Brain Interstitial Nociceptin/Orphanin FQ Levels are Elevated in Parkinson’s Disease Matteo Marti, PhD, 1,2 Silvio Sarubbo, MD, 3 Francesco Latini, MD, 3 Michele Cavallo, MD, 3 Roberto Eleopra, MD, 4 Sara Biguzzi, MD, 4 Christian Lettieri, MD, 4 Carlo Conti, MD, 5 Michele Simonato, MD, 1,2 Silvia Zucchini, PhD, 1,2 Rocco Quatrale, MD, 6 Mariachiara Sensi, MD, 6 Sanzio Candeletti, PhD, 7 Patrizia Romualdi, PhD, 7 and Michele Morari, PhD 1,2 * 1 Department of Experimental and Clinical Medicine, Section of Pharmacology, University of Ferrara, Ferrara, Italy 2 Center for Neuroscience and National Institute of Neuroscience, University of Ferrara, Ferrara, Italy 3 Division of Neurosurgery, S. Anna University Hospital of Ferrara, Ferrara, Italy 4 Clinical and Neurological Department, Angel’s Hospital of Mestre, Venice, Italy 5 Department of Neurosurgery, Angel’s Hospital of Mestre, Venice, Italy 6 Clinical and Neurological Department, S. Anna University Hospital of Ferrara, Ferrara, Italy 7 Department of Pharmacology, University of Bologna, Bologna, Italy Abstract: Expression and release of nociceptin/orphanin FQ (N/OFQ) are elevated in the substantia nigra reticulata of 6- hydroxydopamine-hemilesioned rats, suggesting a pathogenic role for N/OFQ in Parkinson’s disease. In this study, we investigated whether elevation of N/OFQ expression in 6-hy- droxydopamine-hemilesioned rats selectively occurs in sub- stantia nigra and whether hypomotility following acute halo- peridol administration is accompanied by a rise in nigral N/ OFQ levels. Moreover, to prove a link between N/OFQ and idiopathic Parkinson’s disease in humans, we measured N/ OFQ levels in the cerebrospinal fluid of parkinsonian patients undergoing surgery for deep brain stimulation. In situ hybrid- ization demonstrated that dopamine depletion was associated with increase of N/OFQ expression in substantia nigra (com- pacta 1160%, reticulata 1105%) and subthalamic nucleus (145%), as well as reduction in caudate putamen (220%). No change was observed in globus pallidus, nucleus accum- bens, thalamus, and motor cortex. Microdialysis coupled to the bar test allowed to demonstrate that acute administration of haloperidol (0.8 and 3 mg/kg) increased nigral N/OFQ lev- els (maximally of 147% and 153%, respectively) in parallel with akinesia. A correlation with preclinical studies was found by analyzing N/OFQ levels in humans. Indeed, N/OFQ levels were found to be 3.5-fold elevated in the cerebrospi- nal fluid of parkinsonian patients (148 fmol/ml) compared with nonparkinsonian neurologic controls (41 fmol/ml). These data represent the first clinical evidence linking N/ OFQ to idiopathic Parkinson’s disease in humans. They strengthen the pathogenic role of N/OFQ in the modulation of parkinsonism across species and provide a rationale for developing N/OFQ receptor antagonists as antiparkinsonian drugs. Ó 2010 Movement Disorder Society Key words: cerebrospinal fluid; haloperidol; human; noci- ceptin/orphanin FQ; 6-OHDA; Parkinson’s disease Nociceptin/orphanin FQ (N/OFQ) is an opioid-like neuropeptide that activates the N/OFQ peptide (NOP) receptor. N/OFQ and its receptor are widely expressed throughout the rodent and primate central nervous sys- tems and modulate a number of biological functions such as pain, mood, reward, and locomotion. 1,2 N/OFQ has been proposed to play a role in Parkinson’s disease (PD) based on the findings that NOP receptor anta- gonists attenuated motor deficit in models of parkin- sonism, such as the haloperidol-treated rat and mouse, 3–5 the 6-hydroxydopamine (6-OHDA)-hemile- sioned rat, 4,6,7 or the 1-methyl-4-phenyl-1,2,5,6-tetrahy- dropyridine (MPTP)- intoxicated mouse and nonhuman primate. 8,9 Moreover, mice carrying a deletion of the gene encoding for the N/OFQ precursor (ppN/OFQ) were found resistant to the MPTP-induced degenera- Potential conflict of interest: The authors declare no conflict of in- terest. Received 12 January 2010; Revised 19 April 2010; Accepted 30 April 2010 Published online 29 June 2010 in Wiley InterScience (www. interscience.wiley.com). DOI: 10.1002/mds.23271 *Correspondence to: Michele Morari, Department of Experimental and Clinical Medicine, Section of Pharmacology, University of Fer- rara,via Fossato di Mortara 17-19, 44100 Ferrara, Italy E-mail: [email protected] 1723 Movement Disorders Vol. 25, No. 11, 2010, pp. 1723–1732 Ó 2010 Movement Disorder Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Brain Interstitial Nociceptin/Orphanin FQ Levelsare Elevated in Parkinson’s Disease
Matteo Marti, PhD,1,2 Silvio Sarubbo, MD,3 Francesco Latini, MD,3 Michele Cavallo, MD,3
Roberto Eleopra, MD,4 Sara Biguzzi, MD,4 Christian Lettieri, MD,4 Carlo Conti, MD,5
Michele Simonato, MD,1,2 Silvia Zucchini, PhD,1,2 Rocco Quatrale, MD,6 Mariachiara Sensi, MD,6
Sanzio Candeletti, PhD,7 Patrizia Romualdi, PhD,7 and Michele Morari, PhD1,2*
1Department of Experimental and Clinical Medicine, Section of Pharmacology, University of Ferrara, Ferrara, Italy2Center for Neuroscience and National Institute of Neuroscience, University of Ferrara, Ferrara, Italy
3Division of Neurosurgery, S. Anna University Hospital of Ferrara, Ferrara, Italy4Clinical and Neurological Department, Angel’s Hospital of Mestre, Venice, Italy
5Department of Neurosurgery, Angel’s Hospital of Mestre, Venice, Italy6Clinical and Neurological Department, S. Anna University Hospital of Ferrara, Ferrara, Italy
7Department of Pharmacology, University of Bologna, Bologna, Italy
Abstract: Expression and release of nociceptin/orphanin FQ(N/OFQ) are elevated in the substantia nigra reticulata of 6-hydroxydopamine-hemilesioned rats, suggesting a pathogenicrole for N/OFQ in Parkinson’s disease. In this study, weinvestigated whether elevation of N/OFQ expression in 6-hy-droxydopamine-hemilesioned rats selectively occurs in sub-stantia nigra and whether hypomotility following acute halo-peridol administration is accompanied by a rise in nigral N/OFQ levels. Moreover, to prove a link between N/OFQ andidiopathic Parkinson’s disease in humans, we measured N/OFQ levels in the cerebrospinal fluid of parkinsonian patientsundergoing surgery for deep brain stimulation. In situ hybrid-ization demonstrated that dopamine depletion was associatedwith increase of N/OFQ expression in substantia nigra (com-pacta 1160%, reticulata 1105%) and subthalamic nucleus(145%), as well as reduction in caudate putamen (220%).No change was observed in globus pallidus, nucleus accum-
bens, thalamus, and motor cortex. Microdialysis coupled tothe bar test allowed to demonstrate that acute administrationof haloperidol (0.8 and 3 mg/kg) increased nigral N/OFQ lev-els (maximally of 147% and 153%, respectively) in parallelwith akinesia. A correlation with preclinical studies wasfound by analyzing N/OFQ levels in humans. Indeed, N/OFQlevels were found to be �3.5-fold elevated in the cerebrospi-nal fluid of parkinsonian patients (148 fmol/ml) comparedwith nonparkinsonian neurologic controls (41 fmol/ml).These data represent the first clinical evidence linking N/OFQ to idiopathic Parkinson’s disease in humans. Theystrengthen the pathogenic role of N/OFQ in the modulationof parkinsonism across species and provide a rationale fordeveloping N/OFQ receptor antagonists as antiparkinsoniandrugs. � 2010 Movement Disorder SocietyKey words: cerebrospinal fluid; haloperidol; human; noci-
ceptin/orphanin FQ; 6-OHDA; Parkinson’s disease
Nociceptin/orphanin FQ (N/OFQ) is an opioid-like
neuropeptide that activates the N/OFQ peptide (NOP)
receptor. N/OFQ and its receptor are widely expressed
throughout the rodent and primate central nervous sys-
tems and modulate a number of biological functions
such as pain, mood, reward, and locomotion.1,2 N/OFQ
has been proposed to play a role in Parkinson’s disease
(PD) based on the findings that NOP receptor anta-
gonists attenuated motor deficit in models of parkin-
sonism, such as the haloperidol-treated rat and
mouse,3–5 the 6-hydroxydopamine (6-OHDA)-hemile-
sioned rat,4,6,7 or the 1-methyl-4-phenyl-1,2,5,6-tetrahy-
dropyridine (MPTP)- intoxicated mouse and nonhuman
primate.8,9 Moreover, mice carrying a deletion of the
gene encoding for the N/OFQ precursor (ppN/OFQ)
were found resistant to the MPTP-induced degenera-
Potential conflict of interest: The authors declare no conflict of in-terest.
