Arsenic hazards: strategies for tolerance and remediation by plants Rudra D. Tripathi 1 , Sudhakar Srivastava 1 , Seema Mishra 1 , Nandita Singh 1 , Rakesh Tuli 2 , Dharmendra K. Gupta 3 and Frans J.M. Maathuis 3 1 Environmental Science Division, National Botanical Research Institute, Rana Pratap Marg, Lucknow 226001, India 2 Centre for Plant Molecular Biology, National Botanical Research Institute, Rana Pratap Marg, Lucknow-226001, India 3 Department of Biology, Area 9, University of York, York YO10 5DD, UK Arsenic toxicity has become a global concern owing to the ever-increasing contamination of water, soil and crops in many regions of the world. To limit the detri- mental impact of arsenic compounds, efficient strategies such as phytoremediation are required. Suitable plants include arsenic hyperaccumulating ferns and aquatic plants that are capable of completing their life cycle in the presence of high levels of arsenic through the con- certed action of arsenate reduction to arsenite, arsenite complexation, and vacuolar compartmentalization of complexed or inorganic arsenic. Tolerance can also be conferred by lowering arsenic uptake by suppression of phosphate transport activity, a major pathway for arsenate entry. In many unicellular organisms, arsenic tolerance is based on the active removal of cytosolic arsenite while limiting the uptake of arsenate. Recent molecular studies have revealed many of the gene pro- ducts involved in these processes, providing the tools to improve crop species and to optimize phytoremediation; however, so far only single genes have been manipu- lated, which has limited progress. We will discuss recent advances and their potential applications, particularly in the context of multigenic engineering approaches. Introduction Arsenic (As) is ubiquitous in many environments and highly toxic to all forms of life. It occurs predominantly in inorganic form as arsenate (AsV) and arsenite (AsIII). Arsenate is an analogue of phosphate and thus interferes with essential cellular processes such as oxidative phos- phorylation and ATP synthesis, whereas the toxicity of AsIII is due to its propensity to bind to sulfhydryl groups, with consequent detrimental effects on general protein functioning. Water supplies, soils and sediments contaminated with As are the major sources of drinking water and food-chain contamination in numerous countries. This has caused a global epidemic of As poisoning, with many people having developed skin lesions, cancers and other symptoms [1,2]. Geochemical weathering of rocks and microbial activities also contribute to As mobilization in the environment, but human activity has enhanced this process and thus exacer- bated the problem [3]. In some areas of Bangladesh and India, groundwater As concentration has exceeded 2000 mgl À1 [4]. Arsenic can find its way into the grains of plants, such as rice and wheat, and into vegetables and fruit plants [5–8] through irrigation with As-contaminated water (Table 1). Rice is the major crop in areas where severe As contamination occurs [9]; it has been reported to accumulate up to 2 mg kg À1 As in grains [10] and up to 92 mg kg À1 in straw [11]. The diet of many rice consumers is, therefore, under threat from As contamination [6,9]. Two strategies might help to counter the detrimental effect of As: removal of As from the environment and the development of safe crops that can be grown in the pre- sence of As contamination. The physical and chemical technologies that have been applied to remediate As have, so far, not been successful [2]; however, As-hyperaccumu- lating ferns [12–15] and aquatic plants (macrophytes) [16] provide the possibility of developing cost-effective, eco- friendly As phytoremediation fuelled by solar energy [17]. Minimizing the uptake and translocation of As to edible parts would form the basis for improving crops. For both scenarios, understanding the mechanistic details of As uptake and detoxification are necessary. Recent studies have provided fresh insights into these processes in plants, and this review focuses on the poten- tial applications of these findings to reduce As hazards. Arsenic metabolism and tolerance in plants Arsenic forms inorganic and organic complexes in the environment, but the two biologically important species are AsV and AsIII, which are interconvertible depending on the redox status of the environment. Plants cope with AsV and AsIII using mechanisms that are partly similar to those discovered in microbes, in which As detoxification has been extensively studied (Box 1). The main route of AsV uptake in plants is through the phosphate transporters as a phosphate analogue [18], whereas AsIII is transported in the neutral As(OH) 3 form through aquaglyceroporins (Figure 1) [19]. Low-level uptake of organic As species, such as monomethylarsonic acid and dimethylarsinic acid, has also been observed in rice but the underlying transport pathways are unknown [6]. Restricting the influx of As might be an important mechanism to avoid toxicity, and several metallicolous plants (plants adapted to grow preferentially in soils con- taining a higher than normal concentration of a particular metal), such as Holcus lanatus and Cytisus striatus, Review TRENDS in Biotechnology Vol.25 No.4 Corresponding author: Maathuis, F.J.M. ([email protected]). Available online 15 February 2007. www.sciencedirect.com 0167-7799/$ – see front matter ß 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.tibtech.2007.02.003

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Arsenic hazards: strategies fortolerance and remediation by plantsRudra D. Tripathi1, Sudhakar Srivastava1, Seema Mishra1, Nandita Singh1,Rakesh Tuli2, Dharmendra K. Gupta3 and Frans J.M. Maathuis3
1 Environmental Science Division, National Botanical Research Institute, Rana Pratap Marg, Lucknow 226001, India2 Centre for Plant Molecular Biology, National Botanical Research Institute, Rana Pratap Marg, Lucknow-226001, India3 Department of Biology, Area 9, University of York, York YO10 5DD, UK
Review TRENDS in Biotechnology Vol.25 No.4
Arsenic toxicity has become a global concern owing tothe ever-increasing contamination of water, soil andcrops in many regions of the world. To limit the detri-mental impact of arsenic compounds, efficient strategiessuch as phytoremediation are required. Suitable plantsinclude arsenic hyperaccumulating ferns and aquaticplants that are capable of completing their life cycle inthe presence of high levels of arsenic through the con-certed action of arsenate reduction to arsenite, arsenitecomplexation, and vacuolar compartmentalization ofcomplexed or inorganic arsenic. Tolerance can also beconferred by lowering arsenic uptake by suppression ofphosphate transport activity, a major pathway forarsenate entry. In many unicellular organisms, arsenictolerance is based on the active removal of cytosolicarsenite while limiting the uptake of arsenate. Recentmolecular studies have revealed many of the gene pro-ducts involved in these processes, providing the tools toimprove crop species and to optimize phytoremediation;however, so far only single genes have been manipu-lated, which has limited progress. We will discuss recentadvances and their potential applications, particularly inthe context of multigenic engineering approaches.
IntroductionArsenic (As) is ubiquitous in many environments andhighly toxic to all forms of life. It occurs predominantlyin inorganic form as arsenate (AsV) and arsenite (AsIII).Arsenate is an analogue of phosphate and thus interfereswith essential cellular processes such as oxidative phos-phorylation and ATP synthesis, whereas the toxicity ofAsIII is due to its propensity to bind to sulfhydryl groups,with consequent detrimental effects on general proteinfunctioning.
