In Vitro Cell.Dev. Biol.--Plant 34:122-130, April-June 1998 9 1998 Society forIn Vitro Biology 1071-2690/98 $05.00+0.00 REVIEW BENEFITS OF IN VITRO "BIOTIZATION" OF PLANT TISSUE CULTURES WITH MICROBIAL INOCULANTS JERZY NOWAK Department of Plant Science, Nova Scotia Agricultural College, P.O. Box 550, Truro, Nova Scotia, Canada B2N 5E3 (Received 8 August 1997; accepted 18 December 1997; editor T. A. Thorpe) SUMMARY In vitro cocuhure of plant tissue explants with beneficial microorganisms induces developmental and metabolic changes in the derived plantlets which enhance their tolerance to abiotic and biotic stresses. The induced resistance response caused by the inoculants is referred to as "biotization." There is enough experimental evidence with bacteria (bacterization) and vesicular arbuscular mycorrhiza (mycorrhization) inoculations to recommend utilization of this technology in commercial micropropagation. The paper reviews literature on this topic, outlines other uses and potential uses of in vitro plant- microbial cocuhures, and discusses their current constraints. Key words: micropropagation; bacterization; mycorrhization; stress resistance responses. INTRODUCTION Plant propagation technology via tissue culture has been devel- oped over the last 30---40yr as a spinoff of in vitro studies on differ- entiation (Pierik, 1987, 1988). Typically, aseptic explants are grown under low light intensity in small containers, on artificial culture media containing sucrose, mineral salts, vitamins, and growth regu- lators, in concentrations exceeding levels recorded under natural en- vironments (Pierik, 1987; Leifert et al., 1995). Moreover, the culture conditions do not allow for a good gas and moisture exchange (McClelland and Smith, 1990; Jeong et al., 1995; Matthijs et al., 1995). This causes developmental distortions (Ziv, 1991; Van Huy- lenbroeck and Debergh, 1996) and repression or modulation of some metabolic pathways (Phan and Hegedus, 1986; Baccou, 1995; Des- jardins, 1995; Rchid and Baccou, 1995). Plantlets have reduced pho- tosynthetic capacity (Cournac et al., 1991, 1992; Desjardins, 1995), malfunctioning stomata (Ziv et al., 1987; Frommel et al., 1991a; Majada et al., 1995), root system devoid of root hairs, poor cuticle development, and low wax deposits (Pierik, 1987; Lee and Wetzstein, 1988; Preece and Sutter, 1991). Consequently, when transplanted out of the culture vessels, they suffer from severe environmental stresses and substantial losses may occur (Preece and Sutter, 1991; Van Huylenbroeck and Debergh, 1996). Manipulation of acclimation conditions prior to or upon transplanting usually reduced losses (Donnelly et al., 1985; Desjardins et al., 1988; Preece and Sutter, 1991; Desjardins, 1995; Van Huylenbroeck and Debergh, 1996), however, at additional cost to the producer. To minimize these con- straints, in recent years, attempts have been made to develop pho- toautotrophic culture systems with high light intensity and reduced or eliminated sugar in the medium (Kozai et al., 1988; Kozai, 1991; Fujiwara and Kozai, 1995; Jeong et al., 1995). So far, this technology has not gained popularity. During the last decade some researchers have reached for natural inhabitants of plants, epiphytes and endophytes, to enhance adap- tation of tissue culture propagules to environmental stresses (reported by Herman, 1996a, 1996b). The plant-associated microorganisms, endophytes in particular, were always treated as problem-causing contaminants of in vitro cultures (reviewed by Cassells, 1991, and Leifert et al., 1994) and various procedures have been developed for their elimination (Cassells, 1991; Reed and Tanprasert, 1995) or management (Levin et al., 1997). In nature, microorganisms inhabit the interior and exterior of plant organs (Jacobs et al., 1985; Frommel et al., 1993; Leifert et al., 1994; Mclnroy and Kloepper, 1995; Sturz, 1995; Sturz and Christie, 1995; Sturz et al., 1997). Some of these microorganisms, plant-beneficial bacteria and vesicular-arbuscular mycorrhizae in particular, can im- prove plant performance under stress environments and conse- quently, enhance yields (Brown, 1974; Schippers et al., 1987; Kloep- per et al., 1988, 1989; Lynch, 1990; Frommel et al., 1991b; Pfleger and Linderman, 1994; Chen et al., 1995; Datnoff et al., 1995; Glick, 1995; Balachandran et al., 1997; Bethlenfalvay et al., 1997; Creus et al., 1997; Jackson et al., 1997; Lazarovits and Nowak, 1997; Zehn- der et al., 1997). The mechanisms governing these responses have been extensively reviewed (e.g., Kloepper, 1988, 1989; Lynch, 1990; Dowling and O'Gara, 1994; Pfleger and Linderman, 1994; Glick, 1995, 1997; Lazarovits and Nowak, 1997). This review provides an outline of the present status of knowledge on utilization of microbial inoculants in plant tissue culture. IN VITRO "BIOTIZATION" Induction of stress resistance in plant propagules produced in vitro prior to transplanting, is a primary target of several researeh groups attempting utilization of microbial inoeulants in micropropagation (Hooker et al., 1994; Elmeskaoui et al., 1995; Nowak et al., 1995; Balla et al., 1997; Lazarovits and Nowak, 1997; Murphy et al., 1997; Wilhelm et al., 1997). Induced resistance responses to microbial inoeulants are well documented in plants grown in vivo (reviewed by Benhamou, 1996; Benhamou et al., 1996; Hunt and Ryals, 1996; Kloepper et al., 1997). Such responses in vitro have only been ree- 122

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

In Vitro Cell. Dev. Biol.--Plant 34:122-130, April-June 1998 �9 1998 Society for In Vitro Biology 1071-2690/98 $05.00+0.00
REVIEW BENEFITS OF IN VITRO "BIOTIZATION" OF PLANT TISSUE CULTURES WITH
MICROBIAL INOCULANTS
JERZY NOWAK
Department of Plant Science, Nova Scotia Agricultural College, P.O. Box 550, Truro, Nova Scotia, Canada B2N 5E3
(Received 8 August 1997; accepted 18 December 1997; editor T. A. Thorpe)
SUMMARY
In vitro cocuhure of plant tissue explants with beneficial microorganisms induces developmental and metabolic changes in the derived plantlets which enhance their tolerance to abiotic and biotic stresses. The induced resistance response caused by the inoculants is referred to as "biotization." There is enough experimental evidence with bacteria (bacterization) and vesicular arbuscular mycorrhiza (mycorrhization) inoculations to recommend utilization of this technology in commercial micropropagation. The paper reviews literature on this topic, outlines other uses and potential uses of in vitro plant- microbial cocuhures, and discusses their current constraints.
Key words: micropropagation; bacterization; mycorrhization; stress resistance responses.
INTRODUCTION
Plant propagation technology via tissue culture has been devel- oped over the last 30---40 yr as a spinoff of in vitro studies on differ- entiation (Pierik, 1987, 1988). Typically, aseptic explants are grown under low light intensity in small containers, on artificial culture media containing sucrose, mineral salts, vitamins, and growth regu- lators, in concentrations exceeding levels recorded under natural en- vironments (Pierik, 1987; Leifert et al., 1995). Moreover, the culture conditions do not allow for a good gas and moisture exchange (McClelland and Smith, 1990; Jeong et al., 1995; Matthijs et al., 1995). This causes developmental distortions (Ziv, 1991; Van Huy- lenbroeck and Debergh, 1996) and repression or modulation of some metabolic pathways (Phan and Hegedus, 1986; Baccou, 1995; Des- jardins, 1995; Rchid and Baccou, 1995). Plantlets have reduced pho- tosynthetic capacity (Cournac et al., 1991, 1992; Desjardins, 1995), malfunctioning stomata (Ziv et al., 1987; Frommel et al., 1991a; Majada et al., 1995), root system devoid of root hairs, poor cuticle development, and low wax deposits (Pierik, 1987; Lee and Wetzstein, 1988; Preece and Sutter, 1991). Consequently, when transplanted out of the culture vessels, they suffer from severe environmental stresses and substantial losses may occur (Preece and Sutter, 1991; Van Huylenbroeck and Debergh, 1996). Manipulation of acclimation conditions prior to or upon transplanting usually reduced losses (Donnelly et al., 1985; Desjardins et al., 1988; Preece and Sutter, 1991; Desjardins, 1995; Van Huylenbroeck and Debergh, 1996), however, at additional cost to the producer. To minimize these con- straints, in recent years, attempts have been made to develop pho- toautotrophic culture systems with high light intensity and reduced or eliminated sugar in the medium (Kozai et al., 1988; Kozai, 1991; Fujiwara and Kozai, 1995; Jeong et al., 1995). So far, this technology has not gained popularity.