Received 12 January 2010; Revised 19 April 2010; Accepted 30April 2010
Published online 29 June 2010 in Wiley InterScience (www.
interscience.wiley.com). DOI: 10.1002/mds.23271
*Correspondence to: Michele Morari, Department of Experimentaland Clinical Medicine, Section of Pharmacology, University of Fer-rara,via Fossato di Mortara 17-19, 44100 Ferrara, ItalyE-mail: [email protected]
1723
Movement DisordersVol. 25, No. 11, 2010, pp. 1723–1732� 2010 Movement Disorder Society

tion of SN compacta (SNc) dopamine (DA) cells, sug-
gesting that N/OFQ contributes to both the parkinso-
nian symptoms and neurotoxicity associated with the
disease.4 To further endorse a pathogenic role of N/OFQ
in PD, the parkinsonian toxins 6-OHDA4,10 and
MPP110,11 promoted ppN/OFQ expression in rat SN.
Indeed, such upregulation resulted in increased extracel-
lular N/OFQ levels in the SNr of 6-OHDA-lesioned
rats4 possibly leading to exacerbation of the physiologi-
cally inhibitory influence of endogenous N/OFQ on ni-
grostriatal DA transmission and motor behavior.12 De-
spite such compelling preclinical evidence, no relation-
ship between N/OFQ and PD in humans has been
demonstrated so far.2 Nonetheless, lower ppN/OFQ and
NOP expression was found in the hippocampus and cen-
tral amygdala, respectively, of alcoholics,13 and lower
N/OFQ binding was observed in patients with temporal
lobe epilepsy,14 indicating that changes in N/OFQ trans-
mission may be associated with neurologic diseases. To
possibly support this view, N/OFQ plasma levels were
found elevated in Wilson’s disease,15 an autosomal re-
cessive disorder related to defective copper metabolism
in the liver and copper overload in tissues, leading,
among others, to basal ganglia degeneration.16
The aims of this study were 3-fold: (i) to investigate
whether the increase in ppN/OFQ expression following
6-OHDA lesioning is selective for SN or involves
other brain areas, (ii) to demonstrate that functional
parkinsonism is associated with an increase in nigral
N/OFQ levels, and (iii) to provide clinical evidence
that N/OFQ may be relevant for PD in humans. To
pursue these aims, in situ hybridization was employed
to study ppN/OFQ expression in SN, primary (M1)
and secondary (M2) motor cortex, caudate-putamen
(CPu), nucleus accumbens (Ac), globus pallidus (GP),
and subthalamic nucleus (STN) of 6-OHDA-hemile-
sioned rats in comparison with sham-operated rats.
Moreover, microdialysis coupled to the bar test (a test
for akinesia) was used in awake rats to simultaneously
monitor SNr N/OFQ levels and immobility time fol-
lowing haloperidol administration. Finally, N/OFQ lev-
els were measured in brain cerebrospinal fluid (CSF)
samples obtained from PD patients undergoing surgery
for deep brain stimulation and compared with those
measured in nonparkinsonian neurologic controls.
MATERIALS AND METHODS
Animals
Male Sprague-Dawley rats (Harlan, S. Pietro al Nati-
sone, Italy) were used for this study. Four animals were
housed for cage, with free access to food and water, and
kept under environmentally controlled conditions (12-
hour light/dark cycle with light on between 07:00 AM
and 07:00 PM). The experimental protocols were
approved by the Italian Ministry of Health (licence no
194/2008-B) and the Ethical Committee of the Univer-
sity of Ferrara, and adequate measures were taken to
minimize animal pain and the number of animals used.
6-OHDA Lesion
Unilateral lesion of dopaminergic neurons was
induced in isoflurane-anesthetized male rats as previ-
ously described.6 Eight micrograms of 6-OHDA (dis-
solved in 4 ll of saline containing 0.02% ascorbic
acid) were stereotaxically injected into the right medial
forebrain bundle according to the following coordinates
from bregma: anteroposterior-4.4 mm, mediolateral-1.2
mm, dorsoventral-7.8 mm below dura.17 Control rats
(sham operated) were injected with vehicle. Two
weeks after 6-OHDA injection, successfully lesioned
rats (hemiparkinsonian) were selected using the rota-
tional model.18 Rats showing a turning behavior >7
turns/min in the direction ipsilateral to the lesion fol-
lowing intraperitoneal administration of amphetamine
(5 mg/kg) were enrolled in the study. This behavior
has been associated with >95% loss of striatal DA ter-
minals6 and extracellular DA levels.19 Sham-operated
rats were also injected with amphetamine.
