Villarroya, A. Shimada, A. Varela-Echavarría and O. Mora Y. Ghinis-Hozumi, A. González-Gallardo, L. González-Dávalos, A. Antaramian, F. muscle, and adipose tissue Bovine sirtuins: Initial characterization and expression of sirtuins 1 and 3 in liver, doi: 10.2527/jas.2010-3476 originally published online March 18, 2011 2011, 89:2529-2536. J ANIM SCI http://www.journalofanimalscience.org/content/89/8/2529 the World Wide Web at: The online version of this article, along with updated information and services, is located on www.asas.org by guest on September 13, 2014 www.journalofanimalscience.org Downloaded from by guest on September 13, 2014 www.journalofanimalscience.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Villarroya, A. Shimada, A. Varela-Echavarría and O. MoraY. Ghinis-Hozumi, A. González-Gallardo, L. González-Dávalos, A. Antaramian, F.
muscle, and adipose tissueBovine sirtuins: Initial characterization and expression of sirtuins 1 and 3 in liver,
doi: 10.2527/jas.2010-3476 originally published online March 18, 20112011, 89:2529-2536.J ANIM SCI
http://www.journalofanimalscience.org/content/89/8/2529the World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

Bovine sirtuins: Initial characterization and expression of sirtuins 1 and 3 in liver, muscle, and adipose tissue1,2
Y. Ghinis-Hozumi,* A. González-Gallardo,† L. González-Dávalos,‡ A. Antaramian,† F. Villarroya,§ A. Shimada,‡ A. Varela-Echavarría,† and O. Mora‡3
*Programa de Posgrado en Ciencias de la Producción y de la Salud Animal, Universidad Nacional Autónoma de México (UNAM), México City 04510, México; †Instituto de Neurobiología, UNAM, Blvd. Juriquilla 3001,
Querétaro, Qro. 76230, México; ‡Laboratorio de Rumiología y Metabolismo Nutricional (RuMeN), Secretaría de Posgrado, Facultad de Estudios Superiores-Cuautitlán, UNAM, Blvd. B. Quintana 514-D,
Col. Arboledas, Querétaro, Qro. 76140, México; and §Departamento de Bioquímica y Biología Molecular e Instituto de Biomedicina (IBUB), Universitat de Barcelona y CIBER Fisiopatología de la Obesidad y Nutrición,
Av. Diagonal 645, Barcelona, 08028, Spain
ABSTRACT: Sirtuins, the mammalian homologs of the silent information regulator 2 gene of Saccharomy-ces cerevisiae, are members of the NAD+-dependent family of histone deacetylases. In vertebrates, 7 sirtuins have been described, with different cellular localiza-tions and target proteins. Glucose and lipid metabolism are among the processes regulated by these enzymes. In ruminants, gluconeogenesis is the main biochemical pathway by which glucose is obtained. Because sirtuins in bovines have not been studied, the aim of this work was to obtain sequences coding for the 7 sirtuins and determine the expression patterns of sirtuin1 (Sirt1) and sirtuin3 (Sirt3) in the liver, muscle, and adipose tissue of calves and bulls. Using PCR amplification, we obtained sirtuin gene sequences and reported them to the National Center for Biotechnology Information GenBank. Characteristic sequence motifs correspond-ing to the sirtuin catalytic core domain were found, including the active and zinc-binding sites. Relative expression patterns of Sirt1 and Sirt3 in liver, muscle,
and adipose tissue were quantified by real-time PCR, normalizing to the geometric mean of the housekeeping genes cyclophilin A and β-actin. Expression of Sirt1 was less in liver and muscle, whereas it was greater in adipose tissue of adult animals, with statistical dif-ferences (P = 0.0071) only in the latter. In the case of Sirt3, expression was greater in all 3 adult tissues, but statistical differences were found only in liver (P = 0.0141) and muscle (P = 0.0017). The greatest ex-pression was observed in liver for Sirt1 and in muscle for Sirt3, whereas the least expression was in muscle for Sirt1 and in adipose tissue for Sirt3. In other spe-cies, sirtuin expression (both Sirt1 and Sirt3) in liver is reported to be the greatest among these 3 tissues, a pattern different from what we measured. These differ-ences in expression can be associated with metabolic differences between nonruminant and ruminant species. However, further research on the relationship between bovine sirtuins and ruminant metabolism is required for a better understanding of these fields.
Key words: adipose tissue, bovine, expression, liver, muscle, sequence, sirtuin
©2011 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2011. 89:2529–2536 doi:10.2527/jas.2010-3476
INTRODUCTION
Sirtuins, the mammalian homologs of the silent in-formation regulator 2 gene of Saccharomyces cerevisiae, are members of the NAD+-dependent family of histone deacetylases (Yamamoto et al., 2007). At first, research focused on their role as repressors of gene expression, but Schwer et al. (2002) found that they also deacety-late nonhistone proteins, inducing the transcription of several genes involved in metabolism.
In vertebrates, 7 sirtuins have been described, each with different target proteins (Saunders and Verdin, 2007). Sirtuin1 (Sirt1) is present in the nucleus, and
1 This study was supported by grant 55429 from SEP-CONACYT [SEP (Ministry of Public Education)-Consejo Nacional de Ciencia y Tecnología, México City, México].
