3 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21 otras vías de transducción intracelulares relacionadas con los fenómenos de neuroplasticidad, que están cobrando cada vez mayor importancia en la fisiopatología del trastorno bipo- lar. Entre los factores neurotróficos relacionados con el meca- nismo de acción de los reguladores del humor cabe mencionar el BDNF, el ERK o el CREB, así como la proteína bcl-2, dotada de propiedades neuroprotectoras y antiapoptósicas. Palabras clave: Antiepilépticos. Trastorno bipolar. Fisiopatología. Mecanismo de acción. Vías de transduc- ción. Neuroprotección. Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21 Neurobiological basis of the use of antiepileptics in bipolar disorder Currently, antiepileptic drugs are a therapeutic alter- native to lithium when treating bipolar disorder. Among these, carbamazepine, valproate and lamotrigine have been authorized both for the treatment of manic phases and for the prevention of depressive episodes. In this pa- per, we will review the mechanisms of action of these drugs as euthymic agents, despite they have not been completely elucidated yet. At a synaptic level, the anti- convulsivant agents might act blocking the sodium vol- tage-depending channel with the subsequent stabiliza- tion of the neuronal membrane and the inhibtion of the release of excitatory neurotransmitters, modulating the glutamatergic mechanisms of neurotransmission. This is due to the action exerted on the NMDA, kainate or AMPA receptors or potentiating the gabergic mechanisms (increase of action of DAG enzyme, inhibition of GABA- transaminase, selective GABA recaptation or potentia- tion of GABA receptor’s agonist actions). With regard to the complicated molecular mechanisms associated to the signal transduction in postsynaptic neurones and their effect on antiepileptics, there are only significant data on carbamazepine and valproate. The most studied sig- nal transduction systems of secondary messengers are those referring to adenylate ciyclase and to the phos- phoinositol ways, which include A and C proteincinases, respectively. Regarding the euthymics, the most discus- sed hypothesis is the mioinositol depletion. It holds back a whole sequence of transduction events, where the phos- Los fármacos antiepilépticos constituyen en la actuali- dad una alternativa terapéutica al litio en el abordaje del trastorno bipolar. Entre ellos, la carbamazepina, el valproato y la lamotrigina han sido autorizados para el tratamiento, bien de las fases maníacas o para la prevención de episodios depresivos. En el presente trabajo revisaremos los mecanismos de acción de estos fármacos como agentes eutimizantes, a pe- sar de que aún no han sido completamente dilucidados. A nivel sináptico, los agentes anticonvulsivantes pueden ac- tuar bloqueando el canal de sodio dependiente de voltaje, con la consecuente estabilización de la membrana neuronal y la inhibición de la liberación de neurotransmisores excita- torios, modulando los mecanismos de neurotransmisión glutamatérgicos al actuar sobre los receptores de N-metil- D-aspartato (NMDA), kainato o ácido propiónico α-amino- 3-hidroxi-5-metil-4-isixazol (AMPA), o potenciando los gabérgicos (incremento de la acción de la enzima descarbo- xilasa del ácido glutámico (DAG), inhibición de la enzima γ-aminobutírico (GABA)-transaminasa, inhibición selectiva de la recaptación de GABA o potenciación de las acciones agonistas del receptor GABA). Con respecto a los complica- dos mecanismos moleculares relacionados con la transduc- ción de señales en las neuronas postsinápticas y al efecto sobre ellos de los antiepilépticos, únicamente se dispone de datos relevantes con carbamazepina y valproato. Los siste- mas de transducción de la señal de segundos mensajeros más estudiados son los referentes a la adenilatociclasa y a las vías del fosfoinositol, que incluyen a las proteincinasas A y C, respectivamente. En relación con los eutimizantes, la hipótesis más discutida es la de la depleción del mioinositol, que frena toda una cascada de efectos de transducción, en los que están implicados la fosfolipasa C, segundos mensajeros como el diacilglicerol y el inositol-trifosfato, la proteincina- sa C, la proteína MARCKS (myristoylated alanine-rich C kinase substrate) y los genes de respuesta inmediata c-jun, c-fos o Egr-1. Finalmente se comentarán los efectos de los euti- mizantes sobre la vía de la glucógeno-sintetasa-cinasa-3 y Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolar Departamento de Farmacología Facultad de Medicina Universidad de Alcalá de Henares Madrid Correspondencia: Cecilio Álamo González Jesús Aprendiz, 10-C, 5. o A 28007 Madrid Correo electrónico: [email protected] C. Álamo F. López-Muñoz J. A. Guerra

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

3Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
otras vías de transducción intracelulares relacionadas conlos fenómenos de neuroplasticidad, que están cobrando cadavez mayor importancia en la fisiopatología del trastorno bipo-lar. Entre los factores neurotróficos relacionados con el meca-nismo de acción de los reguladores del humor cabe mencionarel BDNF, el ERK o el CREB, así como la proteína bcl-2, dotadade propiedades neuroprotectoras y antiapoptósicas.Palabras clave:Antiepilépticos. Trastorno bipolar. Fisiopatología. Mecanismo de acción. Vías de transduc-ción. Neuroprotección.
Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
Neurobiological basis of the use of antiepileptics in bipolar disorder
Currently, antiepileptic drugs are a therapeutic alter-native to lithium when treating bipolar disorder. Amongthese, carbamazepine, valproate and lamotrigine havebeen authorized both for the treatment of manic phasesand for the prevention of depressive episodes. In this pa-per, we will review the mechanisms of action of thesedrugs as euthymic agents, despite they have not beencompletely elucidated yet. At a synaptic level, the anti-convulsivant agents might act blocking the sodium vol-tage-depending channel with the subsequent stabiliza-tion of the neuronal membrane and the inhibtion of therelease of excitatory neurotransmitters, modulating theglutamatergic mechanisms of neurotransmission. This is due to the action exerted on the NMDA, kainate or AMPA receptors or potentiating the gabergic mechanisms(increase of action of DAG enzyme, inhibition of GABA-transaminase, selective GABA recaptation or potentia-tion of GABA receptor’s agonist actions). With regard tothe complicated molecular mechanisms associated to thesignal transduction in postsynaptic neurones and theireffect on antiepileptics, there are only significant dataon carbamazepine and valproate. The most studied sig-nal transduction systems of secondary messengers arethose referring to adenylate ciyclase and to the phos-phoinositol ways, which include A and C proteincinases,respectively. Regarding the euthymics, the most discus-sed hypothesis is the mioinositol depletion. It holds backa whole sequence of transduction events, where the phos-
Los fármacos antiepilépticos constituyen en la actuali-dad una alternativa terapéutica al litio en el abordaje deltrastorno bipolar. Entre ellos, la carbamazepina, el valproatoy la lamotrigina han sido autorizados para el tratamiento,bien de las fases maníacas o para la prevención de episodiosdepresivos. En el presente trabajo revisaremos los mecanismosde acción de estos fármacos como agentes eutimizantes, a pe-sar de que aún no han sido completamente dilucidados. A nivel sináptico, los agentes anticonvulsivantes pueden ac-tuar bloqueando el canal de sodio dependiente de voltaje,con la consecuente estabilización de la membrana neuronaly la inhibición de la liberación de neurotransmisores excita-torios, modulando los mecanismos de neurotransmisiónglutamatérgicos al actuar sobre los receptores de N-metil-D-aspartato (NMDA), kainato o ácido propiónico α-amino-3-hidroxi-5-metil-4-isixazol (AMPA), o potenciando los gabérgicos (incremento de la acción de la enzima descarbo-xilasa del ácido glutámico (DAG), inhibición de la enzima γ-aminobutírico (GABA)-transaminasa, inhibición selectivade la recaptación de GABA o potenciación de las accionesagonistas del receptor GABA). Con respecto a los complica-dos mecanismos moleculares relacionados con la transduc-ción de señales en las neuronas postsinápticas y al efectosobre ellos de los antiepilépticos, únicamente se dispone dedatos relevantes con carbamazepina y valproato. Los siste-mas de transducción de la señal de segundos mensajerosmás estudiados son los referentes a la adenilatociclasa y alas vías del fosfoinositol, que incluyen a las proteincinasas Ay C, respectivamente. En relación con los eutimizantes, lahipótesis más discutida es la de la depleción del mioinositol,que frena toda una cascada de efectos de transducción, en losque están implicados la fosfolipasa C, segundos mensajeroscomo el diacilglicerol y el inositol-trifosfato, la proteincina-sa C, la proteína MARCKS (myristoylated alanine-rich C kinasesubstrate) y los genes de respuesta inmediata c-jun, c-foso Egr-1. Finalmente se comentarán los efectos de los euti-mizantes sobre la vía de la glucógeno-sintetasa-cinasa-3 y
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolar
Departamento de FarmacologíaFacultad de MedicinaUniversidad de Alcalá de HenaresMadrid
Correspondencia:Cecilio Álamo GonzálezJesús Aprendiz, 10-C, 5.o A28007 MadridCorreo electrónico: [email protected]
C. ÁlamoF. López-MuñozJ. A. Guerra
3-21.qxd 10/4/08 12:11 Página 3

polipase C, secondary messengers such as the diacylgly-cerol and the inositol triphosphate, the proteincinase C,the protein MARCKS and the immediate early responsegenes c-jun, c-fos or Egr-1 are involved. Lastly, we willdiscuss the euthymics effects on the glycogen synthasekinase 3’s way and other intracellular transduction waysrelated to the neuroplasticity phenomena which are gra-dually acquiring a higher significance in the bipolar di-sorder physiopathology. Among the neurotrophic factorsassociated to the mechanism of action of the mood regu-lators we should mention the BDNF, ERK or CREB, aswell as the protein bcl-2 which presents neuroprotectiveand antiapoptotic properties.Key words: Antiepileptics. Bipolar disorder. Phisiopathology. Mechanism of action. Transduction ways.Neuroprotection.
INTRODUCCIÓN
El trastorno bipolar es una de las entidades psiquiátricasque más atención está recibiendo en los últimos años en la li-teratura científica, como hemos puesto de manifiesto en unreciente estudio bibliométrico (López-Muñoz et al., 2006). Setrata de un trastorno del humor grave, de carácter crónico,caracterizado por la aparición, de forma continua o irregular,de episodios alternativos de manía/hipomanía con eutimia/episodios depresivos (Vieta, 2005). El trastorno bipolar, cono-cido clásicamente como enfermedad o psicosis maniacode-presiva, presenta una importante morbimortalidad (Bauer yPfenning, 2005), estimándose que su prevalencia a lo largo dela vida está comprendida entre el 1-2% de la población ma-yor de 20 años (Goodwin y Jamison, 1990). Pese a esta granrelevancia clínica, el arsenal farmacoterapéutico de este tras-torno ha sido muy limitado. En la última década la irrupciónde nuevos agentes farmacológicos, fundamentalmente an-tipsicóticos atípicos y anticonvulsivantes, ha creado nuevasperspectivas en el tratamiento de los pacientes bipolares.
El descubrimiento de las propiedades antimaníacas de lassales de litio en 1949 por parte del psiquiatra australianoJohn Cade abrió la puerta del trastorno bipolar a la terapéu-tica farmacológica (Cade, 1949). Sin embargo, a pesar de lasinteresantes perspectivas que apuntaba el trabajo de Cade,considerado por muchos autores como el punto de arranquede la denominada «revolución de la farmacología psiquiátri-ca» (López-Muñoz et al., 2000), las sales de litio no llegarona ser ampliamente utilizadas en psiquiatría hasta mediadosde la década de 1960 al demostrarse, mediante diversos es-tudios abiertos, controlados con placebo y de diseño dobleciego, los efectos profilácticos del tratamiento prolongadocon litio en los trastornos maníacos recurrentes y depresivos(Schou, 1996). De esta forma, la Food and Drug Adminis-tration aprobó finalmente este fármaco en 1970, 21 añosdespués del clásico artículo de Cade, y hoy día, a pesar desus limitaciones terapéuticas y efectos adversos, el litio si-gue formando parte del armamentarium del trastorno bi-polar (López-Muñoz et al., 2006).
La posterior incorporación de la carbamazepina y el val-proato como agentes reguladores del humor, de forma al-tamente empírica, ya que se justificaba por el clásico afo-rismo de Hipócrates, quien señalaba que «los melancólicosfrecuentemente se hacen epilépticos y los epilépticos me-lancólicos», abrió el campo del empleo de antiepilépticosen el tratamiento de los pacientes bipolares. Sin embargo,con estos tres agentes farmacológicos: litio, carbamazepinay valproato, a pesar de los progresos conseguidos, quedabanproblemas por resolver: muchos pacientes eran refractariosal tratamiento convencional y otros manifestaban proble-mas de tolerabilidad debido a los frecuentes efectos ad-versos de estos fármacos. Asimismo, estos agentes son máseficaces corrigiendo la fase maníaca que la depresiva y loscuadros de ciclación rápida y de manía mixta (disfórica)respondían mal al tratamiento (Dadennes et al., 1995;Keck y McElroy, 1996; Macritchie et al., 2001). Estos défi-cits motivaron la investigación de nuevos agentes en estetrastorno. Así pues, desde 1993 con la introducción clíni-ca de los nuevos antiepilépticos y antipsicóticos el trata-miento del trastorno bipolar ha experimentado un im-portante impulso (Wang et al., 2003; López-Muñoz et al.,2006).
Aunque la carbamazepina ya se venía utilizando en Euro-pa como sustituto del litio o en combinación en casos resis-tentes, el gran auge de la utilización de estos fármacos en elcampo del trastorno bipolar vino con la demostración de laeficacia antimaníaca del valproato, en su forma bivalente(divalproato) (Pope et al., 1991), y su comercialización enEstados Unidos como antimaníaco a mediados de la década de1990. Ello estimuló la evaluación de los antiepilépticos de tercera generación en el tratamiento del trastorno bipo-lar, máxime cuando se había demostrado en el terreno de laepilepsia que estaban dotados de un mejor perfil de efectossecundarios, interacciones, toxicidad y teratogenia. De entreellos, únicamente la lamotrigina, un antiepiléptico derivadode la feniltriazina, ha sido autorizada en este trastorno psi-quiátrico, fundamentalmente para la prevención de los epi-sodios depresivos en pacientes con trastorno bipolar (Bow-den et al., 2003; Calabrese et al., 2003).
El hilo conductor que utilizaremos en este trabajo pararealizar un acercamiento al mecanismo de acción de los an-tiepilépticos como eutimizantes son las sales de litio, queson consideradas, a nivel de investigación, como el patrónde oro o patrón estándar de los eutimizantes. El litio es laprimera sustancia de la que se demostraron propiedadesnormotímicas, con la que más experiencia se tiene y de laque mayor evidencia contrastada se posee sobre su eficaciaen la profilaxis de las recaídas del trastorno bipolar, siendo,además, la más conocida y estudiada desde la perspectiva desu mecanismo de acción eutímico. Las publicaciones sobre elmecanismo de acción de este fármaco se han ido incremen-tando, sobre todo en la última década, hecho particular-mente unido a los avances que se han producido en el ma-yor conocimiento de la etiopatogenia del trastorno bipolar(López-Muñoz et al., 2006).
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
4 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 4
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight

