Cell Tissue Res (1992) 269:289-297 Cell&Tissue Research Springer-Verlag 1992 Calbindin D-28k-positive neurons in the rat olfactory bulb An immunohistochemical study J.G. Brifi6n, J.R. Alonso, R. Ar6valo, E. Garcia-Ojeda, J. Lara, and J. Aij6n Departamento de Biologia Celular, Universidad de Salamanca, E-37007 Salamanca, Spain Received November 7, 1991 / Accepted April 3, 1992 Summary. We have studied the distribution of calbindin D-28k immunoreactivity in the rat olfactory bulb using specific monoclonal antibodies and the avidin-biotin-im- munoperoxidase method. The largest number of positive neurons was located in the periglomerular layer. These neurons were identified as periglomerular cells; they have been described also by other authors as calbindin- positive elements. Close to these neurons, a second pop- ulation of nerve cells was identified as superficial short- axon neurons. The remaining layers showed a smaller number of stained elements. Other labeled neurons were located along the external border of the external plexi- form layer; the scarce neurons marking its internal border were identified as van Gehuchten cells. No immu- noreactive structures were found in the mitral cell layer, although we observed another population of immuno- stained short-axon cells at its internal border. Some reac- tive structures, identified by us as horizontal and vertical cells of Cajal, were located in the boundary zone between the internal plexiform layer and the granule layer. In the white matter, we found a neuronal type characterized by its large size and oriented arborization of varicose dendrites. Key words: Calbindin D-28k Olfactory bulb Calci- um-binding proteins - Immunohistochemistry - Rat (Wistar) The "EF-hand" calcium-binding proteins are calcium- sequestering molecules that regulate calcium concentra- tions in cells over time scales that parallel electric events (Yamamoto et al. 1989). Except for troponin-C, all sub- families and individual proteins of the EF-hand group represented in vertebrates can be found in neuronal or glial populations (Persechini et al. 1989). Calbindin D-28k (CaBP), which belongs to the EF- hand homolog group, is a water-soluble calcium-binding Correspondence to: J.R. Alonso protein of low molecular weight found in different verte- brate tissues and organs, including the enteric and the central nervous systems (Taylor 1974; Baimbridge and Miller 1982; Celio 1990). CaBP may contribute to the regulation of calcium through its capacity to buffer calci- um, and thereby promote or restrict the actions of this ion. It may also facilitate the diffusion of calcium, thus modulating calcium gradients in neurons (Kretsinger et al. 1982). The distribution of CaBP has been described in the rat brain using radioimmunoassay, immunocytochem- istry and mRNA localization (see Baimbridge and Miller 1982; Baimbridge et al. 1982; Feldman and Christakos 1983; Garcia-Segura et al. 1984; Halfisz et al. 1985; S& quier et al. 1988, 1990; Celio 1989, 1990). CaBP is found in specific neuronal populations in various areas of the brain, where it is throught to play a role in these neurons in the regulation of calcium-dependent events (Celio 1989). CaBP has been demonstrated in different types of interneurons (Baimbridge and Miller 1982; Garcia-Se- gura et al. 1984; Celio 1990), mainly located in the peri- glomerular layer of the olfactory bulb. However, these reports are general, referring to the whole brain, and the classification of the bulbar CaBP-positive neurons remains unclear. Furthermore, previously used polyclon- al antibodies against CaBP also recognize another calci- um-binding protein, calretinin, because of its similar pri- mary sequence (Rogers 1987). In this study, we have used a highly specific monoclonal antibody against cal- bindin D-28k and the avidin-biotin immunoperoxidase method in order to obtain a better characterization of CaBP-positive neurons in the olfactory bulb of the rat. Materials and methods Six adult female Wistar rats weighing 200-250 g were used. The animals were perfused through the ascending aorta under deep ketamine anesthesia (50 ml/kg body weight). The vascular system was ringed with 60 ml Ringer solution followed by 400 ml of a fixative mixture containing 4% paraformaldehyde, 0.08% glutaral-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cell Tissue Res (1992) 269:289-297 Cell&Tissue
Research �9 Springer-Verlag 1992
Calbindin D-28k-positive neurons in the rat olfactory bulb
An immunohistochemical study
J.G. Brifi6n, J.R. Alonso, R. Ar6valo, E. Garcia-Ojeda, J. Lara, and J. Aij6n
Departamento de Biologia Celular, Universidad de Salamanca, E-37007 Salamanca, Spain
Received November 7, 1991 / Accepted April 3, 1992
Summary. We have studied the distribution of calbindin D-28k immunoreactivity in the rat olfactory bulb using specific monoclonal antibodies and the avidin-biotin-im- munoperoxidase method. The largest number of positive neurons was located in the periglomerular layer. These neurons were identified as periglomerular cells; they have been described also by other authors as calbindin- positive elements. Close to these neurons, a second pop- ulation of nerve cells was identified as superficial short- axon neurons. The remaining layers showed a smaller number of stained elements. Other labeled neurons were located along the external border of the external plexi- form layer; the scarce neurons marking its internal border were identified as van Gehuchten cells. No immu- noreactive structures were found in the mitral cell layer, although we observed another population of immuno- stained short-axon cells at its internal border. Some reac- tive structures, identified by us as horizontal and vertical cells of Cajal, were located in the boundary zone between the internal plexiform layer and the granule layer. In the white matter, we found a neuronal type characterized by its large size and oriented arborization of varicose dendrites.