Water supplies, soils and sediments contaminated withAs are the major sources of drinking water and food-chaincontamination in numerous countries. This has caused aglobal epidemic of As poisoning, with many people havingdeveloped skin lesions, cancers and other symptoms [1,2].Geochemical weathering of rocks and microbial activitiesalso contribute to As mobilization in the environment, buthuman activity has enhanced this process and thus exacer-bated the problem [3]. In some areas of Bangladesh and
Corresponding author: Maathuis, F.J.M. ([email protected]).Available online 15 February 2007.
www.sciencedirect.com 0167-7799/$ – see front matter � 2006 Elsevier Ltd. All rights reserve
India, groundwater As concentration has exceeded2000 mg l�1 [4]. Arsenic can find its way into the grainsof plants, such as rice and wheat, and into vegetables andfruit plants [5–8] through irrigation with As-contaminatedwater (Table 1). Rice is the major crop in areas wheresevere As contamination occurs [9]; it has been reported toaccumulate up to 2 mg kg�1 As in grains [10] and up to92 mg kg�1 in straw [11]. The diet of many rice consumersis, therefore, under threat from As contamination [6,9].
Two strategies might help to counter the detrimentaleffect of As: removal of As from the environment and thedevelopment of safe crops that can be grown in the pre-sence of As contamination. The physical and chemicaltechnologies that have been applied to remediate As have,so far, not been successful [2]; however, As-hyperaccumu-lating ferns [12–15] and aquatic plants (macrophytes) [16]provide the possibility of developing cost-effective, eco-friendly As phytoremediation fuelled by solar energy[17]. Minimizing the uptake and translocation of As toedible parts would form the basis for improving crops.
For both scenarios, understanding the mechanisticdetails of As uptake and detoxification are necessary.Recent studies have provided fresh insights into theseprocesses in plants, and this review focuses on the poten-tial applications of these findings to reduce As hazards.
Arsenic metabolism and tolerance in plantsArsenic forms inorganic and organic complexes in theenvironment, but the two biologically important speciesare AsV and AsIII, which are interconvertible dependingon the redox status of the environment. Plants cope withAsV and AsIII using mechanisms that are partly similar tothose discovered in microbes, in which As detoxificationhas been extensively studied (Box 1).
The main route of AsV uptake in plants is through thephosphate transporters as a phosphate analogue [18],whereas AsIII is transported in the neutral As(OH)3 formthrough aquaglyceroporins (Figure 1) [19]. Low-leveluptake of organic As species, such as monomethylarsonicacid and dimethylarsinic acid, has also been observed inrice but the underlying transport pathways are unknown[6]. Restricting the influx of As might be an importantmechanism to avoid toxicity, and several metallicolousplants (plants adapted to grow preferentially in soils con-taining a higher than normal concentration of a particularmetal), such as Holcus lanatus and Cytisus striatus,
d. doi:10.1016/j.tibtech.2007.02.003

Table 1. Accumulation of arsenic in food crops and other plants
Food Crop or vegetable or fruit Location Total As
(mg kgS1 dw)
Refs
Rice Bangladesh, China, Taiwan,
United States, Vietnam, West
Bengal, India, Europe
0.08–2.05 [5,9,10,55]
Wheat West Bengal, India 0.74 [7]
Potato, sweet potato, carrot, radish, onion, garlic,
cauliflower, broccoli, cabbage, celery, Chinese
cabbage, lettuce, ginger, tomato, cucumber, egg plant,
spinach and bamboo shoot
China 0.029–0.217 [8]
Snake gourd, ghotkal, taro, green papaya and
elephant foot
Bangladesh 0.338–0.489 [64]
Wheat flour, turmeric powder, beans and green chili West Bengal, India 0.080–0.335 [5]
Review TRENDS in Biotechnology Vol.25 No.4 159
improve As tolerance by constitutive suppression ofhigh-affinity phosphate/AsV transport [20,21].
As part of As detoxification, the majority of AsV isreduced to AsIII by the enzyme arsenate reductase (AR).Recently, AR genes have been identified in plants, includ-ing Arabidopsis (AtAsr/AtACR2), Holcus (HlAsr) andPteris (PvACR2) [22–24]. Many ARs from plants showCDC25 tyrosine phosphatase activity and contain theconserved motif HCX5R. Increased AR activity in plantssuch as H. lanatus might contribute to their As tolerance[22].
Plants exposed to As substantially increase the syn-thesis of glutathione (GSH) and phytochelatins (PCs), thepolymers of GSH [17,25]. Leguminous plants synthesizehomo-PCs (hPCs) in addition to PCs during As stress[17,26]. Augmented phytochelatin (PC) synthesis hasbeen observed in tolerant species, such as H. lanatus[27], and many studies point to the essential role of PCsin both constitutive and adaptive tolerance to As[25,27,28]. Complexation of As with GSH and PCs hasbeen demonstrated in various plants, such as Rauvolfiaserpentina, H. lanatus, P. cretica, Helianthus annuus andBrassica juncea [29–33], and appears to contain AsIII,exclusively. Raab et al. [33] demonstrated 14 different Ascomplexes in H. annuus, including AsIII–PC3, GS–AsIII–PC2, AsIII–GS3, AsIII–(PC2)2 and the newly identifiedmonomethylarsonic–PC2 [MA-AsIII–PC2]. However, inthe hyperaccumulator Pteris vittata most As appears tobe in an unbound, inorganic form [32,34], suggestingthat the role of PCs in As hyperaccumulation might belimited.
There is debate on the form in which As is transportedfrom root-to-shoot: X-ray and high-performance liquidchromatography–inductively coupled plasma mass spec-trometery (HPLC–ICP-MS) [34–37] studies on sporo-phytes of P. vittata have shown that As is translocatedto the shoot mainly as AsV and is stored in the fronds asinorganic AsIII. By contrast, Duan et al. [38] showedevidence that AR activity was found exclusively in theroots of P. vittata and, because most As exists as AsIIIin the fronds, concluded that the majority of As is translo-cated in its reduced form. This was also reported by others[22,39]. However, recent X-ray analyses on intact tissuefrom P. vittata provided strong evidence that, mainly,untransformed AsV is transported to shoots [34], andreduction of AsV was suggested to involve thiolates inthe form of a cylinder around the AsV in the veins.
www.sciencedirect.com
A final step in detoxification involves As sequestrationin the vacuoles of root and shoot tissue. In analogy to yeast,this might be in the form of AsIII–thiol complexes [1,22],although there is, as yet, no in vivo evidence that thisoccurs in plants. Recent research on hyperaccumulatingand tolerant species showed that vacuoles of P. vittata, P.cretica and H. lanatus contain large concentrations ofinorganic AsIII [24,32,34,36]. This suggests that themajority of stored As is not complexed, and that plantsmight have vacuolar AsIII transporters, possibly withsimilarities to bacterial AsIII extrusion pumps (Figure 1).
