Research Report Calbindin and calretinin immunoreactivity in the retina of adult and larval sea lamprey Begoña Villar-Cheda, Xesús Manoel Abalo, Ramón Anadón, María Celina Rodicio ⁎ Department of Cell Biology and Ecology, University of Santiago de Compostela, 15782-Santiago de Compostela, Spain ARTICLE INFO ABSTRACT Article history: Accepted 6 November 2005 Available online 20 December 2005 The presence of calretinin and calbindin immunoreactivity is studied in the retina of larval and adult lamprey and their respective distributions are compared. Calretinin distribution is also studied in the retina of transforming stages. Western blot analysis in brain extracts showed a 29-kDa band with both polyclonal anti-calbindin and anti-calretinin antibodies. Calbindin and calretinin immunoreactivity has shown a partially different distribution. In the adult retina large and small bipolar cells, with respectively stratified or diffuse axons, the inner row of horizontal cells and ganglion cells and/or some amacrine cells were labeled with anti-calretinin antibody. The anti-calbindin antibody labels the same cell types except most of ganglion cells, but the label was less conspicuous. Therefore, the possible existence of these two calcium-binding proteins in the central nervous system of the sea lamprey could be discussed. In the differentiated central retina of larval lampreys, numerous calretinin immunoreactive bipolar and ganglion cells were observed, while, in the lateral retina, only ganglion cells were labeled, accordingly with the lack of differentiation of other neural cell types. CR-ir bipolar cells appeared in the retina by the stage 5 of transformation, i.e. about the time when differentiation of photoreceptors occurs. The comparison of the distribution of calretinin and calbindin between adult and larval central retina of lampreys shows striking differences that could be related to the different functionality of eyes in these two stages of the life cycle of lampreys. In addition, this is the first report on the presence of calcium-binding proteins in the larval and transforming lamprey retina, on the presence of calretinin- and calbindin-immunoreactive horizontal cells in adult lamprey retinas and on the differential stratification of bipolar cell terminals. © 2005 Elsevier B.V. All rights reserved. Theme: Sensory systems Topic: Retina and photoreceptors Keywords: Calcium-binding protein Immunohistochemistry Development Retina Petromyzon marinus Abbreviations: HCL, horizontal cell sublayer HCL1, horizontal cell sublayer 1 of the INL HCL2, horizontal cell sublayer 2 of the INL INL, inner nuclear layer IPL, inner plexiform layer IPLi, inner sublayer of the IPL IPLo, outer sublayer of the IPL Lc, Landolt's club NbL, neuroblastic layer NFL, nerve fiber layer OLM, outer limiting membrane ON, optic nerve ONL, outer nuclear layer PE, pigment epithelium Ph, photoreceptor layer BRAIN RESEARCH 1068 (2006) 118 – 130 ⁎ Corresponding author. Fax: +34 981 596904. E-mail address: [email protected] (M.C. Rodicio). 0006-8993/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2005.11.014 available at www.sciencedirect.com www.elsevier.com/locate/brainres

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res
Research Report
Calbindin and calretinin immunoreactivity in the retina ofadult and larval sea lamprey
Begoña Villar-Cheda, Xesús Manoel Abalo, Ramón Anadón, María Celina Rodicio⁎
Department of Cell Biology and Ecology, University of Santiago de Compostela, 15782-Santiago de Compostela, Spain
A R T I C L E I N F O
⁎ Corresponding author. Fax: +34 981 596904.E-mail address: [email protected] (M.C. Rod
0006-8993/$ – see front matter © 2005 Elsevidoi:10.1016/j.brainres.2005.11.014
A B S T R A C T
Article history:Accepted 6 November 2005Available online 20 December 2005
The presence of calretinin and calbindin immunoreactivity is studied in the retina of larvaland adult lamprey and their respective distributions are compared. Calretinin distributionis also studied in the retina of transforming stages. Western blot analysis in brain extractsshowed a 29-kDa band with both polyclonal anti-calbindin and anti-calretinin antibodies.Calbindin and calretinin immunoreactivity has shown a partially different distribution. Inthe adult retina large and small bipolar cells, with respectively stratified or diffuse axons,the inner row of horizontal cells and ganglion cells and/or some amacrine cells were labeledwith anti-calretinin antibody. The anti-calbindin antibody labels the same cell types exceptmost of ganglion cells, but the label was less conspicuous. Therefore, the possible existenceof these two calcium-binding proteins in the central nervous system of the sea lampreycould be discussed. In the differentiated central retina of larval lampreys, numerouscalretinin immunoreactive bipolar and ganglion cells were observed, while, in the lateralretina, only ganglion cells were labeled, accordingly with the lack of differentiation of otherneural cell types. CR-ir bipolar cells appeared in the retina by the stage 5 of transformation,i.e. about the time when differentiation of photoreceptors occurs. The comparison of thedistribution of calretinin and calbindin between adult and larval central retina of lampreysshows striking differences that could be related to the different functionality of eyes inthese two stages of the life cycle of lampreys. In addition, this is the first report on thepresence of calcium-binding proteins in the larval and transforming lamprey retina, on thepresence of calretinin- and calbindin-immunoreactive horizontal cells in adult lampreyretinas and on the differential stratification of bipolar cell terminals.
© 2005 Elsevier B.V. All rights reserved.
Theme:Sensory systemsTopic:Retina and photoreceptors
Keywords:Calcium-binding proteinImmunohistochemistryDevelopmentRetinaPetromyzon marinus
Abbreviations:HCL, horizontal cell sublayerHCL1,horizontal cell sublayer1of the INLHCL2,horizontal cell sublayer2of the INLINL, inner nuclear layerIPL, inner plexiform layerIPLi, inner sublayer of the IPLIPLo, outer sublayer of the IPLLc, Landolt's clubNbL, neuroblastic layerNFL, nerve fiber layerOLM, outer limitingmembraneON, optic nerveONL, outer nuclear layerPE, pigment epitheliumPh, photoreceptor layer
icio).
er B.V. All rights reserved.
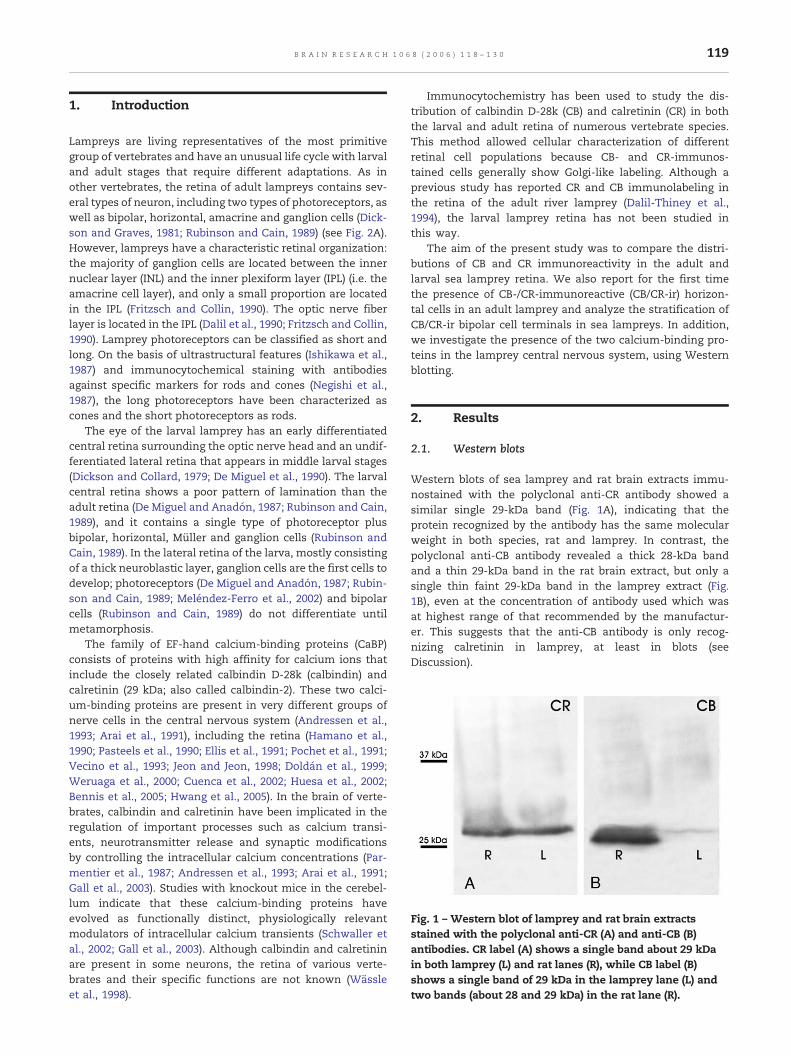
Fig. 1 – Western blot of lamprey and rat brain extractsstained with the polyclonal anti-CR (A) and anti-CB (B)antibodies. CR label (A) shows a single band about 29 kDain both lamprey (L) and rat lanes (R), while CB label (B)shows a single band of 29 kDa in the lamprey lane (L) andtwo bands (about 28 and 29 kDa) in the rat lane (R).