MECANISMOS DE ACCIÓN DE LOS FÁRMACOSANTIEPILÉPTICOS COMO AGENTES EUTIMIZANTES
Desde la perspectiva histórica, las primeras hipótesis so-bre el mecanismo de acción de los agentes eutimizantes sefundamentaron en una posible modificación del transporteiónico (de sodio, básicamente) y en la facultad de alterar laspropiedades electrofisiológicas de las membranas neurona-les, dado que el único agente terapéutico eficaz disponibleera el litio, esto es, un catión. Posteriormente con el descu-brimiento de los mecanismos de neurotransmisión en el sis-tema nervioso central (SNC), ampliamente distribuidos a nivel del sistema límbico, núcleo estriado y circuitos corti-coprefrontales, relacionados con las manifestaciones pro-pias de la enfermedad maniacodepresiva, se concedió mayortrascendencia a diversos aspectos relativos a estos mecanis-mos: su influencia sobre el transporte de colina y aminoáci-dos o bien su capacidad de alterar la transmisión monoami-nérgica, colinérgica o gabérgica (Alamo et al., 1997; Cuencaet al., 2000; Chen y Manji, 2006). En esta línea se centraronlos primeros estudios que analizaron los efectos eutimizan-tes de los fármacos antiepilépticos, fundamentalmente en lamodulación de la neurotransmisión gabérgica y glutamatér-gica.
Sin embargo, la mayor parte de las líneas de investigación enlos últimos años van encaminadas a comprender los mecanis-mos moleculares relacionados con la transducción de señales enlas neuronas postsinápticas. El sistema de transmisión neuronal
y la transducción de señales es una complicada cascada, alta-mente regulada, de acontecimientos bioquímicos en el que con-vergen múltiples sistemas de neurotransmisión, como el meta-bolismo de los fosfoinositoles o la vía de la adenilatociclasa (AC).De esta forma, se ha profundizado en el estudio de la influenciade los fármacos eutimizantes sobre las distintas fracciones delas proteínas G (PG) (Waldmeier, 1990), lo que promueve la acti-vación de factores de transcripción génica a nivel nuclear (Ikno-mov y Manji, 1999; Silva, 2001; Quiroz et al., 2004; Tamayo etal., 2004; Chen y Manji, 2006), así como la síntesis de factoresde crecimiento neuronal, abriendo las puertas a un posible pa-pel neuroprotector, que en el caso del litio ya fue apuntado porD’Mello et al. (1994) y Volonte et al. (1994) con sus estudios encultivos celulares (Wada et al., 2005).
No obstante, a pesar de los avances que han tenido lugaren los últimos años, el conocimiento del mecanismo de ac-ción eutimizante de los agentes anticonvulsivantes viene tro-pezando con varios problemas, como la inexistencia de unmodelo animal homologado de esta patología o el desconoci-miento y la complejidad de la fisiopatología del trastorno bi-polar, en el que coparticipan numerosas vías de neurotrans-misión que parecen estar disreguladas (Smith, 1990; Manji etal., 1995; Lenox et al., 1998). De hecho, a título de ejemplo, lacarbamazepina, un derivado iminodibencílico con una estruc-tura química muy parecida a la de los antidepresivos tricícli-cos, exhibe una serie de acciones comunes al litio que podríanestar implicadas en su efecto eutimizante, pero a la vez mues-tra acciones contrarias a las producidas por el ion (tabla 1), lo
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
5Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
Parámetro Litio Carbamazepina Valpropato
Respuesta de prolactina a triptófano Aumentada Aumentada —Funcionalismo serotoninérgico Aumentado Aumentado AumentadoRespuesta de receptores 5-HT2 Disminuida Aumentada —-Función receptor GABAB Aumentada Aumentada AumentadaRecaptación de noradrenalina Aumentada Disminuida AumentadaRespuesta c-AMP a noradrenalina Disminuida Disminuida DisminuidaRespuesta c-AMP a dopamina Disminuida Disminuida DisminuidaProducción de c-GMP Disminuida Disminuida —Niveles de tiroxina Disminuidos Disminuidos —Secreción de TSH Aumentada Disminuida —Acciones sustancia P Aumentadas Aumentadas —Flujo de calcio Aumentado Disminuido —Actividad proteínas G Disminuida Disminuida —Niveles IP3 Aumentados Disminuidos DisminuidosActividad inositol-fosfatasa Inhibida Aumentada No afectadaActividad PKC Disminuida Disminuida DisminuidaActividad MARCK Disminuida — DisminuidaActividad GSK3β Disminuida — DisminuidaNiveles CREB Aumentados Disminuidos —Actividad AP-1 Aumentada — Aumentada
Tabla 1 Modificaciones bioquímicas inducidas por litio, carbamazepina y valproato
3-21.qxd 10/4/08 12:11 Página 5
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Rectangle
Kiyor
Rectangle
Kiyor
Rectangle
Kiyor
Rectangle
Kiyor
Rectangle
Kiyor
Rectangle
que hablaría a favor de mecanismos de acción diferenciadosentre ambos eutimizantes. Es más, existen agentes anticon-vulsivantes introducidos en clínica cuyo mecanismo de ac-ción antiepiléptico es aún completamente desconocido, como el caso del levetiracetam.
Pese a lo expuesto, a continuación citaremos algunos delos datos publicados en relación con el mecanismo de ac-ción de los fármacos antiepilépticos, haciendo hincapié enlas dianas directas demostradas en el caso de estos agentes,en las modificaciones observadas tras tratamiento crónico y en las acciones que comparten con el litio, que no sólo esel patrón de referencia de los reguladores del humor en laevaluación de la eficacia terapéutica, sino también el agen-te eutimizante que dispone de más estudios específicos re-lativos al conocimiento de su mecanismo íntimo de acción.Sin embargo, a pesar de los avances producidos en los últi-mos años, el mecanismo de acción antimaníaca de los fár-macos anticonvulsivantes no está aún bien determinado yno existen pruebas de si coincide o no con su mecanismo deacción antiepiléptico. De hecho, los antiepilépticos, como seha comentado, afectan de forma diferente a algunos aspec-tos bioquímicos implicados en el hipotético mecanismo an-timaníaco del litio (tabla 1).
Efectos sinápticos implicados en el mecanismo de acción eutimizante de los fármacos antiepilépticos
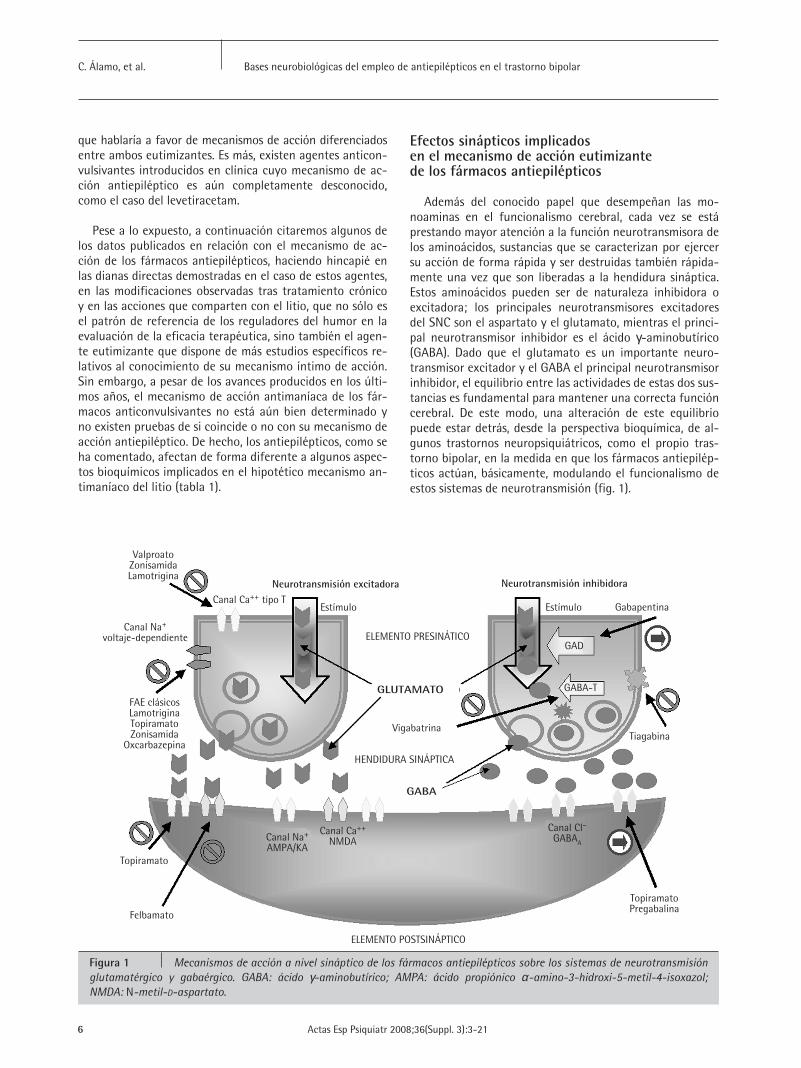
Además del conocido papel que desempeñan las mo-noaminas en el funcionalismo cerebral, cada vez se estáprestando mayor atención a la función neurotransmisora delos aminoácidos, sustancias que se caracterizan por ejercersu acción de forma rápida y ser destruidas también rápida-mente una vez que son liberadas a la hendidura sináptica.Estos aminoácidos pueden ser de naturaleza inhibidora oexcitadora; los principales neurotransmisores excitadoresdel SNC son el aspartato y el glutamato, mientras el princi-pal neurotransmisor inhibidor es el ácido γ-aminobutírico(GABA). Dado que el glutamato es un importante neuro-transmisor excitador y el GABA el principal neurotransmisorinhibidor, el equilibrio entre las actividades de estas dos sus-tancias es fundamental para mantener una correcta funcióncerebral. De este modo, una alteración de este equilibriopuede estar detrás, desde la perspectiva bioquímica, de al-gunos trastornos neuropsiquiátricos, como el propio tras-torno bipolar, en la medida en que los fármacos antiepilép-ticos actúan, básicamente, modulando el funcionalismo deestos sistemas de neurotransmisión (fig. 1).
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
6 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
Neurotransmisión excitadora Neurotransmisión inhibidora
ValproatoZonisamidaLamotrigina
Canal Ca++ tipo TEstímulo
Canal Na+
voltaje-dependiente
FAE clásicosLamotriginaTopiramatoZonisamida
Oxcarbazepina
ELEMENTO PRESINÁTICO
GLUTAMATO
HENDIDURA SINÁPTICA
GABA
Vigabatrina
Estímulo Gabapentina
Tiagabina
GAD
GABA-T
TopiramatoPregabalina
Topiramato
Felbamato
Canal Na+
AMPA/KA
Canal Ca++
NMDACanal Cl–GABAA
ELEMENTO POSTSINÁPTICO
Figura 1 Mecanismos de acción a nivel sináptico de los fármacos antiepilépticos sobre los sistemas de neurotransmisiónglutamatérgico y gabaérgico. GABA: ácido γ-aminobutírico; AMPA: ácido propiónico α-amino-3-hidroxi-5-metil-4-isoxazol; NMDA: N-metil-D-aspartato.
3-21.qxd 10/4/08 12:11 Página 6

