This article was downloaded by: [Northeastern University] On: 23 November 2014, At: 10:52 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Bioremediation Journal Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/bbrm20 Cadmium-Resistance Plasmid Affected Cd +2 Uptake More Than Cd +2 Adsorption in Klebsiella oxytoca Khaled M. Khleifat a , Khaled Nawayseh a , Nawel R. Adjeroud a , Ali M. Khlaifat b , Isam H. Aljundi c & Khaled A. Tarawneh a a Biology Department , Mutah University , Mutah, Karak, Jordan b Department of Community Health, Faculty of Nursing , Mutah University , Mutah, Karak, Jordan c Chemical Engineering Department , Mutah University , Mutah, Karak, Jordan Published online: 25 Nov 2009. To cite this article: Khaled M. Khleifat , Khaled Nawayseh , Nawel R. Adjeroud , Ali M. Khlaifat , Isam H. Aljundi & Khaled A. Tarawneh (2009) Cadmium-Resistance Plasmid Affected Cd +2 Uptake More Than Cd +2 Adsorption in Klebsiella oxytoca , Bioremediation Journal, 13:4, 159-170 To link to this article: http://dx.doi.org/10.1080/10889860903080739 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by: [Northeastern University]On: 23 November 2014, At: 10:52Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Bioremediation JournalPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/bbrm20
Cadmium-Resistance Plasmid Affected Cd+2 UptakeMore Than Cd+2 Adsorption in Klebsiella oxytocaKhaled M. Khleifat a , Khaled Nawayseh a , Nawel R. Adjeroud a , Ali M. Khlaifat b , Isam H.Aljundi c & Khaled A. Tarawneh aa Biology Department , Mutah University , Mutah, Karak, Jordanb Department of Community Health, Faculty of Nursing , Mutah University , Mutah, Karak,Jordanc Chemical Engineering Department , Mutah University , Mutah, Karak, JordanPublished online: 25 Nov 2009.
To cite this article: Khaled M. Khleifat , Khaled Nawayseh , Nawel R. Adjeroud , Ali M. Khlaifat , Isam H. Aljundi & KhaledA. Tarawneh (2009) Cadmium-Resistance Plasmid Affected Cd+2 Uptake More Than Cd+2 Adsorption in Klebsiella oxytoca ,Bioremediation Journal, 13:4, 159-170
To link to this article: http://dx.doi.org/10.1080/10889860903080739
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of theContent. Any opinions and views expressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon andshould be independently verified with primary sources of information. Taylor and Francis shall not be liable forany losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use ofthe Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Bioremediation Journal, 13(4):159–170, 2009Copyright ©c 2009 Taylor and Francis Group, LLCISSN: 1088-9868DOI: 10.1080/10889860903080739
Cadmium-Resistance Plasmid AffectedCd+2 Uptake More Than Cd+2 Adsorption
in Klebsiella oxytoca
Khaled M. Khleifat,1
Khaled Nawayseh,1
Nawel R. Adjeroud,1
Ali M. Khlaifat,2
Isam H. Aljundi,3
and Khaled A. Tarawneh1
1Biology Department,2Department of CommunityHealth, Faculty of Nursing, and3Chemical EngineeringDepartment, Mutah University,Mutah, Karak, Jordan
ABSTRACT A bacterial strain was isolated from Petra City Wastewater Treat-ment Plant. This isolate was identified as Klebsiella oxytoca based on 16S rDNAanalysis. A single plasmid (>23 kb) was detected in this strain and transformedinto Esherichia coli JM83. The transformed E. coli cells exhibited elevated re-sistance to cadmium as compared to parental plasmid-free cells. The sodiumdodecyl sulfate (SDS)-treated cells showed higher efficiency in plasmid curingthan the ethidium bromide–treated cells. The ethidium bromide–cured cellsgrew only in a 10 µg/ml Cd+2 minimal tolerable concentration, whereas theSDS-treated cells had no growth in any of the Cd concentrations tested (2,5, 10, 20, 30, 40, and 50 ppm). Contrary to the Freundlich model, the Lang-muir model gave a good fit to the Cd biosorption data by K. oxytoca cells.Plasmid curing caused 80%, 82%, and 70% inhibition in the Cd biosorption,adsorption, and uptake, respectively. Furthermore, the absence of lysine decar-boxylase (LDC) activity in the cured strain strongly implies that the structuralgene-encoding LDC in this bacterium is plasmid encoded. After curing of theplasmid, 100% of the antibiotic-resistant loci were observed as chromosomalencoded. All of the results shown above indicated that the Cd resistance isplasmid mediated.
KEYWORDS adsorption, Cd+2 biosorption, Klebsiella oxytoca, plasmid curing, uptake
INTRODUCTIONMuch research has been devoted to microbial detoxification of heavy met-
als. In most cases, these detoxification mechanisms are plasmid encoded.Among the other mechanisms mentioned are decreased membrane perme-ability (Pan-Hou et al., 1981; Aiking et al., 1982, 1984), uptake, and intracel-lular or extracellular precipitation. As a response to the heavy metal challenge,either metal-induced adaptive cell protection evolves, which requires newlysynthesized proteins (Banjerdkji et al., 2003), or multiple-metal ion-resistantbacteria evolve, which contain a variety of plasmid-encoded metal-resistant de-terminants, e.g., Staphylococcus aureus (Novick and Roth, 1968) and Alcaligeneseutrophus [Ralstonia metallidurans] strain CH34 (Mergeay et al., 1985). Bacterial
Address correspondence to Khaled M.Khleifat, Department of Biology,Mutah University, Mutah, P. O. Box (7),Karak, 61710, Jordan. E-mail:alkh [email protected]
159
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

biomass has been known to readily adsorb or accumu-late metal ions (Romero et al., 1999).
This ability of metal bioaccumulation by some gram-negative bacterial species such as Escherichia coli (Cohenet al., 1991), and Pseudomonas aeruginosa (Hassen et al.,1998) was established in the production of intracellu-lar cadmium-binding proteins. Furthermore, anothergram-negative rod, Alcaligenes eutrophus [Ralstonia met-allidurans] strain CH34, is known to be biosorbent ofheavy metals metals (Diels and Mergeay, 1990; Nies,1995). Among heavy metals that are toxic, with longresidence times and biological half-lives, cadmium inparticular constitutes a major problem in industrializednations (Nas et al., 2005), because its presence in the en-vironment significantly endangers public health (Diels,1997).
In gram-negative bacteria such as Esherichia coli,the active uptake of cadmium is coupled with theZn+2 transport system (Hao et al., 1999). Once insidethe cell, the Cd+2 may become toxic either throughsingle-stranded breakage of cellular DNA (Mitra andBernstein, 1978), or by interfering with the respirationand growth of the bacteria (Holmes et al., 1997). Themechanism of Cd resistance in gram-negative bacte-ria is mainly based on the action of an efflux systemcalled RND that has diverse functions, such as the resis-tance/nodulation of legumes/cell division (Saier et al.,1994).
This active transporter is a proton-driven export sys-tem (Rensing et al., 1997). Because comparable dataare lacking for other E. coli species, it seemed pertinentto extend these investigations to Klebsiella oxytoca usingcadmium. Therefore, this paper reports our studies onCd+2 removal (detoxification) by K. oxytoca isolatedfrom Petra Wastewater Treatment Plant. Petra city isone of the world’s most famous archaeological sites, lo-cated in southern Jordan. This investigation was carriedout to determine whether the location of the cadmiumresistance is chromosomal or plasmid mediated. Thecorrelation between Cd+2 biosorption, uptake, and ad-sorption, and Cd-mediated plasmid was studied. Mod-eling the cadmium biosorption data was also carriedout.