119B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
1. Introduction
Lampreys are living representatives of the most primitivegroup of vertebrates and have an unusual life cycle with larvaland adult stages that require different adaptations. As inother vertebrates, the retina of adult lampreys contains sev-eral types of neuron, including two types of photoreceptors, aswell as bipolar, horizontal, amacrine and ganglion cells (Dick-son and Graves, 1981; Rubinson and Cain, 1989) (see Fig. 2A).However, lampreys have a characteristic retinal organization:the majority of ganglion cells are located between the innernuclear layer (INL) and the inner plexiform layer (IPL) (i.e. theamacrine cell layer), and only a small proportion are locatedin the IPL (Fritzsch and Collin, 1990). The optic nerve fiberlayer is located in the IPL (Dalil et al., 1990; Fritzsch and Collin,1990). Lamprey photoreceptors can be classified as short andlong. On the basis of ultrastructural features (Ishikawa et al.,1987) and immunocytochemical staining with antibodiesagainst specific markers for rods and cones (Negishi et al.,1987), the long photoreceptors have been characterized ascones and the short photoreceptors as rods.
The eye of the larval lamprey has an early differentiatedcentral retina surrounding the optic nerve head and an undif-ferentiated lateral retina that appears in middle larval stages(Dickson and Collard, 1979; De Miguel et al., 1990). The larvalcentral retina shows a poor pattern of lamination than theadult retina (De Miguel and Anadón, 1987; Rubinson and Cain,1989), and it contains a single type of photoreceptor plusbipolar, horizontal, Müller and ganglion cells (Rubinson andCain, 1989). In the lateral retina of the larva, mostly consistingof a thick neuroblastic layer, ganglion cells are the first cells todevelop; photoreceptors (De Miguel and Anadón, 1987; Rubin-son and Cain, 1989; Meléndez-Ferro et al., 2002) and bipolarcells (Rubinson and Cain, 1989) do not differentiate untilmetamorphosis.
The family of EF-hand calcium-binding proteins (CaBP)consists of proteins with high affinity for calcium ions thatinclude the closely related calbindin D-28k (calbindin) andcalretinin (29 kDa; also called calbindin-2). These two calci-um-binding proteins are present in very different groups ofnerve cells in the central nervous system (Andressen et al.,1993; Arai et al., 1991), including the retina (Hamano et al.,1990; Pasteels et al., 1990; Ellis et al., 1991; Pochet et al., 1991;Vecino et al., 1993; Jeon and Jeon, 1998; Doldán et al., 1999;Weruaga et al., 2000; Cuenca et al., 2002; Huesa et al., 2002;Bennis et al., 2005; Hwang et al., 2005). In the brain of verte-brates, calbindin and calretinin have been implicated in theregulation of important processes such as calcium transi-ents, neurotransmitter release and synaptic modificationsby controlling the intracellular calcium concentrations (Par-mentier et al., 1987; Andressen et al., 1993; Arai et al., 1991;Gall et al., 2003). Studies with knockout mice in the cerebel-lum indicate that these calcium-binding proteins haveevolved as functionally distinct, physiologically relevantmodulators of intracellular calcium transients (Schwaller etal., 2002; Gall et al., 2003). Although calbindin and calretininare present in some neurons, the retina of various verte-brates and their specific functions are not known (Wässleet al., 1998).
Immunocytochemistry has been used to study the dis-tribution of calbindin D-28k (CB) and calretinin (CR) in boththe larval and adult retina of numerous vertebrate species.This method allowed cellular characterization of differentretinal cell populations because CB- and CR-immunos-tained cells generally show Golgi-like labeling. Although aprevious study has reported CR and CB immunolabeling inthe retina of the adult river lamprey (Dalil-Thiney et al.,1994), the larval lamprey retina has not been studied inthis way.
The aim of the present study was to compare the distri-butions of CB and CR immunoreactivity in the adult andlarval sea lamprey retina. We also report for the first timethe presence of CB-/CR-immunoreactive (CB/CR-ir) horizon-tal cells in an adult lamprey and analyze the stratification ofCB/CR-ir bipolar cell terminals in sea lampreys. In addition,we investigate the presence of the two calcium-binding pro-teins in the lamprey central nervous system, using Westernblotting.
2. Results
2.1. Western blots
Western blots of sea lamprey and rat brain extracts immu-nostained with the polyclonal anti-CR antibody showed asimilar single 29-kDa band (Fig. 1A), indicating that theprotein recognized by the antibody has the same molecularweight in both species, rat and lamprey. In contrast, thepolyclonal anti-CB antibody revealed a thick 28-kDa bandand a thin 29-kDa band in the rat brain extract, but only asingle thin faint 29-kDa band in the lamprey extract (Fig.1B), even at the concentration of antibody used which wasat highest range of that recommended by the manufactur-er. This suggests that the anti-CB antibody is only recog-nizing calretinin in lamprey, at least in blots (seeDiscussion).

120 B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
2.2. Antibody binding
As noted, immunocytochemistry with the monoclonal anti-CR and anti-CB antibodies (see Experimental procedures) didnot label any structures in either the retina or the brain of thesea lamprey (data not shown). Instead, the use of the rabbitpolyclonal antibodies against CR and CB led to immunolabel-ing of numerous neurons and fibers throughout the brain ofboth larvae and adults (not shown) and of some neurons ofthe adult and larval retinae. Therefore, all our immunocyto-chemical results were obtained with these polyclonal antibo-dies, as they are described below.
2.3. Calretinin immunoreactivity in adult sea lampreyretina
The organization of the adult sea lamprey retina is somewhatdifferent from that observed in other vertebrates, as indicatedin the Introduction section and observed in Fig. 2A. Briefly, thephotoreceptor layer consists of two types of photoreceptorcell, short and long, distinguishable by the different locationand appearance of their inner and outer segments. The innernuclear layer consists of two layers of large horizontal cells,outer (HCL1) and inner (HCL2), and an inner region with ama-crine cells and displaced ganglion cells, some of which aregiant. The thick inner plexiform layer shows the bundles ofoptic fibers in its outer region, limiting with the INL, andscarce neurons (orthotopic ganglion cells, displaced amacrinecells) are located close to the inner limiting membrane (seeFritzsch and Collin, 1990).
In the adult retina, several cell typeswere distinguishedwiththe anti-CR antibody (Figs. 2B, 3): large and small bipolar cells,horizontal cells, amacrine cells and ganglion cells. The mostconspicuous elements were the large bipolar cells, which wereintensely labeled. Their perikarya were located in the outer partof the INL (INLo) external to the HCL1 cells, having a pyramidalappearance with the apex of the pyramid (inner fibers) towardthe inner part of the INL (INLi) (Figs. 2B, 3A). Most of themexhibited a thick apical process, Landolt's club, which runs tothe outer limitingmembrane (OLM) (Fig. 3A). These large bipolarcells gave rise to 3–5 thick dendrites that coursed horizontally in
Fig. 2 – (A) Hematoxylin eosin-stained section of the adult sea lastructure. (B) Schematic drawing of the CR/CB-ir cells of the retinh, horizontal cell; lb, large bipolar cell; sb, small bipolar cell. For
the outer plexiform layer (OPL), branching abundantly and giv-ing off thin ascending appendages ending at the base of thephotoreceptors (Figs. 3A–C). Dendrite branching was better ob-served in tangential sections to this layer, whereas ascendingappendages were better seen in vertical sections. A thick axonran to the IPL where it often showed a bulbous protrusion andgave rise to several coarsely beaded horizontal collaterals thatwere stratified in the outer part of the IPL (IPLo) (Figs. 3A, E, J, K).These collaterals are apposedwith terminal varicosities of othersimilar bipolar cells, forming a characteristic CR-ir network(Figs. 3D, K). This network was better appreciable in tangentialsections of the IPL (Fig. 3D).
