Endogenous nociceptin modulates diet preference independent of motivation and reward Miwako Koizumi 1 , Barbara Cagniard, Niall P. Murphy ⁎ Molecular Neuropathology Group, RIKEN Brain Science Institute, Wakoshi, Saitama 351-0198, Japan abstract article info Article history: Received 12 September 2008 Received in revised form 4 December 2008 Accepted 10 December 2008 Keywords: Opioid Nociceptin Orphanin FQ Feeding Reward Motivation Conditioned place preference Palatable Previous studies show that the opioid peptide nociceptin stimulates food intake. Here, we studied nociceptin receptor knockout (NOP KO) mice in various behavioral paradigms designed to differentiate psychological and physiological loci at which endogenous nociceptin might control feeding. When presented a choice under food restriction, NOP KO mice displayed reduced preference for high sucrose diet, but lower intake of high fat diet under no-choice conditions. These responses were absent under ad libitum feeding conditions. Conditioned place preference to high fat diet under food-deprived conditions was unaltered in NOP KO mice, suggesting no difference in reward responses. Furthermore, operant food self-administration under a variety of conditions showed no genotype-dependent differences, suggesting no differences in the motivational properties of food. Taste reactivity to sucrose was unchanged in NOP KO mice, though NOP KO mice had altered aversive reactions to quinine solutions under ad libitum feeding, suggesting minor differences in the affective impact of palatable and unpalatable tastants. Although NOP KO mice re-fed following food- deprivation showed normal increases in plasma glucose and insulin, multidimensional scaling analysis showed that the relationship between these measures, body weight and plasma leptin was substantially disrupted in NOP KO, particularly in fasted mice. Additionally, the typical positive relationship between body weight and plasma leptin was considerably weaker in NOP KO mice. Together, these findings suggest that endogenous nociceptin differentially modulates diet preference depending on macronutrient content and homeostatic state, independently of the motivating, rewarding or orosensory properties of food, but may involve metabolic or postingestive processes. © 2008 Elsevier Inc. All rights reserved. 1. Introduction Changes in dietary patterns, particularly due to increases in readily available foods that are both cheap and have high sugar and fat contents, are believed to be a major factor underlying the increasing prevalence of obesity in Western nations [1]. Amongst the multitude of neurochemical systems involved in regulating feeding behavior, endogenous opioid peptides have been well established [2,3]. The actions of endogenous opioids may be due to effects on fundamental post-ingestive mechan- isms regulating food intake, or effects on higher psychological processes such as motivation and reward (see [2,4,5]). For instance, endogenous opioid peptides such as endorphin and enkephalin have well-estab- lished roles in mediating the rewarding properties of stimuli such as addictive drugs. Likewise, numerous studies suggest that endogenous opioids mediate the rewarding properties of palatable foods, such as those high in sugar or fat content [6]. Nociceptin (also known as orphanin FQ, abbreviated here to N/OFQ) is a member of the endogenous opioid family (see [7]). Similar to other endogenous opioids, N/OFQ is implicated in rewarded and reinforced behaviors (see [8]). Previous work has led to the hypothesis that N/OFQ generally inhibits reward processes and drug-seeking behavior (see [9] for references), possibly by suppressing activity in neurochemical systems mediating incentive motivation [10] or the hedonic impact of stimuli [11]. Like many neuropeptides, N/OFQ is particularly concen- trated in brain areas controlling food intake such as the hypothalamus [12,13]. When administered intracerebroventricularly (i.c.v.), N/OFQ stimulates food intake [14,15]. Specifically, it appears that activation of the N/OFQ receptor (termed the “NOP” receptor) initiates and maintains feeding behavior, though a role for N/OFQ in inhibiting processes terminating feeding behavior has also been suggested [16]. Virtually all studies to date have investigated the effects of exogenous N/OFQ on feeding-related behavior. Thus, the purpose of the current study was to seek evidence of a physiological role of endogenous N/OFQ in feeding behavior, particularly in modulating the motivating and reward- ing properties of food, by studying NOP receptor KO mice (NOP KO). This is particularly topical as obesity rates are increasing in human Physiology & Behavior 97 (2009) 1–13 Abbreviations: N/OFQ, nociceptin/orphanin FQ. ⁎ Corresponding author. Tel.: +81 48 467 7126. E-mail address: [email protected] (N.P. Murphy). 1 Current address: Shimojo Implicit Brain Function Project, ERATO, Japan Science and Technology Agency, NTT Atsugi Research and Development Center, Kanagawa 243-0198, Japan. 0031-9384/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.physbeh.2008.12.008 Contents lists available at ScienceDirect Physiology & Behavior journal homepage: www.elsevier.com/locate/phb

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Physiology & Behavior 97 (2009) 1–13
Contents lists available at ScienceDirect
Physiology & Behavior
j ourna l homepage: www.e lsev ie r.com/ locate /phb
Endogenous nociceptin modulates diet preference independent of motivationand reward
Miwako Koizumi 1, Barbara Cagniard, Niall P. Murphy ⁎Molecular Neuropathology Group, RIKEN Brain Science Institute, Wakoshi, Saitama 351-0198, Japan
Abbreviations: N/OFQ, nociceptin/orphanin FQ.⁎ Corresponding author. Tel.: +81 48 467 7126.
E-mail address: [email protected] (N.P. Murphy).1 Current address: Shimojo Implicit Brain Function Pro
Technology Agency, NTT Atsugi Research and DevelopmeJapan.
0031-9384/$ – see front matter © 2008 Elsevier Inc. Adoi:10.1016/j.physbeh.2008.12.008
a b s t r a c t
a r t i c l e i n f oArticle history:
Previous studies show that Received 12 September 2008Received in revised form 4 December 2008Accepted 10 December 2008Keywords:OpioidNociceptinOrphanin FQFeedingRewardMotivationConditioned place preferencePalatable
the opioid peptide nociceptin stimulates food intake. Here, we studied nociceptinreceptor knockout (NOP KO) mice in various behavioral paradigms designed to differentiate psychologicaland physiological loci at which endogenous nociceptin might control feeding. When presented a choiceunder food restriction, NOP KO mice displayed reduced preference for high sucrose diet, but lower intake ofhigh fat diet under no-choice conditions. These responses were absent under ad libitum feeding conditions.Conditioned place preference to high fat diet under food-deprived conditions was unaltered in NOP KO mice,suggesting no difference in reward responses. Furthermore, operant food self-administration under a varietyof conditions showed no genotype-dependent differences, suggesting no differences in the motivationalproperties of food. Taste reactivity to sucrose was unchanged in NOP KO mice, though NOP KO mice hadaltered aversive reactions to quinine solutions under ad libitum feeding, suggesting minor differences in theaffective impact of palatable and unpalatable tastants. Although NOP KO mice re-fed following food-deprivation showed normal increases in plasma glucose and insulin, multidimensional scaling analysisshowed that the relationship between these measures, body weight and plasma leptin was substantiallydisrupted in NOP KO, particularly in fasted mice. Additionally, the typical positive relationship between bodyweight and plasma leptin was considerably weaker in NOP KO mice. Together, these findings suggest thatendogenous nociceptin differentially modulates diet preference depending on macronutrient content andhomeostatic state, independently of the motivating, rewarding or orosensory properties of food, but mayinvolve metabolic or postingestive processes.
© 2008 Elsevier Inc. All rights reserved.
1. Introduction
Changes in dietary patterns, particularly due to increases in readilyavailable foods that are both cheap and have high sugar and fat contents,are believed to be amajor factor underlying the increasing prevalence ofobesity inWesternnations [1]. Amongst themultitude of neurochemicalsystems involved in regulating feeding behavior, endogenous opioidpeptides have been well established [2,3]. The actions of endogenousopioids may be due to effects on fundamental post-ingestive mechan-isms regulating food intake, or effects on higher psychological processessuch as motivation and reward (see [2,4,5]). For instance, endogenousopioid peptides such as endorphin and enkephalin have well-estab-lished roles in mediating the rewarding properties of stimuli such asaddictive drugs. Likewise, numerous studies suggest that endogenous
ject, ERATO, Japan Science andnt Center, Kanagawa 243-0198,
ll rights reserved.
opioids mediate the rewarding properties of palatable foods, such asthose high in sugar or fat content [6].
Nociceptin (also known as orphanin FQ, abbreviated here to N/OFQ)is a member of the endogenous opioid family (see [7]). Similar to otherendogenous opioids, N/OFQ is implicated in rewarded and reinforcedbehaviors (see [8]). Previouswork has led to the hypothesis that N/OFQgenerally inhibits reward processes and drug-seeking behavior (see [9]for references), possibly by suppressing activity in neurochemicalsystems mediating incentive motivation [10] or the hedonic impact ofstimuli [11]. Like many neuropeptides, N/OFQ is particularly concen-trated in brain areas controlling food intake such as the hypothalamus[12,13]. When administered intracerebroventricularly (i.c.v.), N/OFQstimulates food intake [14,15]. Specifically, it appears that activation oftheN/OFQ receptor (termed the “NOP” receptor) initiates andmaintainsfeeding behavior, though a role for N/OFQ in inhibiting processesterminating feeding behavior has also been suggested [16].
Virtually all studies to date have investigated the effects of exogenousN/OFQ on feeding-related behavior. Thus, the purpose of the currentstudywas to seek evidence of aphysiological role of endogenousN/OFQ infeeding behavior, particularly in modulating the motivating and reward-ing properties of food, by studying NOP receptor KO mice (NOP KO).This is particularly topical as obesity rates are increasing in human

Table 1Composition of laboratory-made diets used in consumption studies under food-deprived and ad libitum feeding conditions.
Constituent Normal High sucrose High fat
Casein (milk) 20 20 20Cornstarch 59.8 14.8 34.8Sucrose 5 50 5Corn oil 5 5 5Shortening 0 0 25Mineral mix AIN93G 4 4 4Vitamin mix AIN93G 1 1 1Cellulose 5 5 5L-cystine 0.18 0.18 0.183-butyl-hydroquinone 0.0008 0.0008 0.0008kcal/100 g 384.2 384.2 509.2
2 M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
populations, and targeting the N/OFQ-NOP receptor system may bebeneficial in managing pathological feeding behavior [17]. Thus, westudied NOP KOmice in a battery of behavioral responses to high sucroseor high fat diets (which presumably have different palatabilities)designed to assess these psychological processes. Furthermore, as fooddeprivation increases the motivation to consume, and the palatability offood [18], many of the responses we studied were compared under free-feeding and food-deprived conditions. We predicted that NOP KO micewould generally consume more of, and prefer palatable diets (i.e. thosewithhigh sugaror fat contents) andworkharder toobtain them,basedonthe prediction that such dietswould bemore rewarding in the absence ofendogenous N/OFQ effects [9]. However, we found that NOP KO micegenerally consumed less, not more, of high sucrose and high fat diets,though only when food deprived. Furthermore, NOP KO mice did notdiffer in any behavioral tests aimed at identifying differences inmotivation or reward, suggesting that the observed effects were duepredominantly to an effect of endogenous N/OFQ on post-ingestiveprocesses.
