*Department of Pharmacology and Pharmacotherapy, Faculty of Pharmaceutical Sciences, University of Copenhagen, Denmark Department of Neuroscience, Medical Faculty, NTNU-Norwegian University of Science and Technology, Norway Glucose is the primary energy substrate for the adult mammalian brain, but lactate and ketone bodies can be used as alternative energy substrates (Owen et al. 1967; Schurr et al. 1988, 1997; Dalsgaard 2006). Ketone bodies include b- hydroxybutyrate, acetoacetate and acetone, of which the latter is generally considered of little metabolic significance. When glucose availability is too low to meet brain require- ments, such as during starvation, the liver generates ketone bodies from the catabolism of fatty acids (Owen et al. 1967). The ketone bodies enter the brain via the monocarboxylate transporters (MCT) and during prolonged starvation it may even replace glucose as the primary fuel in adult brain (Owen et al. 1967). It is well known that starvation or a ketogenic diet diminishes the number of seizures in some patients with epilepsy. Although its biochemical basis is not clarified, several hypotheses have been suggested (Morris 2005; Gasior et al. 2006; Bough and Rho 2007; Ma et al. 2007). One of these hypotheses involves an altered brain amino acid metabolism (Hawkins et al. 1971; Erecinska et al. 1996; Daikhin and Yudkoff 1998; Yudkoff et al. 2001; Melo et al. 2006). Another recently proposed hypothesis suggests an altered ATP distribution in b-hydroxybutyrate nourished cells affecting K ATP channels (Ma et al. 2007). Glucose and b-hydroxybutyrate are both metabolized to acetyl-CoA, however, the initial catabolic steps take place in different compartments; i.e., glucose is mainly metabolized via glycolysis in the cytosol and subsequently pyruvate is metabolized in the mitochondria, whereas b-hydroxybutyrate directly enters the mitochondria. Thus, the required activity of the malate–aspartate shuttle (MAS) transferring reducing equivalents generated in glycolysis is negligible when b- hydroxybutyrate is metabolized and this could affect the Received November 21, 2008; revised manuscript received April 1, 2009; accepted April 8, 2009. Address correspondence and reprint requests to Dr Trine Meldgaard Lund, Faculty of Pharmaceutical Sciences, University of Copenhagen, Universitetsparken 2, 2100 Copenhagen, Denmark. E-mail: [email protected] Abbreviations used: b-HOB, b-hydroxybutyrate; LC-MS, liquid chromatography mass spectrometry; MAS, malate–aspartate shuttle; MCL, molecular carbon labeling; MCT, monocarboxylate transporter. Abstract Ketone bodies serve as alternative energy substrates for the brain in cases of low glucose availability such as during starvation or in patients treated with a ketogenic diet. The ketone bodies are metabolized via a distinct pathway confined to the mitochondria. We have compared metabolism of [2,4- 13 C]b-hydroxybutyrate to that of [1,6- 13 C]glucose in cul- tured glutamatergic neurons and investigated the effect of neuronal activity focusing on the aspartate–glutamate homeostasis, an essential component of the excitatory activity in the brain. The amount of 13 C incorporation and cellular content was lower for glutamate and higher for aspartate in the presence of [2,4- 13 C]b-hydroxybutyrate as opposed to [1,6- 13 C]glucose. Our results suggest that the change in aspartate–glutamate homeostasis is due to a decreased availability of NADH for cytosolic malate dehydrogenase and thus reduced malate–aspartate shuttle activity in neurons using b-hydroxybutyrate. In the presence of glucose, the glutamate content decreased significantly upon activation of neurotransmitter release, whereas in the presence of only b- hydroxybutyrate, no decrease in the glutamate content was observed. Thus, the fraction of the glutamate pool available for transmitter release was diminished when metabolizing b- hydroxybutyrate, which is in line with the hypothesis of for- mation of transmitter glutamate via an obligatory involvement of the malate–aspartate shuttle. Keywords: aspartate, energy, glutamate, ketone bodies, metabolism, b-hydroxybutyrate. J. Neurochem. (2009) 110, 80–91. JOURNAL OF NEUROCHEMISTRY | 2009 | 110 | 80–91 doi: 10.1111/j.1471-4159.2009.06115.x 80 Journal Compilation Ó 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91 Ó 2009 The Authors

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

*Department of Pharmacology and Pharmacotherapy, Faculty of Pharmaceutical Sciences, University of Copenhagen, Denmark
�Department of Neuroscience, Medical Faculty, NTNU-Norwegian University of Science and Technology, Norway
Glucose is the primary energy substrate for the adultmammalian brain, but lactate and ketone bodies can be usedas alternative energy substrates (Owen et al. 1967; Schurret al. 1988, 1997; Dalsgaard 2006). Ketone bodies include b-hydroxybutyrate, acetoacetate and acetone, of which thelatter is generally considered of little metabolic significance.When glucose availability is too low to meet brain require-ments, such as during starvation, the liver generates ketonebodies from the catabolism of fatty acids (Owen et al. 1967).The ketone bodies enter the brain via the monocarboxylatetransporters (MCT) and during prolonged starvation it mayeven replace glucose as the primary fuel in adult brain (Owenet al. 1967).
It is well known that starvation or a ketogenic dietdiminishes the number of seizures in some patients withepilepsy. Although its biochemical basis is not clarified,several hypotheses have been suggested (Morris 2005;Gasior et al. 2006; Bough and Rho 2007; Ma et al. 2007).One of these hypotheses involves an altered brain amino acidmetabolism (Hawkins et al. 1971; Erecinska et al. 1996;Daikhin and Yudkoff 1998; Yudkoff et al. 2001; Melo et al.
2006). Another recently proposed hypothesis suggests analtered ATP distribution in b-hydroxybutyrate nourishedcells affecting KATP channels (Ma et al. 2007). Glucose andb-hydroxybutyrate are both metabolized to acetyl-CoA,however, the initial catabolic steps take place in differentcompartments; i.e., glucose is mainly metabolized viaglycolysis in the cytosol and subsequently pyruvate ismetabolized in the mitochondria, whereas b-hydroxybutyratedirectly enters the mitochondria. Thus, the required activityof the malate–aspartate shuttle (MAS) transferring reducingequivalents generated in glycolysis is negligible when b-hydroxybutyrate is metabolized and this could affect the
Received November 21, 2008; revised manuscript received April 1,2009; accepted April 8, 2009.Address correspondence and reprint requests to Dr Trine Meldgaard
Lund, Faculty of Pharmaceutical Sciences, University of Copenhagen,Universitetsparken 2, 2100 Copenhagen, Denmark.E-mail: [email protected] used: b-HOB, b-hydroxybutyrate; LC-MS, liquid
chromatography mass spectrometry; MAS, malate–aspartate shuttle;MCL, molecular carbon labeling; MCT, monocarboxylate transporter.
Abstract
Ketone bodies serve as alternative energy substrates for the
brain in cases of low glucose availability such as during
starvation or in patients treated with a ketogenic diet. The
ketone bodies are metabolized via a distinct pathway confined
to the mitochondria. We have compared metabolism of
[2,4-13C]b-hydroxybutyrate to that of [1,6-13C]glucose in cul-
tured glutamatergic neurons and investigated the effect of
neuronal activity focusing on the aspartate–glutamate
homeostasis, an essential component of the excitatory activity
in the brain. The amount of 13C incorporation and cellular
content was lower for glutamate and higher for aspartate in
the presence of [2,4-13C]b-hydroxybutyrate as opposed to
[1,6-13C]glucose. Our results suggest that the change in
aspartate–glutamate homeostasis is due to a decreased
availability of NADH for cytosolic malate dehydrogenase and
thus reduced malate–aspartate shuttle activity in neurons
using b-hydroxybutyrate. In the presence of glucose, the
glutamate content decreased significantly upon activation of
neurotransmitter release, whereas in the presence of only b-
hydroxybutyrate, no decrease in the glutamate content was
observed. Thus, the fraction of the glutamate pool available
for transmitter release was diminished when metabolizing b-
hydroxybutyrate, which is in line with the hypothesis of for-
mation of transmitter glutamate via an obligatory involvement
of the malate–aspartate shuttle.
Keywords: aspartate, energy, glutamate, ketone bodies,
metabolism, b-hydroxybutyrate.
J. Neurochem. (2009) 110, 80–91.
JOURNAL OF NEUROCHEMISTRY | 2009 | 110 | 80–91 doi: 10.1111/j.1471-4159.2009.06115.x
80 Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91� 2009 The Authors

homeostasis of essential neuroactive amino acids in the cell.In particular, the balance between aspartate and glutamate inglutamatergic neurons might be affected since synthesis oftransmitter glutamate entails the operation of one of thecarriers of MAS, i.e. the ketodicarboxylate carrier(Palaiologos et al. 1988).