Además, algunos agentes antiepilépticos también estándotados de actividad sobre el funcionalismo aminérgico, loque puede suponer un cierto papel en su mecanismo de ac-ción eutimizante. Tal es el caso de la lamotrigina, capaz debloquear los receptores postsinápticos serotoninérgicos 5-hi-droxitriptamina-3 (5-HT3) (Ketter et al., 1997). En este senti-do, la lamotrigina parece tener también un papel reguladorde la recaptación de monoaminas, básicamente de serotoni-na y sólo a concentraciones elevadas, que podría llegar atraducirse clínicamente en un incremento de la neurotransmi-sión serotoninérgica. Esta modulación de la neurotransmisiónserotoninérgica podría contribuir también a explicar la uti-lidad de la lamotrigina en la estabilización de los episodiosmaníacos y/o mixtos (incluyendo cicladores rápidos), así co-mo su eficacia en el tratamiento de la depresión bipolar yunipolar (Southam et al., 1998). Otras teorías proponen quelamotrigina puede potenciar el funcionalismo dopaminérgi-co, así como también ocasionar una regulación a la baja(down regulation) de la densidad de los receptores β-adre-nérgicos, una propiedad típica de los fármacos antidepresi-vos (Southam et al., 1998).
Con respecto a la carbamazepina, también se han descri-to una serie de efectos sobre los sistemas de neurotransmi-sión aminérgicos, como una disminución del recambio dedopamina, una disminución de la liberación de noradrenali-na o un incremento en los niveles sinápticos de serotonina(Post et al., 1992). Asimismo, el valproato ocasiona un aumento de la disponibilidad de noradrenalina en la hendidurasináptica por un aumento de la liberación de esta catecola-mina y un incremento de la neurotransmisión serotoninérgicamerced a la hiposensibilización de los receptores 5-HT1A(Shiah et al., 1997). En el caso concreto de este último fár-maco, se ha observado que su administración se asocia conuna disminución de las concentraciones de factor liberadorde corticotrofina (CRF) en la amígdala y el hipocampo, me-canismo que puede estar relacionado también con su efectoeutimizante (Tamayo et al., 2004).
Por otro lado, en las últimas décadas se ha concedidogran importancia al papel de los canales iónicos presentesen las membranas neuronales en la etiopatogenia de diver-sas patologías psiquiátricas. Algunos canales iónicos formanparte de un tipo de receptores, denominados ionotrópicos,que integran unos complejos proteicos, responsables de lageneración y mediación de señales de y entre membranascelulares excitables, como es el caso de las membranas delas neuronas. Suelen denominarse en función de su permea-bilidad y selectividad para un determinado ion (p. ej., canalde sodio o de cloro) y responden a cambios en el potencialde membrana, a ligandos extracelulares y a segundos men-sajeros. Son ejemplos de canales iónicos dependientes devoltaje los canales de calcio, potasio y sodio, y ejemplos de canales relacionados con ligandos extracelulares, el canal decalcio ligado al receptor glutamatérgico N-metil-D-asparta-to (NMDA) o el canal de cloro relacionado con el receptordel GABAA. Un ejemplo de canal ligado a un segundo men-sajero es el canal de calcio ligado a inositol-trifosfato. En
este sentido también los canales iónicos desempeñan un pa-pel importante en el mecanismo de acción de los fármacosantiepilépticos: de hecho existen agentes que inhiben los ca-nales de sodio dependientes de voltaje, fármacos que facilitanla acción del GABA sobre el canal de cloro y agentes que inhiben el efecto del glutamato sobre el receptor NMDA, eincluso fármacos que combinan algunas de estas propieda-des, como el topiramato (Corbella et al., 2000).
Disminución del funcionalismo del sistemade aminoácidos excitatorios
El glutamato, principal neurotransmisor excitador cere-bral, es un aminoácido que se encuentra a concentracioneselevadas dentro del tejido nervioso y se caracteriza por pro-ducir, incluso a dosis muy bajas, una excitación de la mem-brana neuronal en preparaciones in vitro. El glutamato seorigina a partir de una variedad de sustancias precursoras,incluidas la glucosa y el aminoácido glutamina, en las mito-condrias, organelas muy abundantes en las terminacionesnerviosas. La liberación del glutamato a partir de las neuro-nas parece ser un fenómeno dependiente del calcio, aunquein vitro también se ha demostrado su liberación en respues-ta a otros estímulos, tales como niveles elevados de potasio.Una vez liberado de la neurona presináptica, el glutamatoes rápidamente eliminado, principalmente mediante su cap-tación por los astrocitos, donde el glutamato se transformaotra vez en el aminoácido glutamina. Cuando existe una al-teración de la eliminación del glutamato, éste tiende a acu-mularse dentro del espacio sináptico y a concentracioneselevadas es neurotóxico. El glutamato actúa sobre dos tiposde receptores: unos receptores ionotrópicos ligados a cana-les, que incluyen al receptor NMDA, kainato y ácido propió-nico α-amino-3-hidroxi-5-metil-4-isoxazol (AMPA), y otrosreceptores metabotrópicos (mGlu), de los cuales existen almenos ocho subtipos clasificados en tres grupos.
El receptor NMDA es un complejo relacionado con el ca-nal sodio/calcio que presenta muchos loci moduladores: unlocus para el NMDA, otro para el cinc, dos loci situados en elcanal ionóforo, uno para el magnesio y otro para la fencicli-dina, una estructura para poliaminas y un locus para la gli-cina. Igual que otros receptores glutamatérgicos, como elreceptor AMPA y el receptor para el kainato, está formadopor cinco subunidades. El canal de calcio del receptor NMDAno se activa en la transmisión sináptica normal, ya que estábloqueado por iones de magnesio. El glutamato sólo activael canal de calcio del receptor NMDA cuando la neurona seha despolarizado parcialmente, desplazando a los iones demagnesio. Se ha dado una gran importancia a los receptoresNMDA en el desarrollo de fármacos antiepilépticos, ya queel aumento del tono glutamatérgico es convulsionante y sudisminución suele ser anticonvulsionante. La reducción deltono glutamatérgico en el receptor NMDA puede conseguir-se antagonizando competitivamente la acción del glutama-to sobre el receptor, antagonizando competitivamente el si-tio de la glicina y antagonizando de forma no competitiva
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
7Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 7
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight
Kiyor
Highlight

la apertura del canal. Sin embargo, algunas de estas estrate-gias no han tenido efecto anticonvulsionante en humanos olo han tenido, pero con un índice terapéutico inaceptable.
En la atrofia y muerte neuronal observada en el hipo-campo ante situaciones sostenidas de estrés, uno de los fac-tores etiopatogénicos más vinculados a los trastornos delhumor, también influye la acción de los aminoácidos excita-torios sobre el receptor NMDA. Es conocido, además, que elestrés continuado produce atrofia de dendritas apicales deneuronas piramidales CA3 (Duman, 2000; Manji et al., 2000).Este efecto parece secundario al incremento de aminoácidosexcitatorios en el hipocampo y es bloqueado por el antiepi-léptico fenitoína, que bloquea la liberación de glutamato ylos canales T del calcio, estimulados durante la liberacióndel citado aminoácido (McEwen y Magarinos, 1997). En estesentido parece ser que el estrés actuaría impidiendo la re-captación de glutamato más que facilitando o incremen-tando su liberación. Sea cual fuere la causa del incrementosináptico de glutamato, el bloqueo de receptores NMDApreviene la atrofia dendrítica producida por el estrés y po-dría asociarse a efectos terapéuticos positivos en pacientescon trastorno bipolar.
La importancia del receptor NMDA en el espectro bipolarviene dada, además, por la capacidad que exhiben diversosantidepresivos de modular este receptor (Skolnick et al.,1996). Por otra parte destaca que existen interrelacionesimportantes entre el receptor NMDA y el factor neurotrófi-co derivado del cerebro (BDNF; brain-derived neurotrophicfactor). Es de hecho conocido que esta neurotrofina ejerceun efecto protector neuronal y que, por el contrario, el re-ceptor NMDA, mediante la facilitación de la entrada de cal-cio en la célula, ejerce un efecto agresivo sobre la neurona.Los antidepresivos incrementan, vía CREB, los niveles deBDNF, y se ha podido demostrar que la exposición de célulasgranulares a BDNF reduce los niveles de proteína y delARNm del receptor NMDA-2A y NMDA-2C. Además, las mo-dificaciones receptoriales NMDA inducidas por el BDNF seacompañan de una disminución de la entrada de calcio enla neurona, de modo similar a como lo realizan los antago-nistas del receptor NMDA (Dildy y Leslie, 1989; Brandoli etal., 1998).
Otra vía explorada en relación con los aminoácidos exci-tatorios es la administración de agentes capaces de actuarsobre otros receptores de estos neurotransmisores. El canalde sodio del receptor glutamatérgico AMPA participa en latransmisión normal del estímulo excitador. Este canal no erauna diana importante para la búsqueda de antiepilépticos.Sin embargo, al descubrirse que las alteraciones en algunade sus subunidades permiten la entrada de calcio, el interéspor este canal ha aumentado notoriamente. De hecho, la la-motrigina, al contrario que el valproato y el litio, incremen-ta la expresión de las subunidades GluR1 y GluR2 del recep-tor AMPA, de forma dependiente de la dosis, en cultivos deneuronas de hipocampo (Du et al., 2007). Estos hechos, jun-to con la observación de que la imipramina y la fluoxetina
en tratamiento prolongado aumentan la expresión de GluR1del receptor AMPA, se han relacionado con las propiedadesantidepresivas de la lamotrigina y con las antimaníacas delvalproato en el marco del trastorno bipolar. Del mismo mo-do, ha cobrado interés el canal de sodio del receptor del kai-nato, muy similar al AMPA, y se han buscado fármacos quesean capaces de antagonizarlo. El topiramato es un ejemploen este sentido.
Tanto la carbamazepina como el valproato son agentesantiepilépticos capaces de disminuir el funcionalismo gluta-matérgico, bien actuando directamente sobre este tipo dereceptores (NMDA, AMPA y kainato) o aumentando el meta-bolismo de este neurotransmisor mediante la activación dela enzima glutamino-sintetasa a nivel de las células de glía(Li et al., 2002; Tamayo et al., 2004). En concreto, la carba-mazepina, a título de ejemplo, inhibe la liberación de gluta-mato inducida por veratridina (Olpe et al., 1985) y 4-amino-piridina (Cramer et al., 1994), disminuye, a concentracionesterapéuticas, la despolarización neuronal producida por laactivación del receptor NMDA (Lancaster y Davies, 1992) y re-duce las lesiones cerebrales perinatales inducidas por NMDAen la rata (Martin y Regan, 1988).
Del mismo modo, se han descrito posibles efectos positi-vos de algunos nuevos antiepilépticos, como lamotrigina,topiramato y gabapentina, en la modulación de este sistemade aminoácidos excitatorios. La lamotrigina inhibe la libera-ción de glutamato al bloquear los canales de sodio y calcio(Stefani et al., 1996; Wang et al., 1996) en la neurona presi-náptica, como después se comentará, pero también atenúala neurotoxicidad inducida por la inyección intraestriatal dekainato (McGeer y Zhu, 1990). Con respecto al topiramato,es conocido su papel antagonista de los receptores AMPA-kainato (Rosenfeld, 1997), pero también es capaz de dismi-nuir la liberación presináptica de glutamato (Hanaya et al.,1998), mientras que la gabapentina reduce la despolariza-ción de la membrana ocasionada por la administración exó-gena de glutamato (Calabresi et al., 1999).
Potenciación de la neurotransmisión gabérgica
El GABA es el principal neurotransmisor inhibidor delSNC. Precisamente, la unión del GABA a los receptores de lamembrana postsináptica genera un potencial inhibidor pos-tsináptico. En otras palabras, el GABA hace menos sensible ala activación a la neurona postsináptica. El GABA, que seencuentra en casi todos los órganos de la anatomía, se sin-tetiza en las neuronas por descarboxilación del glutamatogracias a la enzima descarboxilasa del ácido glutámico(DAG). La gabapentina, un fármaco antiepiléptico análogoestructural del GABA, actúa incrementando su síntesis alpotenciar la acción de esta enzima DAG, aumentando, se-cundariamente, la tasa de este neurotransmisor inhibidor enla hendidura sináptica (Goldlust et al., 1995). Un efecto pa-recido ha sido postulado también para el valproato (Loscher,1999).
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
8 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 8

La liberación del GABA, como la del glutamato, parece serdependiente de calcio. Además, al igual que el glutamato, elGABA es eliminado de la sinapsis mediante su captación porparte de los astrocitos. En el interior de éstos es transaminadopor la enzima GABA-transaminasa y luego oxidado para for-mar succinato. La inhibición de la GABA-transaminasa produ-ce una acumulación de GABA a nivel de la sinapsis, con la con-secuente inhibición de la actividad cerebral. Precisamente estesistema de inhibición de la GABA-transaminasa, conocido enel caso del valproato (Loscher, 1999), es uno de los mecanis-mos de acción investigados en el desarrollo de nuevos fárma-cos antiepilépticos, como es el caso de la vigabatrina. Así pues,en este sentido, el ácido valproico y su sal sódica (valproato)son agentes anticonvulsivantes cuyo mecanismo de acción,aunque parcialmente desconocido, parece basarse en el incre-mento de los niveles de GABA (Loscher, 1999), al igual que eltopiramato (Kuzniecky et al. 1998).
El receptor del GABA es una estructura bioquímica com-pleja, aunque muy bien caracterizada, en la que se encuen-tra también el receptor para las benzodiazepinas y barbitú-ricos, sustancias que se pueden considerar «imitadoras» y«potenciadoras» de los efectos inhibidores del GABA. Estecomplejo receptorial pertenece a una superfamilia de recep-tores ligados a canales iónicos de respuesta rápida. El re-ceptor GABAA está constituido por un complejo proteínicode cinco subunidades, alineadas simétricamente, formandoen su seno un canal que atraviesa la membrana neuronalpor el que penetra el cloro (Bormann, 2000). Mediante estu-dios de clonación se ha podido determinar la presencia demúltiples subunidades diferentes, denominadas α, β, γ, δ, ρ,que podrían dar lugar a más de 10.000 posibles combinacio-nes de receptores GABAA. En concreto, la estimulación delreceptor GABAA aumenta la entrada del ion cloro al interiorde la célula, lo que ocasiona una hiperpolarización de lamembrana neuronal (Alamo et al., 2000). El valproato po-tencia las acciones agonistas del receptor GABAA (Olpe etal., 1998) e incrementa la densidad de receptores GABAB enel hipocampo (Motohashi, 1992). En relación con los nuevosagentes antiepilépticos, como la gabapentina y la pregabali-na, al actuar potenciando mecanismos gabérgicos (funda-mentalmente potenciando la liberación de GABA) es asumi-ble su potencial eficacia en pacientes con trastorno bipolar.
El funcionalismo del sistema gabérgico también ha sidoexplorado en otros sentidos, como es la inhibición selectivade la recaptación de GABA (Lydiard, 2003). En este sentido,la tiagabina es el único antiepiléptico disponible en tera-péutica que actúa inhibiendo el transportador de membra-na GAT-1 del GABA. De esta forma, su eficacia antimaníacateórica se justificaría en el efecto antikindling secundario ala potenciación del funcionalismo gabérgico.
Movimientos iónicos implicados en el mecanismo de acción eutimizante de los antiepilépticos
El canal de sodio dependiente de voltaje es el responsablede generar los potenciales de acción en respuesta a la des-
polarización parcial de la membrana neuronal. Desde elpunto de vista bioquímico, este canal está formado por unasubunidad α, una subunidad β1 y una subunidad β2. La sub-unidad α es operativa por sí sola, mientras que las subuni-dades β1 y β2 parecen tener un papel modulador; la β1 esnecesaria para la inactivación del receptor y la β2 posee unpapel modulador menor (Errington et al., 2005). Este canales el lugar de acción de la mayor parte de los fármacos an-tiepilépticos clásicos, como la carbamazepina y el valproato,así como de algunos nuevos, como la lamotrigina, la oxcar-bazepina o el topiramato, que al inhibir este canal estabili-zan la membrana, ocasionando, de esta forma, una inhibi-ción de la liberación presináptica de neurotransmisoresexcitatorios (glutamato y aspartato) (Wang et al., 1996).Además, en el caso concreto de la lamotrigina y de la carba-mazepina se ha demostrado que este efecto sólo tiene lugarcuando el fármaco actúa sobre neuronas hiperexcitadas, ca-reciendo de efecto sobre aquellas otras que presentan unaactividad electrofisiológica normal (Ketter et al., 1997). Setrataría, pues, de una especie de efecto antikindling o «an-tiencendido», fenómeno caracterizado por la aparición de res-puestas conductuales o convulsivas tras la repetición de estí-mulos subumbrales continuados. Este mecanismo de acciónpodría explicar parte de la eficacia de lamotrigina en el ma-nejo del trastorno bipolar, aunque el mecanismo exacto delefecto eutimizante es aún desconocido (Farber et al., 2002).
Por su parte, la activación de los canales de calcio depen-dientes de voltaje se acompaña del aumento del calcio in-tracelular y la disminución del calcio extracelular. Este canalse ha asociado con la liberación de neurotransmisores, conla despolarización sostenida de la neurona, con fenómenosde potenciación prolongada, con los fenómenos de «autoen-cendido» (kindling) y con fenómenos de excitotoxicidad (Post,2002). La entrada de calcio en las neuronas a nivel presináp-tico facilita la liberación de neurotransmisores; a nivel pos-tsináptico produce la despolarización mantenida que se ob-serva en los cambios paroxísticos de despolarización de lascélulas que actúan como marcapasos. Se han descrito, almenos, cinco tipos de canales de calcio: L, N, T, P y Q, que sediferencian en su conductancia, duración de la corriente yvelocidad de inactivación. En concreto, tanto la lamotriginacomo el valproato se caracterizan por disminuir la entradaintraneuronal de calcio, actuando específicamente sobrecanales de tipo T (Post et al., 1992; Stefani et al., 1996; Ta-mayo et al., 2004). Un fenómeno común observado con di-ferentes reguladores del humor, como las sales de litio, lacarbamazepina, el valproato, el topiramato o la lamotrigina,además de otros fármacos, como los antagonistas del calcio,es la disminución de la entrada de calcio en la neurona, locual puede ser realizado a través de diferentes mecanismosque llevan todos a la disminución de la entrada del ion. Lacarbamazepina ejerce esta disminución de la entrada de cal-cio por su acción directa sobre el receptor NMDA.
Finalmente, el canal de potasio dependiente de voltajeparticipa en la repolarización e hiperpolarización de lamembrana neuronal y sus alteraciones pueden provocar un
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
9Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 9