During the last decade some researchers have reached for natural inhabitants of plants, epiphytes and endophytes, to enhance adap-
tation of tissue culture propagules to environmental stresses (reported by Herman, 1996a, 1996b). The plant-associated microorganisms, endophytes in particular, were always treated as problem-causing contaminants of in vitro cultures (reviewed by Cassells, 1991, and Leifert et al., 1994) and various procedures have been developed for their elimination (Cassells, 1991; Reed and Tanprasert, 1995) or management (Levin et al., 1997).
In nature, microorganisms inhabit the interior and exterior of plant organs (Jacobs et al., 1985; Frommel et al., 1993; Leifert et al., 1994; Mclnroy and Kloepper, 1995; Sturz, 1995; Sturz and Christie, 1995; Sturz et al., 1997). Some of these microorganisms, plant-beneficial bacteria and vesicular-arbuscular mycorrhizae in particular, can im- prove plant performance under stress environments and conse- quently, enhance yields (Brown, 1974; Schippers et al., 1987; Kloep- per et al., 1988, 1989; Lynch, 1990; Frommel et al., 1991b; Pfleger and Linderman, 1994; Chen et al., 1995; Datnoff et al., 1995; Glick, 1995; Balachandran et al., 1997; Bethlenfalvay et al., 1997; Creus et al., 1997; Jackson et al., 1997; Lazarovits and Nowak, 1997; Zehn- der et al., 1997). The mechanisms governing these responses have been extensively reviewed (e.g., Kloepper, 1988, 1989; Lynch, 1990; Dowling and O'Gara, 1994; Pfleger and Linderman, 1994; Glick, 1995, 1997; Lazarovits and Nowak, 1997). This review provides an outline of the present status of knowledge on utilization of microbial inoculants in plant tissue culture.
IN VITRO "BIOTIZATION"
Induction of stress resistance in plant propagules produced in vitro prior to transplanting, is a primary target of several researeh groups attempting utilization of microbial inoeulants in micropropagation (Hooker et al., 1994; Elmeskaoui et al., 1995; Nowak et al., 1995; Balla et al., 1997; Lazarovits and Nowak, 1997; Murphy et al., 1997; Wilhelm et al., 1997). Induced resistance responses to microbial inoeulants are well documented in plants grown in vivo (reviewed by Benhamou, 1996; Benhamou et al., 1996; Hunt and Ryals, 1996; Kloepper et al., 1997). Such responses in vitro have only been ree-
122

BIOTIZATION IN TISSUE CULTURE 123
ognized very recently (Herman, 1996b) and are referred to as "biot- ization" (reported by Herman, 1996a). I propose to define this term as follows: Biotization is a metabolic response of in vitro-grown plant material to a microbial inoculant(s), leading to the developmental and physiological changes enhancing biotic and abiotic stress resis- tance of the derived propagules. Establishment of artificial symbiotic associations (see Varga et al., 1994, and Preininger et al., 1997) could also belong to this category.
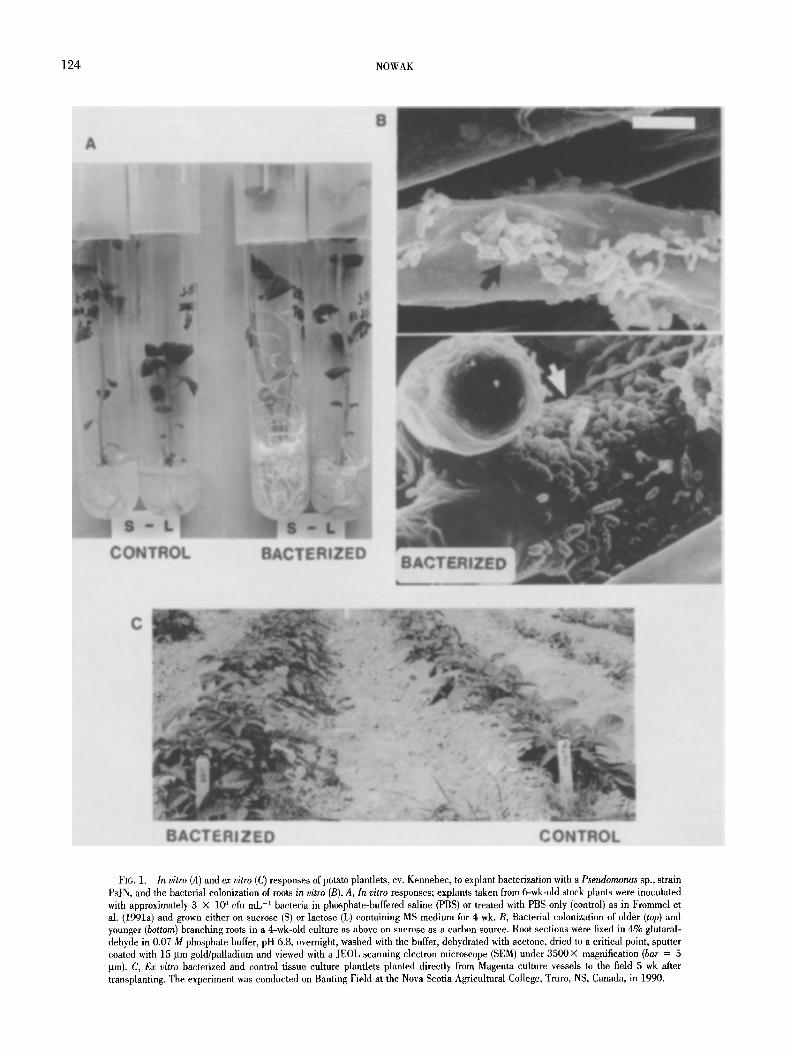
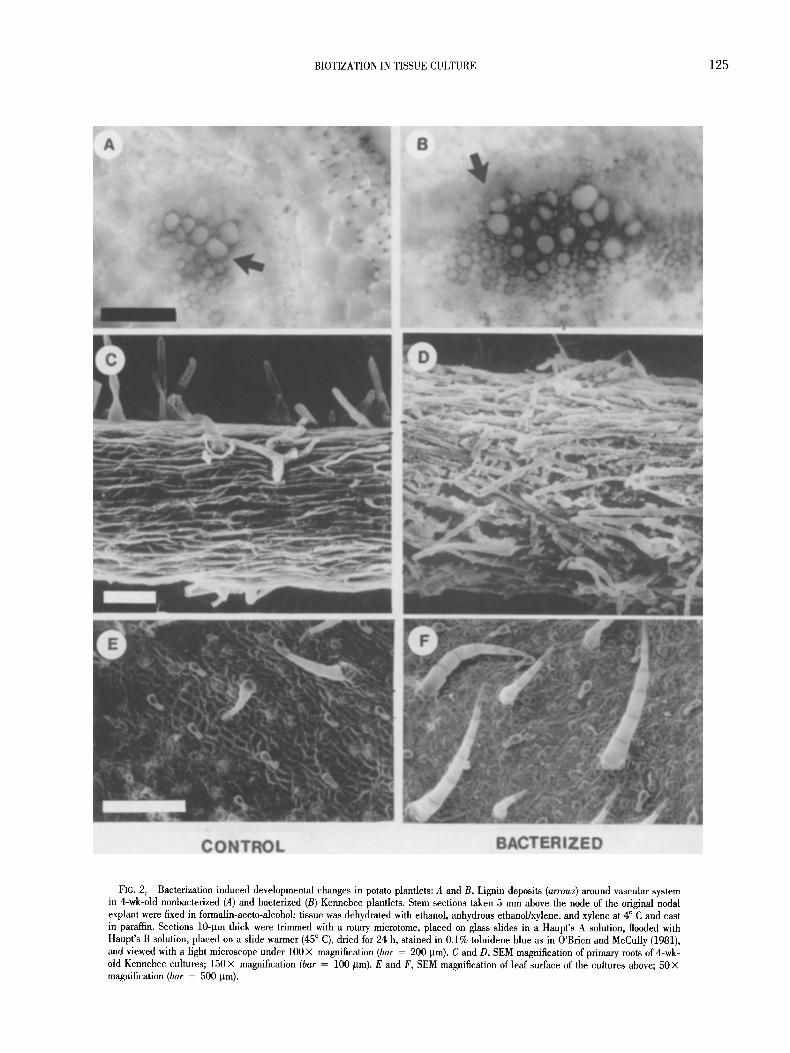
Bacterization. Eleven yr ago we initiated in vitro bacterization of potato plantlets to enhance their transplant stress tolerance and elim- inate an expensive greenhouse hardening step, which even now is commonly used by pre-elite seed potato producers (first reported by Herman in 1987). A pseudomonad bacterium, originally isolated as a contaminant from surface-sterilized onion roots in a study of Glo- mus vesiculiferum effect on in vitro growth of potato plantlets (Nowak et al., 1997), appears to be the most effective plant growth promoter we have come across so far (Conn et al., 1997; J. Nowak and C. Smith, unpublished). The bacterium (Pseudomonas spp. strain PsJN) does not grow on a standard plant tissue culture medium without the presence of the plant, and forms endophytic and epiphytic popula- tions when cocultured, either with potato (Frommel et al., 1991a) or tomato (Pillay and Nowak, 1997). Thus, in clonally propagated ma- terial there is no need for further reinoculation (Frommel et al., 1991a; Nowak et al., 1997). The bacterium stimulates plantlet growth (Frommel et al., 1991a; Liu et al., 1995; Nowak et al., 1995; Pillay and Nowak, 1997; Bensalim et al., 1998) and induces developmental changes (Frommel et al., 1991a), leading to better water management (Nowak et al., 1995; Lazarovits and Nowak, 1997) and enhanced resistance to low levels of pathogens (Nowak et al., 1995; Richards, 1997; Stewart, 1997; Sharma and Nowak, 1998) upon transplanting. Typical modifications are presented in Figs. 1 and 2. The inoculated plantlets have a massive, well-branched root system (Fig. 1 A), with bacteria cells grouped in small clusters along the older roots (top portion of Fig. 1 B) and forming tight cover around newly emerging root branches (bottom part of Fig. 1 B). After 4 wk in culture, com- pared to the nonbacterized controls, the bacterized plantlets are de- velopmentally more advanced, have sturdier stems, with more lignin deposits around the vascular system (Fig. 2 A vs B), more root hairs (Fig. 2 C vs D) and more and larger leaf hairs (Fig. 2 E vs F). Stomata function of the bacterized in vitro plantlets closely resembles that of greenhouse-hardened transplants. Five rain after leaf detachment from in vitro-grown plantlets, 90.1% of the stomata were closed in the bacterized versus 8.3% in the nonbacterized controls; the figures for the greenhouse-hardened transplants were 89.3 and 88.7% for the bacterized and controls, respectively (Frommel et al., 1991a). Improvement of water relations under osmotic stress was also re- corded in wheat seedlings cocuhured with AzospiriUum brasiliense strain SP245 (Creus et al., 1997). Live bacterial cells were required to induce these responses. Neither growth promotion nor develop- mental changes were induced, either by the autoclaved bacteria (Frommel et al., 1991a; Creus et al., 1997) or by the culture medium (Frommel et al., 1991a). Moreover, the growth stimulation pattern in tomato was closely related to the endophytic bacterial colonization (Pillay and Nowak, 1997). There was no relationship between colo- nization of the root surface and growth promotion in plantlets of to- mato (Pillay and Nowak, 1997), potato (J. Nowak, S. K. Asiedu, and C. Smith, unpublished), or other vegetable crops (Liu et al., 1995).
