Analysis of the plant growth-promoting properties encoded by the genome of the rhizobacterium Pseudomonas putida BIRD-1 Amalia Roca, 1† Paloma Pizarro-Tobías, 1† Zulema Udaondo, 1,2† Matilde Fernández, 1 Miguel A. Matilla, 1 M. Antonia Molina-Henares, 2 Lázaro Molina, 2,3 Ana Segura, 2 Estrella Duque 2 and Juan-Luis Ramos 2 * 1 Polígono Industrial Juncaril, Bio-Iliberis R&D, 18210 Peligros, Granada, Spain. 2 Estación Experimental del Zadín, CSIC, 18008 Granada, Granada, Spain. 3 CIDERTA, Universidad de Huelva, Huelva, Spain. Summary Pseudomonas putida BIRD-1 is a plant growth- promoting rhizobacterium whose genome size is 5.7 Mbp. It adheres to plant roots and colonizes the rhizosphere to high cell densities even in soils with low moisture. This property is linked to its ability to synthesize trehalose, since a mutant deficient in the synthesis of trehalose exhibited less tolerance to des- iccation than the parental strain. The genome of BIRD-1 encodes a wide range of proteins that help it to deal with reactive oxygen stress generated in the plant rhizosphere. BIRD-1 plant growth-promoting rhizobacteria properties derive from its ability to enhance phosphorous and iron solubilization and to produce phytohormones. BIRD-1 is capable of solu- bilizing insoluble inorganic phosphate forms through acid production. The genome of BIRD-1 encodes at least five phosphatases related to phosphorous solu- bilization, one of them being a phytase that facilitates the utilization of phytic acid, the main storage form of phosphorous in plants. Pyoverdine is the siderophore produced by this strain, a mutant that in the FvpD siderophore synthase failed to grow on medium without supplementary iron, but the mutant was as competitive as the parental strain in soils because it captures the siderophores produced by other microbes. BIRD-1 overproduces indole-3-acetic acid through convergent pathways. Introduction The rhizosphere is a very dynamic environment in which complex interactions take place between plant roots and microorganisms (Molina et al., 2000; Uroz et al., 2010; Blom et al., 2011). Management of the rhizosphere repre- sents a genuine opportunity for biotechnology since it can increase the levels of crop production while reducing the use of fertilizers and agrochemicals in agriculture. Plant growth-promoting rhizobacteria (PGPR) efficiently colo- nize the rhizosphere of a wide range of plants and stimu- late plant growth through direct or indirect mechanisms (Kloepper et al., 1980; Glick, 1995; Somers and Vander- leyden, 2004; Lugtenberg and Kamilova, 2009; Matilla et al., 2010; Blom et al., 2011). Direct plant growth pro- motion is often mediated by the synthesis of phytohor- mones or enhanced nutrition, e.g. by increasing the bioavailability of mineral nutrients such as phosphorous, nitrogen and iron (Lugtenberg and Kamilova, 2009), or by providing amino acids and other nutritional factors (Simmons et al., 1997; Compant et al., 2010; Vial et al., 2011). Many PGPR produce siderophores, which are small metal binding molecules which mainly chelate Fe(III) and other metals with lower affinity (Schalk et al., 2011). The siderophore–metal complexes are preferen- tially used by microbes (van Loon, 2007), but they can also be taken up by plants (Lemanceau et al., 2009; Gamalero and Glick, 2011). Plant growth promotion by PGPR can also be mediated by a number of indirect effects: (i) by avoiding the action of soil-borne pathogens through competition for iron, (ii) through the synthesis of antibiotics, antifungals and pesticides (Hammer et al., 1997; Lugtenberg and Kamilova, 2009; Pérez et al., 2011; Ahemad and Khan, 2012) and (iii) through the production of biocides such as hydrogen cyanide and fungal cell wall-degrading enzymes, e.g. chitinase and b-1,3- glucanase (Zhang and Yuen, 2000; Haas and Keel, 2003; Meschke and Schrempf, 2010; Malfanova et al., 2011). Some non-pathogenic rhizobacteria have been reported to suppress diseases in plants by triggering induced sys- temic resistance (ISR), a plant response that depends on Received 31 August, 2012; revised 17 October, 2012; accepted 23 October, 2012. *For correspondence. E-mail juanluis.ramos@ eez.csic.es; Tel. (+34) 958 181 608; Fax (+34) 958 135 740. † These co-authors have equally contributed to this study. Environmental Microbiology (2012) doi:10.1111/1462-2920.12037 © 2012 Society for Applied Microbiology and Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Analysis of the plant growth-promoting propertiesencoded by the genome of the rhizobacteriumPseudomonas putida BIRD-1
Amalia Roca,1† Paloma Pizarro-Tobías,1†
Zulema Udaondo,1,2† Matilde Fernández,1
Miguel A. Matilla,1 M. Antonia Molina-Henares,2
Lázaro Molina,2,3 Ana Segura,2 Estrella Duque2 andJuan-Luis Ramos2*1Polígono Industrial Juncaril, Bio-Iliberis R&D, 18210Peligros, Granada, Spain.2Estación Experimental del Zadín, CSIC, 18008Granada, Granada, Spain.3CIDERTA, Universidad de Huelva, Huelva, Spain.
Summary
Pseudomonas putida BIRD-1 is a plant growth-promoting rhizobacterium whose genome size is5.7 Mbp. It adheres to plant roots and colonizes therhizosphere to high cell densities even in soils withlow moisture. This property is linked to its ability tosynthesize trehalose, since a mutant deficient in thesynthesis of trehalose exhibited less tolerance to des-iccation than the parental strain. The genome ofBIRD-1 encodes a wide range of proteins that help itto deal with reactive oxygen stress generated in theplant rhizosphere. BIRD-1 plant growth-promotingrhizobacteria properties derive from its ability toenhance phosphorous and iron solubilization and toproduce phytohormones. BIRD-1 is capable of solu-bilizing insoluble inorganic phosphate forms throughacid production. The genome of BIRD-1 encodes atleast five phosphatases related to phosphorous solu-bilization, one of them being a phytase that facilitatesthe utilization of phytic acid, the main storage form ofphosphorous in plants. Pyoverdine is the siderophoreproduced by this strain, a mutant that in the FvpDsiderophore synthase failed to grow on mediumwithout supplementary iron, but the mutant was ascompetitive as the parental strain in soils becauseit captures the siderophores produced by other
microbes. BIRD-1 overproduces indole-3-acetic acidthrough convergent pathways.
Introduction
The rhizosphere is a very dynamic environment in whichcomplex interactions take place between plant roots andmicroorganisms (Molina et al., 2000; Uroz et al., 2010;Blom et al., 2011). Management of the rhizosphere repre-sents a genuine opportunity for biotechnology since it canincrease the levels of crop production while reducing theuse of fertilizers and agrochemicals in agriculture. Plantgrowth-promoting rhizobacteria (PGPR) efficiently colo-nize the rhizosphere of a wide range of plants and stimu-late plant growth through direct or indirect mechanisms(Kloepper et al., 1980; Glick, 1995; Somers and Vander-leyden, 2004; Lugtenberg and Kamilova, 2009; Matillaet al., 2010; Blom et al., 2011). Direct plant growth pro-motion is often mediated by the synthesis of phytohor-mones or enhanced nutrition, e.g. by increasing thebioavailability of mineral nutrients such as phosphorous,nitrogen and iron (Lugtenberg and Kamilova, 2009), or byproviding amino acids and other nutritional factors(Simmons et al., 1997; Compant et al., 2010; Vial et al.,2011). Many PGPR produce siderophores, which aresmall metal binding molecules which mainly chelateFe(III) and other metals with lower affinity (Schalk et al.,2011). The siderophore–metal complexes are preferen-tially used by microbes (van Loon, 2007), but they canalso be taken up by plants (Lemanceau et al., 2009;Gamalero and Glick, 2011). Plant growth promotion byPGPR can also be mediated by a number of indirecteffects: (i) by avoiding the action of soil-borne pathogensthrough competition for iron, (ii) through the synthesis ofantibiotics, antifungals and pesticides (Hammer et al.,1997; Lugtenberg and Kamilova, 2009; Pérez et al., 2011;Ahemad and Khan, 2012) and (iii) through the productionof biocides such as hydrogen cyanide and fungal cellwall-degrading enzymes, e.g. chitinase and b-1,3-glucanase (Zhang and Yuen, 2000; Haas and Keel, 2003;Meschke and Schrempf, 2010; Malfanova et al., 2011).Some non-pathogenic rhizobacteria have been reportedto suppress diseases in plants by triggering induced sys-temic resistance (ISR), a plant response that depends on
Received 31 August, 2012; revised 17 October, 2012; accepted 23October, 2012. *For correspondence. E-mail [email protected]; Tel. (+34) 958 181 608; Fax (+34) 958 135 740. †Theseco-authors have equally contributed to this study.
bs_bs_banner
Environmental Microbiology (2012) doi:10.1111/1462-2920.12037
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd

jasmonic acid and on the plant’s ethylene signalling path-ways (van Loon and Bakker, 2005; Bakker et al., 2007).Several PGPR are commercially available for applicationas crop inoculants and for biofertilization, phytostimula-tion, biocontrol and rhizoremediation (Nakkeeran et al.,2005; Berg, 2009; Segura et al., 2009; Kroll et al., 2010).