2. Methods
2.1. Animals
Experimental protocols used throughout the study were approvedby the institutional review committee and were in accord with NIHethics guidelines. Adult male wild-type or NOP KO mice (aged 9 to30weeks, see specific experiments for details), maintained on a C57BL6background, were generated by mating heterozygous male and femalepairs from a colony maintained in the RIKEN Brain Science Instituteanimal facility from a previously described NOP KO line [19] kindlyprovided by Prof. Hiroshi Takeshima of Tohoku University. Duringbreeding and maintenance in the colony, mice were housed in cagesamongst same generation siblings of mixed genotypes in groups of twoto five animals per cage under a 12 h:12 h light/dark cycle (lights on8:00), where they received standard lab chow (CRF-1, Charles River,Wilmington, MA) and water ad libitum. Mice were habituated to singlehousing for two to seven days before the start of the testing protocol. Allbehavioral experiments were conducted between 13:00 and 19:00. Ourprevious studies show that these mice differ in several reward-relatedresponses [9], and are insensitive to the suppressive effect of N/OFQ onmidbrain dopamine activity [10]. Furthermore, i.c.v. administration of3 nmol ofN/OFQ towild-type satiatedmice stimulates a robust intake oflaboratory chow, whereas this effect is absent in NOP KO mice(unpublished observation), confirming the absence of the NOP receptorgene and the C57BL6 strain as a reliable model.
2.2. Experiment 1: Voluntary feeding under food-deprived conditions anddrinking behavior under free-feeding conditions
To assess the role of endogenous N/OFQ in food consummatorybehavior, voluntary feeding of high sucrose (50% sucrose) and high fat(30% fat) diet was monitored under food-deprived conditions. The
composition of the diets used for these studies are shown in Table 1.Additionally, the role of endogenous N/OFQ in drinking of solutions ofsucrose, saccharin and oil was evaluated using a two-bottle choicedrinking test. By comparing preference for sweet solutions that eithercontained (sucrose) or were deficient (saccharin) in calorific value, weaimed to distinguish post-gustatory effects.
Mice (12 per genotype) aged 19 to 30 weeks were used in thisstudy. Feeding experiments and two-bottle choice drinking tests wereconducted in the home cage and lasted 4 h starting from 15:00 h. Forthe feeding experiment, micewere food deprived from 19:00 onwardsthe day before the start of intake tests. Body weight was measuredprior to each test session. Animals were presented an excess of diets inplastic cups (4.5 cm in diameter) placed on the floor of the cage. Theposition of cups was randomized to avoid place bias. During the 4 hexperiment, mice did not have access to the normal chow but waterwas provided ad libitum throughout the whole experiment. Thefollowing parameters were recorded: 1) energy (food) intake,expressed as kcal per 100 g of body weight, 2) percentage of originalbody weight and 3) preference for high sucrose or high fat diet,calculated as (the amount of ingestant consumed/total amount ofboth ingestants consumed)×100%.
For the two-bottle choice test, sucrose (0.3 and 1.5 M, respectively10 and 50% sucrose; Oriental Yeast Co.), saccharin (0.003 and 0.006M;Sigma) and corn oil (100%; Ajinomoto Co., Tokyo, Japan) werepresented versus distilled water in graduated 15 ml tubes equippedwith a sipper for 4 h, under ad libitum feeding. Sucrose and saccharinwere presented in an increasing concentration order to avoid thedevelopment of a preference for the highest concentration beforepresentation of the lowest concentration. Solution preference wascalculated as (the amount of ingestant consumed/total amount ofboth ingestants consumed)×100%.
The testing sequence was as follows: Mice were shifted toalternating presentation of normal and high sucrose diets for sixdays (starting with the normal diet) followed by a choice test ofnormal versus high sucrose diets for three days. Under ad libitumfeeding conditions, mice were habituated to ad libitum drinking fromtwo graduated cylinders located in the roof of the home cage for5 days. Drinking tests under 20 h fluid restriction were conducted onfour consecutive days in the following order: 0.003 M saccharin, 0.3 Msucrose, 0.006 M saccharin and 1.5 M sucrose. Following eight days ofrecovery, a similar procedure was repeated with normal and high fatdiets. Two-bottle choice oil drinking was conducted on twoconsecutives days and data averaged for analysis.
2.3. Experiment 2: Food consumption and food preference undernon-food-deprived conditions
These studies were similar to those of Experiment 1 with theexception that mice were not food deprived and that no study ofdrinking behavior was incorporated in the protocol. Mice (8 wild-typeand 7 NOP KO) aged 21 to 28 weeks were used for this study. Basalbody weight and food consumption were assessed for five days priorto beginning the experiment. Mice were then shifted to alternatingpresentation of normal and high sucrose diets for 4 h per day for sixdays followed by a choice test of normal versus high sucrose diets forthree days as described above. Following six days on 24 h access tonormal lab chow, the same procedure was repeated with normal andhigh fat diets. During the 4 h diet intake or choice tests, mice did nothave access to the normal chow.
2.4. Experiment 3: Taste reactivity to palatable and unpalatable tastants
Studies by Berridge and colleagues suggest that the microstructureof taste reactions induced by intraoral infusion of taste stimuli directlyreflect the affective aspects of these taste stimuli (i.e. their palatability,[20]). Thus, taste reactions were studied in NOP KO mice by observing

3M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
orofacial reactions in response to experimenter-delivered sweet(sucrose) and bitter (quinine) solutions. These reactions were thencategorized as being positive (i.e. indicative of a positive affectiveimpact) or aversive (i.e. indicative of a negative affective impact).
Mice (8 wild-type and 7 NOP KO) aged 10 to 14 weeks were usedfor this study. Mice were anesthetized by intraperitoneal injection ofketamine (100 mg/kg) and xylazine (10 mg/kg), and were implantedwith chronic bilateral oral cannula (polyethylene PE-50 tubing). Theintraoral end of the cannula was heat-flared to an approximatediameter of 3 mm. A piece of 1.2 mm diameter stainless tubingsharpened at one end was used to insert cannulae lateral to the firstmaxillary molar. Cannulae were routed subcutaneously posterior tothe eye, to exit the top of the head where they were fixed to a screwanchored to the skull using dental cement (Yamahachi Dental,Gamagoorishi, Japan). To avoid obstruction of cannulae during therecovery period, a silk thread was inserted and the loose end of thecannula was heat-sealed until the start of infusions. Mice receivedpostoperative penicillin (0.1 ml; 10,000 U) and were allowed at least7 days of recovery.
The taste reactivity observation chamber was a Plexiglas cylinder(30 cm high×13 cm diameter). A mirror located beneath atransparent Plexiglas floor reflected a ventral view of the mouse andallowed videotaping of the mouth of the animal using a digital videocamera (DCR PC101, Sony Corporation, Japan) directed at the mirror.Mice were habituated to the testing chamber for 20 mins for threedays before testing and habituated to the taste reactivity procedurewith three oral infusions of drinking water on the last two days ofhabituation.
For taste reactivity testing, a delivery line constructed of PE-50polyethylene tubing was connected to one of the oral cannulae by apolyethylene PE-100 nozzle and connected to a 1 ml syringe mountedon an automated syringe pump (Harvard Apparatus, Holliston, MA).The entire length of tubing was filled with the appropriate solutionand a small amount of solution was infused into the mouth of theanimal to verify the patency of the cannula. Mice were adapted to thetest chamber for 7 min. They then received three infusions, 7 minapart. Each infusion was 0.1 ml in volume and lasted 1 min, duringwhich time mice were videotaped.
Mice were tested with three concentrations of sucrose (0.01, 0.1and 1 M in ascending order) and quinine (0.03, 0.3 and 3 mM inascending order) under ad libitum feeding conditions and food-deprivation conditions. The order of presentation of tastants wasrandom, but identical between feeding conditions. Tastants werepresented individually on two consecutive days under each feedingcondition, interspersed by three days for establishing fooddeprivation.
Behavioral analysis of taste reactions was performed frame-by-frame (30 frames/s) by an observer blind to treatment using videoediting software (Final Cut Pro, Apple Inc, Cupertino, CA). Scoring wasperformed manually and recorded using software for recordingobservational data (ODlog, Macropod Software, http://www.macro-podsoftware.com/). Categorization of responses was adapted from[20] and [21]. Positive reactions scored were rhythmic tongueprotrusion (rhythmic extension of the tongue), non-rhythmic tongueprotrusions (single lateral or midline extension of the tongue) andpaw licking. Aversive reactions recorded were gapes (large opening ofthe mouth with a retraction of the lower lip), forelimb flails (rapidwaving of one or both forelimb), headshakes (rapid lateral movementof the head), facewiping (wipes over the facewith the paws) and chinrubbing (pushing the chin against the wall or the floor of the testchamber). To be sure that each component made an equal contribu-tion to the final positive, aversive or neutral scores, reactions thatoccurred in continuous bouts were scored in time bins. Componentscharacterized by bouts of moderate duration, such as rhythmic tongueprotrusion and chin rubbing, were scored in 2 s bins (continuousrepetitions within 2 s scored as one occurrence). Components that
typically have longer bout durations, such as paw licking and facewipingwere similarly scored in 5 s bins. Other reactions that can occuras single behaviors were scored as separate occurrences (non-rhythmic tongue protrusions, gapes, forelimb flails, headshakes).These time bins help equate scores of different reactions within aresponse category, so that more perseverative reactions do notobscure rare but informative reactions.
2.5. Experiment 4: Food self-administration using an operant apparatus
In order to study the influence of endogenous N/OFQ onmotivation to obtain food, mice were tested under two reinforcementschedules, as well as two feeding conditions (food restricted or adlibitum) for each schedule of reinforcement in a food self-administra-tion apparatus. They were first tested under a progressive ratio (PR)schedule for presentation of either control (20 mg pellet normalgrain-based diet, formula 5TUM, 3.26 kcal/g, TestDiet, Richmond, IN,USA) or sucrose pellets (formula 5TUT, 3.56 kcal/g). The PR schedule,in which the instrumental response requirement to obtain reinforce-ment is gradually increased throughout the session, measures howhard animals are willing to work for food reward [22]. The strength ofmotivation is determined by the highest ratio (breakpoint) that theanimal sustains. Then, similar to the voluntary food preference studydescribed above, mice were presented a choice of two levers thatdelivered control or sucrose pellets during the same session under afixed ratio (FR) schedule of reinforcement, whereby animals arerequired to lever press a constant number of times to obtain foodreward. The purpose of this experiment was to evaluate theinvolvement of effort on modulation of sucrose preference byendogenous N/OFQ.
For these experiments, mice operant conditioning chambers(O'Hara and Co., Tokyo, Japan) were used (dimensions,19×10×10 cm, W×D×H) housed in sound attenuating boxes. Aretractable lever and food receptacle were located at both ends of thechambers. Lever pressing and food delivery were synchronized andrecorded using Med PC software (MedAssociates, St. Albans, VT).Throughout the experiment, food self-administration sessions lasted1 h or until a subject obtained 50 rewards (pellets) on FR schedules. Inthe case of PR schedule testing, sessions terminated either auto-matically after 90 min or whenever a 5 min unrewarded periodoccurred. Mice (7 per genotype), aged 12 to 19 weeks, were tested fivedays aweek and results are expressed as the average of the five days oftesting for each condition.