Key words: Calbindin D-28k Olfactory bulb Calci- um-binding proteins - Immunohistochemistry - Rat (Wistar)
The " E F - h a n d " calcium-binding proteins are calcium- sequestering molecules that regulate calcium concentra- tions in cells over time scales that parallel electric events (Yamamoto et al. 1989). Except for troponin-C, all sub- families and individual proteins of the EF-hand group represented in vertebrates can be found in neuronal or glial populations (Persechini et al. 1989).
Calbindin D-28k (CaBP), which belongs to the EF- hand homolog group, is a water-soluble calcium-binding
Correspondence to: J.R. Alonso
protein of low molecular weight found in different verte- brate tissues and organs, including the enteric and the central nervous systems (Taylor 1974; Baimbridge and Miller 1982; Celio 1990). CaBP may contribute to the regulation of calcium through its capacity to buffer calci- um, and thereby promote or restrict the actions of this ion. It may also facilitate the diffusion of calcium, thus modulating calcium gradients in neurons (Kretsinger et al. 1982).
The distribution of CaBP has been described in the rat brain using radioimmunoassay, immunocytochem- istry and mRNA localization (see Baimbridge and Miller 1982; Baimbridge et al. 1982; Feldman and Christakos 1983; Garcia-Segura et al. 1984; Halfisz et al. 1985; S& quier et al. 1988, 1990; Celio 1989, 1990). CaBP is found in specific neuronal populations in various areas of the brain, where it is throught to play a role in these neurons in the regulation of calcium-dependent events (Celio 1989).
CaBP has been demonstrated in different types of interneurons (Baimbridge and Miller 1982; Garcia-Se- gura et al. 1984; Celio 1990), mainly located in the peri- glomerular layer of the olfactory bulb. However, these reports are general, referring to the whole brain, and the classification of the bulbar CaBP-positive neurons remains unclear. Furthermore, previously used polyclon- al antibodies against CaBP also recognize another calci- um-binding protein, calretinin, because of its similar pri- mary sequence (Rogers 1987). In this study, we have used a highly specific monoclonal antibody against cal- bindin D-28k and the avidin-biotin immunoperoxidase method in order to obtain a better characterization of CaBP-positive neurons in the olfactory bulb of the rat.
Materials and methods
Six adult female Wistar rats weighing 200-250 g were used. The animals were perfused through the ascending aorta under deep ketamine anesthesia (50 ml/kg body weight). The vascular system was ringed with 60 ml Ringer solution followed by 400 ml of a fixative mixture containing 4% paraformaldehyde, 0.08 % glutaral-

290
dehyde and 15% saturated picric acid (Somogyi and Takagi 1982) in 0.1 M phosphate buffer, pH 7.2 (PB).
After perfusion, the brains were removed, the olfactory bulbs dissected out and stored in the same fixative without glutaralde- hyde for 2 h. The bulbs were then transferred to 30% sucrose (v/v) in PB at 4 ~ C overnight. These blocks were washed in several changes of PB and cut at a thickness of 30 pm perpendicularly to the longitudinal axis of the brain using a cryostat. The sections were washed in PB and stored at 4 ~ C overnight.
The immunohistochernical procedure was carried out on free floating sections and involved the use of the avidin-biotin-peroxi- dase complex method (Hsu et al. 1981). The sections were preincu- bated in 0.01% Triton X-100 in PB for 2 h at room temperature. Thereafter, the sections were incubated with a monoclonal anti- body against calbindin D-28k diluted at 1:1000 in PB for 48 h at 4 ~ C. This antibody has been fully characterized (Cello et al. 1990) and widely used in different studies of the central nervous system (see Cello 1990). The sections were washed in PB and incu- bated with biotinylated anti-mouse immuno-gammaglobulin (Vec- tastain ABC kit, Vector Laboratories, Burlingame, USA) diluted 1:250, for 3 h at 20 ~ C, and then in Vectastain ABC reagent diluted 1:250, for 2 h. Tissue-bound peroxidase was visualized using 0.07% diaminobenzidine and 0.003% hydrogen peroxide in 0.1 M TRIS buffer (pH 7.6) for 5-10 rain, under visual control.
The specificity of the CaBP-staining was controlled by omitting the specific antibody in the first incubation step. Additionally, in- terference by endogenous peroxidases was ruled out by staining some sections with chromogen and hydrogen peroxide. No residual immunoreaction was found in either controls. Finally, the sections were dehydrated in an increasing ethanol series, mounted in Entel- lan and studied under the light microscope.
Results
Light-microscopic examination of CaBP-stained sec- tions of the rat olfactory bulb revealed positive somata, dendritic arborizations, axons and axonic collaterals with a laminated distribution (Figs. 1, 2 a). No immuno- reactivity was found in the controls.
Olfactory nerve layer (ONL)
No immunostaining was visible in this layer. The axons arising f rom the olfactory receptors, the olfactory fibers, were CaBP-immunonegat ive (Figs. 1, 2 a).
Glomerular layer (GL)
This layer showed the highest number of CaBP-positive neurons. We observed neuronal bodies and fibers with strong staining in the glomerular region and, more rare- ly, in the periglomerular region (Figs. 1, 2c-e).