Bacteria of the genus Pseudomonas are frequentrhizosphere colonizers and some strains of the speciesPseudomonas fluorescens, Pseudomonas putida, Pseu-domonas mendocina and Pseudomonas stutzeri havebeen shown to possess PGPR capabilities (reviewed byWu et al., 2010). Strains of the species P. putida areacknowledged as one of the most efficient phosphate-solubilizing cultivable bacteria (Rodríguez and Fraga,1999; Vyas and Gulati, 2009; Daniels et al., 2010), andamong them, the BIRD-1 strain is particularly efficient.This strain was isolated from a garden soil in a culturemedium without iron addition and with insoluble inorganicphosphate as a source of phosphorous (Matilla et al.,2011). The genome sequence of P. putida BIRD-1 wasobtained using the 454 pyrosequencing technology(Matilla et al., 2011). The complete genome sequence ofP. putida BIRD-1 is made up of a 61.74% GC circularchromosome of 5 731 541 bp with no plasmid. Its genomesize is similar to that of P. putida strain W619(5 774 330 bp) (Taghavi et al., 2009; Wu et al., 2010),but significantly smaller than that of P. putida strainsKT2440 (6 181 860 bp) (Nelson et al., 2002), F1(5 959 964 bp) (GenBank Accession CP000712) andGB-1 (6 078 430 bp) (GenBank Accession CP000926).The genome of BIRD-1 shows high homology andsynteny with that of KT2440 and that of the F1 strain. Allthree strains share around 80% of all of their genes.
In this study we analysed the genome of P. putidaBIRD-1 in relation to its PGPR properties and the mostrelevant data are presented.
Results and discussion
Adhesion and colonization of plant roots
An early event in the process of plant root colonization bybacteria is their ability to adhere to biotic surfaces – eitherto the seed surface or to the root. With BIRD-1 our assaysshowed that approximately 104 to 105 cfu of P. putidaBIRD-1 cells adhered per corn seed, a similar amount tothat reported for the KT2440 strain (Espinosa-Urgel et al.,2000; Weinel et al., 2002; Molina-Henares et al., 2006;Yousef-Coronado et al., 2008). Eleven genes found to beinvolved in adhesion to abiotic and biotic surfaces inP. putida KT2440 (Duque et al., 2012) were identified inBIRD-1 (Table S1); their degree of identity was in therange of 40% to 100%. Mutants of BIRD-1 in LapA were10-fold less efficient in adhesion to seeds than the paren-
tal strain, and exhibited decreased ability in rhizospherecolonization in competition with the wild-type strain.
Proliferation of BIRD-1 in the plant rhizosphere isfavoured by the ability of the strain to use compoundssecreted by the plant, among which are the 20 proteino-genic amino acids, organic acids (citric, lactic), sugars(glucose, fructose) and flavonoids. Phenomic assays witha wide battery of carbon sources revealed that P. putidaBIRD-1 is able to use a wide range of amino acids ascarbon or nitrogen sources and has the complete set ofgenes required for the metabolism of proline, glutamate,glutamine, asparagine, tyrosine, valine, branched aminoacids, serine and alanine among the amino acids. Thestrain can use citrate and succinate, chemicals alsopresent in root exudates, as well as a wide range oflignin-derived compounds including hydroxylated aro-matic acids and methoxylated compounds. All of thesepathways are common to KT2440 and gene analysisshowed sequence and gene order conservation, an issuerevised before by Jiménez and colleagues (2002) andBielecki and colleagues (2011).
Plant growth-promoting rhizobacteria properties areparticularly relevant in marginal soils or when plants areexposed to abiotic or biotic stresses. Water scarcity is acommon problem in soils in the Mediterranean basin.We tested the survival of P. putida BIRD-1 in soils withdifferent degrees of humidity using P. putida KT2440 as areference strain for these studies. We introduced approxi-mately 5 ¥ 107 cfu BIRD-1 or KT2440 per gram of soil andsoil moisture was kept at 2%, 10%, 25% and 50%. At amoisture level of > 25% survival of both strains over a16 day period was similar with a slight increase in cfu g-1
reaching levels in the order of 2 to 4 ¥ 108 cfu g-1 soil.When the soil moisture was kept at 2%, BIRD-1 survivedat a higher density than KT2440 (Fig. 1). Therefore,BIRD-1 is more tolerant to desiccation than P. putidaKT2440 (Muñoz-Rojas et al., 2006). Trehalose has beensuggested to be important for the survival of P. putida atlow-humidity rates. Biosynthesis of trehalose in Pseu-domonas can take place through two pathways thatinvolve the conversion of maltose into trehalose viaTreS [trehalose synthases (PPU_BIRD-1_1765) andPPU_BIRD-1_2817 or from maltodextrine catalysed bythe TreY (PPU_BIRD-1_1772)/TreZ (PPU_BIRD-1_1774)proteins]. To test the role of trehalose production in resist-ance to desiccation in BIRD-1 we generated mutants intreZ and treS.
In the low-humidity soil the initial survival of the treZmutant was clearly lower than that of the parental strain orthat of mutants in the treS gene (Fig. 1), suggesting thatthe maltodextrine pathway for the production of trehaloseis relevant for colonization of soils by P. putida.
Matilla and colleagues (2007) showed that reactiveoxygen species are produced in the plant rhizosphere as
2 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

a consequence of plant root cell respiration, and that rootcolonization by bacteria requires the ability to overcomeoxidative stress. To test resistance to oxidative stress inBIRD-1, a series of phenomic analyses were carried out inP. putida BIRD-1 with methylviologen, H2O2, chromate,arseniate and antibiotics such as b-lactams that inducethe formation of reactive oxygen species (Sandermann,2004). The results showed that P. putida BIRD-1 reachedsimilarly high cell densities in cultures without the stressoror with 1 mM methylviologen, 4 mM H2O2, 1 mM Cr2O7
and 100 mg ml-1 ampicillin. The genome of BIRD-1 wasthen analysed in relation to the set of genes involved inoxidative stress responses, and it was found to encodetwo iron–manganese SodA superoxide dismutases(PPU_BIRD-1_0966 and PPU_BIRD-1_0999) and fourcatalases (PPU_BIRD-1_0143, 0518, 2060 and 2848). Inaddition, we searched for the presence of alkylhydroper-oxidases, cytochrome-c peroxidase and glutathione per-oxidase. We found six homologues for the first group(PPU_BIRD-1_1273, 2010, 2150, 2706, 2708, 3265and 3243), a single cytochrome peroxidase (PPU_BIRD-1_2732) and two glutathione peroxidases (PPU_BIRD-1_3741 and 3931). The redundancy of the genesencoding different isoenzymes related to oxidative stressin the genome of P. putida BIRD-1 probably pointstowards the importance of these proteins and their likelysynergic action in the removal of active oxygen species.
Evidence of plant growth-promoting activities encodedby BIRD-1
In another set of assays we tested the stimulatory effect ofBIRD-1 on growth of maize roots on agar plates as amodel. To this end sterilized corn seeds were depositedon the surface of a water agar plate either inoculated ornot with P. putida BIRD-1 (106 cfu cm-2 of surface plate).Germination of corn seeds was monitored over time andthe size of the primary root was observed to be larger forseeds on agar surfaces with bacteria than in the absenceof microbes at any incubation time (Fig. 2), and secondaryroots also developed more regularly in seeds germinatedwith bacteria than without.
Fig. 1. Viable P. putida KT2440, BIRD-1 and a TreZ mutant inrhizosphere soil. Soils were inoculated with ~ 5 ¥ 107 cfu g-1 andsterile pre-germinated corn seeds sown. At the indicated timecfu g-1 rhizosphere soil was determined using three different pots.Black bars (BIRD-1), light grey (KT2440) and dark grey (treZBIRD-1 mutant). Other conditions are as described in Experimentalprocedures.
Fig. 2. Root development of corn seedsgerminated on agar plates. Water agar plateswere prepared, one plate was kept as acontrol (A) and 0.1 ml of a suspensioncontaining 108 cfu ml-1 was spread on top ofthe other plate (B). Once the surfaces weredry, a single sterile corn seed was depositedper agar plate and root developmentmonitored over time. The picture correspondsto a typical situation after 1 week incubationat 22°C.