In Situ Hybridization
Rats were killed by decapitation under light diethyl-
ether anesthesia. Their brains rapidly removed, frozen
in isopentane, cooled in a dry ice/methanol bath, and
stored at 2708C until use. Probes were prepared from
a full-length cDNA insert cloned in a pBluescript
SK(1) plasmid (proN/OFQ from C. Mollereau, Tou-
louse, France). The proN/OFQ plasmid was linearized
with BamH1 and transcribed with T7 RNA polymerase
to obtain antisense riboprobes or linearized with Xho1
and transcribed with T3 RNA polymerase to obtain
sense riboprobes. All riboprobes were obtained by run-
ning the transcription assays in the presence of a[33P]-rUTP, and hydrolyzed to fragments of approximately
200 base pairs with sodium carbonate at 608C.20
Twenty-micrometer coronal sections were cut at
three different brain levels from bregma: 10.70 to
11.20, 20.30 to 20.92, and 22.56 to 23.60.17 Sec-
tions were thaw mounted onto polylysine-coated slides,
fixed in 4% paraformaldehyde, soaked in 33 phos-
phate buffered saline, rinsed in a graded ethanol series,
dried, and stored at 2208C until use. Immediately
before in situ hybridization, they were pretreated with
proteinase K (1 lg/ll, 10 minutes, 378C) and with ace-
1724 M. MARTI ET AL.
Movement Disorders, Vol. 25, No. 11, 2010

tic anhydride (0.25% vol/vol, 10 minutes, and room
temperature). In situ hybridization was performed as
previously described.21 Briefly, sections were incubated
overnight at 528C with 40 ll hybridization mixture:
50% deionized formamide, 23 STE (Sodium-Tris-
EDTA), 53 Denhart’s solution, 100 lg/ml ssDNA,
100 lg/ml tRNA, 0.05% sodium pyrophosphate, and
60 ng/ml [33P]-riboprobe. They were then rinsed in 43SSC, treated with RNAase A (20 lg/ml, 30 minutes,
378C), washed in 13 SSC for 10 minutes, 0.13 SSC
at 528C for 30 minutes, and 0.13 SSC at room tem-
perature for 10 minutes, and dehydrated. Autoradio-
grams were generated by apposing these dried sections
alongside [33P]-riboprobe standards to Kodak BioMax
MR film at 2708C for 30 days.
The mean total optical density within an area of in-
terest was calculated by multiple sampling of that area
in two sections taken from each animal using a digital
analysis system (RBR Altair, Firenze, Italy). Back-
ground optical density in film areas not exposed to sec-
tions was subtracted from the total optical density.
Finally, data have been expressed as percent of optical
density in the control (noninjected) side.
Microdialysis in Haloperidol-Treated Rats
One probe of concentric design (1-mm dialyzing
membrane; AN69; Hospal, Bologna, Italy) was
implanted in SNr (anteroposterior 25.5, mediolateral
6 2.2, VD 28.3) under isoflurane anaesthesia. After
surgery, rats were allowed to recover, and experiments
were run 48 hour after probe implantation. Microdialy-
sis probes were perfused (3 ll/minutes) with a modi-
fied Ringer solution (for composition see above) added
with polypep 0.1%, captopril 300 lM, and BSA
0.3%.22 Haloperidol was administered intraperitoneally
after a 270-minute washout period, at doses causing
akinesia in both rats and mice (0.8 and 3 mg/kg,
respectively), according to our previous studies.3,4
Microdialysis samples were collected every 60
minutes, starting 180 minutes before haloperidol
administration. Samples were mixed with an equal vol-
ume of trifluoroacetic acid (TFA; 1% vol:vol) and
loaded onto C18 cartridges (SEPCOL-1; Bachem Ltd.,
St Helens, United Kingdom). Microcolumns were
washed twice with 3 ml 1% TFA and then eluted with
3 ml 60% acetonitrile in 1% TFA. Eluates were lyoph-
ilized and stored at 2708C until radioimmunoassay.
The adopted extraction procedure was validated by
measuring the recovery of [3H-Leucyl]-N/OFQ (Amer-
sham Biosciences Europe, Freiburg, Germany) under
identical conditions. More than 95% of the tritiated N/
OFQ added to a sample was recovered in the eluate
(90–92% in the first milliliter).
Analysis of Endogenous N/OFQ
N/OFQ-like immunoreactivity (N/OFQ-LI) present
in microdialysis fractions was measured 7 weeks after
6-OHDA lesion by a specific radioimmunoassay (RIA)
according to the method described by Ploj et al.23 Ly-
ophilized samples, reconstituted in 25 ll methanol in
0.1% HCl (1:1), were mixed with 100 ll [125I]-N/OFQ(Bachem Ltd.) and 100 ll N/OFQ antiserum (anti-
serum 96:21, kindly supplied by Dr I. Nylander, Upp-
sala, Sweden). The antiserum, raised against N/OFQ,
shows 0.5% cross-reactivity with the N/OFQ fragment
N/OFQ (1–13) and less than 0.1% cross-reactivity with
nocistatin and the following opioid peptides: dynorphin
A (DYN A), dynorphin B (DYN B), their truncated
fragments or elongated forms DYN A (1–6), DYN A
(1–32), and DYN B (1–29), or with Met-enkephalin,
Met-enkephalinArg6Phe7, Leu-enkephalin, and b-endorphin. The antiserum was used at the appropriate
dilution to give 30% to 34% binding of the [125I]-N/
OFQ added (4,800–5,000 cpm). The labelled peptide
and the antiserum were diluted in a gelatin buffer con-
taining 0.15 M NaCl, 0.02% sodium azide, 0.1% gela-
tin, 0.1% Triton X-100, and 0.1% BSA in a 0.05-M so-
dium phosphate buffer (pH 7.4). RIA tubes were incu-
bated at 48C for 24 h. Charcoal slurry (1 ml/tube) was
used to separate free and antibody bound peptide (15%
horse serum, 3% charcoal, and 0.3% dextran in RIA
buffer). Bound peptide was separated by centrifugation
(5,000g at 48C), and 1 ml aliquots of the supernatants
were counted for 1 minute on a Beckman 5500 gamma
counter (Beckman, Fullerton, CA). The detection limit
of the RIA assay was 1 fmol/tube. RIA curves and
data were analyzed using the GraphPad Prism 4 soft-
ware for Windows (GraphPad Software, San Diego,
CA). The limit of detection of the assay was 1 fmol,
and N/OFQ in vitro recovery was 2.0 6 0.1%.
Behavioral Testing During Microdialysis
To correlate changes of N/OFQ dialysate levels with
motor activity, rats undergoing microdialysis were
challenged in the bar test, a test for akinesia24 as previ-
ously described.3,7 Behavioral testing was performed
every 60 minutes. The bar test was performed essen-
tially as described above. Rat forepaws were placed
alternatively on blocks of increasing heights (3, 6, and
9 cm), and the immobility time was recorded (cut-off
time of 20 seconds).
1725N/OFQ LEVELS IN PARKINSON’S DISEASE
Movement Disorders, Vol. 25, No. 11, 2010

Subjects
Experiments were undertaken with the understanding
and written consent of each subject, with the approval
of the local Ethical Committees of the Hospitals of
Ferrara and Mestre, and in compliance with national
registration and the Code of Ethical Principles for
Medical Research Involving Human Subjects of the
World Medical Association (Declaration of Helsinki).
Two groups of patients were enrolled in the study:
patients suffering from idiopathic PD (n 5 20) and
nonparkinsonian neurologic controls (n 5 17). Patients
with idiopathic PD were diagnosed by neurologists at
the two Hospitals involved in the study using the Diag-
nostic Criteria of Gelb et al.25 Unified Parkinson’s dis-
ease rating scale (UPDRS, Part III: Motor section) was
used to quantify parkinsonian disabilities. CSF (usually
�0.5 ml) was obtained from PD patients subjected to
craniotomy (under local anaesthesia) performed to
implant an electrode for deep brain stimulation. All PD
patients have been off-treatment (L-dopa and dopami-
nomimetics) for at least 24 hours before surgery. In
nonparkinsonian neurologic controls, CSF (�0.5 ml)
was obtained by either lumbar puncture (n 5 7) or cra-
nial drainage (n 5 10). Subjects were affected by dif-
ferent pathologies: tumors, brain haemorrhage, normo-
tensive hydrocephalus, head trauma, paraplegy and
amyotrophic lateral sclerosis. Brain CSF was collected
during craniotomy or, in a fewer cases, via ventricular
catheter to reduce intracranial pressure.