How to improve arsenic phytoremediationIncreasing As chelation
Many efforts have been made to increase thephytoremediation efficiency of plants. One of the strategiesto improve heavy metal and/or metalloid remediation wasto increase synthesis of chelators such as GSH and PCs.Reports on overexpression of phytochelatin synthase (PCS)showed promising results for both bacteria and plants [17].In one study, constitutive overexpression of AtPCS1resulted in a substantial increase in As resistance, witha 20–100 times greater biomass in transgenic plants afterexposure to As, but led to Cd hypersensitivity [28]. Despiteresulting in increased PC synthesis, chloroplast-targetedoverexpression of AtPCS1 markedly sensitized transgenicplants to As, whereas cytosolic overexpression of the samegene resulted in increased tolerance of transgenic plants toAs [40].
Thus, it appears that overexpression of PCS on its ownhas limited effects and can easily lead to unwanted proper-ties, such as toxicity. The latter might be partly due to thelimiting supply of essential metabolites such as cysteine, g-glutamylcysteine (g-EC) and GSH, which are needed forthe production of PCs [40]. Indeed, biochemical studieshave demonstrated that induced PC synthesis after Asexposure leads to depletion of GSH [27] and, consequently,oxidative stress [41] – a problem exacerbated by theadditional role of GSH in AsV reduction and AsIII com-plexation [32,33].
The additional overexpression of components involvedin GSH biosynthesis, such as g-glutamylcysteine synthe-tase (g-ECS) and glutathione synthetase (GS), mightprove a more successful approach because this will leadto increased levels of GSH and the end-product, PCs. Liet al. [42,43] overexpressed g-ECS in A. thaliana andfound a 3- to 20-fold greater production of g-EC, GSH
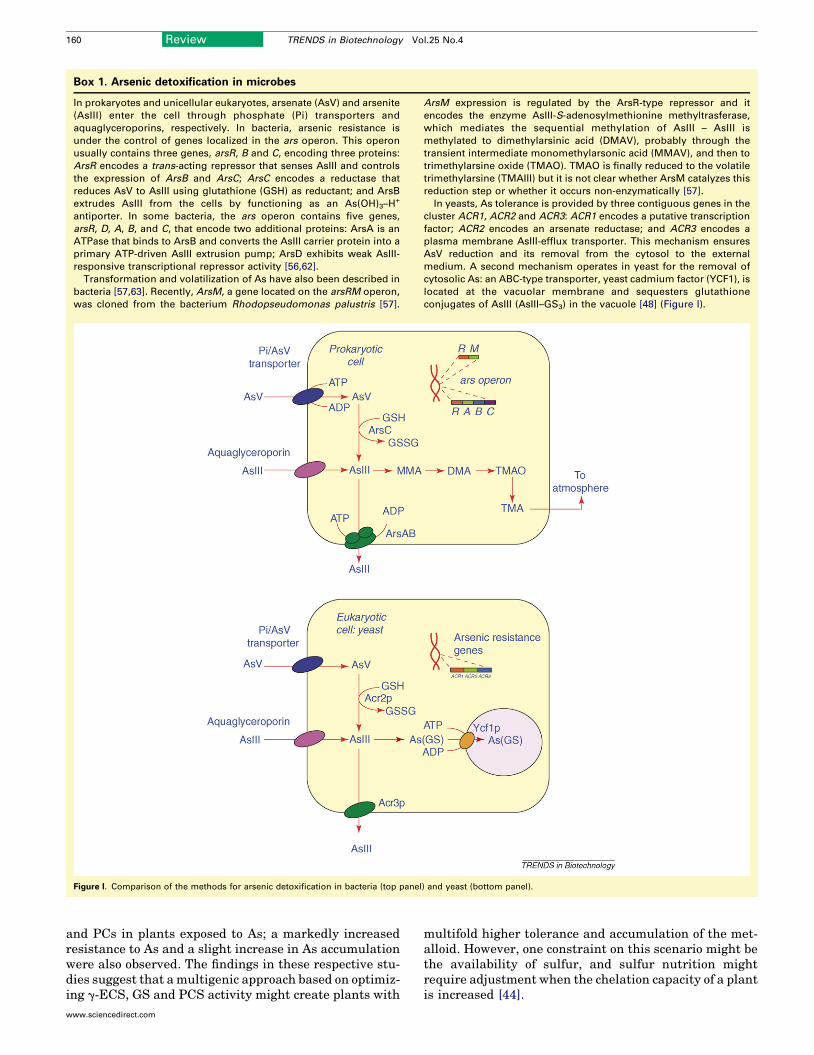
Box 1. Arsenic detoxification in microbes
In prokaryotes and unicellular eukaryotes, arsenate (AsV) and arsenite
(AsIII) enter the cell through phosphate (Pi) transporters and
aquaglyceroporins, respectively. In bacteria, arsenic resistance is
under the control of genes localized in the ars operon. This operon
usually contains three genes, arsR, B and C, encoding three proteins:
ArsR encodes a trans-acting repressor that senses AsIII and controls
the expression of ArsB and ArsC; ArsC encodes a reductase that
reduces AsV to AsIII using glutathione (GSH) as reductant; and ArsB
extrudes AsIII from the cells by functioning as an As(OH)3–H+
antiporter. In some bacteria, the ars operon contains five genes,
arsR, D, A, B, and C, that encode two additional proteins: ArsA is an
ATPase that binds to ArsB and converts the AsIII carrier protein into a
primary ATP-driven AsIII extrusion pump; ArsD exhibits weak AsIII-
responsive transcriptional repressor activity [56,62].
Transformation and volatilization of As have also been described in
bacteria [57,63]. Recently, ArsM, a gene located on the arsRM operon,
was cloned from the bacterium Rhodopseudomonas palustris [57].
ArsM expression is regulated by the ArsR-type repressor and it
encodes the enzyme AsIII-S-adenosylmethionine methyltrasferase,
which mediates the sequential methylation of AsIII – AsIII is
methylated to dimethylarsinic acid (DMAV), probably through the
transient intermediate monomethylarsonic acid (MMAV), and then to
trimethylarsine oxide (TMAO). TMAO is finally reduced to the volatile
trimethylarsine (TMAIII) but it is not clear whether ArsM catalyzes this
reduction step or whether it occurs non-enzymatically [57].
In yeasts, As tolerance is provided by three contiguous genes in the
cluster ACR1, ACR2 and ACR3: ACR1 encodes a putative transcription
factor; ACR2 encodes an arsenate reductase; and ACR3 encodes a
plasma membrane AsIII-efflux transporter. This mechanism ensures
AsV reduction and its removal from the cytosol to the external
medium. A second mechanism operates in yeast for the removal of
cytosolic As: an ABC-type transporter, yeast cadmium factor (YCF1), is
located at the vacuolar membrane and sequesters glutathione
conjugates of AsIII (AsIII–GS3) in the vacuole [48] (Figure I).