MATERIAL AND METHODSMedia and Culture Conditions
The bacterium used in these experiments was Kleb-siella oxytoca, which was already isolated from the
wastewater treatment plant in Petra, Jordan (Adjeroud,2009). Its biochemical identity was reverified using theREMEL kit (RapID ONE and RapID NF Plus systems)procedure as well as the morphological characteris-tics, which were always microscopically checked. Itsidentification was also confirmed by 16S rDNA anal-ysis (Macrogen, Korea). The accession number of nu-cleotide sequence in GenBank is file nuc 1.sqn nuc 1GQ144701. An E. coli JM83 was used as control forkit identification tests. Nutrient broth (NB) (1.0 g meatextract, 2.0 g yeast extract, 5.0 g peptone, and 5.0 gsodium chloride per liter of solution) was used in allexperiments unless otherwise stated.
Plasmid Isolation and TransformationCrude lysates of plasmid DNA were prepared as de-
scribed by Holmes (1984). Agarose gel electrophoresiswas prepared using 0.7% (w/v) agarose in a 89 mMTris-borate buffer (pH 8.5), plus 2.5 mM disodiumEDTA (horizontal slab gel). The size of plasmid wasdetermined using the λ-HindIII cut as a marker. Theisolated plasmid was transformed into E. coli JM83 asdescribed by Sambrook et al. (1989). The transformantwas selected on selective medium plates containing500 µg/ml Cd (Haq et al., 1999).
Plasmid CuringTwo chemical compounds were used for plasmid
curing: sodium dodecyl sulfate (SDS) (El-Mansi et al.,2000) and ethidium bromide (Haq et al., 1999). In theSDS procedure, a 10% w/v (pH 7.4) solution of SDSwas added to nutrient broth (double strength) and thefinal volume was made up to 10 ml in test tubes withdistilled water to give 8%, 7%, 6%, 5%, 4%, 3%, 2%,1%, and 0.5% in the NB (normal strength). Then thedifferent percentage of SDS-containing NB media wereinoculated with adjusted starting inoculum (≈1.75×108 cell/ml) and incubated overnight at 150 rpm shak-ing rate at 30◦C. After incubation, 1 ml of each SDS-containing NB medium was harvested to remove theSDS, washed with 50 mM phosphate buffer (pH 7.4),and resuspended in the same buffer to give an OD420
of 1.0. In the case of ethidium bromide, different con-centrations of ethidium bromide (50 to 300 µg/ml)were used. After filtration and sterilization, ethidiumbromide was added to the autoclaved medium priorto inoculating with bacterial isolates. The cultures wereincubated for 48 h at shaking rate of 200 rpm. For
K. M. Khleifat et al. 160
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

both SDS and ethidium bromide, samples were dilutedwith saline (0.85% NaCl) and spread on plates to pro-duce isolated colonies. The colonies were tooth-pickedand transferred simultaneously onto grids made on NBagar medium plates with and without 70 µg/ml Cd.The colonies shown on NB agar plates without Cd+2
and not on plates containing Cd were counted as theones with their plasmid cured and were further analyzedfor plasmid loss. To assure the curing of plasmid(s),the control (untreated), cured, and transformed bac-terial cells were checked for their antibiotic-resistanceprofile. The antibiotics were used as sensi discs(i.e., amoxycillin, ampicillin, aztreonam, ceftazidime,cephalexin, cephalothin, cephradine, ciprofloxacin,imipenem, nalidixic acid, nitrofurantoin, norfloxacin,ofloxacin, penicillin G, streptomycin, sulfamethoxa-zole, teicoplanin, and tetracycline). To ensure the con-sistency of control and cured bacterial cells for Cd re-sistance, a 70 µg/ml Cd solution containing bacterialcultures was used. Thus any change against either an-tibiotics or Cd was an indication of curing and trans-formation (Haq et al., 1999).
MIC DeterminationMinimal Inhibitory Concentration (MIC) was deter-
mined in duplicate in a NB with 10, 20, 30, 40, 50,60, 70, 80, 90, and 100 ppm Cd+2. The Cd+2 concen-trations were normalized using the cadmium chemicalcompound Cd(NO3)2·4H2O as Cd+2 source. Cultureswere incubated at 37◦C for at least 72 h, checking bacte-rial development by absorbance at 600 nm every 24 h.MIC was estimated as the first dilution that completelyinhibits bacterial growth in the nutrient broth medium.
Cd RemovalThe Cd+2 removal by bacterial cultures was deter-
mined using different concentrations of Cd+2 in nu-trient broth (Wilson and Walker, 1994). Each time,the cultures were spun at 14000 rpm to remove bac-terial cells and the supernatant was used to determineCd+2 concentrations by using a Varian AA-280 ZeemanAtomic Absorption Spectrophotometer, coupled witha GTA-120 (Varian Techtron, Australia). Reduction inthe amount of Cd+2 in the medium after growth ofbacteria was taken as the Cd+2 removal ability of theisolate.
Measurement of CadmiumBiosorption, Adsorption, and Uptake
The biosorption of Cd+2 (uptake and adsorption)was carried out according to the method of Scott (1990)and Kanazawa and Mori (1996) with slight modifica-tions. Cultures of bacterial strains were grown in NBmedium at 37◦C using a shaking incubator with anagitation rate of 150 rpm. Cells were harvested by cen-trifugation at 8000 rpm for 20 min. The supernatantwas then discarded and the cells washed twice with0.85% saline. Harvested biomass (50 mg) was resus-pended in 12 ml solutions containing of 5, 10, 20, 30,40, 50, 60 and 70 µg/ml Cd+2 for 1 h. These solu-tions were filtered with a Millipore filter (0.45 µm poresize) to separate the biomass from the filtrate. Subse-quently, the biomass was washed either with distilledwater or with 0.01 N CH3COONH4 solutions at pH4.0 according to the method of Kanazawa and Mori(1996). After washing, the biomass was reweighed inthe dry state and decomposed by 1% nitric acid for24 h. The Cd+2 content for the latest solution was de-termined by using a Varian AA-280 Zeeman AtomicAbsorption Spectrophotometer, coupled with a GTA-120 (Varian Techtron). The water-washed biomass wasused for measuring the Cd+2 biosorption, whereas theCH3COONH4-washed biomass was used for the Cd+2
uptake. The Cd+2 adsorption was then determinedby subtracting the amount of Cd+2 uptake from theamount of Cd+2 biosorption. All the results of Cd+2
concentrations were normalized as ppm.
Determining the Toxicity Effect of Cdon Bacterial Growth
The bacterial growth medium was composed of NBsupplemented with 0, 5, 10, 20, 30, 40, 50 and 60 ppmCd+2. The cells were incubated in a GFL Model 3032shaker (Burgwedel, Germany) at 37◦C using normal ag-itation rate of 200 rpm. Samples were taken at intervalsto determine the OD at 600 nm. The growth curvesbased on OD600 measurements were plotted againsttime (h).
