Biol. Chem., Vol. 388, pp. 107–117, January 2007 • Copyright by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2007.012 2007/257 Article in press - uncorrected proof Binding of aflatoxins to the 20S proteasome: effects on enzyme functionality and implications for oxidative stress and apoptosis Manila Amici 1 , Valentina Cecarini 1 , Assuntina Pettinari 1 , Laura Bonfili 1 , Mauro Angeletti 1 , Simone Barocci 2 , Massimo Biagetti 2 , Evandro Fioretti 1 and Anna Maria Eleuteri 1, * 1 Department of Molecular, Cellular and Animal Biology, University of Camerino, I-62032 Camerino (MC), Italy 2 Istituto Zooprofilattico Sperimentale Umbria-Marche, I-06100 Perugia (PG), Italy * Corresponding author e-mail: [email protected] Abstract Aflatoxins (AF) are contaminants of improperly stored foods; they are potent genotoxic and carcinogenic com- pounds, exerting their effects through damage to DNA. They can also induce mutations that increase oxidative damage. The goal of this study was to evaluate the pos- sibility that a third mechanism could be involved in the carcinogenic action of aflatoxins, namely, direct binding to key enzymes involved in the regulatory pathways of the cell cycle, thereby modulating enzyme functionality. The 20S constitutive and immunoproteasome peptidase and proteolytic activities were assayed in the presence of aflatoxins B1, G1 and M1. All three toxins activated multiple peptidase activities of the proteasome. Aflatoxin (AF) M1 was the most potent activator of proteasome activity, while the constitutive 20S proteasome was spe- cifically stimulated by AFG1. Furthermore, the effects of AFB1 on cultured hepatoma cells were investigated and the various proteasomal activities determined with cell lysates were differently affected. Taking into account the key role of the proteasome in cellular defense against oxidative stress, the carbonyl group content and the activities of antioxidant enzymes in cell lysates were ana- lyzed. The proapoptotic effect of AFB1 was also inves- tigated by measuring caspase-3 activity and cellular levels of p27 and IkBa. Keywords: aflatoxin; apoptosis; HepG2; oxidative stress; proteasome; proteolytic activity. Introduction Aflatoxins are a group of mycotoxins produced by the fungi Aspergillus flavus and Aspergillus parasiticus, which grow on improperly stored foods such as rice and corn (Smela et al., 2001). There are different types of afla- toxins, but the predominant aflatoxins found as natural contaminants are aflatoxin (AF) B1, AFB2, AFM1, AFM2, AFG1 and AFG2. Aflatoxins B1, B2, G1 and G2 are pres- ent in rice and corn, whereas M1 and M2 are present in milk obtained from livestock that have ingested contam- inated feed and meat. Figure 1 shows the chemical struc- tures of the aflatoxins under investigation (AFB1, AFG1 and AFM1). It is almost impossible to get rid of aflatoxins from con- taminated food owing to the fact that they are highly sta- ble compounds. Although the approved levels of AF are low (a few ppb), they tend to accumulate in target organs, leading to chronic diseases such as carcinogenesis, mutagenesis, immunotoxicity and hepatotoxicity. More- over, AFB1 is considered to be one of the most potent liver carcinogens known in rats and mice (Steyn, 1995). AFM1 is a hydroxylated metabolite of AFB1 produced in the liver and is found in bovine milk; this biotransfor- mation does not change the genotoxic and mutagenic properties of the precursor. AFB1 has an immunosuppressive action that is partic- ularly evident in the cell-mediated immune response (Wogan, 1992). Several studies have also reported an increase in the formation of reactive oxygen species (ROS) in cultured hepatocytes after AFB1 treatment and the use of antioxidants and/or free radical scavengers able to inhibit the hepatotoxic effects (Kodama et al., 1990; Shen et al., 1995, 1996; Lee et al., 2001; Towner et al., 2003; Renzulli et al., 2004; Guerra et al., 2005). The effects of aflatoxins on the proteasome have not been elucidated to date. Because of the importance of protea- somes in several cell processes (Friguet et al., 1994; Grune et al., 1997, 2001; Baumeister et al., 1998; De- Martino and Slaughter, 1999; Keller et al., 2000; Orlowski and Wilk, 2000; Davies, 2001; Szweda et al., 2002), it is possible that many of the actions of aflatoxins may be mediated by their interactions with the pro- teasome. The purpose of this study was to investigate the effects of aflatoxins on the 20S proteasome. The proteasome is a large, barrel-shaped, multicatalytic protease, present in both the nucleus and cytosol. It comprises approximately 1% of total cellular protein and is found in all eukaryotic cells (Tanaka et al., 1986; Hilt et al., 1993; Ciechanover, 1994; Rock et al., 1994; Tanaka, 1995). The catalytic core is composed of 28 subunits, 14 a- type and 14 b-type, organized in four rings. The two inner rings contain b-subunits, and the outer rings contain a-subunits. The three constitutive b-subunits X, Y and Z are responsible for mediating the proteolytic activities of the proteasome and, on exposure to g-interferon, they are replaced by three homologous subunits, LMP7, LMP2 and MECL-1. This new structure, assembling the inducible b-subunits, is termed immunoproteasome (Baumeister et al., 1998; DeMartino and Slaughter, 1999; Orlowski and Wilk, 2000).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biol. Chem., Vol. 388, pp. 107–117, January 2007 • Copyright � by Walter de Gruyter • Berlin • New York. DOI 10.1515/BC.2007.012
2007/257
Article in press - uncorrected proof
Binding of aflatoxins to the 20S proteasome: effects onenzyme functionality and implications for oxidative stressand apoptosis
Manila Amici1, Valentina Cecarini1, AssuntinaPettinari1, Laura Bonfili1, Mauro Angeletti1,Simone Barocci2, Massimo Biagetti2, EvandroFioretti1 and Anna Maria Eleuteri1,*1 Department of Molecular, Cellular and Animal Biology,University of Camerino, I-62032 Camerino (MC), Italy2 Istituto Zooprofilattico Sperimentale Umbria-Marche,I-06100 Perugia (PG), Italy
* Corresponding authore-mail: [email protected]
Abstract
Aflatoxins (AF) are contaminants of improperly storedfoods; they are potent genotoxic and carcinogenic com-pounds, exerting their effects through damage to DNA.They can also induce mutations that increase oxidativedamage. The goal of this study was to evaluate the pos-sibility that a third mechanism could be involved in thecarcinogenic action of aflatoxins, namely, direct bindingto key enzymes involved in the regulatory pathways ofthe cell cycle, thereby modulating enzyme functionality.The 20S constitutive and immunoproteasome peptidaseand proteolytic activities were assayed in the presenceof aflatoxins B1, G1 and M1. All three toxins activatedmultiple peptidase activities of the proteasome. Aflatoxin(AF) M1 was the most potent activator of proteasomeactivity, while the constitutive 20S proteasome was spe-cifically stimulated by AFG1. Furthermore, the effects ofAFB1 on cultured hepatoma cells were investigated andthe various proteasomal activities determined with celllysates were differently affected. Taking into account thekey role of the proteasome in cellular defense againstoxidative stress, the carbonyl group content and theactivities of antioxidant enzymes in cell lysates were ana-lyzed. The proapoptotic effect of AFB1 was also inves-tigated by measuring caspase-3 activity and cellularlevels of p27 and IkBa.
Keywords: aflatoxin; apoptosis; HepG2; oxidativestress; proteasome; proteolytic activity.