Only a few studies have compared the metabolism ofketone bodies with that of glucose. A study using synapto-somes (McKenna et al. 1994) and one using culturedneurons (Lopes-Cardozo et al. 1986), both focus on thecomplete oxidation of the substrates, measuring the produc-tion of CO2, whereas a detailed metabolic mapping of theeffects of these two substrates was not performed. Moreover,in a recent study by Scafidi et al. (2009) b-hydroxybutyratemetabolism was studied in rats in vivo using 13C-labeling. Inanalogy with this the present study was designed to comparemetabolism of [1,6-13C]glucose and [2,4-13C]b-hydroxybu-tyrate to elucidate glutamate and aspartate homeostasis inresting as well as activated cultured cerebellar granuleneurons, which are mainly glutamatergic (Drejer et al. 1982;Sonnewald et al. 2004). Neuronal activity was induced byrepetitive exposure to NMDA selectively causing vesicularrelease of neurotransmitter glutamate (Bak et al. 2003). It haspreviously been shown that glucose can maintain neuro-transmitter homeostasis as the only energy substrate, whereasthis cannot be fulfilled by lactate, during synaptic activityinduced in cultured cerebellar granule neurons (Bak et al.2006). However, whether b-hydroxybutyrate can compensatefor a lack of glucose and maintain neurotransmitter homeo-stasis is not known.
Materials and methods
MaterialsNMRI mice were obtained from Taconic M&B (Ry, Denmark),
plastic tissue culture flasks from NUNC A/S (Roskilde, Denmark)
and fetal calf serum from Harlan Sera-Lab Ltd. (Sussex, UK).
Culture medium, poly-D-lysine (MW > 300 000), L-glutamine, p-aminobenzoic acid (pABA), sodium (R)-(-)-3-hydroxybutyrate were
from Sigma (St Louis, MO, USA) and penicillin from LEO
(Ballerup, Denmark). o-Phthalaldehyde-reagent was obtained from
Agilent Technologies (Nærum, Denmark). Sodium D-b-[2,4-13C]hydroxybutyrate (99% enriched) was from Cambridge
Isotopes Laboratories (Woburn, MA, USA) and D-[1,6-13C]glucose
(98% enriched) from Isotec (St Louis, MO, USA). For amino acid
analysis by the LC-MS method, the EZ:faast LC-MS kit (KHO-
7337) was used and for HPLC, a Gemini column was used, both
from Phenomenex (Torrance, CA, USA). All other chemicals were
of the purest grade available from regular commercial sources.
Cell culturesCerebellar neurons were isolated from 7-day-old mice and cultured as
described by Schousboe et al. (1989). In brief, after dissection, the
tissue was trypsinized (0.25 mg/mL trypsin, 15 min, 37�C) followed
by trituration in a Dnase solution (75 IU/mL) containing soybean
trypsin inhibitor (0.52 mg/mL). The cells were suspended inmodified
Dulbecco’s minimum essential medium (Schousboe et al. 1989)
containing 24.5 mM KCl, 12 mM glucose, 7 lM pABA, 50 lMkainic acid [to inhibit GABAergic cell function (Drejer and
Schousboe 1989)], 0.05 I.U. insulin, 50 000 I.U. penicillin and 10%
fetal calf serum and seeded in poly-D-lysine (50 mg/L) coated 25 cm2
or 80 cm2 culture flasks at a density of 2.75 · 106 or 3.50 · 106 cells/
mL, respectively. The medium was supplemented twice (day 2 and 5)
with an aliquot of glucose (1.2 M) to maintain a minimum
concentration of 12 mM. Cytosine arabinoside was added to a final
concentration of 20 lM after 2 days in culture to prevent astrocytic
proliferation. The cells were cultured for 7–8 days at which point the
cultures consist of 80–90% cells with glutamatergic characteristics
(Drejer and Schousboe 1989).
Superfusion experimentsThe culture medium was replaced with a Mg2 + -free Tris-buffered
saline solution (50 mM Tris, 135 mM NaCl, 5 mM KCl, 1 mM
CaCl2, pH 7.4, 37�C) containing either 1 mM [1,6-13C]glucose or
1 mM [2,4-13C]b-hydroxybutyrate or combinations of labeled and
unlabeled substrates. (It should be noted that Tris-buffer does not
enter the cells and thus does not contribute to intracellular
buffering). Immediately thereafter, the cultures were placed in a
heated (37�C) superfusion system in a sloping position so that the
media could flow in from one end of the flask and be sucked out
from the other. In this system the superfusion medium could be
repetitively alternated as described in detail by Drejer et al. (1987).The cells were covered by a nylon mesh (80 lm) and superfused
with the Tris-buffered saline solution containing the labeled
substrates (37�C, 4 mL/min) for 74 min (control cultures). In some
of the cultures the medium was repetitively changed every third
minute for 30 s to a medium known to induce neuronal activity, i.e.
neurotransmitter release (Bak et al. 2006). This depolarizing
medium contained 300 lM NMDA, 10 lM glycine and an elevated
potassium concentration (15 mM KCl) with an equimolar reduction
in sodium (to 125 mM NaCl). Using this superfusion system the
effect of repetitive depolarization on intermediary metabolism in
neuronal cultures could be investigated in the presence of either
[1,6-13C]glucose or [2,4-13C]b-hydroxybutyrate. The cell cultures
were subsequently washed twice in ice-cold phophate-buffered
saline (137 mM NaCl, 2.7 mM KCl, 7.3 mM Na2HPO4, 1.5 mM
KH2PO4, 0.9 mM CaCl2, 0.5 mM MgCl2, pH 7.4) and metabolites
extracted using 70% v/v ethanol. The cell residues were scraped off
the dish and collected. The cell extract was separated from insoluble
proteins after centrifugation at 20 000 g for 20 min at 4�C. The cellextracts were lyophilized and re-dissolved in water for analysis by
LC-MS and 13C NMR. Protein contents in the pellets were
determined according to Lowry et al. (1951) using bovine serum
albumin as standard.
HPLCAspartate and glutamate were quantified by HPLC using precolumn,
on-line o-phthalaldehyde-reagent-derivatization and fluorescence
detection on a Shimadzu 10A HPLC system equipped with a
Shimadzu RF-10AXL fluorescence detector (Geddes and Wood
1984). The chromatography was performed on a Gemini C18
column (150 · 4.6 mm, particle size 5 lm, pore size 110 A).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91
b-hydroxybutyrate alters glutamate homeostasis | 81

13C NMRLyophilized cell extracts were dissolved in 200 lL D2O containing
0.1% ethylene glycol. Proton decoupled 150.92 MHz 13C NMR
spectra were obtained on a Bruker 600 MHz spectrometer (Bruker
Analytik GmbH, Rheinstetten, Germany). Spectra were accumulated
using a 30� pulse angle, an acquisition time of 1.08, and a 0.5 s
relaxation delay. The number of scans was typically 50 000.
Relevant peaks were integrated and quantified using ethylene glycol
as quantification standard. Peak areas were corrected for nuclear
Overhauser and relaxation effects by applying correction factors.
LC-MSAspartate and glutamate in the cell extracts were derivatized using
Phenomenex EZ:faast amino acid kit (KHO-7337) and analyzed on
a Shimadzu LCMS-2010 mass spectrometer with electrospray
ionization coupled to a Shimadzu 10A VP HPLC system.
Percentage incorporation of 13C in each isotopomer of glutamate
and aspartate was determined after correction for natural abun-
dance as described by Biemann (1962). Mono labeling was
designated (M + 1), double labeling (M + 2), triple labeling
(M + 3) etc. To obtain a measure of the total 13C incorporation
in each amino acid, an average percent of labeling, called percent
molecular carbon labeling (MCL) for each metabolite was
calculated as described by Bak et al. (2006). As an example,
glutamate can have from one to five 13C atoms (designated M + 1
to M + 5). Percentage of each isotopomer (M + 1, M + 2, etc.) is
multiplied by the ratio of labeled carbon atoms (1/5 for M + 1, 2/5
for M + 2 etc.); these values are summed and expressed as the
average percentage of labeling for each metabolite. MCL reflects
total labeling and these values are dependent upon both the acetyl
CoA enrichment and the TCA cycling.
Metabolic pathways and labeling patternsThe position of 13C-labeling in [1,6-13C]glucose and [2,4-13C]b-hydroxybutyrate was chosen to make a comparison possible, since
they are both metabolized to [2-13C]acetyl-CoA (Fig. 1a).
[1,6-13C]Glucose can be converted to [3-13C]pyruvate via glycol-
ysis and subsequently to [2-13C]acetyl-CoA. On the other hand,
[2,4-13C]b-hydroxybutyrate is metabolized to [2-13C]acetyl-CoA
via [2,4-13C]acetoacetate and [2,4-13C]acetoacetyl-CoA.