2 This work is part of the doctoral thesis submitted by the first author to the Universidad Nacional Autónoma de México (UNAM). Yumi Ghinis-Hozumi also thanks CONACYT for a scholarship at Facultad de Estudios Superiores-Cuautitlán, UNAM, and grants SAF2008-01896 from the Ministerio de Ciencia e Innovación, Spain. We also thank Dorothy Pless [INB (Neurobiology Institute)-UNAM, Querétaro, Querétaro, México] for revising the English manuscript.
3 Corresponding author: [email protected] or [email protected]
Received August 31, 2010.Accepted March 4, 2011.
2529
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

among its known targets are transcriptional regulators, such as PPARγ coactivator 1α (Rodgers et al., 2005), the PPAR (Yu and Auwerx, 2009), and the forkhead box, subgroup O transcription factors (Wang and Tong, 2009), as well as enzymes, such as cytosolic acetyl-CoA synthetase (Hallows et al., 2006), glutamate dehydroge-nase (Shoba et al., 2009), and isocitrate dehydrogenase (Schlicker et al., 2008). Sirtuin3 (Sirt3) is mainly a mitochondrial sirtuin; it deacetylates enzymes involved in lipid oxidation, such as mitochondrial acetyl-CoA synthetase (Hallows et al., 2006) and long-chain-acyl-CoA dehydrogenase (Hirschey et al., 2010), thereby in-creasing their activity. Collectively, sirtuins appear to be important metabolic regulators, especially of gluco-neogenesis and lipid oxidation.
In ruminants, gluconeogenesis is the main biochemi-cal pathway used to generate glucose from substrates other than carbohydrates, mainly propionate, glycerol, and AA (Shimada, 2003). Therefore, the present study was conducted to obtain the sequences of the 7 bo-vine sirtuins and to quantify the expression of Sirt1 and Sirt3 in liver, muscle, and white adipose tissue of bulls and of calves up to 3 d of age to determine changes in their expression between the nonruminant and rumi-nant stages of development.
MATERIALS AND METHODS
The experimental protocol was approved by the Ani-mal Care Advisory Committee of the National Autono-mous University of Mexico (UNAM). All procedures were carried out in accordance with the Mexican federal laws for animal care (SAGAR, 1997, 1998; SAGARPA, 2001).
Animals
Six male Holstein calves, between 1 and 3 d of age, were purchased from a dairy farm in the municipality of Pedro Escobedo in the state of Querétaro, México. The animals were killed, in the absence of food deprivation, by stunning with a captive bolt gun, followed by exsan-guination. Six randomly selected crossbred bulls were sampled at the local slaughterhouse in the municipal-ity of Santiago de Querétaro in the state of Querétaro, México. Samples from the liver, semitendinosus and semimembranous muscles, subcutaneous adipose tissue, kidney, and testis were collected from both calves and bulls, immediately frozen in liquid nitrogen, and kept at −70°C until their analysis.
Purification of Total RNA
Total RNA from liver, kidney, and testis tissue samples was purified using a Qiagen RNeasy Mini Kit (Qiagen Inc., Valencia, CA), whereas TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA) was used to isolate RNA from muscle and adipose tissue, with a
subsequent purification using a Qiagen RNeasy Mini Kit according to the manufacturer’s instructions. The RNA obtained were quantified with a NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific Inc., Wilmington, DE) at 260 nm, and purity was assessed using the 260- to 280-nm ratios, which ranged from 1.8 to 2.0. Integrity of the RNA was checked by electropho-resis in a 1% agarose gel.
cDNA Synthesis Through Reverse-Transcriptase PCR
For cDNA synthesis, SuperScript II Reverse Tran-scriptase (Invitrogen Life Technologies) was used with 2 μg of total RNA and Oligo(dT)12–18 primer (Sigma-Genosys, St. Louis, MO), following the manufacturer’s instructions. Before beginning the reaction, RNA was treated with DNase (15-min incubation at room tem-perature and 5-min incubation at 70°C) to reduce ge-nomic DNA contamination.
Bovine Sirtuin mRNA Sequences
Specific primers for the 7 bovine sirtuins were de-signed using the predicted sequences available at the NCBI (National Center for Biotechnology Information) GenBank, derived from the publicly available bovine reference genome (Zimin et al., 2009), and taking care that the products obtained would belong to the cod-ing regions, either partially or totally (Table 1). Primer design followed basic design rules (Dieffenbach et al., 1993). Table 1 shows the designed primers and the predicted and obtained fragment sizes for the 7 sirtu-ins. The PCR products were obtained using previously synthesized first-strand cDNA and a Recombinant Taq DNA Polymerase Kit (Invitrogen Life Technologies). The PCR amplification parameters consisted of an ini-tial denaturation at 94°C for 5 min, 40 cycles of de-naturation at 94°C for 1 min, annealing for 1 min at the specific temperature for each sirtuin primer pair, and extension at 72°C for 1 min, with a final 10-min extension at 72°C, added to help complete unfinished amplifications. The PCR reactions were prepared us-ing the following final concentrations: 1× PCR Buf-fer, 0.2 mM deoxynucleotide 5′-triphosphate mixture, 0.2 μM forward and reverse primers, 0.1 U/μL of Taq DNA Polymerase (Invitrogen Life Technologies), 3 μL of cDNA from either testis [Sirt1, sirtuin2 (Sirt2), and sirtuin4 (Sirt4)] or kidney [Sirt3, sirtuin5 (Sirt5), sir-tuin6 (Sirt6), and sirtuin7 (Sirt7)], either 1 mM (Sirt4 and Sirt7), 2 mM (Sirt1, Sirt2, Sirt3, and Sirt6), or 4 mM (Sirt5) MgCl2, and water to a final reaction volume of 25 μL. Additionally, a standard curve for the volume of cDNA input was performed for Sirt7, and it was determined that 2 μL was optimal. Different annealing temperatures were used for each sirtuin: 50°C for Sirt6, 52°C for Sirt3, 55°C for Sirt2, 58°C for Sirt4, 60°C for Sirt1 and Sirt7, and 64°C for Sirt5. The PCR products
2530 Ghinis-Hozumi et al.
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

of Sirt1, Sirt6, and Sirt7, as well as cloned PCR prod-ucts of Sirt2, Sirt3, Sirt4, and Sirt5 were sequenced in a 310 ABI Prism Sequencer with version 3 Big Dye (Ap-plied Biosystems, Carlsbad, CA).