The small CR-ir bipolar cells are mainly located in the INLi,though some of them were located in the outer half (Figs. 3F–H). Their perikarya were oval and gave rise to a thin dendritethat ran to the OPL where it ramified scarcely close to the baseof the photoreceptors (Fig. 3F) and gave rise to very thin ter-minal appendages. The thin axon gave rise to a diffuse termi-nal arbor that branched mainly in the inner part of the IPL(IPLi) (Figs. 3G, I). The lateral extension of dendritic and axonaltrees is much smaller in small bipolar cells than in largebipolar cells. Occasionally, very thin Landolt's clubs of thesmall bipolar cells were seen coursing to the OLM (Fig. 3H).
The adult sea lamprey retina shows two rows of largehorizontal cells, outer and inner (HCL1 and HCL2, respective-ly), forming a thick sublayer located in the INLo below thelarge bipolar cell perikarya (Fig. 2A). The large HCL2 cellswere moderately CR-ir (Figs. 3A, F), with their perikarya hori-zontally orientated. In tangential sections, these CR-ir HCL2cells showed a lobulate profile and contacted other HCL2 cells,forming a mosaic and leaving small holes between throughwhich processes of bipolar cells and other retinal cell types(for instance, Müller cells) cross this layer. The HCL2 horizon-tal cells also presented faintly CR-ir-thick dendrites thatcoursed between the HCL1 and HCL2 sublayers and gave offprocesses ascending to the OPL among the CR-negative HCL1cells (Fig. 3F). The faint staining of these processes precludeddetailed study of the ending pattern in the OPL.
CR immunoreactivity was also present in cells located inthe IPL and in the INLi (Figs. 3F–H, J). Moderately CR-ir bipolarcells extending long processes horizontally in the IPL and
mprey retina showing the layered organization of thisa of the adult sea lamprey. a, amacrine cell; g, ganglion cell;other abbreviations, see list. Scale bar = 25 μm.

Fig. 3 – Photomicrographs of sections of the adult retina showing the organization of the CR-ir structures. (A) Vertical sectionof the retina showing large bipolar (wide arrows) and type 1 and type 2 horizontal cells (black andwhite asterisks, respectively).Arrowhead points to the Landolt's club. (B) Tangential section of the retina showing the dendrites of a large bipolar neuron inthe OPL. (C) Vertical section showing the ascending appendages of a large bipolar cell. (D) Tangential section of the retinashowing the plexus of large bipolar axon collaterals in the IPLo. (E) Vertical section of the IPLo showing the stratification of largebipolar cell axon collaterals. (F) Vertical section showing CR immunoreactivity in a small bipolar cell (double arrow), HCL2horizontal cells (white asterisks) and amacrine cells (thin arrow). Note that HCL1 horizontal cells are CR-negative (blackasterisk). The arrowhead points to the bifurcation of the small bipolar cell dendrite (arrowhead). (G) Section showing a smallbipolar cell (double arrow), large bipolar cells (thick arrow) and amacrine cells (thin arrow). Note the branching of the smallbipolar cell axon in the IPLi. (H) Detail of a small bipolar cell showing the Landolt's club. (I) Detail of the axonof a small bipolar cell(arrowheads) branching in the IPLi close to the inner limiting membrane. (J) Vertical section showing CR-ir ganglion cells(outlined arrows). (K) Section showing a bundle of CR-ir optic nerve fibers coursing in the outer part of the IPLo. Note thestratified axon collaterals of a large bipolar cell (arrowhead). For abbreviations, see list. Scale bars = 25 μm (A, B, D, F–H, J–K)and 12.5 μm (C, E, I).
121B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
located either in the inner part of the IPL or in the inner part ofthe INL were considered as orthotopic and displaced ganglioncells, respectively (Figs. 2B, 3J). No CR immunoreactivity wasobserved in the displaced giant ganglion cells. The bundles ofoptic fibers that coursed in the outer part of the IPLo (Figs. 2B,
3K) were moderately CR-ir, and the optic nerve also containedCR-ir fibers. Small, moderate to strongly CR-ir rounded cellslocated in the INLi were tentatively considered as amacrinecells (Figs. 3F–H). The branching pattern of processes of thesecells could not be determined.

122 B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
2.4. Calretinin immunoreactivity in larval lamprey retina
The retina of larval sea lampreys consists of two differentregions, central and lateral (Figs. 4A–B). The central retina isthe only part present in small larvae and varies little in exten-sion throughout larval development (De Miguel and Anadón,1987), showing a differentiated photoreceptor layer, two nu-clear layers, the outer (ONL) and inner (INL) nuclear layer, andtwo plexiform layers, the OPL and the IPL (Fig. 4A). CR immu-noreactivity was observed in bipolar and ganglion cells fromsmall (30 mm) to large larvae (120 mm) (Figs. 4C–E), although
Fig. 4 – (A, B) Photomicrographs of vertical sections through the c(120 mm in length) stained with hematoxylin eosin. In panel B,very thick neuroblastic layer into outer and inner parts. (C) VertiCR-ir bipolar cells (thick arrow). Note that most of the retina has50-mm larva showing the CR-ir bipolar cells (thick arrow) and sothrough a part of the retina of a 120-mm larva showing the clearcells (thick arrow) and the lateral retina consisting mostly of acells (outlined arrow) in both retinal parts. Thin arrows, bundlesoutlined arrows, ganglion cells; double arrowhead, Landolt's clucentral (F–G) and lateral (H) retina of a 114-mm larva showing ththrough the optic nerve head (asterisks). Arrowhead, bipolar celbipolar cell axons; outlined arrows, ganglion cells; thick arrowssee list. Scale bars = 25 μm (A–D, F–H), 50 μm (E).
immunostaining intensity increased markedly with develop-ment. In small larvae (30–50 mm), bipolar cell perikarya ap-pear faint to moderately stained (Figs. 4C–D). In large larvae(90–120 mm), bipolar cells were intensely CR-ir, had a pear-shaped soma, and were located in the INLo (Figs. 4C–G). Den-drites and axons of these cells were clearly stained (Figs. 4E–G). Dendrites appeared to branch scarcely at the boundarybetween the INL and ONL, i.e. in the OPL, and some Landolt'sclubs were seen coursing to the OLM (Fig. 4F). The axonsbranched occasionally in the IPL and probably contacted gan-glion cells, which appeared faintly stained (Fig. 4F).
entral (A) and lateral (B) retina of a large sea lamprey larvanote that the primordial horizontal cell layer subdivides thecal section of the eye of a 30-mm larva showing a layer ofa photoreceptor layer. (D) Similar section of the retina of ame CR-ir ganglion cells (outlined arrow). (E) Vertical sectiondistinction between the central retina exhibiting CR-ir bipolarthick neuroblastic layer. Note the presence of CR-ir ganglionof CR-ir optic fibers coursing to the optic nerve head;b. (F–H) Photomicrographs of vertical sections through thee CR-ir structures. In panels F and G, the sections passl dendrites; double arrowhead, Landolt's club; small arrows,, bipolar cells; thin arrow, optic fibers. For abbreviations,

123B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
In the central retina, previous tract-tracing studies fromour laboratory (Anadón et al., 1998; De Miguel et al., 1989)indicate that ganglion cells are located on the INLi andthroughout the whole IPL. In these locations, faintly CR-ircells were observed (Figs. 4D–F). Some ganglion cells are alsopresent in the region immediately surrounding the emer-gence of the optic nerve (Fig. 4G). The intensity of labelingwas clearly higher in bipolar than in ganglion cells and washigher in larger larvae. Faintly labeled cell processes fromganglion cells coursed in the IPL (Figs. 4E–F) and the opticnerve. Owing to the large number of ascending processes ofbipolar cells and the faint staining of ganglion cells, we couldnot determine whether some CR-ir ganglion cells have as-cending processes coursing to the OPL (i.e. biplexiform gangli-on cells). Biplexiform ganglion cells of sea lamprey were firstdescribed with tract-tracing methods in our laboratory (DeMiguel et al., 1989).
The lateral retina is only present in larvae larger than 55–60 mm in length and expands progressively during larvalgrowth (De Miguel and Anadón, 1987). It consists of a neuro-blastic layer with only a narrow primordial IPL with associatedganglion cells on its vitreal side (Fig. 4B). In this retinal part,photoreceptors are lacking until middle stages of metamor-phosis. In premetamorphic larvae (larvae more than 100 mmlong), the neuroblastic layer is thicker and becomes dividedinto inner and outer neuroblastic layers by a thin primordialhorizontal cell sublayer. In larvae with lateral retina, moder-ately CR-ir putative ganglion cells were observed close to theprimordial IPL of the lateral retina, and their axons (opticfibers) were also faintly stained (Fig. 4H).
