Drug Development Research 20:337-348 (1990) Behavioral Contingencies Determine Changes in Drug-Induced Neurotransmitter Turnover James E. Smith and Steven I. Dworkin Departments of Pharmacology and Psychiatry, Louisiana State University School of Medicine, Shreveport, Louisiana Key words: amino acids, biogenic amines, receptors, brain reinforcement mechanisms, ce- rebral glucose metabolism, dopamine, a-aminobutyric acid, norepinephrine, glutamate, intracranial electrical self-stimulation, intravenous opiate self-admini- stration, neuronal networks, rat brain regions, serotonin, 2-deoxyglucoseautora- diography Smith, J.E., and S.I. Dworkin: Behavioral contingencies determine changes in drug-in- duced neurotransmitterturnover. Drug Dev. Res. 20:337-348, 1990. Evidence from several research areas suggests that response-dependent and -indepen- dent presentation of environmental events have different neurobiological consequences. Investigationsof neurotransmitterturnover rates and cholinergic muscarinic receptor den- sities in brain regions of rats receiving contingent and noncontingent intravenous morphine resulted in the identification of two neuronal networks that are involved in response-depen- dent morphine presentation. These circuits are concordant with brain regions showing increasedglucose utilization in animals receiving response-dependent electrical stimulation of either the substantia nigra or ventral tegmental area. These data indicate substantial differences between contingent and noncontingent presentation of environmental events and suggest that neurobiological investigations of reinforcement that use response-inde- pendent delivery of a putative reinforcer must be interpreted with some caution. INTRODUCTION The development of research techniques during the past decade has resulted in a steadily increasing volume of data suggesting behavior to have significant neurochemical consequences [reviewed by Dworkin and Smith, 19891. This discourse evaluates the current evidence for the role of the response contingency in the neurobiological consequences of exposure to environ- Received final version December 20, 1989; accepted January 25, 1990. Address reprint requests to Dr. James E. Smith, Physiology and Pharmacology, Bowman Gray School of Medicine, Wake Forest University, 300 S. Hawthorne Road, Winston-Salem, NC 27103. 0 1990 Wiley-Liss, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Drug Development Research 20:337-348 (1990)
Behavioral Contingencies Determine Changes in Drug-Induced Neurotransmitter Turnover James E. Smith and Steven I. Dworkin
Departments of Pharmacology and Psychiatry, Louisiana State University School of Medicine, Shreveport, Louisiana
Key words: amino acids, biogenic amines, receptors, brain reinforcement mechanisms, ce- rebral glucose metabolism, dopamine, a-aminobutyric acid, norepinephrine, glutamate, intracranial electrical self-stimulation, intravenous opiate self-admini- stration, neuronal networks, rat brain regions, serotonin, 2-deoxyglucose autora- diography
Smith, J.E., and S.I. Dworkin: Behavioral contingencies determine changes in drug-in- duced neurotransmitter turnover. Drug Dev. Res. 20:337-348, 1990.
Evidence from several research areas suggests that response-dependent and -indepen- dent presentation of environmental events have different neurobiological consequences. Investigations of neurotransmitter turnover rates and cholinergic muscarinic receptor den- sities in brain regions of rats receiving contingent and noncontingent intravenous morphine resulted in the identification of two neuronal networks that are involved in response-depen- dent morphine presentation. These circuits are concordant with brain regions showing increased glucose utilization in animals receiving response-dependent electrical stimulation of either the substantia nigra or ventral tegmental area. These data indicate substantial differences between contingent and noncontingent presentation of environmental events and suggest that neurobiological investigations of reinforcement that use response-inde- pendent delivery of a putative reinforcer must be interpreted with some caution.
INTRODUCTION
The development of research techniques during the past decade has resulted in a steadily increasing volume of data suggesting behavior to have significant neurochemical consequences [reviewed by Dworkin and Smith, 19891. This discourse evaluates the current evidence for the role of the response contingency in the neurobiological consequences of exposure to environ-
Received final version December 20, 1989; accepted January 25, 1990.
Address reprint requests to Dr. James E. Smith, Physiology and Pharmacology, Bowman Gray School of Medicine, Wake Forest University, 300 S. Hawthorne Road, Winston-Salem, NC 27103.
0 1990 Wiley-Liss, Inc.

338 Smith and Dworkin
mental events and assesses whether central nervous system (CNS) correlates of response dependency are different from response-independent presentation of the same event. It is generally accepted that there are biological substrates of behavior. However, basic knowledge of such processes is sparse for something that is so intuitively obvious. This results largely from a number of circumstances that include the general difficulties in identifying adequate and appropriate experimental designs and the dynamic and complex nature of the CNS. With regard to the first issue, it is often difficult to define control groups permitting delineation of the neurobiological consequences of specific behavioral events, since response requirements or consummatory responses are intrinsic variables in the experimental design. Thus, the identi- fication of neurobiological changes includes procedural differences that may obviate recogni- tion of behavior-specific CNS events. It is also difficult to assess something when the actual process of measurement modifies the entity being measured [Heisenberg, 19581. Frequently the behavior of the organism, the neuronal processes themselves, or the interaction of behavior with brain processes are modified by the procedures of measurement, again confounding the results. Identification of the neurobiological substrates of specific behaviors is clearly a very difficult challenge for the neurobehavioral scientist. In spite of these difficulties, an expanding body of evidence indicates that the CNS effects of contingently presented environmental events are different from noncontingent presentation of the same event.