Figure I. Comparison of the methods for arsenic detoxification in bacteria (top panel) and yeast (bottom panel).
160 Review TRENDS in Biotechnology Vol.25 No.4
and PCs in plants exposed to As; a markedly increasedresistance to As and a slight increase in As accumulationwere also observed. The findings in these respective stu-dies suggest that a multigenic approach based on optimiz-ing g-ECS, GS and PCS activity might create plants with
www.sciencedirect.com
multifold higher tolerance and accumulation of the met-alloid. However, one constraint on this scenario might bethe availability of sulfur, and sulfur nutrition mightrequire adjustment when the chelation capacity of a plantis increased [44].
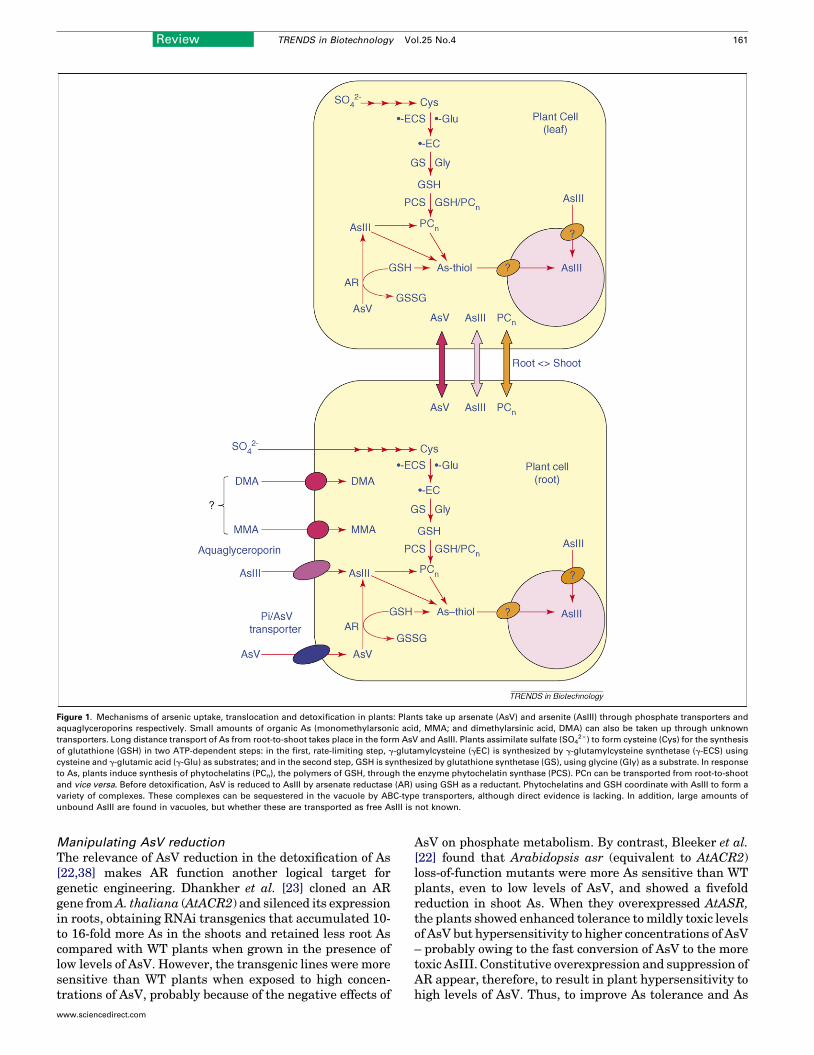
Figure 1. Mechanisms of arsenic uptake, translocation and detoxification in plants: Plants take up arsenate (AsV) and arsenite (AsIII) through phosphate transporters and
aquaglyceroporins respectively. Small amounts of organic As (monomethylarsonic acid, MMA; and dimethylarsinic acid, DMA) can also be taken up through unknown
transporters. Long distance transport of As from root-to-shoot takes place in the form AsV and AsIII. Plants assimilate sulfate (SO42�) to form cysteine (Cys) for the synthesis
of glutathione (GSH) in two ATP-dependent steps: in the first, rate-limiting step, g-glutamylcysteine (gEC) is synthesized by g-glutamylcysteine synthetase (g-ECS) using
cysteine and g-glutamic acid (g-Glu) as substrates; and in the second step, GSH is synthesized by glutathione synthetase (GS), using glycine (Gly) as a substrate. In response
to As, plants induce synthesis of phytochelatins (PCn), the polymers of GSH, through the enzyme phytochelatin synthase (PCS). PCn can be transported from root-to-shoot
and vice versa. Before detoxification, AsV is reduced to AsIII by arsenate reductase (AR) using GSH as a reductant. Phytochelatins and GSH coordinate with AsIII to form a
variety of complexes. These complexes can be sequestered in the vacuole by ABC-type transporters, although direct evidence is lacking. In addition, large amounts of
unbound AsIII are found in vacuoles, but whether these are transported as free AsIII is not known.
Review TRENDS in Biotechnology Vol.25 No.4 161
Manipulating AsV reduction
The relevance of AsV reduction in the detoxification of As[22,38] makes AR function another logical target forgenetic engineering. Dhankher et al. [23] cloned an ARgene fromA. thaliana (AtACR2) and silenced its expressionin roots, obtaining RNAi transgenics that accumulated 10-to 16-fold more As in the shoots and retained less root Ascompared with WT plants when grown in the presence oflow levels of AsV. However, the transgenic lines were moresensitive than WT plants when exposed to high concen-trations of AsV, probably because of the negative effects of
www.sciencedirect.com
AsV on phosphate metabolism. By contrast, Bleeker et al.[22] found that Arabidopsis asr (equivalent to AtACR2)loss-of-function mutants were more As sensitive than WTplants, even to low levels of AsV, and showed a fivefoldreduction in shoot As. When they overexpressed AtASR,the plants showed enhanced tolerance tomildly toxic levelsof AsV but hypersensitivity to higher concentrations of AsV– probably owing to the fast conversion of AsV to the moretoxic AsIII. Constitutive overexpression and suppression ofAR appear, therefore, to result in plant hypersensitivity tohigh levels of AsV. Thus, to improve As tolerance and As

Box 2. Arsenic phytoremediation: does nature know best?
Arsenic (As) toxicity is an increasing menace across the globe. Clean
up of the environment using plants, so-called phytoremediation,
potentially offers an efficient and cost-effective strategy but requires
species that are highly tolerant, show high bioaccumulation of As in
above ground tissue, grow fast and can be used in diverse habitats.
Various efforts have been made to improve the remediation
capacity of plants through genetic engineering, for example,
through overexpression of arsenate reductases and phytochelatin
synthases (see text). Although As tolerance in some transgenics was
enhanced, accumulation factors, particularly in shoot tissue, have
remained low.