2.5. Calretinin immunoreactivity in the retina oftransforming and postmetamorphic lamprey
The lateral retina of the larva suffers dramatic changes duringtransformation, acquiring the adult layering and differentiat-ing the two types of photoreceptors (De Miguel and Anadón,1987; De Miguel, 1988). During the first stages of metamorpho-sis (M1–M3), the IPL becomes considerably thickened. CR-ircells were observed on both sides of the IPL (Fig. 5A), i.e. closeto the inner limitingmembrane (putative ganglion cells) and inthe inner border of the neuroblastic layer (putative amacrinecells and displaced ganglion cells). No CR-ir cells were ob-served in the central region of the thick neuroblastic layer ofM1–M3 stages. In stageM5 of transformation, the retina showsphotoreceptors throughout all its extension. In this retina, CR-ir cells were observed in the outer region of the inner nuclearlayer, by this stage clearly differentiable from the outer nuclearlayer (Figs. 5B–C). A Landolt's club can be sometimes observed(Fig. 5C), indicating that these external CR-ir cells are bipolarcells. Distinction between large and small bipolar cells was notpossible at this stage, although perikarya of putative bipolarcells were located at different heights (Fig. 5C). CR-ir cells wereobserved on both sides of the inner plexiform layer, as indi-cated above for the M3 stage. The IPL of M5 stages alsocontained numerous CR-ir processes.
In recently metamorphosed lampreys, the retina showsthe lamination found in adults, as well as tall photoreceptorsof two types. In the outer part of the INL, the postmeta-morphic retina exhibited two types of CR-ir bipolar cells in-
distinguishable by the size of their perikarya but exhibitingeither outer or inner locations as regards the horizontal cellsublayers (Figs. 5E–G). The presence of axonal terminals in theouter and inner sublayers of the IPL exhibiting a similar mor-phology to that observed in the upstream migrating adult(Figs. 5H, I), which sometimes could be followed to their re-spective perikarya, indicated that these outer and inner cellscorrespond to the large and small CR-ir bipolar cells, respec-tively, as reported in the adult (see Section 2.3). Some CR-irganglion cells in the inner region of the IPL, and CR-ir cells inthe amacrine/ganglion cell sublayer of the INL, were also ob-served (Fig. 5E).
2.6. Calbindin immunoreactivity in adult and larvalretinas
In the retina of upstream migrating adult lamprey, high CBimmunoreactivity was in large and small bipolar cells (Figs.6A–C). The morphology and distribution of these CB-ir cellswere similar to those revealed with the anti-CR antibody. Theinner row of horizontal cells (HCL2) was moderately CB-im-munoreactive (Figs. 6A–C). Most cells located in the INLi wereCB-negative, although a very small proportion were faintlyCB-ir (Fig. 6C). Since occasional optic fibers were faintly CB-ir(Fig. 6B), some of these CB-ir cells might be ganglion cells. Inaddition, some putative orthotopic ganglion cells showedfaint CB immunostaining (Figs. 6A, D). In general, these celltypes showed stronger immunolabeling with the anti-CR an-tibody than with the anti-CB antibody (Fig. 6D). The displacedgiant ganglion cells were CB-negative.
The immunoreactivity to calbindin in the larval retina par-allels that of calretinin, though staining intensity of CB wasgenerally lower than that of CR. First CB-ir bipolar cells wereobserved in the INL of the retina (central retina) of 30–50 mmlarvae (Fig. 7A), corresponding to the bipolar cells observedwith CR immunocytochemistry. A few very faintly stainedcells were also observed near the inner limiting membrane.In large larvae (90–120 mm), the CB-ir bipolar cells of thecentral retina were intensely stained (Figs. 7B–D) and showedan appearance similar to the CR-ir bipolar cells, including thepresence of Landolt's clubs (Fig. 7D). In addition, scarce, faint-ly CB-ir cells were present in the innermost layers of thelateral (Figs. 7E–F) and central retina of large larvae, in similarlocations than the CR-ir cells of the same regions. Bundles ofoptic fibers showed very faint CB staining, suggesting thatthese inner cells were ganglion cells. In the lateral retina ofthese large larvae, no CB-ir cells were observed within thethick neuroblastic layer (Fig. 7E).
The distribution of calbindin in the retina of transformingstages was not investigated.
3. Discussion
3.1. Western blot analysis
CR and CB belong to the large family of EF-hand calcium-binding proteins, and show highly similar sequences, with58% amino acid homology in chick (Rogers, 1987). The presentWestern blot analysis shows that a single 29-kDa protein is
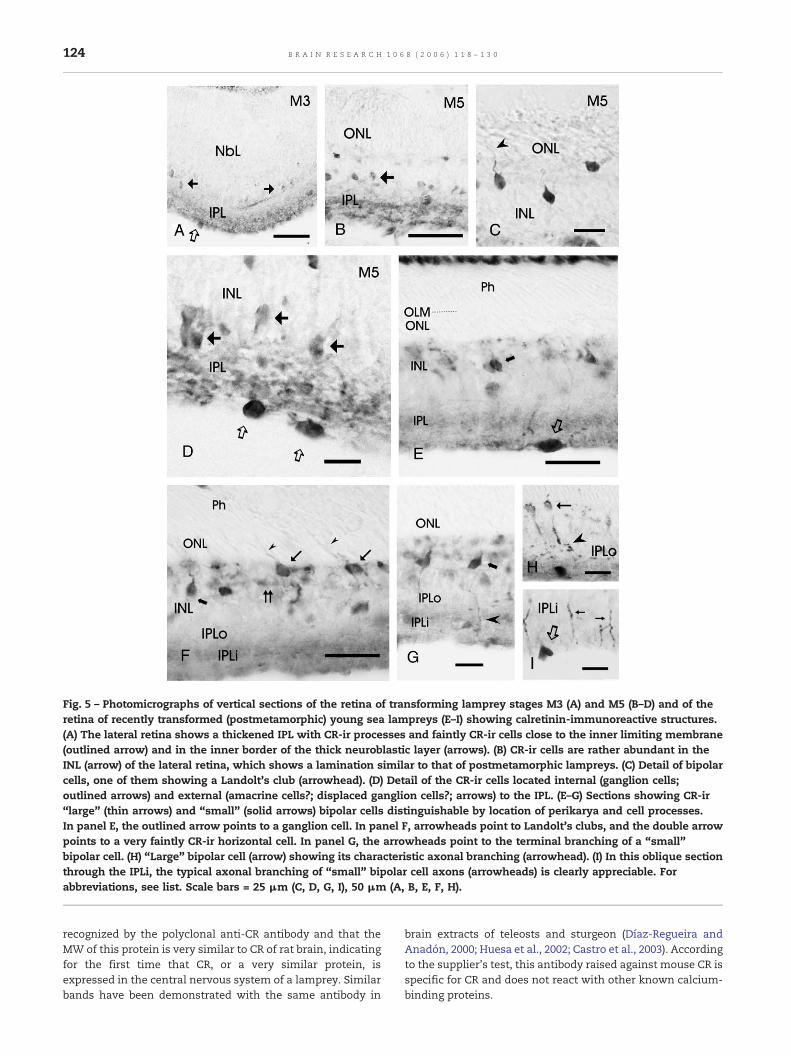
Fig. 5 – Photomicrographs of vertical sections of the retina of transforming lamprey stages M3 (A) and M5 (B–D) and of theretina of recently transformed (postmetamorphic) young sea lampreys (E–I) showing calretinin-immunoreactive structures.(A) The lateral retina shows a thickened IPL with CR-ir processes and faintly CR-ir cells close to the inner limiting membrane(outlined arrow) and in the inner border of the thick neuroblastic layer (arrows). (B) CR-ir cells are rather abundant in theINL (arrow) of the lateral retina, which shows a lamination similar to that of postmetamorphic lampreys. (C) Detail of bipolarcells, one of them showing a Landolt's club (arrowhead). (D) Detail of the CR-ir cells located internal (ganglion cells;outlined arrows) and external (amacrine cells?; displaced ganglion cells?; arrows) to the IPL. (E–G) Sections showing CR-ir“large” (thin arrows) and “small” (solid arrows) bipolar cells distinguishable by location of perikarya and cell processes.In panel E, the outlined arrow points to a ganglion cell. In panel F, arrowheads point to Landolt's clubs, and the double arrowpoints to a very faintly CR-ir horizontal cell. In panel G, the arrowheads point to the terminal branching of a “small”bipolar cell. (H) “Large” bipolar cell (arrow) showing its characteristic axonal branching (arrowhead). (I) In this oblique sectionthrough the IPLi, the typical axonal branching of “small” bipolar cell axons (arrowheads) is clearly appreciable. Forabbreviations, see list. Scale bars = 25 μm (C, D, G, I), 50 μm (A, B, E, F, H).