[2-13C]Acetyl-CoA generated from either [1,6-13C]glucose or
[2,4-13C]b-hydroxybutyrate may condense with unlabeled oxalo-
acetate producing [2-13C]citrate initiating the first turn of label
incorporation into TCA-cycle intermediates and related amino
acids, i.e. glutamate and aspartate (Fig. 1a). The subsequent steps
yield [4-13C]a-ketoglutarate, which may be transaminated to
[4-13C]glutamate giving rise to a singlet in the 13C NMR spectrum
and mono labeling (M + 1) in the mass spectrum. If [4-13C]a-ketoglutarate is decarboxylated to succinyl-CoA for further
TCA-cycle metabolism, labeling of malate and oxaloacetate will
occur in either the C2 or C3 position (due to the symmetry of
succinate). [2/3-13C]Oxaloacetate may be transaminated to
[2/3-13C]aspartate (singlet, M + 1) catalyzed by aspartate amino-
transferase. Alternatively, [2/3-13C]oxaloacetate may be condensed
with either labeled or unlabeled acetyl-CoA, giving rise to
double labeling (M + 2), i.e. [3,4-13C] or [2,4-13C]glutamate
(Fig. 1b) or mono labeling (M + 1), i.e. [2-13C] or [3-13C]gluta-
mate, respectively. Triple labeling (M + 3) may arise from three
consecutive condensations of [2-13C]acetyl-CoA. Incorporation of13C, labeling patterns plus integrals for glutamate and aspartate
were analyzed by LC-MS and 13C NMR in cell extracts. The
cellular content of glutamate and aspartate was determined by
HPLC.
[2,4-13C]ββ-Hydroxybutyrate
[2,4-13C]Acetoacetate
[2,4-13C]Acetoacetyl-CoA [ 2-13C]Acetyl-CoA
[2-13C]Citrate
[4-13C]Isocitrate
[4-13C]α-Ketoglutarate
[2/3-13C]Succinate
[3-13C]Succinyl-CoA
[2/3-13C]Malate
[2/3-13C]Oxaloacetate
TCA Cycle
(a)
[4-13C]Glutamate
[2/3-13C]Fumarate
[1,6-13C]Glucose
[3-13C]Pyruvate
[2/3-13C]Aspartate
TCAcycle
12345
Oxaloacetate
(b)
1234
[2-13C]Acetyl-CoA
12
1234
1. turn
2. turn3. turn
1234
1.turn
2.turn
3.turn
1.turn
2.turn
3.turn
Aspartate
Glutamate
Fig. 1 Metabolic scheme showing the metabolism of [2,4-13C]b-hy-
droxybutyrate and [1,6-13C]glucose and the position of 13C-labeling in
aspartate and glutamate subsequent to condensation of unlabeled
oxaloacetate and [2-13C]acetyl-CoA, i.e. the first turn of the TCA cycle
(a). A simplified scheme showing labeling patterns (as black circles) in
glutamate and aspartate subsequent to condensation of unlabeled
oxaloacetate and [2-13C]acetyl-CoA and the following turns in which
labeled oxaloacetate condenses with [2-13C]acetyl-CoA (b).
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91� 2009 The Authors
82 | T. M. Lund et al.

Data analysisGlutamate is labeled from the TCA-cycle intermediate a-ketoglu-tarate by transamination, primarily by aspartate aminotransferase
having a very high activity (Drejer et al. 1985; Yudkoff et al. 1994),thus, the labeling pattern of glutamate reflects that of the TCA-cycle
intermediate a-ketoglutarate (Bak et al. 2006). From 13C NMR data
a TCA-cycling ratio was calculated as the sum of labeling in
glutamate C2 and C3, generated in the second and subsequent turns
of the TCA cycle, divided by labeling in C4 formed whenever
[2-13C]acetyl-CoA condenses with oxaloacetate. Thus, as this is a
relative value, where the labeling in the numerator as well as the
labeling in the denominator are proportional to enrichment in the
acetyl-CoA pool, the enrichment in the acetyl-CoA pool is
eliminated and the TCA-cycling ratio reflects activity in the TCA
cycle independently of enrichment in the acetyl-CoA pool.
StatisticsResults are presented as means ± standard error of the mean (SEM).
Statistically significant differences were determined using Student’s
t-test where indicated and otherwise one or two way ANOVA and pair
wise multiple comparison by the Holm-Sidak method. The level for
statistically significant differences was p < 0.05.
Results
Cellular content of aspartate and glutamateThe cellular content of the two neuroactive amino acidsglutamate and aspartate was determined in cell extracts ofcultured cerebellar granule neurons superfused with glucoseor b-hydroxybutyrate or with a combination of the twosubstrates (Table 1). Neurons receiving only glucose orglucose in combination with b-hydroxybutyrate had adecreased content of glutamate upon depolarization, butneurons receiving only b-hydroxybutyrate exhibited no
significant changes upon depolarization. The content ofaspartate was not affected by depolarization under any of thethree substrate conditions.
Under resting conditions neurons exposed to only b-hydroxybutyrate exhibited a cellular content of aspartatewhich was 10 nmol/mg protein higher than that observed inneurons exposed to only glucose, and the content ofglutamate was decreased correspondingly to a valuesignificantly lower than that found in neurons providedwith glucose. When applying a combination of the twosubstrates, the amount of glutamate again increased and theamount of aspartate was reduced to the level observed inneurons receiving only glucose. The differences observed,between the three different substrate combinations undercontrol conditions, were almost eliminated by repetitivedepolarization, in which case no difference was found inthe glutamate content. Furthermore, no difference wasfound in the aspartate content when comparing glucose andb-hydroxybutyrate, however, a lower content was foundusing a combination of the two substrates compared withb-hydroxybutyrate alone.
Since the two amino acids glutamate and aspartate areinterconverted very rapidly by transamination, the sum of thetwo was calculated to test whether the total pool stayedconstant under the different conditions. Indeed it did, nodifference was found for either the control or the depolarizedcondition among the three different substrates; glucose, b-hydroxybutyrate or the combination (Table 1). However,there was a significant decrease from control to depolarizedfor both glucose and the combination of the two substrates,whereas when employing b-hydroxybutyrate as substrate nosignificant decrease was found in the total pool of glutamateplus aspartate upon depolarization.