By contrast, natural hyperaccumulators, such as the fern Pteris
vittata, can store arsenic in frond tissue at concentrations that are 10
to 30 times higher than those in the soil it grows in. P. vittata grows
relatively fast in its natural warm and humid habitat but its
application to other environments is limited. Identifying further
species would greatly enhance the feasibility to use natural
hyperaccumulators in arsenic phytoremediation.
Genetic engineering potentially provides an alternative route to
introduce hyperaccumulation traits into other plant species. How-
ever, many of the details of hyperaccumulation remain to be
elucidated, and developing molecular tools for species such as P.
vittata might be required to unravel the mechanisms of this
phenomenon. Nevertheless, hyperaccumulation is likely to be
sustained by the action of many gene products, including those
involved in arsenic uptake, arsenate reduction, root-to-shoot
translocation, vacuolar sequestration and aspects of sulfur and
phosphorus metabolism. Genetic engineering of single genes is,
therefore, unlikely to yield major progress in providing hyperaccu-
mulating plants, and an integrated approach is needed to optimize
the various aspects of plant arsenic tolerance and accumulation.
162 Review TRENDS in Biotechnology Vol.25 No.4
accumulation without affecting phosphate metabolism,overexpression of AR might need to be limited to the rootsand might need the simultaneous upregulation of thecapacity for AsIII chelation, root-to-shoot transport andsubsequent vacuolar sequestration.
Uptake, root-to-shoot translocation and vacuolar
sequestration of As
Efficient uptake and translocation from root-to-shootcontribute greatly to hyperaccumulation of As in P. vittata[12,45]. Not only is farmore As taken up but an exceedinglyhigh proportion of As is translocated to shoot tissue. Thisfact emphasizes the importance of the transportersinvolved in As uptake, root-to-shoot translocation andvacuolar sequestration during hyperaccumulation. In gen-eral, the constitutive expression of genes that encodevarious transporters, and the biosynthesis of chelators,is higher in hyperaccumulator plants compared withnon-accumulators [46].
For the long-distance movement of As, no or littleinformation is available on the nature of the transporterinvolved. For AsV, transport is considered to occur throughphosphate carriers, and the levels of expression of theArabidopsis phosphate transporters Ph1;1 and Ph1;4affect As tolerance [47]. However, no plant transporterhas yet been identified that could mediate transport ofAsIII from root-to-shoot; furthermore, the role of chelatorsfor the transport of As from root-to-shoot has not beendescribed. Pickering et al. [29] and Raab et al. [33] ident-ified unbound AsV and AsIII in xylem sap of Brassicajuncea and Helianthus annuus, but no As–PC complexes[33]. Thus, Raab et al. [33] postulated that AsV and AsIIIare the main species that are transported from root-to-shoot, not the As–PC complexes.
During the final step of detoxification, inorganic andcomplexed As can be sequestered into vacuoles. In yeast,vacuolar accumulation of AsIII–GS3 complexes ismediated by an ABC-type transporter [48]. In plants, invitro transport of GSH-complexed AsIII across tonoplastvesicles of A. thaliana has been demonstrated [22] and wasalso postulated to be carried out by an ABC transporter.However, no report of in vivo transport of either AsIII–GS3
or As–PC complexes in plants is available. Similarly, notransport processes that could mediate the vacuolar depo-sition of inorganic forms of As have been identified. Tonget al. [49] proposed that approaches using overexpressionof vacuolar transporters might prove successful because,even with a modest increase in vacuolar transportcapacity, substantially more substrate would be seques-tered without the need for large amounts of chelatingpeptide.
An integrated approach
The mechanisms discussed above have all been shown tooccur in plants and, to a lesser or greater extent, areamenable to manipulation to improve As phytoremedia-tion. There might be further processes that have beenunderinvestigated, such as As methylation [8,9,50] andvolatilization. Arsenic methyltransferase activity has beenrecorded in vitro from leaf extracts of Agrostis tenuis [50],and the potential to engineer plant As volatilization,
www.sciencedirect.com
possibly using bacterial genes (Box 1), certainly meritsfurther exploration.
More importantly, the above studies indicate thatmanipulation of single or small numbers of genes is unli-kely to result in plants that resemble natural hyperaccu-mulators (Box 2) and that the simultaneous optimization ofinterlinked processes is crucial. For example, Dhankheret al. [51] found substantially greater As tolerance andaccumulation inA. thaliana after combined overexpressionof ArsC and g-ECS relative to overexpression of g-ECS orArsC alone. However, we might have to intervene in theconstitutive or organ-specific expression of dozens of genesthat participate in As uptake, chelator synthesis, vacuolarsequestration and AsV reduction (Figure 2a). This mightrequire the identification of key regulatory genes and theimplementation of novel technologies such as trait pyra-miding and gene stacking.
Growing crops in the presence of arsenicThe accumulation of As in food (Table 1) is at alarmingproportions [6] and is mainly due to irrigation with As-contaminated water. To restrict As contamination of edibleplant parts, such as cereal grain, preventing root-to-shoottranslocation would suffice (Figure 2b). However, forroot crops, this strategy fails for obvious reasons and analmost complete cessation of As uptake would be required.Promoting As translocation to shoots and its subsequentvolatilization into the atmosphere might be another routefor increasing food safety. Although many crop species areaffected byAs contamination, wewill limit our focus to rice,the principal crop in severely As-contaminated areas.