In 8 of 11 experiments conducted with potato plantlets trans- planted directly from culture vessels to the field between 1987 and
1992, bacterized plantlets had significantly better survival than non- bacterized controls (Nowak et al., 1995). Improved survival has also been recorded by Balla et al. (1997) with rooted black locust (Robinia pseudoacacia L.) plantlets inoculated with different Rhizobium iso- lates selected for growth vigor from a population of root nodule bac- teria obtained from five Robinia varieties. The bacterized potato transplants had also higher vigor during the first few weeks after planting to the field (e.g., Fig. 1 C). However, tuber yield responses in the field varied from yr to yr. In most experiments, there was an earlier tuber set and an enhancement of tuber number and yield in the bacterized treatments when compared to the nonbacterized (J. Nowak, S. K. Asiedu, and coworkers, unpublished). High precipi- tation or a very severe drought usually reduced this effect. Green- house experiments also demonstrated that plants derived from dual cultures of potato and the pseudomonad bacterium had a larger root system, set stolons and tuberized earlier, and gave better tuber yield than nonbacterized controls (Dunbar, 1997). Both in vitro and ex vitro benefits of bacterization depended on plant species, cultivar, and growth conditions (Nowak et al., 1995, 1997; Dunbar, 1997; Laza- rovits and Nowak, 1997; Pillay and Nowak, 1997; Bensalim et al., 1998).
Agrobacterium rhizogenes stimulation of adventitious rooting in culture is a well-known phenomenon (see McAfee et al., 1993; Bums and Schwarz, 1996). Rooting stimulation by bacteria isolates consid- ered to be culture contaminants (other than A. rhizogenes) have been reported only recently. In our group, dramatic root growth promotion was induced by a pseudomonad bacterium cocuhured with herba- ceous plants, potato (Frommel et al., 1991a; Lazarovits and Nowak, 1997), tomato (Pillay and Nowak, 1977), watermelon (Liu et al., 1995), and cucumber and pepper (Nowak et al., 1995). Bums and Schwarz (1996) had good success with induction of root development on slash pine seedling explants. The authors were able to root 85, 50, and 15% of explants of 6, 16, and 21 wk of age, respectively, when cocultured with their bacterium isolate. None of the explants rooted in the nonbacterized control groups. Similar to our observa- tions with herbaceous plants, Burns and Schwarz (1996) reported that the bacterium-induced roots resemble "seedling" roots rather than "hairy" roots, which are known to be induced by A. rhizogenes. Contrary to our observations that cocuhured plantlets had always well-branched, massive, but usually shorter root systems than non- inoculated controls, Lifshitz et al. (1987) reported dramatic elonga- tion of roots in Brassica campestris seedlings by a strain of Pseudomonas putida.
In vitro cocultivation of soybean cotyledon explants with two strains of Pseudomonas maltophilia, stimulated development of nod- ular callus with high regeneration potential (Yang et al., 1991). Im- provement of somatic embryogenesis by selected bacterial inoculants in genotypes recalcitrant to regeneration has further been reinforced by a study by Visser-Tenyenhuis et al. (1994). In a study by Shetty et al. (1995), shoot explant bacterization of oregano with a Pseudo- monas spp. prevented vitrification. The oregano plantlets cocuhured with the bacterium had lowered water content and contained more phenolics and chlorophyll than nonbacterized controls. Similar re- sponses were recorded in our studies with potato (Nowak et al., 1997) and vegetable crops (Nowak et al., 1995; Pillay and Nowak, 1997). Bacterized plantlets were greener, had elevated levels of cytokinins (Lazarovits and Nowak, 1997), phenylalanine ammonia lyase (PAL), and free phenolics (chlorogenic and caffeic acids in particular) (No- wak et al., 1997; Richards, 1997) and contained more lignin (From-
124 NOWAK
A B
CONTROL BACTERIZED
C
BACTERIZED CONTROL
FIG. 1. In vitro (A) and ex vitro (C) responses of potato plantlets, cv. Kennebec, to explant bacterization with a Pseudomonas sp., strain PsJN, and the bacterial colonization of roots in vitro (B). A, In vitro responses; explants taken from 6-wk-old stock plants were inoculated with approximately 3 • 10 a cfu mL -1 bacteria in phosphate-buffered saline (PBS) or treated with PBS only (control) as in Frommel et al. (1991a) and grown either on sucrose (S) or lactose (L) containing MS medium for 4 wk. B, Bacterial colonization of older (top) and younger (bottom) branching roots in a 4-wk-old culture as above on sucrose as a carbon source. Root sections were fixed in 4% glutaral- dehyde in 0.07 M phosphate buffer, pH 6.8, overnight, washed with the buffer, dehydrated with acetone, dried to a critical point, sputter coated with 15 Ilm gold/palladium and viewed with a ]EOL scanning electron microscope (SEM) under 3500 • magnification (bar = 5 p.m). C, Ex vitro bacterized and control tissue culture plantlets planted directly from Magenta culture vessels to the field 5 wk after transplanting. The experiment was conducted on Banting Field at the Nova Scotia Agricultural College, Trurn, NS, Canada, in 1990.