Table 1 Intracellular content of glutamate,
aspartate and the sum of the two amino
acids in cerebellar neurons after superfu-
sion with either 1 mM glucose, 1 mM b-
hydroxybutyrate or the combination of the
two substrates
Substrate Condition
Amino acid content (nmol/mg protein)
Glutamate Aspartate Glutamate + aspartate
Glucose Control 41.2 ± 1.9 8.0 ± 0.7 49.3 ± 1.8
Depolarized 29.9 ± 1.8* 9.0 ± 0.1 38.9 ± 1.7*
b-HOB Control 31.1 ± 1.7# 18.1 ± 2.6# 49.1 ± 3.1
Depolarized 26.8 ± 1.8 13.8 ± 3.1 40.6 ± 3.8
Glucose + b-HOB Control 44.9 ± 3.0¤ 4.6 ± 0.5¤ 49.6 ± 3.3
Depolarized 29.9 ± 1.1* 6.3 ± 0.3¤ 36.3 ± 1.1*
Cultured cerebellar neurons were superfused (see Materials and methods) with either 1 mM glu-
cose, 1 mM b-hydroxybutyrate (b-HOB) or the combination of the two and repetitively depolarized
with 300 lM NMDA, 10 lM glycine and 15 mM K+. The cell extracts were subsequently analyzed
with HPLC and cellular contents of glutamate and aspartate determined. Results are mean ± SEM,
with n = 4 cultures for each condition with all cultures from the same batch. All conditions were
repeated in other batches with similar results. Statistically significant differences between control
and repetitively depolarized cells are indicated with an asterisk and differences between substrates
are indicated as follows; #: between glucose and b-hydroxybutyrate, ¤: between b-hydroxybutyrate
and the combination of the two substrates, all determined by two way ANOVA and pair wise com-
parison by the Holm-Sidak method (p < 0.05).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91
b-hydroxybutyrate alters glutamate homeostasis | 83

Incorporation of 13C into glutamate and aspartatedetermined by 13C NMRTypical 13C NMR spectra obtained from cell extracts ofcerebellar neurons superfused with either [1,6-13C]glucose or
[2,4-13C]b-hydroxybutyrate are presented in Fig. 2. [Forexplanation of 13C-labeling and splitting patterns, seeMaterials and methods (Fig. 1) and Waagepetersen et al.(1998)]. The spectra were integrated and the intracellular 13Ccontents of glutamate (C4, C2 and C3), aspartate (C2 andC3) as well as b-hydroxybutyrate (C2 and C4) are shown inTable 2. Upon depolarization using glucose as substrate,significantly lower 13C contents were found in both thesinglet and the doublet of glutamate C4 and C2 as well as inthe doublet and doublet of doublets of glutamate C3. Incontrast, the singlets of aspartate C2 and C3 were notsignificantly changed upon depolarization. These singletswere the only quantifiable peaks of aspartate in the spectra,compatible with a low cellular content of aspartate in culturessuperfused in buffer containing only [1,6-13C]glucose.Depolarization resulted in similar changes for glutamatewhen utilizing [2,4-13C]b-hydroxybutyrate, in which case theC4 singlet and doublet, the C2 doublet as well as the C3doublet and doublet of doublets decreased. However, thedecrease was not as pronounced as that observed in neuronsreceiving [1,6-13C]glucose. When utilizing [2,4-13C]b-hydroxybutyrate, a complex coupling pattern was detectable
Table 2 Intracellular content 13C in glutamate, aspartate and b-hydroxybutyrate from cultured cerebellar neurons after superfusion with either
[1,6-13C]glucose or [2,4-13C]b-hydroxybutyrate during repeated depolarization and control conditions
Substrate Condition
Content of 13C-labeled isotopomer (nmol/mg protein)
Glu C4 Glu C2 Glu C3
[4-13C]
Singlet
[3,4-13C]
Doublet
[2-13C]
Singlet
[2,3-13C]
Doublet
[1,2,3-13C]
Doublet of
doublets
[3-13C]
Singlet
[2,3-13C];
[3,4-13C]
Doublet
[2,3,4-13C]
Doublet of
doublets
[1,6-13C]glucose Control 16.3 ± 1.0 12.8 ± 0.5 9.5 ± 0.4 7.2 ± 0.2 ND 1.9 ± 0.2 6.9 ± 0.2 6.7 ± 0.6
Depolarized 3.9 ± 0.4* 4.0 ± 0.3* 3.7 ± 0.4* 3.5 ± 0.4* ND 2.1 ± 0.4 2.6 ± 0.4* 2.5 ± 0.2*
[2,4-13C]b-HOB Control 7.7 ± 1.0� 9.5 ± 1.8 3.6 ± 0.4� 4.3 ± 0.5� 1.9 ± 0.4 1.1 ± 0.1� 4.7 ± 0.8� 5.1 ± 1.0
Depolarized 4.8 ± 0.4* 3.6 ± 0.3* 2.5 ± 0.3 2.1 ± 0.2*� 1.2 ± 0.1 1.3 ± 0.1 2.6 ± 0.3* 1.9 ± 0.2*
Asp C3 Asp C2 b-HOB C2 b-HOB C4
[3-13C]
Singlet
[2,3-13C]
Doublet
[1,2,3-13C]
Doublet of
doublets
[2-13C]
Singlet
[2,3-13C]
Doublet
[1,2,3-13C]
Doublet of
doublets
[2,4-13C]
Doublet
[2,4-13C]
Doublet
[1,6-13C]glucose Control 1.1 ± 0.1 ND ND 1.2 ± 0.1 ND ND ND ND
Depolarized 0.9 ± 0.1 ND ND 1.0 ± 0.1 ND ND ND ND
[2,4-13C]b-HOB Control 2.1 ± 0.3� 2.8 ± 0.4 1.4 ± 0.3 2.7 ± 0.5� 4.2 ± 0.7 1.9 ± 0.3 13.6 ± 1.9 13.8 ± 2.0
Depolarized 1.7 ± 0.2� 1.3 ± 0.2 ND 1.6 ± 0.2 2.7 ± 0.9 ND 19.9 ± 1.9 20.4 ± 1.6
Cultured cerebellar neurons were superfused (see Materials and methods) in the presence of either 1 mM [1,6-13C]glucose or 1 mM [2,4-13C]b-
hydroxybutyrate. The effect of repetitive neuronal depolarization using pulses (30 s) of 300 lM NMDA and 10 lM glycine in the presence of 15 mM
K+ was investigated using NMR spectroscopy on extracted cellular metabolites. Results are means ± SEM, n = 4 for each group; control or
depolarized with either [1,6-13C]glucose or [2,4-13C]b-hydroxybutyrate. Statistically significant differences (p < 0.05) between groups were
determined by two way ANOVA and pair wise comparison by the Holm-Sidak method. Differences between control and repetitively depolarized cells
are indicated by * and differences between cells superfused with either 1 mM [1,6-13C]glucose or 1 mM [2,4-13C]b-hydroxybutyrate by �. Glu,
glutamate; Asp, aspartate; b-HOB, b-hydroxybutyrate. ND, not detectable.
25 30 35 40 45 50 55 ppm
Glu C2
Asp C2 Asp C3
Glu C4
Glu C3
25 30 35 40 45 50 55 ppm
Glu C2
Asp C2
β-HOB C2
Asp C3
Glu C4
Glu C3
β-HOB C4
Glucose
β-Hydroxybutyrate
Lac C3
Fig. 2 13C NMR spectra of cell extracts from cultured cerebellar
neurons (for details, see Materials and methods) superfused with ei-
ther [2,4-13C]b-hydroxybutyrate or [1,6-13C]glucose for 74 min. Glu,
glutamate; Asp, aspartate; Lac, lactate; b-HOB, b-hydroxybutyrate.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91� 2009 The Authors
84 | T. M. Lund et al.

for aspartate C2 and C3, i.e. singlet, doublet and doublet ofdoublets, during the resting condition. Upon depolarization,the splitting pattern, i.e. doublet of doublets, which isobtained during extensive TCA-cycle metabolism incorpo-rating [2-13C]acetyl-CoA, disappeared leaving only a singletand a doublet for aspartate C2 and C3. Depolarizationresulted in a slightly although not significantly higher contentof [2,4-13C]b-hydroxybutyrate.
When utilizing [2,4-13C]b-hydroxybutyrate comparedwith [1,6-13C]glucose, a significantly lower content of 13Cwas found in the C2, C3 and C4 singlets as well as the C2and C3 doublets of glutamate in the control situation and inthe C2 doublet of glutamate in the depolarizing condition.However, significantly higher contents of 13C were found inaspartate C2 and C3 singlets under control as well as in theC3 singlet under depolarizing conditions using [2,4-13C]b-hydroxybutyrate compared with [1,6-13C]glucose.
TCA-cycle metabolismFor evaluation of TCA-cycle metabolism, a cycling ratiowas calculated from the labeling patterns in glutamate asdetermined by 13C NMR (see Materials and methods forcalculations). The values for the two substrates duringresting and depolarizing conditions are presented inTable 3. Depolarization induced an approximately 60%increase in TCA-cycling ratio using [1,6-13C]glucose,whereas the TCA-cycling ratio was unaffected when[2,4-13C]b-hydroxybutyrate was the substrate. Furthermore,the cycling ratio in the depolarized condition employing[2,4-13C]b-hydroxybutyrate was significantly lower thanwhen using [1,6-13C]glucose.