The sequences obtained were searched by BLAST (Basic Local Alignment Search Tool; Altschul et al., 1997) to confirm the identity of the obtained sirtuin products. Different alignment tools, namely, ClustalW (Larkin et al., 2007), Jufo (Meiler et al., 2001, Mei-ler and Baker, 2003), and Strap (Gille and Frömmel, 2001), were used to compare the sequences obtained with the sirtuin gene and protein sequences from other species available in GenBank.
Sirt1 and Sirt3 Expression Profiles
Real-time PCR primers were designed for Sirt1 and Sirt3 (Table 1), and their expression profiles in liver, muscle, and adipose tissue were analyzed by real-time PCR using a Roche LightCycler 2.0 Instrument (Roche Applied Science, Mannheim, Germany) and LightCy-cler FastStart DNA Master Sybr Green I (Roche Ap-plied Science). Expression stability of 3 selected house-keeping genes [cyclophilin A (PPIA), 18S rRNA, and β-actin] was evaluated using both geNorm (Biogazelle
NV, Ghent, Belgium) and the Excel add-in Normfinder (MDL, Aarhus, Denmark). The stability values ob-tained with geNorm were 1.567, 1.643, and 1.891, and with Normfinder they were 0.031, 0.033, and 0.048 for PPIA, β-actin, and 18S rRNA, respectively. Comple-mentary DNA standard curves were prepared using se-rial dilutions to obtain the reaction efficiencies for each gene, which were 2.154 for Sirt1, 2.247 for Sirt3, 2.117 for PPIA, and 2.298 for β-actin, as given by the Light-Cycler Software 4.0 (Roche Applied Science). There-fore, quantification is expressed relative to the geomet-ric mean of PPIA and β-actin (Vandesompele et al., 2002) using the 2−ΔCt method (where Ct refers to cycle threshold; Livak and Schmittgen, 2001; Schmittgen and Livak, 2008).
The real-time PCR program used consisted of a pre-incubation at 95°C for 2 min, 55 quantification cycles consisting of a 10-s denaturation at 95°C, a 10-s an-nealing at 58°C, and an 11-s extension at 72°C. The amplification program was followed by a melt curve analysis, which consisted of 1 min of annealing at 65°C, a temperature increase of 0.1°C/s up to 95°C, and a fi-nal cooling cycle at 40°C for 30 s. The melt curves were performed to ensure purity of the amplified product, which was confirmed by a single peak after 80°C.
Table 1. Polymerase chain reaction primers for bovine sirtuins (Sirt)
SirtuinReference sequence accession number Primer position,1 5′ → 3′
Predicted/obtained fragment size
PCR product type
Sirt1 NM_001192980.1 Forward: ATACACTGGAGCAGGTT 164 bp/173 bp Real-time1031–1047Reverse: TTCATCAGCTGGGCATCTAG1175–1194
Sirt2 NM_001113531.1 Forward: GAGAAGATCTTCTCCGA 650 bp/648 bp Conventional716–732Reverse: GAAATGTTCTGAGCTCCCCA1346–1365
Sirt3 XM_002683729.1 Forward: AAGCATGGCGTTGTTTCCTC 1093 bp/616 bp Conventional46–65Reverse: AAACGTGCCCGAGCTCTTC1120–1138
Sirt3 XM_002683729.1 Forward: GCTAGGTTCCTGCTGCATCT 264 bp Real-time746–765Reverse: GATGAGGTCCTGGATGTCGT990–1009
Sirt4 NM_001075785.1 Forward: AGAGATGTTGCTTTGCCGCC 859 bp/594 bp Conventional266–285Reverse: AGAGATGTTGCTTTGCCGCC1105–1124
Sirt5 NM_001034295.1 Forward: TGGGGAGAGGAATGCAACAC 1094 bp/1094 bp Conventional88–107Reverse: TTGGATTCCAGCCCAGCAAT1162–1181
Sirt6 NM_001098084.1 Forward: TTAACTTGGGGCCTATGGAG 161 bp/231 bp Real-time1258–1277Reverse: TTAGGCTGGAAATCAGGGAG1399–1418
Sirt7 NM_001075217.1 Forward: CGTAACCTGGTGCTTGACCA 133 bp/133 bp Real-time1205–1224Reverse: TCAGGGTGAAAATGTCACCC1318–1337
1Below each primer: initial nucleotide to final nucleotide position.
2531Bovine sirtuins
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

Statistical Analysis
Data were analyzed as a 2 × 3 factorial arrangement of treatments (calves or bulls, and liver, muscle, or adi-pose tissue) in a completely randomized design using the GLM procedure (SAS Inst. Inc., Cary, NC), with the significance level set at P ≤ 0.05. Least squares means ± SEM were used to analyze differences between developmental stages and tissues for each sirtuin.