2.5.1. FR1 trainingUnder food-deprivation (10–15% reduction of baseline weight),
mice were first trained under a FR1 schedule of reinforcement withboth levers extended but only presses on one of the levers (activelever) were reinforced by the delivery of control diet pellets. Duringthe first two days of training, food pellets were also delivered into thefood receptacle on a variable time 60 s schedulewith intervals rangingbetween 0 and 120 s. When mice reached a criterion of at least 40pellets earned and a percentage of lever pressing on the active lever of70% on two consecutive days, they were shifted to a PR schedule ofreinforcement. A shaping procedure was applied to two mice (one pergenotype) that had difficulty learning the task.
2.5.2. PR scheduleMice were initially tested under a PR5 schedule for three days. In
this schedule, the first lever press is reinforced, and then the numberof presses required to obtain a reward is increased by five followingeach reinforcement. However, as the number of rewards obtainedwith a PR5 schedule was unusually low (less than 10 rewards persession), mice were shifted to a PR2 schedule.
Under food-deprivation (10–15% reduction of baseline weight),mice were tested on PR2 for one week with control pellets, and then

4 M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
one week with sucrose pellets. At the end of this testing period, micewere returned to ad libitum access to food and 10 days later, testedagain for one week on a PR2 schedule with control pellets as a rewardfollowed by one week of testing with the sucrose pellets. Theparameters recorded were the breakpoint (defined as the last ratiocompleted before mice ceased pressing the lever), the number ofpresses on the inactive lever as a measure of general activity, and thepercentage of original body weight as a result of food-deprivation.
2.5.3. Choice procedureThe choice procedure started two weeks after the end of the PR
experiment. One NOP KO mouse was removed from the experimentdue to illness. In this procedure, mice had the choice to lever press toobtain a control pellet on one side of the chamber, or lever press toobtain a sucrose pellet on the opposite side of the chamber. Thelocations of the control and sucrose pellets were randomized betweenmice. The schedule of reinforcement was FR3 for each lever. Before thebeginning of the choice testing, under food-deprivation, mice receivedseparate training sessions on each lever (one session per day, only onelever extended at a time). There were four presentation sessions perlever with the following schedule of reinforcement: one session onFR1, one session on FR2 and two sessions on FR3. Mice were thentested on the choice procedure first for two weeks under food-deprivation, as well as for another week after 10 days of ad libitumfood access. The percentage of sucrose preference was calculated(number of sucrose reward obtained/total number of rewardobtained)×100%, as well as the percentage of food-deprivation.
2.6. Experiment 5: Conditioned place preference for high fat food underfood-deprivation
The conditioned place preference procedure provides a robustmeasure of the rewarding effects of unconditioned stimuli such asfood [23]. In this paradigm, the opportunity to consume food is pairedwith distinctive environmental cues, leading the animal to form alearned association. When tested in the absence of food, animalsvoluntarily approach those cues, thus expressing a “preference”indicative of the associated rewarding properties. Here, the strengthof conditioned place preference to access to a high fat diet in a food-deprived state was compared between wild-type and NOP KO mice.
Mice (10 wild-type and 7 NOP KO) aged 9 to 17 weeks were usedfor this study. Mice were tested for the establishment of placeconditioning to presentation of a high fat diet in a food-deprived state(target level 85% of original body weight) that started four days priorto the first day of the place conditioning procedure and continuedthroughout the remainder of the experiment. A commerciallyavailable 26.6% fat formula (26.7% protein, 46.7% carbohydrate,4.73 kcal/g) designed by Research Diets (New Brunswick, NJ) wasused. The place conditioning apparatus was a two compartmentdesign consisting of 25×25×20 cm (W×D×H) locomotor activitymonitoring boxes divided into two equal sized compartments(25×12.5×20 cm) by a divider variably containing a 5×5 cm doorin the center of the base. One compartment had a vertical stripespattern and a metal floor with small holes. The other compartmenthad a horizontal stripes pattern, and a metal floor with larger holes.Horizontal locomotor activity and location within the box wasautomatically recorded throughout all sessions by locomotor activitymonitors (Truscan, Coulbourn Instruments, Allentown, PA, USA) thatmeasured locomotion by breaking of infrared photosensors (16×16array, covering an area of 25×25 cm). On the day prior toconditioning, all animals were tested to determine any individualinitial preference for either of the two compartments when given freeaccess for 20 min (termed “pretest”). Conditioning started thefollowing day using an unbiased experimental design. On the firstconditioning day, micewere confined to one of the two compartmentswithout food for 40 min. Mice were then returned to the home cage
and the daily food ration provided at least 1 h later. The following day,each animal was confined to the opposite compartment to that of theprevious day for 40 minwith excess of commercially supplied high fatdiet. No additional food was given in the home cage. This process wasrepeated three more times such that mice received a total of eightconditioning sessions (four without food, four with food). Preferenceto the food-paired environment was tested the following day (termed“test”) during a 20 min session identical to the pretest conditions.Establishment of place conditioning was defined as a statisticallysignificant increase in time spent between the pretest and test in thecompartment in which mice received food. Food intake in the placeconditioning apparatus and percentage of original body weight as aresult of food deprivation were also calculated.
2.7. Experiment 6: Effect of re-feeding on blood measures following fooddeprivation
This experiment was designed to seek endocrine and biochemicalsupport for a role of endogenous N/OFQ in modulating voluntaryconsumption of high sucrose or high fat diets following a period offood deprivation. Mice (six per genotype and feeding condition) usedin this part of the study were those used for studying the effects ofvarious diets on food intake under food-deprivation conditions(Experiment 1). These mice were allowed 10 days to recover fromthe previous food deprivation periods and bottle choice testing. Oneday before the experiment, mice were food-deprived from 19:30 honwards. At 13:00 to 18:00 h (randomized across genotype andfeeding group) on the following day, animals were weighed andpresented with an excess of either normal, high sucrose or high fatdiet in their home cages. Additional groups of mice were presented nofood, and thus remained fasted. Thirty minutes later, food consumedwas measured, mice were sacrificed under halothane anesthesia andblood immediately aspirated from the heart and cooled on ice. Bloodglucose content was immediately measured using a commercialmonitor (Accu-Chek, Roche, Japan), and the remaining blood samplewas placed in a heparinized tube, centrifuged (5000 rpm,15min, 4 °C)and plasma stored at −80 °C until assay for insulin and leptin usingcommercially availably ELISA kits (Morinaga Institute of BiologicalScience, Kanagawa Japan).
2.8. Data analysis
Genotypes were compared by Student's t-test or repeatedmeasures analysis of variance (ANOVA) when appropriate. In thecase of significant main effects found by ANOVA, specific groups ortimes were compared using Student's t-test as post-hoc analysis. Dataare expressed as the mean±standard error of the mean throughoutthe manuscript. Statistical analyses were performed using StatViewsoftware (Statview, Berkley, CA, USA). p values less than 0.05 wereconsidered statistically significant. Correlations between biochemicaland behavioral measures were determined using a Spearman rankcorrelation. Relationships between measures in Experiment 6 weredetermined by multidimensional scaling analyses performed usingMatlab software (The Mathworks, Natick, MA, USA) and plotted on atwo dimensional plane.
3. Results
3.1. Experiment 1: Voluntary feeding under food-deprived conditions anddrinking behavior under free-feeding conditions
As shown in Fig. 1a, both NOP KO and wild-type mice showed aprogressive increase in intake of normal and high sucrose diets whenpresented on alternating days under non-choice conditions (genotype:F1, 22=0.12, p=0.73; time: F5, 110=96.33; pb0.0001). Notably,abrupt increases in intake were recorded each time normal diet
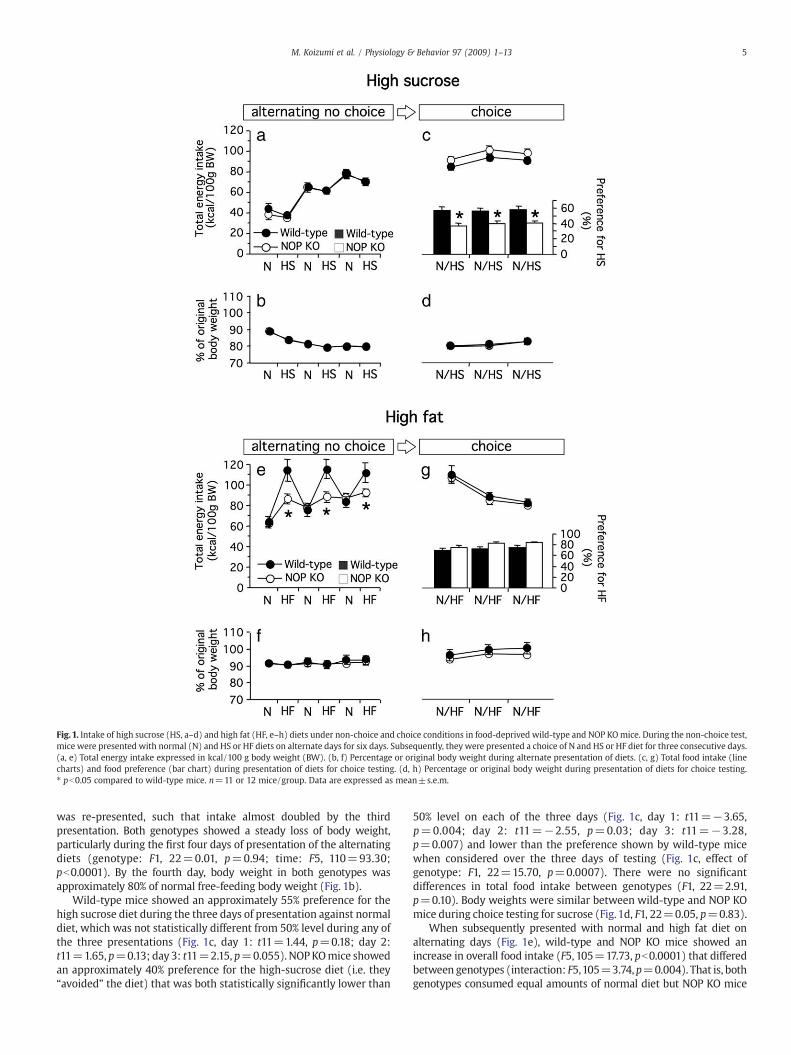
Fig. 1. Intake of high sucrose (HS, a–d) and high fat (HF, e–h) diets under non-choice and choice conditions in food-deprived wild-type and NOP KOmice. During the non-choice test,mice were presented with normal (N) and HS or HF diets on alternate days for six days. Subsequently, they were presented a choice of N and HS or HF diet for three consecutive days.(a, e) Total energy intake expressed in kcal/100 g body weight (BW). (b, f) Percentage or original body weight during alternate presentation of diets. (c, g) Total food intake (linecharts) and food preference (bar chart) during presentation of diets for choice testing. (d, h) Percentage or original body weight during presentation of diets for choice testing.⁎ pb0.05 compared to wild-type mice. n=11 or 12 mice/group. Data are expressed as mean±s.e.m.
5M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
was re-presented, such that intake almost doubled by the thirdpresentation. Both genotypes showed a steady loss of body weight,particularly during the first four days of presentation of the alternatingdiets (genotype: F1, 22=0.01, p=0.94; time: F5, 110=93.30;pb0.0001). By the fourth day, body weight in both genotypes wasapproximately 80% of normal free-feeding body weight (Fig. 1b).