ModelingA commonly used model for describing heavy metal
biosorption is the Langmuir model. It suggests that ad-sorption occurs on a homogeneous surface by mono-layer sorption without interaction between adsorbed
161 Cd+2 Biosorption, Uptake, and Adsorption by Klebsiella oxytoca
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014
molecules (Langmuir, 1918). This model can be writ-ten as:
qe = qm KLCe
1 + KLCe(1)
where qe is the equilibrium cadmium ion concentrationon the biomass (mg/g), Ce is the equilibrium cadmiumion concentration in the solution (mg/l), qm is themonolayer biosorption capacity of the biomass (mg/g),and KL is the Langmuir biosorption constant (L/mg)relating the free energy of biosorption. Another widelyused isotherm to describe nonlinear biosorption is theFreundlich model. This model proposes a monolayersorption with a heterogeneous energetic distribution ofactive sites, accompanied by interactions between ad-sorbed molecules. The Freundlich model can be writtenas (Febrianto et al., 2009):
qe = K f C1/ne (2)
where K f is a constant relating the biosorption capacityand 1/n is an empirical parameter relating the biosorp-tion intensity, which varies with the heterogeneity ofthe biomass.
RESULTS AND DISCUSSIONSMIC of Cd for Different Bacteria
From monitoring the Cd+2 resistance by differentgram-negative bacteria isolated from the Petra wastewa-ter treatment plant, Klebsiella oxytoca isolate showed themost efficient resistance to cadmium, based on the hy-pergrowth observed, indicating that removal of Cd+2
was dependent on bacterial species (Adjeroud, 2009).In K. aerogenes, previous reports (Aiking et al., 1982,1984) showed that the actual cellular contents of cad-mium, phosphate, and sulfide depended very much onthe particular strain and on the growth conditions. Theclassification of strains as either resistant or sensitive isnot as clear-cut as it seems because such classificationis easily confused by different kinds of physiological re-sponses, especially in batch culture. Therefore, a labellike sensitive or resistant is useful only when accompa-nied by a detailed description of the growth conditions.Therefore, K. oxytoca was used for further Cd+2 removalstudies.
0
20
40
60
80
100
120
0 100 200 300
Ethidium bromide concentration (ppm)
Sur
viva
l (%
)
(a)
0
20
40
60
80
100
0 2 4 6 8
SDS concentration (%)
Sur
viva
l (%
)
(b)
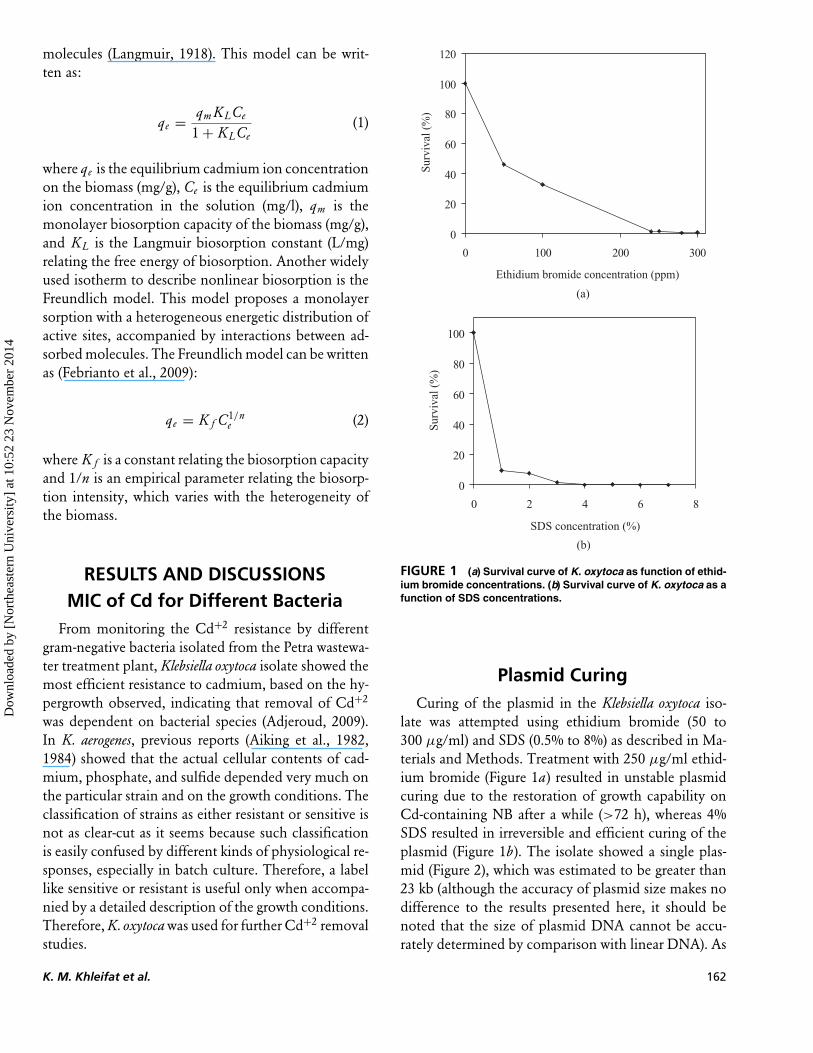
FIGURE 1 (a) Survival curve of K. oxytoca as function of ethid-ium bromide concentrations. (b) Survival curve of K. oxytoca as afunction of SDS concentrations.
Plasmid CuringCuring of the plasmid in the Klebsiella oxytoca iso-
late was attempted using ethidium bromide (50 to300 µg/ml) and SDS (0.5% to 8%) as described in Ma-terials and Methods. Treatment with 250 µg/ml ethid-ium bromide (Figure 1a) resulted in unstable plasmidcuring due to the restoration of growth capability onCd-containing NB after a while (>72 h), whereas 4%SDS resulted in irreversible and efficient curing of theplasmid (Figure 1b). The isolate showed a single plas-mid (Figure 2), which was estimated to be greater than23 kb (although the accuracy of plasmid size makes nodifference to the results presented here, it should benoted that the size of plasmid DNA cannot be accu-rately determined by comparison with linear DNA). As
K. M. Khleifat et al. 162
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

FIGURE 2 Agarose gel electrophoresis pattern of plasmid DNAisolated from wild-type K. oxytoca and its plasmid-cured deriva-tive. Lane 1, λ DNA cleaved with HindIII; Lane 2, K. oxytoca show-ing one plasmid (≈23 kb); lanes 3 and 4 were plasmid-curedderivative of K. oxytoca treated with ethidium bromide and 7%SDS, respectively; lane 5, plasmid shown in lane 2 was purifiedfrom transformed E. coli.
can be seen in Figure 1, the K. oxytoca strain appearedto have completely lost its original plasmid, which isevidence that the purified plasmid matched that ofthe wild-type. Thus the loss of Cd resistance in curedstrain was used as sign of curing (Figure 2, Table 1).Out of 235 colonies transformed each for SDS- andethidium bromide–treated cells, 192 and 117 colonieswere cured, respectively, showing in this manner 81%and 50% curing for the SDS- and ethidium bromide–treated bacterial isolate, respectively. Colony size of thecured cells was smaller than that of uncured cells. Fur-thermore, after repeated subculturing, the SDS-curedcells maintained their characteristics and did not re-vert to their parental status. In order to prove thatthe cured K. oxytoca strain was indeed a derivative ofK. oxytoca, biochemical identity tests were carried outroutinely and the results obtained are summarized inTable 1. All API biochemical tests except LDC (lysinedecarboxylase) showed substantiated evidence as char-acteristics of both the wild-type and the cured strains.It should be pointed out that the negativity of LDCcharacter was obtained in SDS-treated isolates only.By comparison with SDS, ethidium bromide appearedto be less effective as a curing agent. It has been re-ported that natural, large plasmids are stable and diffi-cult to remove from the cells (Joset et al., 1993). Thisis further supported by the observation that irreversiblecuring of the plasmid in K. oxytoca was difficult to
TABLE 1 Biochemical Characteristics for K. oxytoca and itsPlasmid-Cured Derivative (Ethidium Bromide– and SDS-TreatedCells)
EthidiumBiochemical Uncured bromide-treated SDS-treatedtests K. oxytoca cells (250 ppm) cells (4%)
URE − − −ADH − − −ODC − − −LDC + + −TET − − −LIP − − −KSF + + +SBL + + +GUR − − −ONPG − − −βGLU + + +βXYL − − −NAG − − −MAL + + +PRO − − −GGT − − −PYR + + +ADON − − −IND + + +OXI − − −
Note. URE: urea; ADH: arginine; ODC: ornithine; LDC: lysine;TET: aliphatic thiol; LIP: fatty acid ester; KSF: sugar aldehyde; SBL:sorbitol; GUR: p-nitrophenyl-β, D-glucoronide; ONPG: o-nitrophenyl-β,D-galactoside; βGLU: p-nitrophenyl-β,D-glucoside; βXYL: p-nitrophenyl-β,D-xyloside; NAG: p-nitrophenyl-N-acetyl-β,D-glucosaminide; MAL:malonate; PRO: proline-β-naphthylamide; GGT: γ -glutamyl-β-naphthylamide; PYR: pyrrolidonyl-β- naphthylamide; ADON: adonitol;IND: tryptophan; OXI: oxidase test.