Analysis of Endogenous N/OFQ
CSF samples were mixed with an equal volume of
TFA (1% vol:vol) and stored at 2808C until use. Sam-
ples were then lyophilized and processed for RIA as
described above.
Data Presentation and Statistical Analysis
In microdialysis studies, N/OFQ release was
expressed as N/OFQ concentration in a 60-minute dial-
ysate fraction and reported as N/OFQ-like immunore-
activity (N/OFQ-LI fmol/fraction). N/OFQ-like immu-
noreactivity in CSF sample was expressed as femto-
moles per milliliter of analyzed CSF sample. Statistical
analysis was performed by the Student’s t test for
unpaired data (Figs 2 and 4) or by repeated-measure
one-way ANOVA (Fig. 3). If ANOVA yielded a sig-
nificant F score, then post hoc analysis was performed
by contrast analysis to determine group differences. If
a significant time 3 treatment interaction was found,
the sequentially rejective Bonferroni test was used
(implemented on excel spreadsheet) to determine spe-
cific differences (i.e., at the single time-point level)
between groups. P values <0.05 were considered to be
statistically significant.
Materials
Haloperidol was purchased from Tocris Neuramin
(Bristol, United Kingdom), amphetamine and 6-OHDA
bromide from Sigma Chemical Company (St Louis,
MO). All drugs were freshly dissolved in isosmotic sa-
line solution just before use.
RESULTS
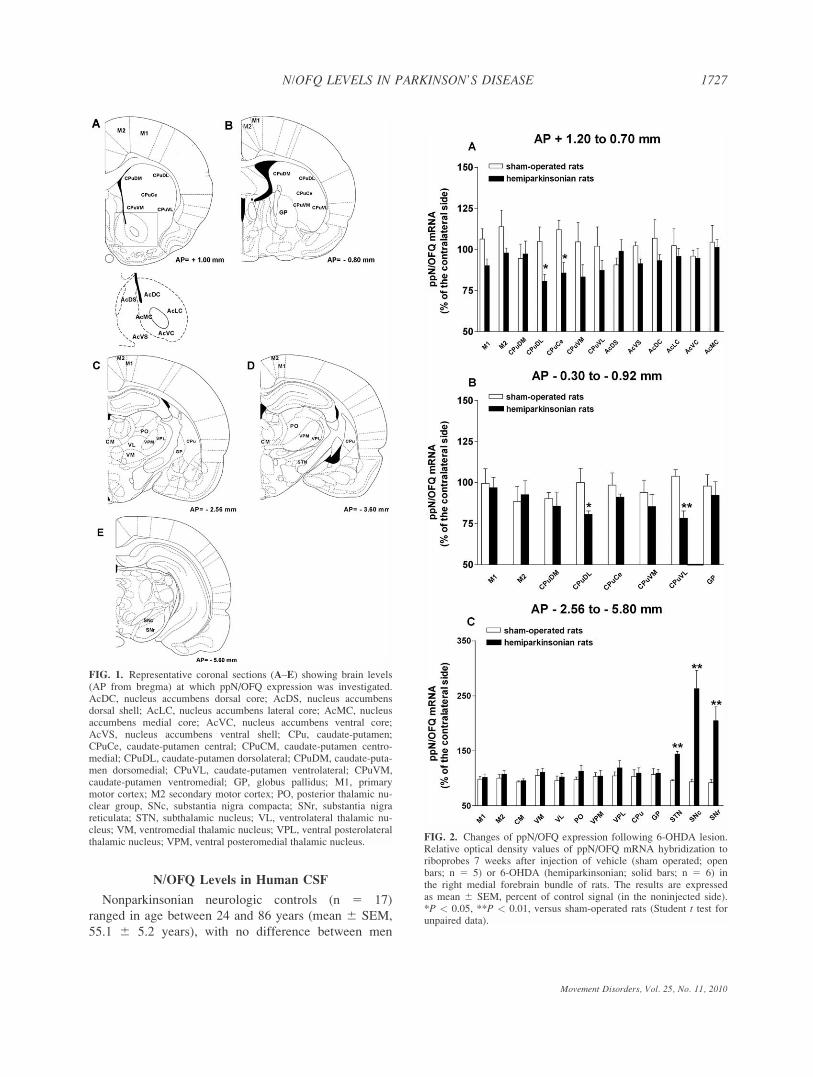
In Situ Hybridization
In situ hybridization analysis revealed distribution
patterns of ppN/OFQ mRNA consistent with those
described in other studies.26,27 ppN/OFQ gene expres-
sion was studied 7 weeks after unilateral injection of
vehicle or 6-OHDA. In sham-operated rats, constitutive
ppN/OFQ gene expression was found in all areas
examined (Fig. 1), highest levels being found in motor
cortex whereas lowest in STN. The rank order of
expression was M1 5 M2 > thalamus > GP 5 SNr 5SNc > CPu 5 Ac > STN. No changes were observed
between the injected and noninjected side in vehicle-
injected animals. Conversely, in the lesioned side of 6-
OHDA-treated rats, reduced expression (220%) was
observed in the dorsolateral, central, and ventrolateral
portions of the CPu (Fig. 2A, B) whereas significant
increases (Fig. 2C) were observed in STN (145%),
SNc (1160%) and SNr (1105%).
N/OFQ Levels in the SNr of
Haloperidol-Treated Rats
Haloperidol caused significant increases in immobil-
ity time compared with saline-treated animals. Immo-
bility time was almost maximally elevated (50.4 6 0.6
seconds; Fig. 3A) 1 hour after intraperitoneal adminis-
tration of 0.8 mg/kg haloperidol, then stabilized above
threshold levels (60 seconds). A similar response was
observed following haloperidol 3 mg/kg (Fig. 3A),
with the difference that immobility time was already
maximal after 1 hour. In line with akinesia values, N/
OFQ levels rose in the first hour after haloperidol (0.8
mg/kg; Fig 3B), peaked (147%) after 2 hours, and
then displayed a tendency to decline. A quali-quantita-
tively similar response was observed following halo-
peridol 3 mg/kg (Fig. 3B).