RESULTS
Bovine Sirtuin mRNA Sequences
From the PCR amplifications described above, we obtained fragments of the gene sequences and of the protein coding regions for Sirt1 to Sirt4 and the com-plete coding region for Sirt5. In the case of Sirt6 and Sirt7, the fragments obtained correspond to the 3′ end region. All these sequences are available at NCBI Gen-Bank (accession numbers GQ166647 to GQ166653).
ClustalW (Larkin et al., 2007) analysis of the bovine sirtuin gene sequences (Sirt5) showed identities that ranged from 72 to 94% with Homo sapiens, 67 to 89% with Mus musculus, 68 to 79% with Rattus norvegicus, 81 to 96% with Sus scrofa, 52 to 87% with Gallus gal-lus, and 67 to 95% with Equus caballus, whereas pre-dicted proteins had identities ranging from 70 to 96% with H. sapiens, 66 to 94% with M. musculus, 51 to 96% with R. norvegicus, 76 to 96% with S. scrofa, 56 to 91% with G. gallus, and 67 to 96% with E. caballus. The gene sequences obtained for Sirt6 and Sirt7 are very short fragments; hence, the identities of these with their orthologs are very small, with the exception of S. scrofa (78 and 85%, respectively) and E. caballus (with 80% identity with Sirt7; Table 2).
After alignment of the bovine sirtuin sequences against S. cerevisiae Sir2 with different alignment tools (ClustalW, Jufo, and Strap), secondary structures were predicted using the Geneious software (Drummond et al., 2010) and are presented in Figure 1, which shows
the conserved sirtuin core domain motifs, including substrate, NAD+, and zinc binding sites.
Sirt1 and Sirt3 Expression Profiles
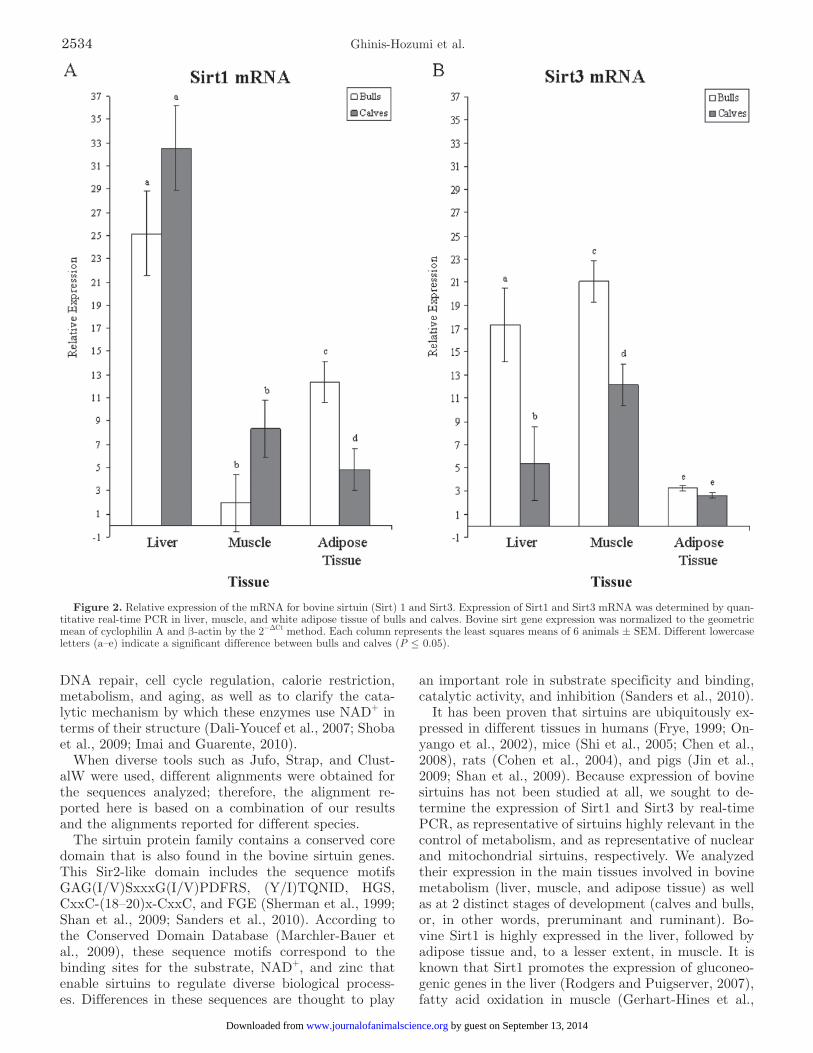
Real-time PCR was performed to determine the ex-pression of bovine Sirt1 and Sirt3 genes in liver, muscle, and adipose tissue of both calves and bulls. The rela-tive expression results were obtained using the 2−ΔCT method after normalization with the geometric means of PPIA and β-actin. As shown in Figure 2A, when comparing expression of Sirt1 of bulls against calves, the expression of Sirt1 in liver and muscle was some-what greater in calves than in bulls, although no sta-tistical differences were found; in adipose tissue, it was greater in bulls than in calves, and this difference was significant (P = 0.0071). In the case of Sirt3 (Figure 2B), expression was greater in all 3 adult tissues when compared with those of calves. In liver and muscle, sta-tistical differences were found (P = 0.0141 and 0.0017, respectively), whereas in adipose tissue there was no statistical difference.
