Bioscience Reports, Vol. 7, No. 4, 1987 Annexins--New Family of Ca 2 +-Regulated- Phospholipid Binding Protein Michael J. Geisow 1, John H. Walker 2, Catherine Boustead 2 and Willie Taylor 3 Received May 6, 1987 KEY WORDS: Annexin; Ca2+-binding protein; calelectrin; calpactin; phospholipid-binding protein; tyrosine kinase. Calcium and phospholipid binding proteins have been identified and localized by immunocytochemistry in a wide range of cells and tissues. Two of these proteins (calpactins) also bind F-actin and are substrates for tyrosine kinases. The similar membrane-binding properties of these molecules arise from conserved amino acid sequences and a model is proposed for the tertiary structure of a common calcium and phospholipid binding domain. BACKGROUND A new group of calcium-binding proteins which associate reversibly with biomembranes has recently been identified in many different cells and tissues (Owens and Crumpton, 1984; Geisow and Walker, 1986). The key common feature of these proteins is their ability to bind to phospholipid bilayers in the presence of micromolar free calcium concentrations. This binding appears to be both specific for calcium (as opposed to other divalent cations) and for acidic phospholipids. No binding takes place to pure lecithin bilayers. A subsidiary common feature of these proteins is their ability, at more elevated levels of calcium, to potentiate the aggregation of phospholipid vesicles as well as natural membranes. This property was first reported for an adrenal medullary protein (Creutz et al., 1978) named "synexin" (Gr. synexis-- n~leeting). In keeping with this idea we use the generic term "Annexin" to refer to the group as a whole. In this review we describe the biochemistry and cell biology of the 1 Protein Chemistry, Delta Biotechnology Ltd., Castle Boulevard, Nottingham NG7 1FD. 2 Biochemistry Department, Leeds University. a Department of Crystallography, Birkbeck College, Malet Street, London NW1. 289 0144-8463/87/0400.0289505.00/0 1987PlenumPublishing Corporation

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Bioscience Reports, Vol. 7, No. 4, 1987
Annexins--New Family of C a 2 +-Regulated- Phospholipid Binding Protein
Michael J. Geisow 1, John H. Walker 2, Catherine Boustead 2 and Willie Taylor 3
Received May 6, 1987
KEY WORDS: Annexin; Ca2+-binding protein; calelectrin; calpactin; phospholipid-binding protein; tyrosine kinase.
Calcium and phospholipid binding proteins have been identified and localized by immunocytochemistry in a wide range of cells and tissues. Two of these proteins (calpactins) also bind F-actin and are substrates for tyrosine kinases. The similar membrane-binding properties of these molecules arise from conserved amino acid sequences and a model is proposed for the tertiary structure of a common calcium and phospholipid binding domain.
BACKGROUND
A new group of calcium-binding proteins which associate reversibly with biomembranes has recently been identified in many different cells and tissues (Owens and Crumpton, 1984; Geisow and Walker, 1986). The key common feature of these proteins is their ability to bind to phospholipid bilayers in the presence of micromolar free calcium concentrations. This binding appears to be both specific for calcium (as opposed to other divalent cations) and for acidic phospholipids. No binding takes place to pure lecithin bilayers. A subsidiary common feature of these proteins is their ability, at more elevated levels of calcium, to potentiate the aggregation of phospholipid vesicles as well as natural membranes. This property was first reported for an adrenal medullary protein (Creutz et al., 1978) named "synexin" (Gr. synexis-- n~leeting). In keeping with this idea we use the generic term "Annexin" to refer to the group as a whole. In this review we describe the biochemistry and cell biology of the
1 Protein Chemistry, Delta Biotechnology Ltd., Castle Boulevard, Nottingham NG7 1FD. 2 Biochemistry Department, Leeds University. a Department of Crystallography, Birkbeck College, Malet Street, London NW1.
