortega-Gutiérrez, F., 1999. Review of Paleo-zoic stratigraphy of México and its role in the Gondwana–Laurentia connections.In: Ramos, V.A., Keppie, J.D. (Eds.), Laurentia–Gondwana Connections Before Pan-gaea: Geological Society of America, Special Paper, 336, pp. 211–226.
Schmidt, F., 1888. Über eine neu entdeckte unterkambrische Fauna in Estland. Mémoirsde l'Académie Impériale des Sciences, St.-Pétersbourg, (VII), 36, pp. 1–27.
Schröder, S., Grotzinger, J.P., Amthor, J.E., Matter, A., 2005. Carbonate deposition andhydrocarbon reservoir development at the Precambrian–Cambrian boundary: theAra Group in South Oman. Sedimentary Geology 180, 1–28.
Scotese, C.R., Bambach, R.K., Barton, C., Van der Voo, R., Ziegler, A.M., 1979. Palaeozoicbase maps. Journal of Geology 87, 217–277.
Sdzuy, K., 1961. Das Kambrium Spaniens. Teil II: Trilobiten. 1. Abschnitt. Akademie derWissenschaft und Literatur, Mainz, Abhandlungen der mathematisch-naturwissenschaftlichen Klasse, 7, pp. 499–594.
Sdzuy, K., 1962. Über das Entzerren von Fossilien (mit Beispielen aus derunterkambrischen Saukianda-Fauna). Paläontologische Zeitschrift 36, 275–284.
Sdzuy, K., 1978. The Precambrian–Cambrian boundary beds in Morocco (PreliminaryReport). Geological Magazine 115, 83–94.
Sdzuy, K., 1981. Der Beginn des Phanerozoikums — paläobiologische undstratigraphische Probleme. Natur und Museum 111, 390–399.
Sdzuy, K., Geyer, G., 1988. The base of the Cambrian in Morocco. In: Jacobshagen, V.H.(Ed.), The Atlas System of Morocco. Studies on Its Geodynamic Evolution: LectureNotes in Earth Sciences, 15, pp. 91–106.
Seilacher, A., 1955. Spuren und Fazies im Unterkambrium. In: Schindewolf, O.H.,Seilacher, A. (Eds.), Beiträge zur Kenntnis des Kambrium in der Salt Range (Paki-stan): Akademie der Wissenschaft und Literatur, Abhandlungen dermathematisch-naturwissenschaftlichen Klasse, 10, pp. 373–399.
Seilacher, A., 1956. Der Beginn des Kambriums als biologische Wende. Neues Jahrbuchfür Geologie und Paläontologie (Abhandlungen) 103, 155–180 (in German).
Seilacher, A., 1970. Sedimentary rhythms and trace fossils in Paleozoic sandstones ofLibya. In: Kanes, W.H. (Ed.), Geology, Archeology and Prehistory of the Southwest-ern Fezzan, Libya. Petroleum Exploration Society of Libya, 11th Annual Field Con-ference 1969, Tripoli, pp. 117–123.
Seilacher, A., Buatois, L.A., Mángano, M.G., 2005. Trace fossils in the Ediacaran–Cambriantransition: behavioral diversification, ecological turnover and environmental shift.Palaeogeography, Palaeoclimatology, Palaeoecology 227, 323–356.
Shaler, N.S., Foerste, A.F., 1888. Preliminary description of North Attleboro fossils.Harvard Museum of Comparative Zoology Bulletin 16, 27–41.
Sheehan, P.M., 1996. A new look at ecologic evolutionary units. Palaeogeography,Palaeoclimatology, Palaeoecology 127, 21–32.

171E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Shen, Y., Schidlowski, M., 2000. New C isotope stratigraphy from southwest China: im-plications for the placement of the Precambrian–Cambrian boundary on theYangtze Platform and global correlations. Geology 28, 623–626.
Shen, B., Xiao, S., Dong, L., Zhou, C., Liu, J., 2007. Problematic macrofossils fromEdiacaran successions in the North China and Chaidan blocks: implications fortheir evolutionary roots and biostratigraphic significance. Journal of Paleontolo-gy 81, 1396–1411.
Singh, P., Shukla, D.S., 1981. Fossils from the Lower Tal: their age and its bearing on thestratigraphy of Lesser Himalaya. Geosciences Journal 11, 157–176.
Smith, A.G., 2001. Paleomagnetically and tectonically based global maps for Vendian toMid-Ordovician time. In: Zhuravlev, A.Yu, Riding, R. (Eds.), The Ecology of theCambrian Radiation. Columbia University Press, New York, pp. 11–46.