NEUROTRANSMITTER TURNOVER RATES WITH CONTINGENT AND NONCONTINGENT MORPHINE PRESENTATION
A series of studies were initiated to identify the neurotransmitter turnover rate changes that occur in intravenous morphine self-administering rats. Groups of three male littermate rats were implanted with jugular catheters; two of the littermates in each triad were made physi- cally dependent on morphine with hourly infusions while the third rat received yoked injec- tions of the vehicle [Smith et al., 19821. One of the two physically dependent littermates was allowed to self-administer morphine with continuous access by lever pressing initially on a fixed ratio one (FR 1) schedule of presentation that was rapidly increased to FR 10, while the other physically dependent animal received identical yoked infusions of morphine and the third littermate, yoked infusions of the vehicle. This design permitted identification of neurotrans- mitter turnover rate changes resulting from the response contingent versus noncontingent presentation of drug in the absence of differences in the density of infusions and with mini- mized differences in the response requirement. The density of morphine infusion between the two rats receiving the drug in each litter was identical, since infusions occurred concurrently. The response requirement was also minimized, with the 10 responses for each infusion oc- curring on the average of once every 2 hr. When stable rates of intake were obtained, permitting the calculation of an accurate interinfusion interval (6-10 weeks of 24-hr access), the litter was pulse-labeled with high specific radioactivity precursors to selected neurotrans- mitters (3H-tryptophan, 'H-tyrosine, and ''C-glucose in one experiment and 'H-choline in another). The litter was killed by immersion in liquid nitrogen at the time that the self- administering rat was predicted to end the postinjection pause, which was also one of two time points on the linear decay in radioactivity curve for the neurotransmitters being evaluated (at 60 and 90 min for the biogenic amine and amino acid neurotransmitters and at 5 and 10 min for acetylcholine). The brains were removed at -2O"C, stored at -7O"C, dissected at -2O"C, and pulverized in liquid nitrogen. The pulverized frozen tissue was extracted with formic acid/acetone and the content and specific radioactivity of the pulse labeled neurotransmitters determined.
Biogenic Amine Neurotransmitters
Only the statistically significant differences in the turnover rates of the designated neurohumor follow, presented as the percentage of turnover rates in the vehicle controls.
Drug-Induced Neurotransmitter Turnover 339
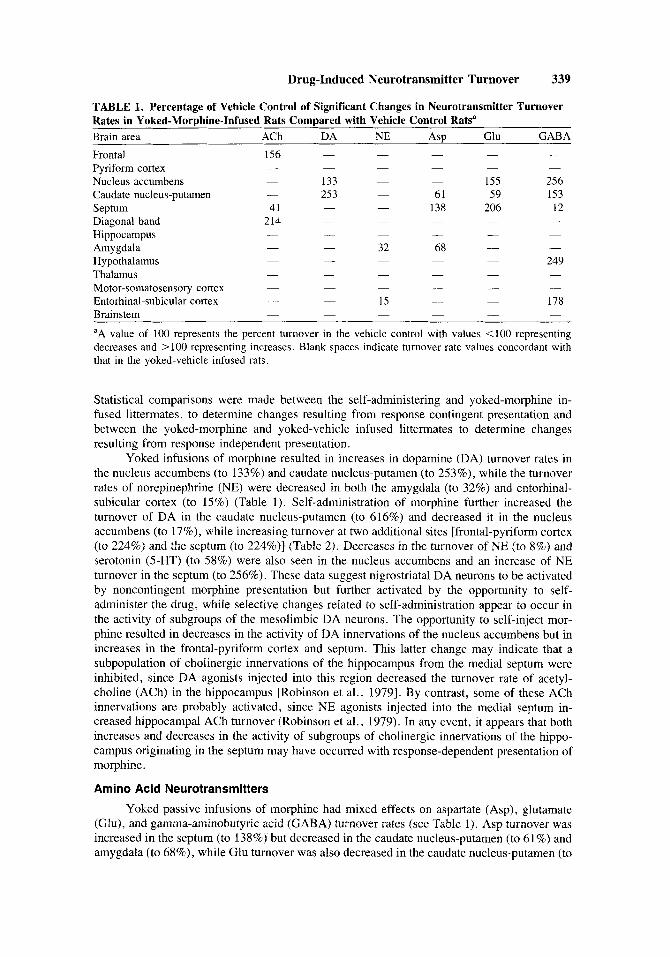
TABLE 1. Percentage of Vehicle Control of Significant Changes in Neurotransmitter Turnover Rates in Yoked-Morphine-Infused Rats Compared with Vehicle Control Ratsa Brain area ACh DA NE ASP Glu GABA Frontal 156 Pyriform cortex Nucleus accumbens - 133 - - 155 256 Caudate nucleus-putamen - 253 - 61 59 153
Diagonal band 214 Hippocampus Amygdala - 32 68 - -
- 249 Thalamus -
- 178 Entorhinal-subicular cortex - - Brainstem aA value of 100 represents the percent turnover in the vehicle control with values <lo0 representing decreases and > 100 representing increases. Blank spaces indicate turnover rate values concordant with that in the yoked-vehicle infused rats.