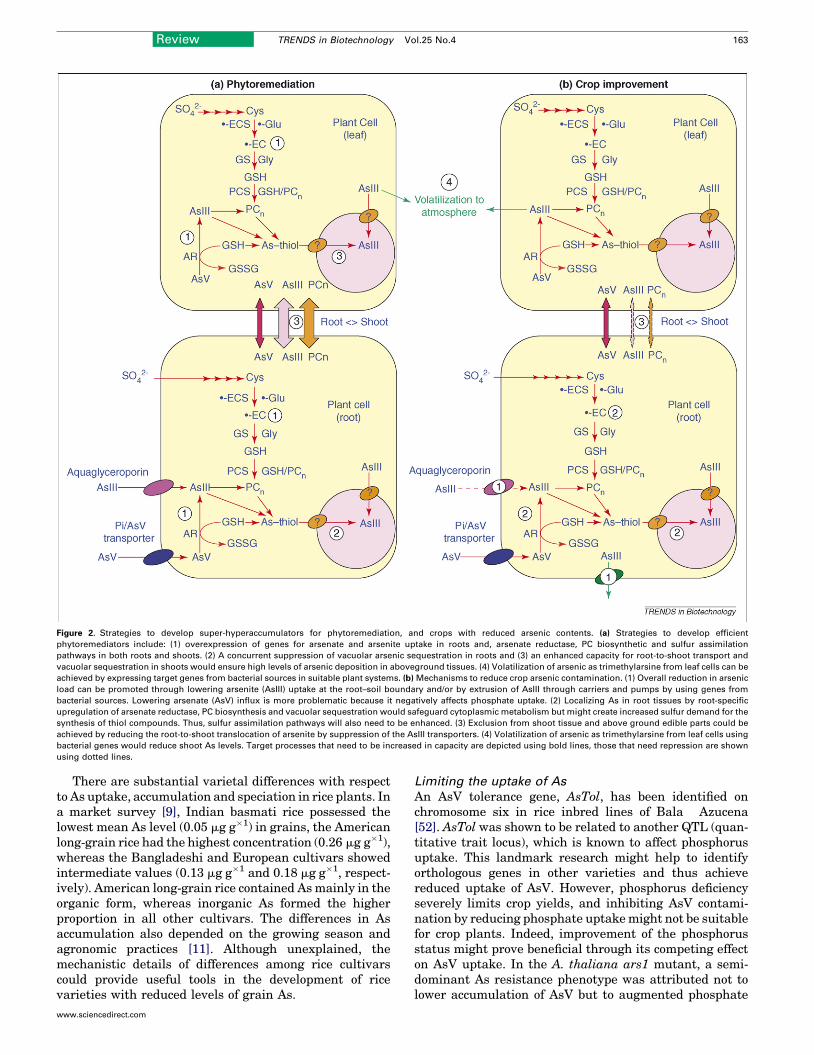
Figure 2. Strategies to develop super-hyperaccumulators for phytoremediation, and crops with reduced arsenic contents. (a) Strategies to develop efficient
phytoremediators include: (1) overexpression of genes for arsenate and arsenite uptake in roots and, arsenate reductase, PC biosynthetic and sulfur assimilation
pathways in both roots and shoots. (2) A concurrent suppression of vacuolar arsenic sequestration in roots and (3) an enhanced capacity for root-to-shoot transport and
vacuolar sequestration in shoots would ensure high levels of arsenic deposition in aboveground tissues. (4) Volatilization of arsenic as trimethylarsine from leaf cells can be
achieved by expressing target genes from bacterial sources in suitable plant systems. (b) Mechanisms to reduce crop arsenic contamination. (1) Overall reduction in arsenic
load can be promoted through lowering arsenite (AsIII) uptake at the root–soil boundary and/or by extrusion of AsIII through carriers and pumps by using genes from
bacterial sources. Lowering arsenate (AsV) influx is more problematic because it negatively affects phosphate uptake. (2) Localizing As in root tissues by root-specific
upregulation of arsenate reductase, PC biosynthesis and vacuolar sequestration would safeguard cytoplasmic metabolism but might create increased sulfur demand for the
synthesis of thiol compounds. Thus, sulfur assimilation pathways will also need to be enhanced. (3) Exclusion from shoot tissue and above ground edible parts could be
achieved by reducing the root-to-shoot translocation of arsenite by suppression of the AsIII transporters. (4) Volatilization of arsenic as trimethylarsine from leaf cells using
bacterial genes would reduce shoot As levels. Target processes that need to be increased in capacity are depicted using bold lines, those that need repression are shown
using dotted lines.
Review TRENDS in Biotechnology Vol.25 No.4 163
There are substantial varietal differences with respectto As uptake, accumulation and speciation in rice plants. Ina market survey [9], Indian basmati rice possessed thelowest mean As level (0.05 mg g�1) in grains, the Americanlong-grain rice had the highest concentration (0.26 mg g�1),whereas the Bangladeshi and European cultivars showedintermediate values (0.13 mg g�1 and 0.18 mg g�1, respect-ively). American long-grain rice contained Asmainly in theorganic form, whereas inorganic As formed the higherproportion in all other cultivars. The differences in Asaccumulation also depended on the growing season andagronomic practices [11]. Although unexplained, themechanistic details of differences among rice cultivarscould provide useful tools in the development of ricevarieties with reduced levels of grain As.
www.sciencedirect.com
Limiting the uptake of As
An AsV tolerance gene, AsTol, has been identified onchromosome six in rice inbred lines of Bala � Azucena[52]. AsTol was shown to be related to another QTL (quan-titative trait locus), which is known to affect phosphorusuptake. This landmark research might help to identifyorthologous genes in other varieties and thus achievereduced uptake of AsV. However, phosphorus deficiencyseverely limits crop yields, and inhibiting AsV contami-nation by reducing phosphate uptakemight not be suitablefor crop plants. Indeed, improvement of the phosphorusstatus might prove beneficial through its competing effecton AsV uptake. In the A. thaliana ars1 mutant, a semi-dominant As resistance phenotype was attributed not tolower accumulation of AsV but to augmented phosphate

164 Review TRENDS in Biotechnology Vol.25 No.4
uptake [53]. Similarly, high levels of phosphate improvedtolerance to AsV in WT Arabidopsis, a phenomenon alsoobserved in several bacterial species [3] that maintainhigh-uptake rates for phosphate, even in the presence of100-fold ratios of AsV:phosphate. In our field study on therice varieties Saryu-52, Sabha-5204 and Pant-4 that weresubjected to low doses of fly-ash (FA) containing As, no Asaccumulation was detected in the grains and low accumu-lation levels were found in leaves [54] – this was thought tobe because of the presence of phosphate inFA-amended soil.
Recently, iron plaque formation due to the oxidation offerrous to ferric iron and the precipitation of iron oxide onthe root surface has been shown to cause a decrease in Asconcentration in rice. This was interpreted to be the resultof extensive binding of As to the iron plaque, thus limitingAs uptake. Different rice cultivars showed differentcapacities to form iron plaques, thus providing furthermeans to modulate As uptake in rice through selectionof proper rice genotypes [55].
Removal of As from specific cells and tissues
Extrusion of As from cells [56] as well as its volatilization tothe atmosphere [57] are mechanisms that are well charac-terized in bacterial As tolerance (Box 1). Some of themicrobial genes responsible for volatilization have beensuccessfully applied to improve selenium and mercuryphytoremediation [58,59]. Similar approaches might provesuccessful in reducing As build up in edible plant partssuch as grains. A high level of root AsIII extrusion capacitythrough ArsAB- and ACR3-like mechanisms in the plasmamembrane, coupled to the action of gene products such asArsM in shoot tissue – to promote As volatilization – mightbe desirable in this respect.
Lee et al. [60] tested whether the E. coli ZntA gene,which encodes a PbII/CdII/ZnII transporting ATPase thatpumps metals out of the cell, could be useful for developingplants with reduced metal content. Arabidopsis plantstransformed with ZntA showed a reduction in heavy metalcontent and improved resistance to Pb and Cd. Moreover,the transgenic protoplasts showed faster release of pre-loaded Cd than WT protoplasts. Recently, Yang et al. [61]found an aquaglyceroporin (AqpS) in place of ArsB in thears operon of the legume symbiont Sinorhizobiummeliloti.Co-expression of S. meliloti AqpS and ArsC in a strain ofE.coli that lacks the ars operon complemented AsV sensi-tivity, suggesting the involvement of aquaglyceroporins inAsIII extrusion rather than AsIII uptake. This offers newpossibilities related to As detoxification and removal.Sequence comparison of AqpS with plant aquaglyceropor-ins might result in the identification of specific isoformsthat could participate in AsIII extrusion from plant cells.