estado de hiperexcitabilidad neuronal. En el caso concretode la carbamazepina se ha descrito un aumento de la con-ductancia del potasio, lo que se ha asociado a sus efectosanticonvulsivantes (Post et al., 1992).
Pese a que el litio utiliza en pequeña proporción la bom-ba Na+/K+ ATPasa dependiente, esta relación ha sido am-pliamente estudiada como mecanismo posible de acción delion. En este sentido se ha podido detectar una disminuciónde la actividad de la bomba Na+/K+ ATPasa eritrocitaria enpacientes bipolares (Hokin-Neaverson y Jefferson, 1989a),que se normaliza tras tratamiento crónico con litio (Hokin-Neaverson y Jefferson, 1989b; Dubovsky, 1995). Además,experimentalmente en un método de conducta motora, quepuede constituir un buen modelo de trastorno bipolar, se haobservado que la oubaína, inhibidor de esta bomba, disminuyela actividad motora en la rata, fenómeno que se pueden preve-nir con el pretratamiento continuado con litio (Li et al., 2002).
Los datos expuestos parecen sugerir la implicación de labomba Na+/K+ dependiente en la fisiopatología de la enfer-medad bipolar, así como en el mecanismo de acción del li-tio. De esta forma, el déficit de la citada bomba provocaría,por un lado, un aumento de la excitabilidad neuronal, quesería responsable del proceso maníaco y, por otro, una dis-minución de la liberación de neurotransmisores, que se co-rrespondería con la fase depresiva (El-Mallakh y Wyatt,1995). Sin embargo, no debemos olvidar que la hipótesis cita-
da se basa en un modelo periférico, el eritrocito, o en estudiosexperimentales en SNC de rata y por tanto la extrapolaciónde estos datos a los humanos puede ser especulativa (Lenoxet al., 1998). Esta cautela viene avalada por la observaciónde que la expresión genética de subunidades de esta bombano es idéntica en todos los tejidos, existiendo incluso dife-rencias regionales dentro del propio SNC (Arystarkhoua etal., 1996; Lecuona et al., 1996; Malick et al., 1996).
Eventos intraneuronales implicadosen el mecanismo de acción eutimizantede los fármacos antiepilépticos
La investigación bioquímica sobre los mecanismos respon-sables del efecto eutimizante de diferentes fármacos, funda-mentalmente del litio, se centró en la cascada de aconteci-mientos que participan en la transducción de las señales,desde los estudios a nivel de primeros mensajeros, como lainteracción neurotransmisor-receptor, hasta, más reciente-mente, los de mecanismos de transducción de la informaciónmás allá de los receptores, que involucran el acoplamiento delos receptores con los transductores de la señal (fig. 2). En estesentido, la familia de PG heterotriméricas es un punto crucialde convergencia en la transmisión de señales a partir de variosprimeros mensajeros hasta una serie de acontecimientos celu-lares en cascada: enzimas intracelulares efectoras como se-gundos mensajeros y canales iónicos.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
10 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
N1 N2
R R R R R
GiGs Gx
PIP2
DAGPLC
IP3
CA++
CA++
PKCaMPKCC-Jun
AMPc
CREBFOS
AC
ATP
FDEs
5-AMP
PKA
1.er mensajero
2.o mensajero
3.er mensajero
4.o mensajero
Acciones tróficasSupervivencia neuronalRemodelado sináptico
Figura 2 Mecanismos receptoriales y vías de transducción intraneuronales asociadas a los sistemas de la adenilatocicla-sa, los fosfoinositoles y mediadas por calcio relacionadas con el mecanismo de acción de los fármacos eutimizantes.
3-21.qxd 10/4/08 12:11 Página 10

De todos los agentes reguladores del humor, el litio es,sin lugar a dudas, el fármaco más estudiado a este nivel ydel que más datos se dispone en relación a sus efectos bio-químicos intraneuronales. Por este motivo discutiremos losmecanismos compartidos, así como los diferenciales, del ioncon los fármacos anticonvulsivantes, fundamentalmente conla carbamazepina y el valproato, ya que los efectos postsi-nápticos de los nuevos antiepilépticos son prácticamentedesconocidos, salvo datos puntuales, como en el caso de lalamotrigina (Li et al., 2002).
Aunque aún se desconoce el mecanismo neuroquímicoexacto que subyace en los efectos terapéuticos eutimizantesdel litio y de los clásicos anticonvulsivantes, las teorías másrecientes postulan la alteración de la transducción de la se-ñal neuronal más allá de los receptores de membrana, loque explicaría el efecto regulador del humor a largo plazode estos fármacos, sobre todo en lo referente a la preven-ción de episodios maníacos y/o depresivos. En 1988 se infor-mó por primera vez que las PG participaban en el mecanis-mo bioquímico que subyace a la acción terapéutica del litio(Avissar et al., 1988). La inhibición por este ion de la funciónde los receptores β-adrenérgicos y muscarínicos acoplados alas PG se sugirió como un locus molecular para los efectosterapéuticos antimaníacos y antidepresivos del mismo(Avissar et al., 1988). Es importante resaltar que la interac-ción del litio con las PG es la más contrastada hipótesisacerca del mecanismo de su actividad terapéutica como es-tabilizador del estado de ánimo. En este sentido se postulaque el litio estabilizando las PGs actuaría como antimaníacoy por el contrario estabilizando las PGi se comportaría comoantidepresivo. Además, se ha constatado que otros trata-mientos empleados en el trastorno bipolar, como la carba-mazepina o la terapia electroconvulsiva, afectan también ala función de las PG (Schreiber y Avissar, 2003).
Por otra parte existen teorías basadas en los efectos del litiosobre los sistemas de transducción de la señal de segundosmensajeros, que comprenden la AC y las vías del fosfoinositol,que incluyen a las proteincinasas A y C (PKA y PKC, respectiva-mente) (Williams y Harwod, 2000). Asimismo, el litio inhibe deforma directa a la glucógeno-sintetasa-cinasa-3 (GSK3), unacinasa que actúa contracorriente y pertenece al grupo de lascinasas distales activadas por mitógenos (MAP-cinasas). Estacinasa, que funciona como intermediaria en numerosas víasintracelulares que participan en diferentes funciones del SNC,es la diana también de otros estabilizadores del humor, comoel valproato, y por ello el objetivo de investigación para nue-vos eutimizantes (Stambolic et al., 1996; Cuenca et al., 2000;Gould et al., 2002; Quiroz et al., 2004; Gould et al., 2003;Gould, 2006).
El papel de las proteínas G
La vía de transducción asociada a PG está regulada porun punto regulador proximal, que es el propio receptor quese acopla a la PG, y un punto regulador distal que se corres-
ponde con la actividad de la enzima glutamiltranspeptidasa(GTPasa). Recientemente se ha confirmado que ambos pun-tos reguladores pueden guardar relación con la fisiopatolo-gía de los trastornos del estado de ánimo y podrían ser dia-na del mecanismo de acción de los antidepresivos. Casitodos los receptores acoplados a proteínas G (GPCR) estánestrictamente regulados por un mecanismo desensibilizantecomún. Últimamente se ha podido establecer que la fosfori-lación del receptor por las cinasas de receptores acoplados aPG (GRK) es el paso inicial y crítico en el desacoplamientodel receptor de la PG, que conduce a la atenuación o desen-sibilización de la señalización GPCR. Tras la fosforilación porlas GRK, los GPCR se unen a una familia de proteínas solu-bles, denominadas arrestinas porque «arrestan» la señaliza-ción GPCR. Las arrestinas se unen a las regiones de GPCRque también son los determinantes principales para la inter-acción con la PG, desacoplando así el GPCR de las PG.
Cuando este tema se ha estudiado desde un enfoque ge-nómico funcional se han identificado una serie de genescandidatos involucrados en la patogenia de los trastornosdel estado de ánimo, entre los que destaca el que codifica lacinasa-3 del receptor acoplado a la PG (GRK3), que tambiénestá disminuido en líneas de células linfoblastoides proce-dentes de un subgrupo de pacientes con trastorno bipolar(Niculescu et al., 2000). Recientemente se ha encontradoque un polimorfismo de nucleótido único en la región pro-motora de la GRK3 se asocia con el trastorno bipolar (Ba-rrett et al., 2003).
Así pues, cada vez existen más evidencias que sugierenque las PG están involucradas en la patogenia de los tras-tornos del estado de ánimo, incluyendo el trastorno bipolar(Schreiber y Avissar, 2003). Estas evidencias se basan en va-rios puntos, como los efectos clínicos del litio y de otros fár-macos en pacientes con trastorno bipolar o las alteracionesen la densidad y/o la función de las PG en elementos desangre periférica, observándose un aumento de ambos pa-rámetros en los pacientes con trastorno bipolar. También lasalteraciones en los niveles de PG en los tejidos obtenidos deautopsias de pacientes con trastorno bipolar apoyan estarelación.
Por otro lado, las proteínas reguladoras de la señalizaciónde la PG (RGS) forman una familia de señalización multifun-cional. Un papel importante de estas proteínas es la modula-ción de la señalización a través de las vías de la PG, mediante laadherencia a las PGα que se unen a la GTP y acortan la du-ración de la señalización de la PG, actuando sobre todo co-mo proteínas activadoras de la GTPasa (GAP). Así, estas pro-teínas reducen la duración de la señalización intracelular demuchos GPCR pertenecientes a sistemas de neurotransmi-sión tan diversos como el dopaminérgico, el gabérgico o elglutamatérgico, por lo que también podrían participar en el mecanismo de acción eutimizante de aquellos agentesantiepilépticos que actúan sobre dichos sistemas de neuro-transmisión.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
11Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 11