289 0144-8463/87/0400.0289505.00/0 �9 1987 Plenum Publishing Corporation

290 Geisow, Walker, Boustead and Taylor
Table 1 Nomenclature of the annexins
Mol. wt. (Kd) pI Names
67-73 5.7-5.9
56-57 5.7 47 6.1-6.9
3540 6.5-6.8 36 4.8-5.5
34-39 7.4-7.8
28-33 5.4--5.8
p70 (liver/adrenal/placenta), p68 (lymphocyte), 67 Kd calcemedin (smooth muscle), Protein III (liver), synhibin, chromobidin XV Synexin ti, chromobindin XVIt Synexin I, chromobindin VI p35, calpactin II, lipocortin I, lipomodulin, chromobindin IX calelectrin p36 (p39, p34), calpactin I, lipocortin II, Protein I (intestine), p33 (lymphocyte), chromobindin VIII p32.5, endonexin (liver/adrenal/placenta), protein II (intestine), p28 (lymphocyte), chromobindin IV
annexin family and recent advances in their structure, which points the way to understanding their functions and interactions within the cell.
Because members of the annexin family have been isolated and characterised in quite different contexts, a confusing terminology exists. Recent immunological and amino acid sequence data has almost completely resolved this and a nomenclature key is indicated in Table 1. A brief historical description will also be helpful in relating the names shown in this table.
Creutz (1983) and Geisow and Burgoyne (1982) described unknown proteins with the ability to bind to chromaffin granules from adrenal medulla, at micromolar free calcium concentrations. Creutz called these proteins chromobindins and clearly distinguished a subset of these which bound to phospholipid as opposed to integral membrane protein components. At about this time an analogous protein was isolated from the electric organ of the ray Torpedo marmorata (Walker, 1982) and called calelectrin. Antisera raised to denatured calelectrin was found to cross-react with a number of mammalian proteins, in particular adrenal chromaffin granule-binding and liver proteins (Siidhof et al., 1984; Geisow et al., 1984). Calelectrin was also found to be a phospholipid-binding and vesicle aggregation protein (Siidhof et al., 1982; Siidhof et al., 1985). The unnamed mammalian equivalents were referred to as mammalian caMectrins. Like calelectrin, the mammalian proteins could be purified from crude cell or tissue homogenates by Ca 2 § -dependent affinity chromatography on phenyl or phenothiazine-substituted Sepharose. (Siidhof et al., 1984) and this approach was used to isolate proteins called calcimedins from smooth muscle homogenates (Moore and Dedman, 1982) and from mouse mammary epithelial cells (Braslau et aI., 1984).
Bovine liver yielded two major proteins, p70 and p32.5 (endonexin) by this method. The former protein is almost certainly identical with the previously-named synhibin (Pollard and Scott, 1982), a 68 Kd liver protein which competes with synexin for binding to chromaffin granules. Minor proteins of molecular weights 56 Kd and 47 Kd, present in both tissues and called Synexin II and I have been less well characterised, but in view of their reported properties are included in Table 1 as members of the "annexin family".
Essentially the same proteins have been isolated from purified plasma membrane fractions after extraction with EGTA. In most cases a non-ionic detergent was used to

Annexins 291
remove excess lipid from the preparations to give a detergent-insoluble lipid-protein residue (the cytoskeleton).
Lymphocyte plasma membrane cytoskeleton extracted with EGTA yielded large amounts of p68, but also proteins corresponding to p36 and endonexin (see Table 1) (Owens and Crumpton, 1984; Davis and Crumpton, 1985). Intestinal epithelial cells proved to contain large amounts of p36 and endonexin in association with the brush border membranes (Gerke and Weber, 1984). In a key series of studies, Gerke and Weber showed that p36 corresponded to a previously known substrate for retroviral protein tyrosine kinases, described variously as p39, p36 or p34 (for review see Cooper and Hunter, 1983). This protein was shown to bind F-actin and possibly fodrin in a Ca 2 +-dependent manner (Gerke and Weber, 1985a) as well as to liposomes composed of phosphatidylserine (Glenney, 1985; Johnsson et aI., 1986). Independently, a Mr 35,000 Kd calcium-dependent membrane-binding protein (p35) was identified in the human epidermoid carcinoma cell line A431 (Fava and Cohen, 1984) and as a major component of human placenta (Valentine-Braun et al., 1986; Haigler et al., 1987). This protein was a substrate for the epidermal growth factor (EGF) protein tyrosine kinase which is rich in these two biological sources. The clear demonstration of both p36 and p35 as distinct proteins, co-expressed in the A431 cell line was made by Glenney (1986b) together with the demonstration that p35, like p36, also binds actin.