achieve using ethidium bromide (Peiris and Dlamini,1999).
Rubin and Rosenblum (1971) used ethidium bro-mide, which causes loss of the known penicillinaseplasmid markers of S. aureus, but subsequent work hasrevealed that complete loss of the penicillinase plasmidis dependent on the strain. Strains in which the plasmidhas been artificially introduced by transduction appearto lose the entire plasmid when subjected to a cur-ing treatment, whereas naturally occurring penicillinaseplasmid–bearing strains appear to retain a large frag-ment, losing only the known resistance markers (Rubinand Rosenblum, 1971).
To determine whether the Cd-resistance characterwas plasmid dependent, the same plasmid isolatedfrom K. oxytoca was transformed into a competentE. coli JM83 strain. The transfer of plasmid carrying
163 Cd+2 Biosorption, Uptake, and Adsorption by Klebsiella oxytoca
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

Cd+2 resistance into the recipient strain was confirmedby carrying out electrophoresis. The gel patterndemonstrated that transformed E. coli harbored a singleplasmid with mobility identical to that of the donorstrain’s plasmid (Figure 2). Prior to transformation, theE. coli strain was inhibited by 400 µg/ml Cd+2, whereasthe Cd+2 minimal tolerable concentration (MTC) forthe transformed E. coli strain was 500 µg/ml. Cd resis-tance in K. oxytoca was found to be plasmid mediatedas confirmed by transformation studies, because resis-tance against metals is frequently coded by the plasmidspresent in bacteria (Ohman, 1988). Considerable at-tention has been given to the positive biotechnologicalrole of heavy metal–resistant plasmids as they aretransformed into other cells (Horitsu et al., 1986;Haq et al., 1999; Pan-Hou et al., 1981; Aiking et al.,1984), but very few studies addressed the question asto whether it might play a negative role in reducing thebasal level of heavy metals biosorption capacity presentin plasmid-free heterologous host cells. The decreasein heavy metals biosorption in transformed bacterialcells could be because of more growth drawbacksthan plasmid-free cells. Such growth limitations are at-tributed to disturbances in cellular metabolism that areimposed by the energy burden on plasmid replication(Cheah et al., 1987; Khosravi et al., 1990). The limita-tion in Cd+2 accumulation caused by the presence ofexternal plasmids that in turn cause growth weaknessin cellular metabolism and possible alterations in Cd-resistance/sensitivity mechanisms might be overcomeby the cell mechanisms’ capacity for elimination ofCd+2 conferred by the receiving of precise Cd-mediatedplasmid.
Antibiotics ResistanceParental and cured bacterial isolates were compared
for their resistance against different antibiotics in or-der to determine the location of antibiotic-resistantgenes (Haq et al., 1999). The comparison before andafter curing is shown in Table 2. Uncured K. oxy-toca cells were sensitive to aztreonam, ceftazidime,cephalexin, cephalothin, cephradine, ciprofloxacin,imipenem, nalidixic acid, nitrofurantoin, norfloxacin,ofloxacin, streptomycin, sulfamethoxazole, and tetracy-cline, intermediate to amoxicillin, and resistant to ampi-cillin, penicillin G, and teicoplanin. The cured cellsshowed almost the same results as the parental cells.Therefore, this is indicative of the chromosomal an-
TABLE 2 Antibiotic Resistance Profile of K. oxytoca and itsPlasmid-Cured Derivative (Ethidium Bromide– and SDS-TreatedCells)
CuredK. oxytoca
(at 250 ppm CuredUncured ethidium K. oxytoca
Antibiotic K. oxytoca bromide) (at 4% SDS)
Amoxicillin I R RAmpicillin R R RAztreonam S S SCeftazidime S S SCephalexin S S SCephalothin S S SCephradine S S SCiprofloxacin S S SImipenem S S SNalidixic acid S S SNitrofurantoin S S SNorfloxacin S S SOfloxacin S S SPenicillin G R R RStreptomycin S S SSulfamethoxazole S S STeicoplanin R R RTetracycline S S S
Note. I, intermediate; R, resistant; S, sensitive.
tibiotic gene–resistant dependence. Recently, K. oxytocahas been found to harbor acquired plasmid-mediatedβ-lactamases significantly less often (10%) than K . pneu-moniae (Hadziyannis et al., 2000).