Acciones de los antiepilépticos sobre el sistema de la adenilatociclasa
El mecanismo por el cual el litio inhibe in vitro la AC pa-rece ser directo, actuando sobre la unidad catalítica de laenzima, ya que inhibe la estimulación producida por dosagentes que actúan a través de mecanismos diferentes: unanálogo estable de la GTP, que activa la subunidad α, elguanil-imidodifosfato, así como por el complejo calcio-caldmodulina. Puesto que in vitro estos efectos son antago-nizados por el magnesio, podría decirse que el litio competi-ría directamente con aquel ion por la unión en un punto dela unidad catalítica de la AC (Mork y Geisler, 1989; Mork etal., 1992; Manji et al., 1995). La carbamazepina también escapaz de ejercer una inhibición directa de la AC (Chen et al.,1996).
La complejidad del mecanismo de acción de los eutimi-zantes a este nivel es manifiesta, ya que existen datos queindican que las acciones a largo plazo sobre la AC puedenser opuestas a las observadas tras su administración aguda.Colin et al. (1991) han demostrado que en tratamiento cró-nico el litio aumenta la expresión de algunas de las formasde la AC (tipos I y II) y sus correspondientes ARNm. Este in-cremento podría significar una sensibilización del sistemagenerador de adenosina-monofosfato cíclico (AMPc) y po-dría, dentro del terreno de las hipótesis, explicar el alto ries-go de recaídas maníacas tras la suspensión del tratamientocon litio (Manji et al., 1995).
El efecto global del litio sobre la producción de AMPc de-penderá de la preponderancia de los distintos subtipos dePG y AC en las diferentes zonas del SNC y de la capacidaddel ion de afectar a los mismos, a las concentraciones alcan-zadas en el SNC. De hecho, su efecto sobre la AC estimuladapor la hormona antidiurética o por la hormona liberadorade tirotropina es manifiesto, incluso clínicamente, pero suacción inhibidora sobre la acumulación de AMPc, inducidapor varios neurotransmisores y hormonas en el SNC, pareceproducirse a concentraciones más elevadas que las necesa-rias para afectar al sistema de fosfoinositol (Lenox et al.,1998). En este sentido, se ha podido constatar que la admi-nistración crónica de litio disminuye la actividad de AMPcpor dos vías diferentes: disminuyendo la actividad de la PGsy aumentando la concentración de PGi (Ebstein et al., 1976;Post et al., 1992; Nestler et al., 1995; Belmaker, 2004). Lacarbamazepina también disminuye la actividad del AMPc,tanto basal como estimulada (Lewin y Bleck, 1977). Sin em-bargo, la administración crónica de carbamazepina (21-28 días) disminuye la actividad de la PGi y la PGs (Lesch etal., 1991; Post et al., 1992), pero incrementa los niveles deAMPc específicamente en la corteza frontal, lo que indicatambién una variabilidad regional en los efectos de este fár-maco (Wiborg et al., 1997).
Resulta interesante destacar también que la carbamaze-pina, al igual que el litio, es capaz de inhibir la actividad dela AC basal y, en mayor medida, la estimulada por forskolina
a través de un mecanismo de acción directo sobre la AC osobre factores íntimamente ligados a la misma, sin afectar alas PG (Chen et al., 1996). Estos datos sugieren la coopera-ción de la inhibición de la AC en el mecanismo de acción delos reguladores del humor.
Finalmente debe comentarse que la carbamazepina, alcontrario que el litio y el valproato, inhibe la actividad delfactor de transcripción CREB (Manji et al., 1996) y disminu-ye la expresión de c-fos inducida por forskolina (Divish etal., 1991), lo que resalta la importancia de la vía de señaliza-ción del AMPc como diana de acción de la carbamazepina.Sin embargo, tampoco hay que descartar su papel a travésde la vía de los fosfoinositoles, en tanto que en cultivos deastrocitos ocasiona una regulación al alza (up regulation) de los receptores de adenosina tipo 1 (Biber et al., 1999).
Hipótesis de la depleción del mioinositol (inositol)
Entre las múltiples hipótesis propuestas para explicar elmecanismo de acción del litio, la que implica los niveles demioinositol (inositol isómero de la glucosa presente de for-ma ubicua en el interior celular) es una de las más popularesy se basa en la capacidad del litio para inhibir de forma di-recta y a concentraciones compatibles con dosis terapéuticas(0,6-1,2 nM) (Layden et al., 2000) algunas de las enzimas fos-fomonoesterasas implicadas en el ciclo del fosfoinositol(Manji et al., 1999: Cuenca et al., 2000; Quiroz et al., 2004;).Esta acción también ha sido descrita para el valproato (O’Donnell et al., 2000), lo que permite apuntalar el interésde esta hipótesis.
La inositol-monofosfato-fosfatasa (IMPasa) y la inositol-polifosfato-1-fosfatasa (IPPasa) son enzimas implicadas enla síntesis de novo y en el reciclaje del inositol, componentenecesario en la vía de transducción del fosfoinositol. La bi-fosfato-nucleotidasa (BPNasa) hidroliza al inositol-1-4-bifosfato, de forma similar a como lo realiza la IPPasa, por loque se postula un papel similar en el mecanismo de accióndel ion. Por otra parte, esta enzima, denominada tambiénPAP fosfatasa, actúa sobre nucleótidos bifosforilados, comoel 3-fosfoadenosina-5-fosfato (PAP), produciendo adenosi-na-5-fosfato (AMP). La inhibición de esta enzima por el litioparece inhibir diversas sulfotransferasas que regulan la ho-meostasis del sodio, por lo que parece estar implicada en ladiabetes insípida observada con el tratamiento crónico conlitio (Spiegelberg et al., 1999; Cuenca et al., 2000; Agam etal., 2003; Quiroz et al., 2004). Todas estas enzimas tienen encomún una pequeña cadena de aminoácidos y una estruc-tura terciaria que capta iones metálicos y participa en lafunción catalítica de la enzima (Alamo et al., 1997; Cuencaet al., 2000; Quiroz et al., 2004; Berry et al., 2004).
La vía de los fosfoinositoles está relacionada con una se-rie de receptores de membrana acoplados a la proteína Gq11,como el 5-HT2, α1 y los muscarínicos M1, M3 y M5, así comolas vías relacionadas con las neurotrofinas, y con los recep-
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
12 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 12

tores de tirosincinasas (TkR), cuya estimulación provoca laactivación de la fosfolipasa C (PLC). Esta enzima media en lahidrólisis del fosfolípido de membrana, fosfoinositol 4,5 bi-fosfato (PIP2) para formar los segundos mensajeros: diacil-glicerol y el inositol trifosfato (IP3), que son los encargadosde modular una amplia gama de eventos intracelulares. ElIP3 debe ser desfosforilado, lo que hacen las IPPasa, paraformar inositol, precursor indispensable para la síntesis y re-ciclaje del PIP2 de las membranas, lo que mantiene activo elciclo de fosfoinositoles, ya que el inositol no se encuentra li-bre a nivel celular. El paso limitante en este reciclaje es laIMPasa, que convierte el IP3 en inositol (Cuenca et al., 2000;Quiroz et al., 2004). En relación con la carbamazepina, se hadescrito que es capaz de disminuir los niveles de IP3 (Li et al.,2002), lo que podría tener cierta relevancia en relación consu posible efecto antimaníaco.
El litio, a concentraciones terapéuticas (Hallcher y Sherman,1980), inhibe de forma directa y no competitiva la IMPasa(Parthasarathy et al., 1993) y secundariamente la IPPasa,provocando una depleción de mioinositol (Berridge et al.,1982; Silva, 2001) y un acúmulo de diacilglicerol en el SNC(Allison y Stewart, 1971). En consecuencia, ante la carenciade inositol tiene lugar una disminución de la formación dePIP2, lo que impide que se active la cascada de efectos se-cundarios a la estimulación de los citados receptores aco-plados a la PGq11 (Quiroz et al., 2004). Sin embargo, al con-trario que el litio, la carbamazepina parece incrementar laactividad de la IMPasa (Post et al., 1992), lo que hace pensaren otras vías paralelas en relación a su mecanismo de acción.
El tejido cerebral es especialmente sensible al litio por laincapacidad de las neuronas de captar inositol al encontrar-se protegidas por la barrera hematoencefálica. El hecho deque la inhibición de la IMPasa por el litio sea de tipo nocompetitivo sugiere que el ion se une al complejo enzima-sustrato y que un incremento del sustrato no sólo no superala inhibición, sino que la incrementa. Este hecho explica queel efecto del litio sea más importante en los sistemas con al-to recambio de fosfoinositol y prácticamente neutro en losnormofuncionantes (Nashorski et al., 1991). Por tanto, se-gún esta hipótesis, la acción del litio estaría mediada por lacarencia de inositol libre y su selectividad se debería a su ac-ción preferencial en las vías neuronales más activas.
Mecanismos de transducción relacionados con el ciclo de los fosfoinositoles
La hipótesis de la depleción del inositol no explica por sísola las propiedades estabilizadoras del humor del litio, porlo que la depleción de inositol se interpreta como un efectoinicial que pone en marcha una cascada de señales que es-tarían en relación con los efectos terapéuticos retardadosdel ion (Manji et al., 1996; Silva, 2001).
La activación de las vías de los fosfoinositoles desempeñaun papel fundamental en la transducción de señales secun-
darias al estímulo de diversos GPCR. Así, mediante la PGq11se activa la PLC, que se encarga de la hidrólisis del PIP2 demembrana, produciéndose los segundos mensajeros IP3 ydiacilglicerol (Cuenca et al., 2000; Gould y Manji, 2002; Qui-roz et al., 2004). El IP3, el primero de los segundos mensajerosde la cascada de los fosfoinositoles, se une a sus receptores in-tracelulares, especialmente del retículo endoplasmático, li-berando el calcio intracelular, que se acopla a la caldmodu-lina (CaM), estimulando a la proteincinasa dependiente decaldmodulina (PKCaM), responsable de regular la actividadde proteínas presentes en canales iónicos, proteínas estruc-turales, así como partícipe en la regulación de la apoptosis yde diversos factores de transcripción (Soderling, 2000; Qui-roz et al., 2004). Si se produce la depleción del inositol porel litio, al inhibir de forma directa a la IMPasa y la IPPasa, nose produce el recambio del fosfoinositol y, por tanto, se fre-na toda esta cascada de efectos de transducción (Quiroz etal., 2004).
El incremento inicial en los niveles de IP3 por el litio a concentraciones terapéuticas es debido también a un aumento en la liberación de glutamato por inhibición de laproteína recaptadora del mismo, lo cual permite un incre-mento en los niveles de calcio intraneuronal y secundaria-mente la activación de la PKC y la elevación de los nivelesde IP3. Antagonistas glutamatérgicos que bloquean la per-meabilidad de los canales asociados al receptor NMDA, co-mo dizocilpina, ketamina y magnesio, inhiben en forma se-lectiva la acumulación de IP3 a pesar de la administraciónde litio (Dixon et al., 1994). Este efecto glutamatérgico ini-cial podría ser de relevancia en los efectos antidepresivosdel litio, pero surge la pregunta de cómo puede esta acciónser conciliada con el control de los episodios maníacos y suprofilaxis. Dixon y Hokin (1997) plantean que la elevacióncrónica del IP3 puede hiporregular (down regulation) los re-ceptores de este compuesto ubicados a nivel de la membra-na del retículo endoplasmático, disminuyendo la liberaciónintracelular de calcio y, por tanto, la actividad de la PKC. Es-tos mismos autores probaron posteriormente que la admi-nistración de litio en ratas por espacio de 1 a 2 semanasproducía una hiperregulación (up regulation) de las proteí-nas recaptadoras de glutamato. Este efecto modulador ade-más parece ser dependiente de los niveles plasmáticos delcatión y ofrecería, según los autores, una explicación al me-canismo eutimizante del litio.
La hipótesis de la depleción de inositol por el litio tienesus puntos de apoyo, pero también algunas contradicciones.Entre los primeros, basados en diversos estudios, tanto encultivos celulares como en modelos animales, nos encontra-mos con la disminución de niveles de inositol en cerebro deroedores y de mioinositol en humanos bajo tratamiento conlitio. Asimismo, dosis elevadas de inositol pueden inhibir laconducta motora inducida por el litio en la rata y las con-vulsiones inducidas por el litio asociado a pilocarpina (Bel-maker et al., 1996). Además, algunos estudios sugieren laatractiva posibilidad de que otros eutimizantes, en especialel valproato y la carbamazepina, regulen también la cascada
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
13Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 13

de señales del ciclo del fosfoinositol. En este sentido se sabeque estos eutimizantes disminuyen la expresión y la funcióndel ARNm del SMIT1 (transportador de alta afinidad delmiosinositol), que en la actualidad constituye una nuevadiana para el estudio de potenciales reguladores del humor(van Calker et al., 2000; Quiroz et al., 2004). Asimismo, encultivos celulares específicos para valorar la estabilidad deconos de crecimiento neuronal se ha podido comprobar quelos tres eutimizantes comentados inhiben el colapso de es-tos conos y además aumentan la superficie del área de cre-cimiento de los mismos, siendo estos efectos revertidos porel inositol (Williams et al., 2002). Esta hipótesis explicaría laspropiedades estabilizadoras de los reguladores del humor enpacientes maníacos en los que el nivel de mioinositol es su-perior al observado en los controles (Cecil et al., 2002; Da-vanzo et al., 2003).
Sin embargo, esta teoría tiene algunas matizaciones. Enestudios iniciales se señalaba que el litio a niveles plasmáti-cos terapéuticos provocaba en animales de laboratorio unadisminución en la concentración del PIP2, el IP3 y el diacil-glicerol. Sin embargo, estudios más recientes han demostra-do que este efecto inhibidor del litio se manifiesta, en parti-cular, en presencia de altas concentraciones de agonistascolinérgicos y, por tanto, en un sistema hiperfuncionante.Sin embargo, en condiciones basales se observa un efectoopuesto, permitiendo la acumulación de IP3. Del mismo mo-do parece discrepante la observación de que algunos efec-tos crónicos del mioinositol sean similares a los observadoscon litio (Manji et al., 1996). Por otra parte, ratones caren-tes (knockout) del transportador de inositol (SMIT1) y portanto con niveles bajos de inositol intracelular presentan ni-veles de PIP2 normales (Berry et al., 2004). Además, pareceparadójico que los niveles de inositol se encuentren reduci-dos, no incrementados, en la corteza frontal de pacientesbipolares post mortem (Moore et al., 1997). Asimismo, la hi-pótesis de la depleción del inositol no ha podido probarse enhumanos por la ausencia de inhibidores específicos de laIMPasa que penetren a través de la barrera hematoencefáli-ca y que actúen a nivel del SNC (Quiroz et al., 2004). De he-cho, estudios recientes de neuroimagen por resonanciamagnética, en pacientes con depresión bipolar, ponen demanifiesto que el tratamiento prolongado con litio no mo-difica los niveles de mioinositol debido, probablemente, auna regulación al alza de la IMPasa (Patel et al., 2006).
El diacilglicerol es el otro de los dos segundos mensajerosimplicados en el ciclo de los fosfoinositoles. La importanciade este mensajero viene dada por la capacidad que tiene deactuar sobre su diana primaria, la PKC, la cual ha sido obje-to de múltiples investigaciones sobre su papel en el trastor-no bipolar y en el mecanismo de acción del litio y otros eutimizantes. La PKC es una enzima ampliamente distribui-da en el cerebro, donde tiene un papel fundamental por sucapacidad de fosforilar numerosos sustratos proteicos. Sibien su papel fisiológico en la regulación del humor no estádilucidado, se sabe que la actividad de la PKC está aumenta-da de forma significativa en pacientes maníacos en relación
a sus niveles en sujetos sanos, pacientes con depresión ma-yor o en pacientes esquizofrénicos. Además, tanto el valpro-ato como el litio reducen su actividad a través de diferentesmecanismos, lo que deja entrever un papel importante de laPKC en la patología bipolar (Cuenca et al., 2000; Sela, 2004).
La proteína MARCKS es un sustrato primario de la PKC.La MARKCS se une a la caldmodulina, fenómeno dependien-te del calcio, y enlaza la actina con la membrana plasmática,participando en los fenómenos de reestructuración del ci-toesqueleto (Watson y Lenox, 1996). La administración cróni-ca de litio a concentraciones compatibles con las terapéuticasdisminuye la fosforilización de la MARCKS, mediada por laPKC, y es capaz de hiporregularla en el hipocampo (Lenox etal., 1996). Este efecto puede ser prevenido y revertido conaltas concentraciones de inositol, por lo que se postula quees un fenómeno secundario a la inhibición de la IMPasa y dela IPPasa por el ion (Watson y Lenox, 1996). Por tanto, la hi-porregulación de la MARCKS inducida por el litio influye enprocesos de tanta trascendencia como la reestructuracióndel citoesqueleto, que puede intervenir en los cambiosadaptativos relacionados con la transducción de la señal y laliberación de neurotransmisores (Lenox et al., 1996). La apli-cación de valproato a cultivos celulares también ocasionauna regulación a la baja de los niveles de MARCKS, e inclusode forma más rápida que con el litio, aunque parece que losmecanismos bioquímicos responsables de este efecto son di-ferentes, ya que el valproato no actúa a través de la vía delPIP2 (Watson et al., 1998).
La capacidad de modulación de la actividad enzimáticade la PKC es un efecto farmacológico descrito con diversosagentes terapéuticos, incluidos la mayor parte de los fárma-cos eutimizantes, que de esta forma pueden regular la acti-vidad de genes de respuesta temprana y de diversos factoresde transcripción, como la proteína activadora 1 (AP-1). Tan-to el litio como el valproato regulan la unión de la AP-1 alADN dependiendo del tiempo de administración y de lasconcentraciones cerebrales alcanzadas (Yuan et al., 1998).Un estudio específico realizado en cultivo de células de neu-roblastoma humano puso de manifiesto que el prototipo deeutimizante, el litio, era capaz de reducir un 40% la activi-dad de la AP-1 estimulada por carbacol, cifras superadaspor la carbamazepina, que la redujo en un 46% (Pacheco yJope, 1999). La carbamazepina también ha confirmado unasupresión de la expresión de proteínas Fos inducidas pormetanfetamina en un modelo experimental de manía (Leeet al., 2000).
El valproato es el anticonvulsivante más estudiado en re-lación con su capacidad de regulación de la expresión géni-ca. Así, se ha confirmado que incrementa la expresión de losgenes de respuesta inmediata c-jun y c-fos (Asghari et al.,1998), y en tratamiento crónico reduce la activación deotros factores de transcripción de genes de respuesta rápi-da, como el Egr-1, también ligado como la AP-1, a la vía dela PKC (Grimes y Jope, 1999). Wang et al. (1999) han puestode manifiesto que tanto el valproato como la carbamazepi-
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
14 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 14