Neuronal bodies appeared to be strongly labeled, al- though variations in the intensity of this staining were found in neurons with similar morphologies. Most of the immunosta ined cells in this layer were small spherical or ovoid neurons with diameters ranging between 7 and 9 gm.
The dendritic arborizations were extensively stained. They were formed by one main dendritic t runk that nor- mally branched close to the cellular body into two or, occasionally, three secondary dendrites. These secondary
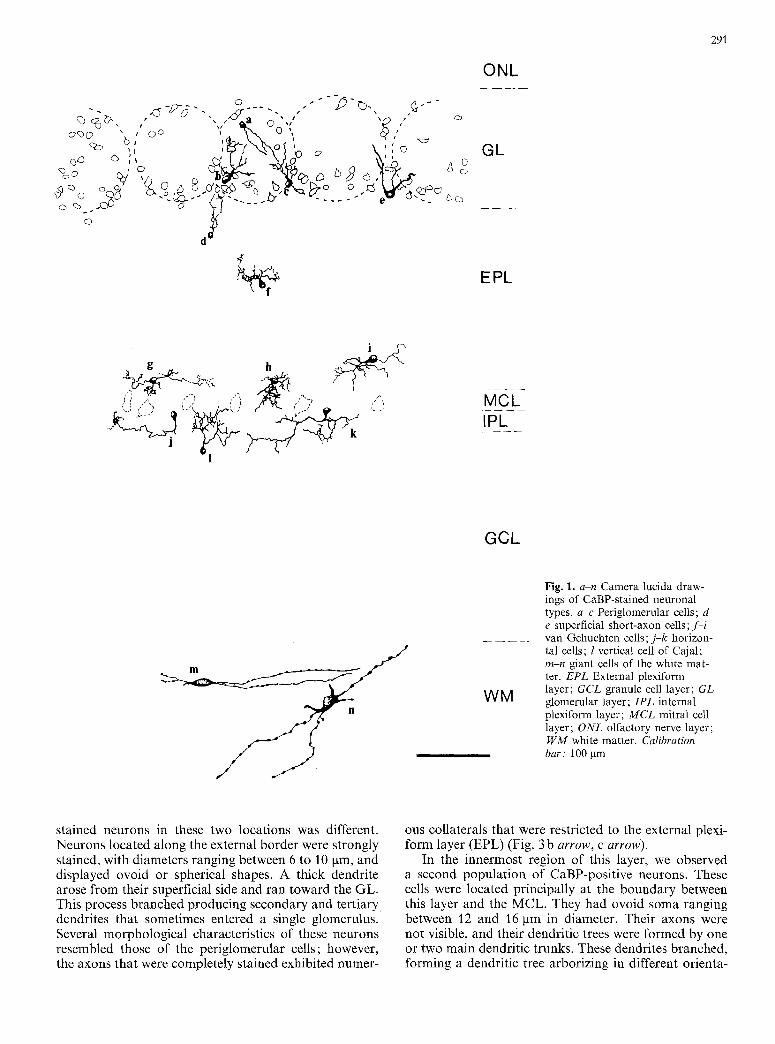
dendrites branched several times forming a small, dense dendritic tree that did not exhibit a specific orientation pat tern with regard to the olfactory bulb lamination (Figs. 1 a-c, 2c). A group of these arborizations clearly constituted a glomerulus, with a variable number of cell bodies located in its periphery (Figs. 1, 2b). Dendritic arborizations of immunoreact ive cells in the same glo- merulus intermingled with each other producing a dense network; however, it was not possible to follow the ex- tensions originating f rom a given cell. Normally, the complete dendritic tree of one cell was included within a single glomerulus. More rarely, the dendrites of a CaBP-positive neuron branched into two or more glo- meruli. In many cases, it was possible to follow the axon- al extensions of the stained neurons. The initial axonal segment generally arose f rom a point on the cell body opposite the main dendrite. More rarely, we were able to follow an axon along most of its length. These axons appeared to extend into the periglomerular layer but it was not possible to identify their targets (Fig. 2c). We classified these neurons as periglomerular cells by their size, morphology and location; most, but not all, were stained after CaBP-immunocytochemistry. A study of sections counterstained with toluidine blue revealed small CaBP-immunonegat ive somata around the glo- meruli.
Within this same layer, we observed immunostained neuronal bodies with a morphology different f rom that previously described (Figs. 1 d, e, 2d, e, 3 b, c). These neurons were less abundant than the periglomerular cells, al though with similar strong immunostaining. The CaBP-stained superficial short-axon cells seemed to rep- resent a heterogenous population, varying in size, mor- phology and location (Fig. I d, e). Generally, the neuro- nal somata were ovoid and slightly larger (10-15 gm in diameter) than those of periglomerular neurons. The main difference was in their dendritic arborization; these dendrites remained in the periglomerular zone and did not project into a glomerulus. The pr imary processes (normally two) of these cells arose f rom opposite poles of the fusiform cell bodies (Figs. 1 e, 2d, e open arrow) and extended into the periglomerular region. Sometimes, the dendrites of these neurons projected into the periglo- merular region of more than two glomeruli (Fig. I e). These pr imary dendrites branched, giving rise to second- ary and tertiary dendrites, but did not form a dendritic field as dense as those arising f rom the periglomerular cells. In this cell type, the axons were stained for shorter distances than in the periglomerular cells, and we did not observe any beading along their course. Because of these morphological characteristics and their location, we identified these cells as superficial short-axon cells. Other types of short-axon cells were also CaBP-immuno- stained, as described in the following layers.