Cd Removal CapabilityCd removal ability of the bacterial isolates was
checked by estimation of the amount of Cd in themedium after removal of bacterial cells at the end ofthe growth and processing period. The results showedthat bacterial isolate was efficient in removal of Cd(Figure 3a and b). The relationship between initialbroth Cd+2 concentration, cellular Cd+2 concentra-tion (Cd+2 bioaccumulation) and bacterial growth ofK oxytoca was investigated (Figure 3a and b). The cellu-lar Cd+2 concentration of bacterial isolate (Figure 3a)inoculated into nutrient broth containing 30 µg/ml Cdvaried throughout its growth curve (Figure 3b). In thefirst 5 h, the cellular Cd+2 concentration was almost 0.After 10 h of growth, the cellular Cd+2 concentrationwas 45 µg/mg Cd+2 biomass. It roses to a maximum
K. M. Khleifat et al. 164
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014
0
60
120
180
240
300
360
420
0 10 20 30 40 50 60
Incubation time (h)
Cd
conc
entr
atio
n (µ
g/m
g bi
omas
s)
(a)
0
0.4
0.8
1.2
1.6
0 2 4 6 8 9 10 12 24 30 36 50 54
Incubation time (h)
Abs
orba
nce
at 6
00 n
m
0
20
40
60
80
100
Cd
rem
oval
(%
)
Bacterial growth
Cd concentration
(b)
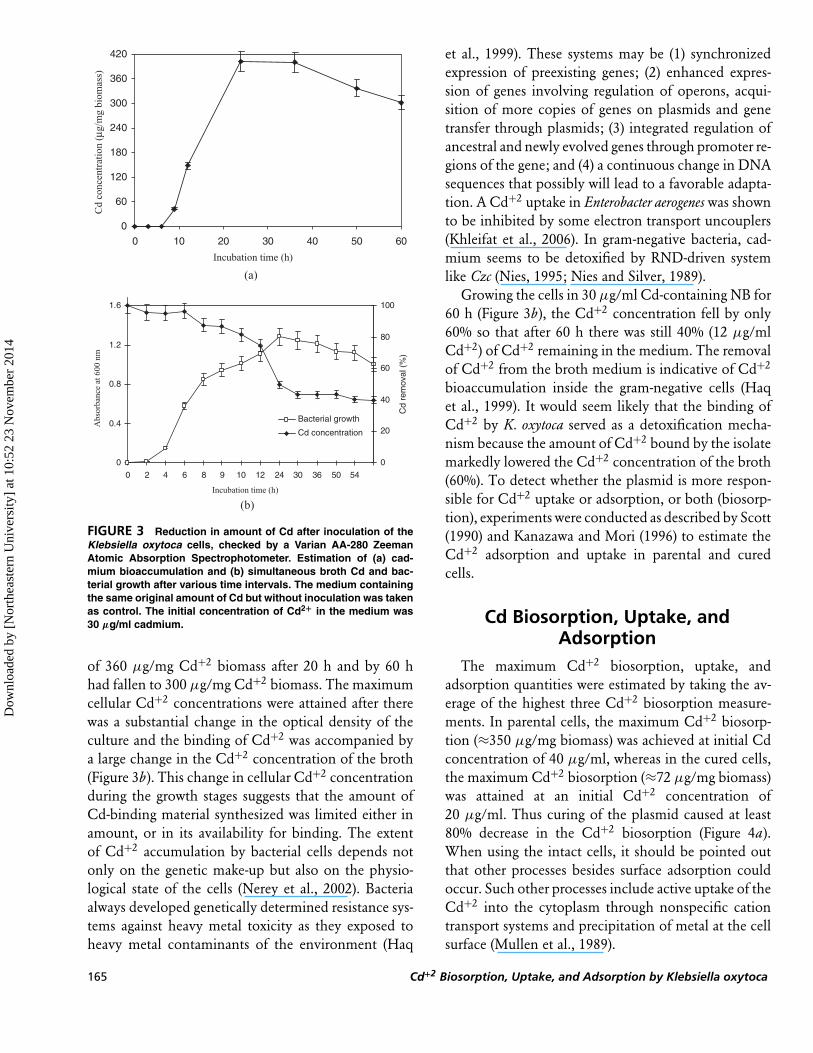
FIGURE 3 Reduction in amount of Cd after inoculation of theKlebsiella oxytoca cells, checked by a Varian AA-280 ZeemanAtomic Absorption Spectrophotometer. Estimation of (a) cad-mium bioaccumulation and (b) simultaneous broth Cd and bac-terial growth after various time intervals. The medium containingthe same original amount of Cd but without inoculation was takenas control. The initial concentration of Cd2+ in the medium was30 µg/ml cadmium.
of 360 µg/mg Cd+2 biomass after 20 h and by 60 hhad fallen to 300 µg/mg Cd+2 biomass. The maximumcellular Cd+2 concentrations were attained after therewas a substantial change in the optical density of theculture and the binding of Cd+2 was accompanied bya large change in the Cd+2 concentration of the broth(Figure 3b). This change in cellular Cd+2 concentrationduring the growth stages suggests that the amount ofCd-binding material synthesized was limited either inamount, or in its availability for binding. The extentof Cd+2 accumulation by bacterial cells depends notonly on the genetic make-up but also on the physio-logical state of the cells (Nerey et al., 2002). Bacteriaalways developed genetically determined resistance sys-tems against heavy metal toxicity as they exposed toheavy metal contaminants of the environment (Haq
et al., 1999). These systems may be (1) synchronizedexpression of preexisting genes; (2) enhanced expres-sion of genes involving regulation of operons, acqui-sition of more copies of genes on plasmids and genetransfer through plasmids; (3) integrated regulation ofancestral and newly evolved genes through promoter re-gions of the gene; and (4) a continuous change in DNAsequences that possibly will lead to a favorable adapta-tion. A Cd+2 uptake in Enterobacter aerogenes was shownto be inhibited by some electron transport uncouplers(Khleifat et al., 2006). In gram-negative bacteria, cad-mium seems to be detoxified by RND-driven systemlike Czc (Nies, 1995; Nies and Silver, 1989).
Growing the cells in 30 µg/ml Cd-containing NB for60 h (Figure 3b), the Cd+2 concentration fell by only60% so that after 60 h there was still 40% (12 µg/mlCd+2) of Cd+2 remaining in the medium. The removalof Cd+2 from the broth medium is indicative of Cd+2
bioaccumulation inside the gram-negative cells (Haqet al., 1999). It would seem likely that the binding ofCd+2 by K. oxytoca served as a detoxification mecha-nism because the amount of Cd+2 bound by the isolatemarkedly lowered the Cd+2 concentration of the broth(60%). To detect whether the plasmid is more respon-sible for Cd+2 uptake or adsorption, or both (biosorp-tion), experiments were conducted as described by Scott(1990) and Kanazawa and Mori (1996) to estimate theCd+2 adsorption and uptake in parental and curedcells.
Cd Biosorption, Uptake, andAdsorption
The maximum Cd+2 biosorption, uptake, andadsorption quantities were estimated by taking the av-erage of the highest three Cd+2 biosorption measure-ments. In parental cells, the maximum Cd+2 biosorp-tion (≈350 µg/mg biomass) was achieved at initial Cdconcentration of 40 µg/ml, whereas in the cured cells,the maximum Cd+2 biosorption (≈72 µg/mg biomass)was attained at an initial Cd+2 concentration of20 µg/ml. Thus curing of the plasmid caused at least80% decrease in the Cd+2 biosorption (Figure 4a).When using the intact cells, it should be pointed outthat other processes besides surface adsorption couldoccur. Such other processes include active uptake of theCd+2 into the cytoplasm through nonspecific cationtransport systems and precipitation of metal at the cellsurface (Mullen et al., 1989).