Molecular 13C-labeling in glutamate and aspartatedetermined by LC-MSTo obtain more detailed information about the relativeimportance of glucose and b-hydroxybutyrate for maintain-ing aspartate–glutamate homeostasis, a series of experimentswere performed in which either [1,6-13C]glucose or[2,4-13C]b-hydroxybutyrate was used alone or in combina-tion with its unlabeled counterpart. Using mass spectrometry,the MCL was determined for glutamate and aspartate in cellextracts of cerebellar neurons during resting or depolarizingconditions. As shown in Fig. 3a labeling in glutamatedecreased significantly, whereas no significant change wasfound in aspartate upon depolarization in the presence of[1,6-13C]glucose only. However, when only [2,4-13C]b-hydroxybutyrate was present a significantly lower incorpo-ration into aspartate was found upon depolarization(Fig. 3b). Comparing the MCL for aspartate under restingconditions for the two different substrates it was significantlyhigher (p = 0.01), i.e. 44%, using [2,4-13C]b-hydroxybuty-rate compared with 28% using [1,6-13C]glucose. When using1 mM [1,6-13C]glucose and 1 mM unlabeled b-hydroxy-butyrate no significant change was found for either glutamateor aspartate upon depolarization (Fig. 3c), whereas adecrease in labeling was found in both amino acids whenb-hydroxybutyrate was the labeled compound in combina-tion with unlabeled glucose (Fig. 3d). A dilution of thelabeled acetyl-CoA pool and thus a lower MCL would beexpected using a combination of unlabeled and labeledsubstrate as observed comparing results obtained by utiliza-tion of labeled glucose with and without unlabeledb-hydroxybutyrate (Fig. 3a and c). Surprisingly, no reduc-tion was found in the MCL for glutamate under neitherresting nor depolarizing conditions when comparing thesituation with [2,4-13C]b-hydroxybutyrate alone (Fig. 3b)with that where labeled b-hydroxybutyrate was used togetherwith unlabeled glucose (Fig. 3d). However, the additionalpresence of glucose caused a shift in the labeling fromaspartate to glutamate, as aspartate had a significantly higherlabeling in the control situation when b-hydroxybutyrate wasused alone (aspartate 44.0 ± 2.8%; glutamate 28.3 ± 3.5%,p = 0.01), whereas glutamate had the highest labeling whena combination of the two substrates was used (aspartate28.3 ± 1.9%; glutamate 33.4 ± 1.0%, p = 0.04) (Fig. 3b andd). In the two experiments with either 1 mM labeled glucoseand 1 mM unlabeled b-hydroxybutyrate (Fig. 3c) or viceversa (Fig. 3d), the cultures had been exposed to exactly thesame conditions, only the position of 13C label beingchanged from glucose to b-hydroxybutyrate. 13C from[2,4-13C]b-hydroxybutyrate (Fig. 3d) was more readilyincorporated into glutamate and aspartate under both controland depolarizing conditions than 13C-labeling from[1,6-13C]glucose (Fig. 3c) when both substrates were presentconcurrently (p < 0.05 for all four comparisons). However,depolarization caused a significantly lower incorporation of
Table 3 TCA-cycling ratio determined from the 13C-labeling in gluta-
mate
Substrate Condition TCA-cycling ratioGlu(C2þC3)
Glu(C4)
[1,6-13C]glucose Control 1.12 ± 0.03
Depolarized 1.81 ± 0.12*
[2,4-13C]b-HOB Control 1.21 ± 0.04
Depolarized 1.37 ± 0.02�
Cultured cerebellar neurons were superfused (see Materials and
methods) with either 1 mM [1,6-13C]glucose or 1 mM [2,4-13C]b-
hydroxybutyrate (b-HOB) and repetitively depolarized with 300 lM
NMDA, 10 lM glycine and 15 mM K+. The cell extracts were subse-
quently analyzed with NMR and the cycling ratio was calculated from
the glutamate peaks in the spectrum. The content of 13C labeling in
glutamate from the second turn and onwards was divided by labeling
in C4 originating from the first turn. Results are means of cycling ratio
± SEM, n = 4 for each condition. Statistically significant difference
(p < 0.05) between control and repetitively depolarized cells is indi-
cated with an asterisk and difference between [1,6-13C]glucose and
[2,4-13C]b-hydroxybutyrate is indicated by �, as determined by two
way ANOVA and pair wise comparison by the Holm-Sidak method.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91
b-hydroxybutyrate alters glutamate homeostasis | 85

13C from [2,4-13C]b-hydroxybutyrate into both glutamateand aspartate, whereas incorporation of 13C coming from[1,6-13C]glucose seemed to increase, although this was notsignificant. Altogether this gives a relative shift in thepreference of substrate towards glucose upon depolarization.Still having in mind that the cultures were exposed to thesame substrates in the two experiments (Fig. 3c and d), thisshift can be further emphasized by calculating the relativecontribution of 13C in glutamate or aspartate (% MCL)coming from [1,6-13C]glucose compared with the sum of13C incorporation coming from both [1,6-13C]glucose and[2,4-13C]b-hydroxybutyrate, e.g. (7.2/(7.2 + 33.42)) · 100% =
17.8% for glutamate in the control situation. Thus, upondepolarization a shift was seen in the preference of substratetowards glucose as the relative contribution of 13C-labelingfrom [1,6-13C]glucose in glutamate increased from17.8 ± 0.9 to 31.8 ± 1.4% (Student’s t-test, p < 0.001) andin aspartate from 17.6 ± 1.0 to 36.6 ± 4.4% (Student’s t-test,p = 0.002).
Molecular distribution of 13C in glutamate and aspartatefrom [2,4-13C]b-hydroxybutyrateTo further elucidate b-hydroxybutyrate metabolism and itsimpact on aspartate–glutamate homeostasis a detailed map-
Glutamate
0
10
20
30
40
50 (a) (c)
(d) (b)
M + 1 M + 2 M + 3
M + 1 M + 2 M + 3 M + 1 M + 2 M + 3
M + 4 M + 1 M + 2 M + 3 M + 4
Lab
elin
g (
%)
Lab
elin
g (
%)
Lab
elin
g (
%)
Lab
elin
g (
%)
*
Glutamate
0
10
20
30
40
50
* *
*
*
*
*
§ §
§
Aspartate
0
10
20
30
40
50
* *
*
Aspartate
0
10
20
30
40
50
*
*
§ §
§ §
[2,4 -13C]β-hydroxybutyrate [2,4 -13C]β-hydroxybutyrate + glucose
Fig. 4 Percentage 13C-labeling in each isotopomer, monolabeling
(M + 1), double-labeling (M + 2) etc. in glutamate and aspartate in cell
extracts of cerebellar granule cells (for details, see Materials and
methods) superfused with either 1 mM [2,4-13C]b-hydroxybutyrate (a
and b) or 1 mM [2,4-13C]b-hydroxybutyrate plus 1 mM unlabeled
glucose (c and d) under resting (bars in grey) and depolarizing con-
ditions (bars in black). Bars indicate mean ± SEM and the number
of individual cultures was 6–8 originating from 2–3 different experi-
ments. *Significantly different from the corresponding resting condi-
tion, §significantly different from the same isotopomer without glucose
(i.e. a versus c; b versus d) (p < 0.05) as determined by two way ANOVA
and pair wise multiple comparison by the Holm-Sidak method.
[1,6-13C]glucose
0 10 20 30 40 50
Glutamate Aspartate M
CL
(%
)
*
[2,4-13C]β-hydroxybutyrate
0 10 20 30 40 50
Glutamate Aspartate
MC
L (
%) *
[1,6-13C]glucose + β-hydroxybutyrate
0 10 20 30 40 50
Glutamate Aspartate
MC
L (
%)
§ § § §
[2,4-13C]β-hydroxybutyrate + glucose
0 10 20 30 40 50
Glutamate Aspartate
MC
L (
%) *
*
§ §
(a)
(c)
(b)
(d)
Fig. 3 Molecular carbon labeling (% MCL) of glutamate and aspartate in
cell extracts of cerebellar granule cells (for details, see Materials and
methods) superfused with 1 mM [1,6-13C]glucose (a), 1 mM [2,4-13C]b-
hydroxybutyrate (b), 1 mM [1,6-13C]glucose and 1 mM b-hydroxybuty-
rate (c) and 1 mM [2,4-13C]b-hydroxybutyrate and 1 mM glucose (d)
under resting (bars in grey) and depolarizing conditions (bars in black).
Bars indicate mean ± SEM and the number of individual cultures was 6–
8 originating from two to three different experiments. *Significantly dif-
ferent from the corresponding resting condition, §significantly different
from the same condition without unlabeled counterpart (i.e. c versus a; d
versus b) (p < 0.05) as determined by two way ANOVA and pair wise
multiple comparison by the Holm-Sidak method.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91� 2009 The Authors
86 | T. M. Lund et al.

ping of the labeling patterns for glutamate and aspartate wasperformed (Fig. 4). Employing [2,4-13C]b-hydroxybutyrateas substrate a significantly higher labeling was found inM + 1 for both glutamate and aspartate, whereas M + 2 andM + 3 decreased for aspartate upon depolarization (Fig. 4aand 4b). When [2,4-13C]b-hydroxybutyrate was combinedwith unlabeled glucose, M + 1 increased for glutamate upondepolarization, whereas M + 2, M + 3 and M + 4 decreased(Fig. 4c), as also observed for M + 2 and M + 3 of aspartate(Fig. 4d). An augmented labeling in M + 1 and a lowerlabeling in M + 2, M + 3 and M + 4 is compatible with adilution of labeling in the acetyl-CoA pool. Labeling inglutamate increased by adding unlabeled glucose for M + 2and M + 3 in the control situation as well as for M + 2 in thedepolarized situation (Figs 4a and 4c). As opposed to this,labeling in aspartate was reduced for M + 3 in the controland for all isotopomers in the depolarized condition whenunlabeled glucose was added. (Fig. 4d compared with 4b).Thus, the impact of the presence of glucose in combinationwith [2,4-13C]b-hydroxybutyrate was a higher labeling inglutamate and a reduced labeling in aspartate.
Discussion
Glucose versus b-hydroxybutyrate as energy substratesGlucose is the primary energy substrate for the brain; but inthe brain of suckling rats and neonate humans, ketone bodiesare used as precursors for synthesis of amino acids and lipids(Morris 2005). However, most patients with inborn errors ofketogenesis develop normally, suggesting that the role forketone bodies is as an alternative energy substrate to glucoseduring development and also during diseases or prolongedfasting (Morris 2005). Thus, a ketogenic diet can be used totreat patients with GLUT1 deficiency syndrome, whichcauses an impaired glucose transport into the brain. In thesepatients, ketone bodies compensate for the low brain glucoseconcentration and are used as an alternative fuel, whicheffectively restores brain energy metabolism (Klepper et al.2005). A ketogenic diet has also shown remarkable effects inthe treatment of drug-resistant childhood epilepsy (Gasioret al. 2006; Bough and Rho 2007; Yellen 2008) and in ananimal model of traumatic brain injury infusion of b-hydroxybutyrate 1 h after injury led to a significant decreasein edema (Scafidi et al. 2009). However, underlying mech-anisms need still to be further explored (Yudkoff et al. 2007).