1726 M. MARTI ET AL.
Movement Disorders, Vol. 25, No. 11, 2010
N/OFQ Levels in Human CSF
Nonparkinsonian neurologic controls (n 5 17)
ranged in age between 24 and 86 years (mean 6 SEM,
55.1 6 5.2 years), with no difference between men
FIG. 1. Representative coronal sections (A–E) showing brain levels(AP from bregma) at which ppN/OFQ expression was investigated.AcDC, nucleus accumbens dorsal core; AcDS, nucleus accumbensdorsal shell; AcLC, nucleus accumbens lateral core; AcMC, nucleusaccumbens medial core; AcVC, nucleus accumbens ventral core;AcVS, nucleus accumbens ventral shell; CPu, caudate-putamen;CPuCe, caudate-putamen central; CPuCM, caudate-putamen centro-medial; CPuDL, caudate-putamen dorsolateral; CPuDM, caudate-puta-men dorsomedial; CPuVL, caudate-putamen ventrolateral; CPuVM,caudate-putamen ventromedial; GP, globus pallidus; M1, primarymotor cortex; M2 secondary motor cortex; PO, posterior thalamic nu-clear group, SNc, substantia nigra compacta; SNr, substantia nigrareticulata; STN, subthalamic nucleus; VL, ventrolateral thalamic nu-cleus; VM, ventromedial thalamic nucleus; VPL, ventral posterolateralthalamic nucleus; VPM, ventral posteromedial thalamic nucleus. FIG. 2. Changes of ppN/OFQ expression following 6-OHDA lesion.
Relative optical density values of ppN/OFQ mRNA hybridization toriboprobes 7 weeks after injection of vehicle (sham operated; openbars; n 5 5) or 6-OHDA (hemiparkinsonian; solid bars; n 5 6) inthe right medial forebrain bundle of rats. The results are expressedas mean 6 SEM, percent of control signal (in the noninjected side).*P < 0.05, **P < 0.01, versus sham-operated rats (Student t test forunpaired data).
1727N/OFQ LEVELS IN PARKINSON’S DISEASE
Movement Disorders, Vol. 25, No. 11, 2010

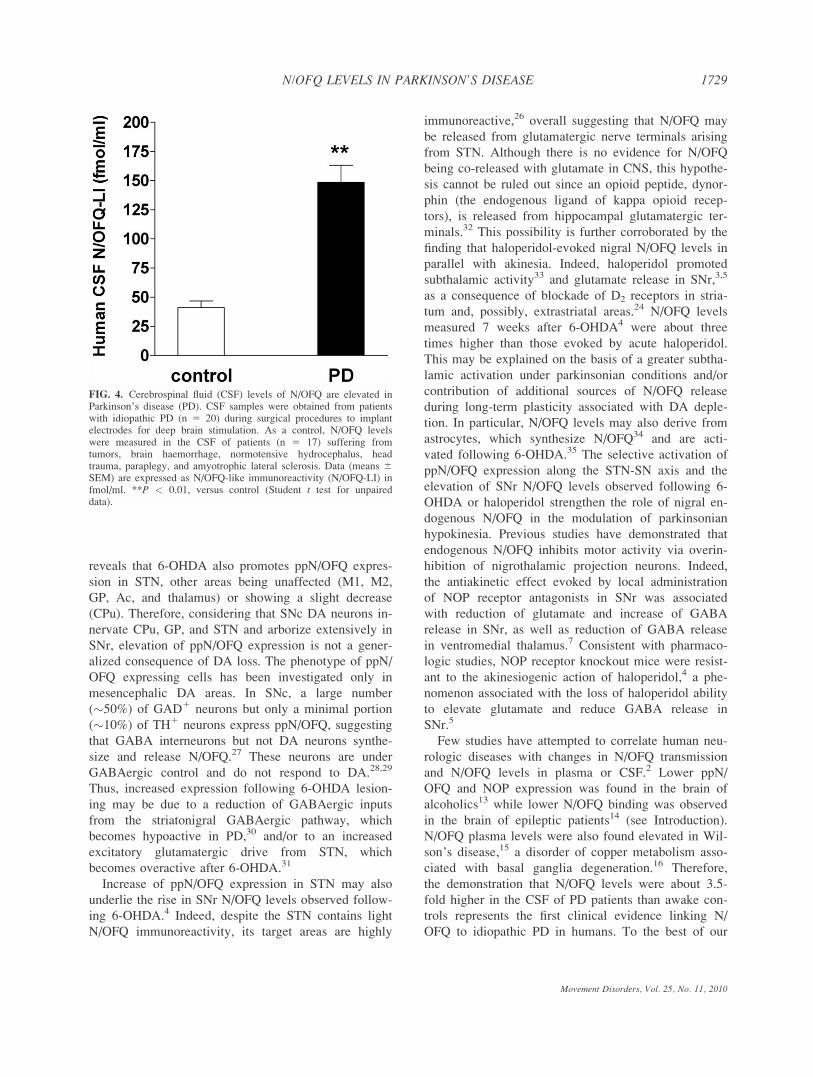
(55.0 6 8.2, n 5 8) and women (54.8 6 6.2; n 5 9;
Table 1). N/OFQ levels in the CSF of control patients
were 41.1 6 5.6 fmol/ml. Higher levels were found in
women (50.1 6 8.0 fmol/ml) than in men (30.9 6 6.3
fmol/ml), but this difference did not reach statistical
significance (P 5 0.084). Samples were obtained from
brain drainage (10 patients) or lumbar puncture (7
patients). N/OFQ levels in brain (33.0 6 4.9 fmol/ml)
and lumbar (52.6 6 10.5 fmol/ml) samples were not
statistically different. PD patients (n 5 20; 11 men and
9 women) ranged in age from 43 to 71 years (mean
60.1 6 1.9 years; Table 1). Male and female subjects
were homogeneous for age (60.5 6 2.5 and 60.6 6 2.6
years, respectively) and age at PD onset (49.2 6 2.3
and 48.2 6 2.3 years, respectively), both being
affected by PD by more than 11 years (Table 1). They
had advanced and severe PD according to both the
UPDRS III (46.1 6 4.1) and the Hoehn and Yahr
(2.9 6 0.1) scales, with no difference between men
(47.5 6 4.6 and 2.9 6 0.1, respectively) and women
(45.0 6 4.5 and 2.8 6 0.1, respectively). N/OFQ lev-
els were about 3.5-fold higher (148.5 6 14.6 fmol/ml)
in the CSF of PD patients than controls (41.1 6 5.6
fmol/ml), with no significant gender difference (Table
1 and Fig. 4).
DISCUSSION
6-OHDA lesioning has been previously reported to
stimulate ppN/OFQ expression in SN.4,10,27 This study
FIG. 3. Haloperidol simultaneously elevated extracellular N/OFQ levels and akinesia in naive rats. Microdialysis coupled to the bar test wasused to simultaneously monitor immobility time (in seconds; A) and extracellular N/OFQ levels in substantia nigra reticulata (SNr; B) of naiverats following administration of haloperidol (0.8 and 3 mg/kg, intraperitoneal) or saline. Behavioral testing and sample collection was performedevery hour. Data (mean 6 SEM of 9 determinations) are expressed as seconds of immobility (A) and N/OFQ-like immunoreactivity (N/OFQ-LI)in fmol/fraction (B). **P < 0.05, versus saline. ##P < 0.05 versus haloperidol 3 mg/kg (repeated-measure ANOVA followed by contrast analysisand the sequentially rejective Bonferroni’s test).