PGPR properties 3
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
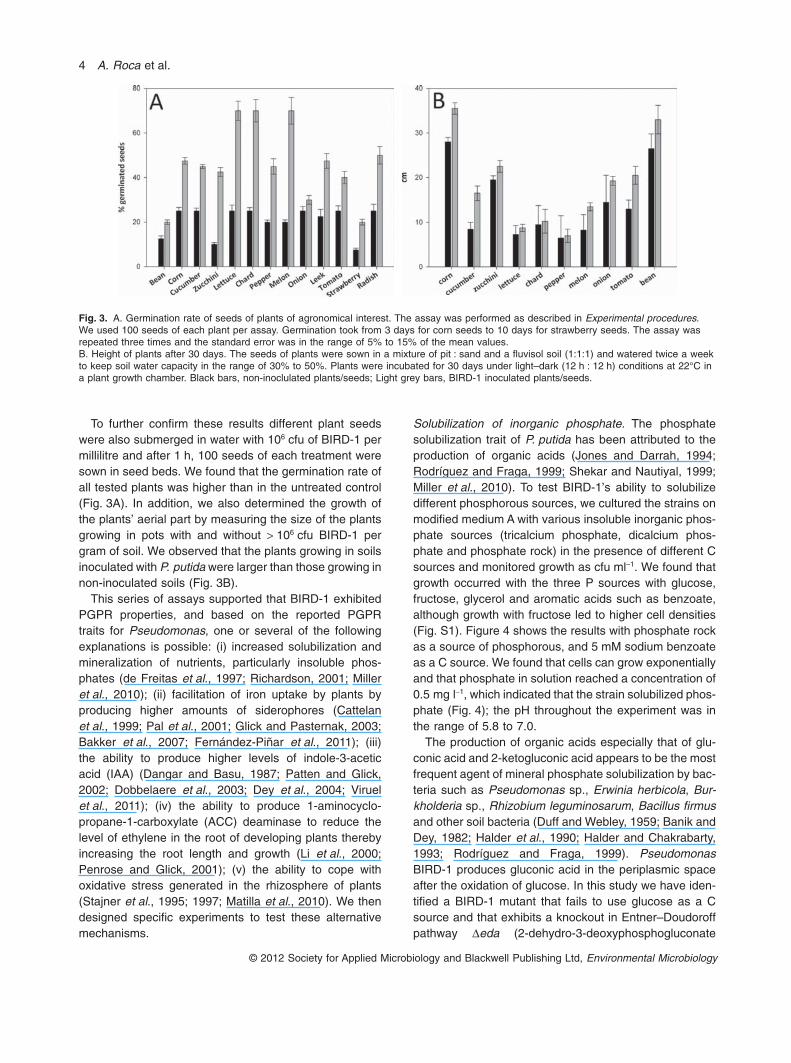
To further confirm these results different plant seedswere also submerged in water with 106 cfu of BIRD-1 permillilitre and after 1 h, 100 seeds of each treatment weresown in seed beds. We found that the germination rate ofall tested plants was higher than in the untreated control(Fig. 3A). In addition, we also determined the growth ofthe plants’ aerial part by measuring the size of the plantsgrowing in pots with and without > 106 cfu BIRD-1 pergram of soil. We observed that the plants growing in soilsinoculated with P. putida were larger than those growing innon-inoculated soils (Fig. 3B).
This series of assays supported that BIRD-1 exhibitedPGPR properties, and based on the reported PGPRtraits for Pseudomonas, one or several of the followingexplanations is possible: (i) increased solubilization andmineralization of nutrients, particularly insoluble phos-phates (de Freitas et al., 1997; Richardson, 2001; Milleret al., 2010); (ii) facilitation of iron uptake by plants byproducing higher amounts of siderophores (Cattelanet al., 1999; Pal et al., 2001; Glick and Pasternak, 2003;Bakker et al., 2007; Fernández-Piñar et al., 2011); (iii)the ability to produce higher levels of indole-3-aceticacid (IAA) (Dangar and Basu, 1987; Patten and Glick,2002; Dobbelaere et al., 2003; Dey et al., 2004; Viruelet al., 2011); (iv) the ability to produce 1-aminocyclo-propane-1-carboxylate (ACC) deaminase to reduce thelevel of ethylene in the root of developing plants therebyincreasing the root length and growth (Li et al., 2000;Penrose and Glick, 2001); (v) the ability to cope withoxidative stress generated in the rhizosphere of plants(Stajner et al., 1995; 1997; Matilla et al., 2010). We thendesigned specific experiments to test these alternativemechanisms.
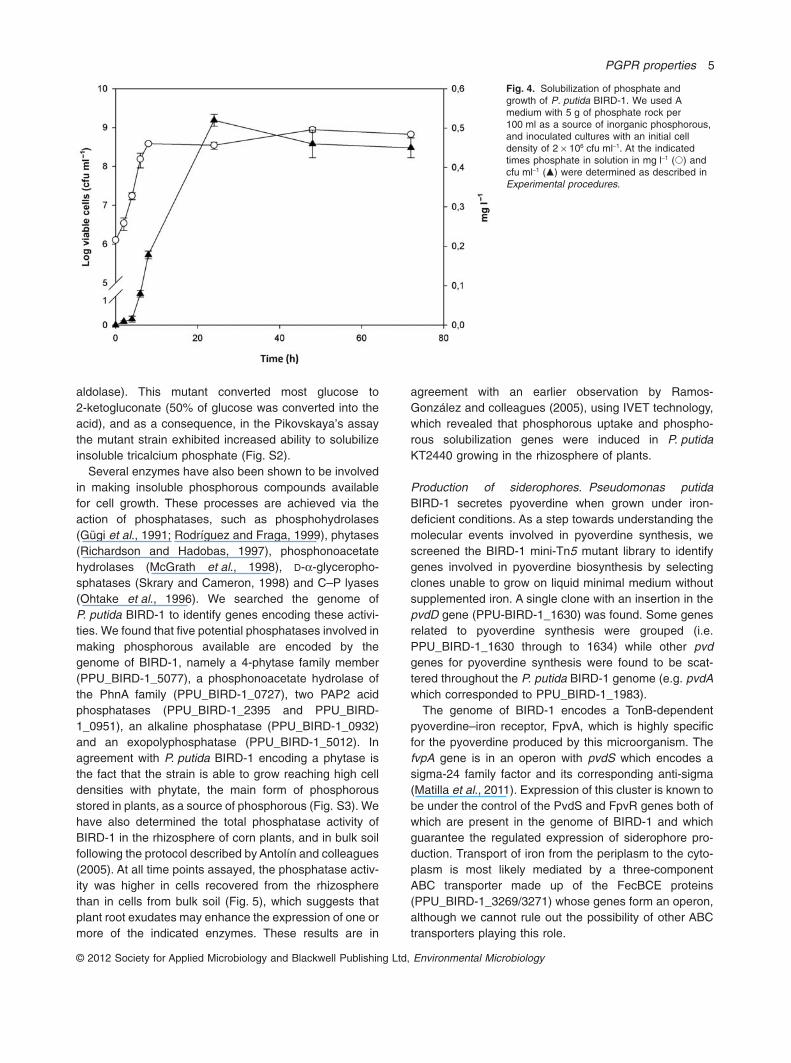
Solubilization of inorganic phosphate. The phosphatesolubilization trait of P. putida has been attributed to theproduction of organic acids (Jones and Darrah, 1994;Rodríguez and Fraga, 1999; Shekar and Nautiyal, 1999;Miller et al., 2010). To test BIRD-1’s ability to solubilizedifferent phosphorous sources, we cultured the strains onmodified medium A with various insoluble inorganic phos-phate sources (tricalcium phosphate, dicalcium phos-phate and phosphate rock) in the presence of different Csources and monitored growth as cfu ml-1. We found thatgrowth occurred with the three P sources with glucose,fructose, glycerol and aromatic acids such as benzoate,although growth with fructose led to higher cell densities(Fig. S1). Figure 4 shows the results with phosphate rockas a source of phosphorous, and 5 mM sodium benzoateas a C source. We found that cells can grow exponentiallyand that phosphate in solution reached a concentration of0.5 mg l-1, which indicated that the strain solubilized phos-phate (Fig. 4); the pH throughout the experiment was inthe range of 5.8 to 7.0.
The production of organic acids especially that of glu-conic acid and 2-ketogluconic acid appears to be the mostfrequent agent of mineral phosphate solubilization by bac-teria such as Pseudomonas sp., Erwinia herbicola, Bur-kholderia sp., Rhizobium leguminosarum, Bacillus firmusand other soil bacteria (Duff and Webley, 1959; Banik andDey, 1982; Halder et al., 1990; Halder and Chakrabarty,1993; Rodríguez and Fraga, 1999). PseudomonasBIRD-1 produces gluconic acid in the periplasmic spaceafter the oxidation of glucose. In this study we have iden-tified a BIRD-1 mutant that fails to use glucose as a Csource and that exhibits a knockout in Entner–Doudoroffpathway Deda (2-dehydro-3-deoxyphosphogluconate
Fig. 3. A. Germination rate of seeds of plants of agronomical interest. The assay was performed as described in Experimental procedures.We used 100 seeds of each plant per assay. Germination took from 3 days for corn seeds to 10 days for strawberry seeds. The assay wasrepeated three times and the standard error was in the range of 5% to 15% of the mean values.B. Height of plants after 30 days. The seeds of plants were sown in a mixture of pit : sand and a fluvisol soil (1:1:1) and watered twice a weekto keep soil water capacity in the range of 30% to 50%. Plants were incubated for 30 days under light–dark (12 h : 12 h) conditions at 22°C ina plant growth chamber. Black bars, non-inoclulated plants/seeds; Light grey bars, BIRD-1 inoculated plants/seeds.
4 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
aldolase). This mutant converted most glucose to2-ketogluconate (50% of glucose was converted into theacid), and as a consequence, in the Pikovskaya’s assaythe mutant strain exhibited increased ability to solubilizeinsoluble tricalcium phosphate (Fig. S2).