The identification of p35 with lipocortin (a previously poorly defined molecule with phospholipase A z inhibitory activity, reviewed by Flower et al. (1984) was made by Pepinsky and Sinclair (1986). This finding was quickly followed by the publication of complete amino acid sequences for p35 by Pepinsky's group (Wallner et al., 1986) and for p36 by Pepinsky at Biogen and Hunter at the Salk (Huang et al., 1986; Saris et al., 1986) resulting from cloning the appropriate cDNAs. A further important result from the cDNA work was the finding of single genes for these two proteins, neither with hydrophobic sequences expected of secreted proteins. The origin of this protein in extracellular medium containing elicited macrophages (Blackwell et al., 1980, 1982) is therefore unknown.
Partial amino acid sequence analyses of annexins (protein II, p36, endonexin and calelectrin) had identified a short amino-acid sequence homology present in multiple copies within each protein (Geisow et al., 1986). It became clear from examination of the complete sequences of p35 and p36 that these proteins were homologous over repeated stretches of about 70 amino acids. There are probably four of these repeats in each of the above proteins.
CELLULAR DISTRIBUTION AND LOCALIZATION OF ANNEXINS
Calelectrin
Calelectrin is present throughout the cells of the electric fish (Torpedo marmorata) cholinergic electromotor tissue (Fiedler and Walker, 1985). The most significant localizations are at the cytoplasmic surface of the post-synaptic acetyl-choline re, ceptor-bearing membrane and in association with synaptic vesicles and neurofilaments (Fig. 1). Association with these organelles is Ca2+-dependent, since EGTA applied to tissue sections removed immunoreactive calelectrin.
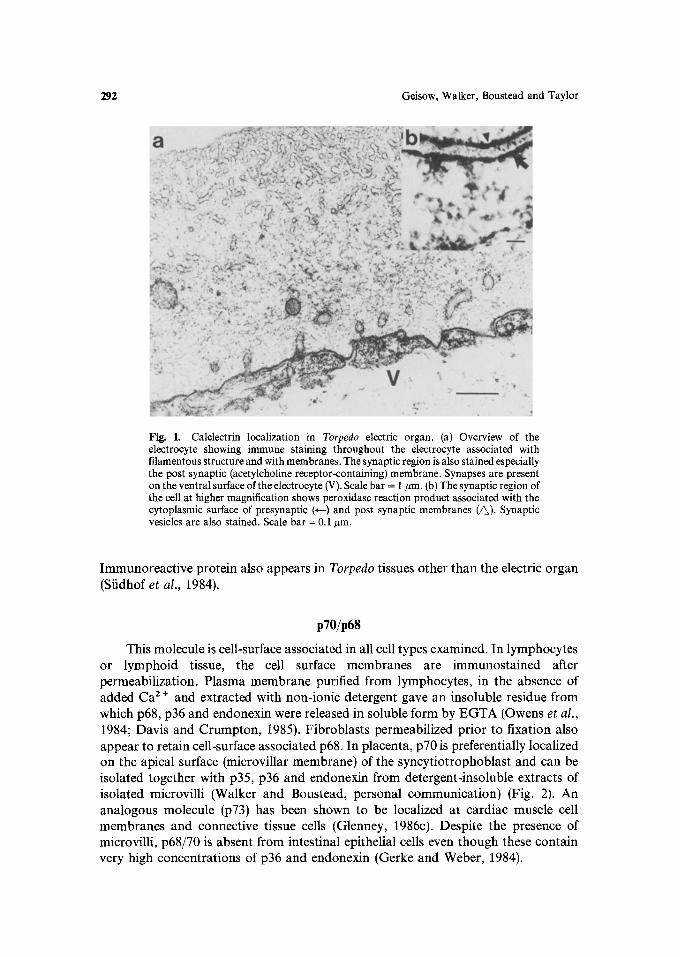
292 Geisow, Walker, Boustead and Taylor
Fig. 1. Calelectrin localization in Torpedo electric organ. (a) Overview of the electrocyte showing immune staining throughout the electrocyte associated with filamentous structure and with membranes. The synaptic region is also stained especially the post synaptic (acetylcholine receptor-containing) membrane. Synapses are present on the ventral surface of the electrocyte (V). Scale bar = 1 #m. (b) The synaptic region of the cell at higher magnification shows peroxidase reaction product associated with the cytoplasmic surface of presynaptic (~--) and post synaptic membranes (~). Synaptic vesicles are also stained. Scale bar = 0.1 #m.