In the present study, we tried to elucidate a part of thismechanism by studying the metabolic changes at the cellularlevel when changing energy substrate from glucose to b-hydroxybutyrate or by using combinations of the twosubstrates. We used cultured glutamatergic neurons andinvestigated the effect of inducing neuronal activity, which isknown to stimulate neuronal oxidative metabolism (Baket al. 2006).
Effect of depolarization on oxidative metabolismDuring neuronal activity, sodium ions enter and potassiumions leave the cell, changing the membrane potential. As thesodium–potassium-ATPase restores the potential at theexpense of energy, increased metabolic activity is expectedupon re-polarization (Clarke and Sokoloff 1999). In thepresent study, a higher TCA-cycling ratio was found upondepolarization monitored by the use of [1,6-13C]glucose(Table 3), indicating increased oxidative metabolism. This issupported by an increased CO2 production during depolar-ization (Peng and Hertz 1993).
It could be speculated that the increased oxidation mayinvolve glutamate as anaplerotic substrate, as we found adecrease in the glutamate content (Table 1). However,addition of unlabeled substrate, such as glutamate, into thepool of TCA-cycle intermediates via a-ketoglutarate haspreviously been shown to reduce the TCA-cycling ratio(Waagepetersen et al. 1998). In contrast, the TCA-cyclingratio calculated on the basis of 13C incorporation from b-hydroxybutyrate was unaltered upon depolarization. Thisfinding is in line with a study in conscious rats by Cruzet al. (2005) showing that tracer amounts of [14C]hydroxy-butyrate do not register an increase in energy metabolismduring acoustic activation, as opposed to increases inglucose and acetate utilization. Moreover, in the presentstudy, the MCL, i.e. total labeling, of aspartate wasmarkedly decreased indicating a reduced metabolism ofb-hydroxybutyrate in neurons exposed to depolarizingconditions (Fig. 3b). This was further emphasized by thedetailed labeling patterns of glutamate and aspartateobtained by MS, indicating a decreased enrichment in theacetyl-CoA pool upon depolarization (Fig. 4a and b). Theintracellular concentration of calcium increases in depolar-ized neurons, which in turn activates the mitochondrialCa2+-uniporter (Kirichok et al. 2004). An augmented intra-mitochondrial Ca2+ concentration activates the pyruvate,isocitrate, and a-ketoglutarate dehydrogenases, potentiallygiving rise to an elevated mitochondrial NADH/NAD+ ratio(Nichols and Denton 1995; Pardo et al. 2006). The reducedcatabolism of b-hydroxybutyrate may be caused by aninhibition of b-hydroxybutyrate dehydrogenase by theincreased level of NADH and it may be compatible withthe higher cellular content of b-hydroxybutyrate found indepolarized neurons, although this was not statisticallysignificant. However, it seems in contrast to the finding byMaalouf et al. (2007) that ketone bodies increase NADHoxidation in isolated mitochondria. This apparent discrep-ancy may be explained by Maalouf et al. (2007) using acombination of b-hydroxybutyrate and acetoacetate clearlyeliminating any involvement of the b-hydroxybutyratedehydrogenase. Moreover, the effect observed in thepresent study is likely induced by increased mitochondrial[Ca2+], a condition which was not tested by Maalouf et al.(2007).
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91
b-hydroxybutyrate alters glutamate homeostasis | 87

Although 13C from b-hydroxybutyrate was more readilyincorporated into glutamate and aspartate than from 13Cglucose when a combination of the two substrates were used,we found that depolarization shifted the preference slightlytowards glucose, as an increased glucose and a decreased b-hydroxybutyrate metabolism was found during depolarizingconditions. This may support the conclusion by Bak et al.(2006) using an analogous preparation of glutamatergicneurons that glucose is necessary to maintain neurotrans-mitter homeostasis during synaptic activity.
Interaction between glucose and b-hydroxybutyratemetabolismThe present study showed an interesting interaction betweenglucose and b-hydroxybutyrate metabolism when usingcombinations of the two substrates, where one of them was13C-labeled and the other unlabeled. The MCL in glutamateand aspartate, originating from [1,6-13C]glucose, was con-siderably reduced when also unlabeled b-hydroxybutyratewas present (Fig. 3a and c). This could be a consequence ofdilution caused by an increase in the pool of unlabeledacetyl-CoA. However, using [2,4-13C]b-hydroxybutyrate,only a slight reduction was observed in the labeling ofaspartate and that of glutamate was unaffected by addition ofunlabeled glucose (Fig. 3b and d); the latter finding arguesagainst a simple dilution effect in the acetyl-CoA pool.Similar results were obtained in synaptic terminals fromweanling rats, where unlabeled glucose did not decrease the14CO2 production from 14C-labeled b-hydroxybutyrate,whereas 14CO2 production from 14C-labeled glucose wasdecreased by b-hydroxybutyrate (McKenna et al. 1994).This observation may be explained by an inhibition ofenzymes in glucose metabolism prior to acetyl-CoA forma-tion or a competition between pyruvate and b-hydroxybuty-rate for entry into the mitochondria. In favor of the firstsuggestion, b-hydroxybutyrate has been shown to inhibit theoxidative decarboxylation of pyruvate in rat brain mitochon-dria (Booth and Clark 1981), and in vivo studies have shownthat i.v. infusion of b-hydroxybutyrate caused a markedlyincreased lactate production in the brain, indicating inhibitionof pyruvate oxidation (Ruderman et al. 1974; Nehlig 2004).A likely mechanism involves allosteric feedback inhibitionby acetyl-CoA of the pyruvate dehydrogenase complex assuggested by Nehlig (2004). Whether a competition betweenb-hydroxybutyrate and pyruvate for entry into mitochondriavia MCT takes place remains uncertain, as little is knownabout the distribution and characteristics of MCT in the innermitochondrial membrane.
The aspartate–glutamate homeostasisThe brain has a well-regulated interplay between excitationand inhibition of neurons. Excessive excitatory activity and/or low inhibitory activity is thought to lead to seizures, whichis consistent with the repeated observation that glutamate
receptor agonists or GABA receptor antagonists generateseizures in animal models (Croucher and Bradford 1989;Kondziella et al. 2002). As opposed to pharmacologicalcorrection of epileptic pathology, nutrition-based generationof ketone bodies - as in the ketogenic diet – may seek toestablish excitatory-inhibitory balance via an alteration ofneuronal intermediary metabolism (Yudkoff et al. 2007).Relevant to this, evidence is presented that the aspartate–glutamate homeostasis in glutamatergic neurons was affecteddifferently by b-hydroxybutyrate or glucose.
Glucose and b-hydroxybutyrate are metabolized in differ-ent pathways and cellular compartments which may affectaspartate–glutamate homeostasis. In the presence of b-hydroxybutyrate, the aspartate content was higher and thatof glutamate lower, than in the presence of glucose. Duringglycolysis, MAS is active, transporting reducing equivalentsto mitochondria (Fig. 5a), whereas b-hydroxybutyrate ismetabolized to acetyl-CoA directly in the mitochondria(Fig. 5b). In the latter case, the activity of cytosolic malatedehydrogenase reducing oxaloacetate to malate may bediminished, and consequently the exchange of malate and a-ketoglutarate via the ketodicarboxylate carrier of the MASmay be decreased (Fig. 5b). Such scenario may potentiallylead to more of the mitochondrial a-ketoglutarate enteringthe TCA cycle, and thus enhancing the formation ofoxaloacetate via TCA-cycle reactions and subsequentlyaspartate via aspartate aminotransferase; a pathway describedby Hertz et al. (1992) as the truncated TCA cycle. The factthat b-hydroxybutyrate metabolism consumes succinyl-CoAin the step from acetoacetate to acetoacetyl-CoA may furtheraccelerate the truncated TCA cycle. Depending upon thecontinued uptake of glutamate into the mitochondria eithervia the glutamate-hydroxyl-carrier or the aspartate–glutamateexchanger, the high activity of the truncated TCA cycle maylead to a limited accessibility of a-ketoglutarate in thecytosol for synthesis of glutamate. Interestingly, the biosyn-thesis of particularly the transmitter pool of glutamate hasbeen shown to be dependent upon the operation of theketodicarboxylate carrier (Palaiologos et al. 1988). More-over, an enlarged pool of aspartate was found in cerebellargranule cells exposed to phenylsuccinate, an inhibitor of theketodicarboxylate carrier (Passarella et al. 1987; Palaiologoset al. 1988). The MCL of aspartate was considerably higherunder resting conditions, using [2,4-13C]b-hydroxybutyratecompared with using [1,6-13C]glucose. This supports thenotion that the increased cellular aspartate content observedin neurons receiving only [2,4-13C]b-hydroxybutyrate isgenerated via the truncated TCA cycle. A similar shift in thecontent of glutamate and aspartate has been found both in ratbrain synaptosomes and in cerebellar granule cells metab-olizing glutamine when changing from a condition withglucose to a condition without glucose in which theproduction of cytosolic NADH and MAS activity is limited(Yudkoff et al. 1994; Peng et al. 2007). However, in these
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91� 2009 The Authors
88 | T. M. Lund et al.