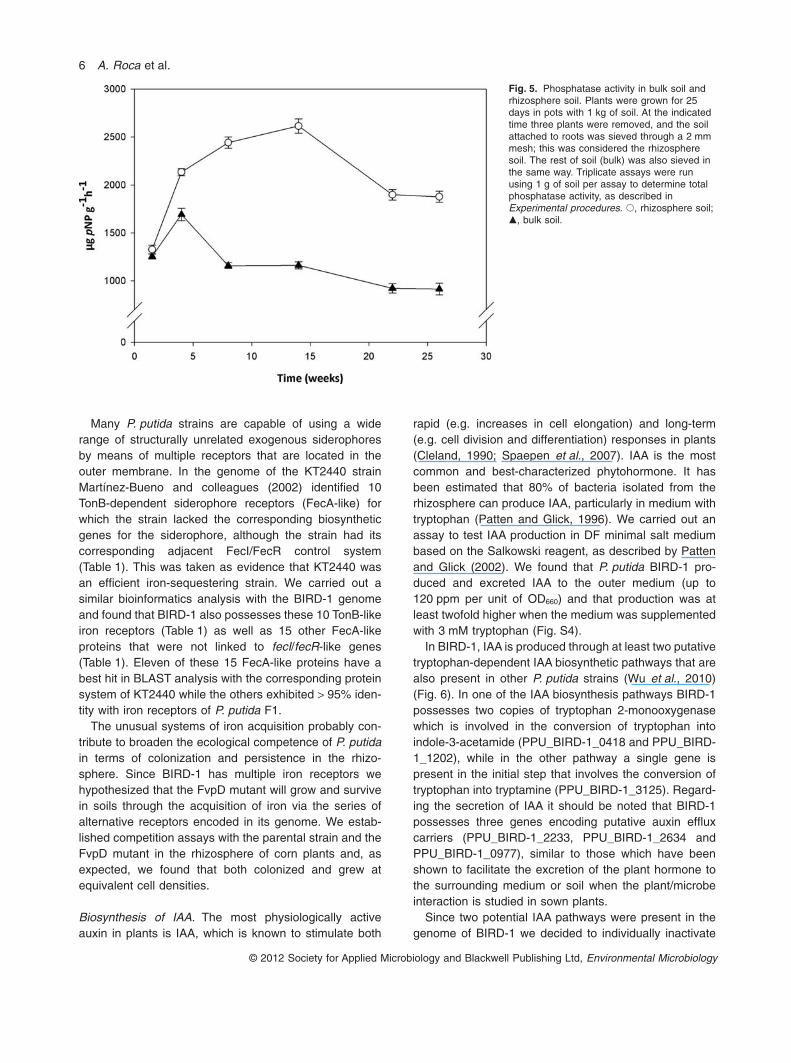
Several enzymes have also been shown to be involvedin making insoluble phosphorous compounds availablefor cell growth. These processes are achieved via theaction of phosphatases, such as phosphohydrolases(Gügi et al., 1991; Rodríguez and Fraga, 1999), phytases(Richardson and Hadobas, 1997), phosphonoacetatehydrolases (McGrath et al., 1998), D-a-glyceropho-sphatases (Skrary and Cameron, 1998) and C–P lyases(Ohtake et al., 1996). We searched the genome ofP. putida BIRD-1 to identify genes encoding these activi-ties. We found that five potential phosphatases involved inmaking phosphorous available are encoded by thegenome of BIRD-1, namely a 4-phytase family member(PPU_BIRD-1_5077), a phosphonoacetate hydrolase ofthe PhnA family (PPU_BIRD-1_0727), two PAP2 acidphosphatases (PPU_BIRD-1_2395 and PPU_BIRD-1_0951), an alkaline phosphatase (PPU_BIRD-1_0932)and an exopolyphosphatase (PPU_BIRD-1_5012). Inagreement with P. putida BIRD-1 encoding a phytase isthe fact that the strain is able to grow reaching high celldensities with phytate, the main form of phosphorousstored in plants, as a source of phosphorous (Fig. S3). Wehave also determined the total phosphatase activity ofBIRD-1 in the rhizosphere of corn plants, and in bulk soilfollowing the protocol described by Antolín and colleagues(2005). At all time points assayed, the phosphatase activ-ity was higher in cells recovered from the rhizospherethan in cells from bulk soil (Fig. 5), which suggests thatplant root exudates may enhance the expression of one ormore of the indicated enzymes. These results are in
agreement with an earlier observation by Ramos-González and colleagues (2005), using IVET technology,which revealed that phosphorous uptake and phospho-rous solubilization genes were induced in P. putidaKT2440 growing in the rhizosphere of plants.
Production of siderophores. Pseudomonas putidaBIRD-1 secretes pyoverdine when grown under iron-deficient conditions. As a step towards understanding themolecular events involved in pyoverdine synthesis, wescreened the BIRD-1 mini-Tn5 mutant library to identifygenes involved in pyoverdine biosynthesis by selectingclones unable to grow on liquid minimal medium withoutsupplemented iron. A single clone with an insertion in thepvdD gene (PPU-BIRD-1_1630) was found. Some genesrelated to pyoverdine synthesis were grouped (i.e.PPU_BIRD-1_1630 through to 1634) while other pvdgenes for pyoverdine synthesis were found to be scat-tered throughout the P. putida BIRD-1 genome (e.g. pvdAwhich corresponded to PPU_BIRD-1_1983).
The genome of BIRD-1 encodes a TonB-dependentpyoverdine–iron receptor, FpvA, which is highly specificfor the pyoverdine produced by this microorganism. ThefvpA gene is in an operon with pvdS which encodes asigma-24 family factor and its corresponding anti-sigma(Matilla et al., 2011). Expression of this cluster is known tobe under the control of the PvdS and FpvR genes both ofwhich are present in the genome of BIRD-1 and whichguarantee the regulated expression of siderophore pro-duction. Transport of iron from the periplasm to the cyto-plasm is most likely mediated by a three-componentABC transporter made up of the FecBCE proteins(PPU_BIRD-1_3269/3271) whose genes form an operon,although we cannot rule out the possibility of other ABCtransporters playing this role.
Fig. 4. Solubilization of phosphate andgrowth of P. putida BIRD-1. We used Amedium with 5 g of phosphate rock per100 ml as a source of inorganic phosphorous,and inoculated cultures with an initial celldensity of 2 ¥ 106 cfu ml-1. At the indicatedtimes phosphate in solution in mg l-1 (�) andcfu ml-1 (�) were determined as described inExperimental procedures.
PGPR properties 5
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
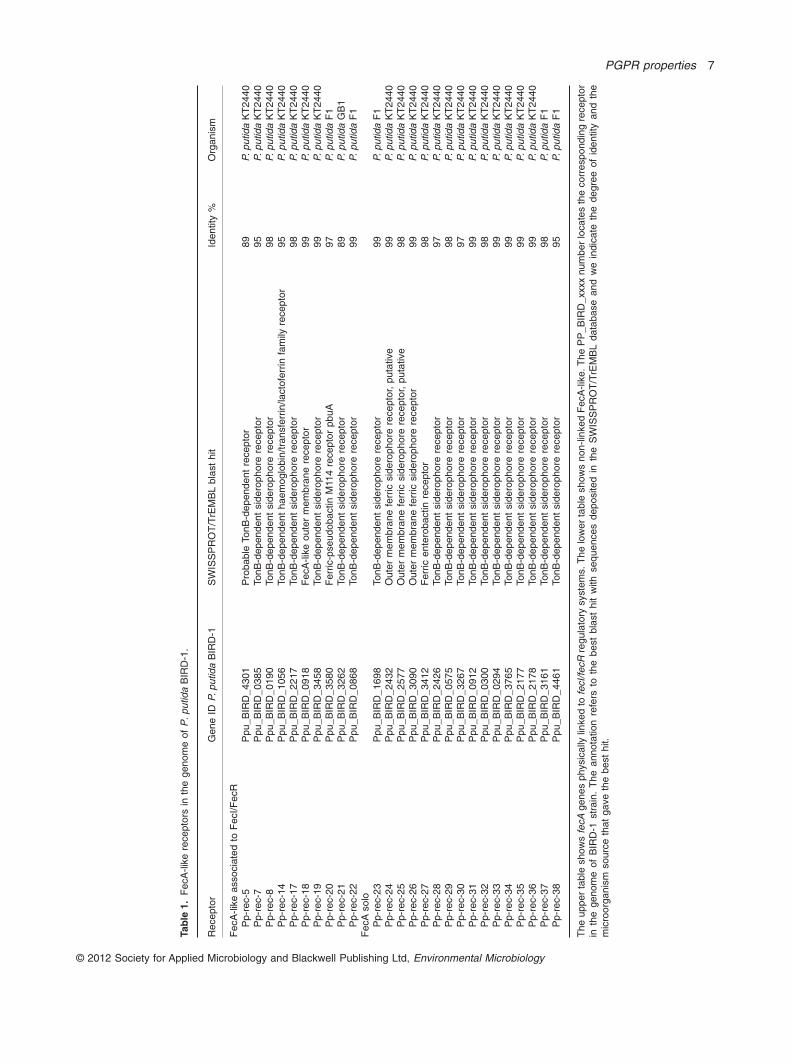
Many P. putida strains are capable of using a widerange of structurally unrelated exogenous siderophoresby means of multiple receptors that are located in theouter membrane. In the genome of the KT2440 strainMartínez-Bueno and colleagues (2002) identified 10TonB-dependent siderophore receptors (FecA-like) forwhich the strain lacked the corresponding biosyntheticgenes for the siderophore, although the strain had itscorresponding adjacent FecI/FecR control system(Table 1). This was taken as evidence that KT2440 wasan efficient iron-sequestering strain. We carried out asimilar bioinformatics analysis with the BIRD-1 genomeand found that BIRD-1 also possesses these 10 TonB-likeiron receptors (Table 1) as well as 15 other FecA-likeproteins that were not linked to fecI/fecR-like genes(Table 1). Eleven of these 15 FecA-like proteins have abest hit in BLAST analysis with the corresponding proteinsystem of KT2440 while the others exhibited > 95% iden-tity with iron receptors of P. putida F1.