- - - - - - - - - - -
Septum 41 - - 138 206 12 - - - - -
- - - - - -
-
- Hypothalamus - - -
Motor-somatosensory cortex - - - - - - - - - -
-
- 15 - - - - - -
Statistical comparisons were made between the self-administering and yoked-morphine in- fused littermates, to determine changes resulting from response contingent presentation and between the yoked-morphine and yoked-vehicle infused littermates to determine changes resulting from response independent presentation.
Yoked infusions of morphine resulted in increases in dopamine (DA) turnover rates in the nucleus accumbens (to 133%) and caudate nucleus-putamen (to 253%), while the turnover rates of norepinephrine (NE) were decreased in both the amygdala (to 32%) and entorhinal- subicular cortex (to 15%) (Table 1). Self-administration of morphine further increased the turnover of DA in the caudate nucleus-putamen (to 616%) and decreased it in the nucleus accumbens (to 17%), while increasing turnover at two additional sites [frontal-pyriform cortex (to 224%) and the septum (to 224%)] (Table 2). Decreases in the turnover of NE (to 8%) and serotonin (5-HT) (to 58%) were also seen in the nucleus accumbens and an increase of NE turnover in the septum (to 256%). These data suggest nigrostnatal DA neurons to be activated by noncontingent morphine presentation but further activated by the opportunity to self- administer the drug, while selective changes related to self-administration appear to occur in the activity of subgroups of the mesolimbic DA neurons. The opportunity to self-inject mor- phine resulted in decreases in the activity of DA innervations of the nucleus accumbens but in increases in the frontal-pyriform cortex and septum. This latter change may indicate that a subpopulation of cholinergic innervations of the hippocampus from the medial septum were inhibited, since DA agonists injected into this region decreased the turnover rate of acetyl- choline (ACh) in the hippocampus [Robinson et al., 19791. By contrast, some of these ACh innervations are probably activated, since NE agonists injected into the medial septum in- creased hippocampal ACh turnover (Robinson et al., 1979). In any event, it appears that both increases and decreases in the activity of subgroups of cholinergic innervations of the hippo- campus originating in the septum may have occurred with response-dependent presentation of morphine.
Amino Acid Neurotransmitters
Yoked passive infusions of morphine had mixed effects on aspartate (Asp), glutamate (Glu), and gamma-aminobutyric acid (GABA) turnover rates (see Table 1). Asp turnover was increased in the septum (to 138%) but decreased in the caudate nucleus-putamen (to 61%) and amygdala (to 68%), while Glu turnover was also decreased in the caudate nucleus-putamen (to
340 Smith and Dworkin
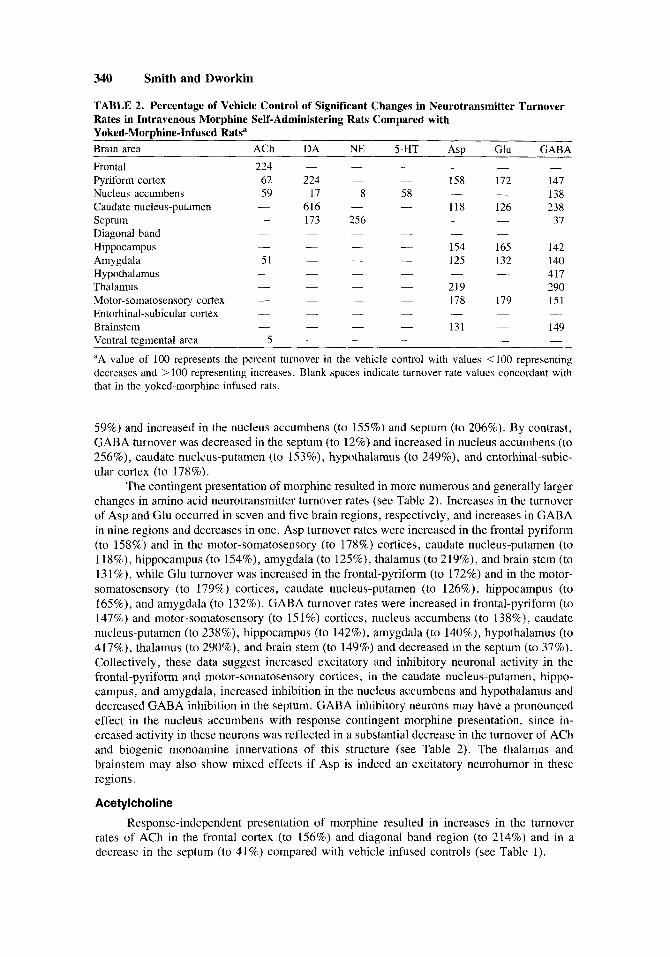
TABLE 2. Percentage of Vehicle Control of Significant Changes in Neurotransmitter Turnover Rates in Intravenous Morphine Self-Administering Rats Compared with Yoked-MorDhine-Infused Ratsa
~ ~~
Brain area ACh DA NE 5-HT ASP Glu GABA Frontal 224 - Pyriform cortex 62 224 - 158 172 147
138 Nucleus accumbens 59 17 8 58 616 - - 118 126 238 Caudate nucleus-putamen -
37 Septum - 173 256 - - Diagonal band - -
- 154 165 142 Hippocampus - - - 125 132 140 Amygdala 51
- 417 Hypothalamus - - - - - 290 Thalamus
- 178 179 151 Motor-somatosensory cortex - - Entorhinal-subicular cortex - - - - - - -
149 Brainstem - - - 131 -
Ventral tegmental area 5 -
- - - - - -
- -
-
- - - - - -
- -
- - - - 219 -
-
-
- - - - -
aA value of 100 represents the percent turnover in the vehicle control with values <I00 representing decreases and > 100 representing increases. Blank spaces indicate turnover rate values concordant with that in the yoked-morphine infused rats.