Wild-type mice showed an approximately 55% preference for thehigh sucrose diet during the three days of presentation against normaldiet, which was not statistically different from 50% level during any ofthe three presentations (Fig. 1c, day 1: t11=1.44, p=0.18; day 2:t11=1.65, p=0.13; day 3: t11=2.15, p=0.055). NOPKOmice showedan approximately 40% preference for the high-sucrose diet (i.e. they“avoided” the diet) that was both statistically significantly lower than
50% level on each of the three days (Fig. 1c, day 1: t11=−3.65,p=0.004; day 2: t11=−2.55, p=0.03; day 3: t11=−3.28,p=0.007) and lower than the preference shown by wild-type micewhen considered over the three days of testing (Fig. 1c, effect ofgenotype: F1, 22=15.70, p=0.0007). There were no significantdifferences in total food intake between genotypes (F1, 22=2.91,p=0.10). Body weights were similar between wild-type and NOP KOmice during choice testing for sucrose (Fig. 1d, F1, 22=0.05, p=0.83).
When subsequently presented with normal and high fat diet onalternating days (Fig. 1e), wild-type and NOP KO mice showed anincrease in overall food intake (F5, 105=17.73, pb0.0001) that differedbetween genotypes (interaction: F5,105=3.74,p=0.004). That is, bothgenotypes consumed equal amounts of normal diet but NOP KO mice
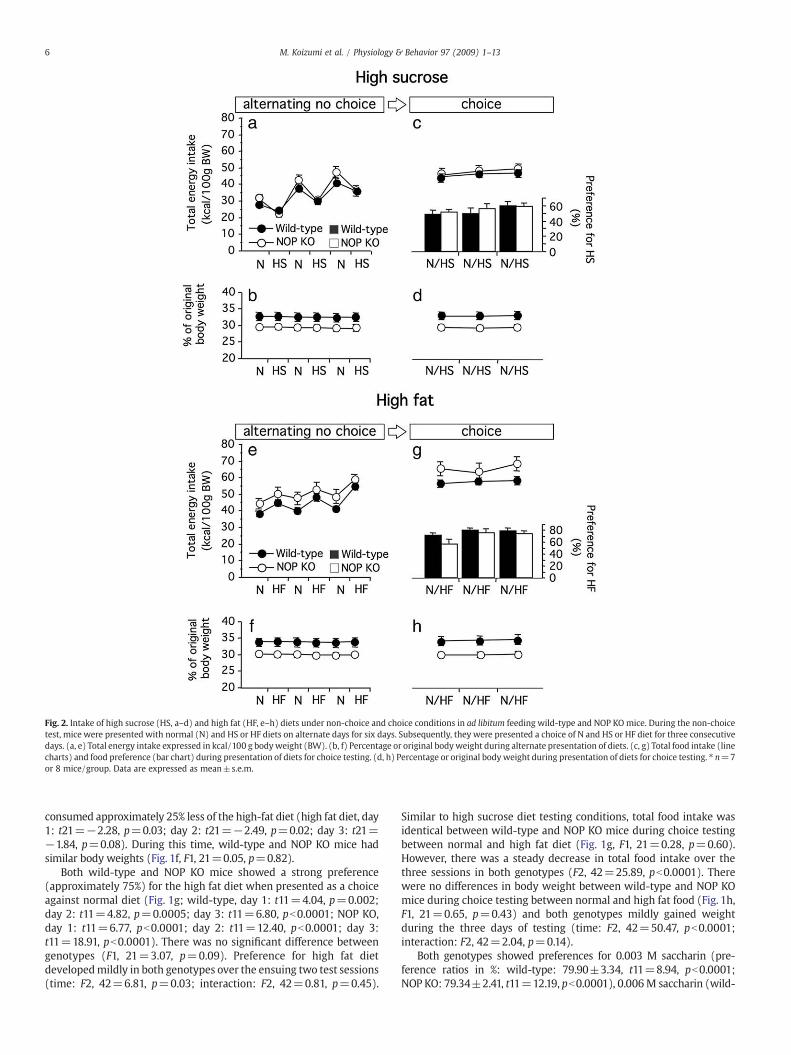
Fig. 2. Intake of high sucrose (HS, a–d) and high fat (HF, e–h) diets under non-choice and choice conditions in ad libitum feeding wild-type and NOP KOmice. During the non-choicetest, mice were presented with normal (N) and HS or HF diets on alternate days for six days. Subsequently, they were presented a choice of N and HS or HF diet for three consecutivedays. (a, e) Total energy intake expressed in kcal/100 g body weight (BW). (b, f) Percentage or original body weight during alternate presentation of diets. (c, g) Total food intake (linecharts) and food preference (bar chart) during presentation of diets for choice testing. (d, h) Percentage or original body weight during presentation of diets for choice testing. ⁎ n=7or 8 mice/group. Data are expressed as mean±s.e.m.
6 M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
consumed approximately 25% less of the high-fat diet (high fat diet, day1: t21=−2.28, p=0.03; day 2: t21=−2.49, p=0.02; day 3: t21=−1.84, p=0.08). During this time, wild-type and NOP KO mice hadsimilar body weights (Fig. 1f, F1, 21=0.05, p=0.82).
Both wild-type and NOP KO mice showed a strong preference(approximately 75%) for the high fat diet when presented as a choiceagainst normal diet (Fig. 1g; wild-type, day 1: t11=4.04, p=0.002;day 2: t11=4.82, p=0.0005; day 3: t11=6.80, pb0.0001; NOP KO,day 1: t11=6.77, pb0.0001; day 2: t11=12.40, pb0.0001; day 3:t11=18.91, pb0.0001). There was no significant difference betweengenotypes (F1, 21=3.07, p=0.09). Preference for high fat dietdevelopedmildly in both genotypes over the ensuing two test sessions(time: F2, 42=6.81, p=0.03; interaction: F2, 42=0.81, p=0.45).
Similar to high sucrose diet testing conditions, total food intake wasidentical between wild-type and NOP KO mice during choice testingbetween normal and high fat diet (Fig. 1g, F1, 21=0.28, p=0.60).However, there was a steady decrease in total food intake over thethree sessions in both genotypes (F2, 42=25.89, pb0.0001). Therewere no differences in body weight between wild-type and NOP KOmice during choice testing between normal and high fat food (Fig. 1h,F1, 21=0.65, p=0.43) and both genotypes mildly gained weightduring the three days of testing (time: F2, 42=50.47, pb0.0001;interaction: F2, 42=2.04, p=0.14).
Both genotypes showed preferences for 0.003 M saccharin (pre-ference ratios in %: wild-type: 79.90±3.34, t11=8.94, pb0.0001;NOP KO: 79.34±2.41, t11=12.19, pb0.0001), 0.006M saccharin (wild-
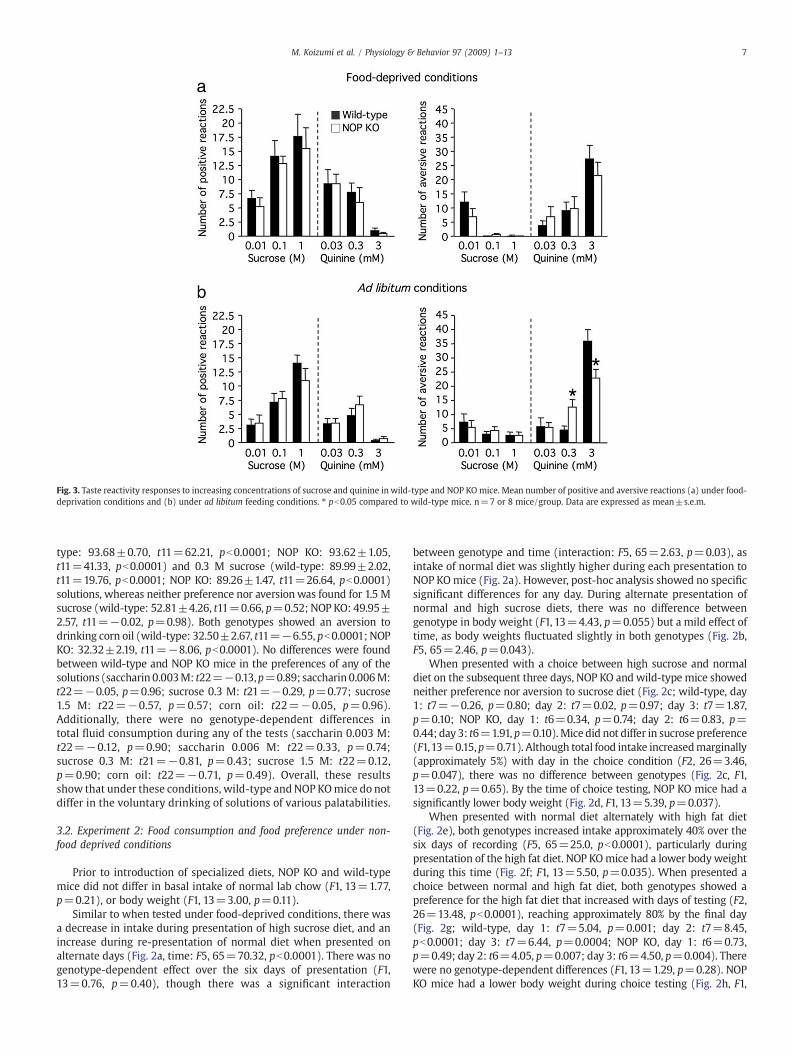
Fig. 3. Taste reactivity responses to increasing concentrations of sucrose and quinine in wild-type and NOP KO mice. Mean number of positive and aversive reactions (a) under food-deprivation conditions and (b) under ad libitum feeding conditions. ⁎ pb0.05 compared to wild-type mice. n=7 or 8 mice/group. Data are expressed as mean±s.e.m.
7M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
type: 93.68±0.70, t11=62.21, pb0.0001; NOP KO: 93.62±1.05,t11=41.33, pb0.0001) and 0.3 M sucrose (wild-type: 89.99±2.02,t11=19.76, pb0.0001; NOP KO: 89.26±1.47, t11=26.64, pb0.0001)solutions, whereas neither preference nor aversionwas found for 1.5 Msucrose (wild-type: 52.81±4.26, t11=0.66, p=0.52; NOP KO: 49.95±2.57, t11=−0.02, p=0.98). Both genotypes showed an aversion todrinking corn oil (wild-type: 32.50±2.67, t11=−6.55, pb0.0001; NOPKO: 32.32±2.19, t11=−8.06, pb0.0001). No differences were foundbetween wild-type and NOP KO mice in the preferences of any of thesolutions (saccharin 0.003M: t22=−0.13,p=0.89; saccharin 0.006M:t22=−0.05, p=0.96; sucrose 0.3 M: t21=−0.29, p=0.77; sucrose1.5 M: t22=−0.57, p=0.57; corn oil: t22=−0.05, p=0.96).Additionally, there were no genotype-dependent differences intotal fluid consumption during any of the tests (saccharin 0.003 M:t22=−0.12, p=0.90; saccharin 0.006 M: t22=0.33, p=0.74;sucrose 0.3 M: t21=−0.81, p=0.43; sucrose 1.5 M: t22=0.12,p=0.90; corn oil: t22=−0.71, p=0.49). Overall, these resultsshow that under these conditions, wild-type and NOP KOmice do notdiffer in the voluntary drinking of solutions of various palatabilities.