External plexiform layer (EPL)
This layer showed few stained neurons in the zones close to both the adjacent strata, the G L and the mitral cell layer (MCL) (Figs. 1, 3a-d) . The morphology of the
-4 ;",,,"" ,"
oo o I, O'I h \ ) o ~ ~ " . .
o ~�9 " ' ~ " d - -~ ' " . . . . D" . . . . . " e " ' -
cb
bC~
ONL
GL
EPL
291
MCL IPL
GCL
WM
Fig. 1. a-n Camera lucida draw- ings of CaBP-stained neuronal types, a-c Periglomerular cells; & e superficial short-axon cells; f - i van Gehuchten cells; j-k horizon- tal cells; l vertical cell of Cajal; m-n giant cells of the white mat- ter. EPL External plexiform layer; GCL granule cell layer; GL glomerular layer; IPL internal plexiform layer; M C L mitral cell layer; ONL olfactory nerve layer; W M white matter. Calibration bar: 100 Ixm
stained neurons in these two locations was different. Neurons located along the external border were strongly stained, with diameters ranging between 6 to 10 pm, and displayed ovoid or spherical shapes. A thick dendrite arose from their superficial side and ran toward the GL. This process branched producing secondary and tert iary dendrites that sometimes entered a single glomerulus. Several morphological characteristics of these neurons resembled those of the periglomerular cells; however, the axons that were completely stained exhibited numer-
ous collaterals that were restricted to the external plexi- form layer (EPL) (Fig. 3 b arrow, c arrow).
In the innermost region of this layer, we observed a second population of CaBP-positive neurons. These cells were located principally at the boundary between this layer and the MCL. They had ovoid soma ranging between 12 and 16 pm in diameter. Their axons were not visible, and their dendritic trees were formed by one or two main dendritic trunks. These dendrites branched, forming a dendritic tree arborizing in different orienta-
292

tions. We identified them as another type of short-axon neurons, viz., van Gehuchten cells (Figs. 1 f-i, 3 a arrows, d).
Mitral cell layer (MCL)
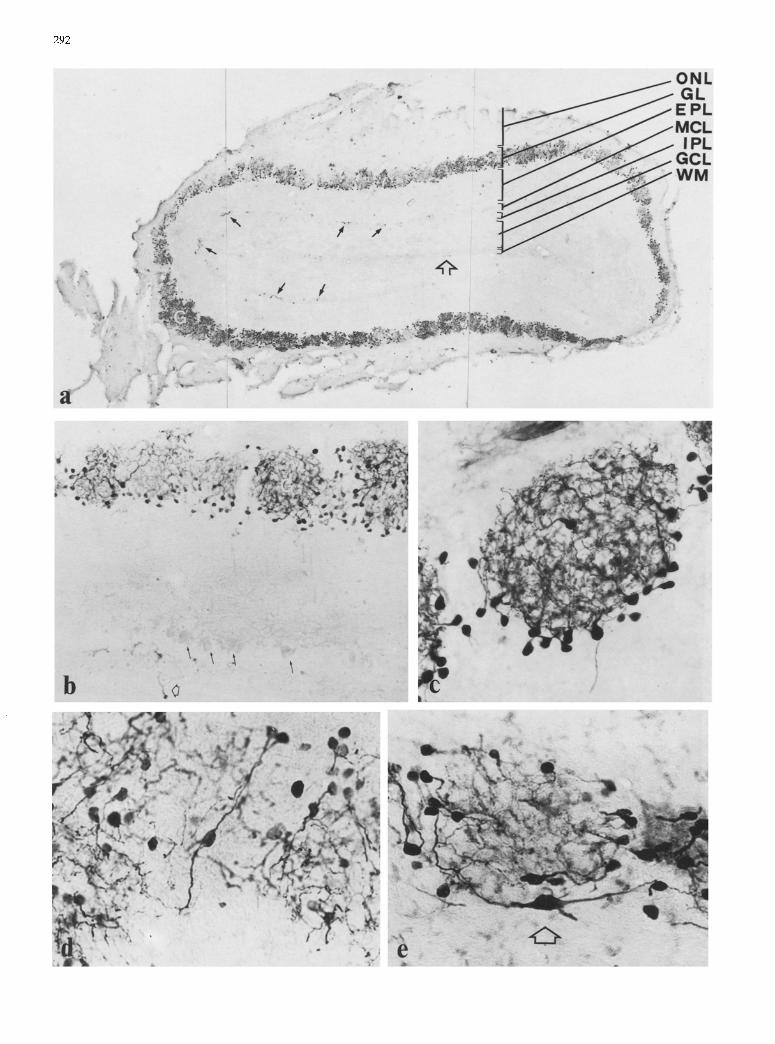
No neuronal body was stained in this layer. After careful examination, it was possible to observe the immunoneg- ative profiles of mitral cells, allowing an exact delimita- tion of the EPL and internal plexiform layer (IPL) (Figs. 1, 2b arrows). Immunopositive short-axon cells were found close to the mitral neurons in both plexiform layers (Figs. 1, 2a, b).