124 B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
recognized by the polyclonal anti-CR antibody and that theMW of this protein is very similar to CR of rat brain, indicatingfor the first time that CR, or a very similar protein, isexpressed in the central nervous system of a lamprey. Similarbands have been demonstrated with the same antibody in
brain extracts of teleosts and sturgeon (Díaz-Regueira andAnadón, 2000; Huesa et al., 2002; Castro et al., 2003). Accordingto the supplier's test, this antibody raised against mouse CR isspecific for CR and does not react with other known calcium-binding proteins.

Fig. 6 – (A–D) Photomicrographs of vertical sections of the adult retina showing CB-immunoreactive cells. Note CBimmunostaining of large bipolar cells (thick arrows in A and C), small bipolar cells (double arrow in B), horizontal cells (whiteasterisks in A–C) and amacrine cells (thin arrows in A and C). A few faintly CB-stained ganglion cells (outlined arrow in D) werealso present. Fig. 6C depicts the retina near the margin, where HCL1 cells are very scarce or are lacking. Arrowheads (A–C)indicate ascending dendritic appendages. For abbreviations, see list. Scale bars = 25 μm.
125B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
The polyclonal anti-CB antibody recognized a single 29-kDa protein band in lamprey brain extracts, while in ratbrain extracts, it recognizes both a thick 28-kDa band and athin 29-kDa band, suggesting that in the rat it recognizedcalbindin but also calretinin, in line with previous reports(Pasteels et al., 1990; Pochet et al., 1991). This is in agreementwith the supplier's indication that this antiserum shows 10%cross reaction with CR in immunoblotting. In the lampreybrain extracts, unlike in rat, the MW of the faint band stainedby the anti-CB polyclonal antibody corresponded to that of thesingle band demonstrated by the CR antibody.
In the sea lamprey, CR and CB immunoreactivities showrather similar cellular distribution in the retina (presentresults) and in brain (unpublished observations). These resultscast doubt about whether the two calcium-binding proteinsare actually expressed in the lamprey central nervous systemor whether only a single CR/CB protein is recognized by theantibodies used here. Although immunocytochemical resultshave previously been used as an argument in favor of thepresence of the two proteins in the river lamprey retina(Dalil-Thiney et al., 1994), the CB antibodies used in this previ-ous study showed the sameproblemof cross-reactionwith CR,and these authors did not perform Western blotting in lam-prey extracts. In what follows, we nevertheless consider the
two immunoreactivities separately, though it should be bornein mind that neither CB nor CR have been sequenced inlamprey.
3.2. Comparison between calretinin and calbindinimmunoreactivities in the larval and adult lamprey retina
Both CR and CB immunoreactivities showed different patternsin the adult retina and the differentiated central region of thelarval retina. In adults, CR and CB immunoreactivities werepresent in bipolar, horizontal and amacrine cells, while, in thedifferentiated central retina of larvae, only bipolar cells wereCR- and CB-ir. In addition, CR-ir ganglion cells were observedin both adult and larval retinas, but only a few ganglion cellswere CB-ir.
Horizontal cells were only CR- and CB-ir in adult retina,although morphologically identified horizontal cells havebeen described in larvae (De Miguel, 1988; Rubinson andCain, 1989), and the presence of gap junctions between themhas been reported on the basis of electron microscopy (DeMiguel, 1988). Putative amacrine cells have been reported inlarge larvae on the basis of Golgi methods (Rubinson and Cain,1989), but no cells of the central or lateral larval retina showimmunoreactivity with typical amacrine cell markers such as

Fig. 7 – (A) Photomicrograph of a vertical section showing CB-ir bipolar cells (thick arrow) in the retina of a 50-mm larva. (B)Photomicrograph of a vertical section of the eye of a 120-mm larva showing that CB-ir bipolar cells (thick arrow) are restricted tothe central retina. The arrowhead points to the optic nerve. Note faint CB immunoreactivity in the optic nerve. (C) Detail of thecentral retina showing CB-ir bipolar cells (thick arrows) and their axon terminals (arrowheads). (D) Dendrites and Landolt'sclubs (double arrowheads) of CB-ir bipolar cells of a 120-mm larva. (E) Vertical section through the lateral retina of a 120-mmlarva showing scarce CB-ir cells in its inner region. (F) Detail of E showing CB-ir ganglion cells close to the inner limitingmembrane. In panels A, B and D, note that the pigment epithelium is highly pigmented in the dorsal region of the retina,pigment lacking in the ventral region. For abbreviations, see list. Scale bars = 25 μm (C, D, F), 50 μm (A, E), 100 μm (B).
126 B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
GABA (Anadón et al., 1998), dopamine (Yáñez and Anadón,1994) and ChAT (Pombal et al., 2003). The lack of CR- and CB-ir amacrine cells in the retina of larval lampreys is consistentwith the lack of other neurochemical markers of amacrinecells.
In the lateral retina of larvae (only present in larvae morethan 60 mm in body length), only putative ganglion cells wereCR-ir. CR expression in these ganglion cells probably startsshortly after they differentiate and send axons to the optictectum (De Miguel et al., 1990), in line with the early expres-sion of calcium-binding proteins in ganglion cells of othervertebrates (Ellis et al., 1991; Vecino et al., 1993; Doldán et al.,1999; Nag and Wadhwa, 1999). Developmental studies inhuman (Nag and Wadhwa, 1999), chick (Ellis et al., 1991) andteleosts (Vecino et al., 1993; Doldán et al., 1999) retinas haveshown that CR and CB expression is associated with morpho-logical differentiation of neurons, and it is accepted that theirappearance in the retina occurs concomitantly with the es-tablishment of the early laminar organization. Although thelateral retina of large premetamorphic larvae shows somedegree of lamination (inner and outer neuroblastic layers sep-arated by primordial horizontal cells; present results) andamacrine and horizontal cells have been detected by Golgimethods (Rubinson and Cain, 1989), only putative ganglioncells were CB- and/or CR-ir, indicating that the other CB-and/or CR-ir retinal cells observed in adults have not yet dif-ferentiated these neurochemical features.
The immunohistochemical analysis of transforminglampreys indicates that the onset of CR expression in bipolarcells of the lateral retina is delayed until middle stages ofmetamorphosis (M5). In this stage, differentiation of photore-ceptors occurs throughout all the retina (De Miguel and Ana-dón, 1987; De Miguel, 1988), indicating that CR expression inbipolar cells is closely related to the onset of visual function.Instead, expression of CR in ganglion cells of the lateral retinawas observed in the retina of large larvae and transformingstages, preceding the maturation of retinal circuits in thisspecial region of the developing lamprey retina, suggestingthat these neurons are functional. The presence of ganglioncells projecting to the brain, and of a system of GABAergiccentrifugal fibers, has already been reported in the lateralretina of the larva (De Miguel et al., 1989, 1990; Anadón et al.,1998; Meléndez-Ferro et al., 2002), though their functionalsignificance is not clear.
In contrast with that observed in adult sea lamprey, only afew CR-ir ganglion cells were CR-ir in the retina of the adultriver lamprey (Dalil-Thiney et al., 1994). In the sea lamprey,only faint CB staining was seen in some ganglion cells thatsometimes exhibited staining of their nuclei (see Fig. 5F), astaining pattern reminding of that reported for calretinin inrat cerebellum (Arai et al., 1993). CR-ir ganglion cells havebeen reported in the retina of most vertebrates studied, in-cluding chondrosteans (Huesa et al., 2002), teleosts (Vecino etal., 1993; Doldán et al., 1999; Weruaga et al., 2000), amphibians

127B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
(Gabriel et al., 1998), reptiles (Bennis et al., 2005), birds (Ellis etal., 1991; Pasteels et al., 1990) and manymammals (Pasteels etal., 1990; Goebel and Pourcho, 1997; Volgyi et al., 1997; Jeonand Jeon, 1998). The presence of CB-ir ganglion cells has alsobeen reported in the retina of some amphibians (Hamano etal., 1990; Pasteels et al., 1990; Pochet et al., 1991; Gabriel et al.,1998). Since most vertebrates studied, including the sea lam-prey, express CR in ganglion cells, this strongly suggests thatthis is a primitive feature.