Introduction
Aflatoxins are a group of mycotoxins produced by thefungi Aspergillus flavus and Aspergillus parasiticus, whichgrow on improperly stored foods such as rice and corn(Smela et al., 2001). There are different types of afla-toxins, but the predominant aflatoxins found as naturalcontaminants are aflatoxin (AF) B1, AFB2, AFM1, AFM2,AFG1 and AFG2. Aflatoxins B1, B2, G1 and G2 are pres-
ent in rice and corn, whereas M1 and M2 are present inmilk obtained from livestock that have ingested contam-inated feed and meat. Figure 1 shows the chemical struc-tures of the aflatoxins under investigation (AFB1, AFG1and AFM1).
It is almost impossible to get rid of aflatoxins from con-taminated food owing to the fact that they are highly sta-ble compounds. Although the approved levels of AF arelow (a few ppb), they tend to accumulate in target organs,leading to chronic diseases such as carcinogenesis,mutagenesis, immunotoxicity and hepatotoxicity. More-over, AFB1 is considered to be one of the most potentliver carcinogens known in rats and mice (Steyn, 1995).
AFM1 is a hydroxylated metabolite of AFB1 producedin the liver and is found in bovine milk; this biotransfor-mation does not change the genotoxic and mutagenicproperties of the precursor.
AFB1 has an immunosuppressive action that is partic-ularly evident in the cell-mediated immune response(Wogan, 1992). Several studies have also reported anincrease in the formation of reactive oxygen species(ROS) in cultured hepatocytes after AFB1 treatment andthe use of antioxidants and/or free radical scavengersable to inhibit the hepatotoxic effects (Kodama et al.,1990; Shen et al., 1995, 1996; Lee et al., 2001; Towneret al., 2003; Renzulli et al., 2004; Guerra et al., 2005). Theeffects of aflatoxins on the proteasome have not beenelucidated to date. Because of the importance of protea-somes in several cell processes (Friguet et al., 1994;Grune et al., 1997, 2001; Baumeister et al., 1998; De-Martino and Slaughter, 1999; Keller et al., 2000; Orlowskiand Wilk, 2000; Davies, 2001; Szweda et al., 2002), it ispossible that many of the actions of aflatoxins may bemediated by their interactions with the pro-teasome.
The purpose of this study was to investigate the effectsof aflatoxins on the 20S proteasome. The proteasome isa large, barrel-shaped, multicatalytic protease, present inboth the nucleus and cytosol. It comprises approximately1% of total cellular protein and is found in all eukaryoticcells (Tanaka et al., 1986; Hilt et al., 1993; Ciechanover,1994; Rock et al., 1994; Tanaka, 1995).
The catalytic core is composed of 28 subunits, 14 a-type and 14 b-type, organized in four rings. The two innerrings contain b-subunits, and the outer rings containa-subunits. The three constitutive b-subunits X, Y and Zare responsible for mediating the proteolytic activities ofthe proteasome and, on exposure to g-interferon, theyare replaced by three homologous subunits, LMP7,LMP2 and MECL-1. This new structure, assembling theinducible b-subunits, is termed immunoproteasome(Baumeister et al., 1998; DeMartino and Slaughter, 1999;Orlowski and Wilk, 2000).

108 M. Amici et al.
Article in press - uncorrected proof
Figure 1 Chemical structure of aflatoxins B1, M1 and G1.
In the eukaryotic proteasome, three different proteo-lytic components can be distinguished: the chymotryp-sin-like (ChT-L) component related to the b5 subunit (X/LMP7), the trypsin-like (T-L) component associated withthe b2 subunit (Z/MECL1), and the peptidyl-glutamylpeptide hydrolase (PGPH) expressed by the b1 subunit(Y/LMP2). In addition, two other activities have beencharacterized: the BrAAP (branched-chain amino acidpreferring) and the SNAAP (small neutral amino acid pre-ferring) activities (Orlowski, 1990; Cardozo, 1993; Bau-meister et al., 1998; Orlowski and Wilk, 2000). It has beendemonstrated that BrAAP activity is expressed by sub-unit b5i (LMP7) when it replaces subunit b5 (X) (Eleuteriet al., 1997; Orlowski et al., 1997).
The 20S proteasome also plays a major role in the reg-ulation of transcription factors, such as NF-kB; in fact,its inhibitor, IkB, is degraded by the complex (Adams,2003). In this regard, it has been demonstrated that afungal toxin, gliotoxin, has an inhibitory effect on 20Sproteasome functionality, in particular ChT-L peptidaseactivity (Kroll et al., 1999).
Therefore, aflatoxins might exert their cytotoxicitythrough various pathways, including promoting an in-crease in oxidative stress and modifying the proteasomefunctionality.
Results
Effects of aflatoxins on 20S proteasome activities
We investigated the effect of aflatoxins AFB1, AFM1 andAFG1 on the peptidase and proteolytic activities of con-stitutive and interferon-g (IFN-g)-inducible 20S protea-somes isolated from bovine brain and thymus, respec-tively. The effects of AFB1 on HepG2 cells were alsoevaluated over time. Even though the most potent afla-toxin in our model was AFM1 (as shown below), wechose to treat cells with AFB1 because it is the toxinproduced by fungi, and therefore is the main foodcontaminant.
Figure 2A,B shows the ChT-L, T-L, PGPH and BrAAPactivities of both proteasomes in the presence of increas-ing toxin concentrations (from 0 to 1.0 mM). Aflatoxinconcentrations were chosen based on their acceptablelimits, which are within the nanomolar range, and theirbioaccumulation over time in target organs (Williams etal., 2004).
An overall activation effect was evident, with 0.1 mM
being the lowest activating aflatoxin concentration: theChT-L component was the most sensitive to the toxins,especially to AFM1 (Figure 2A,B). The T-L activity did notchange in the presence of AFB1, while it increased in theconstitutive proteasome upon AFG1 and AFM1 treatment(Figure 2A). The PGPH component was only significantlyactivated in the constitutive proteasome in the presenceof AFM1 (Figure 2A). The BrAAP activity was never influ-enced by the toxins (Figure 2A,B); control assays showedthat none of the three toxins affected aminopeptidase-N(AP-N) activity (data not shown). This enzyme is used ina coupled enzyme reaction to measure BrAAP activity. Infact, since the 20S proteasome cleaves bonds betweenP1 and P2 amino acid residues, AP-N is responsible forcleavage of the arylamide bond between the P1 aminoacid residue and the aromatic amine, releasing a freefluorophore group (Orlowski et al., 1993).
AFM1 induced the most remarkable effect, eventhough it was neither proteasome- nor component-spe-cific. AFG1 did not affect proteasomes functionality,except for the T-L component in the constitutive protea-some (Figure 2A) and ChT-L in the immunoproteasome(Figure 2B). Moreover, interactions between AFM1 and20S proteasomes were observed to be reversible. In fact,upon dialyzing solutions of the enzymes and aflatoxin,the 20S proteasomes completely recovered their enzy-matic activity (Table 1).
Kinetic studies were also performed to estimatechanges in Km and Vmax upon treatment with 0.8 mM
AFM1 of the constitutive proteasome T-L component andthe immunoproteasome ChT-L. Results obtained fromkinetic data fitting for increasing substrate concentrationsaccording to the Michaelis-Menten equation are sum-marized in Table 2. Treatment with AFM1 did not affectVmax values, whereas a decrease in Km values wasinduced for both ChT-L and T-L, with a higher effect forthe former.