The unusual systems of iron acquisition probably con-tribute to broaden the ecological competence of P. putidain terms of colonization and persistence in the rhizo-sphere. Since BIRD-1 has multiple iron receptors wehypothesized that the FvpD mutant will grow and survivein soils through the acquisition of iron via the series ofalternative receptors encoded in its genome. We estab-lished competition assays with the parental strain and theFvpD mutant in the rhizosphere of corn plants and, asexpected, we found that both colonized and grew atequivalent cell densities.
Biosynthesis of IAA. The most physiologically activeauxin in plants is IAA, which is known to stimulate both
rapid (e.g. increases in cell elongation) and long-term(e.g. cell division and differentiation) responses in plants(Cleland, 1990; Spaepen et al., 2007). IAA is the mostcommon and best-characterized phytohormone. It hasbeen estimated that 80% of bacteria isolated from therhizosphere can produce IAA, particularly in medium withtryptophan (Patten and Glick, 1996). We carried out anassay to test IAA production in DF minimal salt mediumbased on the Salkowski reagent, as described by Pattenand Glick (2002). We found that P. putida BIRD-1 pro-duced and excreted IAA to the outer medium (up to120 ppm per unit of OD660) and that production was atleast twofold higher when the medium was supplementedwith 3 mM tryptophan (Fig. S4).
In BIRD-1, IAA is produced through at least two putativetryptophan-dependent IAA biosynthetic pathways that arealso present in other P. putida strains (Wu et al., 2010)(Fig. 6). In one of the IAA biosynthesis pathways BIRD-1possesses two copies of tryptophan 2-monooxygenasewhich is involved in the conversion of tryptophan intoindole-3-acetamide (PPU_BIRD-1_0418 and PPU_BIRD-1_1202), while in the other pathway a single gene ispresent in the initial step that involves the conversion oftryptophan into tryptamine (PPU_BIRD-1_3125). Regard-ing the secretion of IAA it should be noted that BIRD-1possesses three genes encoding putative auxin effluxcarriers (PPU_BIRD-1_2233, PPU_BIRD-1_2634 andPPU_BIRD-1_0977), similar to those which have beenshown to facilitate the excretion of the plant hormone tothe surrounding medium or soil when the plant/microbeinteraction is studied in sown plants.
Since two potential IAA pathways were present in thegenome of BIRD-1 we decided to individually inactivate
Fig. 5. Phosphatase activity in bulk soil andrhizosphere soil. Plants were grown for 25days in pots with 1 kg of soil. At the indicatedtime three plants were removed, and the soilattached to roots was sieved through a 2 mmmesh; this was considered the rhizospheresoil. The rest of soil (bulk) was also sieved inthe same way. Triplicate assays were runusing 1 g of soil per assay to determine totalphosphatase activity, as described inExperimental procedures. �, rhizosphere soil;�, bulk soil.
6 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
Tab
le1.
Fec
A-li
kere
cept
ors
inth
ege
nom
eof
P.p
utid
aB
IRD
-1.
Rec
epto
rG
ene
IDP.
putid
aB
IRD
-1S
WIS
SP
RO
T/T
rEM
BL
blas
thi
tId
entit
y%
Org
anis
m
Fec
A-li
keas
soci
ated
toF
ecI/F
ecR
Pp-
rec-
5P
pu_B
IRD
_430
1P
roba
ble
TonB
-dep
ende
ntre
cept
or89
P.pu
tida
KT
2440
Pp-
rec-
7P
pu_B
IRD
_038
5To
nB-d
epen
dent
side
roph
ore
rece
ptor
95P.
putid
aK
T24
40P
p-re
c-8
Ppu
_BIR
D_0
190
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or98
P.pu
tida
KT
2440
Pp-
rec-
14P
pu_B
IRD
_105
6To
nB-d
epen
dent
haem
oglo
bin/
tran
sfer
rin/la
ctof
errin
fam
ilyre
cept
or95
P.pu
tida
KT
2440
Pp-
rec-
17P
pu_B
IRD
_221
7To
nB-d
epen
dent
side
roph
ore
rece
ptor
98P.
putid
aK
T24
40P
p-re
c-18
Ppu
_BIR
D_0
918
Fec
A-li
keou
ter
mem
bran
ere
cept
or99
P.pu
tida
KT
2440
Pp-
rec-
19P
pu_B
IRD
_345
8To
nB-d
epen
dent
side
roph
ore
rece
ptor
99P.
putid
aK
T24
40P
p-re
c-20
Ppu
_BIR
D_3
580
Fer
ric-p
seud
obac
tinM
114
rece
ptor
pbuA
97P.
putid
aF
1P
p-re
c-21
Ppu
_BIR
D_3
262
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or89
P.pu
tida
GB
1P
p-re
c-22
Ppu
_BIR
D_0
868
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or99
P.pu
tida
F1
Fec
Aso
loP
p-re
c-23
Ppu
_BIR
D_1
698
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or99
P.pu
tida
F1
Pp-
rec-
24P
pu_B
IRD
_243
2O
uter
mem
bran
efe
rric
side
roph
ore
rece
ptor
,pu
tativ
e99
P.pu
tida
KT
2440
Pp-
rec-
25P
pu_B
IRD
_257
7O
uter
mem
bran
efe
rric
side
roph
ore
rece
ptor
,pu
tativ
e98
P.pu
tida
KT
2440
Pp-
rec-
26P
pu_B
IRD
_309
0O
uter
mem
bran
efe
rric
side
roph
ore
rece
ptor
99P.
putid
aK
T24
40P
p-re
c-27
Ppu
_BIR
D_3
412
Fer
ricen
tero
bact
inre
cept
or98
P.pu
tida
KT
2440
Pp-
rec-
28P
pu_B
IRD
_242
6To
nB-d
epen
dent
side
roph
ore
rece
ptor
97P.
putid
aK
T24
40P
p-re
c-29
Ppu
_BIR
D_0
575
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or98
P.pu
tida
KT
2440
Pp-
rec-
30P
pu_B
IRD
_326
7To
nB-d
epen
dent
side
roph
ore
rece
ptor
97P.
putid
aK
T24
40P
p-re
c-31
Ppu
_BIR
D_0
912
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or99
P.pu
tida
KT
2440
Pp-
rec-
32P
pu_B
IRD
_030
0To
nB-d
epen
dent
side
roph
ore
rece
ptor
98P.
putid
aK
T24
40P
p-re
c-33
Ppu
_BIR
D_0
294
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or99
P.pu
tida
KT
2440
Pp-
rec-
34P
pu_B
IRD
_376
5To
nB-d
epen
dent
side
roph
ore
rece
ptor
99P.
putid
aK
T24
40P
p-re
c-35
Ppu
_BIR
D_2
177
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or99
P.pu
tida
KT
2440
Pp-
rec-
36P
pu_B
IRD
_217
8To
nB-d
epen
dent
side
roph
ore
rece
ptor
99P.
putid
aK
T24
40P
p-re
c-37
Ppu
_BIR
D_3
161
TonB
-dep
ende
ntsi
dero
phor
ere
cept
or98
P.pu
tida
F1
Pp-
rec-
38P
pu_B
IRD
_446
1To
nB-d
epen
dent
side
roph
ore
rece
ptor
95P.
putid
aF
1
The
uppe
rta
ble
show
sfe
cAge
nes
phys
ical
lylin
ked
tofe
cI/fe
cRre
gula
tory
syst
ems.
The
low
erta
ble
show
sno
n-lin
ked
Fec
A-li
ke.T
heP
P_B
IRD
_xxx
xnu
mbe
rlo
cate
sth
eco
rres
pond
ing
rece
ptor
inth
ege
nom
eof
BIR
D-1
stra
in.
The
anno
tatio
nre
fers
toth
ebe
stbl
ast
hit
with
sequ
ence
sde
posi
ted
inth
eS
WIS
SP
RO
T/T
rEM
BL
data
base
and
we
indi
cate
the
degr
eeof
iden
tity
and
the
mic
roor
gani
smso
urce
that
gave
the
best
hit.