studies the lack of glucose is compensated for by glutamineas an anaplerotic substrate and probably pyruvate recyclingto form acetyl-CoA (Olstad et al. 2007), whereas in ourstudies using b-hydroxybutyrate acetyl-CoA is not a limitingfactor.
In line with the hypothesis presented above re-establish-ment of the activities of cytosolic malate dehydrogenase andthe ketodicarboxylate carrier was expected when cerebellarneurons were superfused in the combined presence ofglucose and b-hydroxybutyrate and, indeed, the aspartate–glutamate homeostasis was restored (Table 1). The additionalpresence of glucose caused a shift in the labeling fromaspartate to glutamate, as aspartate had a significantly higherlabeling when b-hydroxybutyrate was used alone, whereasglutamate had the highest labeling when a combination of thetwo substrates was used (comparing Fig. 3b with 3d andemphasized in detail in Fig. 4). Thus, as discussed earlier, theaddition of glucose activated the MAS, reconstitutingtransport of a-ketoglutarate out of the mitochondria, whichsupports subsequent formation of transmitter glutamate viatransamination at the expense of aspartate formation. Inagreement with this, the energy substrate available, i.e.glucose, b-hydroxybutyrate or both, was found to have animpact on how cultured granule neurons responded to therepetitive exposure to NMDA leading to vesicular release of
the neurotransmitter pool. In the presence of glucose, eitheralone or in combination with b-hydroxybutyrate the gluta-mate content as well as the total pool of glutamate plusaspartate decreased significantly upon activation of neuro-transmitter release (Table 1). In contrast to this no significantdecrease was observed in the presence of only b-hydroxy-butyrate, neither in the glutamate content nor in the total poolof glutamate plus aspartate (Table 1). As mentioned abovethe ketodicarboxylate carrier, an obligatory entity of theMAS, has been shown to be functionally involved in thesynthesis of transmitter glutamate (Palaiologos et al. 1988),which might explain a diminished pool of transmitterglutamate and a subsequent limited vesicular release inneurons receiving b-hydroxybutyrate as the only substrate.
Conclusion
Cultured glutamatergic neurons metabolized b-hydroxybu-tyrate more readily than glucose, when both substrates wereavailable. This was evident during resting conditions as wellas during induced neuronal activity, although glucosemetabolism increased while that of b-hydroxybutyratedecreased due to depolarization. Interestingly, neuronsmetabolizing b-hydroxybutyrate as energy substrate, had analtered aspartate–glutamate homeostasis, with an increased
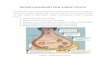
Fig. 5 The activity of the individual processes involved in the malate–
aspartate shuttle is affected under conditions when glucose is present
(a) and when only b-hydroxybutyrate is present (b). In the latter case, no
reducing equivalents are formed in glycolysis and the activity of the
cytosolic malate dehydrogenase and subsequently the ketodicarboxy-
late carrier may decrease. This may result in an increased flow of in-
tramitochondrial a-ketoglutarate into the TCA cycle and thus enhancing
the formation of oxaloacetate, the latter being transaminated to aspar-
tate. Such scenario might explain an increased intracellular content of
aspartate in neurons receiving b-hydroxybutyrate only. The shift from
glutamate to aspartate via the truncated TCA cycle is indicated with bold
arrows. AcAc, acetoacetate; AcAc-CoA, acetoacetyl-CoA; Ac-CoA,
acetyl-CoA; Asp, aspartate; cAAT, cytosolic aspartate aminotransfer-
ease; Cit, citrate; ETC, electron transport chain; Glu, glutamate; b-HOB,
b-hydroxybutyrate; a-KG, a-ketoglutarate; mATT, mitochondrial
aspartate aminotransferease; OAA, oxaloacetate; Suc, succinate; Suc-
CoA, succinyl-CoA. Note that the equations are not stoichiometric as
acetoacetyl-CoA reacts with CoA and yields two acetyl-CoA.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91
b-hydroxybutyrate alters glutamate homeostasis | 89

aspartate and a correspondingly decreased glutamate contentcompared with those metabolizing glucose. We suggest thatthese changes are caused by a decreased activity of the MASin neurons metabolizing b-hydroxybutyrate; a malfunction-ing MAS has previously been shown to limit the amountof transmitter glutamate available for vesicular release(Palaiologos et al. 1988). Thus, our observation supportsthe hypothesis of reduced excitatory transmission playing arole in the anti-convulsive effect of the ketogenic diet (Patelet al. 1988). In patients treated with the ketogenic diet, thebrain has access to both glucose and b-hydroxybutyrate, acondition in which the current experiments were unable toshow any significant reduction in neuronal glutamate con-tent. However, a shift towards ketone based energy metab-olism may have a capacity to tune down the overallexcitatory capacity of glutamatergic neurons potentiallyleading to an anti-convulsive effect.
Acknowledgments
Ms. Lene Vigh, Mette Clausen and Johanne Kroon Hansen are
cordially acknowledged for expert technical support. This study was
supported by grants from The Danish State Medical Research
Council (271-07-0262), the Horslev, Alfred Benzon and Lundbeck
Foundations.
References
BakL.K., SchousboeA. andWaagepetersenH. S. (2003)Characterizationof depolarization-coupled release of glutamate from cultured mousecerebellar granule cells using DL-threo-beta-benzyloxyaspartate(DL-TBOA) to distinguish between the vesicular and cytoplasmicpools. Neurochem. Int. 43, 417–424.
Bak L. K., Schousboe A., Sonnewald U. and Waagepetersen H. S.(2006) Glucose is necessary to maintain neurotransmitter homeo-stasis during synaptic activity in cultured glutamatergic neurons.J. Cereb. Blood Flow Metab. 26, 1285–1297.
Biemann K. (1962) The mass spectra of isotopically labeled molecules,in Mass Spectrometry; Organic Chemical Applications, pp. 223–227. McGraw-Hill, New York.
Booth R. F. and Clark J. B. (1981) Energy metabolism in rat brain:inhibition of pyruvate decarboxylation by 3-hydroxybutyrate inneonatal mitochondria. J. Neurochem. 37, 179–185.
Bough K. J. and Rho J. M. (2007) Anticonvulsant mechanisms of theketogenic diet. Epilepsia 48, 43–58.
Clarke D. D. and Sokoloff L. (1999)Circulation and energy metabolismof the brain, in Basic Neurochemistry (Siegel G. J., Agranoff B. W.,Albers R. W., Fisher S. K. and Uhler M. D., eds), pp. 637–669.Lippincott Williams and Wilkins, Philadelphia.
Croucher M. J. and Bradford H. F. (1989) Kindling of full limbic sei-zures by repeated microinjections of excitatory amino acids intothe rat amygdala. Brain Res. 501, 58–65.
Cruz N. F., Lasater A., Zielke H. R. and Dienel G. A. (2005) Activationof astrocytes in brain of conscious rats during acoustic stimulation:acetate utilization in working brain. J. Neurochem. 92, 934–947.
Daikhin Y. and Yudkoff M. (1998) Ketone bodies and brain glutamateand GABA metabolism. Dev. Neurosci. 20, 358–364.
Dalsgaard M. K. (2006) Fuelling cerebral activity in exercising man.J. Cereb. Blood Flow Metab. 26, 731–750.
Drejer J. and Schousboe A. (1989) Selection of a pure cerebellar granulecell culture by kainate treatment. Neurochem. Res. 14, 751–754.
Drejer J., Larsson O. M. and Schousboe A. (1982) Characterization of L-glutamate uptake into and release from astrocytes and neuronscultured from different brain regions. Exp. Brain Res. 47, 259–269.
Drejer J., Larsson O. M., Kvamme E., Svenneby G., Hertz L. andSchousboe A. (1985) Ontogenetic development of glutamatemetabolizing enzymes in cultured cerebellar granule cells and incerebellum in vivo. Neurochem. Res. 10, 49–62.
Drejer J., Honore T. and Schousboe A. (1987) Excitatory amino acid-induced release of 3H-GABA from cultured mouse cerebral cortexinterneurons. J. Neurosci. 7, 2910–2916.