Internal plexiform layer and granule cell layer (IPL and GCL)
A CaBP-immunostained population was located at the boundary between these layers, and was formed by me- dium-sized (14-16 gin) neurons with ovoid or piriform cell bodies and highly ramified dendritic processes (Figs. l j-l, 3e, f) that projected into both layers. Two different populations could be differentiated. One had predominantly ovoid or irregular somata and three or more dendritic trunks oriented parallel to the MCL. We identified these neurons as horizontal cells (Figs. l j, k, 3f). The second population of stained neurons in this region was similar in size to the aforementioned class, but their dendrites extended from two opposite regions at either end of the long axis of the fusiform soma, and were oriented in such a way that most of the branches ramified predominantly toward the olfactory bulb lamination (Figs. 11, 3 e). These stained neurons ap- peared to be homologous to the vertical cells of Cajal. The stained segment of the axon sometimes showed col- laterals forming a small arborization near to the cell body (Fig. 3 e, arrow).
Granule cells, the most abundant neurons in the ol- factory bulb, were always CaBP-negative.
White matter (WM)
Close to the ventricle, we found a rarely described CaBP- stained population (Figs. 1 m, n, 4a, b). The cell bodies
Fig. 2. a Photographic montage of a coronal section through a CaBP-stained olfactory bulb showing strong staining in the GL and scattered stained neurons in the IPL-MCL (arrows) and WM (open arrow), x 35. b CaBP-staining of the external layers. Note the positive glomeruli (G) whereas EPL and MCL are practically immunonegative. Negative profiles of the mitral cells (arrows) and a positive neuron in the IPL (open arrow) can be distinguished. x 115. e Immunopositive periglomerular cells forming a glomeru-
lus. Note the complete staining of their dendritic trunks, x 400. d, e Superficial short-axon cells surrounding the periglomerular cells�9 Note the larger size of these neurons (open arrow) with respect to the periglomerular cells, and the characteristic shapes of their somata and dendritic trunks, x 570
G
O
t ~
r m O ct~
"C "-~
o
o
;>
o o o o o o ~ o ~ o o ~
o
o
O
O o
293
o
" a
o~
o ~
...a
,--A
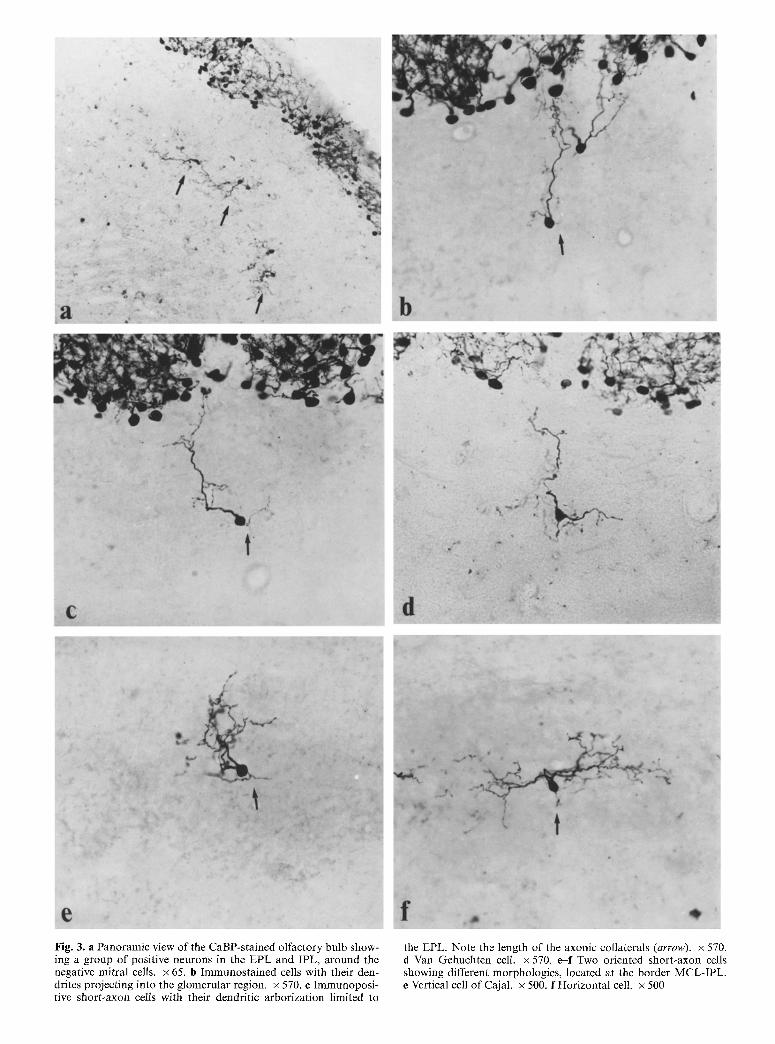
Fig. 3. a Panoramic view of the CaBP-stained olfactory bulb show- ing a group of positive neurons in the EPL and IPL, around the negative mitral cells, x 65. b Immunostained cells with their den- drites projecting into the glomerular region. • 570. r Immunoposi- tive short-axon cells with their dendritic arborization limited to
the EPL. Note the length of the axonic collaterals (arrow). • 570. d Van Gehuchten cell. x 570. e--f Two oriented short-axon cells showing different morphologies, located at the border MCL-IPL. e Vertical cell of Cajal. x 500. f Horizontal cell. x 500
295