BIOTIZATION IN TISSUE CULTURE 125
CONTROL BACTERIZED
FIG. 2. Bacterization induced developmental changes in potato plantlets: A and B, Lignin deposits (arrows) around vascular system in 4-wk-old nonbacterized (A) and bacterized (B) Kennebec plantlets. Stem sections taken 5 mm above the node of the original nodal explant were fixed in formalin-aceto-alcohol; tissue was dehydrated with ethanol, anhydrous ethanol/xylene, and xylene at 4 ~ C and cast in paraffin. Sections 10-/.tin thick were trimmed with a rotary microtome, placed on glass slides in a Haupt's A solution, flooded with Haupt's B solution, placed on a slide warmer (45 ~ C), dried for 24 h, stained in 0.1% toluidene blue as in O'Brien and McCully (1981), and viewed with a light microscope under 100 X magnification (bar = 200 p_m). C and D, SEM magnification of primary roots of 4-wk- old Kennebec cultures; 150 • magnification (bar = 100 ~tm). E and F, SEM magnification of leaf surface of the cultures above; 50 X magnification (bar = 500 p_m).

126 NOWAK
mel et al., 1991a; Richards, 1997). Cocuhuring chestnut with a strain L25 of Bacillus subtilis, Wilhelm et al. (1997) also found production of pathogenesis-related proteins, extracellular chitinase, and 13-1,3 gluconase isoenzymes which did not appear in wounded control plants. The above are typical symptoms of induced resistance re- sponses (Dixon et al., 1994; Hunt and Ryals, 1996). In our study with potatoes, elevated PAL activity in bacterized plants, compared to nonbacterized, was found during the first 4 wk of growth; there was no difference in its level in 6-wk-old plantlets. However, when explants were taken from 6-wk-old bacterized and control stock plants, PAL activity in the newly derived shoots was higher in the prebacterized treatment (Nowak et al., 1997; Richards, 1997). This indicates a more rapid plant response to cutting in the prebacterized tissues. The speed and degree of plant response to stress signals is recognized as a critical factor in plant resistance to pathogen attack (Behamou, 1996). Unlike the Bacillus strain in the study by Wilhelm et al. (1997), our Pseudomonas bacterium did not show any antago- nistic effects on fungal pathogens in in vitro plate bioassay. Moreover, bacterized potato were resistant to low levels of Verticillium albo- atrum (Nowak et al., 1995; Stewart, 1997) and tomato plants to V. dahliae (Sharma and Nowak, 1998). Plantlet bacterization, upon transplanting, did not induce such responses (Stewart, 1997; Sharma and Nowak, 1998). As the in vitro growth responses depended on the degree of endophytic colonization and a certain threshold of the bac- teria concentration was required to trigger such response (Pillay and Nowak, 1997), Sharma and Nowak (1998) postulated similar induc- tion patterns for the resistance to pathogens. There is some evidence that a pathway of responses to biotic and abiotic stresses in plants can be universal (reviewed by Sheen, 1996, and Harmon, 1997).
Mycorrhization. Benefits of inoculation of tissue culture plantlets with endomycorrhizal fungi have been advocated by several authors (Pierik, 1988; Ch~vez and Ferrera-Cerrato, 1990; Ponton et al., 1990; Hooker et al., 1994; Elmeskaoui et al., 1995). It is well established that root colonization by vesicular-arbuscular mycorrhizae (VAM) im- proves plant nutritional status, disease resistance, and water man- agement (see references in Powell and Bagyaraj, 1984; Hooker et al., 1994; Elmeskaoui et al., 1995). In an attempt to harness this poten- tial for the improvement of the performance of tissue culture plant- lets, several researchers inoculated rooted shoots upon transplanting (Ch~vez and Ferrera-Cerrata, 1990; Ponton et al., 1990; Wang et al., 1993 and references therein). The benefits of mycorrhization de- pended on the growing medium (Ponton et al., 1990) and the degree of root colonization (Wang et al., 1993). As the VAM stimulation of plantlet growth was frequently expressed only after acclimatization, Wang et al. (1993) suggested development of a culture system into which the mycorrhizal fungi are introduced in vitro, during the root- ing stage. Challenges of the axenic cultivation of VAM fungi were overcome in studies by B~card and Fortin (1988) and B6card and Pich~ (1989). The authors were able to successfully grow the fungi in vitro on Ri T-DNA transformed carrot roots, under elevated CO2. Capitalizing on this technique, Y. Desjardins's group developed a "tripartite" culture system for in vitro inoculation of strawberries (Desjardins, 1995; Elmeskaoui et al., 1995) and vegetable crops (Y. Desjardins, personal communication).
The most effective treatment consisted of a 30-d-old VA (Glomus intraradices) mycorrhizal-transformed carrot root culture and straw- berry shoots in cellulose plugs. After root induction, the plugs were placed on the surface of the mycorrhized root culture and grown in a growth chamber, under 5000 ppm CO2 for 20 d (Elmeskaoui et al.,
1995). All plantlets were successfully colonized and exhibited larger root systems, better shoot growth, and higher (more negative) osmotic potential than nonmycorrhizal controls. The authors suggest that the enhancement of osmotic potential can be an important preadaptation step prior to full acclimation of plantlets for transplanting. Indeed, the work by Creus et al. (1997) demonstrated that a higher potential recorded in wheat seedlings cocultured with Azospirillum allowed them to withstand osmotic stress much better than noninoculated controls with lower osmotic potential.
Another mycorrhization system was developed by A. Cassell's group (Murphy et al., 1997). The authors grew strawberry microplants in a polyurethane foam substrate under photoautotrophic conditions, with reduced sucrose concentration in the medium. The foam tear- away strips seem to be very useful as they can be made to fit any culture vessel and the VAM spores can be placed directly in the planting holes. The mycorrhized plantlets had better establishment and produced more runners than nonmycorrhized controls. Murphy et al. (1997) also suggest that a combination of VAM and beneficial bacteria can perhaps be even more efficient than VAM alone.
Recent reports by Walley and Germida (1997) indicate that util- ization of such combinations should be approached with caution. In their study with five plant growth-promoting pseudomonads and a VAM isolate NT4 of Glomus clarum in a gnotobiotic system, dual inoculation with VAM and bacteria usually reduced the magnitude of the plant growth response as compared to the single inoculant. Moreover, two bacteria strains inhibited G. clarum NT4 spore ger- mination.
Other potential inoculations. Cyanobacterial and fungal culture ex- tracts have been proven to induce developmental changes and path- ogen resistance in tissue-cultured material when added to the me- dium. Both dialysates and nondialysates of hot water extracts from filamentous cyanobacteria, Nostoc sp. and Anabaena sp., and uni- cellular strains of Synechococcus sp. and Xenococcus sp., promoted somatic embryogenesis and plantlet regeneration in Daucus carota L. (Wake et al., 1991). Higher molecular compounds (nondialysates) from the unicellular strains were more effective than the smaller ones (dialysates). Further fractionation of the extract from Synechococcus sp. strain NKBG 042902 by centrifugation allowed the authors to concentrate this high molecular elicitor (Wake et al., 1992). When added to an embryogenic culture of carrot it induced chlorophyll accumulation (5 X more than in nontreated cultures) and adjusted the chlorophyll a/b ratio close to the zygotic embryo level. Plantlet conversion was 60% with this compound and only 28% without its addition.
Yu et al. (1990) demonstrated the stabilizing effect of Verticillium albo-atrum culture filtrate on somatic embryogenesis in alfalfa cell suspension culture. While control cultures lost their somatic em- bryogenesis after six subcultures, the culture filtrate-treated ones (20% vol/vol) still maintained a relatively high number of embryos through all subcultures. Shoot regeneration from potato tuber discs was also stimulated by a culture filtrate of Alternaria solani (Lynch et al., 1991). The authors advocate it for use in in vitro selection and genetic transformation of this species.