Smith, A.G., Hurley, A.M., Briden, J.C., 1981. Phanerozoic paleocontinental world maps.Cambridge Earth Science Series (83 pp.).
Sokolov, B.S., 1972. The Vendian Period in Earth history. Editor not listedMezhdunarodnyi geologicheskiy congress, XXIV Sessiya. : Dokladov sovetskykhgeologov. Problema, 7. Nauka, Moscow (79–86. (in Russian)).
Sokolov, B.S., Zhuravleva, A.B. (Eds.), 1983. Yarusnoe raschelenie nizhnego kembriyaSibiri. Atlas okamenelostey. Nauka, Moscow (284 pp. (in Russian)).
Staplin, F.L., Jansonius, J., Pocock, S.A.J., 1965. Evaluation of some acritarchous hystrichospheregenera. Neues Jahrbuch der Geologie und Paläontologie, Abhandlungen 123, 167–201.
Steiner, M., 1994. Die neoproterozoischen Megaalgen Südchinas. BerlinerGeowissensch. Abhandlungen E, 15 (146 pp. (in German)).
Steiner, M., Zhu, M.Y., Weber, B., Geyer, G., 2001. Early Cambrian of eastern Yunnan:trilobite-based biostratigraphy and related faunas. Acta Palaeontologica Sinica 40,63–69 (Supplement).
Steiner, M., Li, G.X., Qian, Y., Zhu, M.Y., 2004. Lower Cambrian small shelly fossils ofnorthern Sichuan and southern Shaanxi (China), and their biostratigraphic signif-icance. Geobios 37, 259–275.
Steiner, M., Li, G.X., Qian, Y., Zhu, M.Y., Erdtmann, B.-D., 2007. Neoproterozoic to EarlyCambrian small shelly fossil assemblages and a revised biostratigraphic correlationof the Yangtze Platform (China). Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 254, 67–99.
Steiner, M., Li, G., Ergaliev, G., 2011. Toward a subdivision of the traditional ‘LowerCambrian. Museum of Northern Arizona Bulletin 67, 306–308.
Strauss, H., DeMarais, D.J., Hayes, J.M., Summons, R.E., 1992. The carbon isotope record.In: Schopf, J.W., Klein, C. (Eds.), The Proterozoic Biosphere. Cambridge UniversityPress, Cambridge, pp. 117–127.
Sukhov, S.S., 1997. Cambrian depositional history of the Siberian craton: evolution ofthe carbonate platforms and basins. Yanxiang Guidi 17, 27–39.
Sundberg, F.A., McCollum, L.B., 2000. Ptychopariid trilobites of the Lower–Middle Cam-brian boundary interval, Pioche Shale, southeastern Nevada. Journal of Paleontolo-gy 74, 604–630.
Sundberg, F.A., McCollum, L.B., McCollum, M.B., 2011. Stop 7C. Stratigraphy of the Lau-rentian uppermost Dyeran Stage to Skullrockian Stage, Emigrant Formation, atSplit Mountain, Esmeralda County, Nevada. Museum of Northern Arizona Bulletin67, 246–252.
Suvorova, N.P., 1960. Nadsemeystvo Redlichioidea. In: Chernysheva, N.E. (Ed.), Osnovypaleontologii. Chlenistonogie, trilobitoobraznye i rakoobraznye: Gosudarstvennoenauchno-tekhnicheskoe izdat'elstvo, Moscow, pp. 64–72 (in Russian).
Sysoiev, V.A., 1968. Stratigrafiya i khioliti drevneyskikh sloev nizhnego kembriyaSibiriskoy platformy. Akademiya Nauk SSSR, Yakutskiy filial' Sibirskogo otdeleniyainstitut geologii, Yakutsk (67 pp.).
Tanoli, S.K., Pickerill, R.K., 1988. Lithostratigraphy of the Cambrian–Lower Ordovician SaintJohn Group, southern New Brunswick. Canadian Journal of Earth Sciences 25, 669–690.
Tate, R., 1892. The Cambrian fossils of South Australia. Transactions of the Royal Societyof South Australia 15, 165–185.
Terfelt, F., Ahlberg, P., 2010. Pseudagnostus rugosus Ergaliev, 1980: a key agnostoid spe-cies for intercontinental correlation of upper Furongian (Cambrian) strata. Geolog-ical Magazine 147, 789–796.
Terfelt, F., Bagnoli, G.E., Stouge, S., 2011a. Re-evaluation of the conodont Iapetognathusand implications for the base of the Ordovician System. Lethaia 45, 227–237.