Immunoreactive protein also appears in Torpedo tissues other than the electric organ (Sfidhof et al., 1984).
p70/p68
This molecule is cell-surface associated in all cell types examined. In lymphocytes or lymphoid tissue, the cell surface membranes are immunostained after permeabilization. Plasma membrane purified from lymphocytes, in the absence of added Ca 2 § and extracted with non-ionic detergent gave an insoluble residue from which p68, p36 and endonexin were released in soluble form by EGTA (Owens et al., 1984; Davis and Crumpton, 1985). Fibroblasts permeabilized prior to fixation also appear to retain cell-surface associated p68. In placenta, p70 is preferentially localized on the apical surface (microvillar membrane) of the syncytiotrophoblast and can be isolated together with p35, p36 and endonexin from detergent-insoluble extracts of isolated microvilli (Walker and Boustead, personal communication) (Fig. 2). An analogous molecule (p73) has been shown to be localized at cardiac muscle cell membranes and connective tissue cells (Glenney, 1986c). Despite the presence of microvilli, p68/70 is absent from intestinal epithelial cells even though these contain very high concentrations of p36 and endonexin (Gerke and Weber, 1984).
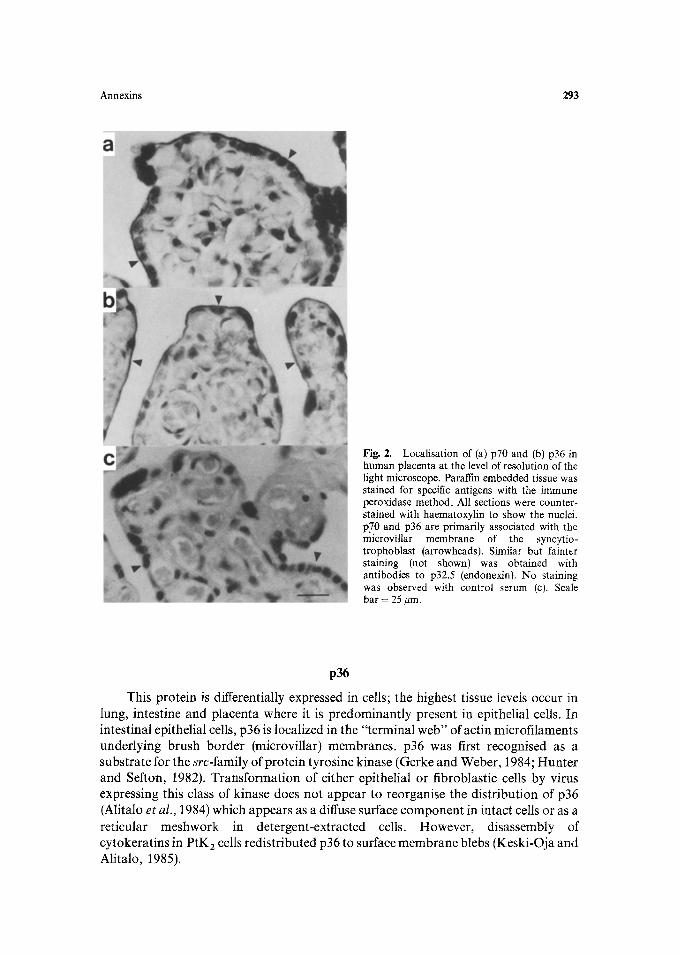
Annexins 293
Fig. 2. Localisation of (a) p70 and (b) p36 in human placenta at the level of resolution of the light microscope. Paraffin embedded tissue was stained for specific antigens with the immune peroxidase method. All sections were counter- stained with haematoxylin to show the nuclei. p70 and p36 are primarily associated with the microvillar membrane of the syncytio- trophoblast (arrowheads). Similar but fainter staining (not shown) was obtained with antibodies to p32.5 (endonexin). No staining was observed with control serum (c). Scale bar = 25 #m.
p36
This protein is differentially expressed in cells; the highest tissue levels occur in lung, intestine and placenta where it is predominant ly present in epithelial cells. In intestinal epithelial cells, p36 is localized in the "terminal web" of actin microfilaments underlying brush border (microvillar) membranes, p36 was first recognised as a substrate for the src-family of protein tyrosine kinase (Gerke and Weber, 1984; Hunte r and Sefton, 1982). Transformat ion of either epithelial or fibroblastic cells by virus expressing this class of kinase does not appear to reorganise the distribution of p36 (Alitalo et al., 1984) which appears as a diffuse surface componen t in intact cells or as a reticular meshwork in detergent-extracted cells. However , disassembly of cytokerat ins in P tK 2 cells redistributed p36 to surface membrane blebs (Keski-Oja and Alitalo, 1985).