Additionally, when comparing the expression of Sirt1 against that of Sirt3 (Figure 2A and 2B), it can be observed that expression of both sirtuins in adult liver samples was not statistically different; this was also the case for calf muscle and adipose tissue. In contrast, the relative expression of Sirt1 in calf liver, when normal-ized to the geometric mean of the housekeeping genes, was 32.52, whereas Sirt3 expression was only 5.36, a 6-fold difference; this was also the case for adult muscle, with an 11-fold difference (1.94 vs. 21.07, respectively), and adipose tissue, with a 3.8-fold difference (12.37 vs. 3.24, respectively). However, these results raise a note of caution about the differences in expression profiles because the data were not statistically analyzed.
DISCUSSION
In the past decade, sirtuins have been extensively studied to better understand their role in gene silencing,
Table 2. Sirtuin (Sirt) gene and protein homology between the bovine and other spe-cies (% identity)
SirtHomo sapiens
Mus musculus
Rattus norvegicus
Sus scrofa
Gallus gallus
Equus caballus
Gene or protein
Sirt1 94 89 — 96 87 95 Gene96 94 96 96 91 96 Protein
Sirt2 87 79 79 91 52 68 Gene86 82 83 93 65 76 Protein
Sirt3 72 67 68 81 55 73 Gene70 66 51 76 56 67 Protein
Sirt4 74 78 79 87 54 67 Gene91 89 87 91 69 91 Protein
Sirt5 83 80 78 88 60 83 Gene87 85 83 87 74 91 Protein
Sirt6 33 15 18 78 6 12 GeneSirt7 62 55 49 85 9 80 Gene
2532 Ghinis-Hozumi et al.
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

Fig
ure
1. A
lignm
ent
of y
east
Sir
2 an
d bo
vine
sir
tuin
s. B
ovin
e si
rtui
ns c
onta
in t
he c
ore
dom
ain
of t
he s
irtu
in fam
ily. T
his
Sir2
-lik
e do
mai
n in
clud
es t
he 4
con
serv
ed c
yste
ines
of th
e pu
tative
zin
c fin
ger
(+),
whi
ch g
ener
ally
has
the
seq
uenc
e C
xxC
-(18
–20)
x-C
xxC
; N
AD
+-b
indi
ng s
ites
(|)
, co
nsis
ting
of th
e se
quen
ces
GA
G(I
/V)S
xxxG
(I/V
)PD
FR
S an
d (Y
/I)T
QN
ID; th
e se
quen
ce H
GS,
whi
ch
cont
ains
the
act
ive
site
res
idue
(m
arke
d w
ith
a st
ar);
and
sub
stra
te b
indi
ng s
ites
(■
), w
hich
inc
lude
the
seq
uenc
e FG
E, am
ong
othe
rs. Su
bstr
ate
bind
ing
site
s ar
e di
ffer
ent
for
each
sir
tuin
and
are
th
ough
t to
det
erm
ine
subs
trat
e sp
ecifi
city
and
cat
alyt
ic a
ctiv
ity
of t
he e
nzym
e. T
hese
mot
ifs a
re d
iagn
ostic
for
mem
bers
of th
is g
ene
fam
ily. Sc
er =
Sac
char
omyc
es c
erev
isia
e; B
tau
= B
os t
auru
s.
2533Bovine sirtuins
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from
DNA repair, cell cycle regulation, calorie restriction, metabolism, and aging, as well as to clarify the cata-lytic mechanism by which these enzymes use NAD+ in terms of their structure (Dali-Youcef et al., 2007; Shoba et al., 2009; Imai and Guarente, 2010).
When diverse tools such as Jufo, Strap, and Clust-alW were used, different alignments were obtained for the sequences analyzed; therefore, the alignment re-ported here is based on a combination of our results and the alignments reported for different species.
The sirtuin protein family contains a conserved core domain that is also found in the bovine sirtuin genes. This Sir2-like domain includes the sequence motifs GAG(I/V)SxxxG(I/V)PDFRS, (Y/I)TQNID, HGS, CxxC-(18–20)x-CxxC, and FGE (Sherman et al., 1999; Shan et al., 2009; Sanders et al., 2010). According to the Conserved Domain Database (Marchler-Bauer et al., 2009), these sequence motifs correspond to the binding sites for the substrate, NAD+, and zinc that enable sirtuins to regulate diverse biological process-es. Differences in these sequences are thought to play
an important role in substrate specificity and binding, catalytic activity, and inhibition (Sanders et al., 2010).
It has been proven that sirtuins are ubiquitously ex-pressed in different tissues in humans (Frye, 1999; On-yango et al., 2002), mice (Shi et al., 2005; Chen et al., 2008), rats (Cohen et al., 2004), and pigs (Jin et al., 2009; Shan et al., 2009). Because expression of bovine sirtuins has not been studied at all, we sought to de-termine the expression of Sirt1 and Sirt3 by real-time PCR, as representative of sirtuins highly relevant in the control of metabolism, and as representative of nuclear and mitochondrial sirtuins, respectively. We analyzed their expression in the main tissues involved in bovine metabolism (liver, muscle, and adipose tissue) as well as at 2 distinct stages of development (calves and bulls, or, in other words, preruminant and ruminant). Bo-vine Sirt1 is highly expressed in the liver, followed by adipose tissue and, to a lesser extent, in muscle. It is known that Sirt1 promotes the expression of gluconeo-genic genes in the liver (Rodgers and Puigserver, 2007), fatty acid oxidation in muscle (Gerhart-Hines et al.,
Figure 2. Relative expression of the mRNA for bovine sirtuin (Sirt) 1 and Sirt3. Expression of Sirt1 and Sirt3 mRNA was determined by quan-titative real-time PCR in liver, muscle, and white adipose tissue of bulls and calves. Bovine sirt gene expression was normalized to the geometric mean of cyclophilin A and β-actin by the 2−ΔCt method. Each column represents the least squares means of 6 animals ± SEM. Different lowercase letters (a–e) indicate a significant difference between bulls and calves (P ≤ 0.05).