PGPR properties 7
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

each of the pathways through the generation of mutantsby site-directed mutagenesis. For the tryptophan 2-monoxygenase single and double mutants were gener-ated using pCHESI-Km and pCHESI-Gm. A single mutantin tryptophan decarboxylase was also constructedusing pCHESI-Km, and a double mutant inactivating tryp-tophan decarboxylase and one of the tryptophan2-monooxygenases was constructed. We then measuredthe capacity of the mutants to produce IAA using thequantitative assay. The results showed that after 24 hincubation in DF minimal medium the single and doublemutants produced amounts that were about a half of thatproduced by the wild-type strain. These IAA levels werestill high enough to allow for a faster growth of maize rootsthan in the absence of bacteria (see Fig. S5). Theseresults are intriguing and thus we are further analysingIAA biosynthesis and IAA metabolism in this strain.
Inactivation of the ACC deaminase. Ethylene is a plantgrowth regulator that affects plant growth, developmentand senescence. ACC is present in plant root exudates
(Reid, 1987) and some PGPR have been proposed topromote plant growth by lowering the levels of ethylenethrough the activity of the enzyme ACC deaminase, whichhydrolyses ACC, the immediate biosynthetic precursor ofethylene in plants (Yang and Hoffman, 1984). The prod-ucts of this hydrolysis, ammonia and a-ketobutyrate, canbe used by the bacterium as a source of nitrogen andcarbon for growth (Klee et al., 1991). In this way, thebacterium acts as a sink for ACC and thus lowers ethylenelevels in plants, preventing some of the potentially delete-rious consequences of high ethylene concentrations(Glick et al., 1998; Steenhoudt and Vanderleyden, 2000;Saleem et al., 2007). We found that BIRD-1 has an ORF(PPU_BIRD-1_3642) that exhibits high homology to theACC deaminase of different microorganisms. We inacti-vated the PPU_BIRD-1_3642 allele using site-directedmutagenesis and found that its absence did not influenceplant root development under standard growth conditionsin soil in controlled chambers (Fig. S6), which indicatedthat ACC deaminase did not play a significant role in thedevelopment of maize roots.
Fig. 6. Synthesis of IAA based on the annotation of the BIRD-1 genome.
8 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Concluding remarks
Several mechanisms by which PGPR promote plantgrowth have been described in the literature. In thegenome of P. putida BIRD-1 we have identified many ofthe genes that are involved in this phenotype. Concord-antly, BIRD-1 is able to solubilize organic and inorganicphosphate, to synthesize plant hormones, to producesiderophores and to survive the oxidative stress imposedby the rhizosphere. Furthermore, the BIRD-1 genomealso encodes all of the functions required to adhere toseeds and to colonize roots – we have shown that it is agood colonizer of maize, tomato, pepper, zucchini andstrawberry roots. BIRD-1 possesses a complete setof chemotaxis genes (Z. Udaondo and J.-L. Ramos,unpubl. results) and responds to amino acids in root exu-dates as a signal (A. Roca and P. Pizarro-Tobías, unpubl.results). The combination of all of these properties makesit an excellent microbe for utilization in agriculture. BIRD-1promotes the growth of the plant by synthesizing IAA andother chemicals. Furthermore, its ability to survive dessi-cating conditions grants this strain an additional propertyof exceptional utility for its use in non-irrigated farming.We view the Pseudomonas–plant pair as a mutualisticinteraction in which the microorganism solubilizes P andFe and also produces plant growth hormones that favourplant growth, while the plants release up to 20% of fixedCO2 as root exudates that can be used by Pseudomonasto proliferate, so that at the end a beneficial interaction isestablished due to the concomitant high microbial celldensities in the plant rhizosphere.
Pseudomonas putida BIRD-1 possesses an excep-tional arsenal of mechanisms to flourish in the rhizo-sphere of plants and to promote plant growth. Morestudies are necessary to firmly establish these abilities infield tests and to determine the PGPR efficiency in a lesscontrolled environment.
Experimental procedures
Strains, plasmids and culture media
The bacterial strains cloning vectors and plasmids used inthis study have been described before (Molina-Henareset al., 2009; Matilla et al., 2010). Pseudomonas putidaBIRD-1 and its mutant derivatives were routinely grown in M9minimal medium supplemented with glucose [0.5% (w/v)] asthe carbon source (Abril et al., 1989). Cultures were incu-bated at 30°C and shaken on an orbital platform operating at200 strokes per minute. When required, antibiotics were usedat the following final concentrations (in mg ml-1): ampicillin,100; chloramphenicol, 30; kanamycin, 50; and tetracycline,20. Escherichia coli strains were grown at 37°C in LB broth(Sambrook et al., 1989) with shaking.
When indicated P. putida cells were grown on modified Amedium (Nautilaya, 1999) that lacks a phosphorous sourceand whose specific composition per litre was NH4Cl, 267 mg;
MgSO4 ¥ 7H2O, 410 mg; KCl, 300 mg; NaCl, 200 mg, 1 ml ofan aqueous solution of iron citrate (6 g l-1) and 0.5% (w/v)glucose, or 10 mM sodium benzoate as the carbon sources.As a phosphorous source we added either phosphate rock(5 g l-1) or a mixture of insoluble inorganic phosphates madeup of CaHPO4 ¥ 2H2O and Ca5(OH)(PO4)3.
To screen the ability of P. putida strains to use inorganicand organic insoluble phosphate as a phosphorous sourcePikovskaya agar plates (Naik et al., 2008) and PSM (phytatescreening medium) agar solid medium were used (Jorqueraet al., 2008). The reaction was positive when a clear halosurrounding the bacterial colonies was formed after 3–7 daysof incubation at 30°C.
Chrome azurol S agar solid medium was used to screensiderophore production (Alexander and Zuberer, 1991). Thereaction was considered positive when an orange halo sur-rounding the bacterial colony was formed after 3–7 days ofincubation at 30°C.
To screen the ability of P. putida strains to produce IAA onsolid medium, amended LB and the Salkowski’s reagentwere used (Naik et al., 2008). The composition of theamended LB per litre was 10 g of bactotryptone, 5 g of yeastextract, 10 g of NaCl, 5 mM L-tryptophan, 600 mg of sodiumdodecyl sulfate (SDS), 10 ml of glycerol and 15 g of agar. Thecomposition of the Salkowski’s reagent per litre was 2% v/v of0.5 M FeCl3 dissolved in 35% perchloric acid. Quantificationof IAA in liquid medium was performed after growth of thestrains for 24 h at 30°C with different concentrations ofL-tryptophan (0, 100, 250, 500 and 1000 mg l-1, as described)(Patten and Glick, 2002).
Quantification of soluble phosphate in culture medium
Determination of soluble phosphate in solution was per-formed as described by Murphy and Riley (1958). Superna-tants at 0, 2, 4, 8, 24, 48 and 72 h from P. putida BIRD-1cultures grown in phosphate rock (5 g l-1) were mixed with areagent whose composition per 250 ml was 125 ml of H2SO4
(5N), 37.5 ml of 4% ammonium molybdate solution and 75 mlof 0.1 M ascorbic acid. Forty millilitres of each supernatantwas mixed with 8 ml of the reagent solution and incubated for30 min in a 60°C water bath. After cooling, absorbance at620 nm was measured. The amount of soluble phosphate(mg l-1) present in the culture supernatants was determinedby comparing absorbance measures with a PO4
3- standardcurve.
Siderophore production
Strains were inoculated in M9 minimal medium supple-mented with glucose (25 mM). Iron citrate (6 mg l-1) wasadded to the control cultures (plus iron) but not to the iron-deprived ones. When the cultures reached an OD at 660 nmof around 1, they were illuminated with ultraviolet light in aLKB 2011-002 MacroVue transilluminator (LKB Bromma,Sweden) and photographed (GelDoc 2000, Bio-Rad). Torecord the UV-visible spectrum (220–700 nm) of the culturesupernatant, 1 ml of culture was withdrawn, cells removed bycentrifugation (14 000 g ¥ 5 min) and the supernatant used tomeasure the absorption spectrum.
PGPR properties 9
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Toxic compound resistance assays
Individual colonies of P. putida BIRD-1 and mutant strainswere picked from freshly cultured LB plates, streaked onto LBmedium plates, supplemented with the suitable antibiotic andgrown overnight at 30°C. The biomass of this overnight platewas recovered from the plate surface and resuspended in15 ml of LB liquid medium to an OD660 of 0.1. Microwell platewells were filled with 180 ml of the above cell suspension andwith 20 ml of the different 10 ¥ concentrated solutions ofthe stressors. Samples were incubated and data recordingswere processed using a Bioscreen C MBR analyser asdescribed before (Daniels et al., 2010). Toxic compoundstested were: K2Cr2O7 (12.5 mg ml-1), H2O2 (0.004%), methylviologen (100 mM), tert-butyl hydroperoxide (0.00078%),KCN (0.325 mg ml-1) and ampicillin (100 mg ml-1).
Surface sterilization, germination of seeds and rootcolonization assay
These assays were performed as described by Ramos-González and colleagues (2005). Corn seeds were surface-sterilized by rinsing with sterile deionized water, washingfor 10 min with 70% (v/v) ethanol, then for 15 min with10% (v/v) bleach, and followed thorough rinsing withsterile deionized water. Surface-sterilized seeds were pre-germinated on water agar [2.25% agar (w/v)] at 30°C in thedark for 48 h.