CB and/or CR immunoreactivity was found in bipolar cellsin both the larval central retina and the adult retina. While inthe adult there are two very different types of CR/CB-ir bipolarcells, large and small, this distinction was not observed inlarvae. The large and small bipolar cells seem to correspondto the type-1 and type-2 bipolar cells reported in river lamprey(Öhman, 1976), respectively. Large bipolar cells were also CR-and CB-ir in the river lamprey retina (Dalil-Thiney et al., 1994),but stratification of their axons in the IPL is not as clear as inthe sea lamprey. Large bipolar cells of lampreys resemble thedisplaced bipolar cells of the sturgeon (Huesa et al., 2002) andthe type-a1 bipolar cells of the dogfish (Witkovsky and Stell,1973), and all three show stratified endings in the scleral halfof the IPL (the sublamina a of Ramón y Cajal, 1911). However,while the endings are monostratified in sturgeon and dogfish,in sea lamprey, they are distributed in two or three sublayers.In addition, the contacts between stratified terminals of largebipolar cells give them a plexus appearance similar to thatreported in sturgeon (Huesa et al., 2002), which suggests thepresence of electrical coupling similar to that reported in thecarp retina (Cuenca et al., 1993). In other vertebrates, bipolarcells with stratified axons in the scleral IPL are physiologicallyof the “off” type (Famiglietti et al., 1977). On the other hand,the wide dendritic trees and numerous dendritic appendagesof large bipolar cells in the OPL of sea lamprey suggest thatthey contact with many photoreceptors. Such appendageswere not reported in river lamprey (Dalil-Thiney et al., 1994).Morphologically similar bipolar cells of teleosts with endingsin the OFF sublayer contact both cones and rods (Scholes andMorris, 1973; Stell et al., 1977; Sherry and Yazulla, 1993), but,whether the large bipolar cells of lampreys contact one or thetwo types of photoreceptor remains to be investigated.
Small CR- and CB-ir bipolar cells of sea lamprey showeddiffuse axon terminals in the inner half of the IPL. Althoughsmall CR-ir bipolar cells have been observed in the river lam-prey L. fluviatilis (Dalil-Thiney et al., 1994), in the sea lamprey,the small bipolar cells were immunoreactive to CR and CB,while, in the river lamprey, small bipolar cells were notreported with CB immunocytochemistry (Dalil-Thiney et al.,1994). Whether these interspecific differences are real or sim-ply reflect better staining of these cells in sea lamprey is notknown. In other vertebrates, bipolar cells with diffuse term-inals in the vitreal part of the IPL are physiologically of the “on”type (Famiglietti et al., 1977). The connections and physiologyof bipolar cells in lamprey are largely unknown, but our resultssuggest a morphological and functional similarity to those ofother vertebrates studied. Stratification of the lamprey IPL inON and OFF sublaminae has also been reported using cholineacetyltransferase immunocytochemistry (Pombal et al., 2003).
Dendritic appendages of CR- and CB-ir bipolar cells of lar-vae contact with photoreceptors, and their axons course to-
ward the ganglion cells. Well-developed synaptic contactsbetween the base of photoreceptors and dendrites of bipolarcells in the OPL, and bipolar terminals with synaptic ribbonscontacting ganglion cells in the IPL, have been reported inprevious studies (Dickson and Collard, 1979; De Miguel,1988). Owing to the considerable differences in cell size andappearance between larval and adult bipolar cells, it was notpossible to assign larval cells to one or other of the two bipolarcell types seen in adults.
In the larval retina, the anti-CR and anti-CB antibodies didnot label horizontal cells, whereas the vitreal row of horizon-tal cells was stained with these antibodies in the adult retina.This suggests that there are important functional differencesbetween adult and larval cells. The fact that CR and CB immu-noreactivities were restricted to the vitreal row of horizontalcells in the adult retina also suggests that CR and CB play animportant role in their functionality. This type of horizontalcell receives inputs from short and long photoreceptors, whilethe cells of the scleral row synapse with long photoreceptors(Teranishi et al., 1982). The most striking difference observedbetween the two species of lamprey studied is the lack of CRand CB immunoreactivity in horizontal cells of the river lam-prey (Dalil-Thiney et al., 1994). The dilutions of antibodiesused here were adjusted appropriately, as noted in that ourphotographs have no background staining, excluding that thisdifference between lamprey species was caused by technicalproblems. Our results in sea lamprey showing expression ofboth proteins in horizontal cells contradict the hypothesis ofthese authors that absence of these proteins in horizontalcells is the “primitive vertebrate state” (Dalil-Thiney et al.,1994). Moreover, the finding of CB and CR expression in sealamprey HCs suggests that their absence in river lampreyhorizontal cells is a derived characteristic.
CR and/or CB immunoreactivities have been reported inhorizontal cells of some teleosts (Doldán et al., 1999; Weruagaet al., 2000), urodeles (Pasteels et al., 1990; Deng et al., 2001),chameleon (Bennis et al., 2005), chick (Pasteels et al., 1990) andseveral species of mammals (Pasteels et al., 1990; Goebel andPourcho, 1997; Volgyi et al., 1997; Jeon and Jeon, 1998; Wässleet al., 1998; Jeon et al., 2001; Cuenca et al., 2002; Bytyqi andLayer, 2005; Loeliger and Rees, 2005), but not in trout (Vecinoet al., 1993; Weruaga et al., 2000) and other mammals (Jeonand Jeon, 1998; Nag and Wadhwa, 1999; Hwang et al., 2005).Transient expression of calretinin has been reported in hori-zontal cells of the human retina (Nag andWadhwa, 1999). Thesignificance of these between-species variations is unclear,although it has been attributed to different mechanisms forCa2+ buffering. No expression of CR or CB was found in photo-receptors of the river lamprey (Dalil-Thiney et al., 1994) or thesea lamprey (present results).
In the adult lamprey retina, most amacrine and ganglioncells are located in the INL, but some of them are located tothe vitreal part of the IPL (orthotopic ganglion cells, displacedamacrine cells). Accordingly, topographic criteria are not use-ful for distinguishing between these two types of cell. In thepresent study, the moderately CR-ir cells located near theinner limiting membrane or in the IPLi were considered asprobable ganglion cells because fascicles of optic fibers alsoshowed moderate CR immunoreactivity. Since no stronglylabeled optic fibers were observed in optic fascicles, we

128 B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
considered the strongly CR-ir cells observed in the IPL to beprobable displaced amacrine cells. This subset of amacrinecells does not appear to correspond to the cholinergic ama-crine cells whose processes form two asymmetrical inner andouter subplexuses in the IPL (Pombal et al., 2003). The pres-ence of CB- and CR-ir amacrine/ganglion cells has also beenreported in the INL of the river lamprey (Dalil-Thiney et al.,1994), although the immunoreactive cells could not be un-equivocally assigned to these cell types. In sturgeon, CR-ircells are present in the IPL, inner INL and ganglion cell layer,though all have been identified as ganglion cells (Huesa et al.,2002).
In conclusion, comparison of CR/CB immunoreactivitiesbetween adult and larval sea lamprey retinas reveals impor-tant differences that appear to be related to the deep func-tional differences between larval and adult eyes. In adultlampreys, the retina forms part of a typical camera-type eye(as in practically all other vertebrates). However, the larval eyelacks a differentiated lens, and the retina has only simpleneuronal circuitry, without functional amacrine or horizontalcells. It has been suggested that the larval eye is simply anocellus-like light detector without image-forming capability(Meléndez-Ferro et al., 2002).
4. Experimental procedures
4.1. Animals
Larvae (30–135 mm in length; n = 7), transforming stages (stages 1,3 and 5 and young postmetamorphic, determined according toYouson and Potter, 1979; n = 6) and upstream migrating adults(65–75 cm in length; n = 7) of sea lamprey (Petromyzon marinus L.)were used. Larvae were caught in the River Ulla (northwesternSpain), and adults were purchased from a local supplier. Allexperiments conformed to European Community guidelines onanimal experimentation.