Successful application of microbial culture filtrates in control of plant tissue culture contaminants was demonstrated by Hussain et al. (1994). The acetone-precipitated fraction of Bacillus subtilis Ehrnberg (strain 2) and Trichoderma viride Pers. (strain A) exhibited high antifungal activity, whereas extracts from Pseudomonasfluores- ceace Migula (strain X) had high antibacterial activity. The bacterial

BIOTIZATION IN TISSUE CULTURE 127
extracts did not reduce the performance of uncontaminated cultures of Nicotiana tabacum L. over four subcultures and reduced the growth of accidental contaminants. Heat-stable mycelial extracts of another Trichoderma species, a nonpathogenic T. longibrachiatum, were capable of triggering induced resistance responses in tobacco seedlings to Phytophthora parasitica var. nicotianae (race 0) (Chang et al., 1997). It induced expression of pathogenesis-related proteins, PR-16 and osmotin (PR-5) on a higher level than an extract prepared the same way from the pathogenic fungus. Also the elicitation of the defense responses could not be fully achieved by a combination of salicylic acid and methyl jasmonate (Chang et al., 1997).
The above studies further emphasize a potential for use of non- pathogenic organisms in plant tissue culture systems. If the culture conditions are refined, artificial associations with either individual microorganisms or their groups can be created. Such microorganisms not only would act as inducers of the stress resistance responses but also could occupy microsites on host plants, making them unavail- able to pathogens. An example of a successful artificial association between plants and a nitrogen-fixing bacterium has been described by Varga et al. (1994). The authors created a symbiotic culture sys- tem, callus-bacterium, between Daucns carota L. and Azotobacter zettuovii (CRS-H6) which could grow for 4 yr on a nitrogen-free me- dium with lactose as a carbon source. The bacterium, located in intracellular spaces, could also be transmitted to and fix nitrogen in newly regenerated plantlets (Varga et al., 1994; Preininger et al., 1997). Recently the authors were able to establish similar associa- tions also in tomato, potato, wheat, sugar cane, and poplar, using 11 strains of 8 Azotobacter species (Sz. S. Varga, personal communica- tion) and in strawberries with Azomonas insignis (Preininger et al., 1997). In the tomato system, the bacterium could also be transmitted through seeds; approximately 20% of seedlings derived from the seeds of Azotobacter beijerinckii containing tomato demonstrated N 2
fixing activity 2 mo. after germination (Sz. S. Varga, personal com- munication).
A potential also exists for the utilization of traditional "night- mares" of micropropagators, the mycoplasma-like organisms, in re- cent literature termed as 'mollicutes' (Bove and Gamier, 1997), and viruses (Herman, 1996b). Both are known to form very stable asso- ciations with plants and are difficult to eradicate in tissue culture. Reports on mycoplasma promotion of morphogenesis in sandalwood (Santalum album) callus, and tomato mosaic virus stimulation of callus growth and morphogenesis in five Lycopersicon species (see Herman, 1996b) as well as viral modification of plant responses to environmental factors (Balachandran et al., 1997) encourage further investigation of their potential use in tissue culture.
PLANT--MICROBIAL COCULTURES FOR PURPOSES
OTHER THAN PROPAGATION
In vitro bioassays. Stimulation of Brassica campestris root devel- opment in a gnotobiotic growth pouch bioassay by pseudomonad bac- teria (Lifshitz et al., 1987) indicated the usefulness of such a system in screening of rhizosphere bacteria for plant growth promotion with- out interference of contaminants. DeFreitas and Germida (1990) fur- ther demonstrated modification of root morphology and stimulation of root hair development in root tissue culture of wheat by various bacterial species and strains, and Chanway and Nelson (1991) showed soybean callus growth promotion on a vitamin-free medium by one of the two tested strains of Pseudomonas putida. Recently, a
canola and lettuce seeding bioassay was also used to demonstrate root development stimulation of nonlegume plants by Rhizobium leg- uminosarum (Noel et al., 1996). One of the disadvantages of seed- based bioassay is the difficulty with obtaining "clean" cultures. Conn et al. (1997) described a clonal bioassay based on bacterization of potato nodal explants. Such bioassay, modified after Frommel et al. (1991a), provides genetically identical explants which are taken from contaminant-free stock plants of the same developmental stage, is reproducible and allows study of bacteria-plant interactions for many wk. The authors (Conn et al., 1997) were able to successfully assess growth-promoting ability of various bacterial isolates and Tn5 mu- tants of a nonfluorescent Pseudomonas species (PsJN). The bioassay can also be used to determine plant genotypic responses to particular strains of bacteria or their combinations.
Plant culture bioassays are also recommended for identification of bacteria for biological control organisms (e.g., a surface-sterilized petunia disc bioassay for Botrytis-controlling bacteria reported by Gould et al., 1996) and for monitoring disease development on roots (e.g., Verticillium development on roots of tissue culture-grown po- tatoes reported by Platt and Bollen, 1993). Hampson et al. (1997) were recently able to establish a successful dual culture system of potato and the infective agent of potato wart disease (Synchytrium endobioticum) which can be used for virulence determination and identification of phenotypes of this pathogen.
Test-tube ecophysiology of plant-microbial interactions. Erratic yield responses of field-grown crops subjected to microbial inocu- lants frustrate the inoculants' users and producers (Rovira, 1991; Lazarovits and Nowak, 1997). One of the limitations preventing us from using their full potential is insufficient knowledge of the dy- namics of microbial interactions in plant rhizosphere and phyllos- phere under varied environmental conditions. Tissue culture allows for the creation of simple model systems for testing single factors and their combinations. A recent study by Pillay and Nowak (1997) demonstrated, for example, that lower inoculum densities were re- quired to maximize root growth promotion (approximately los cfu mL -~) than shoot growth promotion (approximately 3 • l0 s cfu mL-1) in tomatoes cocuhured with a growth-promoting pseudomonas bacterium. Temperature had also a dramatic effect on the degree of plant growth promotion (Pillay and Nowak, 1997; Bensalim et al., 1998) and tissue colonization (Pillay and Nowak, 1997). Compared to room temperature, a temperature elevation stimulated in vitro growth responses to bacterization in most clones of potato (Bensalim et al., 1998) but lowered them in tomatoes (Pillay and Nowak, 1997). In potato, under heat stress, the bacterium facilitated changes in biomass allocation to roots (Bensalim et al., 1998). In both these studies, there were dramatic differences between the responses of different cultivars. This could be partly linked to the genotypic dif- ferences in ethylene accumulation in culture vessels (J. Nowak and D. Stevens, unpublished).
Carbon source in the medium can also have an effect on plant growth responses to bacterization. Pseudomonas sp. strain PsJN, as well as other growth-promoting rhizobacteria tested in our laboratory, including some strains of Rhizobium leguminosarum (J. Nowak, T. Sturz and D. Stevens, unpublished), always stimulated more root growth on sucrose and more shoot growth on lactose-containing me- dium (Fig. 1 A).
A beneficial effect of bacterial combinations on in vitro growth of red clover was demonstrated by Sturz et al. (1997). The authors iden- tified root nodule bacteria (isolated from in vivo-grown plants) be- longing to the genera Agrobacterium, Bacillus, Pseudomonas and

128 NOWAK
Curtobacterium as capable of in vitro seedling growth promotion or nodulation when inoculated either singly (growth promotion) or in combination with Rhizobium leguminosarum BV trifolii (growth pro- motion and/or nodulation). Interestingly, the Rhizobium inoculation alone always caused seedling growth depression compared to non- inoculated controls.
The above-outlined studies are examples of the potential uses of the tissue culture systems for study of allelopathic side effects of different microbial combinations on plant performance, selection of the best strain combinations, determination of inoculum concentra- tions, and plant growth conditions for bacteria utilization (endophytes in particular, as their populations are less susceptible to environ- mental changes) to their full potential. Similar studies are also being conducted with VAM (Desjardins, 1995) and VAM-rhizobacteria combinations in other laboratories (Y. Desjardins, personal commu- nication; Walley and Germida, 1997).