294 Geisow, Walker, Boustead and Taylor
p35
This protein is relatively abundant in placental and lung tissue. It appears to have a cellular localization similar to p36, although there are differences in their tissue distributions. For example, p35 is not found in intestinal epithelial cells.
Endonexin
This molecule appears more widely distributed than the others (Geisow et al., 1984; Siidhof et al., 1984). In all cell types examined, the protein appears to be more generalized or cytoplasmic, however, in placenta the syncytiotrophoblast microvilli are strongly stained with specific antisera and the protein is present in isolated microvilli.
PHOSPHORYLATION OF ANNEXINS
Only p35 and p36 have been shown to be substrates for protein kinases. In A431 cells, p35 is the major substrate for the EGF-receptor tyrosine kinase (Fava and Cohen, 1984; Valentine-Braun, 1986; Pepinsky and Sinclair, 1986; Haigler et al., 1987). Conversely, p36 is not a substrate in vitro for the EGF-receptor tyrosine kinase, but is phosphorylated by the Rous sarcoma virus enzyme pp60 v-src (Gerke and Weber, 1984; Glenney and Tack, 1985). p36 is also a substrate for protein kinase C (Khanna et al., 1986; Gould et al., 1986) both in vitro and in vivo. Both p35 and p36 are phosphorylated in vitro within the amino terminal 25 amino acids. The amino terminal sequences are not homologous (Fig. 3) which explains the different substrate specificities.
The extent (Vmax) of phosphorylation on tyrosine of p35 and p36 is increased by free Ca 2 § ions without any increase in Km for the enzyme reaction (Fava and Cohen, 1984; Glenney, 1985). However, the half-maximal concentration of Ca 2 + required for p36 phosphorylation is decreased by two orders of magnitude in the presence of acidic phospholipids (Glenney, 1985). Phosphorylation of p35 at tyr 21 rendered the protein 10-fold more sensitive to tryptic cleavage at lys 26, suggesting that phosphorylation causes a protein conformation change (Schaepfer and Haigler, 1987; Haigler et al., 1987).
C-Kinase pp60~ rc 1 I 10 20 .
p36 M S T V H E I L C K L S L E G D H S T P P S A Y G S V . . �9
p35 Q A W F I E N E E Q E Y V Q T V K S S K G G P G S A V . . . 10 20
EGF-R
Fig. 3. Amino-terminal sequences of p36 and p35 showing poor homology and sites of phosphorylation by the EGF-receptor (EGF-R) and Rous sarcoma virus kinase pp60 vs'c. The serine residue phosphorylated by kinase C is also shown.

Annexins 295
INTERACTION WITH PHOSPHOLIPIDS
None of the annexins bind to liposomes made from phosphatidylcholine (PC) or PC mixtures with sphingomyelin and cholesterol at Ca z + concentrations as high as 1 mM. Addition of the phospholipids phosphatidylserine (PS), phosphatidylinositol (PI) and to a lesser extent phosphatidylethanolamine (PE) promotes Ca 2 +-dependent binding. Binding does not appear to be enhanced by diacylglycerol as observed for protein kinase C (Geisow et al., 1986). The acidic phospholipid cardiolipin promotes the Ca 2 +-dependent binding of a 33 Kd neutrophil protein to liposomes (Utsumi et al., 1986) at sub-micromolar free Ca 2 + concentrations.
p36 binds a minimum of 2 tools Ca / +/mol with a KDiss of 4.5 pM at 200 ~g/ml PS (Glenney, 1986a). p35 binds 4 mols Ca 2 § with a KDiss of 75 pM. Half-maximal binding to PS liposomes occurs at 22 #M Ca 2 +, but this Ca 2 § requirement is reduced 5-fold after phosphorylation of tyr 21 (Schaepfer and Haigler, 1987; Haigler et aI., 1987).