4.2. Western blotting
We were unable to obtain sufficient protein from the lampreyretina for Western blot assays so we used protein extracts fromwhole adult brains. Adult sea lampreys were killed with an over-dose of benzocaine (Sigma, St. Louis, MO), and their brains wereremoved and homogenized at 4 °C in 6 volumes of modified RIPAbuffer (50 mM Tris–HCl, 1 mM EDTA, 150 mM NaCl, 1 mM phenyl-methylsulfonylfluoride, 1% Triton X-100, 0.1% sodium dodecylsul-fate, 5 μg/ml aprotinin, pH 7.4). The homogenate was centrifugedat 20,000 × g at 4 °C. The proteins present in the supernatants wereprecipitated with 6 volumes of 100% methanol followed by 100 × gcentrifugation at 4 °C.
Samples of brain extract resuspended in modified RIPA buffer(minimum 25 μg of total protein) were loaded onto 17% acrylam-ide gels, resolved by SDS-PAGE and then electroblotted in a Mini-Protean 3 cell (Biorad, Hercules, CA) onto 0.2 μm polyvinylidenedifluoride membranes (Biorad). After transfer, nonspecific bindingsites on the membrane were blocked by incubating in 5% pow-dered non-fat milk dissolved in Tris-buffered saline containing0.05% Tween 20 (TBST) for 1 h. After blocking, the membraneswere rinsed in TBST and incubated overnight with rabbit poly-clonal anti-CR (Swant, Bellinzona, Switzerland; code 7699/4) oranti-CB antibodies (Swant; code CB 38) diluted 1:1000 and 1:100in TBST, respectively. The membranes were then rinsed in TBST,incubated with a goat anti-rabbit HRP-conjugated antibody(Biorad, diluted 1:15000), rinsed again in TBST, and incubated
with enhanced chemiluminescent reagent (Biorad Immun-StarHRP kit).
As a control, an adult Sprague–Dawley rat was killed by chlo-roform overdose, its brain removed, and protein extracts obtainedas above. The rat brain extract was processed in parallel with thelamprey extracts in adjacent lines. Precision Plus protein stan-dards (Biorad) were used as molecular weight (MW) markers.
4.3. Immunocytochemistry
Animals were deeply anesthetized with benzocaine and killed bydecapitation. Heads of larvae or eyes of adults were fixed in 4%paraformaldehyde in phosphate buffer (PB), rinsed in phosphate-buffered saline (PBS) and cryoprotected. Samples were then em-bedded in Tissue Tek (Sakura, Torrance, CA) and cut on a cryostat.Sections were processed for immunocytochemistry as follows.Nonspecific binding sites were blocked by incubating with 10%normal goat serum (NGS) (Vector Laboratories, Burlingame, CA).Sections were then incubated overnight in a humid chamber atroom temperature with the anti-CR or anti-CB polyclonal antibo-dies (Swant; diluted 1:3000 and 1:1000 respectively). CR immuno-labeling was visualized by the peroxidase–antiperoxidase (PAP)procedure using a goat anti-rabbit antibody (Dako, Glostrup, Den-mark) and the PAP complex (Sigma), while, for CB immunocyto-chemistry, sections were incubated with a biotinylated goat anti-rabbit antibody (Vector Laboratories, Burlingame, CA) and thenthe Vectastain ABC reagent (Vector). All antibody dilutions werecarried out in PBS containing 0.2% Triton X-100 as detergent and3% NGS. The immunocomplexes were developed by incubationwith 0.6 mg/ml 3-3′-diaminobenzidine (DAB, Sigma) and 0.003%H2O2. The specificity of both antibodies has been verified by thesupplier and tested by Western blotting (see above). As a furthercontrol, sections were processed without the primary antibody orthe primary antibody was replaced by the corresponding preim-mune serum; no immunostaining was observed in these sections.
In addition, the monoclonal antibodies anti-CR (Swant; code6B3), anti-CB (Swant; mab 300) and anti-CB (Sigma; code C9848)were tested, but no immunostaining was observed.
For topographic purposes, hematoxylin eosin-stained sectionsof adult and larval retinas from our collection were also used.
4.4. Microscopy and image processing
Photographs were taken in an Olympus AX-70 photomicroscopeequipped with an Olympus DP-12 color digital camera. The photo-graphs were converted to gray scale and adjusted for brightnessand contrast using Corel Photo-Paint 12 software (Corel, Ottawa,Canada). Photomontage and lettering were done using Corel Draw12.
Acknowledgments
This work was supported by grants from the Spanish Ministryof Science and Technology (BM2000-0283) and Xunta de Gali-cia (PGIDT01PXI20006PN; PGIDT04PXIB020003PR). B.V.-C. re-ceived a predoctoral fellowship from the “Xunta de Galicia”.
R E F E R E N C E S
Anadón, R., Meléndez-Ferro, M., Pérez-Costas, E., Pombal, M.A.,Rodicio, M.C., 1998. Centrifugal fibers are the only GABAergicstructures of the retina of the larval sea lamprey: animmunocytochemical study. Brain Res. 782, 297–302.
Andressen, C.I., Blümcke, I., Celio, M.R., 1993. Calcium binding

129B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
proteins: selective markers of nerve cells. Cell Tissue Res.271, 181–208.
Arai, R., Winsky, L., Arai, M., Jacobowitz, D.M., 1991.Immunohistochemical localization of calretinin in the rathindbrain. J. Comp. Neurol. 310, 21–44.
Arai, R., Jacobowitz, D.M., Deura, S., 1993. Ultrastructurallocalization of calretinin immunoreactivity in lobule V of therat cerebellum. Brain Res. 613, 300–304.
Bennis, M., Versaux-Botteri, C., Repérant, J., Armengol, J.A., 2005.Calbindin, calretinin and parvalbumin immunoreactivity inthe retina of the chameleon (Chamaeleo chamaeleon).Brain Behav. Evol. 65, 177–187.
Bytyqi, A.H., Layer, P.G., 2005. Lamina formation in the Mongoliangerbil retina (Meriones unguiculatus). Anat. Embryol. 209,217–225.
Castro, A., Becerra, M., Manso, M.J., Anadón, R., 2003. Distributionand development of calretinin-like immunoreactivity in thetelencephalon of the brown trout, Salmo trutta fario. J. Comp.Neurol. 467, 254–269.
Cuenca, N., Fernández, E., García, M., De Juan, J., 1993. Dendrites ofrod dominant ON-bipolar cells are coupled by gap junctions incarp retina. Neurosci. Lett. 162, 34–38.
Cuenca, N., Deng, P., Linberg, K.A., Lewis, G.P., Fisher, S.K., Kolb, H.,2002. The neurons of the ground squirrel retina as revealed byimmunostains for calcium binding proteins andneurotransmitters. J. Neurocytol. 31, 649–666.
Dalil, N., Repérant, J., Kenigfest, N., Vesselkin, N., Versaux-Botteri,C., Rio, J.P., 1990. Typology and distribution of ganglion cells inthe retina of lamprey (Lampetra fluviatilis). C. R. Acad. Sci., III 11,403–410.
Dalil-Thiney, N., Pochet, R., Versaux-Botteri, C., Vesselkin, N.,Repérant, J., Nguyen-Legros, J., 1994. Immunohistochemicallocalization of calbindin-D28K and calretinin in the lampreyretina. J. Comp. Neurol. 340, 140–147.
De Miguel, E., 1988. Estudio del sistema visual y oculomotor de lalamprea de mar Petromyzon marinus L. en estado larvario ytransformación de la retina durante la metamorfosis. PhDthesis, Universidad de Santiago de Compostela, Santiago deCompostela.
De Miguel, E., Anadón, R., 1987. The development of retina and theoptic tectum of Petromyzon marinus, L. A light microscopicstudy. J. Hirnforsch. 28, 445–456.
De Miguel, E., Rodicio, M.C., Anadón, R., 1989. Ganglion cells andretinopetal fibers of the larval lamprey retina: an HRPultrastructural study. Neurosci. Lett. 106, 1–6.
De Miguel, E., Rodicio, M.C., Anadón, R., 1990. Organization of thevisual system in larval lampreys: an HRP study. J. Comp.Neurol. 302, 529–542.
Deng, P., Cuenca, N., Doerr, T., Pow, D.V., Miller, R., Kolb, H., 2001.Localization of neurotransmitters and calcium bindingproteins to neurons of salamander and mudpuppy retinas.Vision Res. 41, 1771–1783.
Díaz-Regueira, S., Anadón, R., 2000. Calretinin expression inspecific neuronal systems in the brain of an advanced teleost,the grey mullet (Chelon labrosus). J. Comp. Neurol. 426, 81–105.
Dickson, D.H., Collard, T.R., 1979. Retinal development in thelamprey (Petromyzon marinus L.): premetamorphic ammocoeteeye. Am. J. Anat. 154, 321–336.