na, pero no el litio, incrementan selectivamente la expresiónde una proteína de la familia de las chaperonas, denomi-nada proteína reguladora de la glucosa (GRP78).
Acción de los anticonvulsivantes sobre las vías de transducción controladas por la glucógeno-sintetasa-cinasa 3
La GSK3 es una serina-treonina-cinasa de amplia distri-bución celular (citoplasma, mitocondrias, núcleo). Parecenexistir dos subtipos de GSK3, una forma α y otra forma β(51 y 47 kDa, respectivamente), con funciones similares,aunque no idénticas. La GSK3 está, en condiciones norma-les, fuertemente activada y es desactivada por múltiples víasde transducción. Entre éstas destacan, por ser las mejor es-tudiadas, la vía de la insulina/IGF1 y la de las neurotrofinas,ambas utilizando la fosfoinositol-3-cinasa (PI3K), así comola vía de la Wnt, aunque sobre ella pueden actuar otros mu-chos inhibidores como la PKC, la PKA y la p90RSK (cinasa ri-bosomial S6). La desactivación de la GSK3 por cualquiera deestas vías influye a su vez sobre una gran variedad de siste-mas efectores controlados por esta enzima (fig. 3). Es porello que la acción de la GSK3 a nivel celular es muy ubicua y
depende de la vía que actúe sobre ella, así como de su loca-lización celular y regional (Manji et al., 1999; Quiroz et al.,2004; Chen y Manji, 2006). Además de los comentados,otros sistemas efectores que dependen de la inactivación dela GSK3 incluyen diversos factores de transcripción (C/EBPβHSF-1, c-Jun, Myc, NFATc, C/EBPα, CREB, MITF), proteínasligadas a los microtúbulos (APC, MAP1B, MAP2, humandynamin-like protein, cadena ligera de cinesina, proteínatau), mediadores del ciclo celular (ciclina D, nineína huma-na), proteínas estructurales (axina, presenilina-1), proteínasimplicadas en la adhesión celular (β-catenina, DF3, MUC1,proteína neural de adhesión celular), así como elementos deregulación del metabolismo (eIF2b, glucógeno-sintetasa,ATP-citrato-liasa, cinasa dependiente de AMPc, piruvato-deshidrogenasa) (Gould et al., 2004; Quiroz et al., 2004).
Los trastornos del humor están muy relacionados con ladisfunción de mecanismos serotoninérgicos. En este sentido,estudios de neuroimagen, así como las respuestas neuroen-docrinas por estimulación de receptores 5-HT1A, ponen demanifiesto una disminución del funcionalismo serotoninérgi-co en pacientes depresivos unipolares o bipolares. La adminis-tración de litio, merced a la disminución de la sensibilidad delos receptores somatodendríticos 5-HT1A, incrementa los ni-
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
15Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
Figura 3 Vías de señalización intraneuronales relacionadas con la GSK3 y con diversos factores neurotróficos.
BDNF
TRK TRK Membrana celular
P P
PSOS
SRC GrB2
Ras
Raf
MEK
ERK 1, 2
RSK
CREB
Bcl-2Bcl-xL BDNF Proteínas sustrato para proteincinasa
TKSH2
SH2
SH2
PI-3K
PIP3
AKT
BAD Apoptosis
GSK-3
tau
c-jun
Ciclina D
p-catenina
ISRS PLC
PKC
CREB
BDNFBcl-2Bcl-xL
ERK 1, 2
3-21.qxd 10/4/08 12:11 Página 15

veles de serotonina en la hendidura sináptica (Hotta y Ya-mawaki, 1988; Tamayo et al., 2004), lo que podría explicarla actividad del litio en la fase depresiva del trastorno bipo-lar. Este mecanismo también es compartido por la lamotri-gina (Vinod y Subhash, 2002). Es de destacar que diversosagentes farmacológicos aumentan, al igual que lo hace el li-tio, los niveles de serotonina y ello se acompaña de un in-cremento de la fosforilización de la serina de la GSK3 y portanto su inhibición. Este hecho parece indicar que los agen-tes que mejoran el humor a través de mecanismos serotoni-nérgicos tienen un paso común en la inhibición de la GSK3(Li et al., 2004).
Por otra parte, la hiperactividad inducida por anfetami-nas es considerada como un modelo de manía experimental,que es mediado en gran parte por la actividad de la GSK3.En este modelo, una serie de estabilizadores del humor, in-cluido el litio, antiepilépticos y antipsicóticos, son capacesde atenuar la citada hiperactividad (Beaulieu et al., 2004;Gould et al., 2004). Estos datos apoyan la hipótesis de que lainhibición de la GSK3 está implicada tanto en el efecto anti-depresivo de los eutimizantes como en su eficacia antima-níaca y reguladora del humor (Quiroz et al., 2004).
Efectos neuroprotectores de los reguladores del humor
Aunque los datos neuroanatómicos aún son escasos, existe la suficiente evidencia, procedente básicamente deestudios de neuroimagen, para soportar la hipótesis de laexistencia de alteraciones estructurales en pacientes contrastorno bipolar que podrían considerarse como manifes-taciones de procesos de daño neuronal (Li et al., 2002). Deesta forma, la implicación de las vías de transducción intra-celulares relacionadas con la neuroplasticidad están co-brando cada vez mayor importancia en la fisiopatología deltrastorno bipolar y en la explicación del mecanismo de ac-ción de los reguladores del humor (Goodwin y Jamison,2007; Shaltiel et al., 2007). En este sentido, aunque ya seconocía el efecto neuroprotector del litio, los clásicos anti-epilépticos también parecen estar dotados de esta propie-dad, gracias a la estimulación de la síntesis de diversos fac-tores neurotróficos, lo que contribuiría a prevenir el procesode neurodegeneración asociado al trastorno bipolar (fig. 3).Entre los factores más conocidos en este sentido cabe des-tacar el BDNF, el factor de crecimiento fibroblástico tipo 2(FCF-2), el ERK (extracellular signal-regulated kinase) o elCREB, este último relacionado con los procesos de neurogé-nesis en el hipocampo en sujetos adultos (Manji et al.,2000). Tanto el BDNF como el ERK constituyen dos vías devital importancia en los fenómenos de plasticidad sináptica,que han recibido una creciente atención en relación a la fi-siopatología del trastorno bipolar (Manji y Duman, 2001;Chen y Manji, 2006; Shaltiel et al., 2007). El tratamientocrónico con litio y valproato ocasiona un incremento de lainmunorreactividad al BDNF en la corteza frontal de rata(Einat et al., 2003) e incrementa el fenómeno de neurogé-
nesis en el hipocampo, un efecto mediado, al menos en par-te, por la activación de la vía ERK (Hao et al., 2004).
Por otro lado, la modulación de los niveles de glutama-to ejercida por los agentes eutimizantes de tipo antiepi-léptico podría explicar también su acción neuroprotecto-ra, extrapolable también al litio. En este sentido, Nonakaet al. (1998) demostraron que la administración crónicade litio protegió a células neuronales embrionarias querecibieron 100 veces las concentraciones de glutamatoobservadas en estado de reposo y que usualmente produ-cen apoptosis.
Al efecto neuroprotector del valproato podrían contri-buir también otras modificaciones bioquímicas, como la in-hibición de la histona-deacetilasa (HCAD), enzima responsa-ble de la ramificación axonal durante el neurodesarrollo(Phiel et al., 2001), o el incremento de la actividad de la víaERK-MAP-cinasa tras la activación de las proteínas Ras yRaf (Chen y Manji, 2006). La mayor funcionalidad de estavía se ha asociado a un incremento de la diferenciación ycrecimiento celular en general y a un crecimiento de lasprolongaciones dendríticas en particular.
Estudios en cultivos celulares y en modelos animalesmuestran las propiedades neuroprotectoras del litio frente adiferentes estados nocivos para las neuronas, como puedenser la hiperactivación del receptor NMDA por glutamato, ladeprivación de factores de crecimiento nervioso, la dismi-nución de potasio o el aumento de calcio intraneuronal(Manji et al., 1999; Quiroz et al., 2004). En este efecto neu-roprotector interviene, al menos en parte, la capacidad inhi-bidora de la GSK3, como se pone de manifiesto por un in-cremento de la β-catenina, efecto descrito también en elcaso del valproato (Chen et al., 1999; O’Brien et al., 2004), yuna disminución de la proteína neurotóxica β-amiloide(Phiel et al., 2003).
Estas múltiples dianas de la GSK3 la hacen, lógicamente,participar en múltiples procesos celulares, como la síntesisdel glucógeno, de donde procede su denominación, la trans-cripción genética, efectos relacionados con la plasticidad si-náptica, la apoptosis, la resistencia celular, así como en losritmos circadianos. Estos aspectos tienen un gran interés, yaque el estudio conjunto de datos experimentales indica quela apoptosis de diversos tipos de neuronas guarda relacióncon la activación de la GSK-3. Por el contrario, la inhibiciónde la misma actuaría como factor preventivo de la apopto-sis, aumentaría la plasticidad sináptica y la resistencia celu-lar. En definitiva, la inhibición de la GSK-3 se traduciría porun efecto fundamentalmente neuroprotector (Gould et al.,2004; Gould 2006). Por otra parte, existen evidencias quesugieren la existencia de una relación entre trastornos delhumor y alteraciones de la neuroplasticidad y resistenciacelular, manifestadas tanto en estudios post morten comoin vivo, en los que se observan pérdida de células gliales y/oneuronas, así como atrofia en determinadas zonas cerebra-les (Manji et al., 1999; Quiroz et al., 2004).
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
16 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 16