CONSTRAINTS AND FUTURE DIRECTIONS
The fact that we can culture probably only a few percent or even less of naturally occurring microorganisms (Hugenhohz and Pace, 1996) is one of the challenges of the utilization of microbial inocu- lants in plant production. Development of new culture methods al- lowing establishment of stable associations between plants and ben- eficial organisms in Vitro and ex vitro and understanding of mechanisms of signal recognition and transduction in plant-micro- bial associations under different environments are probably the most critical elements of this challenge. There are also psychological and regulatory barriers set up, for example, in the nuclear seed potato systems which do not allow cocuhuring of tissue culture propagules with microorganisms. In view of the present knowledge of the role of these microorganisms and a substantial body of literature demon- strating either their benefits (reviewed above) or passivity (e.g., Mo- nette, 1986), the regulatory hurdles should be lifted. Such a move would stimulate inflow of industrial research funds. This in turn could stimulate research aiming at the genetic modification of the plant-associative microorganisms, as well as modification of plant genome to target specific groups of such microorganisms. Plant breeding for sustainable agricultural production systems must seri- ously consider such an option. Certain groups of microorganisms could not only benefit a targeted crop but also other crops in a rotation system. In a study with potato, clover, and barley, we have identified some endophytic bacteria, including Rhizobia, which can benefit all three crops (J. Nowak, A. V. Sturz, and B. R. Christie, in preparation). A field study by Bethenfalvay et al. (1997) demonstrate that mycor- rhizal fungi-rhizobacteria combinations not only can affect plants directly but also indirectly, by the enhancement of the proportion of water-stable aggregates in soils.
ACKNOWLEDGMENTS
Microscopy work presented in Figs. 1 and 2 was done by Mr. Jeff Richards, a graduate student (M.Sc.) working under the author's cosupervision. The author would like to thank Ms. Marie Marshall for her contribution in pre- paring the manuscript.
REFERENCES
Baccou, J. C. Effect of photosynthesis on the secondary metabolism of cell cultures. In: Carrr, F.; Chagvardieff, P., ed. Ecophysiology and pho- tosynthetic in vitro cultures. Aix-en-Provence (France): CEA; 1995:71-85.
Balachandran, S.; Hurry, V. M.; Kelley, S. E., et al. Concepts of plant biotic stress. Some insights into the stress physiology of vires-infected plants, from perspective of photosynthesis. Physiol. Plant. 100:203- 213; 1997.
Balla, I.; Vrrtesy, J.; Ktives-Pechy, K., et al. Acclimation results of micro- propagated black locust (Robinia pseudoacacia L.) improved by use of microorganisms. In: Cassells, A. L., ed. Pathogen and microbial contamination management in micropropagation. Dordrecht (NL): Kluwer Acad. Publ.; 1997:351-354.
Brcard, G.; Fortin, J. A. Early events of vesicular-arbuscular mycorrhiza formation on Ri T-DNA transformed roots. New Phytol. 108:211-218; 1988.
B6card, G.; Pichr, Y. Fungal growth stimulation by CO 2 and root exudates in vesicular-arbuscular mycorrhizal symbiosis. Appl. Envir. Microbiol. 55:2320-2325; 1989.
Benhamou, N. Elicitor-induced plant defence pathways. TIPS 1:233-240; 1996.
Benhamou, N.; Kloepper, J. W.; Quadt-Hallman, A., et al. Induction of de- fence-related ultra-structural modifications in pea root tissues inoc- ulated with endophytic bacteria. Plant Physiol. 112:919-929; 1996.
Bensalim, S.; Nowak, J.; Asiedu, S. K. Temperature and pseudomonad bac- terium effects on in vitro and ex vitro performance of 18 clones of potato. Am. Potato J. (in press); 1998.
Bethlenfalvay, G. J.; Andrade, G.; Azc6n-Aquilar, C. Plant and soil responses to mycorrhizal fungi and rhizobacteria in nodulated or nitrate-fertil- ized peas (Pisum sativum L.). Biol. Fertil. Soils 24:164-168; 1997.
Bove, J. M.; Gamier, M. YC:alled and wall-less eubacteria from plants: sieve- tube-restricted plant pathogens. In: Cassells, A., ed. Pathogen and microbial contamination management in micropropagation. Dordrecht (NL): Kluwer Academic Publ.; 1997:45-60.
Brown, M. E. Seed and root bacterization. Annu. Rev. Phytopathol. 12:181- 197; 1974.
Bums, J. A.; Schwarz, O. J. Bacterial stimulation of adventitious rooting on in vitro cultured slash pine (Pinus eliotti Engelm.) seedling explants. Plant Cell Rep. 15:405-408; 1996.
Cassells, A. C. Problems in tissue culture: culture contamination. In: De- bergh, P. C.; Zimmerman, R. M., ed. Micropropagation, technology and application. Dordrecht: Kluwer Acad. Publ.; 1991:31-44.
Chang, P.-F. L.; Xu, Y.; Narasimhan, M. L., et al. Induction of pathogen resistance and pathogenesis-related genes in tobacco by a heat-stable Trichoderma mycelial extract and plant signal messengers. Physiol. Plant. 100:341-352; 1997.
Chanway, C. P.; Nelson, L. M. Tissue culture bioassay for plant growth pro- moting rhizobacteria. Soil Biol. Biochem. 23:331-333; 1991.
CMvez, M. C. G.; Ferrera-Cerrato, R. Effect of vesicular-arbuscular mycor- rhizae on tissue culture-derived plantlets of strawberry. HortScience 25:903-905; 1990.
Chen, C.; Bauske, E. M.; Musson, G., et al. Biological control of fusarium wilt on cotton by use of endophytic bacteria. Biol. Control 5:83-91; 1995.
Conn, K. L.; Nowak, J.; Lazarovits, G. A gnotobiotic bioassay for studying interactions between potato and plant growth-promoting rhizobac- teria. Can. J. Microbiol. 43:801--808; 1997.
Cournac, L.; Cirier, I.; Chagvardieff, P. Improvement of photoautotrophic So- lanum tuberosum plantlet culture by light and CO2: differential de- velopment of photosynthetic characteristics and varietal constraints. Acta Hortic. 319:53-58; 1992.
Cournac, L.; Dimon, B.; Carrier, P., et al. Growth and photosynthetic char- acteristics of Solanum tuberosum plantlets cultivated in vitro in dif- ferent conditions of aeration, sucrose supply and CO2 enrichment. Plant Physiol. 97:112-117; 1991.
Creus, C. M.; Sueldo, R. J.; Barassi, C. A. Water relations in Azospirillum- inoculated wheat seedlings under osmotic stress. Can. J. Bot. (in press): 1998.
Datnoff, L. E.; Nemec, S.; Pemezny, K. Biological control of fusarium crown and root rot of tomato in Florida using Trichoderma harzianum and Glomus intraradices. Biol. Control 5:427-431; 1995.
DeFreitas, J. R.; Germida, J. J. A root tissue culture system to study winter wheat-rhizobacteria interactions. Appl. Microbiol. Biotechnol. 33:589-595; 1990.
Desjardins, Y. Overview of factors influencing photosynthesis of micropro- pagated plantlets and their effect on acclimatization. In: Carrr, F.;

BIOTIZATION IN TISSUE CULTURE 129
Chagvardieff, E, ed. Ecophysiology and photosynthetic in vitro cul- tures. Aix-en-Provenee (France): CEA; 1995:145-160.
Desjardins, Y.; Laforge, E; Lussier, C., et al. Effect of CO2 enrichment and high photosynthetic photon flux on the development of autotrophy and growth of tissue euhured strawberry, raspberry and asparagus plants. Acta Hortie. 230:45-53; 1988.
Dixon, R. A.; Harrison, M. J.; Lamb, C. E Early events in the activation of plant defenee responses. Annu. Rev. Phytopathol. 32:47%501; 1994.
Donnelly, D. J.; Vidaver, W. E.; Lee, K. Y. The anatomy of tissue cultured red raspberry prior to and after transfer to soil. Plant Cell Tissue Organ Cult. 4:43-50; 1985.
Dowling, D. N.; O'Gara, E Metabolites of Pseudomonas involved in the bio- control of plant diseases. TIBTECH 12:133-141; 1994.
Dunbar, C. Utilization of seaweed extract and plant growth promoting rhizo- bacterium in greenhouse production of potato minitubers. M.Sc. the- sis, Dalhousie University, Halifax, NS, Canada; 1997.
Elmeskaoui, A.; Damont, J.-J. E; Pichr, Y., et al. A tripartite euhure system for endomyeorrhizal inoculation of micropropagated strawberry plant- lets in vitro. Mycorrhiza 5:313-319; 1995.
Frommel, M. I.; Nowak, J.; Lazarovits, G. Growth enhancement and devel- opmental modifications of in vitro grown potato (Solanum tuberosum ssp. tuberosum) as affected by a nonfluorescent Pseudomonas sp. Plant Physiol. 96:928-936; 1991a.
Frommel, M. I.; Pazos, G. S.; Nowak, J. Plant-growth stimulation and bio- control of Fusarium wilt by co-inoculation of tomato seeds with Ser- ratia plymuthica and Pseudomonas sp. Fitopathologia 26:66-73; 1991b.