TABLE 1. Clinical features and N/OFQ levels in the cerebrospinal fluid of subjects enrolled in the study
Nonparkinsonian neurologic controls Parkinson’s disease
Total Male Female Total Male Female
N 17 8 9 20 11 9Age (yr) 55.1 6 5.2 55.0 6 8.2 54.8 6 6.2 60.6 6 1.8 60.5 6 2.5 60.7 6 2.6Age at onset (yr) – – – 48.7 6 1.6 49.2 6 2.3 48.2 6 2.3UPDRS III – – – 46.1 6 4.1 47.5 6 4.6 45.0 6 4.5Hoehn & Yahr stage – – – 2.9 6 0.1 2.9 6 0.1 2.8 6 0.1Disease duration (yr) – – – 11.8 6 1.1 11.3 6 1.4 12.5 6 1.6N/OFQ (fmol/ml) 41.1 6 5.6 30.9 6 6.3 55.1 6 8.2 148.5 6 14.6 158.1 6 23.5 136.7 6 15.8
Data are expressed as mean 6 SEM.
1728 M. MARTI ET AL.
Movement Disorders, Vol. 25, No. 11, 2010
reveals that 6-OHDA also promotes ppN/OFQ expres-
sion in STN, other areas being unaffected (M1, M2,
GP, Ac, and thalamus) or showing a slight decrease
(CPu). Therefore, considering that SNc DA neurons in-
nervate CPu, GP, and STN and arborize extensively in
SNr, elevation of ppN/OFQ expression is not a gener-
alized consequence of DA loss. The phenotype of ppN/
OFQ expressing cells has been investigated only in
mesencephalic DA areas. In SNc, a large number
(�50%) of GAD1 neurons but only a minimal portion
(�10%) of TH1 neurons express ppN/OFQ, suggesting
that GABA interneurons but not DA neurons synthe-
size and release N/OFQ.27 These neurons are under
GABAergic control and do not respond to DA.28,29
Thus, increased expression following 6-OHDA lesion-
ing may be due to a reduction of GABAergic inputs
from the striatonigral GABAergic pathway, which
becomes hypoactive in PD,30 and/or to an increased
excitatory glutamatergic drive from STN, which
becomes overactive after 6-OHDA.31
Increase of ppN/OFQ expression in STN may also
underlie the rise in SNr N/OFQ levels observed follow-
ing 6-OHDA.4 Indeed, despite the STN contains light
N/OFQ immunoreactivity, its target areas are highly
immunoreactive,26 overall suggesting that N/OFQ may
be released from glutamatergic nerve terminals arising
from STN. Although there is no evidence for N/OFQ
being co-released with glutamate in CNS, this hypothe-
sis cannot be ruled out since an opioid peptide, dynor-
phin (the endogenous ligand of kappa opioid recep-
tors), is released from hippocampal glutamatergic ter-
minals.32 This possibility is further corroborated by the
finding that haloperidol-evoked nigral N/OFQ levels in
parallel with akinesia. Indeed, haloperidol promoted
subthalamic activity33 and glutamate release in SNr,3,5
as a consequence of blockade of D2 receptors in stria-
tum and, possibly, extrastriatal areas.24 N/OFQ levels
measured 7 weeks after 6-OHDA4 were about three
times higher than those evoked by acute haloperidol.
This may be explained on the basis of a greater subtha-
lamic activation under parkinsonian conditions and/or
contribution of additional sources of N/OFQ release
during long-term plasticity associated with DA deple-
tion. In particular, N/OFQ levels may also derive from
astrocytes, which synthesize N/OFQ34 and are acti-
vated following 6-OHDA.35 The selective activation of
ppN/OFQ expression along the STN-SN axis and the
elevation of SNr N/OFQ levels observed following 6-
OHDA or haloperidol strengthen the role of nigral en-
dogenous N/OFQ in the modulation of parkinsonian
hypokinesia. Previous studies have demonstrated that
endogenous N/OFQ inhibits motor activity via overin-
hibition of nigrothalamic projection neurons. Indeed,
the antiakinetic effect evoked by local administration
of NOP receptor antagonists in SNr was associated
with reduction of glutamate and increase of GABA
release in SNr, as well as reduction of GABA release
in ventromedial thalamus.7 Consistent with pharmaco-
logic studies, NOP receptor knockout mice were resist-
ant to the akinesiogenic action of haloperidol,4 a phe-
nomenon associated with the loss of haloperidol ability
to elevate glutamate and reduce GABA release in
SNr.5
Few studies have attempted to correlate human neu-
rologic diseases with changes in N/OFQ transmission
and N/OFQ levels in plasma or CSF.2 Lower ppN/
OFQ and NOP expression was found in the brain of
alcoholics13 while lower N/OFQ binding was observed
in the brain of epileptic patients14 (see Introduction).
N/OFQ plasma levels were also found elevated in Wil-
son’s disease,15 a disorder of copper metabolism asso-
ciated with basal ganglia degeneration.16 Therefore,
the demonstration that N/OFQ levels were about 3.5-
fold higher in the CSF of PD patients than awake con-
trols represents the first clinical evidence linking N/
OFQ to idiopathic PD in humans. To the best of our
FIG. 4. Cerebrospinal fluid (CSF) levels of N/OFQ are elevated inParkinson’s disease (PD). CSF samples were obtained from patientswith idiopathic PD (n 5 20) during surgical procedures to implantelectrodes for deep brain stimulation. As a control, N/OFQ levelswere measured in the CSF of patients (n 5 17) suffering fromtumors, brain haemorrhage, normotensive hydrocephalus, headtrauma, paraplegy, and amyotrophic lateral sclerosis. Data (means 6SEM) are expressed as N/OFQ-like immunoreactivity (N/OFQ-LI) infmol/ml. **P < 0.01, versus control (Student t test for unpaireddata).
1729N/OFQ LEVELS IN PARKINSON’S DISEASE
Movement Disorders, Vol. 25, No. 11, 2010

knowledge, it is also the first measure of N/OFQ lev-
els in the CSF obtained at the brain level. N/OFQ lev-
els in lumbar samples of nonparkinsonian neurologic
controls (52.6 fmol) were similar to those reported in
a previous study (44.7 fmol)36 and in the same order
of magnitude of those measured in brain samples
(33.0 fmol). However, cranial and lumbar sampling
was never performed in the same subject, making it
difficult to unequivocally prove that N/OFQ levels in
CSF do not have a rostrocaudal gradient as consis-
tently demonstrated for monoamines and their metabo-
lites.37–39 Nevertheless, the lack of difference in N/
OFQ levels between brain and lumbar samples sug-
gests that this is not the case, in line with what has
been observed for opioids (b-lipotropin, b-endor-phin,40 and other neuropeptides such as NPY,41 sub-
stance P42 or ACTH).40 However, the possibility that
lumbar CSF samples are enriched with N/OFQ derived
from spinal cord neurons should be considered due to
the high density of N/OFQ immunoreactivity in this
area.26
Human CSF is produced by choroid plexus and is
in equilibrium with brain interstitial fluid, reflecting
its composition.43 Thus, N/OFQ levels in CSF reflect
neuronal activity/metabolism. ppN/OFQ expression in
the human and rodent brains substantially over-
laps,13,26,44 although, possibly relevant to this study,
highest ppN/OFQ mRNA levels were found in human
STN.44 N/OFQ content was also measured in a num-
ber of brain structures,45 among which the SN (�7.9
fmol/mg tissue), CPu (�4.6 fmol/mg tissue), and GP
(�2.2 fmol/mg tissue). Therefore, enhanced N/OFQ
levels in the CSF of PD patients may be consistent
with increased N/OFQ expression and release in the
basal ganglia complex, as predicted on the basis of
studies in rodent models of parkinsonism. On a quan-
titative basis, the observed increase in N/OFQ levels
(3.5-folds) was quite remarkable, also considering the
CSF volume (�160 ml) and formation rate (0.40 ml/
minutes, leading to complete renewal every 6
hours).46 Although a reduction in CSF turnover occur-
ring with age46 may amplify such difference, the
degree of such increase may further emphasize the
pathologic relevance of this phenomenon. The influ-
ence of N/OFQ in motor control in humans is still
unknown. Studies with NOP receptor antagonists in
naive and parkinsonian nonhuman primates8 have pro-
vided preliminary evidence that endogenous N/OFQ
may act as a physiologic constraint of movement, as
shown in rodents.12,47,48 On this basis, an increase in
N/OFQ levels in human CSF may be considered as
pathogenic, leading to a worsening of motor deficit
and, possibly, also contributing to neurodegeneration
associated with PD.4,11
CONCLUSIONS
Enhancement of ppN/OFQ expression has been
observed in the STN and SN of 6-OHDA-hemilesioned
rats. Moreover, elevation of SNr extracellular N/OFQ
levels was observed following administration of akine-
siogenic doses of haloperidol (a functional model of
parkinsonism) in line with previous observations made
in 6-OHDA-hemilesioned rats4 (a neurodegeneration
model of parkinsonism). The finding that higher N/
OFQ levels were found in the CSF of PD patients
nicely complemented these preclinical data, strengthen-
ing the notion that elevated N/OFQ levels are a feature
of both experimental parkinsonism and idiopathic PD.