Determination of casein degradation by 20Sproteasomes
The proteolytic activity of 20S proteasomes was assayedusing b-casein as a macromolecular substrate. Theeffects of 0.1 mM aflatoxin, the lowest activating concen-tration, on the caseinolytic activity of the constitutive (A)and immunoproteasomes (B) are presented in Figure 3.
The constitutive 20S proteasome completely degradedb-casein in the presence and absence of toxins, even ifAFB1 inhibited the degradation rate within a 30 min incu-bation time. The immunoproteasome showed lower pro-teolytic activity that was enhanced in the presence ofaflatoxins, with AFB1 and AFM1 producing the strongestactivation.
Moreover, the chromatographic profiles obtained in thepresence and absence of aflatoxins showed no differ-

20S proteasome and aflatoxins 109
Article in press - uncorrected proof
Figure 2 Effect of increasing concentrations of aflatoxins B1 (�), M1 (j) and G1 (m) on the peptidase activities of the constitutive(A) and immuno (B) 20S proteasomes.Samples of 5 mg of enzyme were incubated with the appropriate substrate in 50 mM Tris-HCl, pH 8.0 to a final volume of 500 ml.Incubation was carried out at 378C and after 60 min the hydrolyzed 7-amino-4-methyl-coumarin (AMC) and 4-aminobenzoic acid(PABA) were measured (AMC: lexcs365 nm, lems449 nm; PABA: lexcs304 nm, lems664 nm) on a Shimadzu RF5301 spectrofluori-meter. Each data point is the mean and standard error of results from at least four separate experiments.
Table 1 Reversibility of the interaction of AFM1 with the constitutive 20S proteasomeand the immunoproteasome.
ChT-L immunoproteasome T-L constitutive 20Sproteasome
0 mM AFM1 0.8 mM AFM1 0 mM AFM1 0.8 mM AFM1
Before dialysis 101.32"0.33 445.46"12.11 732.49"9.76 1008.7"19.45After dialysis 95.88"0.93 103.43"0.43 769.24"6.84 782.67"11.14
Results are reported as fluorescence arbitrary units.
Table 2 Kinetic constants exhibited by the ChT-L of the immunoproteasome and the T-L of the constitutive 20S protea-some, treated with 0.8 mM AFM1.
AFM1 ChT-L immunoproteasome T-L constitutive 20S proteasome
Vmax (mol min-1) Km (M) Vmax (mol min-1) Km (M)
0 mM 3.2143=10-5"2.89=10-7 7.9431=10-5"2.30=10-6 1.4062=10-4"2.79=10-5 1.0543=10-5"4.01=10-6
0.8 mM 3.6851=10-5"1.55=10-7 1.5264=10-5"3.83=10-7 2.7541=10-4"1.231=10-5 0.4826=10-5"5.49=10-7

110 M. Amici et al.
Article in press - uncorrected proof
Figure 3 Effect of 0.1 mM aflatoxins B1 (�), M1 (j) and G1(m) on casein degradation by the constitutive (A) and immuno(B) 20S proteasomes at different incubation times.Control experiments were performed in the absence of toxins(h). After HPLC elution, the casein peak area (ls210 nm) atdifferent incubation times is reported as percentage of the con-trol (casein peak obtained for 0 min incubation). Each data pointis the mean and standard error for three separate deter-minations.
Figure 4 Cell survival analyzed with the MTT assay.HepG2 cells were treated with 0, 0.5, 1 and 10 mM AFB1 for 3,6, 12, 24 and 48 h. Data are expressed as a percentage of therespective controls in the absence of toxin. Data are the meanand standard error of results from at least six cultures from fiveseparate experiments. *p-0.05, **p-0.01, ***p-0.001 comparedto the corresponding control cells without AFB1.
ences, which means that the pattern of degradationdetermined by RP-HPLC was not altered by aflatoxins.
Cell viability
Treating the cells with increasing AFB1 concentrationscaused essentially no change in HepG2 cell viability, upto 1 mM aflatoxin, with exposed cells not less than 85%of the corresponding controls. At 10 mM AFB1, de-creased cell survival was observed after 24 h of treat-ment, with cell death observed at 48 h (Figure 4).
Effect of increasing AFB1 concentrations on HepG2proteasomal activities
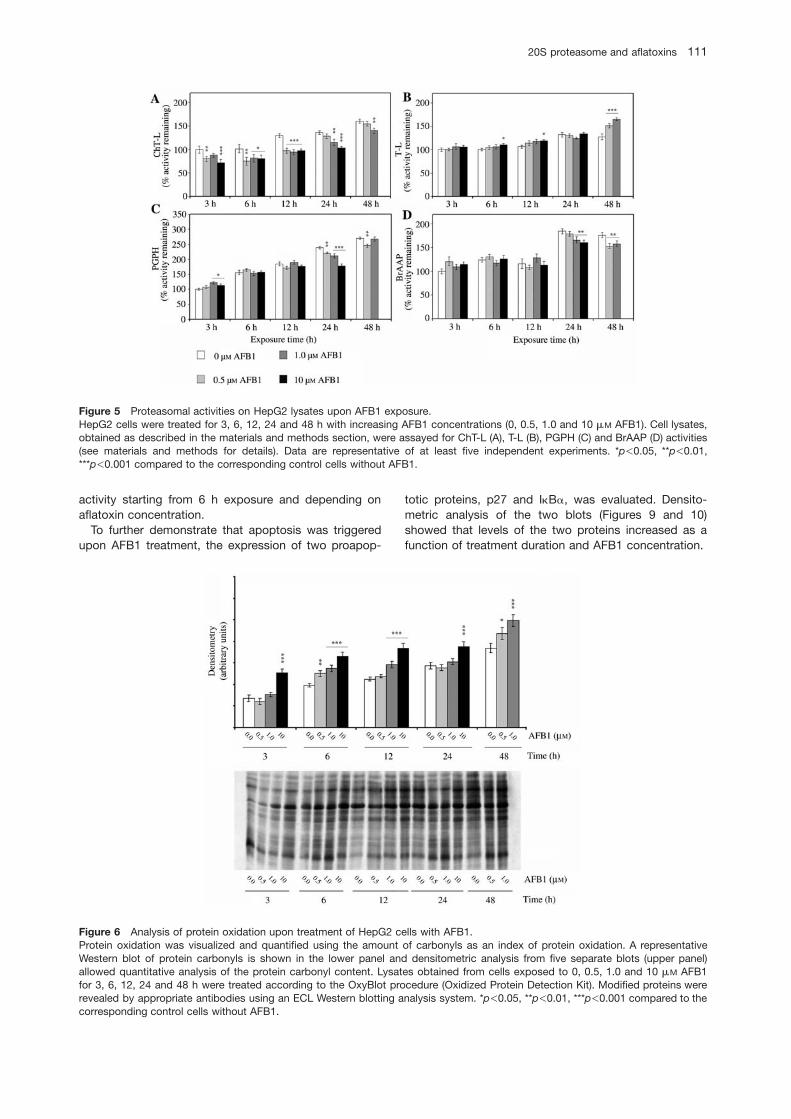
Activity assays were also performed with cell lysates onexposure to 0, 0.5, 1.0 and 10 mM AFB1 for 3, 6, 12, 24and 48 h (Figure 5A–D).