3.2. Experiment 2: Food consumption and food preference under non-food deprived conditions
Prior to introduction of specialized diets, NOP KO and wild-typemice did not differ in basal intake of normal lab chow (F1, 13=1.77,p=0.21), or body weight (F1, 13=3.00, p=0.11).
Similar to when tested under food-deprived conditions, there wasa decrease in intake during presentation of high sucrose diet, and anincrease during re-presentation of normal diet when presented onalternate days (Fig. 2a, time: F5, 65=70.32, pb0.0001). There was nogenotype-dependent effect over the six days of presentation (F1,13=0.76, p=0.40), though there was a significant interaction
between genotype and time (interaction: F5, 65=2.63, p=0.03), asintake of normal diet was slightly higher during each presentation toNOP KOmice (Fig. 2a). However, post-hoc analysis showed no specificsignificant differences for any day. During alternate presentation ofnormal and high sucrose diets, there was no difference betweengenotype in body weight (F1, 13=4.43, p=0.055) but a mild effect oftime, as body weights fluctuated slightly in both genotypes (Fig. 2b,F5, 65=2.46, p=0.043).
When presented with a choice between high sucrose and normaldiet on the subsequent three days, NOP KO and wild-type mice showedneither preference nor aversion to sucrose diet (Fig. 2c; wild-type, day1: t7=−0.26, p=0.80; day 2: t7=0.02, p=0.97; day 3: t7=1.87,p=0.10; NOP KO, day 1: t6=0.34, p=0.74; day 2: t6=0.83, p=0.44; day 3: t6=1.91, p=0.10).Mice did not differ in sucrose preference(F1,13=0.15,p=0.71). Although total food intake increasedmarginally(approximately 5%) with day in the choice condition (F2, 26=3.46,p=0.047), there was no difference between genotypes (Fig. 2c, F1,13=0.22, p=0.65). By the time of choice testing, NOP KO mice had asignificantly lower body weight (Fig. 2d, F1, 13=5.39, p=0.037).
When presented with normal diet alternately with high fat diet(Fig. 2e), both genotypes increased intake approximately 40% over thesix days of recording (F5, 65=25.0, pb0.0001), particularly duringpresentation of the high fat diet. NOP KOmice had a lower bodyweightduring this time (Fig. 2f; F1, 13=5.50, p=0.035). When presented achoice between normal and high fat diet, both genotypes showed apreference for the high fat diet that increased with days of testing (F2,26=13.48, pb0.0001), reaching approximately 80% by the final day(Fig. 2g; wild-type, day 1: t7=5.04, p=0.001; day 2: t7=8.45,pb0.0001; day 3: t7=6.44, p=0.0004; NOP KO, day 1: t6=0.73,p=0.49; day 2: t6=4.05, p=0.007; day 3: t6=4.50, p=0.004). Therewere no genotype-dependent differences (F1, 13=1.29, p=0.28). NOPKO mice had a lower body weight during choice testing (Fig. 2h, F1,
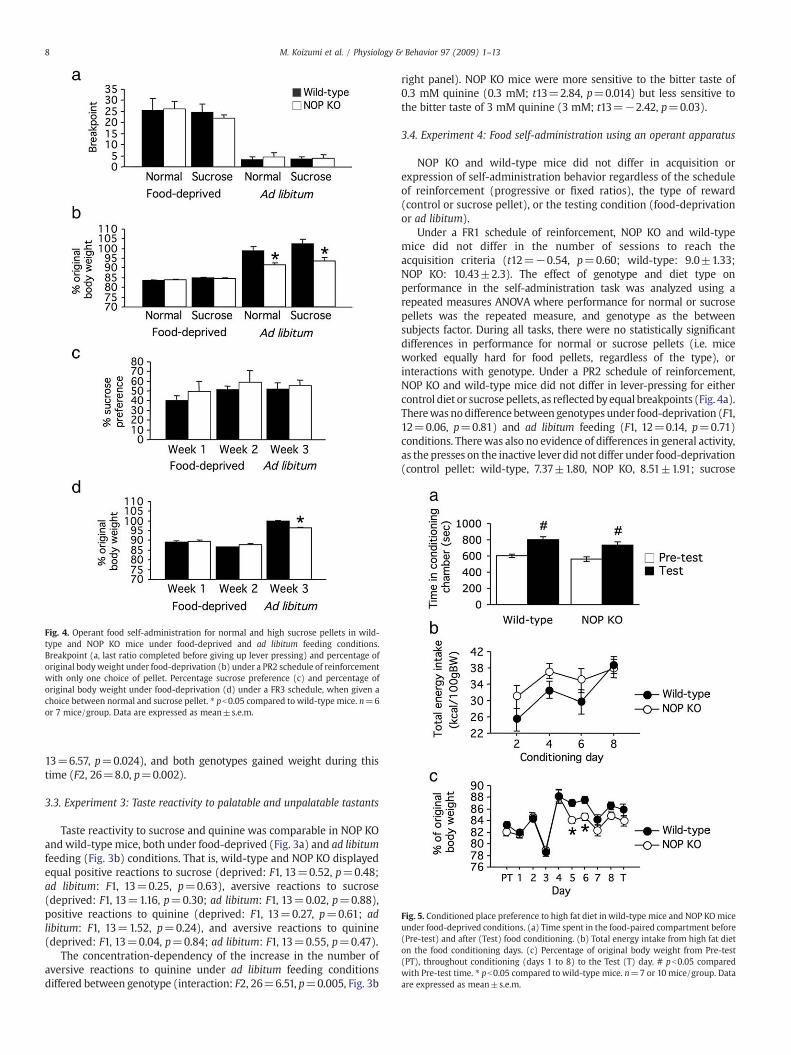
Fig. 4. Operant food self-administration for normal and high sucrose pellets in wild-type and NOP KO mice under food-deprived and ad libitum feeding conditions.Breakpoint (a, last ratio completed before giving up lever pressing) and percentage oforiginal bodyweight under food-deprivation (b) under a PR2 schedule of reinforcementwith only one choice of pellet. Percentage sucrose preference (c) and percentage oforiginal body weight under food-deprivation (d) under a FR3 schedule, when given achoice between normal and sucrose pellet. ⁎ pb0.05 compared to wild-type mice. n=6or 7 mice/group. Data are expressed as mean±s.e.m.
Fig. 5. Conditioned place preference to high fat diet inwild-type mice and NOP KOmiceunder food-deprived conditions. (a) Time spent in the food-paired compartment before(Pre-test) and after (Test) food conditioning. (b) Total energy intake from high fat dieton the food conditioning days. (c) Percentage of original body weight from Pre-test(PT), throughout conditioning (days 1 to 8) to the Test (T) day. # pb0.05 comparedwith Pre-test time. ⁎ pb0.05 compared to wild-type mice. n=7 or 10 mice/group. Dataare expressed as mean±s.e.m.
8 M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
13=6.57, p=0.024), and both genotypes gained weight during thistime (F2, 26=8.0, p=0.002).
3.3. Experiment 3: Taste reactivity to palatable and unpalatable tastants
Taste reactivity to sucrose and quinine was comparable in NOP KOand wild-type mice, both under food-deprived (Fig. 3a) and ad libitumfeeding (Fig. 3b) conditions. That is, wild-type and NOP KO displayedequal positive reactions to sucrose (deprived: F1, 13=0.52, p=0.48;ad libitum: F1, 13=0.25, p=0.63), aversive reactions to sucrose(deprived: F1, 13=1.16, p=0.30; ad libitum: F1, 13=0.02, p=0.88),positive reactions to quinine (deprived: F1, 13=0.27, p=0.61; adlibitum: F1, 13=1.52, p=0.24), and aversive reactions to quinine(deprived: F1, 13=0.04, p=0.84; ad libitum: F1, 13=0.55, p=0.47).
The concentration-dependency of the increase in the number ofaversive reactions to quinine under ad libitum feeding conditionsdiffered between genotype (interaction: F2, 26=6.51, p=0.005, Fig. 3b
right panel). NOP KO mice were more sensitive to the bitter taste of0.3 mM quinine (0.3 mM; t13=2.84, p=0.014) but less sensitive tothe bitter taste of 3 mM quinine (3 mM; t13=−2.42, p=0.03).
3.4. Experiment 4: Food self-administration using an operant apparatus
NOP KO and wild-type mice did not differ in acquisition orexpression of self-administration behavior regardless of the scheduleof reinforcement (progressive or fixed ratios), the type of reward(control or sucrose pellet), or the testing condition (food-deprivationor ad libitum).
Under a FR1 schedule of reinforcement, NOP KO and wild-typemice did not differ in the number of sessions to reach theacquisition criteria (t12=−0.54, p=0.60; wild-type: 9.0±1.33;NOP KO: 10.43±2.3). The effect of genotype and diet type onperformance in the self-administration task was analyzed using arepeated measures ANOVA where performance for normal or sucrosepellets was the repeated measure, and genotype as the betweensubjects factor. During all tasks, there were no statistically significantdifferences in performance for normal or sucrose pellets (i.e. miceworked equally hard for food pellets, regardless of the type), orinteractions with genotype. Under a PR2 schedule of reinforcement,NOP KO and wild-type mice did not differ in lever-pressing for eithercontrol diet or sucrose pellets, as reflectedbyequal breakpoints (Fig. 4a).Therewasnodifference between genotypes under food-deprivation (F1,12=0.06, p=0.81) and ad libitum feeding (F1, 12=0.14, p=0.71)conditions. Therewas also no evidence of differences in general activity,as the presses on the inactive lever did not differ under food-deprivation(control pellet: wild-type, 7.37±1.80, NOP KO, 8.51±1.91; sucrose
9M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
pellet: wild-type, 8.31±3.57, NOP KO, 6.06±1.8; F1, 12=0.03,p=0.86) or ad libitum feeding (control pellet: wild-type, 1.8±0.90,NOP KO, 1.54±0.51; sucrose pellet: wild-type, 2.31±0.98, NOP KO,1.29±0.51; F1, 12=0.37, p=0.55) conditions. The rate of body weightlost (Fig. 4b) under food-deprivation conditions was equal betweengenotypes (F1, 12=0.44, p=0.52). Despite all mice being given freeaccess to food for 10 days before testing again under progressive ratioconditions, NOP KO mice did not fully recover their original bodyweight leading to a significant difference between genotypes in thepercentage of body weight lost (F1, 12=9.44, p=0.01, Fig. 4b).