Frommel, M. I.; Nowak, J.; Lazarovits, G. Treatment of potato tubers with a growth promoting Pseudomonas sp.; bacterium distribution in the rhi- zosphere and plant growth responses. Plant Soil 150:51-60; 1993.
Fujiwara, K.; Kozai, T. Control of environmental factors for plantlet produc- tion-with some mathematical simulation. In: Carre, E; Chagvardieff, E, ed. Ecophysiology and photosynthetic in vitro euhures. Aix-en- Provence (France): CEA; 1995:109-120.
Gliek, B. R. The enhancement of plant growth by free-living bacteria. Can. J. Mierobiol. 41:109-117; 1995.
Gliek, B. R.; Bashan, Y. Genetic manipulation of plant growth-promoting bacteria to enhance biocontrol of phytopathogens. Bioteehnol. Adv. 15:353-378; 1997.
Gould, A. B.; Kobayashi, D. Y.; Bergen, M. S. Identification of bacteria for biocontrol of Botrytis einerea on petunia using petal disc assay. Plant Disease 80:1029-1033; 1996.
Hampson, M. C.; Coombes, J. W.; Debnath, S. C. Dual euhure of Solanum tuberosum and Synchytrium endobioticum (patfiotype 2). Mycologia 89:772-776; 1997.
Harmon, A. C. The calcium connection. TIPS 2:121-122; 1997. Herman, E. B. Contaminants promote potato micropropagation. Agricell Re-
port 9:38; 1987. Herman, E. B. Beneficial effects of bacteria and fungi on plant tissue cultures.
Agricell Rep. 27:26-27; 1996a. Herman, E. B. Microbial contamination of plant tissue cultures. Recent Ad-
vances in Plant Tissue Culture IV. Shrub Oak (NY): Agriteeh Cons., Inc.; 1996b.
Hooker, J. E.; Gianinazzi, S.; Vestberg, M., et al. The application of arbuseular mycorrhizal fungi to mieropropagation systems: an opportunity to re- duce chemical inputs. Agrie. Sci. Finland 3:227-232; 1994.
Hugenholtz, E; Pace, N. R. Identifying microbial diversity in the natural environment; a molecular phylogenetie approach. TIBTECH 14:190- 197; 19%.
Hunt, M. D.; Ryals, J. A. Systemic acquired resistance signal transduetion. Critical Rev. Plant Sci. 15:583-606; 1996.
Hussain, S.; Lane, S. D.; Price, D. N. A preliminary evaluation of the use of microbial euhure filtrates for the control of contaminants in plant tissue culture systems. Plant Cell Tissue Organ Cult. 36:45--51; 1994.
Jackson, A. J.; Waiters, D. R.; Marshall, G. Antagonistic interactions between the foliar pathogen Botrytis fabae and isolates of Penieillium brevi- eompaetum and Cladosporium cladosporioides on faba beans. Biol. Control 8:97-106; 1997.
Jaeobs, M. J.; Bugbee, W. M.; Gabrielson, D. A. Enumeration, location, and characterization of endophytie bacteria within sugar beet roots. Can. J. Bot. 63:1262-1265; 1985.
Jeong, B. R.; Fujiwara, K.; Kozai, T. Environmental control and photoauto- trophic micropropagation. Hort. Rev. 17:125-172; 1995.
Kloepper, J. W.; Lifshitz, R.; Schroth, M. N. Pseudomonas inoculants to ben- efit enhancing production. ISI Atlas of Sciences: Animal Plant Sci. 1:60-64; 1988.
Kloepper, J. W.; Lifshitz, R.; Zablotowicz, R. M. Free-living bacterial inocula for enhancing crop productivity. TIBTECH 7:39---44; 1989.
Kloepper, J. W.; Tuzun, S.; Zehnder, G. W., et al. Multiple disease protection by rhizobacteria that induce systemic resistance--historical perspec- tive. Phytopathology 87:136-137; 1997.
Kozai, T. Micropropagation under photoautotrophic conditions. In: Debergh, P. C.; Zimmerman, R. M., ed. Micropropagation, technology and ap- plication. Dordrecht: Kluwer Acad. Publ.; 1991:447469.
Kozai, T.; Koyama, Y.; Watanabe, I. Multiplication of potato plantlets in vitro with sugar-free medium under high photosynthetic photon flux. Acta Hortic. 230:121-127; 1988.
Lazarovits, G.; Nowak, J. Rhizobacteria for improvement of plant growth and establishment. HortScience 32:188--192; 1997.
Lee, N.; Wetzstein, H. Y. Quantum flux density effects on the anatomy and surface morphology of in vitro and in vivo developed sweetgum leaves. J. Am. Soc. Hort. Sci. 113:167-171; 1988.
Leifert, C.; Morris, C. E.; Waites, W. M. Ecology of microbial saprophytes and pathogens in tissue culture and field-grown plants: reasons for con- tamination problems in vitro. Crit. Rev. Plant Sci. 13:139-183; 1994.
Leifert, C.; Murphy, K. P.; Lumsden, P. J. Mineral and carbohydrate nutrition of plant cell and tissue cultures. Crit. Rev. Plant Sci. 14:83-109; 1995.
Levin, R.; Stav, R.; Alper, Y., et al. A technique for repeated non-axenic sub- culture of plant tissues in a bioreactor on liquid medium containing sucrose. Plant Tissue Cult. Biotech. 3:4145; 1997.
Lifshitz, R.; Kloepper, J. W.; Kozlowski, M., et al. Growth promotion of canola (rapeseed) seedlings by a strain of Pseudomonas putida under gno- tobiotic conditions. Can. J. Microbiol. 33:390-395; 1987.
Liu, Z.; Pillay, V.; Nowak, J. In vitro culture of watermelon and cantaloupe with and without beneficial bacterium. Acta Hortic. 402:58-60; 1995.
Lynch, D. R.; Coleman, M. C.; Lyon, G. D. Effect ofAlternaria solani culture filtrate on adventitious shoot regeneration in potato. Plant Cell Rep. 9:607-610; 1991.
Lynch, J. M., ed. The rhizosphere. Chichester (UK): Wiley; 1990. Majada, J. P.; Fal, M. S.; Sanchez-Tomrs, R. Influence of the in vitro envi-
ronment on the stomatal physiology and morphology of micropropa- gated Dianthus caryophyllu~ cv. Nelken. In: Carre, F.; Chagvardieff, P., ed. Ecophysiology and photosynthetic in vitro cultures. Aix-en- Provence (France): CEA; 1995:141-144.
Matthijs, D. G.; Pascat, B.; Demeester, J., et al. Factors controlling the evo- lution of the gaseous atmosphere during in vitro culture. In: Carre, E; Chagvardieff, P., ed. Ecophysiology and photosynthetic in vitro cul- tures. Aix-en-Provence (France): CEA; 1995:129-140.
McAfee, B. J.; White, E. E.; Pelcher, L. E., et al. Root induction in pine (Pinus) and larch (Lar/x) spp. using Agrobacterium rhizogenes. Plant Cell Tissue Organ Cult. 34:53-62; 1993.
McClelland, M. T.; Smith, M. A. L. Vessel type, closure, and explant orien- tation influence in vitro performance of five woody species. Hort- Science 25:797-800; 1990.
McInroy, J. A.; Kloepper, J. W. Survey of indigenous bacterial endophytes from cotton and sweet corn. Plant Soil 173:337-342; 1995.
Monette, P. L. Micropropagation of kiwi fruit using non-axenic shoot tips. Plant Cell Tissue Organ Cult. 6:73-82; 1986.
Murphy, J.; Mark, L.; Periappuram, C., et al. Microbial characterisation and preparation of inoculum for in vitro mycorrhization of strawberry in autotrophic culture. In: Cassells, A. L., ed. Pathogen and microbial contamination management in micropropagation. Dordrecht (NL): Kluwer Acad. Publ.; 1997:345-350.
Noel, T. C.; Sheng, C.; Yost, C. K., et al. Rhizobium leguminosarum as a plant growth-promoting rhizobacterium: direct growth promotion of canola and lettuce. Can. J. Microbiol. 42:279-283; 1996.
Nowak, J.; Asiedu, S. K.; Bensalim, S., et al. From laboratory to applications: challenges and progress with in vitro dual cultures of potato and ben- eficial bacteria. In: Cassells; A., ed. Pathogen and microbial contam- ination management in micropropagation. Dordrecht (NL): Kluwer Acad. Publ.; 1997:321-329.