59%) and increased in the nucleus accumbens (to 155%) and septum (to 206%). By contrast, GABA turnover was decreased in the septum (to 12%) and increased in nucleus accumbens (to 256%), caudate nucleus-putamen (to 153%), hypothalamus (to 249%), and entorhinal-subic- ular cortex (to 178%).
The contingent presentation of morphine resulted in more numerous and generally larger changes in amino acid neurotransmitter turnover rates (see Table 2 ) . Increases in the turnover of Asp and Glu occurred in seven and five brain regions, respectively, and increases in GABA in nine regions and decreases in one. Asp turnover rates were increased in the frontal-pyriform (to 158%) and in the motor-somatosensory (to 178%) cortices, caudate nucleus-putamen (to 118%), hippocampus (to 154%), amygdala (to 125%), thalamus (to 219%), and brain stem (to 131%), while Glu turnover was increased in the frontal-pyriform (to 172%) and in the motor- somatosensory (to 179%) cortices, caudate nucleus-putamen (to 126%), hippocampus (to 165%), and amygdala (to 132%). GABA turnover rates were increased in frontal-pyriform (to 147%) and motor-somatosensory (to 15 1%) cortices, nucleus accumbens (to 138%), caudate nucleus-putamen (to 238%), hippocampus (to 142%), amygdala (to 140%), hypothalamus (to 417%), thalamus (to 290%), and brain stem (to 149%) and decreased in the septum (to 37%). Collectively, these data suggest increased excitatory and inhibitory neuronal activity in the frontal-pyriform and motor-somatosensory cortices, in the caudate nucleus-putamen, hippo- campus, and amygdala, increased inhibition in the nucleus accumbens and hypothalamus and decreased GABA inhibition in the septum. GABA inhibitory neurons may have a pronounced effect in the nucleus accumbens with response contingent morphine presentation, since in- creased activity in these neurons was reflected in a substantial decrease in the turnover of ACh and biogenic monoamine innervations of this structure (see Table 2 ) . The thalamus and brainstem may also show mixed effects if Asp is indeed an excitatory neurohumor in these regions.
Acetylcholine Response-independent presentation of morphine resulted in increases in the turnover
rates of ACh in the frontal cortex (to 156%) and diagonal band region (to 214%) and in a decrease in the septum (to 41%) compared with vehicle infused controls (see Table 1).

Drug-Induced Neurotransmitter Turnover 341
TABLE 3. Percentage of Vehicle Control of Significant Changes in Density of Cholinergic Muscarinic (QNB) and Benzodiazepine (FLU) Receptors in Membranes Prepared from Brain Areas of Rats Receiving Yoked Infusions of Morphine Compared with Vehicle Control Rats" Brain area QNB FLU Frontal cortex - -
Pyriform cortex 111 - Cingulate cortex 79 -
- Entorhinal-subicular cortex 71 Hippocampus 87 Amygdala - -
Caudate nucleus-putamen - -
"A value of 100 represents the percentage of binding sites in the vehicle control with values < 100 representing decreases and values > 100 representing increases. Blank spaces indicate binding site densities concordant with that in the yoked-vehicle infused rats.
-
Self-administration of morphine resulted in decreases in the turnover rates of ACh in four brain regions and an increase in one (see Table 2). Decreases were seen in the pyriform cortex (to 62%), nucleus accumbens (to 59%), amygdala (to 51%), and ventral tegmental area (to 15%) and a further increase in the frontal cortex (to 224%). These data suggest that frontal cortical cholinergic innervations from the ventral pallidum are activated by intravenous mor- phine but that they are even further activated by the opportunity to self-administer the drug. The decreases in turnover rates in the pyriform cortex, nucleus accumbens, and amygdala suggest that the opportunity to control drug presentation decrements activity in cholinergic neurons innervating these regions from the diagonal band. It appears that these two cholinergic cell nuclei are affected differently by self-administration. The large decrease in ACh turnover in the ventral tegmental area of the self-administering rats represent a decrease in the activity of the cholinergic innervations from the forebrain that pass through the lateral habenulae area. However, these turnover changes represent only the cholinergic neuronal activity occurring during the 5- and 10- min period immediately before the predicted self-infusion hence may not represent overall net changes in neuronal activity during the 2-hr interinfusion interval and definitely not changes that occurred immediately after the last infusion. For this reason, cholinergic muscarinic binding was assessed in membranes prepared from other portions of these same brain regions as another index of cholinergic neuronal activity.