In the second part of the experiment, micewere presented a choicebetween pressing a lever to obtain control pellets, and pressing adifferent lever to obtain sucrose pellets, under a FR3 schedule ofreinforcement. Under these conditions, NOP KO and wild-type micedid not differ in sucrose preference either under food-deprivation (F1,11=0.58, p=0.46) or ad libitum feeding (t11=0.34, p=0.74)conditions (Fig. 4c). However, both genotypes developed a preferencefor sucrose pellets with training under food-deprivation conditionsthat did not differ between genotypes (weeks: F1, 11=31.30,p=0.0002; interaction: F1, 11=0.11, p=0.75). The total number ofpellets obtained (control and sucrose pellets) was equal betweengenotypes, under both food-deprivation when mice obtained thenumber of pellet allowed (limit of 50 pellets; week 1: wild-type,49.49±0.11, NOP KO: 49.80±0.20; week 2: 50.00±0.00 for NOP KO
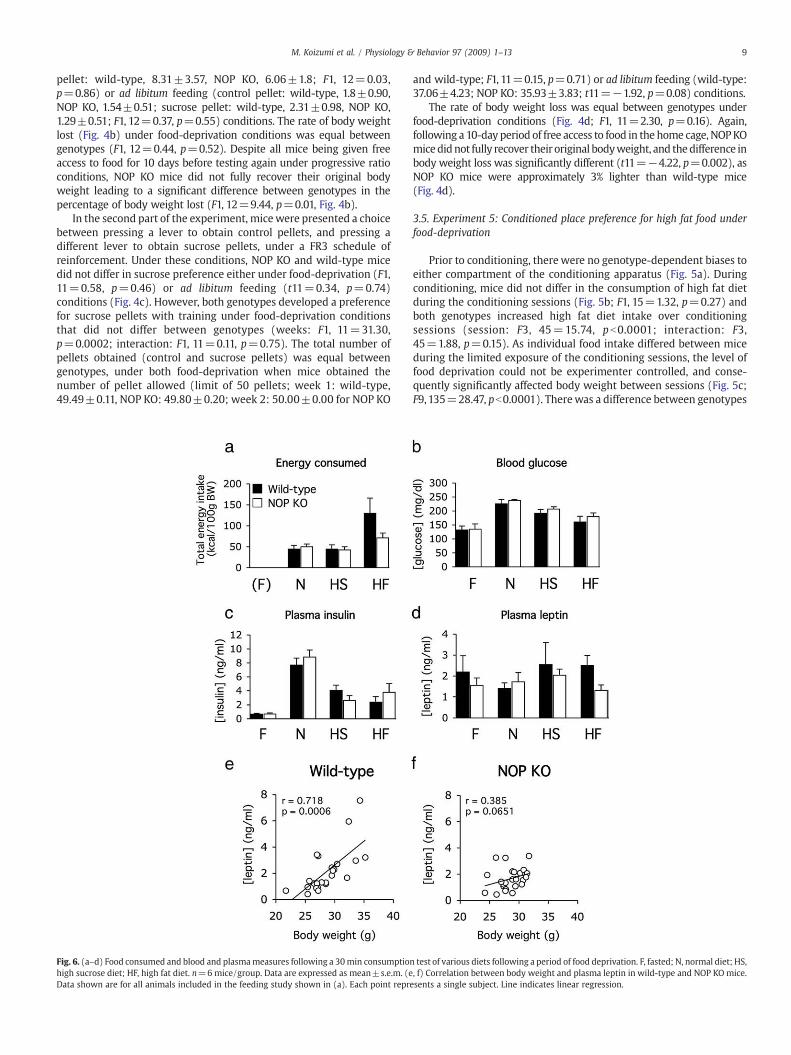
Fig. 6. (a–d) Food consumed and blood and plasmameasures following a 30min consumptionhigh sucrose diet; HF, high fat diet. n=6mice/group. Data are expressed as mean±s.e.m. (eData shown are for all animals included in the feeding study shown in (a). Each point repre
and wild-type; F1, 11=0.15, p=0.71) or ad libitum feeding (wild-type:37.06±4.23; NOP KO: 35.93±3.83; t11=−1.92, p=0.08) conditions.
The rate of body weight loss was equal between genotypes underfood-deprivation conditions (Fig. 4d; F1, 11=2.30, p=0.16). Again,following a 10-day period of free access to food in the home cage, NOPKOmicedidnot fully recover their original bodyweight, and thedifference inbody weight loss was significantly different (t11=−4.22, p=0.002), asNOP KO mice were approximately 3% lighter than wild-type mice(Fig. 4d).
3.5. Experiment 5: Conditioned place preference for high fat food underfood-deprivation
Prior to conditioning, there were no genotype-dependent biases toeither compartment of the conditioning apparatus (Fig. 5a). Duringconditioning, mice did not differ in the consumption of high fat dietduring the conditioning sessions (Fig. 5b; F1, 15=1.32, p=0.27) andboth genotypes increased high fat diet intake over conditioningsessions (session: F3, 45=15.74, pb0.0001; interaction: F3,45=1.88, p=0.15). As individual food intake differed between miceduring the limited exposure of the conditioning sessions, the level offood deprivation could not be experimenter controlled, and conse-quently significantly affected body weight between sessions (Fig. 5c;F9,135=28.47, pb0.0001). Therewas a difference between genotypes
test of various diets following a period of food deprivation. F, fasted; N, normal diet; HS,, f) Correlation between body weight and plasma leptin in wild-type and NOP KO mice.sents a single subject. Line indicates linear regression.
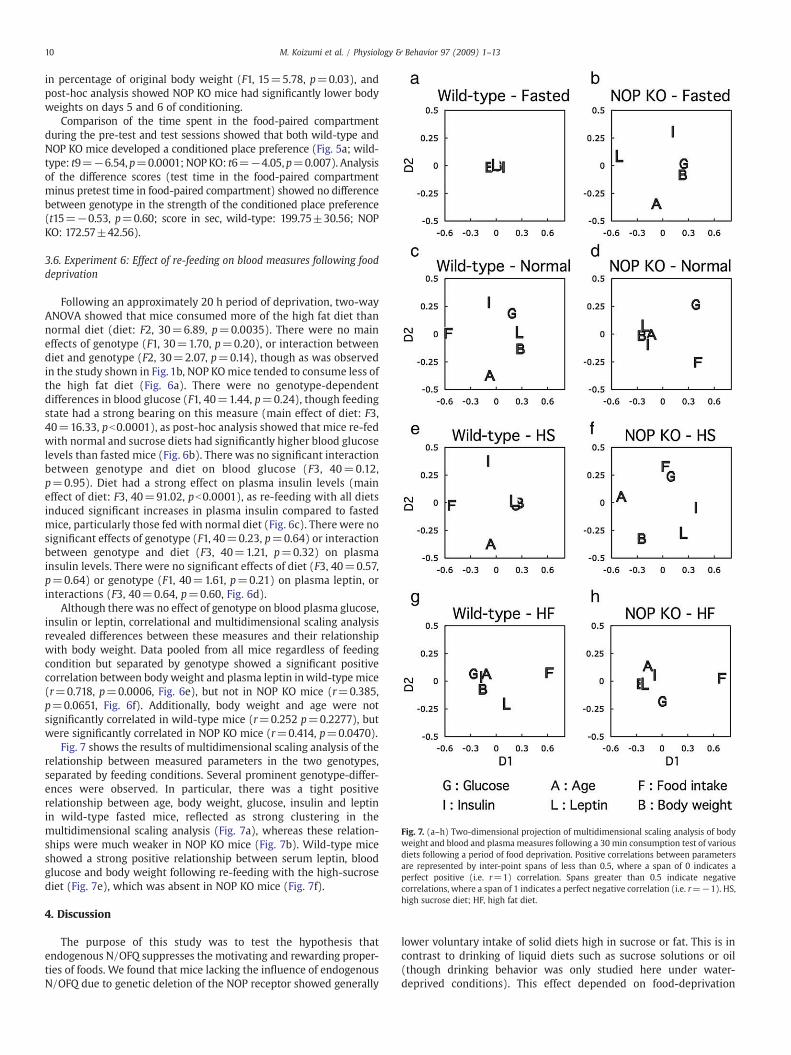
Fig. 7. (a–h) Two-dimensional projection of multidimensional scaling analysis of bodyweight and blood and plasma measures following a 30 min consumption test of variousdiets following a period of food deprivation. Positive correlations between parametersare represented by inter-point spans of less than 0.5, where a span of 0 indicates aperfect positive (i.e. r=1) correlation. Spans greater than 0.5 indicate negativecorrelations, where a span of 1 indicates a perfect negative correlation (i.e. r=−1). HS,high sucrose diet; HF, high fat diet.
10 M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
in percentage of original body weight (F1, 15=5.78, p=0.03), andpost-hoc analysis showed NOP KO mice had significantly lower bodyweights on days 5 and 6 of conditioning.
Comparison of the time spent in the food-paired compartmentduring the pre-test and test sessions showed that both wild-type andNOP KO mice developed a conditioned place preference (Fig. 5a; wild-type: t9=−6.54, p=0.0001; NOPKO: t6=−4.05, p=0.007). Analysisof the difference scores (test time in the food-paired compartmentminus pretest time in food-paired compartment) showed no differencebetween genotype in the strength of the conditioned place preference(t15=−0.53, p=0.60; score in sec, wild-type: 199.75±30.56; NOPKO: 172.57±42.56).
3.6. Experiment 6: Effect of re-feeding on blood measures following fooddeprivation
Following an approximately 20 h period of deprivation, two-wayANOVA showed that mice consumed more of the high fat diet thannormal diet (diet: F2, 30=6.89, p=0.0035). There were no maineffects of genotype (F1, 30=1.70, p=0.20), or interaction betweendiet and genotype (F2, 30=2.07, p=0.14), though as was observedin the study shown in Fig. 1b, NOP KOmice tended to consume less ofthe high fat diet (Fig. 6a). There were no genotype-dependentdifferences in blood glucose (F1, 40=1.44, p=0.24), though feedingstate had a strong bearing on this measure (main effect of diet: F3,40=16.33, pb0.0001), as post-hoc analysis showed that mice re-fedwith normal and sucrose diets had significantly higher blood glucoselevels than fasted mice (Fig. 6b). There was no significant interactionbetween genotype and diet on blood glucose (F3, 40=0.12,p=0.95). Diet had a strong effect on plasma insulin levels (maineffect of diet: F3, 40=91.02, pb0.0001), as re-feeding with all dietsinduced significant increases in plasma insulin compared to fastedmice, particularly those fed with normal diet (Fig. 6c). There were nosignificant effects of genotype (F1, 40=0.23, p=0.64) or interactionbetween genotype and diet (F3, 40=1.21, p=0.32) on plasmainsulin levels. There were no significant effects of diet (F3, 40=0.57,p=0.64) or genotype (F1, 40=1.61, p=0.21) on plasma leptin, orinteractions (F3, 40=0.64, p=0.60, Fig. 6d).
Although there was no effect of genotype on blood plasma glucose,insulin or leptin, correlational and multidimensional scaling analysisrevealed differences between these measures and their relationshipwith body weight. Data pooled from all mice regardless of feedingcondition but separated by genotype showed a significant positivecorrelation between body weight and plasma leptin inwild-type mice(r=0.718, p=0.0006, Fig. 6e), but not in NOP KO mice (r=0.385,p=0.0651, Fig. 6f). Additionally, body weight and age were notsignificantly correlated in wild-type mice (r=0.252 p=0.2277), butwere significantly correlated in NOP KO mice (r=0.414, p=0.0470).