Terfelt, F., Bagnoli, G.E., Stouge, S., 2011b. The Cambrian–Ordovician boundary global bound-ary: an anachronistic point in time. Museum of Northern Arizona Bulletin 67 (308), 309.
Theokritoff, G., 1979. Early Cambrian biogeography in the North Atlantic region.Lethaia 18, 283–293.
Toll, E., von, 1899. Beiträge zur Kenntniss des sibirischen Kambrium. I. [Materialy popoznaniyu Sibirskogo kembriya]. Mémores du Académie Impériale du Sciences,St.-Pétersbourg, Series, VIII (10) (57 pp. (in German and Russian)).
Torsvik, T.H., Smethurst, M.A., Meert, J.G., Van der Voo, R., 1996. Continental break-upand collision in the Neoproterozoic and Palaeozoic: a tale of Baltica and Laurentia.Earth-Science Reviews 40, 229–258.
Van Couvering, J.A., Berggren, W.A., 1977. Biostratigraphic basis of the Neogene timescale. In: Kaufman, E.G., Hazel, J.E. (Eds.), Concepts andMethods of Biostratigraphy.Dowden, Hutchinson & Ross, Stroudsburg, Pennsylvania, pp. 283–306.
Vanguestaine, M., Léonard, R., 2005. New biostratigraphic and chronostratigraphic datafrom the Sautou Formation and adjacent strata (Cambrian, Givonne inlier, RevinGroup, northern France), and some lithostratigraphic and tectonic implications.Geologica Belgica 8, 131–144.
Varlamov, A.I., Rozanov, A.Yu, Khomentovsky, V.V., Shabanov, Yu.Ya, Abaimova, G.P.,Demidenko, Yu.E, Karlova, G.A., Korovnikov, I.V., Luchinina, V.A., Malakhovskaya,Ya.E., Parkhaev, P.Yu, Pegel, T.V., Skorlotova, N.A., Sundukov, V.M., Sukhov, S.S.,Fedorov, A.B., Kipriyanova, L.D., 2008. The Cambrian system of the Siberian Plat-form. Part 1: The Aldan–Lena Region. Field Conference of the Cambrian Stage Sub-division Working Group, Yukatia, Russia, July 20–August 1, 2008. PIN RAS,Moscow, Novosibirsk (300 pp.).
Vidal, G., 1981. Lower Cambrian acritarch stratigraphy in Scandinavia. GeologiskaFöreningens i Stockholm Förhandlingar 103, 183–192.
Vidal, G., Moczydłowska, M., 1992. Patterns of phytoplankton radiation across thePrecambrian–Cambrian boundary. Journal of the Geological Society of London 149,647–654.
Vidal, G., Moczydłowska, M., 1995. The Neoproterozoic of Baltica. Stratigraphy,palaeobiology and general geological evolution. Precambrian Research 73, 197–216.
Vidal, G., Jensen, S., Palacios, T., 1994. Neoproterozoic (Vendian) ichnofossils fromLower Alcudian strata in central Spain. Geological Magazine 131, 169–179.
Vidal, G., Moczydłowska, M., Rudavskaya, V.R., 1995. Constraints on the Early Cambrianradiation and correlation of the Tommotian and Nemakit-Daldynian stages of east-ern Siberia. Journal of the Geological Society of London 152, 499–510.
Volkova, N.A., 1968. Akritarkhi pogranichnykh i nizhnego kembriya Estonskoy. In:Volkova, N.A., Zhuravleva, Z.A., Zabrodin, V.E., Klinger, B.Sh. (Eds.), Problematikipogranichnykh sloev rifeya i kembriya Russkoy platformi, Urala i Kazakhstana.Nauka, Moscow, pp. 8–36 (in Russian).
Volkova, N.A., Kiryanov, V.V., Piscun, L.V., Paskeviciene, L.T., Jankauskas, T.V., 1979.Mikroflora. In: Keller, B.M., Rozanov, A.Yu. (Eds.), Paleontologiyaverkhnedokembriyskikh i kembriyskikh otlozheniy Vostochno-Evropeyskoyplatformi. Nauka, Moscow, pp. 9–38 (in Russian).
Voronova, L.G., Missarzhevsky, V.V., 1969. Nakhodki vodorosley i trubok chervey vpogranichnykh sloyakh kembriya i dokembriya na severe Sibirskoy platformy.Doklady Akademiya Nauk SSSR 184, 207–210 (in Russian).
Vostokova, V.A., 1962. Kembriyskie gastropody Sibirskoy platformy i Taymyra. Sbornikstatey po paleontologii i biostratigrafii, 28, pp. 51–74 (in Russian).