NEUROTRANSMITTER RECEPTOR BINDING WITH CONTINGENT AND NONCONTINGENT MORPHINE PRESENTATION
Membranes were prepared from portions of the pulverized tissue samples and the bind- ing of quinuclidinyl benzilate (QNB) for cholinergic muscarinic receptors and flunitrazepam (FLU) for benzodiazepine receptors assessed in individual tissue samples [Smith et al., 1984bl.
Cholinergic Muscarinic Receptors
Response-independent administration of intravenous morphine to physically dependent rats resulted in a significant increase in the density of QNB binding sites (Bma..) in the pyriform cortex (to 11 1 % of vehicle controls) and a decrease in the cingulate cortex (to 79% of vehicle controls) (Table 3). Neither of these areas showed increases in ACh turnover during the

342 Smith and Dworkin
TABLE 4. Percentage of Vehicle Control of Significant Changes in Density Cholinergic Muscarinic (QNB) and Benzodiazepine (FLU) Receptors in Membranes Prepared from Brain Areas of Rats Intravenously Self-administering Morphine Compared with the Yoked-MorDhine Infused Rats" Brain area QNB FLU Frontal cortex Pyriform cortex Cingulate cortex Entorhinal-subicular cortex
68 - -
87 Hippocampus - -
Amygdala 120 -
Caudate nucleus-putamen - -
"A value of 100 represents the percentage of binding sites in the vehicle control with values < 100 representing decreases and > 100 representing increases. Blank spaces indicate binding site densities concordant with that in the yoked-morphine ve- hicle infused rats.
pulse-label interval, but these receptor changes suggest that a net decrease in release of ACh may have occurred in the pyriform cortex and an increase in the cingulate cortex that resulted in an upregulation and downregulation in binding sites respectively.
The self-administering animals showed significant decreases in binding in the frontal (to 68% of vehicle controls) and entorhinal-subicular (to 87%) cortices and an increase in the amygdala (to 120%) (Table 4). The frontal cortex and amygdala demonstrated significant and opposite changes in the turnover rates of ACh suggesting net changes in release had occurred, while no change in ACh turnover was detected in the entorhinal-subicular cortex when a net increase in release may have occurred as reflected by the decrease in binding sites. These data collectively suggest that increases in the activity of cholinergic innervations of the frontal cortex from the ventral pallidum and of the entorhinal-subicular cortex from the amygdala and decreases in activity of innervations of the amygdala from the preoptic nuclei may have occurred in the self-administering rats.
Benzodiatepine Receptors
The noncontingent presentation of intravenous morphine resulted in a significant de- crease in flunitrazepam binding in the entorhinal-subicular cortex (to 71%) and in the hippo- campus (to 87%) that was also present in the self-administering rats, suggesting increases in the release of endogenous ligands for these binding sites. Opiates decrease some of the anxiety that accompanies aversive stimuli [Jaffe and Martin, 19851. These changes might be related to these actions and do not appear to be involved in the reinforcing effects of morphine as demonstrated with self-administration.
NEURONAL NETWORKS IDENTIFIED WITH CONTINGENT MORPHINE PRESENTATION
These turnover rate and receptor binding data were integrated with information of neurotransmitter specific pathways, resulting in the identification of two neuronal networks that may be involved in the basic mechanisms mediating opiate self-administration.

Drug-Induced Neurotransmitter Turnover 343
ENTORHINAL
Fig. 1. Neuronal circuits mediating effects of response-contingent morphine. These include a frontal cortex-caudate nucleus circuit involving glutamate and acetylcholine pathways with aspartate and GABA interneurons, and a hippocampal-septum-nucleus accumbens-preoptic nuclei-amygdala-entorhinal cortex- hippocampus circuit also involving glutamate and acetylcholine pathways and aspartate and GABA interneurons in the nucleus accumbens and hippocampus.
FRONTAL CORTEX AND HIPPOCAMPUS CIRCUITS
These include a frontal cortex-caudate nucleus-ventral pallidal-frontal cortex circuit and a nucleus accumbens-preoptic nuclei-amygdala- entorhinal cortex-hippocampus-nucleus ac- cumbens circuit (Fig. 1). These circuits appear to involve cholinergic and glutamatergic pathways and aspartatergic interneurons in the frontal cortex, nucleus accumbens, hippocam- pus, and ventral tegmental area and GABAergic interneurons in the caudate nucleus-putamen, frontal cortex, hippocampus, and nucleus accumbens.