Further studies will be undertaken to evaluate the role
of subthalamonigral pathways in generating nigral N/
OFQ levels and, particularly, whether N/OFQ levels
will be modified by deep brain stimulation of STN.
Although the role of N/OFQ in movement control in
humans remains unknown, the striking correlation
between N/OFQ dynamics in PD patients and parkinso-
nian rodents strengthens the idea that endogenous N/
OFQ plays a pathogenic role in parkinsonism across
species and provides a strong rationale for developing
NOP receptor antagonists for PD therapy.4,6–8
Acknowledgments: This work has been supported bygrants from the Italian Ministry of the University (FIRBInternazionalizzazione no RBIN047W33) to M Morari.
Full financial disclosure: Grants, Employment.
Author Roles: 1. Research project: A. Conception, B. Or-ganization, C. Execution; 2. Statistical Analysis: A. Design,B. Execution, C. Review and Critique; 3. Manuscript: A.Writing of the first draft, B. Review and Critique; MatteoMarti: 1A, 1B, 1C, 2B, 2B, 3B; Silvio Sarubbo: 1B, 1C, 3B;Francesco Latini: 1B, 1C; Michele Cavallo: 1A, 1B, 1C, 3B;Roberto Eleopra: 1A, 1B, 1C, 3B; Sara Biguzzi: 1B, 1C;Christian Lettieri: 1B, 1C; Carlo Conti: 1B, 1C; MicheleSimonato: 1A, 1B, 1C, 3B; Silvia Zucchini: 1A, 1B, 1C, 3B;Rocco Quatrale: 1B, 1C; Mariachiara Sensi: 1B, 1C; SanzioCandeletti: 1A, 1B, 1C, 3B; Patrizia Romualdi: 1A, 1B, 1C,3B; Michele Morari: 1A, 1B, 2A, 2B, 2C, 3A, 3B.
REFERENCES
1. Mogil JS, Pasternak GW. The molecular and behavioral pharma-cology of the orphanin FQ/Nociceptin peptide and receptor fam-ily. Pharmacol Rev 2001;53:381–415.
2. Lambert DG. The nociceptin/orphanin FQ receptor: a target withbroad therapeutic potential. Nat Rev Drug Discov 2008;7:694–710.
3. Marti M, Mela F, Guerrini R, Calo G, Bianchi C, Morari M.Blockade of nociceptin/orphanin FQ transmission in rat substan-
1730 M. MARTI ET AL.
Movement Disorders, Vol. 25, No. 11, 2010

tia nigra reverses haloperidol-induced akinesia and normalizesnigral glutamate release. J Neurochem 2004;91:1501–1504.
4. Marti M, Mela F, Fantin M, et al. Blockade of nociceptin/orpha-nin FQ transmission attenuates symptoms and neurodegenerationassociated with Parkinson’s disease. J Neurosci 2005;95:9591–9601.
5. Mabrouk OS, Marti M, Morari M. Endogenous nociceptin/orphanin FQ (N/OFQ) contributes to haloperidol-inducedchanges of nigral amino acid transmission and parkinsonism: acombined microdialysis and behavioral study in naıve and nocicep-tin/orphanin FQ receptor knockout mice. Neuroscience 2010;166:40–48.
6. Marti M, Trapella C, Viaro R, Morari M. The Nociceptin/Orpha-nin FQ Receptor antagonist J-113397 and L-Dopa additivelyattenuate experimental parkinsonism through overinhibition ofthe nigrothalamic pathway. J Neurosci 2007;27:1297–1307.
7. Marti M, Trapella C, Morari M. The novel nociceptin/orphaninFQ receptor antagonist Trap-101 alleviates experimental parkin-sonism through overinhibition of the nigro-thalamic pathway:positive interaction with L-DOPA. J Neurochem 2008;107:1683–1696.
8. Viaro R, Sanchez-Pernaute R, Marti M, Trapella C, Isacson O,Morari M. Nociceptin/orphanin FQ receptor blockade attenuatesMPTP-induced parkinsonism. Neurobiol Dis 2008;30:430–438.
9. Visanji NP, de Bie RM, Johnston TH, McCreary AC, BrotchieJM, Fox SH. The nociceptin/orphanin FQ (NOP) receptor antago-nist J-113397 enhances the effects of levodopa in the MPTP-lesioned nonhuman primate model of Parkinson’s disease. MovDisord 2008;23:1922–1925.
10. Di Benedetto M, Cavina C, D’Addario C, et al. Alterations of N/OFQ and NOP receptor gene expression in the substantia nigraand caudate putamen of MPP1 and 6-OHDA lesioned rats. Neu-ropharmacology 2009;56:761–767.
11. Brown JM, Gouty S, Iyer V, Rosenberger J, Cox BM. Differen-tial protection against MPTP or methamphetamine toxicity in do-pamine neurons by deletion of ppN/OFQ expression. J Neuro-chem 2006;98:495–505.
12. Marti M, Mela F, Veronesi C, et al. Blockade of nociceptin/orphanin FQ receptor signalling in rat substantia nigra pars retic-ulata stimulates nigrostriatal dopaminergic transmission andmotor behaviour. J Neurosci 2004;24:6659–6666.
13. Kuzmin A, Bazov I, Sheedy D, Garrick T, Harper C, BakalkinG. Expression of pronociceptin and its receptor is downregulatedin the brain of human alcoholics. Brain Res 2009;11:1305(suppl):S80–S85.
14. Rocha L, Orozco-Suarez S, Alonso-Vanegas M, et al. Temporallobe epilepsy causes selective changes in mu opioid and nocicep-tin receptor binding and functional coupling to G-proteins inhuman temporal neocortex. Neurobiol Dis 2009;35:466–473.
15. Hantos MB, Szalay F, Lakatos PL, et al. Elevated plasma noci-ceptin level in patients with Wilson disease. Brain Res Bull2002;58:311–313.