In summary, a range of strategies is potentially avail-able to reduce rice As contamination. Molecular targetsinclude reduced As uptake while enhancing selectivity forphosphate uptake. In most non-accumulator plants, theretention of AsIII–thiol complexes in root tissues forms amajor part of copingwith As stress. Thus, crop varieties areneeded that adopt similar strategies to maintain low Aslevels in edible parts. Root specific overexpression of thegenes involved in the reduction of AsV, coupled withchelation and sequestration of AsIII, might result in
www.sciencedirect.com
increased retention of the metalloid in roots and thuspreclude its accumulation in edible parts. Although morespeculative, AsIII extrusion and As volatilization mightalso be helpful in minimizing As load. Progress in thesedirections is unlikely to be swift because multiple targetsmight have to be combined. In the meantime, the Ascontent in rice can be reduced to acceptable levels throughselection of appropriate varieties and management ofagronomic practices.
AcknowledgementsThis work was supported by a project grant by the Department ofBiotechnology, Government of India. SM and SS are thankful to Councilof Scientific and Industrial Research, New Delhi, India for the award ofSenior Research Fellowships. The authors thank Andrew A. Meharg atthe School of Biological Sciences, University of Aberdeen, Aberdeen, UKfor going through the article and making constructive suggestions. Weappreciate discussions with N. Kumar, Scientist, NBRI, Lucknow, Indiaduring preparation of this article.
References1 Dhankher, O.P. (2005) Arsenic metabolism in plants: an inside story.
New Phytol. 168, 503–5052 Mondal, P. et al. (2006) Laboratory-based approaches for arsenic
remediation from contaminated water: recent developments. J.Hazard Mater. 137, 464–479
3 Duker, A.A. et al. (2005) Arsenic geochemistry and health.Environ. Int.31, 631–641
4 Hossain, M.F. (2006) Arsenic contamination in Bangladesh – a review.Agric. Ecosyst. Environ. 113, 1–16
5 Roychowdhury, T. et al. (2002) Survey of arsenic in food compositesfrom an arsenic-affected area of West Bengal, India. Food Chem.Toxicol. 40, 1611–1621
6 Meharg, A.A. (2004) Arsenic in rice – understanding a new disaster forSouth-East Asia. Trends Plant Sci. 9, 415–417
7 Norra, S. et al. (2005) Impact of irrigation with As-rich groundwater onsoil and crops: a geochemical case study in West Bengal delta plain,India. Appl. Geochem. 20, 1890–1906
8 Zhao, R. et al. (2006) Arsenic speciation in moso bamboo shoot – aterrestrial plant that contains organoarsenic species. Sci. TotalEnviron. 371, 293–303
9 Williams, P.N. et al. (2005) Variation in arsenic speciation andconcentration in paddy rice related to dietary exposure. Environ.Sci. Technol. 39, 5531–5540
10 Islam, M.R. et al. (2004) Assessment of arsenic in the water–soil–plantsystems in gangetic flood plains of Bangladesh. Asian J. Plant Sci. 3,489–493
11 Abedin, M.J. et al. (2002) Uptake kinetics of arsenic species in riceplants. Plant Physiol. 128, 1–9
12 Ma, L.Q. et al. (2001) A fern that hyperaccumulates arsenic – a hardy,versatile, fast-growing plant helps to remove arsenic fromcontaminated soils. Nature 409, 579
13 Srivastava, M. et al. (2006) Three new arsenic hyperaccumulatingferns. Sci. Total Environ. 364, 24–31
14 Wang, H.B. et al. (2006) Arsenic uptake and accumulation in fernspecies growing at arsenic-contaminated sites of Southern China: fieldsurveys. Int. J. Phytoremediation 8, 1–11
15 Francessconi, K. et al. (2002) Arsenic species in an arsenichyperaccumulating fern, Pityrogramma calomelanos: a potentialphytoremediator of arsenic-contaminated soils. Sci. Total Environ.284, 27–35
16 Robinson, B. et al. (2006) Arsenic hyperaccumulation by aquaticmacrophytes in the Taupo volcanic zone, New Zealand. Environ.Exp. Bot. 58, 206–215
17 Grill, E. et al. (2006) Role of phytochelatins in phytoremediation ofheavy metals. In Environmental Bioremediation Technologies (Singh,S.N. and Tripathi, R.D., eds), pp. 101–146, Springer
18 Asher, C.J. and Reay, P.F. (1979) Arsenic uptake by barley seedlings.Aust. J. Plant Physiol. 6, 459–466
19 Meharg, A.A. and Jardine, L. (2003) Arsenite transport into paddy rice(Oryza sativa) roots. New Phytol. 157, 39–44

Review TRENDS in Biotechnology Vol.25 No.4 165
20 Meharg, A.A. and Macnair, M.R. (1992) Suppression of the high-affinity phosphate uptake system: a mechanism of arsenatetolerance in Holcus lanatus L. J. Exp. Bot. 43, 519–524
21 Bleeker, P.M. et al. (2003) Mechanisms of arsenate tolerance inCytisusstriatus. New Phytol. 157, 33–38
22 Bleeker, P.M. et al. (2006)Enhanced arsenate reduction byaCDC25-liketyrosine phosphatase explains increased phytochelatin accumulation inarsenate-tolerant Holcus lanatus. Plant J. 45, 917–929
23 Dhankher, O.P. et al. (2006) Hyperaccumulation of arsenic in theshoots of Arabidopsis silenced for arsenate reductase (ACR2). Proc.Natl. Acad. Sci. U. S. A. 103, 5413–5418
24 Ellis, D.R. et al. (2006) A novel arsenate reductase from the arsenichyperaccumulating Pteris vittata. Plant Physiol. 141, 1544–1554
25 Schat, H. et al. (2002) The role of phytochelatins in constitutive andadaptive heavy metal tolerances in hyperaccumulator and non-hyperaccumulator metallophytes. J. Exp. Bot. 53, 2381–2392
26 Gupta, D.K. et al. Accumulation of arsenic in root and shoot affects thegrowth and level of phytochelatins in seedlings of two varieties of Cicerarietinum L. var. CSG-8962 and C-235. J. Environ. Biol. (in press)(http://www.geocities.com/j_environ_biol/)
27 Hartley-Whitaker, J. et al. (2001) Is differential phytochelatinproduction related to decreased arsenate influx in arsenate tolerantHolcus lanatus? New Phytol. 155, 219–225
28 Li, Y. et al. (2004) Overexpression of phytochelatin synthase inArabidopsis leads to enhanced arsenic tolerance and cadmiumhypersensitivity. Plant Cell Physiol. 45, 1787–1797