The ChT-L component was always inhibited (Figure5A); T-L activity showed activation at 10 mM AFB1, whichwas particularly evident at 48 h (Figure 5B). Except forthe 3 h exposure time, PGPH activity was inhibited (Fig-ure 5C), whereas BrAAP activity decreased only after 24and 48 h of treatment (Figure 5D).
Western blotting analysis using antibodies to ‘core’subunits of the human 20S proteasome showed no dif-ferences in the expression levels of the complex in celllysates (data not shown).
Determination of aflatoxin-induced oxidative effect
There is evidence that aflatoxin B1-induced cytotoxicityand carcinogenicity are related to oxidative damageinduced by an increase in the formation of ROS in cul-tured cells treated with AFB1 (Kodama et al., 1990; Shenet al., 1995, 1996; Towner et al., 2003). To evaluate thepotential oxidative effect of AFB1, AFM1 and AFG1 onisolated 20S proteasomes, immunoblotting analyseswere carried out to measure the amount of carbonylgroups in the two complexes treated with increasing tox-ins concentrations (see materials and methods). Theresults revealed that aflatoxins did not affect the oxida-tive state of the two enzymes (data not shown). However,an increase in carbonyl groups in cell lysates was detect-ed as a function of AFB1 concentration (Figure 6).
To further assess the prooxidant effect caused byAFB1, the activities of antioxidant enzymes such as glu-tathione reductase, glutathione S-transferase and cata-lase were measured. As shown in Figure 7A, theglutathione reductase activity was significantly inhibitedafter 24 and 48 h and was related to the toxin concen-tration, contributing to an increase in cellular oxidativestress (Meki et al., 2004). Glutathione S-transferase (Fig-ure 7B), which also plays a key role in AFB1 detoxifica-tion pathways, showed marked activation at longerexposure times, demonstrating an attempt by hepato-cytes to resist ROS and AFB1 toxicity (Rastogi et al.,2006). Catalase (Figure 6C) activity decreased as a func-tion of treatment duration, indirectly enhancing cellularsusceptibility to ROS toxic effects (Rastogi et al., 2001).
Evaluation of proapoptotic effects
Caspase-3, a cysteine protease that plays a major rolein the apoptotic process, is considered a marker of apop-tosis. It has been accepted that oxidative stress inducesapoptosis (Chandra et al., 2000); in fact many com-pounds that are able to induce programmed cell deathare oxidants or stimulators of cellular oxidative metabo-lism (Freeman and Grapo, 1982). The level of caspase-3in AFB1-treated cells compared to control cells wasdetermined. Figure 8 shows an increase in caspase-3
20S proteasome and aflatoxins 111
Article in press - uncorrected proof
Figure 5 Proteasomal activities on HepG2 lysates upon AFB1 exposure.HepG2 cells were treated for 3, 6, 12, 24 and 48 h with increasing AFB1 concentrations (0, 0.5, 1.0 and 10 mM AFB1). Cell lysates,obtained as described in the materials and methods section, were assayed for ChT-L (A), T-L (B), PGPH (C) and BrAAP (D) activities(see materials and methods for details). Data are representative of at least five independent experiments. *p-0.05, **p-0.01,***p-0.001 compared to the corresponding control cells without AFB1.
Figure 6 Analysis of protein oxidation upon treatment of HepG2 cells with AFB1.Protein oxidation was visualized and quantified using the amount of carbonyls as an index of protein oxidation. A representativeWestern blot of protein carbonyls is shown in the lower panel and densitometric analysis from five separate blots (upper panel)allowed quantitative analysis of the protein carbonyl content. Lysates obtained from cells exposed to 0, 0.5, 1.0 and 10 mM AFB1for 3, 6, 12, 24 and 48 h were treated according to the OxyBlot procedure (Oxidized Protein Detection Kit). Modified proteins wererevealed by appropriate antibodies using an ECL Western blotting analysis system. *p-0.05, **p-0.01, ***p-0.001 compared to thecorresponding control cells without AFB1.
activity starting from 6 h exposure and depending onaflatoxin concentration.
To further demonstrate that apoptosis was triggeredupon AFB1 treatment, the expression of two proapop-
totic proteins, p27 and IkBa, was evaluated. Densito-metric analysis of the two blots (Figures 9 and 10)showed that levels of the two proteins increased as afunction of treatment duration and AFB1 concentration.

112 M. Amici et al.
Article in press - uncorrected proof
Figure 7 Activities of antioxidant enzymes in HepG2 cellstreated with AFB1.(A) Glutathione reductase (GR), (B) glutathione S-transferase(GST) and (C) catalase activities were determined in lysatesobtained from cells treated with 0, 0.5, 1.0 and 10 mM AFB1 for3, 6, 12, 24 and 48 h as described under materials and methods.Data are reported as percentage specific activity of the control(3 h and 0 mM AFB1). Results are representative of at least fiveindependent experiments. *p-0.05, **p-0.01, ***p-0.001 com-pared to the corresponding control cells without AFB1.
Figure 8 Caspase-3 activity in HepG2 cells treated with AFB1.Caspase-3 activity was measured in lysates obtained from cellstreated with 0, 0.5, 1.0 and 10 mM AFB1 for 3, 6, 12, 24 and48 h as described under materials and methods. Data arereported as percentage specific activity of the control (3 h and0 mM AFB1). Results are representative of at least five inde-pendent experiments. *p-0.05, **p-0.01, ***p-0.001 comparedto the corresponding control cells without AFB1.
Moreover, the increase in these two proteins could alsobe related to proteasome inhibition, since they are bothsubstrates for the proteasome.
Discussion
The aim of this work was to investigate the possibility ofa direct interaction between aflatoxins and 20S pro-teasomes.
Data obtained in this study show an activating effectof the three aflatoxins on the activities of isolated pro-teasomes, particularly towards ChT-L, T-L and PGPHactivities. The most activating toxin is AFM1, whichshowed a higher effect on the T-L component of the con-stitutive proteasome. AFG1 was able to discriminate
between the two proteasomes. In fact, AFG1 increasedonly the T-L activity of the constitutive complex and theChT-L activity of the IFN-g-inducible proteasome. Fur-thermore, these interactions were demonstrated to bereversible. The lowest activating aflatoxin concentrationwas 0.1 mM, which is higher than the acceptable limits.However, we believe it is still physiologically relevantbecause it is known aflatoxins accumulate over time intarget organs (Williams et al., 2004). Surprisingly, BrAAPactivity of isolated proteasomes was not affected by thethree toxins. Aflatoxins activate three proteasomal com-ponents, but this phenomenon could not be related todirect binding to the three subunits. In fact, aflatoxinsmight either bind to one subunit and then allostericallyactivate the other two, or bind to non catalytic modifiersites, producing an allosteric effect, as has already beendemonstrated for Ritonavir by Schmidtke et al. (1999).
Conversely, an inhibition trend was observed onHepG2 proteasomal activities, which was more evidentat longer exposure times. In fact, except for an increasein the T-L component, all other activities decreased as afunction of aflatoxin concentration. ChT-L activity was themost sensitive and the BrAAP component, different tothe in vitro behavior, showed inhibition at 24 and 48 h. Itis therefore likely that the immunosuppressive action ofaflatoxins may be mediated by altering antigenic peptideproduction (Cardozo et al., 1999). Interestingly, theresults obtained from in vitro experiments partially dis-agree with measurements on cell lysates. This might bebecause the ultimate effect in cells (mainly 20S protea-some inhibition) arises from multiple actions of AFB1 thatalter various cellular activities. Among these, AFB1affects several enzymes such as protein kinase (Van denHeever and Dirr, 1991; Mistry et al., 1996), Ca2q-ATPase(Adebayo et al., 1995) and cyclic nucleotide phospho-diesterase (Bonsi et al., 1999). Furthermore we do notknow which form of the toxin influenced the 20S protea-some in the cells, either AFB1 directly or its metaboliteAFM1.