2534 Ghinis-Hozumi et al.
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

2007), and fat mobilization in adipose tissue (Picard et al., 2004); therefore, Sirt1 may be regulating glucose production in the liver from glycerol coming from adi-pose tissue because of the effect of Sirt1 on fat, whereas in muscle it is regulating oxidative metabolism. Bovine Sirt3 is expressed in muscle, liver, and adipose tissue, in decreasing amounts. Sirtuin 3 is known to regulate mi-tochondrial function (Hirschey et al., 2010) and main-tain basal ATP concentrations (Ahn et al., 2008) in liver, muscle, and adipose tissue, but in the latter, its expression is known to be reduced (Shi et al., 2005).
Compared with what has been reported for other species (Frye, 1999; Cohen et al., 2004; Shi et al., 2005; Chen et al., 2008; Shan et al., 2009), expression profiles of Sirt1 and Sirt3 in bovines are quite different, mainly because in other species both sirtuins are more highly expressed in liver than in muscle and adipose tissue. These results can be explained by the primary metabolic differences between nonruminant and ruminant species; ruminants maintain their blood glucose concentrations in the fed state mainly through active gluconeogenesis (Martin et al., 1973), whereas in nonruminant species, gluconeogenesis is activated only during energy deficit (Nafikov and Beitz, 2007). In gluconeogenesis, glucose is obtained from noncarbohydrate precursors such as VFA, glycerol, and AA. In ruminants, the main sources for gluconeogenesis are propionate, which comes from the rumen, and glycerol, which comes from the adipose tissue, whereas in nonruminant animals, AA and lac-tate provided by the muscle are the main substrates (Shimada, 2003). In the case of ruminants, the question is whether the gluconeogenesis taking place promotes the expression of sirtuins or the reverse, whether the expression of sirtuins favors gluconeogenesis.
In addition to the differences that are related to tissue type, expression of sirtuins changes with age (Alcendor et al., 2007; Ferrara et al., 2008; Shan et al., 2009). In this study, we found an overall distinc-tive profile of developmental regulation of the expres-sion of bovine Sirt3, which was consistently greater in adult than in calf tissues. In contrast, developmental changes in the expression of bovine Sirt1 were tissue-specific: less in adult muscle compared with calf muscle but greater in adult adipose tissue than in calf adipose tissue. The developmental changes observed in the liver were less marked, but a somewhat greater expression in calf liver was observed. According to Howarth et al. (1968), a functional rumen implies a decreased glucose oxidation capacity in the liver and muscle, a greater gluconeogenic capacity in the liver, a decreased glyco-genic capacity in muscle, and a greater activity of fat synthesis-related enzymes in adipose tissue. From this, differences in expression of Sirt1 and Sirt3 in adults and calves can be explained. The calves used for this study consumed milk only twice or three times before euthanasia, undergoing a moderate fasting period. As a result, Sirt1 was probably promoting fatty acid oxi-dation and mitochondrial respiration in the liver and muscle, whereas the small amount of adipose tissue in
calves could explain its decreased expression in adipose, which could actually be its basal expression. Sirtuin3, on the other hand, was probably supporting Sirt1 in the regulation of energy homeostasis in the liver and muscle, with the profile we found in adipose tissue be-ing the basal expression of this sirtuin. However, even when there were metabolic differences between bulls and calves, there was also the possibility that these re-sults were due to breed effects more than to age-related effects, especially because proteomic mapping studies have demonstrated differential expression of proteins from cattle breeds selected for dairy and beef purposes (Miarelli and Signorelli, 2010).
In conclusion, this is the first study concerning bo-vine sirtuins, and we established a tissue-specific, devel-opmentally regulated profile of expression for Sirt1 and Sirt3. Considering the key regulatory role of sirtuins in metabolism reported in other species, and in light of the present findings, further analysis of the expression, function, and role of sirtuin in the regulation of highly differential bovine metabolism is warranted in the near future.
LITERATURE CITED
Ahn, B. H., H. S. Kim, S. Song, I. H. Lee, J. Liu, A. Vassilopoulos, C. X. Deng, and T. Finkel. 2008. A role for the mitochondrial deacetylase Sirt3 in regulating energy homeostasis. Proc. Natl. Acad. Sci. USA 105:14447–14452.
Alcendor, R. R., S. Gao, P. Zhai, D. Zablocki, E. Holle, X. Yu, B. Tian, T. Wagner, S. F. Vatner, and J. Sadoshima. 2007. Sirt1 regulates aging and resistance to oxidative stress in the heart. Circ. Res. 100:1512–1521.
Altschul, S. F., T. L. Madden, A. A. Schäffer, J. Zhang, Z. Zhang, W. Miller, and D. J. Lipman. 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 25:3389–3402.
Chen, D., J. Bruno, E. Easlon, S.-J. Lin, H.-L. Cheng, F. W. Alt, and L. Guarente. 2008. Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev. 22:1753–1757.