For root colonization assays seeds were inoculated withapproximately 5 ¥ 106 cfu ml-1 from a LB medium overnightculture and suspended in M9 salts medium (Sambrook et al.,1989). After incubation without shaking for 30 min at 30°C,seeds were washed in sterile distilled water and planted in50 ml Falcon tubes containing 35 ml of a mixture of sterilepeat and washed sand (3:1 v/v) and were then maintained ina controlled chamber at 24°C and 55–65% humidity with adaily light period of 16 h. At the indicated times plants werecollected, shoots discarded and the roots placed in 50 mlSterilin tubes containing 10 ml of M9 minimal salts medium(Sambrook et al., 1989) and 4 g of glass beads (0.3 mmdiameter). Tubes were vortexed for 1 min and the number ofcfu attached to the surface of the root was estimated bydrop-plating serial dilutions of the supernatant in selectivemedia.
Corn seeds adhesion assays
Strains were cultured overnight at 30°C in LB broth andculture turbidity was adjusted to OD660 = 1 in a final volume of1 ml of M9 minimal salts and the assays conducted asdescribed by Espinosa-Urgel and Ramos (2004).
Corn seeds growth on agar plates
Seeds were sterilized, as described above, and submergedfor 30 min without shaking on overnight cultures (108 cfu ml-1)of the different strains under test. Seeds were rinsed withsterile distilled water to wash out non-attached cells, and thenplaced on water agar plates (agar 2.25% w/v). Growth was
checked after 7 days of incubation in the dark at 28°C. Rootswere then weighed, for fresh weight, and stored in a stove at90°C for 48 h.
Competitive root colonization assays
Surface sterilization, germination of seeds and bacterialinoculation were performed as described above, except thatseedlings were inoculated with a mixture of a SmR BIRD-1strain, as the wild type, and the mutant strain in the specifiedgene. Inocula size differences between the wild-type andmutant strains were less than 2%. At the indicated times,bacterial cells were recovered from the rhizosphere, as speci-fied above. LB agar supplied with rifampin and streptomycin(or kanamycin) was used to select BIRD-1 or the mutantstrain respectively.
Microcosm assays
Sterile maize seedlings were either inoculated or not withapproximately 106 P. putida BIRD-1 cells per entire seedling(Espinosa-Urgel et al., 2000). The seeds were then planted insterilized potting material (Compo Sana Universal) andgrown for 10 days. Then the plantlets were transplanted intoplant pots containing 600 g of sterilized loamy silt soil classi-fied as Humic Haploxerept with the following characteristics:38% sand, 43% silt and 19% clay, pH 7.9, the organic mattercontent was 2.1% and its CaCO3 content was 8%. To deter-mine drought tolerance of wild-type and mutant strains, soilwater content was adjusted to different degrees. Viablemicroorganisms in the rhizosphere soil (soil closely associ-ated to roots) and bulk soil from four selected plants weredetermined, as described above.
Germination of vegetable seeds
Non-sterile vegetable seeds (> 30 per treatment) were sownin seed beds, previously filled with a mixture of non-sterilepeat and washed sand (3:1 v/v) for control treatments. Forthe treated seeds, a 109 cfu ml-1 culture of the assayed strainwas diluted (1:10 v/v), and mixed (1:250 v/wt) with the peat–sand mixture; vegetable seeds were then sown and wateredwith distilled water to achieve appropriate substrate moisture.Seed beds were covered to allow germination in the dark.Approximately 72 h later, when germination was accom-plished and germination rate was calculated, seed beds wereuncovered to allow stem development, and watered, with tapwater, when necessary.
Greenhouse assays
Vegetable seeds of horticultural importance in the Mediter-ranean area (corn, cucumber, zucchini, lettuce, chard,pepper, melon, onion, tomato and bean) were chosen forgreenhouse assays. Seeds were germinated as describedabove, and plants were watered with tap water when nec-essary along the assay. Monitoring of growth was performedevery month by measuring 30 plants of each treatment,stem length and gauge, when necessary. The root length
10 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

was measured after 3 months, and dry weight was calcu-lated for each treatment. Survival of inoculated cells wasscreened every 15 days, by drop-plating serial dilutions ofrhizosphere soil (soil closely associated to roots) in selectivemedia, as described before.
Measurement of phosphatase activity in soil
Phosphatase activity was determined as described byAntolín and colleagues (2005). The amount of p-nitrophenol(PNP) released from 0.5 g of soil from each treatment (bulksoil and non-inoculated corn rhizosphere soil taken as con-trols versus BIRD-1 inoculated corn rhizosphere soil) wasmeasured after incubation at 37°C for 120 min with 0.115 M4-nitrophenyl phosphate-disodium (PNPP) as substrate forthe enzymatic reaction, in 2 ml of maleate buffer (0.1 M,pH 6.5). Then, samples were cooled at 2°C for 15 min tostop enzymatic reaction, and 0.5 ml of 0.5 M CaCl2 and 2 mlof 0.5 M NaOH were added and well mixed. Each samplewas centrifuged at 2000 g for 10 min. A blank experimentwas performed for each assay, in which the substrate wasadded to the soil sample after incubation and before stop-ping the reaction. The amount of PNP per hour releasedfrom each soil sample (mg PNP g-1 h-1) was determined bycomparing absorbance measures with a PNP standardcurve.
In vitro nucleic acid techniques
Total DNA extraction was performed as described previously(Rodríguez-Herva et al., 2007). Plasmid DNA was isolatedusing the Qiagen spin miniprep kit. For DNA digestion, themanufacturer’s instructions were followed (Roche and NewEngland Biolabs). Ligation of DNA and Southern blots wereperformed following the standard protocols (Sambrook et al.,1989). Highly electro-competent cells were prepared as pre-viously reported (Choi et al., 2006) and transformed usingan EC100 electroporator according to the manufacturer’sinstructions. Expand high-fidelity Taq polymerase was used inthe amplification of PCR fragments for cloning.
Construction of mutants
For the construction of mutants, gene fragments between400 and 1000 bp were amplified with the appropriate oligo-nucleotides and cloned into pMBL1-T plasmid (DominionMBL). Plasmids containing the corresponding inserts werethen cut with BamHI (that cuts into the poly-linker of thevector, but not into the insert sequence), and the WKm resist-ance cassette of plasmid pHP45W-Km (Prentki and Kirsch,1984), previously excised with BamHI, was ligated into theBamHI restriction site. Approximately 600 ng of the resultingplasmid was electroporated into P. putida BIRD-1 (Enderleand Farwell, 1998) to allow a single recombination eventbetween the cloned DNA fragment and its homologoussequence. The resulting kanamycin (25 mg ml-1)-resistantclones were selected and analysed by Southern blot. Clonesthat contained the insertion in the appropriate location werekept for further analysis.
Nucleotide sequence accession number
The complete genome sequence of P. putida BIRD-1 is avail-able in GenBank under Accession Number CP002290.
Acknowledgements
Work was supported by Fondo Social Europeo and FEDERFunds through Programa Campus of Junta de Andalucía(Project SV40), awarded by Agencia IDEA to Bio-IliberisR&D, and by Project BIO2010-17227 from the Ministry ofScience and Innovation awarded to CSIC.
References
Abril, M.A., Michán, C., Timmis, K.N., and Ramos, J.L. (1989)Regulator and enzyme specificities of the TOL plasmid-encoded upper pathway for degradation of aromatic hydro-carbons and expansion of the substrate range of thepathway. J Bacteriol 171: 6782–6790.
Ahemad, M., and Khan, S. (2012) Evaluation of plant-growth-promoting activities of rhizobacterium Pseudomonasputida under herbicide stress. Ann Microbiol 62: 1531–1540. DOI: 10.1007/s13213-011-0407-2.
Alexander, D.B., and Zuberer, D.A. (1991) Use of chromeazurol S reagents to evaluate siderophore production byrhizosphere bacteria. Biol Fertil Soils 12: 39–45.
Antolín, M.C., Pascual, I., García, C., Polo, A., andSánchez-Díaz, M. (2005) Growth, yield and solutecontent of barley in soils treated with sewage sludgeunder semiarid Mediterranean conditions. Field CropsRes 94: 224–237.
Bakker, P.A., Pieterse, C.M., and van Loon, L.C. (2007)Induced systemic resistance by fluorescent Pseudomonasspp. Phytopathology 97: 239–243.
Banik, S., and Dey, B.K. (1982) Available phosphate contentof an alluvial soil as influenced by inoculation of someisolated phosphate solubilizing microorganisms. Plant Soil69: 353–364.
Berg, G. (2009) Plant-microbe interactions promoting plantgrowth and health: perspectives for controlled use of micro-organisms in agriculture. Appl Microbiol Biotechnol 84:11–18.
Bielecki, P., Puchalka, J., Wos-Oxley, M.L., Loessner, H.,Glik, J., Kawecki, M., et al. (2011) In vivo expression pro-filing of Pseudomonas aeruginosa infections reveals niche-specific and strain-independent transcriptional programs.PLoS ONE 6: e24235.
Blom, D., Fabbri, C., Connor, E.C., Schiestl, F.P., Klauser,D.R., Boller, T., et al. (2011) Production of plant growthmodulating volatiles is widespread among rhizospherebacteria and strongly depends on culture conditions.Environ Microbiol 13: 3047–3058.