Dickson, H.D., Graves, D.A., 1981. The ultrastructure anddevelopment of the eye. In: Potter, I.C., Hardisty, M.W. (Eds.),The Biology of Lampreys, vol. 3. Academic Press, London,pp. 43–94.
Doldán, M.J., Prego, B., De Miguel, E., 1999. Immunochemicallocalization of calretinin in the retina of the turbot (Psettamaxima) during development. J. Comp. Neurol. 406, 425–432.
Ellis, J.H., Richards, D.E., Rogers, J.H., 1991. Calretinin and calbindinin the retina of the developing chick. Cell Tissue Res. 264,197–208.
Famiglietti Jr., E.V., Kaneko, A., Tachibana, M., 1977. Neuronal
architecture of on and off pathways to ganglion cells in carpretina. Science 198, 1267–1269.
Fritzsch, B., Collin, S.O., 1990. Dendritic distribution of twopopulations of ganglion cells and the retinopetal fibers in theretina of the silver lamprey (Ichthyomyzon unicuspis).Vis. Neurosci. 4, 533–545.
Gabriel, R., Volgyi, B., Pollak, E., 1998. Calretinin-immunoreactiveelements in the retina and optic tectum of the frog,Rana esculenta. Brain Res. 782, 53–62.
Gall, D., Roussel, C., Susa, I., D'Angelo, E., Rossi, P., Bearzatto, B.,Galas, M.C., Blum, D., Schurmans, S., Schiffmann, S.N., 2003.Altered neuronal excitability in cerebellar granule cells of micelacking calretinin. J. Neurosci. 23, 9320–9327.
Goebel, D.J., Pourcho, R.G., 1997. Calretinin in the cat retina:colocalizations with other calcium-binding proteins, GABA andglycine. Vis. Neurosci. 14, 311–322.
Hamano, K., Kiyama, H., Emson, P.C., Manabe, R., Nakauchi, M.,Tohyama, M., 1990. Localization of two calcium bindingproteins, calbindin (28 kD) and parvalbumin (12 kD), in thevertebrate retina. J. Comp. Neurol. 302, 417–424.
Huesa, G., Yáñez, J., Anadón, R., 2002. Calbindin and calretininimmunoreactivities in the retina of a chondrostean, Acipenserbaeri. Cell Tissue Res. 309, 355–360.
Hwang, I.K., Yoo, K.Y., Kim, D.S., Jung, J.Y., Shin, M.C., Seo, K., Kim,K.S., Kang, T.C., Won, M.H., 2005. Comparative study oncalretinin immunoreactivity in gerbil and rat retina.Anat. Histol. Embryol. 34, 129–131.
Ishikawa, M., Takao, M., Washiola, H., Tokunaga, F., Watanabe, H.,Tonosaki, A., 1987. Demonstration of rod and conephotoreceptors in the lamprey retina by freeze replication andimmunofluorescence. Cell Tissue Res. 249, 241–246.
Jeon, M.H., Jeon, C.J., 1998. Immunocytochemical localization ofcalretinin containing neurons in retina from rabbit, cat, anddog. Neurosci. Res. 32, 75–84.
Jeon, Y.K., Kim, S.Y., Jeon, C.J., 2001. Morphology of calretinin andtyrosine hydroxylase-immunoreactive neurons in the pigretina. Mol. Cells 11, 250–256.
Loeliger, M., Rees, S., 2005. Immunocytochemical development ofthe guinea pig retina. Exp. Eye Res. 80, 9–21.
Meléndez-Ferro, M., Villar-Cheda, B., Abalo, X.M., Pérez-Costas, E.,Rodríguez-Muñoz, R., Degrip, W.J., Yáñez, J., Rodicio, M.C.,Anadón, R., 2002. Early development of the retina and pinealcomplex in the sea lamprey: comparativeimmunocytochemical study. J. Comp. Neurol. 442, 250–265.
Nag, T.C., Wadhwa, S., 1999. Developmental expression ofcalretinin immunoreactivity in the human retina and acomparison with two other EF-hand calcium binding proteins.Neuroscience 91, 41–50.
Negishi, K., Teranishi, T., Kuo, C.H., Miki, N., 1987. Two types oflamprey retina photoreceptors immunoreactive to rod- orcone-specific antibodies. Vision Res. 27, 1237–1241.
Öhman, P., 1976. Fine structure of photoreceptors and associatedneurons in the retina of Lampetra fluviatilis (Cyclostomi).Vision Res. 16, 659–662.
Parmentier, M., Glysens, M., Rypens, F., Lawson, D.E.M., Pasteels,J.L., Pochet, R., 1987. Calbindin in vertebrate classes:immunohistochemical localization and Western blot analysis.Gen. Comp. Endocrinol. 65, 399–407.
Pasteels, B., Rogers, J., Blachier, F., Pochet, R., 1990. Calbindin andcalretinin localization in retina from different species.Vis. Neurosci. 5, 1–16.
Pochet, R., Pasteels, B., Seto-Ohshima, A., Bastianelli, E., Kitajima,S., Van Eldik, L.J., 1991. Calmodulin and calbindin localizationin retina from six vertebrate species. J. Comp. Neurol. 314,750–762.
Pombal, M.A., Abalo, X.M., Rodicio, M.C., Anadón, R., González, A.,2003. Choline acetyltransferase-immunoreactive neurons inthe retina of adult and developing lampreys. Brain Res. 993,154–163.

130 B R A I N R E S E A R C H 1 0 6 8 ( 2 0 0 6 ) 1 1 8 – 1 3 0
Ramón y Cajal, S., 1911. Histologie du Système Nerveux del'Homme et des Vertebrés, vol. 2. Maloine, Paris.
Rogers, J.H., 1987. Calretinin: a gene for a novel calcium-bindingprotein expressed principally in neurons. J. Cell Biol. 105,1343–1353.
Rubinson, K., Cain, H., 1989. Neural differentiation in the retina ofthe larval sea lamprey (Petromyzon marinus). Vis. Neurosci. 3,241–248.
Scholes, J., Morris, J., 1973. Receptor–bipolar connectivity patternsin fish retina. Nature 241, 52–54.
Schwaller, B., Meyer, M., Schiffmann, S., 2002. New functions forold proteins: the role of the calcium-binding proteins calbindinD-28k, calretinin and parvalbumin, in cerebellar physiology.Studies with knockout mice. Cerebellum 1, 241–258.
Sherry, D.M., Yazulla, S., 1993. Goldfish bipolar cells and axonterminal patterns: a Golgi study. J. Comp. Neurol. 329,188–200.
Stell, W.K., Ishida, A.T., Lightfoot, D.O., 1977. Structural basis foron- and off-center responses in retinal bipolar cells. Science198, 1269–1271.
Teranishi, T., Negishi, K., Kato, S., 1982. Two types of light-inducedresponse recorded from horizontal cells in the river lampreyretina. Neurosci. Lett. 33, 41–46.
Vecino, E., García-Briñón, J., Velasco, A., Caminos, E., Lara, J., 1993.Calbindin D-28K distribution in the retina of the developingtrout (Salmo fario L.). Neurosci. Lett. 152, 91–95.
Volgyi, B., Pollak, E., Buzas, P., Gabriel, R., 1997. Calretinin inneurochemically well-defined cell populations of rabbit retina.Brain Res. 763, 79–86.
Wässle, H., Peichl, L., Airaksinen, M.S., Meyer, M., 1998. Calcium-binding proteins in the retina of a calbindin-null mutantmouse. Cell Tissue Res. 292, 211–218.
Weruaga, E., Velasco, A., Briñón, J.G., Arévalo, R., Aijón, J., Alonso,J.R., 2000. Distribution of the calcium-binding proteinsparvalbumin, calbindin D-28k and calretinin in the retina oftwo teleosts. J. Chem. Neuroanat. 19, 1–15.
Witkovsky, P., Stell, W.K., 1973. Retinal structure in the smoothdogfish, Mustelus canis: light microscopy of bipolar cells.J. Comp. Neurol. 148, 47–59.
Yáñez, J., Anadón, R., 1994. Are the dopaminergic cells of thelamprey retina interplexiform cells? A dopamine, tyrosinehydroxylase and dopamine beta-hydroxylaseimmunocytochemical study. Neurosci. Lett. 165, 63–66.
Youson, J.H., Potter, I.C., 1979. A description of the stages in themetamorphosis of the anadromous sea lamprey, Petromyzonmarinus L. Can. J. Zool. 57, 1808–1817.
Related Documents