Cohen, H. Y., C. Miller, K. J. Bitterman, N. R. Wall, B. Hekking, B. Kessler, K. T. Howitz, M. Gorospe, R. de Cabo, and D. A. Sinclair. 2004. Calorie restriction promotes mammalian cell sur-vival by inducing the SIRT1 deacetylase. Science 305:390–392.
Dali-Youcef, N., M. Lagouge, S. Froelich, C. Koehl, K. Schoonjans, and J. Auwerx. 2007. Sirtuins: The ‘magnificent seven,’ func-tion, metabolism and longevity. Ann. Med. 39:335–345.
Dieffenbach, C. W., T. M. Lowe, and G. S. Dveksler. 1993. General concepts for PCR primer design. PCR Methods Appl. 3:S30–S37.
Drummond, A. J., B. Ashton, S. Buxton, M. Cheung, J. Heled, M. Kearse, R. Moir, S. Stones-Havas, S. Sturrock, T. Thierer, and A. Wilson. 2010. Geneious version 5.0. Accessed Mar. 4, 2011. http://www.geneious.com.
Ferrara, N., B. Rinaldi, G. Corbi, V. Conti, P. Stiuso, S. Boccuti, G. Rengo, F. Rossi, and A. Filippelli. 2008. Exercise train-ing promotes SIRT1 activity in aged rats. Rejuvenation Res. 11:139–150.
Frye, R. A. 1999. Characterization of five human cDNAs with ho-mology to the yeast SIR2 gene: Sir2-like proteins (sirtuins) me-tabolize NAD and may have protein ADP-ribosyltransferase activity. Biochem. Biophys. Res. Commun. 260:273–279.
Gerhart-Hines, Z., J. T. Rodgers, O. Bare, C. Lerin, S.-H. Kim, R. Mostoslavsky, F. W. Alt, Z. Wu, and P. Puigserver. 2007. Meta-bolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1α. EMBO J. 26:1913–1923.
2535Bovine sirtuins
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

Gille, C., and C. Frömmel. 2001. STRAP: Editor for STRuctural Alignments of Proteins. Bioinformatics 17:377–378.
Hallows, W. C., S. Lee, and J. M. Denu. 2006. Sirtuins deacetylate and activate mammalian acetyl-CoA synthetases. Proc. Natl. Acad. Sci. USA 103:10230–10235.
Hirschey, M. D., T. Shimazu, E. Goetzman, E. Jing, B. Schwer, D. B. Lombard, C. A. Grueter, C. Harris, S. Biddinger, O. R. Ilkayeva, R. D. Stevens, Y. Li, A. K. Saha, N. B. Ruderman, J. R. Bain, C. B. Newgard, R. V. Farese Jr., F. W. Alt, C. R. Kahn, and E. Verdin. 2010. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Na-ture 464:121–125.
Howarth, R. E., R. L. Baldwin, and M. Ronning. 1968. Enzyme ac-tivities in liver, muscle, and adipose tissue of calves and steers. J. Dairy Sci. 51:1270–1274.
Imai, S., and L. Guarente. 2010. Ten years of NAD-dependent SIR2 family deacetylases: Implications for metabolic diseases. Trends Pharmacol. Sci. 31:212–220.
Jin, D., H. J. Tan, T. Lei, L. Gan, X. D. Chen, Q. Q. Long, B. Feng, and Z. Q. Yang. 2009. Molecular cloning and characterization of porcine sirtuin genes. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 153:348–358.
Larkin, M. A., G. Blackshields, N. P. Brown, R. Chenna, P. A. Mc-Gettigan, H. McWilliam, F. Valentin, I. M. Wallace, A. Wilm, R. Lopez, J. D. Thompson, T. J. Gibson, and D. G. Higgins. 2007. Clustal W and Clustal X version 2.0. Bioinformatics 23:2947–2948.
Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25:402–408.
Marchler-Bauer, A., J. B. Anderson, F. Chitsaz, M. K. Derbyshire, C. DeWeese-Scott, J. H. Fong, L. Y. Geer, R. C. Geer, N. R. Gonzales, M. Gwadz, S. He, D. I. Hurwitz, J. D. Jackson, Z. Ke, C. J. Lanczycki, C. A. Liebert, C. Liu, F. Lu, S. Lu, G. H. Marchler, M. Mullokandov, J. S. Song, A. Tasneem, N. Thanki, R. A. Yamashita, D. Zhang, N. Zhang, and S. H. Bryant. 2009. CDD: Specific functional annotation with the Conserved Do-main Database. Nucleic Acids Res. 37:D205–D210.
Martin, R. J., L. L. Wilson, R. L. Cowan, and J. D. Sink. 1973. Ef-fects of fasting and diet on enzyme profiles in ovine liver and adipose tissue. J. Anim. Sci. 36:101–106.
Meiler, J., and D. Baker. 2003. Coupled prediction of protein sec-ondary and tertiary structure. Proc. Natl. Acad. Sci. USA 100:12105–12110.
Meiler, J., M. Mueller, A. Zeidler, and F. Schmaeschke. 2001. Gen-eration and evaluation of dimension reduced amino acid pa-rameter representations by artificial neural networks. J. Mol. Model. 7:360–369.
Miarelli, M., and F. Signorelli. 2010. Differential expression of liver proteins in Chianina and Holstein young bulls. J. Anim. Sci. 88:593–598.