29 Pickering, I.J. et al. (2000) Reduction and coordination of arsenic inIndian mustard. Plant Physiol. 122, 1171–1177
30 Schmoger, M.E.V. et al. (2000) Detoxification of arsenic byphytochelatins in plants. Plant Physiol. 122, 793–801
31 Montes-Bayon, M. et al. (2004) HPLC–ICP-MS and ESI-Q-TOFanalysis of biomolecules induced in Brassica juncea during arsenicaccumulation. J. Anal. At. Spectrom. 19, 153–158
32 Raab, A. et al. (2004) The nature of arsenic–phytochelatin complexes inHolcus lanatus and Pteris cretica. Plant Physiol. 134, 1113–1122
33 Raab, A. et al. (2005) Uptake, translocation and transformation ofarsenate and arsenite in sunflower (Helianthus annuus): formation ofarsenic–phytochelatin complexes during exposure to high arsenicconcentrations. New Phytol. 168, 551–558
34 Pickering, I.J. et al. (2006) Localizing the biochemical transformationsof arsenate in a hyperaccumulating fern. Environ. Sci. Technol. 40,5010–5014
35 Hokura, A. et al. (2006) Arsenic distribution and speciation in anarsenic hyperaccumulator fern by X-ray spectroscopy utilizing asynchrotron radiation source. J. Anal. At., Spectrom. 21, 321–328
36 Ze-Chun, H. et al. (2004) Direct determination of arsenic species inarsenic hyperaccumulator Pteris vittata by EXAFS. Acta Bot. Sin. 46,46–50
37 Zhao, F.J. et al. (2003) The role of phytochelatins in arsenic tolerance inthe hyperaccumulator Pteris vittata. New Phytol. 159, 403–410
38 Duan, G-L. et al. (2005) Characterization of arsenate reductase in theextract of roots and fronds of Chinese Brake Fern, an arsenichyperaccumulator. Plant Physiol. 138, 461–469
39 Quaghbeur,M. andRengel, Z. (2004) Arsenic uptake, translocation andspeciation in pho1 and pho2 mutants of Arabidopsis thaliana. Physiol.Plant. 120, 280–286
40 Picault, N. et al. (2006) Chloroplast targeting of phytochelatin synthasein Arabidopsis: effects on heavy metal tolerance and accumulation.Biochimie 88, 1743–1750
41 Singh, N. et al. (2006) Metabolic adaptations to arsenic-inducedoxidative stress in Pteris vittata L. and Pteris ensiformis L. PlantSci. 170, 274–282
42 Li, Y. et al. (2005) Engineered overexpression of g-glutamylcysteinesynthetase in plants confers high level arsenic and mercury tolerance.Environ. Toxicol. Chem. 24, 1376–1386
www.sciencedirect.com
43 Li, Y. et al. (2006) The shoot-specific expression of g-glutamylcysteinesynthetase directs the long-distance transport of thiol-peptides to rootsconferring tolerance to mercury and arsenic. Plant Physiol. 141, 288–298
44 Cherian, S. and Oliveira, M.M. (2005) Transgenic plants inphytoremediation: recent advances and new possibilities. Environ.Sci. Technol. 39, 9377–9390
45 Singh, N. and Ma, L.Q. (2006) Arsenic speciation, and arsenic andphosphate distribution in arsenic hyperaccumulator Pteris vittata L.and non-hyperaccumulator Pteris ensiformis L. Environ. Pollut. 141,238–246
46 Kramer, U. (2005) Phytoremediation: novel approaches to cleaning uppolluted soils. Curr. Opin. Biotechnol. 16, 133–141
47 Shin, H. et al. (2004) Phosphate transport in Arabidopsis: Pht1;1 andPht1;4 play a major role in phosphate acquisition from both low- andhigh-phosphate environments. Plant J. 39, 629–642
48 Ghosh, M. et al. (1999) Pathway of As(III) detoxification inSachharomyces cerevisiae. Proc. Natl. Acad. Sci. U. S. A. 96, 5001–5006
49 Tong, Y-P. et al. (2004) Vacuolar compartmentalization: a second-generation approach to engineering plants for phytoremediation.Trends Plant Sci. 9, 7–9
50 Wu, J.H. et al. (2002) Methylation of arsenic in vitro by cell extractsfrom bentgrass (Agrostis tenuis): effect of acute exposure of plants toarsenate. Func. Plant Biol. 29, 73–80
51 Dhankher, O.P. et al. (2002) Engineering tolerance andhyperaccumulation of arsenic in plants by combining arsenatereductase and g-glutamylcysteine synthetase expression. Nat.Biotechnol. 20, 1140–1145
52 Dasgupta, T. et al. (2004) An arsenate tolerance gene on chromosome 6of rice. New Phytol. 163, 45–49
53 Lee, D.A. et al. (2003a) ars1, an Arabidopsis mutant exhibitingincreased tolerance to arsenate and increased phosphate uptake.Plant J. 35, 637–646
54 Dwivedi, S. et al. (2006) Growth performance and biochemicalresponses of three rice (Oryza sativa L.) cultivars grown in fly-ashamended soil. Chemosphere 67, 140–151
55 Liu, W-J. et al. (2004) Do phosphorus nutrition and iron plaque alterarsenate (As)-uptake by rice seedlings in hydroponic culture? NewPhytol. 162, 481–488
56 Rosen, B.P. (2002) Biochemistry of arsenic detoxification. FEBS Lett.529, 86–92
57 Quin, J. et al. (2006) Arsenic detoxification and evolution oftrimethylarsine gas by a microbial arsenite S-adenosylmethioninemethyltransferase. Proc. Natl. Acad. Sci. U. S. A. 103, 2075–2080
58 LeDuc, D.L. and Terry, N. (2005) Phytoremediation of toxic traceelements in soil and water. J. Ind. Microbiol. Biotechnol. 32, 514–520
59 Meagher, R.B. and Heaton, A.C. (2005) Strategies for the engineeredphytoremediation of toxic element pollution: mercury and arsenic. J.Ind. Microbiol. Biotechol. 32, 502–513
60 Lee, J. et al. (2003b) Functional expression of a bacterial heavy metaltransporter in Arabidopsis enhances resistance to and decreasesuptake of heavy metals. Plant Physiol. 133, 589–596
61 Yang, H-C. et al. (2005) Novel pathway for arsenic detoxification inthe legume symbiont Sinorhizobium meliloti. J. Bacteriol. 187, 6991–6997
62 Rosen, B.P. (1999) Families of arsenic transporters. Trends Microbiol.7, 207–212
63 Bentley, R. and Chasteen, T.G. (2002) Microbial methylation ofmetalloids: arsenic, antimony and bismuth. Microbiol. Mol. Biol.Rev. 66, 250–271
64 Alam, M.G.M. et al. (2003) Arsenic and heavy metal contamination invegetables grown in Samta village, Bangladesh. Sci. Total Environ.308, 83–96
Related Documents