165 Cd+2 Biosorption, Uptake, and Adsorption by Klebsiella oxytoca
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014
0
50
100
150
200
250
0 10 20 30 40 50 60 70
Cd concentration (µg/mL)
Cd
upta
ke (
µgC
d/m
g bi
omas
s)
Control
Cured
(b)
0
50
100
150
200
0 10 20 30 40 50 60 70Cd concentration (µg/mL)
Cd
adso
rptio
n (µ
gCd/
mg
biom
ass)
Control
Cured
(c)
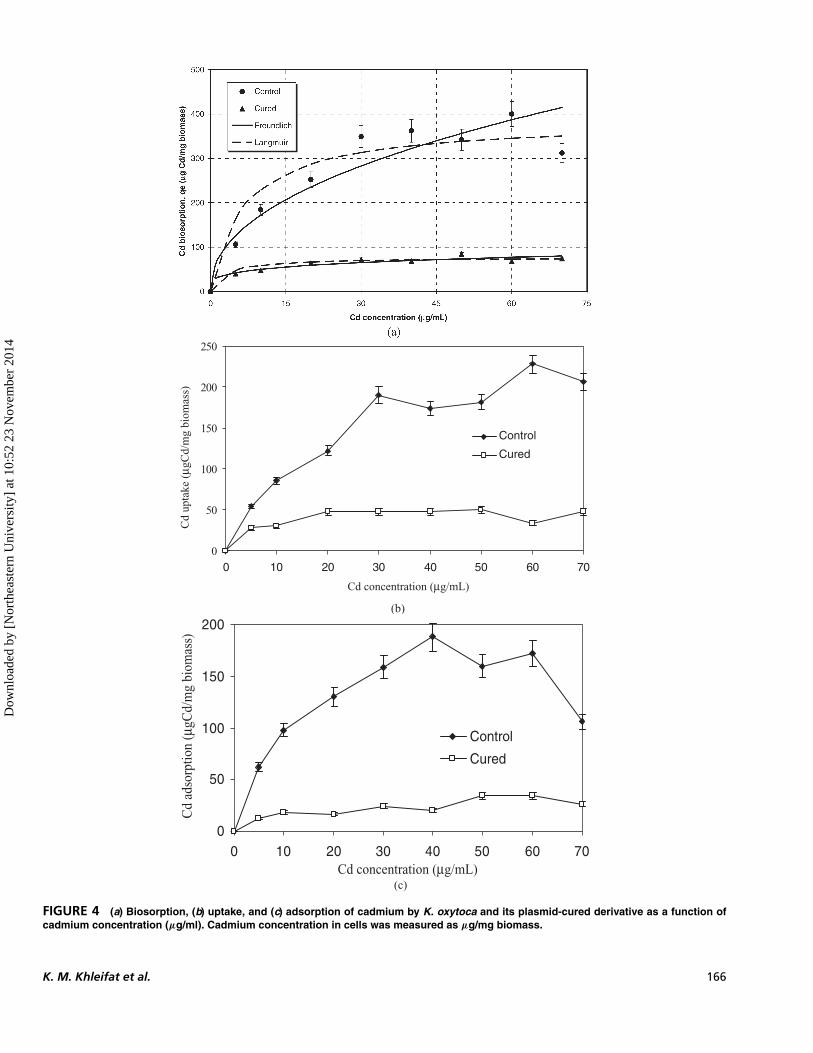
FIGURE 4 (a) Biosorption, (b) uptake, and (c) adsorption of cadmium by K. oxytoca and its plasmid-cured derivative as a function ofcadmium concentration (µg/ml). Cadmium concentration in cells was measured as µg/mg biomass.
K. M. Khleifat et al. 166
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

The maximum Cd+2 uptake achieved was 163 and47 µg/mg biomass for parental and cured-plasmid cells,respectively (Figure 4b). In case of Cd+2 adsorption,the maximum Cd+2 adsorption achieved was 153 and27 µg/mg biomass for parental and cured-plasmidcells (Figure 4c ). Plasmid curing caused 71% and82% inhibition in the Cd+2 uptake and adsorption,respectively.
Comparing the inhibition results of Cd+2 biosorp-tion, uptake, and adsorption in plasmid-cured cells, itis clear that the inhibition in Cd+2 biosorption andadsorption is almost similar (≈80% to 82%); however,the inhibition of Cd+2 uptake in cured cells was ap-proximately 70% as compared with that of parentalcells. Previous studies showed that Cd-sensitive bacte-rial strains lacking plasmids that mediate resistance toCd+2 take up much more Cd+2 from a growth mediumthan do plasmid-positive cells resistant to Cd+2 (Novickand Roth, 1968; Tynecka et al., 1975; Nerey et al.,2002). Bacterial cell surface properties play an impor-tant role in many physical phenomena. Metal biosorp-tion depends upon the available surface negativecharges. Cd2+ biosorption could affect these surfacenegative charges and thus bacterial electric properties.This plasmid seems to be responsible on the numberof exposed negative charges on bacterial surfaces (Vulloet al., 2008). Probably this is why the Cd+2 uptake inthe cured cells had a lower level of Cd+2 inhibition ascompared to that in the Cd+2 adsorption of the samecells. The higher fraction of inhibition in Cd+2 ad-sorption indicated a clearer type-response relationshipbetween the level of Cd+2 adsorption and the plasmidthan for Cd+2 uptake. In other words, the higher theCd+2 resistance, the lower the Cd+2 uptake, and vice-versa. This suggests that the supportive role of basal Cd-biosorption capability is never completely abolished inthe absence of Cd-mediated plasmids. Moreover, cad-mium biosorption (adsorption and uptake) parametersobtained after plasmid curing showed that cadmiumbiosorption mediated by plasmid is in agreement withthe Cd+2 removal capability from the broth duringbacterial growth (Figure 4a and b). No information isknown about Cd2+ uptake and accumulation in K. oxy-toca. However, in E. coli, the accumulation and distribu-tion of Cd2+ was measured by Mitra et al. (1975) andKhazaeli and Mitra (1981). Depending on the growthconditions, most of the accumulated Cd2+ was eitherassociated with the cell envelope (Mitra et al., 1975) or
TABLE 3 Parameters of Langmuir and Freundlich Models
Langmuir constants Freundlich constants
qm (mg/g) K L (L/mg) R2 K f n R2
Control 384.62 0.1461 0.939 60.76 2.212 0.872Cured 76.92 0.3118 0.980 28.36 4.095 0.856
found in the cytoplasmic portions (Khazaeli and Mitra,1981).
The constants of Langmuir model (qm and KL) werecalculated from the slope and intercept of the straightline of plot Ce /qe against Ce , whereas the constants ofFreundlich model (K f and n) were determined fromthe linear plot of log qe versus log Ce . These constantsare shown in Table 3 along with the correlation coeffi-cients. Values of correlation coefficients R2 show thatthe Freundlich model was not able to adequately de-scribe the relationship between the sorbed amount ofcadmium and its equilibrium concentration in the so-lution. However, the Langmuir model fitted best to ourexperimental data. This also can be seen in Figure 4a.The fact that the Langmuir isotherm fits the experi-mental data very well may be due to the homogeneousdistribution of active sites onto biomass surface (Bulutet al., 2007).