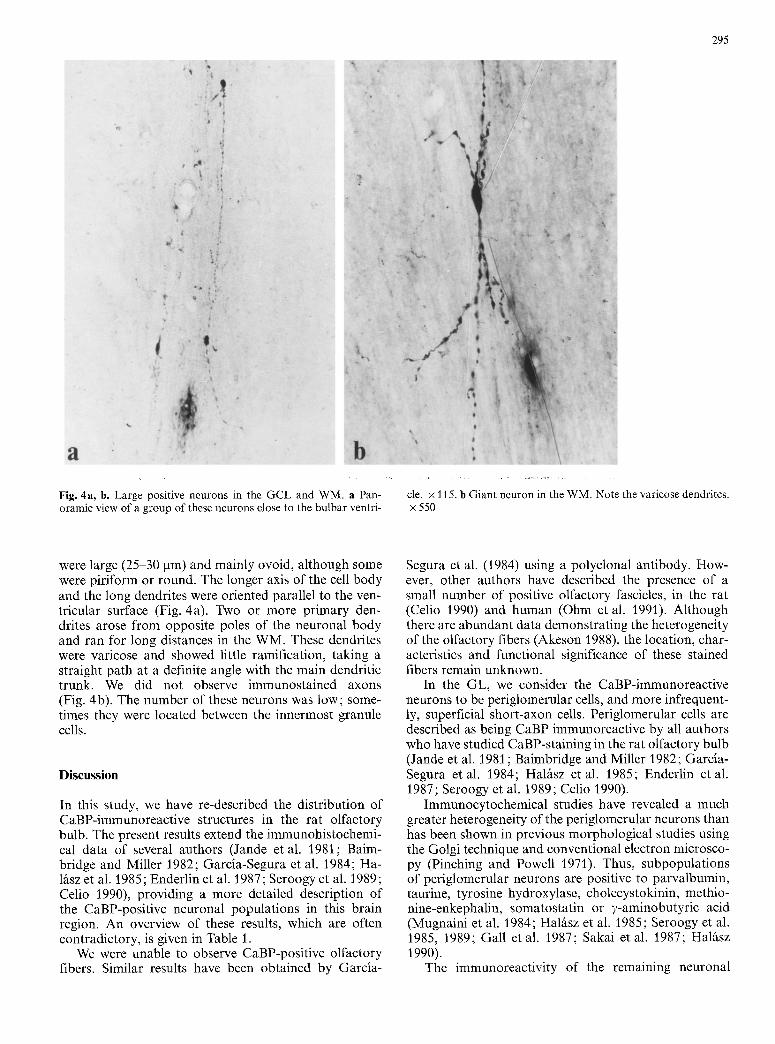
Fig. 4a, b. Large positive neurons in the GCL and WM. a Pan- oramic view of a group of these neurons close to the bulbar ventri-
cle. x 115. b Giant neuron in the WM. Note the varicose dendrites. x 550
were large (25-30 pro) and mainly ovoid, although some were piriform or round. The longer axis of the cell body and the long dendrites were oriented parallel to the ven- tricular surface (Fig. 4a). Two or more primary den- drites arose from opposite poles of the neuronal body and ran for long distances in the WM. These dendrites were varicose and showed little ramification, taking a straight path at a definite angle with the main dendritic trunk. We did not observe immunostained axons (Fig. 4b). The number of these neurons was low; some- times they were located between the innermost granule cells.
Discussion
In this study, we have re-described the distribution of CaBP-immunoreactive structures in the rat olfactory bulb. The present results extend the immunohistochemi- cal data of several authors (Jande et al. 1981; Bairn- bridge and Miller 1982; Garcia-Segura et al. 1984; Ha- lb, sz et al. 1985; Enderlin et al. 1987; Seroogy et al. 1989; Celio 1990), providing a more detailed description of the CaBP-positive neuronal populations in this brain region. An overview of these results, which are often contradictory, is given in Table 1.
We were unable to observe CaBP-positive olfactory fibers. Similar results have been obtained by Garcia-
Segura et al. (1984) using a polyclonal antibody. How- ever, other authors have described the presence of a small number of positive olfactory fascicles, in the rat (Celio 1990) and human (Ohm et al. 1991). Although there are abundant data demonstrating the heterogeneity of the olfactory fibers (Akeson 1988), the location, char- acteristics and functional significance of these stained fibers remain unknown.
In the GL, we consider the CaBP-immunoreactive neurons to be periglomerular cells, and more infrequent- ly, superficial short-axon cells. Periglomerular cells are described as being CaBP immunoreactive by all authors who have studied CaBP-staining in the rat olfactory bulb (Jande et al. 1981; Baimbridge and Miller 1982; Garcia- Segura etal. 1984; Halfisz et al. 1985; Enderlin etal. 1987; Seroogy et al. 1989; Celio 1990).
Immunocytochemical studies have revealed a much greater heterogeneity of the periglomerular neurons than has been shown in previous morphological studies using the Golgi technique and conventional electron microsco- py (Pinching and Powell 1971). Thus, subpopulations of periglomerular neurons are positive to parvalbumin, taurine, tyrosine hydroxylase, cholecystokinin, methio- nine-enkephalin, somatostatin or ?-aminobutyric acid (Mugnaini et al. 1984; Halfisz et al. 1985; Seroogy et al. 1985, 1989; Gall et al. 1987; Sakai et al. 1987; Halfisz 1990).