All annexins inhibit the activity of pancreatic phospholipase A 2 as assayed by the release of [-3H]oleic acid from labelled E. coli membranes (Elsbach et al., 1979). Although there have been a few reports that lipocortins bind to immobilized phospholipase A 2, p36 and p35 clearly inhibit this enzyme by binding the substrate membranes rather than the enzyme (Haigler et al., 1987; Davidson et al., 1987). The inhibition thus depends on inhibitor/substrate rather than inhibitor/enzyme ratio.
A key finding for this class of molecule was the shift in calcium affinity induced by the presence of phospholipid (Glenney, 1985, 1986a) and has led to the suggestion that a l:ernary complex of protein, phospholipid head group and Ca 2 § forms such that the phospholipid directly affects the co-ordination of Ca 2§ (Geisow, 1986). Multiple Ca 2 § sites and repetitive domain structure suggests the potential for binding more than one phospholipid. This probably underlies the potentiation of Ca 2 +- dependent secretion granule and phospholipid vesicle aggregation by annexins (Sfidhof et al., 1982, 1984, 1985; Johnsson et al., 1986). Interaction with phospholipids probably involves the head group rather than the fatty acid-chains. There is no evidence for a hydrophobic interaction as in the case of calmodulin-Ca z+, using fluorescent dyes to probe sites on p70, endonexin and calelectrin (Sfidhof et al., 1985; Sfidhof, 1984). Although purification of these proteins has been reported using Ca 2 + dependent binding to immobilized trifluoperazine or phenyl-Sepharose, 0.1 mM trifluoperazine did not prevent Ca 2 +-dependent binding to liposomes (Sfidhof, 1985).
INTERACTION WITH F-ACTIN
Both p35 and p36 have been shown to bind F-actin at high Ca 2 +-cOncentratiOns (Gerke and Weber, 1984, 1985; Glenney, 1986; Schlaepfer and Haigler, 1987). The interaction of p36 is unique, since this protein alone exists in the form of a tetramer comprising two identical 36Kd chains and two 11 Kd chains (p362pl12). p l l is dimeric and only in the presence of this dimer do the p36 subunits associate (Gerke and Weber, 1984). p l l is homologous with brain glial S-100 protein (Gerke and Weber, 1985b; Glenney and Tack, 1985; Hexham, 1986). p l l contains two "E-F-hand"

296 Geisow, Walker, Boustead and Taylor
potential Ca 2 +-binding sequences (Kretsinger, 1980). However, p36 rather than pl 1 appears to bind Ca 2 § (Gerke and Weber, 1985; Glenney, 1986a). pl 1 binds to p36 within the first 23 amino acids, since loss of this region (Johnsson et al., 1986) or incubation with a synthetic peptide corresponding to the amino terminus (Saris et al., 1986) blocks p l l binding to p36.
This nicely explains the lack ofa pl 1 chain in p35 and failure of this protein to bind p l l dissociated from p362pl12, since the amino terminal sequences are not homologous. The function of p l l is not clear, save that F-actin binding by the tetramer is more efficient than by isolated p36. Since p 11 does not independently bind F-actin, it may enhance p36 binding by associating two p36 monomers. The tetramer appears capable of bundling F-actin (Gerke and Weber, 1984, 1985). The high Ca 2§ requirements for F-actin binding appears to be reduced by phospholipids (Glenney, 1986b).
p36 appears to be a better substrate for Rous sarcoma virus tyrosine kinase as a monomer than in the native tetramer. It is therefore possible that phosphorylation could shift the equilibrium to favour the dissociated form and weaken binding to F- actin in vivo. In this respect it is interesting to note that both p35 and p36 are closely located to cognate kinases in stimulated cells: p35 can be phosphorylated by EGF- receptor which is abundant in placental microvilli and p36 can be phosphorylated by C-kinase and pp6ff .... which are recruited to the cell surface membrane in activated cells.
MODELS FOR PHOSPHOLIPID BINDING
The amino acid sequences clearly demonstrate the presence of a conserved, four- fold repeat in annexins of approximately 70-amino acids. This repeat contains a further two-fold homology. Structural analysis indicates that the 70-amino acid domain is composed of a pair of 0~-helices joined by a short loop and a smaller, 0~-helical linking region (Fig. 4). This predicted structure for the domain is based upon circular dichroism data and secondary and tertiary structure analysis using current prediction algorithms (Taylor and Geisow, 1987). The key aspect of the domain model is its tertiary structural similarity to bovine intestinal calcium binding protein (ICaBP). Since the high resolution crystal structure of ICaBP is known, a model has been
Fig. 4. (a) s-carbon backbone showing the proposed annexin domain structure (solid line) and the actual structure oflCaBP (broken line). (b) Schematic diagram showing location of c~- helicies.