20S proteasome and aflatoxins 113
Article in press - uncorrected proof
Figure 9 Influence of AFB1 treatment on p27 levels.A representative Western blot of p27 is presented (lower panel) and densitometric analysis from five separate blots (upper panel)allowed p27 quantification. HepG2 cells were treated with 0, 0.5, 1.0 and 10 mM AFB1 for 3, 6, 12, 24 and 48 h. Cell lysates wereanalyzed by immunoblotting using a rabbit anti-human p27 polyclonal antibody. Immunoblot detection was carried out using an ECLWestern blotting analysis system with a peroxidase-conjugated secondary antibody. *p-0.05, **p-0.01, ***p-0.001 compared to thecorresponding control cells without AFB1.
Figure 10 Influence of AFB1 treatment on IkBa levels.A representative Western blot of IkB-a is shown in the lower panel and densitometric analysis of five separate blots (upper panel)allowed IkBa quantification. HepG2 cells were treated with 0, 0.5, 1.0 and 10 mM AFB1 for 3, 6, 12, 24 and 48 h. Cell lysates wereanalyzed by immunoblotting using a rabbit anti-human IkBa (C-21):sc-371 polyclonal antibody. Immunoblot detection was carriedout using an ECL Western blotting analysis system with a peroxidase-conjugated secondary antibody. *p-0.05, **p-0.01, ***p-0.001compared to the corresponding control cells without AFB1.
Besides oxidative damage directly inflicted on DNA byfree radicals, there are other mechanisms by which freeradicals can cause cell death; in fact, an increase in oxi-dative proteins not paralleled by their prompt removalleads to the formation of protein aggregates that areextremely cytotoxic. Significant decreases in antioxidantenzyme activities, such as catalase and glutathionereductase, amplify the cytotoxic effect of aflatoxins.
Several studies have demonstrated oxidative damagein cultured cells upon AFB1 exposure (Kodama et al.,1990; Shen et al., 1995, 1996; Towner et al., 2003). Our
data show that none of the three toxins induced anincrease in carbonyl group content in the two isolatedproteasomes, suggesting that the toxins could have apro-oxidant effect that does not involve the proteasome.However, the increase in carbonyl group content inHepG2 lysates obtained from aflatoxin-treated cells wasdependent on AFB1 concentration. 20S proteasome inhi-bition could contribute to the high carbonyl content incell lysates. In fact, it is well established that the 20Scomplex is the proteolytic machinery responsible for pre-venting a build up of oxidized proteins. The results

114 M. Amici et al.
Article in press - uncorrected proof
obtained in this study support the oxidative challengecaused by aflatoxins, although oxidative stress re-sponses and the onset of cell damage occurred to dif-ferent extents according to the exposure duration andaflatoxin concentration.
Furthermore, analysis of validated biomarkers for ap-optosis, such as caspase-3 activation and increased lev-els of p27 and IkB-a, indicated that AFB1 triggers theapoptotic pathway in treated cells. The amount of pro-apoptotic proteins increased as a function of treatmentduration and AFB1 concentration, showing correlationwith the inhibition of ChT-L and BrAAP proteasomalactivities.
In conclusion, aflatoxin-mediated inhibition of the cel-lular 20S proteasomes may contribute to carcinogenesisthrough an increase in oxidative stress due to AFB1 dualeffects: inhibition of proteasome functionality and prooxi-dant action. In this model, the change in proteasome-mediated protein degradation contributes to alteredprotein expression. This may play a key role in the cyto-toxicity induced by aflatoxins, which, as demonstrated inthe present study, causes apoptotic cell death.
Materials and methods
Reagents and chemicals
Substrates for assaying the ChT-L, T-L, PGPH, aminopeptidaseN (AP-N), catalase, glutathione S-transferase, glutathione reduc-tase and caspase-3 activities (Suc-Leu-Leu-Val-Tyr-MCA, Z-Leu-Ser-Thr-Arg-MCA, Z-Leu-Leu-Glu-MCA, L-Leu-pNA, hydro-gen peroxide, 1-chloro-2,4-dinitrobenzene, b-NADPH, oxidizedglutathione, Ac-Asp-Glu-Val-Asp-AMC) were purchased fromSigma-Aldrich S.r.L. (Milano, Italy). The substrate Z-Gly-Pro-Ala-Phe-Gly-pAB to test BrAAP activity was a kind gift from Prof.Orlowski (Department of Pharmacology, Mount Sinai School ofMedicine, New York, USA). Aminopeptidase N (EC 3.4.11.2),used for the coupled assay utilized to detect BrAAP activity(Orlowski and Michaud, 1989), was purified from pig kidney asreported elsewhere (Pfleiderer, 1970; Almenoff and Orlowski,1983). Aflatoxins B1, M1 and G1 were purchased from Sigma-Aldrich and dissolved in dimethyl sulfoxide (DMSO). The mem-branes for Western blot analyses and Centricon YM-10 deviceswere purchased from Millipore (Milano, Italy). Proteins immobi-lized on films were detected with the Enhanced Chemilumines-cence (ECL) system (Amersham Pharmacia Biotech, Milano,Italy). An Oxidized Protein Kit (OxyBlot, Oncor Appligene, Stras-bourg, France) was used to detect carbonyl group levels.
Isolation and purification of 20S proteasomes
Isolation and purification of 20S proteasomes from bovine brainand thymus were carried out as previously reported (Orlowskiand Michaud, 1989; Eleuteri et al., 2000).
From immunoblot analyses using anti-X, Y, Z and anti-LMP2,LMP7, MECL1 antibodies (Biomol International LP, Exeter, UK),we observed that the constitutive 20S proteasome was predom-inantly expressed in the bovine brain whereas the immunopro-teasome was the prevalent form in the bovine thymus (Eleuteriet al., 2000; Amici et al., 2003).
Cell culture and cell extract preparation
The human hepatoblastoma (HepG2, Istituto ZooprofilatticoSperimentale – Lombardia ed Emilia Romagna, Italy) cell line
was maintained in minimum essential medium (MEM) (GibcoInvitrogen Cell Culture, Milano, Italy) supplemented with 10%fetal calf serum, 1% sodium pyruvate, 1% non-essential aminoacids, penicillin and streptomycin. Cells were maintained in a5% CO2 atmosphere at 378C. Cells were grown in 100-mm tis-sue culture dishes at an initial concentration of 2=106 cells/dish.After 24 h of starvation, cells were exposed to different concen-trations (0, 0.5, 1 and 10 mM) of AFB1 for 3, 6, 12, 24 and 48 h.
After removing the medium and washing with cold phosphate-buffered saline (PBS), cells were harvested in 4 ml of PBS andcentrifuged at 1600 g for 5 min. The pellet was resuspended ina lysis buffer (20 mM Tris, pH 7.4, 250 mM sucrose, 1 mM EDTAand 5 mM b-mercaptoethanol) and passed through a 25-gaugeneedle at least ten times. Lysates were centrifuged at 12 000 gfor 15 min and the supernatants were stored at -808C. Proteinconcentration in cell lysates was determined by the method ofBradford (1976) using bovine serum albumin as standard.