Effect of Cd on the Growth CurvesDifferent Cd+2 concentrations were used to assess
the effect on the growth kinetics (Figure 5). It is ob-vious that 5 and 10 µg/ml Cd enhanced the bacterialgrowth in most incubations. However, 20 and 30 µg/mlCd concentrations showed a neutral effect, comparedwith that of Cd-free cultures. And the growth in mediacontaining Cd+2 at a concentrations of 0 to 30 µg/mlCd led to only 2 h of lag period, and the maximaloptical density was not obviously changed. In contrastto these results, the Cd+2 concentrations of more than40 µg/ml Cd markedly inhibited the growth in a Cdconcentration–dependent manner and the cells beganto grow with a 8- to 10-h delay. The prolongation ofthis lag time is probably a question of cell adaptation.Our results were in agreement with those of Doyle et al.(1975), showing that the same Cd+2 concentrations, 5and 10 µg/ml Cd levels, enhanced the growth of E. coliand S. faecalis. The growth stimulation at lower Cd+2
167 Cd+2 Biosorption, Uptake, and Adsorption by Klebsiella oxytoca
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014
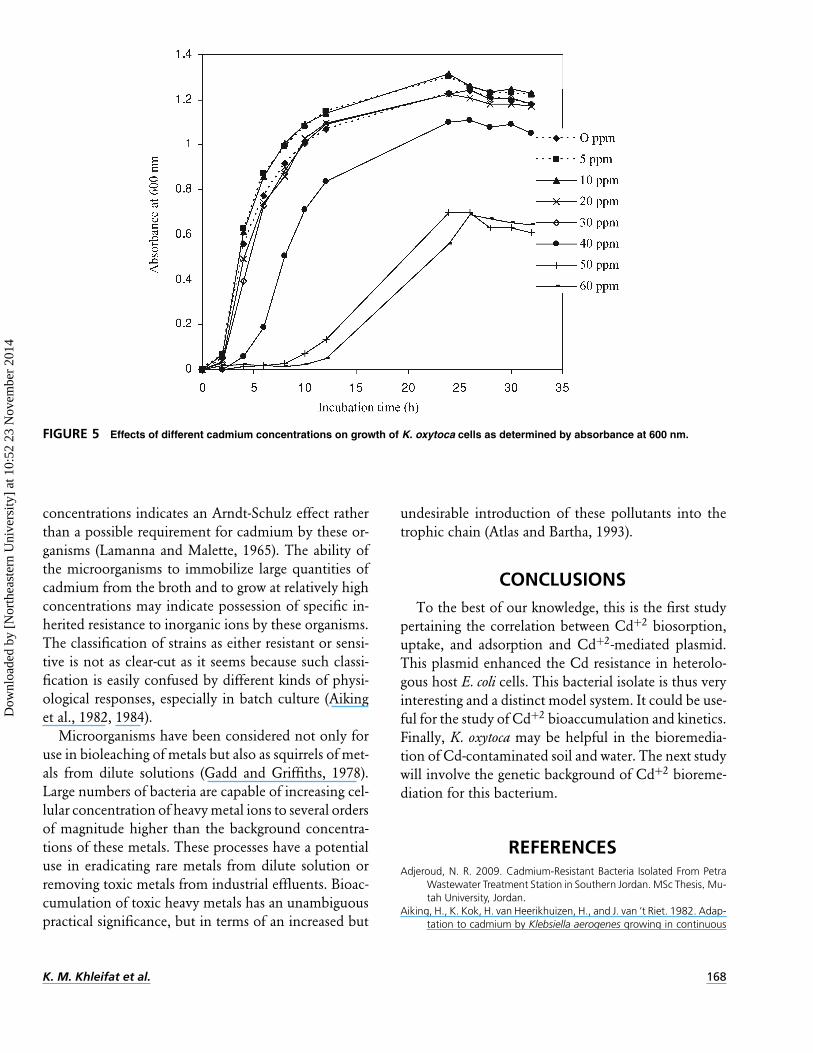
FIGURE 5 Effects of different cadmium concentrations on growth of K. oxytoca cells as determined by absorbance at 600 nm.
concentrations indicates an Arndt-Schulz effect ratherthan a possible requirement for cadmium by these or-ganisms (Lamanna and Malette, 1965). The ability ofthe microorganisms to immobilize large quantities ofcadmium from the broth and to grow at relatively highconcentrations may indicate possession of specific in-herited resistance to inorganic ions by these organisms.The classification of strains as either resistant or sensi-tive is not as clear-cut as it seems because such classi-fication is easily confused by different kinds of physi-ological responses, especially in batch culture (Aikinget al., 1982, 1984).
Microorganisms have been considered not only foruse in bioleaching of metals but also as squirrels of met-als from dilute solutions (Gadd and Griffiths, 1978).Large numbers of bacteria are capable of increasing cel-lular concentration of heavy metal ions to several ordersof magnitude higher than the background concentra-tions of these metals. These processes have a potentialuse in eradicating rare metals from dilute solution orremoving toxic metals from industrial effluents. Bioac-cumulation of toxic heavy metals has an unambiguouspractical significance, but in terms of an increased but
undesirable introduction of these pollutants into thetrophic chain (Atlas and Bartha, 1993).
CONCLUSIONSTo the best of our knowledge, this is the first study
pertaining the correlation between Cd+2 biosorption,uptake, and adsorption and Cd+2-mediated plasmid.This plasmid enhanced the Cd resistance in heterolo-gous host E. coli cells. This bacterial isolate is thus veryinteresting and a distinct model system. It could be use-ful for the study of Cd+2 bioaccumulation and kinetics.Finally, K. oxytoca may be helpful in the bioremedia-tion of Cd-contaminated soil and water. The next studywill involve the genetic background of Cd+2 bioreme-diation for this bacterium.
REFERENCESAdjeroud, N. R. 2009. Cadmium-Resistant Bacteria Isolated From Petra
Wastewater Treatment Station in Southern Jordan. MSc Thesis, Mu-tah University, Jordan.
Aiking, H., K. Kok, H. van Heerikhuizen, H., and J. van ’t Riet. 1982. Adap-tation to cadmium by Klebsiella aerogenes growing in continuous
K. M. Khleifat et al. 168
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

culture proceeds mainly via formation of cadmium sulfide. Appl.Environ. Microbiol. 44:938–944.
Aiking, H., A. Stijnman, C. van Garderen, H. van Heerikhuizen, andJ. van ’t Riet. 1984. Inorganic phosphate accumulation and cad-mium detoxification in Klebsiella aerogenes NCTC 418 grow-ing in continuous culture. Appl. Environ. Microbiol. 47:374–377.
Atlas, R. M., and R. Bartha. 1993. Microbial Ecology, Fundamentals andApplications, 237–238, 410. Canada: Benjamin/Cummings.
Banjerdkij, P., P. Vattananaviboon, and S. Mongkolsuk. 2003.Cadmium-induced adaptive resistance and cross-resistance tozinc in Xanthomonas campestris. Curr. Microbiol. 47:260–262.
Bulut, Y., N. Gozubenli, and H. Aydın. 2007. Equilibrium and kineticsstudies for adsorption of direct blue 71 from aqueous solution bywheat shells. J. Hazard. Mater. 144:300–306.
Cheah, U. E., W. Weigand, and B. C. Stark. 1987. Effect of recombi-nant plasmid size on cellular processes in Escherichia coli. Plasmid18:127–134.
Cohen, I., R. Bitan, and Y. Nitzan. 1991. The effect of zinc and cadmiumions on Escherichia coli. B. Microbios. 68:157–168.
Diels, L. 1997. Methods in Biotechnology, Vol. 2, Bioremediation Proto-cols, ed D. Sheehan, 283–295. Totowa, NJ: Humana Press.
Diels, L., and M. Mergeay. 1990. DNA probe-mediated detection of re-sistant bacteria from soils highly polluted by heavy metals. Appl.Environ. Microbiol. 56:1485–1491.
Doyle, J. J., R. T. Marshall, and A. W. H. Pfander. 1975. Effects of cadmiumon the growth and uptake of cadmium by microorganisms. Appl.Microbiol. 29:562–564.
El-Mansi, M., K. J. Anderson, C. A. Inche, L. K. Knowles, and D. J. Platt.2000. Isolation and curing of the Klebsiella pneumoniae large in-digenous plasmid using sodium dodecyl sulphate. Res. Microbiol.151:201–208.
Febrianto, J., A. J. Kosasih, Y. Sunarso, N. Ju Indraswati, and S. Ismadji.2009. Equilibrium and kinetic studies in adsorption of heavy metalsusing biosorbent: A summary of recent studies. J. Hazard. Mater.162:616–645.
Gadd, G. M., and A. J. Griffiths. 1978. Microorganisms and heavy metaltoxicity. Microb. Ecol. 4:303–317.
Hadziyannis, E., M. Tuohy, L. Thomas, G. W. Procop, J. A. Washington,and G. S. Hall. 2000. Screening and confirmatory testing for ex-tended spectrum beta-lactamases (ESBL) in Escherichia coli, Kleb-siella pneumoniae, and Klebsiella oxytoca clinical isolates. Diagn.Microbiol. Infect. Dis. 36:113–117.
Hao, Z., S. Chen, and D. B. Wilson. 1999. Cloning, expressionand characterization of cadmium and manganese uptake genesfrom Lactobacillus plantarum. Appl. Environ. Microbiol. 65:4746–4752.