Erecinska M., Nelson D., Daikhin Y. and Yudkoff M. (1996) Regulationof GABA level in rat brain synaptosomes: fluxes through enzymesof the GABA shunt and effects of glutamate, calcium, and ketonebodies. J. Neurochem. 67, 2325–2334.
Gasior M., Rogawski M. A. and Hartman A. L. (2006) Neuroprotectiveand disease-modifying effects of the ketogenic diet. Behav. Phar-macol. 17, 431–439.
Geddes J. W. and Wood J. D. (1984) Changes in the amino acid contentof nerve endings (synaptosomes) induced by drugs that alter themetabolism of glutamate and gamma-aminobutyric acid. J. Neu-rochem. 42, 16–24.
Hawkins R. A., Williamson D. H. and Krebs H. A. (1971) Ketone-bodyutilization by adult and suckling rat brain in vivo. Biochem. J. 122,13–18.
Hertz L., Peng L., Westergaard N., Yudkoff M. and Schousboe A. (1992)Neuronal-astrocytic interactions in metaolism of transmitter aminoacids of the glutamate family, in Drug Research Related to Neu-roactive Amino Acids (Schousboe A., Diemer N. H. and Kofod H.,eds),Vol. 32, pp. 30–48. Munksgaard, Copenhagen.
Kirichok Y., Krapivinsky G. and Clapham D. E. (2004) The mitochon-drial calcium uniporter is a highly selective ion channel. Nature427, 360–364.
Klepper J., Scheffer H., Leiendecker B., Gertsen E., Binder S., LeferinkM., Hertzberg C., Nake A., Voit T. and Willemsen M. A. (2005)Seizure control and acceptance of the ketogenic diet in GLUT1deficiency syndrome: a 2- to 5-year follow-up of 15 children en-rolled prospectively. Neuropediatrics 36, 302–308.
Kondziella D., Bidar A., Urfjell B., Sletvold O. and Sonnewald U.(2002) The pentylenetetrazole-kindling model of epilepsy inSAMP8 mice: behavior and metabolism. Neurochem. Int. 40, 413–418.
Lopes-Cardozo M., Larsson O. M. and Schousboe A. (1986) Acetoac-etate and glucose as lipid precursors and energy substrates in pri-mary cultures of astrocytes and neurons from mouse cerebralcortex. J. Neurochem. 46, 773–778.
Lowry O. H., Rosebrough N. J., Farr A. L. and Randall A.J. (1951)Protein measurement with the Folin phenol reagent. J. Biol. Chem.193, 265–275.
Ma W., Berg J. and Yellen G. (2007) Ketogenic diet metabolites reducefiring in central neurons by opening K(ATP) channels. J. Neurosci.27, 3618–3625.
Maalouf M., Sullivan P. G., Davis L., Kim D. Y. and Rho J. M. (2007)Ketones inhibit mitochondrial production of reactive oxygen spe-cies production following glutamate excitotoxicity by increasingNADH oxidation. Neuroscience 145, 256–264.
McKenna M. C., Tildon J. T., Stevenson J. H. and Hopkins I. B. (1994)Energy metabolism in cortical synaptic terminals from weanlingand mature rat brain: evidence for multiple compartments of tri-carboxylic acid cycle activity. Dev. Neurosci. 16, 291–300.
Melo T. M., Nehlig A. and Sonnewald U. (2006) Neuronal-glialinteractions in rats fed a ketogenic diet. Neurochem. Int. 48, 498–507.
Journal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91� 2009 The Authors
90 | T. M. Lund et al.

Morris A. A. (2005) Cerebral ketone body metabolism. J. Inherit. Metab.Dis. 28, 109–121.
Nehlig A. (2004) Brain uptake and metabolism of ketone bodies in animalmodels. Prostaglandins Leukot. Essent. Fatty Acids 70, 265–275.
Nichols B. J. and Denton R. M. (1995) Towards the molecular basis forthe regulation of mitochondrial dehydrogenases by calcium ions.Mol. Cell. Biochem. 149-150, 203–212.
Olstad E., Olsen G. M., Qu H. and Sonnewald U. (2007) Pyruvaterecycling in cultured neurons from cerebellum. J. Neurosci. Res.85, 3318–3325.
Owen O. E., Morgan A. P., Kemp H. G., Sullivan J. M., Herrera M. G.and Cahill G. F. Jr (1967) Brain metabolism during fasting. J. Clin.Invest. 46, 1589–1595.
Palaiologos G., Hertz L. and Schousboe A. (1988) Evidence thataspartate aminotransferase activity and ketodicarboxylate carrierfunction are essential for biosynthesis of transmitter glutamate.J. Neurochem. 51, 317–320.
Pardo B., Contreras L., Serrano A., Ramos M., Kobayashi K., Iijima M.,Saheki T. and Satrustegui J. (2006) Essential role of aralar in thetransduction of small Ca2+ signals to neuronal mitochondria.J. Biol. Chem. 281, 1039–1047.
Passarella S., Atlante A., Barile M. and Quagliariello E. (1987) Aniontransport in rat brain mitochondria: fumarate uptake via the di-carboxylate carrier. Neurochem. Res. 12, 255–264.
Patel S., Chapman A. G., Millan M. H. and Meldrum B. S. (1988)Epilepsy and excitatory amino acid antagonists, in ExcitatoryAmino Acids in Health and Disease (Lodge D., ed), pp. 353–378.John Wiley and sons, London.
Peng L. and Hertz L. (1993) Potassium-induced stimulation of oxidativemetabolism of glucose in cultures of intact cerebellar granule cellsbut not in corresponding cells with dendritic degeneration. BrainRes. 629, 331–334.
Peng L., Gu L., Zhang H., Huang X., Hertz E. and Hertz L. (2007)Glutamine as an energy substrate in cultured neurons during glu-cose deprivation. J. Neurosci. Res. 85, 3480–3486.
Ruderman N. B., Ross P. S., Berger M. and Goodman M. N. (1974)Regulation of glucose and ketone-body metabolism in brain ofanaesthetized rats. Biochem. J. 138, 1–10.
Scafidi S. U., Racz J. R., Bamford P., Shi D., Fiskum G. and McKennaM. C. (2009) Metabolism of 3-b-hydroxybutyrate in brain in brainafter traumatic brain injury (TBI) in young rats, J. Neurochem.108(Suppl. 1), pp 156.
Schousboe A., Meier E., Drejer J. and Hertz L. (1989) Preparation ofprimary cultures of mouse (rat) cerebellar granule cells, in ADissection and Tissue Culture Manual for the Nervous System(Shahar A., de Vellis J., Vernadakis A. and Harber B., eds), pp.183–186. Liss, New York.
Schurr A., West C. A. and Rigor B. M. (1988) Lactate-supported syn-aptic function in the rat hippocampal slice preparation. Science240, 1326–1328.
Schurr A., Payne R. S., Miller J. J. and Rigor B. M. (1997) Brain lactateis an obligatory aerobic energy substrate for functional recoveryafter hypoxia: further in vitro validation. J. Neurochem. 69, 423–426.
Sonnewald U., Olstad E., Qu H., Babot Z., Cristofol R., Sunol C.,Schousboe A. and Waagepetersen H. (2004) First direct demon-stration of extensive GABA synthesis in mouse cerebellar neuronalcultures. J. Neurochem. 91, 796–803.
Waagepetersen H. S., Bakken I. J., Larsson O. M., Sonnewald U. andSchousboe A. (1998) Metabolism of lactate in cultured GABAer-gic neurons studied by 13C nuclear magnetic resonance spectros-copy. J. Cereb. Blood Flow Metab. 18, 109–117.
Yellen G. (2008) Ketone bodies, glycolysis, and KATP channels inthe mechanism of the ketogenic diet. Epilepsia 49(Suppl. 8), 80–82.
Yudkoff M., Nelson D., Daikhin Y. and Erecinska M. (1994) Tricar-boxylic acid cycle in rat brain synaptosomes. Fluxes and interac-tions with aspartate aminotransferase and malate/aspartate shuttle.J. Biol. Chem. 269, 27414–27420.
Yudkoff M., Daikhin Y., Nissim I., Lazarow A. and Nissim I. (2001)Ketogenic diet, amino acid metabolism, and seizure control.J. Neurosci. Res. 66, 931–940.
Yudkoff M., Daikhin Y., Melo T. M., Nissim I., Sonnewald U. andNissim I. (2007) The ketogenic diet and brain metabolism of aminoacids: relationship to the anticonvulsant effect. Annu. Rev. Nutr. 27,415–430.
� 2009 The AuthorsJournal Compilation � 2009 International Society for Neurochemistry, J. Neurochem. (2009) 110, 80–91
b-hydroxybutyrate alters glutamate homeostasis | 91
Related Documents