The immunoreactivity of the remaining neuronal

296
types located in this layer, viz., the superficial short-axon cells and external tufted cells, is more controversial. Our results agree with those of Seroogy et al. (1989) who have shown that CaBP occurs in superficial short-axon cells located at the border between the GL and EPL, and with those of Jande et al. (1981) and Garcia-Segura et al. (1984) who have demonstrated that it is absent in the external tufted cells. The morphological character- istics of the immunopositive cells coincide with those described by Schneider and Macrides (1978) who used the Golgi method; moreover, these CaBP-positive neu- rons frequently show stained, highly branched, local ax- ons, whereas the tufted cells, such as the mitral cells, are projecting neurons. However, other authors have re- ported CaBP-positive external tufted cells in the rat (Ha- 1/tsz et al. 1985; Celio 1990) and in human (Ohm et al. 1991). Golgi-impregnated superficial short-axon cells seem to represent a heterogeneous population (Pinching and Powell 1971), in good agreement with our results.
In the innermost region of this layer, we have found another type of CaBP-stained interneuron. These cells, identified as van Gehuchten cells, have not been referred to by any other author as being CaBP-immunoreactive, but they appear frequently in our sections.
At the border between the IPL and the GCL, we have observed two types of immunoreactive neurons identified as horizontal and vertical cells of Cajal. These two populations belong to the oriented neurons de- scribed by Schneider and Macrides (1978). Celio (1990) has also reported the presence of CaBP-immunostaining in the horizontal and vertical of Cajal cells. The results in the GCL are however contradictory; Baimbridge and Miller (1982) and Enderlin et al. (1987) have not found CaBP-positive neurons in this layer. Furthermore, im- munostained elements in the GCL have been identified as granule cells in the rat (Garcla-Segura et al. 1984) and in human (Ohm et al. 1991), or simply termed gran- ule-layer cells (Halfisz et al. 1985).
A type of CaBP-positive cell characterized by its large size and its oriented varicose dendrites is present in the innermost region of the olfactory bulb, around the ven- tricle. These neurons mentioned previously by Celio (1990) have been described by Mugnaini et al. (1984) as glutamate decarboxylase-immunoreactive elements, but they are only immunoreactive when the animals have been previously treated with colchicine. Ohm et al. (1991) have observed CaBP-stained neurons in the hu- man olfactory bulb; the dendrites of these neurons show a similar branching pattern (although they have not pre- viously been described as varicose) to that of our large neurons. We consider this observation significant be- cause these neurons have not been described in previous studies on the neuronal typology of the olfactory bulb (for reviews, see Macrides and Davis 1983; Halfisz 1990). They are presumably difficult to observe using tech- niques other than immunohistochemical methods.
We therefore conclude that not all cell types described in previous reports have been clearly observed in our sections (e.g., external tufted cells and granule cells). However, on the other hand, we have found different types of CaBP-positive interneurons (short-axon cells,
van Gehuchten cells, horizontal cells and vertical cells of Cajal), and a group of large neurons located around the ventricle whose functional role in the olfactory bulb is unknown. In our material, bulbar projection neurons, mitral and tufted cells, and extrinsic afferents, olfactory and centrifugal fibers, are negative.
Finally, the distribution of CaBP observed in our study is clearly different from that of calretinin as re- ported by Jacobowitz and Winsky (1991). Calretinin has been localized in granule cells (the most abundant im- munopositive neurons), olfactory fibers, periglomerular ceils and, with low staining intensity, in mitral cells. Thus, the only type of neuron where a colocalization of CaBP and calretinin could be expected is the periglo- merular cell.
Acknowledgements. The authors want to express their gratitude to Dr. F. SS.nchez, Miss E.L. Shorten and an anonymous referee for revising the manuscript and Dr. M.R. Celio for his continuous support. This work was supported by grants from the University of Salamanca (" Acciones Concertadas de Investigaci6n") to J.R.A. and J.A.