Nafikov, R. A., and D. C. Beitz. 2007. Carbohydrate and lipid me-tabolism in farm animals. J. Nutr. 137:702–705.
Onyango, P., I. Celic, J. M. McCaffery, J. D. Boeke, and A. P. Feinberg. 2002. SIRT3, a human SIR2 homologue, is a NAD-dependent deacetylase localized to mitochondria. Proc. Natl. Acad. Sci. USA 99:13653–13658.
Picard, F., M. Kurtev, N. Chung, A. Topark-Ngarm, T. Senawong, R. Machado de Oliveira, M. Leid, M. W. McBurney, and L. Guarente. 2004. Sirt1 promotes fat mobilization in white adipo-cytes by repressing PPARγ. Nature 429:771–776.
Rodgers, J. T., C. Lerin, W. Haas, S. P. Gygi, B. M. Spiegelman, and P. Puigserver. 2005. Nutrient control of glucose homeosta-sis through a complex of PGC-1α and SIRT1. Nature 434:113–118.
Rodgers, J. T., and P. Puigserver. 2007. Fasting-dependent glucose and lipid metabolic response through hepatic sirtuin 1. Proc. Natl. Acad. Sci. USA 104:12861–12866.
SAGAR (Secretaría de Agricultura, Ganadería y Desarrollo Rural). 1997. Norma Oficial Mexicana NOM-033-ZOO-1995: Sacrificio humanitario de los animales domésticos y silvestres. Diario Ofi-cial de la Federación. 16 de Julio. México.
SAGAR (Secretaría de Agricultura, Ganadería y Desarrollo Rural). 1998. Norma Oficial Mexicana NOM-051-ZOO-1995: Trato hu-manitario en la movilización de animales. Diario Oficial de la Federación. 23 de Marzo. México.
SAGARPA (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación). 2001. Norma Oficial Mexicana NOM-062-ZOO-1999: Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. Diario Oficial de la Federación. 22 de Agosto. México.
Sanders, B. D., B. Jackson, and R. Marmorstein. 2010. Structural basis for sirtuin function: What we know and what we don’t. Biochim. Biophys. Acta 1804:1604–1616.
Saunders, L. R., and E. Verdin. 2007. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 26:5489–5504.
Schlicker, C., M. Gertz, P. Papatheodorou, B. Kachholz, C. F. W. Becker, and C. Steegborn. 2008. Substrates and regulation mechanisms for the human mitochondrial sirtuins Sirt3 and Sirt5. J. Mol. Biol. 382:790–801.
Schmittgen, T. D., and K. J. Livak. 2008. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3:1101–1108.
Schwer, B., B. J. North, R. A. Frye, M. Ott, and E. Verdin. 2002. The human silent information regulator (Sir)2 homologue hSIRT3 is a mitochondrial nicotinamide adenine dinucleotide-dependent deacetylase. J. Cell Biol. 158:647–657.
Shan, T., Y. Wang, T. Wu, C. Liu, J. Guo, Y. Zhang, J. Liu, and Z. Xu. 2009. Porcine sirtuin 1 gene clone, expression pattern, and regulation by resveratrol. J. Anim. Sci. 87:895–904.
Sherman, J. M., E. M. Stone, L. L. Freeman-Cook, C. B. Brach-mann, J. D. Boeke, and L. Pillus. 1999. The conserved core of a human SIR2 homologue functions in yeast silencing. Mol. Biol. Cell 10:3045–3059.
Shi, T., F. Wang, E. Stieren, and Q. Tong. 2005. SIRT3, a mi-tochondrial sirtuin deacetylase, regulates mitochondrial func-tion and thermogenesis in brown adipocytes. J. Biol. Chem. 280:13560–13567.
Shimada, A. 2003. Animal Nutrition. Ed. Trillas, Mexico City, Mex-ico.
Shoba, B., Z. M. Lwin, L. S. Ling, B. H. Bay, G. W. Yip, and S. D. Kumar. 2009. Function of sirtuins in biological tissues. Anat. Rec. (Hoboken) 292:536–543.
Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. Van Roy, A. De Paepe, and F. Speleman. 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averag-ing of multiple internal control genes. Genome Biol. 3:Re-search0034.1–0034.12.
Wang, F., and Q. Tong. 2009. SIRT2 suppresses adipocyte differen-tiation by deacetylating FOXO1 and enhancing FOXO1’s re-pressive interaction with PPARγ. Mol. Biol. Cell 20:801–808.
Yamamoto, H., K. Schoonjans, and J. Auwerx. 2007. Sirtuin func-tions in health and disease. Mol. Endocrinol. 21:1745–1755.
Yu, J., and J. Auwerx. 2009. The role of sirtuins in the control of metabolic homeostasis. Ann. N. Y. Acad. Sci. 1173:E10–E19.
Zimin, A. V., A. L. Delcher, L. Florea, D. R. Kelley, M. C. Schatz, D. Puiu, R. Hanrahan, G. Pertea, C. P. Van Tassell, T. S. Son-stegard, G. Marçais, M. Roberts, P. Subramanian, J. A. Yorke, and S. L. Salzberg. 2009. A whole-genome assembly of the do-mestic cow, Bos taurus. Genome Biol. 10:R42.
2536 Ghinis-Hozumi et al.
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from

Referenceshttp://www.journalofanimalscience.org/content/89/8/2529#BIBLThis article cites 42 articles, 20 of which you can access for free at:
by guest on September 13, 2014www.journalofanimalscience.orgDownloaded from
Related Documents