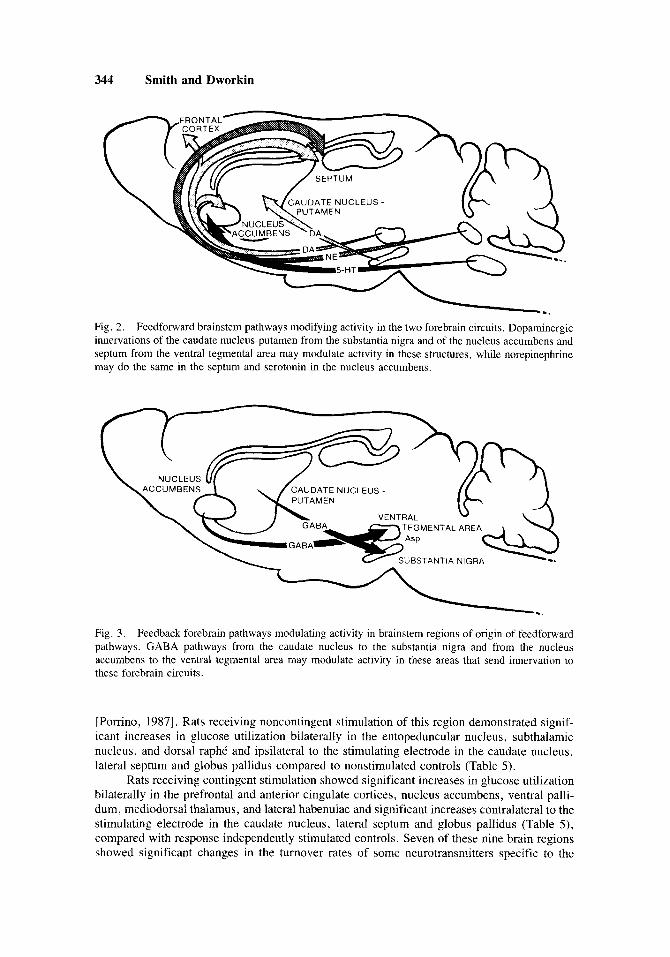
Feedforward Pathways
The activity in these two networks appear to be modulated by feedfonvard pathways from the brainstem (Fig. 2 ) . These could include dopaminergic innervations of the nucleus accumbens, frontal cortex, and septum from the ventral tegmental area and of the caudate nucleus-putamen from the substantia nigra, serotonergic innervations of the nucleus accum- bens from the dorsal raphC nuclei and noradrenergic innervations of the septum from the locus coeruleus . Feedback Pathways
Activity in these brainstem centers may be modulated by feedback pathways from these two networks (Fig. 3). These include GABAergic pathways from the caudate nucleus-putamen innervating the substantia nigra and from the nucleus accumbens innervating the ventral tegmental area. This latter system may include a descending aspartatergic pathway from some unknown forebrain site or aspartatergic interneurons in the ventral tegmental area. Other evidence potentially supporting the involvement of these two neuronal networks in response- contingent presentation (reinforcement processes) has come from experiments on glucose utilization in rats receiving contingent and noncontingent electrical brain stimulation (ICSS).
BRAIN AREAS SHOWING INCREASED GLUCOSE UTILIZATION WITH ICSS Substantia Nigra ICSS
The distribution of 2-deoxyglucose has been evaluated in brain regions of rats receiving either response dependent or response-independent brain stimulation of the substantia nigra
344 Smith and Dworkin
c '1 SEPTUM
CAUDATE NUCLEUS -
Fig. 2. Feedforward brainstem pathways modifying activity in the two forebrain circuits. Dopaminergic innervations of the caudate nucleus-putamen from the substantia nigra and of the nucleus accumbens and septum from the ventral tegmental area may modulate activity in these structures, while norepinephrine may do the same in the septum and serotonin in the nucleus accumbens.
Fig. 3. Feedback forebrain pathways modulating activity in brainstem regions of origin of feedforward pathways. GABA pathways from the caudate nucleus to the substantia nigra and from the nucleus accumbens to the ventral tegmental area may modulate activity in these areas that send innervation to these forebrain circuits.
[Porrino, 19871. Rats receiving noncontingent stimulation of this region demonstrated signif- icant increases in glucose utilization bilaterally in the entopeduncular nucleus, subthalamic nucleus, and dorsal raphC and ipsilateral to the stimulating electrode in the caudate nucleus, lateral septum and globus pallidus compared to nonstimulated controls (Table 5) .
Rats receiving contingent stimulation showed significant increases in glucose utilization bilaterally in the prefrontal and anterior cingulate cortices, nucleus accumbens, ventral palli- dum, mediodorsal thalamus, and lateral habenulae and significant increases contralateral to the stimulating electrode in the caudate nucleus, lateral septum and globus pallidus (Table S ) , compared with response independently stimulated controls. Seven of these nine brain regions showed significant changes in the turnover rates of some neurotransmitters specific to the

Drug-Induced Neurotransmitter Turnover 345
TABLE 5. Brain Areas Showing Increased Glucose Utilization in Rats Receiving Either Response- Contingent or Response-Independent Electrical Stimulation of the Substantia Nigra through a Unilateral Electrodea Bilateral lpsilateral Self-stimulating animals
Prefrontal cortex Anterior cingulate cortex Nucleus accumbens Ventral pallidum Medial dorsal thalamus Lateral habenulae Caudate nucleus Lateral septum Globus pallidus Entopeduncular nucleus Subthalamic nucleus Dorsal raphe
Noncontingent stimulation Entopeduncular nucleus Subthalamic nucleus Dorsal rapht
Caudate nucleus Lateral septum Globus pallidus
"From Porrino 119871, with permission.
self-administration of morphine (see Table 2) and six of these regions are components of the putative neuronal networks proposed to mediate opiate reinforcement (Figs. 1, 2). This is a significant degree of concordance.