Cell viability
Cell viability was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) conversion as previouslydescribed (Mosmann, 1983; Holtsberg et al., 1998). Briefly, fol-lowing experimental treatment, cells were washed in PBS, pH7.5 and then MTT (final concentration 0.5 mg/ml) was added toculture medium without fetal calf serum and incubated for 2 hat 378C. The medium was then removed and replaced with100 ml of DMSO. The optical density was measured at 550 nmin a microtiter plate reader. At least six cultures were utilized foreach time point.
Determination of aflatoxin effects on 20Sproteasome activities
Assays of 20S proteasomes peptidase activities were carried outwith fluorogenic peptides. Suc-Leu-Leu-Val-Tyr-MCA was usedfor chymotrypsin-like activity, Z-Leu-Ser-Thr-Arg-MCA for tryp-sin-like activity, Z-Leu-Leu-Glu-MCA for peptidylglutamyl-pep-tide hydrolase activity, and Z-Gly-Pro-Ala-Phe-Gly-pAB forbranched-chain amino acid preferred activity (Wilk and Orlowski,1983; Orlowski et al., 1993; Eleuteri et al., 2000).
Aflatoxins were dissolved in DMSO and added to the reactionmixture to obtain different concentrations (0, 0.005, 0.01, 0.05,0.1, 0.2, 0.4, 0.6, 0.8 and 1.0 mM). The incubation mixture con-tained the aflatoxin, 5 mg of isolated 20S proteasome, theappropriate substrate and 50 mM Tris-HCl, pH 8.0 up to a finalvolume of 500 ml.
Activity assays with cell lysates (5 mg in the mixture) wereperformed using the substrates listed above; control experi-ments were carried out in the presence of the specific inhibitorsZ-Gly-Pro-Phe-Leu-CHO and lactacystin (5 mM in the reactionmixture).
Tests for BrAAP activity were performed in the presence ofAP-N (EC 3.4.11.2) (Orlowski et al., 1993). Incubation was car-ried out at 378C for 60 min. Hydrolyzed 7-amino-4-methyl-cou-marin (AMC) and 4-aminobenzoic acid (PABA) were thenmeasured (AMC: lexcs365 nm, lems449 nm; PABA: lexcs304nm, lems664 nm) on a Shimadzu RF5301 spectrofluorimeter.
Control assays were also performed to evaluate a possibleeffect of aflatoxins on the proteolytic activity of AP-N: incubationmixtures containing aflatoxin, 8 mg of AP-N, the substrate L-Leu-pNA and 50 mM Tris-HCl, pH 8.0 up to a final volume of500 ml were incubated at 378C. After 10 min of incubation, theactivity was measured (ls405 nm) on a Varian Cary 100Biospectrophotometer.
The reversibility of the interaction between toxins and isolated20S proteasomes was investigated in the presence of the mostactivating toxin, AFM1, at the concentration of 0.8 mM. The

20S proteasome and aflatoxins 115
Article in press - uncorrected proof
ChT-L activity of the immunoproteasome and the T-L componentof the constitutive proteasome were tested. After 30-min incu-bation with 0.8 mM AFM1 in 50 mM Tris-HCl, pH 8.0 at roomtemperature, the 20S proteasomes activities were assayed ac-cording to the procedures described above.
Preincubated enzymes were dialyzed and concentrated withTris-HCl 50 mM using Centricon YM-10 concentrators to removethe toxin. Activity assays were performed with the recoveredenzymes. Parallel experiments were performed in the absenceof aflatoxin, treating the enzymes with DMSO, and the reversi-bility was estimated by comparing the activity of the enzymebefore and after dialysis in the presence or absence of the toxin.
Kinetic studies
Kinetic runs were performed to evaluate the Vmax and Km para-meters for ChT-L activity of the 20S immunoproteasome and T-L activity of the 20S constitutive enzyme at increasing substratesconcentrations: 5–50 mM for Suc-Leu-Leu-Val-Tyr-MCA and0.5–15 mM for Z-Leu-Ser-Thr-Arg-MCA. In a typical kinetic run,1 ml of assay buffer (50 mM Tris-HCl, pH 8.0 and 0.8 mM AFM1)and the appropriate substrate were added to a fluorescencecuvette placed in the jacketed cell holder of a Shimadzu RF5301fluorescence spectrophotometer, maintained at 378C. After thereaction solution had reached thermal equilibrium (ca. 5 min),kinetic runs were started by adding the enzyme, preincubatedwith 0.8 mM AFM1. Reaction progress was monitored by theincrease in fluorescence emission (lexcs365 nm, lems449 nm)and the initial velocity was always calculated after 5 or 10 minreaction, when less than 10% of the substrate has been con-sumed. Control kinetics in the absence of aflatoxin were meas-ured. All kinetic data were obtained from at least four differentexperiments, in which eight different peptide concentrationswere used. The values for Vmax and Km parameters were obtainedby kinetic data fitting according to the Michaelis-Mentenequation.
Determination of casein degradation by 20Sproteasomes
The effect of aflatoxins on the proteolytic activity of isolated pro-teasomal complexes was evaluated using b-casein as proteinsubstrate. 20S proteasomes from bovine brain and thymus werepreincubated for 30 min at room temperature in 50 mM Tris-HCl,pH 8.0 in the presence of 0.1 mM aflatoxin.
Samples of 25.5 mg of treated proteasome were incubatedwith 100 mg of b-casein and 50 mM Tris-HCl, 20 mM MgCl2, pH7.5 to a final volume of 100 ml. The mixtures were kept at 378C,and 20-ml aliquots were withdrawn at different times (from 0 to60 min), acidified with 2 ml of 10% TCA, and subjected to HPLCon a Hamilton PRP-3 column (4.1=150 mm) on a Perkin-ElmerHPLC system. Sample elution and measurement of casein deg-radation were carried out as previously reported (Eleuteri et al.,1997; Amici et al., 2003). The rate of casein degradation wasdetermined by measuring the peak area of casein (ls210 nm).Control experiments were performed in the absence of afla-toxins. Each experimental set was repeated three times and rel-ative mean values and standard errors were calculated.
Immunoblot detection of carbonyl groups
An Oxidized Protein Kit (Oncor Appligene) was used to measurethe increase in carbonyl groups in isolated 20S proteasomes andcell lysates due to possible oxidative effects by aflatoxins. Brief-ly, 15 mg of 20S proteasome was incubated at room temperaturefor 45 min in the dark in 50 mM Tris-HCl, pH 8.00 with eachaflatoxin, dissolved in DMSO, to a final concentration of 0, 0.1and 1.0 mM.
Exposed enzymes and cell lysates (12 mg) were treatedaccording to the OxyBlot kit procedure, run on a 12% poly-acrylamide gel and blotted onto a PVDF membrane. Modifiedproteins were revealed by appropriate antibodies using an ECLWestern blotting analysis system (Amersham-PharmaciaBiotech).