Walcott, C.D., 1889. Descriptive notes on new genera and species from the Lower Cam-brian or Olenellus Zone of North America. Proceedings of the U.S. National Muse-um: , 12, pp. 33–46.
Walcott, C.D., 1890. The fauna of the Lower Cambrian or Olenellus Zone. 10th Report ofthe U.S. Geological Survey 515–774.
Walcott, C.D., 1910. Olenellus and other genera of the Mesonacidae. Cambrian Geologyand Paleontology No. 6: Smithsonian Miscellaneous Collections, 53, pp. 231–422.
Weber, B., Steiner, M., Zhu, M.Y., 2007. Precambrian–Cambrian trace fossils from theYangtze Platform (South China) and the early evolution of bilaterian lifestyles.Palaeogeography, Palaeoclimatology, Palaeoecology 254, 328–349.
Webster, M., Gaines, R.R., Hughes, N.C., 2008. Microstratigraphy, trilobitebiostratinomy, and depositional environment of the “Lower Cambrian” RuinWash Lagerstätte, Pioche Formation, Nevada. Palaeogeography, Palaeoclimatology,Palaeoecology 264, 100–122.
Westrop, S.R., Landing, E., 2011. Lower Cambrian (Branchian) eodiscoid trilobites fromthe lower Brigus Formation, Avalon Peninsula, Newfoundland, Canada. Memoirs ofthe Association of Australasian Palaeontologists 42, 209–262.
Westrop, S.R., Adrain, J.M., Landing, E., 2011. The Cambrian (Sunwaptan, Furongian)agnostoid arthropod Lotagnostus Whitehouse, 1936, in Laurentian and AvalonianNorth America: systematics and biostratigraphic significance. Bulletin ofGeosciences, Czech Geological Survey 86, 569–594.
Wilson, J.P., Grotzinger, J.P., Fischer, W.W., Hand, K.P., Jensen, S., Knoll, A.H., Abelson, J.,Metz, J.M., McLoughlin, N., Cohen, P.A., Tice, M.M., 2012. Deep-water incised valleydeposits at the Ediacaran–Cambrian boundary in southwestern Namibia containabundant Treptichnus pedum. Palaios 27, 252–273.
Woodward, H., 1884. Note on the remains of trilobites from South Australia. GeologicalMagazine, New Series, Decade III, 1, pp. 342–344.
Xiang, L.W., Zhou, T.M., 1987. Class Trilobita. In: Wang, X.F. (Ed.), Biostratigraphy ofthe Yangtze Gorges Area, 2, Early Palaeozoic. Geological Publishing House, Beijing,pp. 294–335 (in Chinese, with English abstract).
Xing, Y.S., Luo, H.L., 1984. Precambrian–Cambrian boundary candidate, Meishucun,Jinning, Yunnan, China. Geological Magazine 121, 143–154.
Xing, Y.S., Ding, Q.X., Luo, H.L., He, T.G., Wang, Y.G., 1983. The Sinian–Cambrian boundary ofChina and its related problems. Bulletin of the Institute of Geology, Chinese Academyof Geological Sciences, 10 (262 pp. (in Chinese, with English summary)).
Yegorova, L.I., Savitsky, V.E., 1969. Stratigrafiya i biofatsii kembriya Sibirskoy platformy(Zapadnoe Prianabar'e). Trudy Sibirskoy nauchno-issledovatel'skiy institutgeologii, geofizikii i mineral'nogo syr'ya (SNIIGGiMS), 43 (407 pp.).
Yin, L.M., Geer, T., Bian, L., Xie, X., 2011. Diversified microfossils found from the Early–Middle Cambrian strata in eastern Guizhou Province, China. Museum of NorthernArizona Bulletin 67, 314.
Yuan, J.L., Zhao, Y.L., 1999. Subdivision and correlation of Lower Cambrian in southwestChina, with a discussion of the age of Early Cambrian series biota. ActaPalaeontologica Sinica 38, 116–131 (Supplement, in Chinese).
Yuan, J.L., Zhao, Y.L., Li, Y., 2001. Biostratigraphy of oryctocephalid trilobites. ActaPalaeontologica Sinica 40, 143–156 (Supplement, in Chinese).
Yuan, J.J., Zhu, X.J., Lin, J.P., Zhu, M.Y., 2011. Tentative correlation of Cambrian Series 2between South China and other continents. Bulletin of Geosciences 86, 397–404.
Zang, W.L., 1992. Sinian and Early Cambrian floras and biostratigraphy on the SouthChina Platform. Palaeontographica Abteilung B 224, 75–119.