Haq, R., S. K. Zaidi, and A. R. Shakoori. 1999. Cadmium resistant Enter-obacter cloacae and Klebsiella sp. isolated from industrial effluentsand their possible role in cadmium. World J. Microbiol. Biotechnol.15:283–290.
Hassen, A., N. Jedidi, M. Cherif, and A. M’Hiri, Boudabous and O. V. A.Cleemput. 1998. Mineralization of nitrogen in a clayey loamy soilamended with organic wastes enriched with Zn, Cu and Cd. Biores.Technol. 64:39–45.
Holmes, D. S. 1984. Improved rapid technique for screening re-combinantDNA plasmids in E. coli. Biotechniques. 2:68–69.
Holmes, J. D., D. J. Richardson, S. Saed, R. Evans-Gowing, D. A. Russell,and J. R. Sodeau. 1997. Cadmium-specific formation of metal sul-fide ‘Q particles’ by Klebsiella pneumonia. Microbiology 143:2521–2530.
Horitsu, H., K. Yamamoto, S. Wachi, and A. Fukuchi. 1986. Plasmid–determined cadmium resistance in Pseudomonas putida GAM-I iso-lated from soil. J. Bacteriol.165:334–335.
Joset, F., J. Guespin-Michel, and O. L. Butler. 1993. Prokaryotic Genet-ics: Genome Organisation Transfer and Plasticity, 51–78. Oxford:Blackwell Scientific.
Kanazawa, S., and K. Mori. 1996. Isolation of cadmium-resistant bacte-ria and their resistance mechanisms. Part II: Cadmium biosorptionby Cd-resistant and sensitive bacteria. Soil Sci. Plant Nut. 42:731–736.
Khazaeli, M. B., and R. S. Mitra. 1981. Cadmium-binding componentin Escherichia coli during accommodation to low levels of this ion.Appl. Environ. Microbiol. 41:46–50.
Khleifat, K. M., M. M. Abboud, and A. H. Al-Mustafa. 2006. Effect ofVitreoscilla hemoglobin gene (vgb) and metabolic inhibitors on cad-mium uptake by the heterologous host Enterobacter aerogenes.Process. Biochem. 41:930–934.
Khosravi, M., W. Ryan, D. A. Webster, and B. C. Stark. 1990. Variationof oxygen requirement with plasmid size in recombinant E. coli.Plasmid. 23:138–143.
Lamanna, C., and M. Malette. 1965. Basic Bacteriology-Its Biological andChemical Background, 3rd ed., 897–902. Baltimore: Williams andWilkins.
Langmuir, I. 1918. The adsorption of gases on plane surfaces of glass,mica and platinum. J Am. Chem. Soc. 40:1361–1403.
Mergeay, M., D. Nies, H. G. Schlegel, J. Gerits, P. Charles, and F. Vangi-jsegem. 1985. Alcaligenes eutrophus CH34 is a facultative chemoli-totroph with plasmid-bound resistance to heavy metals. J. Bacteriol.162:328–334.
Mitra, R. S., and I. A. Bernstein. 1978. Single-strand breakage in DNA ofEscherichia coli exposed to Cd+2. J. Bacteriol. 133:75–80.
Mitra, R. S., R. H. Gray, B. Chin, and I. A. Bernstein. 1975. Molecularmechanisms of accommodation in Escherichia coli to toxic levels ofCd2+. J. Bacteriol. 121:1180–1188.
Mullen, M. D., D. C. Wolf, F. G. Ferris, T. J. Beveridge, C. A. Flemming,and G. W. Bailley. 1989. Bacterial sorption of heavy metals. Appl.Environ. Microbiol. 55:3143–3149.
Nas, N., H. Young, N. Ahmed, and J. Gadd. 2005. Cadmium accumulationand DNA homology with metal resistance genes in sulfate-reducingbacteria. Appl. Environ. Microbiol. 71:4610–4618.
Nerey, M. D. C., S. E. Pichuantes, C. P. Saavedra, M. A. Araya,J. C. Tantalean, and C. C. Vasquez. 2002. Expression of Bacillusstearothermophilus LV cadmium resistance genes in Escherichiacoli causes hypersensitivity to cadmium chloride. Curr Microbiol.45:187–190.
Nies, D. H. 1995. The cobalt, zinc, and cadmium efflux system CzcABCfrom Alcaligenes eutrophus functions as a cation-proton antiporterin Escherichia coli. J. Bacteriol. 177:2707–2712.
Nies, D. H., and S. Silver. 1989. Plasmid-determined inducible efflux is re-sponsible for resistance to cadmium, zinc, and cobalt in Alcaligeneseutrophus. J. Bacteriol. 171:896–900.
Nies, D. H., and S. Silver. 1995. Ion efflux systems involved in bacterialmetal resistances. J. Indust. Microbiol. 14:186–199.
Novick, R. P., and C. Roth. 1968. Plasmid-linked resistance to in-organic salts in Staphylococcus aureus. J. Bacteriol. 95:1335–1342.
Ohman, D. E. 1988. Experiments in Gene Manipulation. Englewood Cliffs,NJ: Prentice Hall.
Pan-Hou, H. S., M. Nishimoto, and N. Imura. 1981. Possible role of mem-brane proteins in mercury resistence of Enterobacter aerogenes.Arch. Microbiol. 130:93–95.
Peiris, P. S., and A. M. Dlaminiy. 1999. Involvement of a plasmidin exopolysaccharide production by Klebsiella oxytoca. Microbiol.Biotechnol. 15:193–196.
Rensing, C., T. Pribyl, and D. H. Nies. 1997. New functions for thethree subunits of the czcCBA cation-proton-antiporter. J. Bacteriol.22:6871–6879.
Romero, M. C., E. M. Gatti, and D. E. Bruno. 1999. Effects of heavy metalson microbial activity of water and sediment communities. World J.Microbiol. Biotechnol. 15:179–184.
Rubin, S. J., and E. D. Rosenblum. 1971. Effects of the recipient strainand ultraviolet irradiation on transduction kinetics of the penicilli-nase plasmid of Staphylococcus aureus. Bacteriology. 108:1 192–1199.
169 Cd+2 Biosorption, Uptake, and Adsorption by Klebsiella oxytoca
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014

Sair, M. H., R. Tam, A. Reizer, and J. Reizer. 1994. Two novel familiesof bacterial membrane proteins concerened with nodulation, celldivision and transport. Mol. Microbiol. 11:841–847.
Sambrook, J., T. Maniatis, and E. F. Fritsch. 1989. Molecular Cloning—A Laboratory Manual. Cold Spring Habor, NY: Cold Spring HarborLaboratory Press.
Scott, J. A. 1990. Sites of cadmium uptake in bacteria used for biosorp-tion.Appl. Environ. Microbiol. 33:221–225.
Tynecka, Z., T. Zajac, and Z. Gos. 1975. Plasmid dependent impermeabilitybarrier to cadmium ions in Staphylococcus aureus. Acta Microbiol.Pol. Ser. A. 7:11–20.
Vullo, D. L., H. M. Ceretii, M. A. Daniel, S. A. M. Ramirez, and A. Zalts.2008. Cadmium, zinc and copper biosorption mediated by Pseu-domonas veronii 2E. Biores. Technol. 99:5574–5581.
Wilson, K., and J. Walker. 1994. Principles and Techniques of PracticalBiochemistry, 4th ed. Cambridge, UK: Cambridge University Press.
K. M. Khleifat et al. 170
Dow
nloa
ded
by [
Nor
thea
ster
n U
nive
rsity
] at
10:
52 2
3 N
ovem
ber
2014
Related Documents