Activity of antioxidant enzymes
The activity of antioxidant enzymes was measured in fresh celllysates using a Varian Cary 100Bio spectrophotometer at a con-stant temperature of 258C. Catalase (EC 1.11.1.6) was deter-mined by following the decrease in absorbance at 240 nm(´s0.04 mM-1 cm-1) due to hydrogen peroxide consumption (Clai-borne and Fridovich, 1979). The reaction was performed usinga solution of 20 mM hydrogen peroxide in 50 mM Tris-HCl, pH7.5 containing 40 mg of total cellular protein in a final volume of1 ml. Glutathione S-transferase (GST; EC 2.5.1.18) activity wasmeasured at 340 nm using 1-chloro-2,4-dinitrobenzene (CDNB)as substrate (´s9.6 mM-1 cm-1); the assay was carried out in100 mM potassium phosphate, pH 6.5, 1 mM reduced gluta-thione (GSH) and 1 mM CDNB, containing 40 mg of total cellularprotein (Habig et al., 1974). Glutathione reductase (GR; EC1.6.4.2) was determined at 340 nm, following the oxidation ofNADPH during the reduction of oxidized glutathione (GSSG;´s6.22 mM-1 cm-1). The reaction was carried out in 100 mM
potassium phosphate, pH 7.5, 1 mM EDTA containing 40 mg oftotal cellular protein, adding 0.1 mM NADPH and 1 mM GSSG.
Caspase-3 activity
Caspase-3 activity was assayed in cell lysates (20 mg of totalcellular protein) using Ac-Asp-Glu-Val-Asp-7-amino-4-methyl-coumarin (20 mM final concentration) as substrate. The assaywas performed in 50 mM Tris-HCl, pH 7.5, 50 mM NaCl, 1 mM
EDTA, 5 mM b-mercaptoethanol, 5 mM CaCl2, 0.1% CHAPS.Incubation was carried out at 378C for 60 min and then hydro-lyzed 7-amino-4-methyl-coumarin (AMC) was measured(lexcs365 nm, lems449 nm) on a Shimadzu RF5301 spec-trofluorimeter.
Western blotting
Cell lysates were resolved on 12% SDS-PAGE and electroblot-ted onto PVDF membranes. Membranes with transferred pro-teins were incubated with rabbit anti-human IkBa(C-21):sc-371(Santa Cruz Biotechnology, Inc., Milano, Italy), rabbit anti-humanp27 (Inalco S.p.A., Milano, Italy) polyclonal antibodies and rabbitpolyclonal antibodies to ‘core’ subunits of the human 20S pro-teasome (Biomol International LP). The immunoblot detectionwas carried out using an ECL Western blotting analysis system(Amersham Pharmacia Biotech) using peroxidase conjugatedsecondary antibodies (Sigma-Aldrich).
Statistical analysis
Values are expressed as mean and standard error of resultsobtained from separate experiments. Student’s t-test was usedto compare mean differences between control and treatedgroups.
Acknowledgments
This work was supported by a grant from the Istituto Zooprofi-lattico Sperimentale Umbria-Marche (Perugia), Ricerca Corrente2004.

116 M. Amici et al.
Article in press - uncorrected proof
References
Adams, J. (2003). Potential for proteasome inhibition in the treat-ment of cancer. Drug Discov. Today 8, 307–315.
Adebayo, A.O., Okunade, G.W., and Olorunsogo, O.O. (1995).The anticalmodulin effect of aflatoxin B1 on purified eryth-rocyte Ca2q-ATPase. Biosci. Rep. 15, 209–220.
Almenoff, J. and Orlowski, M. (1983). Membrane-bound kidneyneutral metalloendopeptidase: interaction with synthetic sub-strates, natural peptides, and inhibitors. Biochemistry 22,590–599.
Amici, M., Lupidi, G., Angeletti, M., Fioretti, E., and Eleuteri, A.M.(2003). Peroxynitrite-induced oxidation and its effects on iso-lated proteasomal systems. Free Radic. Biol. Med. 34, 987–996.
Baumeister, W., Walz, J., Zuhl, F., and Seemuller, E. (1998). Theproteasome: paradigm of a self-compartmentalizing prote-ase. Cell 92, 367–380.
Bonsi, P., Agusti-Tocco, G., Palmery, M., and Giorgi, M. (1999).Aflatoxin B1 is an inhibitor of cyclic nucleotide phosphodies-terase activity. Gen. Pharmacol. 32, 615–619.
Bradford, M.M. (1976). A rapid and sensitive method for thequantification of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72, 248–254.
Cardozo, C. (1993). Catalytic components of the bovine pituitarymulticatalytic proteinase complex (proteasome). EnzymeProtein 47, 296–305.
Cardozo, C., Michaud, C., and Orlowski, M. (1999). Componentsof the bovine pituitary multicatalytic proteinase complex (pro-teasome) cleaving bonds after hydrophobic residues. Bio-chemistry 38, 9768–9777.
Chandra, J., Samali, A., and Orrenius, S. (2000). Triggering andmodulation of apoptosis by oxidative stress. Free Radic. Biol.Med. 29, 323–333.
Ciechanover, A. (1994). The ubiquitin-proteasome proteolyticpathway. Cell 79, 13–21.
Claiborne, A. and Fridovich, I. (1979). Purification of the o-dia-nisidine peroxidase from Escherichia coli B. Physicochemicalcharacterization and analysis of its dual catalytic and peroxi-datic activities. J. Biol. Chem. 254, 4245–4255.
Davies, K.J.A. (2001). Degradation of oxidized proteins by the20S proteasome. Biochimie 83, 301–310.
DeMartino, G.N. and Slaughter, C.A. (1999). The proteasome, anovel protease regulated by multiple mechanisms. J. Biol.Chem. 274, 22123–22126.
Eleuteri, A.M., Kohanski, R.A., Cardozo, C., and Orlowski, M.(1997). Bovine spleen multicatalytic proteinase complex (pro-teasome). Replacement of X, Y, and Z subunits by LMP7,LMP2, and MECL1 and changes in properties and specificity.J. Biol. Chem. 272, 11824–11831.
Eleuteri, A.M., Angeletti, M., Lupidi, G., Tacconi, R., Bini, L., andFioretti, E. (2000). Isolation and characterization of bovinethymus multicatalytic proteinase complex. Protein Expr.Purif. 18, 160–168.
Freeman, B.A. and Grapo, J.D. (1982). Biology of disease: freeradical and tissue injury. Lab. Invest. 47, 412–426.
Friguet, B., Szweda, L.I., and Stadtman, E.R. (1994). Suscepti-bility of glucose-6-phosphate dehydrogenase modified by 4-hydroxy-2-nonenal and metal-catalyzed oxidation to pro-teolysis by the multicatalytic protease. Arch. Biochem. Bio-phys. 311, 168–173.
Grune, T., Reinheckel, T., and Davies, K.J.A. (1997). Degradationof oxidized proteins in mammalian cells. FASEB J. 11, 526–534.
Grune, T., Klotz, L.O., Gieche, J., Rudeck, M., and Sies, H.(2001). Protein oxidation and proteolysis by the nonradicaloxidants singlet oxygen or peroxynitrite. Free Radic. Biol.Med. 30, 1243–1253.
Guerra, M.C., Galvano, F., Bonsi, L., Speroni, E., Costa, S., Ren-zulli, C., and Cervellati, R. (2005). Cyanidin-3-O-b-glucopy-ranoside, a natural free-radical scavenger against aflatoxinB1- and ochratoxin A-induced cell damage in a human hepa-toma cell line (Hep G2) and a human colonic adenocarcino-ma cell line (CaCo-2). Br. J. Nutr. 94, 211–220.
Habig, W.H., Pabst, M.J., and Jakoby, W.B. (1974). GlutathioneS-transferases. The first enzymatic step in mercapturic acidformation. J. Biol. Chem. 249, 7130–7139.