Fig. 7 shows the results of multidimensional scaling analysis of therelationship between measured parameters in the two genotypes,separated by feeding conditions. Several prominent genotype-differ-ences were observed. In particular, there was a tight positiverelationship between age, body weight, glucose, insulin and leptinin wild-type fasted mice, reflected as strong clustering in themultidimensional scaling analysis (Fig. 7a), whereas these relation-ships were much weaker in NOP KO mice (Fig. 7b). Wild-type miceshowed a strong positive relationship between serum leptin, bloodglucose and body weight following re-feeding with the high-sucrosediet (Fig. 7e), which was absent in NOP KO mice (Fig. 7f).
4. Discussion
The purpose of this study was to test the hypothesis thatendogenous N/OFQ suppresses the motivating and rewarding proper-ties of foods. We found that mice lacking the influence of endogenousN/OFQ due to genetic deletion of the NOP receptor showed generally
lower voluntary intake of solid diets high in sucrose or fat. This is incontrast to drinking of liquid diets such as sucrose solutions or oil(though drinking behavior was only studied here under water-deprived conditions). This effect depended on food-deprivation

11M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
state and whether diets were presented alone or as a choice. Despitemany studies showing central administration of N/OFQ stimulatesfeeding behavior in general (see [17,24] for references), there were nodifferences in food intake or body weight in NOP KO mice maintainedunder ad libitum feeding of standard laboratory chow compared towild-type mice.
As other opioid peptides do not appear tomodulate taste sensationitself, but rather orosensory reward [25], we speculated that thealtered consumption of diets with specific macronutrient contents inNOP KO mice directly related to the rewarding impact of the diets.However, no evidence was found to support this speculation. In fact,evidence against a role of endogenous N/OFQ in orosensory rewardwas considerable. That is, NOP KO mice did not differ in sucrose, oil orsaccharin drinking, in the strength of conditioned place preference toa high fat diet, or taste reactions to intraoral infusion of sucrosesolutions. There were also no differences in operant food self-administration, suggesting endogenous N/OFQ does not modulatethe motivating properties of food. Taken together, these studiessuggest that endogenous N/OFQ modulates feeding responses at apost-gustatory level that is dissociated from the motivational andrewarding properties of the food.
In the current study, no genotype-dependent differences werefound in satiated mice, but only under conditions of fooddeprivation, and furthermore, these differences were only inresponse to diets with high sucrose or fat contents. A role forendogenous N/OFQ in feeding under specific dietary conditions hasbeen suggested by studies showing food deprivation decreases N/OFQ and NOP receptor expression in hypothalamic and amygdaloidnuclei [26], which the authors suggest is inconsistent with a directhyperphagic action of endogenous N/OFQ. In particular, theseauthors report that although N/OFQ stimulated laboratory chowintake in freely feeding rats, this effect is no further enhanced infood-deprived rats, suggesting that any role of N/OFQ in consump-tion of regular lab chow may not actually be specific to the food-deprived state. Based on the findings reported here, we speculatethat endogenous N/OFQ activity may be down regulated duringperiods of food deprivation that allows some other process to actthat manifests itself as changes in diet preference. As these previousstudies [26] focused on regular laboratory chow, further studies arerequired to unite these studies with those presented here, whichfocused on diets high in sucrose or fat.
Administration of NOP receptor antagonists alone could poten-tially reveal physiological functions of endogenous N/OFQ, though noeffects on free feeding have been observed in any studies reported todate [27–29]. However, Bodner and colleagues found that antisenseoligodeoxynucleotides targeted at the NOP receptor reduce feedinginduced by food deprivation [30,31], agreeing with the current studythat endogenous N/OFQ modulates feeding at least in the food-deprived state. The conclusion that the effects of N/OFQ on feedingbehavior are largely dissociated from the affective properties of food issupported by previous studies showing that unlike many other opioidpeptides [32], i.c.v. administration of N/OFQ does not increase intakeof preferred foods in rats, but rather stimulates general increases inintake of all diets [14]. However, these studies noted that N/OFQadministration does induce hyperphagia in fat-preferring rats,suggesting a differential role of N/OFQ in feeding behavior, assupported by the findings here.
In the current study, we noted that wild-type and NOP KO micemaintained similar bodyweights despite NOP KOmice consuming lesshigh fat diet during food deprivation, though paradoxically, theyregained their body weight more slowly following food deprivation.Previous studies suggest several possible explanations for such aneffect. One possibility is that NOP KOmice accommodated the reducedfood availability by decreasing physical activity in order to reduceenergy expenditure. Previous studies show that NOP KO micemaintained on ad libitum access to a regular diet do not differ in
locomotor activity [33]. Behavioral activity under food restriction orduring feeding with non-standard diets is yet to be reported, thoughwe have noted that NOP KO mice do not increase their locomotionduring feeding with assorted highly palatable foods meant for humanconsumption (i.e. “cafeteria feeding”, unpublished observation). It isinteresting to note that agonism of the NOP receptor delays gastricemptying, by acting at both central and peripheral levels [34,35] andstimulates gastric acid secretion [36]. Thus, absorption of nutrients inNOP KO mice may differ from wild-type mice, which may contributeto some of the effects observed here.
In the current study, analysis of metabolic and hormonal markersin the blood following re-feeding with various diets after a period offood deprivation showed no overall differences in glucose and insulinresponses. However, more detailed analysis showed that the relation-ship between these factors and leptin was irregular in NOP KO mice.We also noted, that glucose, insulin and leptin levels all appeared to beatypically dissociated from body weight in NOP KOmice under fastingconditions. Numerous studies show a positive predictive relationshipbetween body weight and circulating leptin concentrations (e.g.[37,38]). Whilst this relationshipwas observed here inwild-typemice,it was absent in NOP KO mice. This emphasizes the possibility of amajor disengagement between fundamental hormonal signaling thatoccurs in the absence of endogenous N/OFQ tone. Although identify-ing the precise cause of these aberrant relationships, and how they arereflected in feeding behavior is likely to be challenging, this could be afruitful area of future study.
Several previous studies have addressed the question of themechanism by which N/OFQ induces its hyperphagic effects, par-ticularly on a neuroanatomical level. These have identified severalbrain regions at which N/OFQ stimulates food intake, including thenucleus accumbens, ventral medial hypothalamus [39], paraven-tricular nucleus, arcuate nucleus and nucleus tractus solitarius [40].Ciccocioppo and colleagues have identified a strong interactionbetween N/OFQ effects and neurochemical systems that underliestress responses. In an elegant series of studies, these authorsshowed that N/OFQ acts as a functional antagonist of corticoreleas-ing factor to reverse stress induced anorexia by an action mediatedat least partially in the bed nucleus of the stria terminalis [29,41–43]. Furthermore, these authors have recently shown that centralinjection of corticoreleasing factor increases NOP receptor expres-sion in the bed nucleus of the stria terminalis [44]. A growingnumber of studies suggest a role of N/OFQ in modulating stress (e.g.[45–47]) and anxiety (e.g. [48,49] responses. Food restriction is aclear stressor, and so the altered responses seen under food-deprived conditions in the current study may reflect wider aberrantresponses to stress in NOP KO mice, as shown in several previousreports such as higher levels of anxiety-related behavior whenassayed on an elevated plus maze [46,48,50].
Recent studies increasingly highlight integrative processesbetween central and peripheral sites in the control of reward.However, the lack of evidence of a motivational or reward-baseddeficit in the current study suggests endogenous N/OFQmay controlfeeding responses at a peripheral site. Although the N/OFQprecursor gene is almost exclusively expressed in central sites[51,52], it is also expressed in parts of the periphery, particularly insmooth muscle, peripheral ganglia, and immune cells [53]. Periph-eral roles of N/OFQ have been suggested in a number ofphysiological processes [54], especially in cardiovascular functionand diuresis [55]. Indeed, in our previous studies, we found thatfemale NOP KO mice drink approximately 50% more water thanwild-type mice [9]. Thus, the possibility that the behavioral andmetabolic differences seen in NOP KO mice are due to loss ofendogenous N/OFQ action at predominantly peripheral sites is notout of the question. As far as we are aware, there is no directevidence to support this hypothesis, and this may also constitute aninteresting avenue of study in the future.

12 M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
Finally, it is worth noting that the highest level of N/OFQ expressionoccurs early in development [56], and thus some of the effects observedin the current study may not be due to an immediate loss of N/OFQaction, but as a consequence of atypical development resulting from adeficit of N/OFQ, or unnatural compensation for such a deficit.
In summary, the current study tested the hypothesis that NOP KOmicewould differ in consumption of diets high in sucrose or fat, due toa fundamental involvement of endogenous N/OFQ in psychologicalprocesses such as motivation and reward. This hypothesis was basedon previous studies showing an involvement of endogenous N/OFQ inthe response to rewarding drugs (see [9]) and recognition of manyshared mechanisms between food and drug reward [57,58]. We founda role of endogenous N/OFQ in controlling the consumption of dietshigh in sucrose or fat, though surprisingly; we found very littleevidence that this effect involved higher psychological processes suchas motivation or reward.
Acknowledgements
We thank Ms. Reiko Takahashi for scoring of taste reactivity, Dr.Shuichi Kimura for helpful comments on the study, Dr. Michel Vidal-Naquet for assistance with multidimensional scaling analysis, Dr.Nobuhiko Kojima for providing experimental space and Dr. HiroshiTakeshima for progenitor mice. This workwas supported by the RIKENBrain Science Institute and a Grant-in-Aid for Scientific Research #18500256.
References
[1] Astrup A, Dyerberg J, Selleck M, Stender S. Nutrition transition and its relationshipto the development of obesity and related chronic diseases. Obes Rev 2008;9(Suppl 1):48–52.
[2] Yeomans MR, Gray RW. Opioid peptides and the control of human ingestivebehaviour. Neurosci Biobehav Rev 2002;26(6):713–28.
[3] Bodnar RJ. Endogenous opioids and feeding behavior: a 30-year historicalperspective. Peptides 2004;25(4):697–725.
[4] Barbano MF, Cador M. Opioids for hedonic experience and dopamine to get readyfor it. Psychopharmacology (Berl) 2007;191(3):497–506.
[5] Baldo BA, Kelley AE. Discrete neurochemical coding of distinguishable motiva-tional processes: insights from nucleus accumbens control of feeding. Psycho-pharmacology (Berl) 2007;191(3):439–59.
[6] Olszewski PK, Levine AS. Central opioids and consumption of sweet tastants: whenreward outweighs homeostasis. Physiol Behav 2007;91(5):506–12.
[7] Meis S. Nociceptin/orphanin FQ: actions within the brain. Neuroscientist 2003;9(2):158–68.
[8] Murphy NP. Nociceptin/orphanin FQ, hedonic state and the response to abuseddrugs. Nihon Shinkei Seishin Yakurigaku Zasshi 2004;24(5):295–8.
[9] Sakoori K, Murphy NP. Endogenous nociceptin (orphanin FQ) suppresses basalhedonic state and acute reward responses to methamphetamine and ethanol, butfacilitates chronic responses. Neuropsychopharmacology 2008;33(4):877–91.
[10] Koizumi M, Midorikawa N, Takeshima H, Murphy NP. Exogenous, but notendogenous nociceptin modulates mesolimbic dopamine release in mice.J Neurochem 2004;89(1):257–63.