En este contexto desde el año 1996 se sabe que el litio esun inhibidor directo de la GSK3 (Klein y Melton, 1996;Stambolic et al., 1996) al competir con el magnesio (Ryves yHarwood, 2001). El litio aumenta la fosforilización de la se-rina 9 de la región aminoterminal de la enzima, lo que pro-duce su inhibición. Cabe destacar que este efecto se produ-ce a concentraciones compatibles con las alcanzadas enclínica humana, lo que habla a favor de su trascendencia te-rapéutica desde el punto de vista de la neuroprotección.Además, el efecto in vivo del litio se ve amplificado al aumentar la actividad de la PKC y de la Akt, inhibidoras dela GSK3 al fosforilar la serina (Kirshenboim et al., 2004).También el valproato ejerce un efecto inhibidor de la GSK3a concentraciones terapéuticas (Chen et al., 1999), al igualque la lamotrigina (Li et al., 2002). Por el contrario, la car-bamazepina carece de efectos directos o indirectos sobre laactividad GSK3 (Harwood y Agam, 2003).
Por otro lado, tanto el litio como el valproato aumentanlos niveles de factores de transcripción como el CREB y debcl-2, una proteína dotada de propiedades neuroprotecto-ras y antiapoptóticas (Chen et al., 1999). Ambos agentes,además, inhiben la proteína proapoptótica p53, que activa asu vez a la proteína BAX relacionada con la muerte celular através de la vía de las caspasas (Yuan et al., 2001). Asimismo,los niveles de BDNF también se encuentran incrementados enratas tratadas con litio (Fukumoto et al., 2001).
En estudios de laboratorio, mediante cultivo de células cor-ticales del cerebro de rata, se ha comprobado que el valproato,al igual que el litio, posee efectos directos sobre la neurogéne-sis, incrementado el número de neuronas BrdU-positivas (Haoet al., 2004). Desde la perspectiva clínica, Moore et al. (2000)han confirmado, mediante el empleo de resonancia magnéticaespectroscópica, que el litio aumenta los niveles cerebrales deN-acetil-aspartato, un marcador de viabilidad neuronal, ensujetos diagnosticados de trastorno bipolar, efecto correla-cionado con un aumento del volumen de la sustancia gris. Sinembargo, estos resultados no han podido ser replicados en pa-cientes tratados con valproato (Silverstone et al., 2003).
Finalmente, en relación con los nuevos agentes antiepi-lépticos, los datos disponibles son muy limitados, destacan-do únicamente los resultados obtenidos en estudios de dañocerebral con lamotrigina y topiramato. Los efectos neuro-protectores de estos dos fármacos se han puesto de mani-fiesto en modelos animales de isquemia (Smith y Meldrum,1995; Yang et al., 1998), y en el caso concreto de la lamotri-gina, en estudios experimentales sobre daño neuronal indu-cido por excitotoxinas (Adams et al., 1996) y por toxinas mi-tocondriales (Schulz et al., 1996).
BIBLIOGRAFÍA
Adams J, Collaco-Moraes Y, de Belleroche J. Cyclooxygenase-2 in-duction in cerebral cortex: an intracellular response to synapticexcitation. J Neurochem 1996;66:6-13.
Agam G, Shaltiel G. Possible role of 3’(2’)-phosphoadenosine-5’-phosphate phosphatase in the etiology and therapy of bipolar di-sorder. Prog Neuropsychopharmacol Biol Psychiatry 2003;27:723-7.
Alamo C, López-Muñoz F, Cuenca E. Sales de litio y otros reguladoresdel humor. En: Gómez-Jarabo G, editor. Farmacología de la con-ducta: manual básico para psicoterapeutas y clínicos. Madrid:Síntesis, 1997; p. 247-65.
Alamo C, López-Muñoz F, Cuenca E. Fármacos ansiolíticos. En: Cervi-lla JA, García-Ribera C, editores. Fundamentos biológicos en psi-quiatría. Barcelona: Masson, 2000; p. 299-314.
Allison J, Stewart M. Reduced brain inositol in lithium treated rats.Nature New Biol 1971;233:267-8.
Arystarkhoua E, Sweadner K. Isoform-specific monoclonal antibodiesto Na,K-ATPase alfa subunits: evidence for a tissue-specific pos-ttranslational modification of the alfa subunit. J Biol Chem 1996;271:23407-17.
Asghari V, Wang J, Reiach J. Differential effects of mood stabilizerson Fos/Jun proteins and AP-1 DNA binding activity in humanneuroblastoma SH-SY5Y cells. Brain Res Mol Brain Res 1998;58:95-102.
Avissar S, Schreiber G, Danon A. Lithium inhibits adrenergic and cho-linergic increases in GTP binding in rat cortex. Nature 1988;331:440-2.
Barrett T, Hauger R, Kennedy J. Evidence that a single nucleotidepolymorphism in the promoter of the G protein receptor kinase 3gene is associated with bipolar disorder. Mol Psychiatry 2003;8:546-7.
Bauer M, Pfenning A. Epidemiology of bipolar disorders. Epilepsia 2005;46:8-13.
Beaulieu J, Sotnikova T, Yao W. Lithium antagonizes dopaminede-pendent behaviors mediated by an AKT/glycogen synthase kinase 3signaling cascade. Proc Natl Acad Sci USA 2004;101:5099-104.
Belmaker R. Bipolar disorder. New Engl J Med 2004;351:476-86.Belmaker R, Bersudsky Y, Agam G. How does lithium work on manic
depression? Clinical and psychological correlates of the inositoltheory. Annu Rev Med 1996;47:47-56.
Berridge M, Downes C, Hanley M. Lithium amplifies agonist-depen-dent phosphatidylinositol responses in brain and salivary glands.Biochem J 1982;206:587-95.
Berry G, Buccafusca R, Greer J. Phosphoinositide deficiency due toinositol depletion is not a mechanism of lithium action in brain.Mol Genet Metab 2004;82:87-92.
Biber K, Fiebich B, Gebicke-Harter P. Carbamazepine-induced upre-gulation of adenosine A1-receptors in astrocyte cultures affectscoupling to the phosphoinositol signalling pathway. Neurops-ychopharmacology 1999;20:271-8.
Bormann J. The ABC of GABA receptors. Trends Pharmacol Sci 2000;21:16-9.
Bowden CL, Calabrese JR, Sach G. A placebo-controlled 18 monthstrial of lamotrigine and lithium maintenance treatment in re-cently manic or hypomanic patients with bipolar I disorder. ArchGen Psychiatry 2003;60:392-400.
Brandoli C, Sanna A, De Bernardi M. BDNF and basic fibroblastgrowth factor down regulated NMDA receptor function in cere-bellar granule cells. J Neurosci 1998;18:7953-61.
Cade J. Lithium salts in the treatment of psychotic excitement. Med JAust 1949;2:349-52.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
17Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 17

Calabrese J, Vieta E, Shelton M. Lastest maintenance data on lamo-trigine in bipolar disorder. Eur Neuropsychopharmacol 2003;13(Suppl. 2):57-66.
Calabresi P, Centonze D, Marfia G. An in vitro electrophysiologicalstudy on the effects of phenytoin, lamotrigine and gabapentinon stiatal neurons. Br J Pharmacol 1999;126:689-96.
Cecil K, DelBello MP, Morey R, Strakowski SM. Frontal lobe differen-ces in bipolar disorder as determined by proton MR spectroscopy.Bipolar Disord 2002;4:357-65.
Chen G, Manji H. The extracellular signal-regulated kinase pathway:an emerging promising target for mood stabilizers. Curr OpinPsychiatry 2006;19:313-23.
Chen G, Pan B, Hawver D. Attenuation of cyclic AMP production bycarbamazepine. J Neurochem 1996;67:2079-87.
Chen B, Wang J, Hill B. Lithium and valproate differentially regulatebrain regional expression of phosphorylated CREB and c-Fos.Brain Res Mol Brain Res 1999;70:45-53.
Colin S, Chang H, Mollner S. Chronic lithium regulates the expres-sions of adenylate cyclase and Gi-protein alfa subunit in rat ce-rebral cortex. Proc Natl Acad Sci USA 1991;88:10634-7.
Corbella B, Vieta E, Benabarre A. Utilización del topiramato comonuevo estabilizador del humor. Psiquiatr Biol 2000;7:268-74.
Cramer C, Stagnitto M, Knowles M. Kainic acid and 4-aminopyridineseizures models in mice: evaluation of efficacy of anti-epilepticagents and calcium antagonists. Life Sci 1994;54:271-5.
Cuenca E, Alamo C, López-Muñoz F. Mecanismo de acción de las sales delitio. En: Álvarez E, Pérez J, Pérez V, editores. Clínica de litio. Teoría ypráctica. Barcelona: Publicaciones Permanyer, 2000; p. 11-30.
Dardennes R, Even C, Bange F. Comparison of carbamazepine and li-thium in the prophylaxis of bipolar disorders: a meta-analysis. BrJ Psychiatry 1995;166:378-81.
Davanzo P, Yue K, Thomas M. Proton magnetic resonance spectros-copy of bipolar disorder versus intermittent explosive disorder inchildren and adolescents. Am J Psychiatry 2003;160:1442-52.
Dildy J, Leslie S. Ethanol inhibits NMDA induced increases in free in-tracellular Ca++ in dissociated brain cells. Brain Res 1989;499:383-7.
Divish M, Sheftel G, Boyle A. Differential effect of lithium on fosprotooncogene expression mediated by receptor and post-recep-tor activators of protein kinase C, and cyclic adenosine mono-phosphate: model for its antimanic action. J Neurosci Res 1991;28:40-8.
Dixon J, Hokin L. The antibipolar drug valproate mi- mies lithium ¡ustimulating glutamate release and inositol 1,4,5-trisphosphateaeeumulation in brain corta slices but not accumulation of Inosi-tol monophosphates and bisphosphates. Proc Natl Acad Sci USA1997;94:4757-60.
Dixon J, Los G, Hokin L. Lithium stimulates glutamate «release» andinositol 1,4,5-trisphosphate accumulation via activation of theN-methyl-D-aspartate receptor in monkey and mouse cerebralcortex slices. Proc Natl Acad Sci USA 1994;91:8358-62.
D’Mello S, Anelli R, Calissano P. Lithium induces apoptosis in imma-ture cerebellar granule cells but promotes survival of matureneurons. Exp Cell Res 1994;211:332-8.
Du J, Wei Y, Chen Z. The anticonvulsants lamotrigine, riluzole andvalproate differentially regulate AMPA receptor trafficking: rela-tionship to clinical effects in mood disorders. Neuropsychophar-macology 2007;32:793-802.
Dubovsky S. Calcium channel antagonists as novel agents for manicdepressive disorder. En: Nemeroff C, Schatzberg, editores. Ameri-can Psychiatry Textbook of Psychopharmacology. Washington:American Psychiatric Press, 1995; p. 377-90.
Duman R. Molecular and cellular determinants of stress an antide-pressant treatment. En: Reith MEA, editor. Cerebral signal trans-duction: from first to fourth messengers. Totowa: Humana PressInc., 2000; p. 223-47.
Ebstein R, Belmaker R, Grunhaus L. Lithium inhibition of adrenaline-stimulated adenylate cyclase in humans. Nature 1976;259:411-3.
Einat H, Yuan P, Gould T. The role of the extracellular signal-regula-ted kinase signaling pathway in mood modulation. J Neurosci2003;23:7311-6.
El-Mallakh R, Wyatt R. The Na,K-ATPase hypothesis for bipolar ill-ness. Biol Psychiatry 1995;37:235-44.
Errington A, Madison K, Kumar A. Voltage-gated sodium channelsand their roles in drug action. Curr Anaesth Crit Care 2005;16:81-7.
Farber N, Jiang X, Heinkel C. Antiepileptic drugs and agents that in-hibit voltage-gated sodium channels prevent NMDA antagonistneurotoxicity. Mol Psychiatry 2002;7:726-33.
Fukumoto T, Morinobu S, Okamoto Y. Chronic lithium treatment in-creases the expression of brain-derived neurotrophic factor inthe rat brain. Psychopharmacology (Berl) 2001;158:100-6.
Goldlust A, Su T, Welty D. Effects of anticonvulsant drug gabapentinon the enzymes in metabolic pathways of glutamate and GABA.Epilepsy Res 1995;22:1-11.
Goodwin F, Jamison K. Manic-depressive illness. New York: OxfordUniversity Press, 1990.
Goodwin F, Jamison K. Manic-depressive illness: bipolar disordersand recurrent depression. New York: Oxford University Press,1990.
Gould T. Targeting glycogen synthase kinase-3 as an approach to de-velop novel mood-stabilising medications. Exp Opin Ther Targets2006;10:377-92.
Gould T, Manji H. Signaling networks in the pathophysiology andtreatment of mood disorders. J Psychosom Res 2002;53:687-97.
Gould T, Zarate C, Manji H. Glycogen synthase kinase-3: a target fornovel bipolar disorder treatments. J Clin Psychiatry 2004;65:10-21.
Grimes C, Jope R. Cholinergic stimulation of early growth response-1DNA binding activity requires protein kinase C and mitogen-acti-vated protein kinase kinase activation and is inhibited by sodiumvalproate in SH-SY5Y cells. J Neurochem 1999;73:1384-92.
Hallcher L, Sherman W. The effects of the lithium ion and otheragents on the activity of myo-inositol 1-phosphatase from bovi-ne brain. J Biol Chem 1980;255:10896-901.
Hanaya R, Sasa M, Ujihara H. Suppression by topiratame of epilepti-form burst discharges in hippocampal CA3 neurons of sponta-neously epileptic rat in vitro. Brain Res 1998;789:274-82.
Hao Y, Creson T, Zhang L. Mood stabilizer valproate promotes ERKpathway-dependent cortical neuronal growth and neurogenesis.J Neurosci 2004;24:6590-9.
Harwood A, Agam G. Search for a common mechanism of mood sta-bilizers. Biochem Pharmacol 2003;66:179-89.
Hokin-Neaverson M, Jefferson J. Deficient erythrocyte NaK-ATPaseactivity in different affective states in bipolar affective disorderand normalization by lithium therapy. Neuropsychobiology 1989a;22:18-25.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
18 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 18