Hilt, W., Heinemeyer, W., and Wolf, D.H. (1993). Studies on theyeast proteasome uncover its basic structural features andmultiple in vivo functions. Enzyme Protein 47, 189–201.
Holtsberg, F.W., Steiner, M.R., Keller, J.N., Mark, R.J., Mattson,M.P., and Steiner, S.M. (1998). Lysophosphatidic acid induc-es necrosis and apoptosis in hippocampal neurons. J. Neu-rochem. 70, 66–76.
Keller, J.N., Huang, F.F., Zhu, H., Yu, J., Ho, Y.S., and Kindy, T.S.(2000). Oxidative stress-associated impairment of protea-some activity during ischemia-reperfusion injury. J. Cereb.Blood Flow Metab. 20, 1467–1473.
Kodama, M., Inoue, F., and Akao, M. (1990). Enzymatic and non-enzymatic formation of free radicals from aflatoxin B1. FreeRadic. Res. Commun. 10, 137–142.
Kroll, M., Arenzana-Seisdedos, F., Bachelerie, F., Thomas, D.,Friguet, B., and Conconi, M. (1999). The secondary fungalmetabolite gliotoxin targets proteolytic activities of the pro-teasome. Chem. Biol. 6, 689–698.
Lee, S.E., Campbell, B.C., Molyneux, R.J., Hasegawa, S., andLee, H.S. (2001). Inhibitory effects of naturally occurring com-pounds on aflatoxin B1 biotransformation. J. Agric. FoodChem. 49, 5171–5177.
Meki, A.M.A., Esmail, E.E.F., Hussein, A.A., and Hassanein, H.M.(2004). Caspase-3 and heat shock protein-70 in rat livertreated with aflatoxin B1: effect of melatonin. Toxicon 43,93–100.
Mistry, K.J., Krishna, M., Pasupathy, K., Murthy, V., and Bhat-tacharya, R.K. (1996). Signal transduction mechanism inresponse to aflatoxin B1 exposure: protein kinase C activity.Chem. Biol. Interact. 100, 177–185.
Mosmann, T. (1983). Rapid colorimetric assay for cellular growthand survival: application to proliferation and cytotoxicityassays. J. Immunol. Methods 65, 55–63.
Orlowski, M. (1990). The multicatalytic proteinase complex, amajor extralysosomal proteolytic system. Biochemistry 29,10289–10297.
Orlowski, M. and Michaud, C. (1989). Pituitary multicatalytic pro-teinase complex. Specificity of components and aspects ofproteolytic activity. Biochemistry 28, 9270–9278.
Orlowski, M. and Wilk, S. (2000). Catalytic activities of the 20Sproteasome, a multicatalytic proteinase complex. Arch. Bio-chem. Biophys. 383, 1–16.
Orlowski, M., Cardozo, C., and Michaud, C. (1993). Evidence forthe presence of five distinct proteolytic components in thepituitary multicatalytic proteinase complex. Properties of twocomponents cleaving bonds on the carboxyl side ofbranched chain and small neutral amino acids. Biochemistry32, 1563–1572.
Orlowski, M., Cardozo, C., Eleuteri, A.M., Kohanski, R., Kam,C.M., and Powers, J.C. (1997). Reactions of w14Cx-3,4-dichlo-roisocoumarin with subunits of pituitary and spleen multica-talytic proteinase complexes (proteasomes). Biochemistry36, 13946–13953.
Pfleiderer, G. (1970). Isolation of an aminopeptidase from kidneyparticles. Methods Enzymol. 19, 514–521.
Rastogi, R., Srivastava, A.K., and Rastogi, A.K. (2001). Longterm effect of aflatoxin B1 on lipid peroxidation in rat liverand kidney: effect of picrovil and silymarin. Phytother. Res.15, 307–310.
Rastogi, S., Dogra, R.K.S., Khanna, S.K., and Das, M. (2006).Skin tumorigenic potential of aflatoxin B1 in mice. FoodChem. Toxicol. 44, 670–677.

20S proteasome and aflatoxins 117
Article in press - uncorrected proof
Renzulli, V, Galvano, F., Pierdomenico, L., Speroni, E., and Guer-ra, M.C. (2004). Effects of rosmarinic acid against aflatoxinB1 and ochratoxin-A-induced cell damage in a human hepa-toma cell line (Hep G2). J. Appl. Toxicol. 24, 289–296.
Rock, K.L., Gramm, C., Rothstein, L., Clark, K., Stein, R., Dick,L., Hwangand, D., and Goldberg, A.L. (1994). Inhibitors of theproteasome block the degradation of most cell proteins andthe generation of peptides presented on MHC class I mole-cules. Cell 78, 761–771.
Schmidtke, G., Holzhutter, H.G., Bogyo, M., Kairies, N., Groll,M., De Giuli, R., Emch, S., and Groettrup, M. (1999). How aninhibitor of the HIV-I protease modulates proteasome activity.J. Biol. Chem. 274, 35734–35740.
Shen, H.M., Ong, C.N., and Shi C.Y. (1995). Involvement of reac-tive oxygen species in aflatoxin B1-induced cell injury in cul-tured rat hepatocytes. Toxicology 99, 115–123.
Shen, H.M., Shi, C.Y., Shen, Y., and Ong, C.N. (1996). Detectionof elevated reactive oxygen species level in cultured rathepatocytes treated with aflatoxin B1. Free Radic. Biol. Med.21, 139–146.
Smela, M.E., Currier, S.S., Bailey, E.A., and Essigmann, J.M.(2001). The chemistry and biology of aflatoxin B1: from muta-tional spectrometry to carcinogenesis. Carcinogenesis 22,535–545.
Steyn, P.S. (1995). Mycotoxins, general view, chemistry andstructure. Toxicol. Lett. 82–83, 843–851.
Szweda, P.A., Friguet, B., and Szweda, L.I. (2002). Proteolysis,free radicals, and aging. Free Radic. Biol. Med. 33, 29–36.
Tanaka, K. (1995). Molecular biology of proteasomes. Mol. Biol.Rep. 21, 21–26.
Tanaka, K., Ii, K., Ichihara, A., Waxman, L., and Goldberg, A.L.(1986). A high molecular weight protease in the cytosol of ratliver. I. Purification, enzymological properties, and tissue dis-tribution. J. Biol. Chem. 26, 115197–15203.
Towner, R.A., Qian, S.Y., Kadiiska, M.B., and Mason, R.P. (2003).In vivo identification of aflatoxin-induced free radicals in ratbile. Free Radic. Biol. Med. 35, 1330–1340.
Van den Heever, L.H. and Dirr, H.W. (1991). Effect of aflatoxinB1 on human platelet protein kinase C. Int. J. Biochem. 23,839–843.
Wilk, S. and Orlowski, M. (1983). Evidence that pituitary cation-sensitive neutral endopeptidase is a multicatalytic proteasecomplex. J. Neurochem. 40, 842–849.
Williams, J.H., Phillips, T.D., Jolly, P.E., Stiles, J.K., Jolly, C.M.,and Aggarwal, D. (2004). Human aflatoxicosis in developingcountries: a review of toxicology, exposure, potential healthconsequences, and interventions. Am. J. Clin. Nutr. 80,1106–1122.
Wogan, G.N. (1992). Aflatoxins as risk factors for hepatocellularcarcinoma in humans. Cancer Res. 52, 2114s–2118s.
Received June 1, 2006; accepted July 10, 2006
Related Documents