Zang, W., Moczydłowska, M., Jago, J.B., 2007. Lower Cambrian acritarch assemblagezones in South Australia and global correlation. Memoirs of the Association of Aus-tralasian Palaeontologists 33, 141–177.
Zhang, W.T. [“Chang”], 1966. On the classification of Redlichiacea, with description ofnew families and new genera. Acta Palaeontologica Sinica 14, 135–184 (in Chineseand English).
Zhang, W.T. [“Chang”], 1980. A review of the Cambrian of China. Journal of the Geolog-ical Society of Australia 27, 137–150.
Zhang, W.T. [“Chang”], 1987. World's oldest Cambrian trilobites from eastern Yun-nan. In: Nanjing Institute of Geology and Palaeontology, Academia Sinica(Ed.), Stratigraphy and Palaeontology of Systemic Boundaries in China. :

172 E. Landing et al. / Earth-Science Reviews 123 (2013) 133–172
Precambrian–Cambrian Boundary, 1. Anhui Science and Technology PublishingHouse, Hefei, pp. 1–16.
Zhang, [“Chang”], W.T., Yuan, K.X., Zhou, Zh.Y., Qian, Y., Wang, Z.Zh, 1979. The Cambri-an of southwestern China. . Editor not named Carbonate Biostratigraphy of South-western China. Science Press, Beijing (in Chinese).
Zhang, W.T., Babcock, L.E., Xiang, L.W., Sun, W.G., Luo, H.L., Jiang, Z.W., 2001. LowerCambrian stratigraphy of Chengjiang, eastern Yunnan, China, with special noteson Chinese Parabadiella, Moroccan Abadiella and Australian Abadiella huoi. ActaPalaeontologica Sinica 40, 294–309.
Zhu, M.Y., Li, G.X., Zhang, J.M., Steiner, M., Qian, Y., Jiang, Z.W., 2001a. Early Cambrianstratigraphy of east Yunnan, southwestern China: a synthesis. Acta PalaeontologicaSinica 40, 4–39.
Zhu, M.Y., Zhang, J.M., Steiner, M., Li, G.X., 2001b. Precambrian–Cambrian boundarystratigraphy at Meishucun, Yunnan Province, China: base of the Cambrian andthe first apearence of trilobite. In: Peng, S.C., Babcock, L.E., Zhu, M.Y. (Eds.), Cambri-an System of China: Palaeoworld, 13, pp. 226–235.
Zhu, M.Y., Zhang, J.M., Li, G.X., Yang, A.H., 2004. Evolution of carbon isotopes in theCambrian of China: implications for Cambrian subdivisions and trilobite mass ex-tinctions. Geobios 37, 287–310.
Zhu, M.Y., Li, G.X., Hu, S.X., Zhao, F.C., 2005. Ediacaran–Cambrian boundary stratigraphyat Meishucun, Jinning County, and fossil quarries of the Early Cambrian Chengjiangbiota near Haikou, Kunming City, Yunnan, China. In: Peng, S.C., Babcock, L.E., Zhu,M.Y. (Eds.), Cambrian System of China and Korea. Guide to Field Excursions. Uni-versity of Science and Technology, China Press, Hefei, pp. 13–29.
Zhu, M.Y., Babcock, L.E., Peng, S.C., 2006. Advances in Cambrian stratigraphy andpaleontology: integrating correlation techniques, paleobiology, taponomy andpaleoenvironmental reconstruction. Palaeoworld 15, 217–222.
Zhu, M.Y., Yang, A.H., Li, G.X., Yuan, J.J., 2008. A working model for subdivision of thelower half Cambrian. In: Voronin, T.A. (Ed.), The Siberian Platform, westernYukutia. 13th International Field Conference of the Cambrian Stage SubdivisionWorking Group. SNIGGiMS, Novosibirsk, p. 204 (205).
Zhuravlev, A.Yu., Liñan, E., Gámez Vintaned, J.A., Debrenne, F., Fedorov, A.B., 2012. Newfinds of skeletal fossils in the terminal Neoproterozoic of the Siberian Platform andSpain. Acta Palaeontologica Polonica 57, 205–224.
Zhuravleva, I.T., Korshunov, V.I., Rozanov, A.Yu, 1969. Atdabanskiy yarus i egoobosnovanie po arkheotsiatam v stratotipicheskom razreze. Materialypaleontologii i biostratigrafii nizhnego kembriya Sibiri i Dal'neogo Vostoka.Nauka, Moscow, pp. 3–59 (in Russian).
Related Documents