130 NOWAK
Nowak, J.; Asiedu, S. K.; Lazarovits, G., et al. Enhancement of in vitro growth and transplant stress tolerance of potato and vegetable plants co- cultured with a plant growth promoting rhizobacterium. In: Carre, E; Chagvardieff, P., ed. Ecophysiology and photosynthetic in vitro cul- tures. Aix-en-Provence (France): CEA; 1995:173-180.
O'Brien, T. P.; McCully, M. E. The study of plant structure. Principles and selected methods. Melbourne (Australia): Termarcarphi Pty Ltd.; 198t.
Pfleger, E L.; Linderman, R. G., ed.. Mycorrhizae and plant health. St. Paul (MN): Am. Phytopathol. Soc.; 1994.
Phan, C. T.; Hegedus, P. Possible metabolic basis for the developmental anomaly observed in in vitro culture called 'vitreous plants'. Plant Cell Tissue Organ Cult. 6:83-94; 1986.
Pierik, R. L. M. In vitro culture of higher plants. Dordrecht (NL): Martinus Nijhoff Publ.; 1987.
Pierik, R. L. M. In vitro culture of higher plants as a tool in propagation of horticultural crops. Acta Hort. 226:25-40; 1988.
Pillay, V. K.; Nowak, J. Inoculum density, temperature and genotype effects on epiphytic and endophytic colonization and in vitro growth pro- motion of tomato (Lycopersicon esculentum L.) by a pseudomonad bac- terium. Can. J. Microbiol. 43:354-361; 1997.
Platt, H. W.; Bollen, J. J. Use of tissue culture potato plantlets for investi- gations of diseases of subterranean plant parts. Plant Dis. 77:1112- 1113; 1993.
Ponton, E; Pich~, Y.; Parent, S., et al. The use of vesicular-arbuscular my- corrhizae in Boston fern production: I. Effects of peat-based mixes. HortScience 25:183-189; 1990.
Powell, C. L.; Bagyaraj, D. J., ed.. VA mycorrhiza. Boca Raton (FL): CRC Press, Inc.; 1984.
Preece, J. E.; Sutter, E. G. Acclimation of micropropagated plants to the greenhouse and field. In: Debergh, P. C.; Zimmerman, R. H., ed. Micropropagation, technology and application. Dordrecht (NL): Klu- wer Acad. Publ.; 1991:71-93.
Preininger, E.; Zatyko, J.; Szucs, P., et al. In vitro establishment of nitrogen- fixing strawberry (Fragaria • annassa) via artificial symbiosis with Azomonas insignis. In Vitro Cell. Dev. Biol. 33P:190-194; 1997.
Rchid, H.; Baccou, J. C. Effect of light and sucrose on the ultrastructure of plastids and on the growth and steroidic sapogenin production of Tri- gonella foenum-graecum L. cell suspension cultures. In: Carre, E; Chagvardieff, P., ed. Ecophysiology and photosynthetic in vitro cul- tures. Aix-en-Provence (France): CEA; 1995:87-92.
Reed, B. M.; Tanprasert, P. Detection and control of bacterial contaminants of plant tissue cultures. A review of recent literature. Plant Tissue Culture Biotech. 1:137-142; 1995.
Richards, J. Induced resistance responses in potato inoculated in vitro with a plant growth promoting pseudomonad bacterium. M.Sc. thesis, Dal- housie University, Halifax, NS, Canada; 1997.
Rovira, A. D. Rhizosphere research---85 years progress and frustration. In: Keister, D. L.; Cregan, P. B., ed. Dordrecht (NL): Kluwer Acad. Publ.; The rhizosphere and plant growth. 1991:3-13.
Schippers, B.; Bakker, A. W.; Bakker, P. A. H. M. Interactions of deleterious and beneficial rhizosphere microorganisms and the effect of cropping practices. Ann. Rev. Phytopathol. 25:339-358; 1987.
Sharma, V. K.; Nowak, J. Verticillium wilt suppression in tomato with pseu- domonad bacterium. Can. J. Microbiol. (in press); 1998.
Sheen, J. Ca2+-dependent protein kinases and stress signal transduction in plants. Science 274:1900-1902; 1996.
Shetty, K.; Curtis, O. F.; Levin, R. E., et al. Prevention of vitrification asso- ciated with in vitro shoot culture of oregano (Origanum vulgare) by Pseudomonas spp. J. Plant Physiol. 147:447-451; 1995.
Stewart, A. H. Suppression of verticillium wilt in potatoes with a plant growth promoting rhizobacterium. M.Sc. thesis, Dalhousie University, Hali- fax, NS, Canada; 1997.
Sturz, A. V. The role of endophytic bacteria during seed piece decay and potato tuberization. Plant Soil 175:257-263; 1995.
Sturz, A. V.; Christie, B. R. Endophytic bacterial systems governing red clover growth and development. Ann. Appl. Biol. 126:285-290; 1995.
Sturz, A. V.; Christie, B. R.; Matheson, B. G., et al. Biodiversity of endophytic bacteria which colonize red clover nodules, roots, stems and foliage and their influence on host growth. Biol. Fert. Soils 25:13-19; 1997.
Van Huylenbroeck, J. M.; Debergh, P. C. Physiological aspects in acclima- tization of micropropagated plantlets. Plant Tissue Cult. Biotech. 2:136-141; 1996.
Varga, Sz. S.; Kortinyi, P.; Preininger, E., et at. Artificial associations between Daocus and nitrogen-fixing Azotobacter ceils in vitro. Physiol. Plant. 90:786-790; 1994.
Visser-Tenyenhuis, C.; Odumern, J.; Saxena, P. K., et al. Modulation of so- matic embryogenesis in hypocotyl derived cultures of geranium (Pel- argonium • horturum Bailey) cv. Ringo Rose by a bacterium. In Vitro Cell. Dev. Biol. 30P:140-143; 1994.
Wake, H.; Akasaka, A.; Umetsu, H., et al. Promotion of plantlet formation from somatic embryos of carrot treated with a high molecular weight extract from marine cyanobacterium. Plant Cell Rep. 11:62-65; 1992.
Wake, H.; Umetsu, H.; Ozeki, Y., et al. Extracts of marine cyanobacteria stimulated somatic embryogenesis ofDaucus carota L. Plant Cell Rep. 9:655-658; 1991.
Walley, E L.; Germida, J. J. Response of spring wheat (Triticum aestivum) to interactions between Pseudomonas species and Glomus clarum NT4. Biol. Fertil. Soils 24:365-367; 1997.
Wang, H.; Parent, S.; Gosselin, A., et al. Study of vesicular-arbuscular my- corrhizal peat-based substrates on symbiosis establishment, accli- matization and growth of three micropropagated species. J. Am. Soc. Hort. Sci. 118:896-901; 1993.
Wilhelm, E.; Arthofer, W.; Schafleitner, R., et al. Bacillus subtilis an endo- phyte of chestnut (Castanea sativa) as antagonist against chestnut blight (Cryphonectaria parasitica). In: Cassells, A., ed. Pathogen and microbial contamination management in micropropagation. Dordrecht (NL): Kluwer Acad. Publ.; 1997:331-337.
Yang, Y.-S.; Wada, K.; Goto, M., et al. In vitro formation of nodular calli in soybean (Glycine max L.) induced by co-cultivated Pseudomonas mal- tophilia. Japan. J. Breed. 41:5954504; 1991.
Yu, K.; Christie, B. R.; Paulus, K. E Effects of Verticillium albo-atrum culture filtrate on somatic embryogenesis in alfalfa. Plant Cell Rep. 8:509- 511; 1990.
Zehnder, G.; Kloepper, J.; Tuzun, S., et al. Insect feeding on cucumber me- diated by rhizobacteria-induced plant resistance. Entomol. Exp. Appl. 83:81~35; 1997.
Ziv, M. Vitrification: morphological and physiological disorders of in vitro plants. In: Debergh, P. C.; Zimmerman, R. H., ed. Micropropagation, technology and applications. Dordrecht (NL): Kluwer Acad. Publ.; 1991:45-69.
Ziv, M.; Schwartz, A.; Fleminger, D. Malfunctioning stomata in vitreous leaves of carnation (Dianthus caryophyllus) plants propagated in vitro; im- plications for hardening. Plant Sci. 52:127-134; 1987.
Related Documents