16. Madsen E, Gitlin JD. Copper and iron disorders of the brain.Annu Rev Neurosci 2007;30:317–337.
17. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates.Sydney: Academic Press, 1982.
18. Ungerstedt U, Arbuthnott GW. Quantitative recording of rota-tional behavior in rats after 6-hydroxy-dopamine lesions of thenigrostriatal dopamine system. Brain Res. 1970;24:485–493.
19. Marti M, Mela F, Bianchi C, Beani L, Morari M. Striatal dopa-mine-NMDA receptor interactions in the modulation of glutamaterelease in the substantia nigra pars reticulata in vivo: oppositerole for D1 and D2 receptors. J Neurochem 2002;83:635–644.
20. Bregola G, Zucchini S, Rodi D, et al. Involvement of the neuro-peptide nociceptin/orphanin FQ in kainate seizures. J Neurosi2002;22:10030–10038.
21. Simonato M, Bregola G, Donatini A, et al. Kindled seizure-induced c-fos and prodynorphin mRNA expressions are unrelatedin the rat brain. Eur J Neurosi 1996;8:2064–2067.
22. Aparicio LC, Candeletti S, Binaschi A, et al. Kainate seizuresincrease nociceptin/orphanin FQ release in the rat hippocampusand thalamus: a microdialysis study. J Neurochem 2004;91:30–37.
23. Ploj K, Roman E, Gustavsson L, Nylander I. Basal levels andalcohol-induced changes in nociceptin/orphanin FQ, dynorphin,and enkephalin levels in C57BL/6J mice. Brain Res Bull2000;53:219–226.
24. Sanberg PR, Bunsey MD, Giordano M, Norman AB. The cata-lepsy test: its ups and downs. Behav Neurosci 1988;102:748–759.
25. Gelb DJ, Oliver E, Gilman S. Diagnostic criteria For ParkinsonDisease. Arch Neurol 1999;56:33–39.
26. Neal CR Jr, Mansour A, Reinscheid R, Nothacker HP, Civelli O,Watson SJ Jr. Localization of orphanin FQ (nociceptin) peptideand messenger RNA in the central nervous system of the rat. JComp Neurol 1999;406:503–547.
27. Norton CS, Neal CR, Kumar S, Akil H, Watson SJ. Nociceptin/orphanin FQ and opioid receptor-like receptor mRNA expressionin dopamine systems. J Comp Neurol 2002;444:358–368.
28. Lacey MG, Mercuri NB, North RA. Two cell types in rat sub-stantia nigra zona compacta distinguished by membrane proper-ties and the actions of dopamine and opioids. J Neurosci1989;9:1233–1241.
29. Yung WH, Hausser MA, Jack JJ. Electrophysiology of dopami-nergic and non-dopaminergic neurones of the guinea-pig substan-tia nigra pars compacta in vitro. J Physiol 1991;436:643–667.
30. Albin RL, Young AB, Penney JB. The functional anatomy of ba-sal ganglia disorders. Trends Neurosi 1989;12:366–375.
31. Hassani OK, Mouroux M, Feger J. Increased subthalamic neuro-nal activity after nigral dopaminergic lesion independent of disin-hibition via the globus pallidus. Neuroscience 1996;72:105–115.
32. Conner-Kerr TA, Simmons DR, Peterson GM, Terrian DM. Evi-dence for the corelease of dynorphin and glutamate from rat hip-pocampal mossy fiber terminals. J Neurochem 1993;61:627–636.
33. Cobb WS, Abercrombie ED. Relative involvement of globus pal-lidus and subthalamic nucleus in the regulation of somatoden-dritic dopamine release in substantia nigra is dopamine-depend-ent. Neuroscience 2003;119:777–786.
34. Buzas B, Symes AJ, Cox BM. Regulation of nociceptin/orphaninFQ gene expression by neuropoietic cytokines and neurotrophic fac-tors in neurons and astrocytes. J Neurochem 1999;72:1882–1889.
35. Stromberg I, Bjorklund H, Dahl D, Jonsson G, Sundstrom E,Olson L. Astrocyte responses to dopaminergic denervations by 6-hydroxydopamine and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-dine as evidenced by glial fibrillary acidic protein immunohisto-chemistry. Brain Res Bull 1986;17:225–236.
36. Raffaeli W, Samolsky DBG, Landuzzi D, et al. Nociceptin levelsin the cerebrospinal fluid of chronic pain patients with or withoutintrathecal administration of morphine. J Pain Symptom Manage2006;32:372–377.
37. Almay BG, Haggendal J, von Knorring L, Oreland L. 5-HIAAand HVA in CSF in patients with idiopathic pain disorders. BiolPsychiatry 1987;22:403–412.
38. Eklundh T, Eriksson M, Sjoberg S, Nordin C. Monoamine pre-cursors, transmitters and metabolites in cerebrospinal fluid: a pro-spective study in healthy male subjects. J Psychiatr Res 1996;30:201–208.
39. LeWitt PA, Galloway MP, Matson W, Milbury P, McDermottM, Srivastava DK, Oakes D. Markers of dopamine metabolismin Parkinson’s disease. The Parkinson Study Group Neurology1992;42:2111–2117.
40. Facchinetti F, Petraglia F, Cicero S, Nappi G, Valentini M, Genaz-zani AR. No gradient exists between lumbar and ventricular cere-brospinal fluid beta-endorphin. Neurosci Lett 1987;77:349–352.
41. Berrettini WH, Nurnberger JI Jr, Di Maggio DA. NeuropeptideY immunoreactivity in human cerebrospinal fluid Peptides. 1986;7:455–458.
42. Nutt JG, Mrox EA, Leeman SE, Williams AC, Engel WK, ChaseTN. Substance P in human cerebrospinal fluid: reductions in pe-
1731N/OFQ LEVELS IN PARKINSON’S DISEASE
Movement Disorders, Vol. 25, No. 11, 2010

ripheral neuropathy and autonomic dysfunction. Neurology 1980;30:1280–1285.
43. Abbott NJ. Evidence for bulk flow of brain interstitial fluid: sig-nificance for physiology and pathology. Neurochem Int 2004;45:545–552.
44. Nothacker HP, Reinscheid RK, Mansour A, et al. Primary struc-ture and tissue distribution of the orphanin FQ precursor. ProcNatl Acad Sci U S A 1996;93:8677–8682.
45. Witta J, Palkovits M, Rosenberger J, Cox BM. Distribution ofnociceptin/orphanin FQ in adult human brain. Brain Res 2004;997:24–29.
46. Johanson CE, Duncan JA III, Klinge PM, Brinker T, Stopa EG,Silverberg GD. Multiplicity of cerebrospinal fluid functions: Newchallenges in health and disease. Cerebrospinal Fluid Res2008;14:5–10.
47. Candeletti S, Ferri S. Effects of an antisense oligonucleotide topronociceptin and long-term prevention of morphine actions bynociceptin. Peptides 2000;21:1119–1124.
48. Marti M, Viaro R, Guerrini R, Franchi G, Morari M. Nociceptin/orphanin FQ modulates motor behavior and primary motor cortexoutput through receptors located in substantia nigra reticulata.Neuropsychopharmacology 2009;34: 341–355
Movement Disorders, Vol. 25, No. 11, 2010
1732 M. MARTI ET AL.
Related Documents