Ventral Tegmental Area ICSS
This same laboratory has also investigated brain glucose utilization in rats receiving contingent and noncontingent stimulation of the ventral tegmental area [Porrino et al., 19841. Rats receiving noncontingent stimulation of these region showed increased glucose utilization bilaterally in the locus coeruleus and dorsal raphC nucleus and ipsilaterally to the stimulating electrode in the lateral septum, mediodorsal thalamus and hippocampus CA3 region compared to nonstimulated controls (Table 6).
In the rats receiving response-contingent stimulation, increases in activity occurred bilaterally in the nucleus accumbens, bed nucleus of the stria terminalis and medial bracial nucleus, ipsilaterally to the stimulating electrode in the medial prefrontal cortex, central amygdala and basolateral amygdala, and contralaterally to the stimulating electrode in the lateral septum, mediodorsal thalamus and CA3 area of the hippocampus (Table 6). Increases in activity that occurred bilaterally in the locus coeruleus and dorsal raphC and ipsilaterally to the stimulating electrode in the lateral septum, mediodorsal thalamus and CA3 area of the hippocampus were related to the electrical stimulation itself since these were seen in the noncontingently stimulated rats as well. Again, six of the nine areas where increases in glucose utilization occurred that appear to be related to the response contingent presentation of rein- forcing electrical brain stimulation also showed changes in neurotransmitter turnover rates correlated with the response contingent presentation of intravenous morphine (Table 2) and are part of the putative networks proposed to mediate this behavior (see Figs. 1, 2).

346 Smith and Dworkin
TABLE 6. Brain Regions Showing Increases in Glucose Utilization in Rats Receiving Either Response-Contingent or Response-Independent Electrical Stimulation of the Ventral Tegmental Area through a Unilateral Electrode.a Bilateral Iusilateral Self-stimulating animal
Nucleus accumbens Bed nucleus of stria terminalis Medial parabracial nucleus Lateral septum Mediodorsal thalamus Hippocampus (CA3) Locus coeruleus Dorsal raphi
Noncontingent stimulation Locus coeruleus Dorsal raphC
Lateral septum Mediodorsal thalamus Hippocampus (CA3)
“From Porrino 119871, with permission.
INTRINSIC DIFFERENCES BETWEEN CONTINGENT AND NONCONTINGENT DRUG Presentation
These neurochemical differences between response contingent and response independent presentation of stimuli that have the capacity to serve as reinforcers under appropriate condi- tions suggest that data from experimental procedures that utilize noncontingent presentation to investigate brain mechanisms of reinforcement should be interpreted with caution. These include conditioned place preference and drug discrimination. Conditioned place preference (CPP) is a procedure with which a learned preference is thought to represent the development of a positive valence by previously neutral environmental stimuli paired with the noncontin- gent presentation of a drug that produces a “pleasant” effect. It is assumed that the admin- istration of a substance has intrinsic hedonic value in the absence of the behavior of the animal. Noncontingent presentation is thought to have the same positive effect as contingent presen- tation (reinforcement). Thus, “reward” neurons are activated independent of the control of the organism. However, as the foregoing data suggests, to conclude that response independent presentation of any potential reinforcer is pleasing may be in error. The drug discrimination procedure is subject to the same criticisms since noncontingent drug presentation occurs. With this procedure, drug cues are used to occasion a particular response that results in the presen- tation of a nondrug reinforcer. The procedure uses the entire stimulus properties of the drug, which could include “pleasing” effects, but the response-independent presentation of the drug precludes conclusions concerning reinforcing actions. Self-administration is the only nonhu- man laboratory model that can be used to directly address the reinforcing effects of a drug. However, the self-administration environment is a complex ambience with all of the behav- ioral effects of a drug present. This includes response contingent presentation as well as the stimulus properties, rate effects, effects on ingestive behaviors, and other behavioral actions of the drug. Thus, the self-administered drug has stimulus properties that can set the occasion for the emission of other responses, can increase or decrease the rates of behaviors including subsequent self-infusions, can modulate the intake of food or water which in turn can further increase or decrease drug intake or can modulate other unspecified physiological functions in

Drug-Induced Neurotransmitter Turnover 347
either a direct or indirect manner. Studies to investigate the neurobiological mechanisms of drug reinforcement must control for this myriad of drug actions when self-administration is chosen as the model.