Hokin-Neaverson M, Jefferson J. Erythrocyte sodium pump activityin bipolar affective disorder and other psychiatric disorders. Neu-ropsychobiology 1989b;22:1-7.
Hotta I, Yamawaki S. Possible involvement of presynaptic hippocam-pus of rat. Neuropharmacology 1988;27:987-92.
Ikonomov O, Manji H. Molecular mechanism underlying mood stabi-lization in manic-depressive illness: the phenotype challenge. AmJ Psychiatry 1999;156:1506-14.
Keck P, McElroy S. Outcome in the pharmacologic treatment of bipo-lar disorder. J Clin Psychopharmacol 1996;16:15S-23S.
Ketter T, Frye M, Cora-Locatelli G. Pharmacology and pharmacokine-tics of new anticonvulsants. American Psychiatric AssociationMeeting Syllabus 1997:84-5.
Kirshenboim N, Plotkin B, Shlomo S. Lithium-mediated phosphoryla-tion of glycogen synthase kinase-3beta involves PI3 kinase-de-pendent activation of protein kinase C-alpha. J Mol Neurosci2004;24:237-45.
Klein P, Melton D. A molecular mechanism for the effect of lithiumon development. Proc Natl Acad Sci USA 1996;93:8455-549.
Kuzniecky R, Hetherington H, Ho S. Topiramate increases cerebralGABA in healthy humans. Neurology 1998;51:627-9.
Layden B, Diven C, Minadeo N. Li+:Mg2+ competition at therapeuticintracellular Li+ levels in human neuroblastoma SH-SY5Y cells.Bipolar Disord 2000:2:200-4.
Lancaster J, Davies J. Carbamazepine inhibits NMDA-induced depola-rizations in cortical wedges prepared from DBA/2 mice. Experien-tia 1992;48:751-3.
Lecuona E, Luquin S, Avila J. Expressions of the beta-1 and beta-2(AMOG) subunits of the Na, K-ATPase in neural tissues: cellular anddevelopmental distribution patterns. Brain Res Bull 1996;40:167-74.
Lee Y, Hamamura T, Ohashi K. Carbamazepine suppresses metham-pheramine-induced Fos expresión in a regionally specific mannerin the rat brain. Possible neuronal substrates responsible for an-timanic effects of mood stabilizers. Neuropsychopharmacology2000;22:530-7.
Lenox R, McNamara R, Watterson J. Myristoylated alanine-rich C ki-nase substrate (MARCKS): a molecular target for the therapeuticaction of mood stabilizers in the brain? J Clin Psychiatry 1996;57:S23-S31.
Lenox R, Mcnamara R, Papke R. Neurobiology of lithium: an update. J Clin Psychiatry 1998;59:37-47.
Lesch K, Aulakh C, Tolliver T. Differential effects of long-term lithiumand carbamazepine administration on Gs alpha and Gi alpha pro-tein in rat brain. Eur J Pharmacol 1991;207:355-9.
Lewin E, Bleck V. Cyclic AMP accumulation in cerebral cortical slices:effect of carbamazepine, phenobarbital, and phenytoin. Epilepsia1977;18:237-42.
Li X, Ketter T, Frye M. Synaptic, intracellular, and neuroprotective me-chanism of anticonvulsants: are they relevant for the treatmentand course of bipolar disorders? J Affect Disord 2002;69:1-14.
Li X, Zhu W, Roh M. In vivo regulation of glycogen synthase kinase-3beta (GSK3beta) by serotonergic activity in mouse brain. Neu-ropsychopharmacology 2004;29:1426-31.
López-Muñoz F, Alamo C, Cuenca E. La «década de oro» de la psico-farmacología (1950-1960): trascendencia histórica de la intro-ducción clínica de los psicofármacos clásicos. Psiquiatria.COM(revista electrónica), Sep, 4; 2000. Disponible en URL: http://www.psiquiatria.com/psiquiatria/revista/47/1800/?++interactivo.
López-Muñoz F, Vieta E, Rubio G. Bipolar disorder as an emergingpathology in the scientific literature: a bibliometric approach. J Affect Disord 2006;92:161-70.
Loscher W. Valproate: a reappraisal of its pharmacodynamics proper-ties and mechanisms of action. Prog Neurobiol 1999;58:31-59.
Lydiard R. The Role of GABA in anxiety disorders. J Clin Psychiatr2003;64(Suppl. 3):21-7.
Macritchie K, Geddes J, Scott J. Valproic acid, valproate and divalpro-ex in the maintenance treatment of bipolar disorder (CochraneReview). Cochrane Database Syst Rev 2001;3:CD003196.
Malick N, Canfield V, Beckers M. Identification of mammalian Na,K-ATPase subunit. J Biol Chem 1996;271:22574-758.
Manji H, Duman R. Impairments of neuroplasticity and cellular resi-lience in severe mood disorders: implications for the develop-ment of novel therapeutics. Psychopharmacol Bull 2001;35:5-49.
Manji H, Potter W, Lenox R. Signal transduction pathways: moleculartargets for lithium’s actions. Arch Gen Psychiatry 1995;52:531-43.
Manji H, Chen G, Hsiao J. Regulation of signal transduction pathwaysby mood-stabilizing agents: implications for the delayed onset oftherapeutic efficacy. J Clin Psychiatry 1996a;57(Suppl. 13):34-46.
Manji H, Bersudsky U, Chen G. Modulation of PKC isozymes andsubstrates by lithium: the role of myo-inositol. Neuropharmaco-logy 1996b;15:370-81.
Manji H, Moore G, Chen G. Lithium at 50: have the neuroprotectiveeffects of this unique cation been overlooked? Biol Psychiatry1999;46:929-40.
Manji H, Moore G, Rajkowska G. Neuroplasticity and cellular resilien-ce in mood disorders. Mol Psychiatry 2000a;5:578-93.
Manji H, Moore G, Chen G. Clinical and preclinical evidence for theneurotrophic effects of mood stabilizers: implications for the pa-thophysiology and treatment of manic-depressive illness. BiolPsychiatry 2000b;48;740-54.
Martin M, Regan C. The anticonvulsant sodium valproate specificallyinduces the expression of a rat glial heat shock protein which isidentified as the collagen type IV receptor. Brain Res 1988;459:131-7.
McEwen B, Magarinos A. Stress effects on morphology and functionof the hippocampus. Ann New York Acad Sci 1997;821:271-84.
McGeer E, Zhu S. Lamotrigine protects against kainate but not ibote-nate lesions in rat striatum. Neurosci Lett 1990;112:348-51.
Moore G, Bebchuck J, Manji H. Proton MRS in manic depressive ill-ness: monitoring of lithium induced modulation of brain myo-inositol. Soc Neurosci 1997;23:335-6.
Moore G, Bebchuk J, Hasanat K. Lithium increases N-acetyl-aspartatein the human brain: in vivo evidence in support of bcl-2’s neuro-trophic effects? Biol Psychiatry 2000;48:1-8.
Mork A, Geisler A. The effects of lithium in vitro and ex vivo onadenylate cyclase in brain are exerted by distinct mechanisms.Neuropharmacology 1989;28:307-11.
Mork A, Geisler A, Hollund P. Effects of lithium on second messengersystems in the brain. Pharmacol Toxicol 1992;71:4-17.
Motohashi N. GABA receptor alterations after chronic lithium admi-nistration. Comparison with carbamazepine and sodium valproa-te. Prog Neuro-Psychopharmacol Biol Psychiatry 1992;16:571-9.
Nashorski S, Ragan C, Challis R. Lithium and the phosphoinositidecycle: an example of uncompetitive inhibition and its pharmaco-logical consequences. Trends Pharmacol Sci 1991;12:297-303.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
19Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 19

Nestler E, Terwilliger R, Duman R. Regulation of endogenous rybosi-lation by acute and chronic lithium in rat brain. J Neurochem1995;64:2319-24.
Niculescu A, Segal D, Kuczenski R. Identifying a serious of candidategenes for mania and psychosis: a convergent functional geno-mics approach. Physiol Genomics 2000;9:83-91.
Nonaka S, Chuang D. Neuroprotective effects of chronic lithium onfocal cerebral ischemia in rats. Neuroreport 1998;9:2081-4.
O’Brien W, Harper A, Jove F. Glycogen synthase kinase-3beta ha-ploinsufficiency mimics the behavioral and molecular effects oflithium. J Neurosci 2004;24:6791-98.
O’Donnell T, Rotzinger S, Nakashima T. Chronic lithium and sodiumvalproate both decrease the concentration of myo-inositol andincrease the concentration of inositol monophosphates in ratbrain. Brain Res 2000;880:84-91.
Olpe H, Baudry M, Jones R. Electrophysiological and neurochemicalinvestigations on the action of carbamazepine on the rat hippo-campus. Eur J Pharmacol 1985;110:71-80.
Olpe H, Steinmann M, Pozza M. Valproate enhances GABA-A media-ted inhibition of locus coeruleus neurones in vitro. Neunyn Schmiedebergs Arch Pharmacol 1988;338:655-7.
Pacheco M, Jope R. Modulation of carbachol-stimulated AP-1 DNAbinding activity by therapeutic agents for bipolar disorder in hu-man neuroblastoma SH-SY5Y cells. Brain Res Mol Brain Res 1999;72:138-46.
Parthasarathy L, Vadnal R, Ramesh T. Myo-inositol monophosphatasefrom rats testes: purification and properties. Arch BiochemBiophys 1993;304:94-101.
Patel N, DelBello M, Cecil K. Lithium treatment effects on myo-inosi-tol in adolescents with bipolar depression. Biol Psychiatry 2006;60:998-1004.
Phiel C, Zhang F, Huang F. Histone deacetylase is a direct target ofvalproic acid, a potent anticonvulsant, mood stabilizer, and tera-togen. J Biol Chem 2001;276:36734-41.
Phiel C, Wilson C, Lee V. GSK-3alpha regulates production of Alzhei-mer’s disease amyloid-beta peptides. Nature 2003;423:435-9.
Pope H, McElroy S, Keck P. Valproate in the treatment of acute ma-nia. A placebo-controlled study. Arch Gen Psychiatry 1991;48:62-8.
Post R. Do the epilepsies, pain syndromes, and affective disordersshare common kindling-like mechanisms? Epilepsy Res 2002;50:203-19.
Post R, Weiss, S, Chuang D. Mechanism of action of anticonvulsi-vants in affective disorders: comparison with lithium. J ClinPsychopharmacol 1992;12:238-5.
Quiroz J, Singh J, Gould T. Emerging experimental therapeutics forbipolar disorder: clues from the molecular pathophysiology. MolPsychiatry 2004;9:756-76.
Rosenfeld W. Topiramate: a review of preclinical, pharmacokinetic,and clinical data. Clin Ther 1997;19:1294-308.
Ryves W, Harwood A. Lithium inhibits glycogen synthase kinase-3 bycompetition for magnesium. Biochem Biophys Res Commun2001;280:720-5.
Schou M. Lithium. En: Ban TA, Ray OS, editores. A history of the CINP.Brentwood: JM Productions, 1996; p. 401-3.
Schreiber G, Avissar S. Application of G proteins in the moleculardiagnosis of psychiatric disorders. Exp Rev Mol Diagn 2003;3:89-100.
Schulz J, Matthews R, Henshaw D. Neuroprotective strategies for treat-ment of lesions produced by mitochondrial toxins: implicationsfor neurodegenerative diseases. Neuroscience 1996;71:1043-8.
Sela B. On the relevance of protein kinase C to lithium therapy in bipolar disorder. Harefuah 2004;143:414-6.
Shaltiel G, Chen G, Manji H. Neurotrophic signaling cscades in thepathophysiology and treatment of bipolar disorder. Curr OpinPharmacol 2007;7:22-6.
Shiah I, Yatham L, Lam R. Effects of divalproex sodium on 5-HT1A re-ceptor function in healthy human males: hypothermic, hormo-nal, and behavioural responses to ipsapirone. Neuropsychophar-macology 1997;17:382-90.
Silva H. Mechanism of action of mood stabilizers. Ver Chil Neuro-Psi-quiatr 2001;39:219-30.
Silverstone P, Wu R, O’Donnell T. Chronic treatment with lithium, butnot sodium valproate, increases cortical N-acetyl-aspartate con-centrations in euthymic bipolar patients. Int Clin Psychopharma-col 2003;18:73-9.
Skolnick P, Layer R, Popik P. Adaptation of N-methyl-D-aspartate(NMDA) receptors following antidepressant treatment: implica-tions for the pharmacotherapy of depression. Pharmacops-ychiatry 1996;29:23-6.
Smith D. Strategies in behavioral research on affective disorders:principles, problems and pitfalls. Pharmacol Toxicol 1990;66:S139-143.
Smith S, Meldrum B. Cerebroprotective effect of lamotrigine afterfocal ischemia in rats. Stroke 1995;26:117-21.
Soderling TR. CaM-Kinases: modulators of synaptic plasticity. CurrOpin Neurobiol 2000;10:375-80.
Southam E, Kirkby D, Higgins G. Lamotrigine inhibits monoamine uptake in vitro and modulates 5-hydroxytryptamine uptake inrats. Eur J Pharmacol 1998;358:19-24.
Spiegelberg B, Xiong J, Smith J. Cloning and characterization of amammalian lithium-sensitive bisphosphate 3’-nucleotidase inhi-bited by inositol 1,4 bisphosphate. J Biol Chem 1999;274:13619-28.
Stambolic V, Ruel L, Woodgett J. Lithium inhibits glycogen synthasekinase-3 activity and mimics wingless signalling in intact cells.Curr Biol 1996;6:1664-8.
Stefani A, Sjpadoni F, Siniscalchi A. Lamotrigine inhibits Ca2+ cur-rents in cortical neurons: functional implications. Eur J Pharma-col 1996;307:113-6.
Tamayo J, Zárate C, Vieta E. Medicamentos para los trastornos bipolares:una revisión de sus diferencias terapéuticas y farmacológicas. Par-te II: de la sinapsis a los genes. Actas Esp Psiquiatr 2004;32:18-37.
Van Calker D, Belmaker R. The high affinity inositol transportsystem-implications for the pathophysiology and treatment ofbipolar disorder. Bipolar Disord 2000;2:102-7.
Vieta E. Mood stabilization in the treatment of bipolar disorder: focuson quetiapine. Hum Psychopharmacol Clin Exp 2005;20:225-36.
Vinod K, Subhash M. Lamotrigine induced selective changes in 5-HT1A
receptor mediated response in rat brain. Neurochem Int 2002;40:315-9.
Volonte C, Ciotti M, Merlo D. LiCl promotes survival of GABAergicneurons from cerebellum and cerebral cortex: LiC induces survi-val of GABAergic neurons. Neurosci Letters 1994;172:6-10.
Wada A, Yokoo H, Yanagita T. Lithium: potential therapeutics againstacute brain injures and chronic neurodegenerative diseases. J Pharmacol Sci 2005;99:307-21.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
20 Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 20

Waldmeier P. Mechanisms of action of lithium in affective disorders:a status report. Pharmacol Toxicol 1990;66:S121-32.
Wang S, Huang C, Hsu K. Presynaptic inhibition of excitatory neuro-transmission by lamotrigine in the rat amygdalar neurons.Synapse 1996;24:248-55.
Wang J, Brown C, Young L. Differential display PCR reveals novel tar-gets for the mood-stabilizing drug valproate including the mole-cular chaperone GRP78. Mol Pharmacol 1999;55:521-7.
Wang P, Ketter T, Becker O. New anticonvulsant medication uses inbipolar disorder. CNS Spectrum 2003;8:930-2, 941-7.
Watson D, Lenox R. Chronic lithium induced down-regulation ofMARCKS in inmortalized hippocampal cells: potentiation bymuscarinic receptor activation. J Neurochem 1996;67:767-77.
Watson D, Watterson J, Lenos R. Sodium valproate down-regulatesthe myristoylated alanine-rich C kinase substrate (MARCKS) in in-mortalized hippocampal cells: a property of protein kinase C-me-diated mood stabilizers. J Pharmacol Exp Ther 1998;285:307-16.
Wiborg O, Druger T, Jakobsen S. Region-selective effects of long-term lithium and carbamazepine administration on cyclic AMPlevels in rat brain. Pharmacol Toxicol 1999;84:88-93.
Williams R, Harwod A. Lithium therapy and signal transduction.Trends Pharmacol Sci 2000;21:61-4.
Williams R, Cheng L, Mudge A. A common mechanism of action forthree mood-stabilizing drugs. Nature 2002;417:292-5.
Yang Y, Shuaib A, Li Q. Neuroprotection by delayed administration oftopiramate in a rat model of middle cerebral artery embolization.Brain Res 1998;804:169-76.
Yuan P, Chen G, Huang L. Lithium stimulates gene expression throughthe AP-1 transcription factor pathway. Brain Res Mol Brain Res1998;58:225-30.
Yuan P, Huang L, Jiang Y. The mood stabilizer valproic acid activatesmitogen-activated protein kinases and promotes neurite growth.J Biol Chem 2001;276:31674-83.
Bases neurobiológicas del empleo de antiepilépticos en el trastorno bipolarC. Álamo, et al.
21Actas Esp Psiquiatr 2008;36(Suppl. 3):3-21
3-21.qxd 10/4/08 12:11 Página 21
Related Documents