[11] Wagner EJ, Ronnekleiv OK, Grandy DK, Kelly MJ. The peptide orphanin FQ inhibitsbeta-endorphin neurons and neurosecretory cells in the hypothalamic arcuatenucleus byactivating an inwardly-rectifying K+conductance. Neuroendocrinology1998;67(2):73–82.
[12] Neal Jr CR, Mansour A, Reinscheid R, Nothacker HP, Civelli O, Watson Jr SJ.Localization of orphanin FQ (nociceptin) peptide and messenger RNA in thecentral nervous system of the rat. J Comp Neurol 1999;406(4):503–47.
[13] Gao Q, Horvath TL. Neurobiology of feeding and energy expenditure. Annu RevNeurosci 2007;30:367–98.
[14] Olszewski PK, Grace MK, Sanders JB, Billington CJ, Levine AS. Effect of nociceptin/orphanin FQ on food intake in rats that differ in diet preference. PharmacolBiochem Behav 2002;73(3):529–35.
[15] Pomonis JD, Billington CJ, Levine AS. Orphanin FQ, agonist of orphan opioidreceptor ORL1, stimulates feeding in rats. Neuroreport 1996;8(1):369–71.
[16] Bomberg EM, Grace MK, Levine AS, Olszewski PK. Functional interaction betweennociceptin/orphanin FQ and alpha-melanocyte-stimulating hormone in theregulation of feeding. Peptides 2006;27(7):1827–34.
[17] Przydzial MJ, Heisler LK. Nociceptin/orphanin FQ peptide receptor as a therapeutictarget for obesity. Mini Rev Med Chem 2008;8(8):796–811.
[18] Barbano MF, Cador M. Various aspects of feeding behavior can be partiallydissociated in the rat by the incentive properties of food and the physiologicalstate. Behav Neurosci 2005;119(5):1244–53.
[19] Nishi M, Houtani T, Noda Y, Mamiya T, Sato K, Doi T, et al. Unrestrained nociceptiveresponse and disregulation of hearing ability in mice lacking the nociceptin/orphaninFQ receptor. Embo J 1997;16(8):1858–64.
[20] Berridge KC. Measuring hedonic impact in animals and infants: microstructure ofaffective taste reactivity patterns. Neurosci Biobehav Rev 2000;24(2):173–98.
[21] Kiefer SW, Hill KG, Kaczmarek HJ. Taste reactivity to alcohol and basic tastes inoutbred mice. Alcohol Clin Exp Res 1998;22(5):1146–51.
[22] Hodos W. Progressive ratio as a measure of reward strength. Science1961;134:943–4.
[23] Tzschentke TM. Measuring reward with the conditioned place preference (CPP)paradigm: update of the last decade. Addict Biol 2007;12(3–4):227–462.
[24] Olszewski PK, Levine AS. Minireview: characterization of influence of centralnociceptin/orphanin FQ on consummatory behavior. Endocrinology 2004;145(6):2627–32.
[25] Kelley AE, Bakshi VP, Haber SN, Steininger TL, Will MJ, Zhang M. Opioidmodulation of taste hedonics within the ventral striatum. Physiol Behav2002;76(3):365–77.
[26] Rodi D, Polidori C, Bregola G, Zucchini S, Simonato M, Massi M. Pro-nociceptin/orphanin FQ and NOP receptor mRNA levels in the forebrain of food deprived rats.Brain Res 2002;957(2):354–61.
[27] Economidou D, Policani F, Angellotti T, Massi M, Terada T, Ciccocioppo R. Effect ofnovel NOP receptor ligands on food intake in rats. Peptides 2006;27(4):775–83.
[28] Polidori C, Calo G, Ciccocioppo R, Guerrini R, Regoli D, Massi M. Pharmacologicalcharacterization of the nociceptin receptor mediating hyperphagia: identificationof a selective antagonist. Psychopharmacology (Berl) 2000;148(4):430–7.
[29] Ciccocioppo R, Biondini M, Antonelli L, Wichmann J, Jenck F, Massi M. Reversalof stress- and CRF-induced anorexia in rats by the synthetic nociceptin/orphanin FQ receptor agonist, Ro 64-6198. Psychopharmacology (Berl)2002;161(2):113–9.
[30] Hadjimarkou MM, Khaimova E, Pan YX, Rossi GC, Pasternak GW, Bodnar RJ.Feeding induced by food deprivation is differentially reduced by opioid receptorantisense oligodeoxynucleotide probes in rats. Brain Res 2003;987(2):223–32.
[31] Hadjimarkou MM, Singh A, Kandov Y, Israel Y, Pan YX, Rossi GC, et al. Opioidreceptor involvement in food deprivation-induced feeding: evaluation of selectiveantagonist and antisense oligodeoxynucleotide probe effects in mice and rats.J Pharmacol Exp Ther 2004;311(3):1188–202.
[32] Glass MJ, Billington CJ, Levine AS. Opioids and food intake: distributed functionalneural pathways? Neuropeptides 1999;33(5):360–8.
[33] Uezu K, Sei H, Sano A, Toida K, Suzuki-Yamamoto T, Houtani T, et al. Lack ofnociceptin receptor alters body temperature during resting period in mice.Neuroreport 2004;15(5):751–5.
[34] Broccardo M, Guerrini R, Petrella C, Improta G. Gastrointestinal effects ofintracerebroventricularly injected nociceptin/orphaninFQ in rats. Peptides2004;25(6):1013–20.
[35] Broccardo M, Agostini S, Petrella C, Guerrini R, Improta G. Central and peripheralrole of the nociceptin/orphaninFQ system on normal and disturbed colonic motorfunction and faecal pellet output in the rat. Neurogastroenterol Motil 2008;20(8):939–48.
[36] Ishihara S, Minowa S, Tsuchiya S, Horie S, Watanabe K, Murayama T. Gastric acidsecretion stimulated by centrally injected nociceptin in urethane-anesthetizedrats. Eur J Pharmacol 2002;441(1–2):105–14.
[37] Mallamaci F, Tripepi G, Zoccali C. Leptin in end stage renal disease (ESRD): a linkbetween fat mass, bone and the cardiovascular system. J Nephrol 2005;18(4):464–8.
[38] Maffei M, Halaas J, Ravussin E, Pratley RE, Lee GH, Zhang Y, et al. Leptin levels inhuman and rodent: measurement of plasma leptin and ob RNA in obese andweight-reduced subjects. Nat Med 1995;1(11):1155–61.
[39] Stratford TR, Holahan MR, Kelley AE. Injections of nociceptin into nucleusaccumbens shell or ventromedial hypothalamic nucleus increase food intake.Neuroreport 1997;8(2):423–6.
[40] Polidori C, de Caro G, Massi M. The hyperphagic effect of nociceptin/orphanin FQin rats. Peptides 2000;21(7):1051–62.
[41] Ciccocioppo R, Cippitelli A, Economidou D, Fedeli A, Massi M. Nociceptin/orphaninFQ acts as a functional antagonist of corticotropin-releasing factor to inhibit itsanorectic effect. Physiol Behav 2004;82(1):63–8.
[42] Ciccocioppo R, Fedeli A, Economidou D, Policani F, Weiss F, Massi M. The bednucleus is a neuroanatomical substrate for the anorectic effect of corticotropin-releasing factor and for its reversal by nociceptin/orphanin FQ. J Neurosci 2003;23(28):9445–51.
[43] Ciccocioppo R, Martin-Fardon R, Weiss F, Massi M. Nociceptin/orphanin FQinhibits stress- and CRF-induced anorexia in rats. Neuroreport 2001;12(6):1145–9.
[44] Rodi D, Zucchini S, Simonato M, Cifani C, Massi M, Polidori C. Functional antagonismbetween nociceptin/orphanin FQ (N/OFQ) and corticotropin-releasing factor (CRF)in the rat brain: evidence for involvement of the bed nucleus of the stria terminalis.Psychopharmacology (Berl) 2008;196(4):523–31.
[45] Reiss D, Wolter-Sutter A, Krezel W, Ouagazzal AM. Effects of social crowding onemotionality and expression of hippocampal nociceptin/orphanin FQ systemtranscripts in mice. Behav Brain Res 2007;184(2):167–73.
[46] Ouagazzal AM, Moreau JL, Pauly-Evers M, Jenck F. Impact of environmentalhousing conditions on the emotional responses of mice deficient for nociceptin/orphanin FQ peptide precursor gene. Behav Brain Res 2003;144(1–2):111–7.
[47] Griebel G, Perrault G, Sanger DJ. Orphanin FQ, a novel neuropeptide with anti-stress-like activity. Brain Res 1999;836(1–2):221–4.
[48] Gavioli EC, Rizzi A, Marzola G, Zucchini S, Regoli D, Calo G. Altered anxiety-relatedbehavior in nociceptin/orphanin FQ receptor gene knockout mice. Peptides2007;28(6):1229–39.

13M. Koizumi et al. / Physiology & Behavior 97 (2009) 1–13
[49] Fernandez F, Misilmeri MA, Felger JC, Devine DP. Nociceptin/orphanin FQincreases anxiety-related behavior and circulating levels of corticosterone duringneophobic tests of anxiety. Neuropsychopharmacology 2004;29(1):59–71.
[50] Reinscheid RK, Civelli O. The orphanin FQ/nociceptin knockout mouse: abehavioral model for stress responses. Neuropeptides 2002;36(2–3):72–6.
[51] Mollereau C, Simons MJ, Soularue P, Liners F, Vassart G, Meunier JC, et al. Structure,tissue distribution, and chromosomal localization of the prepronociceptin gene.Proc Natl Acad Sci U S A 1996;93(16):8666–70.
[52] Nothacker HP, Reinscheid RK, Mansour A, Henningsen RA, Ardati A, Monsma Jr FJ,et al. Primary structure and tissue distribution of the orphanin FQ precursor. ProcNatl Acad Sci U S A 1996;93(16):8677–82.
[53] Mollereau C, Mouledous L. Tissue distribution of the opioid receptor-like (ORL1)receptor. Peptides 2000;21(7):907–17.
[54] Chiou LC, Liao YY, Fan PC, Kuo PH, Wang CH, Riemer C, et al. Nociceptin/orphaninFQ peptide receptors: pharmacology and clinical implications. Curr Drug Targets2007;8(1):117–35.
[55] Kapusta DR, Burmeister MA, Calo G, Guerrini R, Gottlieb HB, Kenigs VA. Functionalselectivity of nociceptin/orphanin FQ peptide receptor partial agonists oncardiovascular and renal function. J Pharmacol Exp Ther 2005;314(2):643–51.
[56] Ikeda K, WatanabeM, Ichikawa T, Kobayashi T, Yano R, Kumanishi T. Distribution ofprepro-nociceptin/orphanin FQ mRNA and its receptor mRNA in developing andadult mouse central nervous systems. J Comp Neurol 1998;399(1):139–51.
[57] Trinko R, Sears RM, Guarnieri DJ, DiLeone RJ. Neural mechanisms underlyingobesity and drug addiction. Physiol Behav 2007;91(5):499–505.
[58] Volkow ND, Wise RA. How can drug addiction help us understand obesity?Nat Neurosci 2005;8(5):555–60.
Related Documents