GENERAL REINFORCEMENT CIRCUITS
It is improbable that unique and separate neuronal systems evolved to mediate the reinforcing properties of drugs. It is more likely that these substances activate neuronal sys- tems that mediate these processes for other reinforcers in much the same way as ICSS. The circuits proposed above could be portions of an endogenous system that mediates general reinforcement processes. There is a great deal of concordance between these circuits and the areas showing increased glucose utilization with response contingent electrical stimulation of the substantia nigra and ventral tegmental area. However, if it were this simple, why do lesions of components of these circuits appear to affect drug intake and not responding maintained by concurrent food or water presentation (Dworkin’s chapter in this volume)? This could result from a number of factors. It is likely that the involvement of conditioned stimuli will prove central to these differential effects and that, indeed, these may be portions of circuits dedicated to the processes of reinforcement. However, it is premature to draw such conclusions from the limited data currently available.
CONCLUSIONS
Reinforcement may be differentiated from reward phenomena. Reward has been used to describe immutable hedonic value or valence that occurs after the presentation of a pleasant event. Stimuli are thought to possess these properties independent of the behavior of the organism. Thus, the response-independent presentation of this stimulus is thought to be pleas- ant or rewarding. Reinforcement is a process that involves the increased probability of the occurrence of a behavior that preceded the presentation of a reinforcing event (Skinner, 1938). One may not preclude the response-contingent aspect of reinforcement and assume that an environmental event has intrinsic reinforcing properties independent of the organisms’s be- havior. The data and literature presented above demonstrate that response-dependent and response-independent presentation of an event clearly have different neurochemical conse- quences. Limited evidence suggests that under some circumstances response-independent presentation of an event that has previously been under the control of the organism may actually be aversive [Steiner et al., 19691. Noncontingent presentation of such electrical stimulation also alters glucose utilization in different brain regions than those that are affected when the stimulation is delivered response dependently [Porrino et al., 1984; Porrino, 19871. This suggests increased neuronal activity to occur in a number of brain regions of rats receiving the stimulation response contingently. Neurotransmitter turnover rate and receptor binding studies have also demonstrated differential effects of response-dependent and response-inde- pendent morphine presentation [Smith et al., 1980, 1982, 1984a,b]. Specific neuronal circuits are activated by the response contingent delivery of intravenous morphine. These neuronal networks may be portions of the systems that mediate the CNS components that occur after the contingent presentation of other environmental events that have putative reinforcing proper- ties.
ACKNOWLEDGMENTS
This paper was supported in part by research grants DA-01999, DA-03628, DA-03631, DA-03832, and DAK0500114 from the U.S. Public Health Service. We would like to thank Charlene Griffin for assistance in preparing this manuscript.

348 Smith and Dworkin
REFERENCES
Dworkin, S.I. and Smith, J.E.: The assessment of neurochemical correlates of behavior. In Boulton, A.A. Baker, G.B. and A.J. Greenshaw, A.J. (eds.): “Neuromethods: Psychopharmacology.” I: In press.
Heisenberg, W.: “Physics and Philosophy.” New York, Harper & Row, 1958. Jaffe, J.H. and Martin, W.R.: Opioid analgesics and antagonists. In Gilman, A., Goodman, L., Rall,
T.W. and Murad, F. (eds.): “The Pharmacological Basis of Therapeutics.” New York: Macmillan Publishing Co., 1985, pp. 491-531.
Pomno, L.J.: Cerebral metabolic changes associated with activation of reward systems. In Engel, J. and Oreland, L. (eds.): “Brain Reward Systems and Abuse.” New York: Raven Press, 1987, pp. 51-60.
Porrino, L.J., Esposito, R.U., Seeger, T.F., Crane, A.M., Pert, A. and Sokoloff, L.: Metabolic mapping of the brain during rewarding self-stimulation. Science 224:306-309, 1984.
Robinson, S.E., Malthe-Sorenssen, D., Wood, P.S. and Commission&, J.: Dopaminergic control of the septal-hippocampal cholinergic pathway. J. Pharmacol. Exp. Ther. 280:476-479, 1979.
Skinner, B.F.: “The Behavior of Organisms.” Englewood Cliffs, New Jersey: Prentice-Hall, 1938. Smith, J .E., Co, C., Freeman, M.E. and Lane, J.D.: Brain neurotransmitter turnover correlated with
morphine-seeking behavior in rats. Pharmacol. Biochem. Behav. 16:509-519, 1982. Smith, J.E., Co, C., Freeman, M.E., Sands, M.P. and Lane, J.D.: Neuro-transmitter turnover in the
striatum of rat brains is correlated with morphine self-administration. Nature (Lond.) 287: 152- 154, 1980.
Smith, J.E., Co, C. and Lane, J.D.: Limbic acetylcholine turnover rates correlated with rat morphine- seeking behaviors. Pharmacol. Biochem. Behav. 20:429-441, 1984a.
Smith, J.E., Co, C. and Lane, J.D.: Limbic muscarinic cholinergic and benzodiazepine receptor changes with chronic intravenous morphine and self-administration. Pharmacol. Biochem. Behav. 20: 443-450, 1984b.
Steiner, S.S., Beer, B. and Shaffer, M.M.: Escape from self-produced rates of brain stimulation. Science 163:90-91, 1969.
Related Documents