Annexins 297
Fig. 5. Proposed interaction of the annexin domain with a phospholipid head group in the bilayer.
constructed using molecular graphics. This model has been of particular value in drawing attention to the possibility of binding a Ca 2 § ion at the first short loop in each domain. The low affinity in the absence of phospholipids would be expected for a non "E-F-hand" structure (Tufty and Kretsinger, 1975). However, the lack of the extended main chain normally present in the E-F-hand loop would now permit access of a phospholipid head group to the Ca 2 § ion to form a ternary complex as described previously (Fig. 5). Many conserved amino acids in the 70-residue domain are rationalized by such a model and further experiments need to be carried out to establish the credentials of this suggested mode of binding.
F U N C T I O N S OF T H E ANNEXINS
The localization of many of these proteins at membrane specializations like the pre- and post-synaptic regions of nerve terminals and epithelial cell brush borders suggests a structural or regulatory role in these regions. The phospholipid binding properties would allow the retention of certain phospholipids within a microdomain. Since inner bilayer lipids are predominantly bound, lipid "flipping" across the membrane, especially during Ca 2 + influx, might be restricted. By forming a protein "shell" on the inner surface of the microdomain, inappropriate membrane contact leading to fusion or damaging phospholipase activity might be regulated. Further studies, possibly involving engineered versions of the current molecules, will be needed to clarify the roles of members of this new family of Ca 2 + and phospholipid binding molecules.
REFERENCES
Alitalo, K., Ralston, R. R. and Keski-Oja, J. (1984). Exp. Cell. Res. 150:177. Blackwell, G. J., Carnuccio, R., Di Rosa, M., Flower, R. J., Parente, L. and Persico, P. (1980). Nature
287:147-149. Blackwell, G. J., Carnuccio, R., Di Rose, M., Flower, R. J., Langham, S. J., Paente, L., Persico, P., Russell-
Smith, N. C. and Stone, D. (1982). J. Pharmae. 76:185-194. Braslau, D. L., Ringo, D. L. and Rocha, V. (1984). Exp. Cell. Res. 155:213-221. Cooper, J. A. and Hunter, T. (1983). Curr. Topics Microbiol. lmmunol. 107:125-161. Creutz, C. E., Dowing, L. G., Sando, J. J., Villar-Plasi, C., Whipple, J. H. and Zaks, W. J. (1983). J. Biol.
Chem. 258:14664-14674.

298 Geisow, Walker, Boustead and Taylor
Creutz, C. E., Pazoles, C. J. and Pollard, H. B. (1978). J. Biol. Chem. 253:2858-2866. Davidson, F. F., Dennis, E. A., Powell, M. and Glenney, J. R. (1987). d. Biol. Chem. 262:1698-1705. Davis, A. A. and Crumpton, M. J. (1985). Biochem. Biophys. Res. Commun. 128:571-577. Elsbach, P., Weiss, J., Franson, R. C., Beckerdite-Quagliata, S., Schneider, A. and Harris, L. (1979). J. Biol.