References
Akeson RA (/988) Primary olfactory neuron subclasses. In: Mar- golis FL, Getchell TV (eds) Molecular neurobiology of the ol- factory system. Molecular, membranous and cytological stu- dies. Plenum Press, New York, pp 297-318
Baimbridge KG, Miller JJ (1982) Irnmunohistochemical localiza- tion of calcium binding protein in the cerebellum, hippocampal formation and olfactory bulb of the rat. Brain Res 245 : 223-229
Baimbridge KG, Miller J J, Parkes CO (1982) Calcium-binding pro- tein distribution in the rat brain. Brain Res 239:519-525
Cello MR (1989) Calcium binding proteins in the brain. Arch Ital Anat Embriol 94:227--236
Celio MR (1990) Calbindin D-28k and parvalbumin in the rat nervous system. Neuroscience 35:375-475
Cello MR, Baier W, Sch~rer E, Gregersen H J, Viragh P de, Nor- man AW (1990) Monoclonal antibodies directed against the calcium binding protein calbindin D-28k. Cell Calcium 11 : 599- 602
Enderlin S, Norman AW, Celio Mr (1987) Ontogeny of the calcium binding protein calbindin D-28k in the rat nervous system. Anat Embryol 177 : 15-28
Feldman SC, Christakos S (:1983) Vitamin D-dependent calcium- binding protein in rat brain: biochemical and immunocyto- chemical characterization. Endocrinology 112: 290-302
Gall CM, Hendry SHC, Seroogy KB, Jones EG, Haycock JW (1987) Evidence for coexistence of GABA and dopamine in neurons of the rat olfactory bulb. J Comp Neurol 266:307-318
Garcia-Segura LM, Baetens D, Roth J, Norman AW, Orci L (1984) Immunohistochemical mapping of calcium-binding protein im- munoreactivity in the rat central nervous system. Brain Res 296 : 75-86
Halfisz N (:1990) The vertebrate olfactory system. Chemical neu- roanatomy, function and development. Akad6miai Kiad6, Bu- dapest
Halfisz N, H6kfelt T, Norman AW, Goldstein M (1985) Tyrosine hydroxylase and 28k-vitamin D-dependent calcium binding protein are localized in different subpopulations of periglomer- ular cells of the rat olfactory bulb. Neurosci Lett 61 ::103-107
Hsu SM, Raine L, Fanger H (198:1) The use of avidin biotin peroxi- dase complex (ABC) in immunoperoxidase techniques. A com- parison between ABC and unlabeled antibody (peroxidase) pro- cedures. J Histochem Cytochem 29 : 57%590

297
Jacobowitz DM, Winsky L (1991) Immunocytochemical tocaliza- tion of calretinin in the forebrain of the rat. J Comp Neurol 304:198-218
Jande SS, Maler L, Lawson DEM (1981) Immunohistochemical mapping of vitamin D-dependent calcium binding protein in brain. Nature 294: 765-767
Kretsinger RH, Mann JE, Simmonds JG (1982) Model of facili- tated diffusion of calcium by the intestinal calcium-binding pro- tein. In: Norman AW, Schaefer K, Grigoleit HG, Herrath DV (eds) Vitamin D, chemical, biochemical and clinical endocrinol- ogy of calcium metabolism, de Gruyter, Berlin, pp 233-248
Macrides F, Davis BJ (1983) The olfactory bulb. In: Emson PC (ed) Chemical neuroanatomy. Raven Press, New York, pp 391- 426
Mugnaini E, Wouterlood FG, Dahl AL, Oertel WH (1984) Immu- nocytochemical identification of GABAergic neurons in the main olfactory bulb of the rat. Arch Ital Biol 122:83-113
Ohm TG, Miiller H, Braak E (1991) Calbindin-D-28k-like immu- noreactive structures in the olfactory bulb and anterior olfacto- ry nucleus of the human adult: distribution and cell typology - partial complementarity with parvalbumin. Neuroscience 42:823-840
Persechini A, Nancy DM, Kretsinger RH (1989) The EF-hand family of calcium-modulated proteins. Trends Neurosci 12 : 462-467
Pinching AJ, Powell TPS (1971) The neuron types of the glomeru- lar layer of the olfactory bulb. J Cell Sci 9:305-345
Rogers JH (1987) Calretinin: a gene for a novel calcium-binding protein expressed principally in neurons. J Cell Biol 105:1343- 1353
Sakai M, Yoshida M, Karasawa N, Teramura-Ito M, Nagatsu T, Nagatsu I (1987) Coexistence of taurine-like and tyrosine hyd-
roxytase-like immunoreactivities in some periglomerular cells of the rat main olfactory bulb. Biogenic Amines 4:457-464
Schneider SP, Macrides F (1978) Laminar distributions of inter- neurons in the main olfactory bulb of the adult hamster. Brain Res Bull 3 : 73-82
S~quier JM, Hunziker W, Richards G (1988) Localization of cal- bindin D-28k mRNA in rat tissues by in situ hybridization. Neurosci Lett 86:155-160
S6quier JM, Hunziker W, Andressen C, Celio MR (1990) Calbindin D-28k: protein and mRNA localization in the rat brain. Eur J Neurosci 2:1118-1126
Seroogy KB, Brecha N, Gall C (1985) Distribution of cholecystok- inin-like immunoreactivity in the rat main olfactory bulb. J Comp Neurol 239 : 373-383
Seroogy K, H6kfelt T, Buchan A, Brown JC, Terenius L, Norman AW, Goldstein M (1989) Somatostatin-like immunoreactivity in rat main olfactory bulb: extent of coexistence with neuropep- tide Y-, tyrosine hydroxylase- and vitamin D-dependent calci- um binding protein-like immunoreactivities. Brain Res 496:389-396
Somogyi P, Takagi M (1982) A note on the use of picric acid- paraformaldehyde-glutaraldehyde fixative for correlated light and electron microscopic immunocytochemistry. Neuroscience 7:1779-1784
Taylor AN (1974) Chick brain calcium-binding protein: compari- son with intestinal vitamin-D induced calcium-binding protein. Arch Biochem Biophys 161 : 100-108
Yamamoto T, Carr PA, Baimbridge KG, Nagy JI (1989) Parvalbu- min- and calbindin D28k-immunoreactive neurons in the super- ficial layers of the spinal cord dorsal horn of rat. Brain Res Bull 23 : 493-508
Related Documents