Chem. 254:11000-11009. Fava, R. A. and Cohen, S. (1984). J. Biol. Chem. 259:2636-2645. Fiedler, W. and Walker, J. H. (1985). Eur. J. Cell. Biol. 38:34~1. Flower, R. J., Wood, J. N. and Parente, L. (1984). Adv. lnflamm. Res. 7:61-69. Geisow, M. J. (1986). FEBS Lett. 203:99-102. Geisow, M. J. and Walker, J. H. (1986). Trends Biochem. Sci. 11:420~23. Geisow, M. J. and Burgoyne, E. D. (1982). J. Neurochem. 38:1735-1741. Geisow, M. J., Childs, J., Dash, B., Harris, A., Panayotou, G., Siidhof, T. and Walker, J. H. (1984). EMBO
J. 3:2969-2974. Geisow, M. J., Fritsche, U., Hexharn, J. M., Dash, B. and Johnson, T. (1986). Nature 320:636438. Gerke, V. and Weber, K. (1984). EMBO J. 3:227-233. Gerke, V. and Weber, K. (1985a). J. Biol. Chem. 280:1688-1695. Gerke, V. and Weber, K. (1985b). EMBO J. 4:2917-2920. Glenney, J. R. (1985). FEBS Lett. 192:79-82. Glenney, J. R. (1986a). d. Biol. Chem. 261:7247-7252. Glenney, J. R. (1986b). Proc. Natl. Acad. Sci. 83:4258-4262. Glenney, J. R. (1986c). Exp. Cell. Res. 162:183-190. Glenney, J. R. and Tack, B. F. (t985). Proc. Natl. Acad. Sci. 82:7884--7888. Gould, K. L., Woodgett, J. R., Isacke, C. M. and Hunter, T. (1986). Mol. and Cell. Biol. 6:2738-2744. Haigler, H. T., Schlaepfer, D. D. and Burgess, W. H. (1987). J. Biol. Chem. in press. Hexham, J. M., Totty, N. F., Waterfield, M. D. and Crumpton, M. J. (1986). Biochem. Biophys. Res.
Commun. 134:248-254. Huang, K.-S. et al. (1986). Cell 46:191-199. Hunter, T. (1986). Cell 4:201-212. Hunter, T. and Sefton, B. M. (1982). In: Molecular Actions of Toxins, Viruses and Interferon (P. Cohen and S.
Van Heyningen (Eds.), Elsevier/North Holland, New York, pp. 337-370. Johnsson, N., Vandekerkhove, J., Van Damme, J. and Weber, K. (1986). FEBS Lett. 198:361-364. Keski-Oja, J. and Alitalo, K. (1985). Exp. Cell. Res. 158:86-94. Khanna, N. C., Tokada, M., Chong, S. M. and Waisman, D. M. (1986). Biochem. Biophys. Res. Commun.
137:397-403. Kretsinger, R. H. A. (1980). CRC Crit. Rev. Biochem. 8:86-94. Kristensen, T., Saris, C. J. M., Hunter, T., Hicks, L. J., Noonan, D. J., Glenney, J. R. and Tack, B. F. (1986).
Biochemistry 25:4497-4503. Moore, P. B. and Dedrnan, J. R. (1982). J. Biol. Chem. 257:9663-9667. Owens, R. J. and Crumpton, M. J. (1984). Biochem. Essays 1:61~53. Owens, R. J., Gallagher, C. J. and Crumpton, M. J. (1984). EMBO J. 3:945-952. Pepinsky, R. B. and Sinclair, L. K. (1986). Nature 321:81-84. Pollard, H. B. and Scott, J. H. FEBS Lett. 150:201-206. Reaymaekers, L., Whytack, F. and Casteels, R. (1985). Biochem. Biophys. Res. Commun. 132:526-532. Saris, C. J. M., Tack, B. F., Kristensen, T., Gienney, J. R. and Hunter, T. (1986). Cell 4:201-212. Schlaepfer, D. D. and Haigler, H. T. (1987). J. Biol. Chem. in press. Siidhof, T. C. (1984). Biochem. Biophys. Res. Commun. 123:100-107. Si~dhof, T. C., Walker, J. H. and Ocbrocki, J. (1982). EMBO J. 1:1167-1170. Siidhof, T. C., Ebbecke, M., Walker, J. H., Fritsche, U. and Boustead, C. (1984). Biochemistry 23:1103-
1109. Siidhof, T. C., Walker, J. H. and Fritsche, U. (1985). J. Neurochem. 44:1302-1307. Szebenyi, D. M. E., Obendorf, S. K. and Moffat, K. (1981). Nature 294:327-331. Taylor, V. V. and Geisow, M. J. (1987). Protein Engineering 3 (in press). Tufty, R. M. and Kretsinger, R. H. (1975). Science 187:167-169. Utsurni, K., Sato, E., Okimasu, E., Miyahara, M. and Takahasi, R. (1986). FEBS Lett. 201:277-281. Valentine-Braun, K. A., Northrup, J. K. and Hollenberg, M. D. (1986). Proc. Natl. Acad. Sci. USA 83:236-
240. Walker, J. H. (1982). J. Neurochem. 39:815-823. Wallner, B. P. et al. (1986). Nature 320:77-81.
Related Documents