Cattelan, A.J., Hartel, P.G., and Fuhrmann, J.J. (1999)Screening for plant growth rhizobacteria to promote earlysoybean growth. Soil Sci Soc Am J 63: 1670–1680.
Choi, K.H., Kumar, A., and Schweizer, H.P. (2006) A 10-minmethod for preparation of highly electrocompetent Pseu-domonas aeruginosa cells: application for DNA fragmenttransfer between chromosomes and plasmid transforma-tion. J Microbiol Methods 64: 391–397.
PGPR properties 11
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Cleland, R.E. (1990) Auxin and cell elongation. In Plant Hor-mones and Their Role in Plant Growth and Development.Davies, P.J. (ed.). Dordrecht, the Netherlands: Kluwer, pp.132–148.
Compant, S., Clément, C., and Sessitsch, A. (2010) Plantgrowth-promoting bacteria in the rhizo- and endosphere ofplants: their role, colonization, mechanisms involved andprospects for utilization. Soil Biol Biochem 42: 669–678.
Dangar, T.K., and Basu, P.S. (1987) Studies on plant growthsubstances, IAA metabolism and nitrogenase activity inroot nodules of Phaseolus aureus Roxb. var. mungo. BiolPlant 29: 350–354.
Daniels, C., Godoy, P., Duque, E., Molina-Henares, M.A.,Torre, J., del Arco, J.M., et al. (2010) Global regulation offood supply by Pseudomonas putida DOT-T1E. J Bacteriol192: 2169–2181.
De Freitas, J.R., Banerjee, M.R., and Germida, J.J. (1997)Phosphate solubilizing rhizobacteria enhance the growthand yield but not phosphorus uptake of canola (Brassicanapus L.). Biol Fertil Soils 24: 358–364.
Dey, R., Pal, K.K., Bhatt, D.M., and Chauhan, S.M. (2004)Growth promotion and yield enhancement of peanut(Arachis hypogaea L) by application of plant growth pro-moting rhizobacteria. Microbiol Res 159: 371–394.
Dobbelaere, S., Vanderleyden, J., and Okon, Y. (2003) Plantgrowth promoting effects of diazotrophs in the rhizosphere.Crit Rev Plant Sci 22: 107–149.
Duff, R.B., and Webley, D.M. (1959) 2-Ketogluconic acid as anatural chelator produced by soil bacteria. Chem Ind 1939:1376–1377.
Duque, E., Torre, J., Bernal, P., Molina-Henares, M.A., Ala-minos, M., Espinosa-Urgel, M., et al. (2012) Identificationof reciprocal adhesion genes in pathogenic and nonpatho-genic Pseudomonas. Environ Microbiol [Epub ahead ofprint]. DOI: 10.1111/j.1462-2920.2012.02732.x.
Enderle, P.J., and Farwell, M.A. (1998) Electroporation offreshly plated Escherichia coli and Pseudomonas aerugi-nosa cells. Biotechniques 25: 954–958.
Espinosa-Urgel, M., and Ramos, J.L. (2004) Cell density-dependent gene contributes to efficient seed colonizationby Pseudomonas putida KT2440. Appl Environ Microbiol70: 5190–5198.
Espinosa-Urgel, M., Salido, A., and Ramos, J.L. (2000)Genetic analysis of functions involved in adhesion of Pseu-domonas putida to seeds. J Bacteriol 182: 2363–2369.
Fernández-Piñar, R., Cámara, M., Dubern, J.F., Ramos, J.L.,and Espinosa-Urgel, M. (2011) The Pseudomonas aerugi-nosa quinolone quorum sensing signal a the multicellularbehaviour of Pseudomonas putida KT2440. Res Microbiol162: 773–781.
Gamalero, E., and Glick, B.R. (2011) Mechanisms used byplant growth-promoting bacteria. In Bacteria in Agrobiol-ogy: Plant Nutriente Management. Maheshwari, D.K. (ed.).Berlin, Germany: Springer, pp. 17–46.
Glick, B.R. (1995) The enhancement of plan growth by free-living bacteria. Can J Microbiol 41: 109–117.
Glick, B.R., and Pasternak, J.J. (2003) Plant growth promot-ing bacteria. In Molecular Biotechnology Principles andApplications of Recombinant DNA, 3rd edn. Glick, B.R.,and Pasternak, J.J. (eds). Washington, DC, USA: ASMPress, pp. 436–454.
Glick, B.R., Penrose, D.M., and Li, J. (1998) A model forlowering plant ethylene concentration by plant growth pro-moting rhizobacteria. J Theor Biol 190: 63–68.
Gügi, B., Orange, N., Hellio, F., Burini, J.F., Guillou, C.,Leriche, F., and Guespin-Michel, J.F. (1991) Effect ofgrowth temperature on several exported enzyme activitiesin the psychrotropic bacterium Pseudomonas fluorescens.J Bacteriol 173: 3814–3820.
Haas, P., and Keel, C. (2003) Regulation of antibiotic produc-tion in root colonizing Pseudomonas spp. and relevance forbiological control of plant disease. Annu Rev Phytopathol41: 117–153.
Halder, A.K., and Chakrabarty, P.K. (1993) Solubilization ofinorganic phosphate by Rhizobium. Folia Microbiol 38:325–330.
Halder, A.K., Mishra, A.K., Bhattacharya, P., and Chakrabar-thy, P.K. (1990) Solubilization of rock phosphate by Rhizo-bium and Bradyrhizobium. J Gen Appl Microbiol 36:1–92.
Hammer, P.E., Hill, D.S., Lam, S.T., van Pée, K.H., and Ligon,J.M. (1997) Four genes from Pseudomonas fluorescensthat encode the biosynthesis of pyrrolnitrin. Appl EnvironMicrobiol 63: 2147–2154.
Jiménez, J.I., Miñambres, B., García, J.L., and Díaz, E.(2002) Genomic analysis of the aromatic catabolic path-ways from Pseudomonas putida KT2440. Environ Micro-biol 4: 824–841.
Jones, D.L., and Darrah, P.R. (1994) Role of root derivedorganic acids in the mobilization of nutrients from the rhizo-sphere. Plant Soil 166: 247–257.
Jorquera, M.A., Hernández, M.T., Renger, Z., Marschner, P.,and Mora, M. (2008) Isolation of culturable phosphobacte-ria with both phytate-mineralization and phosphate-solubilization activity from the rhizosphere of plants grownin a volcanic soil. Biol Fertil Soils 44: 1025–1034.
Klee, H.J., Hayford, M.B., Kretzmer, K.A., Barry, G.F., andKishore, G.M. (1991) Control of ethylene synthesis byexpression of a bacterial enzyme in transgenic tomatoplants. Plant Cell 3: 1187–1193.
Kloepper, J.W., Schroth, M.N., and Miller, T.D. (1980) Effectsof rhizosphere colonization by plant growth-promotingrhizobacteria on potato plant development and yield. Phy-topathology 70: 1078–1082.
Kroll, J., Klinter, S., Schneider, C., Vob, I., and Steinbüchel,A. (2010) Plasmid addiction systems: perspectives andapplications in biotechnology. Microb Biotechnol 3: 634–657.
Lemanceau, P., Bauer, P., Kraemer, S., and Briat, J.F. (2009)Iron dynamics in the rhizosphere as a case study for ana-lysing interactions between soils, plants and microbes.Plant Soil 321: 513–535.
Li, J., Ovakin, D.H., Charles, T.C., and Glick, B.R. (2000) AnACC deaminase minus mutant of Entreobacter cloacaeUW4 no longer promotes root elongation. Curr Microbiol41: 101–105.
van Loon, L.C. (2007) Plant responses to plant growth-promoting rhizobacteria. Eur J Plant Pathol 119: 243–254.
van Loon, L.C., and Bakker, P.A.H. (2005) Induced systemicresistance a mechanism of disease suppression by rhizo-bacteria. In PGPR: Biocontrol and Biofertilization. Siddiqui,
12 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Z.A. (ed.). Dordrecht, the Netherlands: Springer Science,pp. 39–66.
Lugtenberg, B., and Kamilova, F. (2009) Plant-growth-promoting rhizobacteria. Annu Rev Microbiol 63: 541–556.
McGrath, J.W., Hammerschmidt, F., and Quinn, J.P. (1998)Biodegradation of phosphonomycin by Rhizobium huakuiiPMY1. Appl Environ Microbiol 64: 356–358.
Malfanova, N., Kamilova, F., Validov, S., Shcherbakov, A.,Chebotar, V., Tikhonovich, I., and Lugtenberg, B. (2011)Characterization of Bacillus subtilis HC8, a novel plant-beneficial endophytic strain from giant hogweed. MicrobBiotechnol 4: 523–532.
Martínez-Bueno, M., Tobes, R., Rey, M., and Ramos, J.L.(2002) Detection of multiple extracitoplasmatic function(ECF) sigma factors in the genome of Pseudomonasputida KT2440 and their counterparts in Pseudomonasaeruginosa PAO1. Environ Microbiol 4: 842–855.
Matilla, M.A., Espinosa-Urgel, M., Rodriguez-Herva, J.J.,Ramos, J.L., and Ramos-Gonzalez, M.I. (2007) Genomicanalysis reveals the major driving forces of bacterial life inthe rhizosphere. Genome Biol 8: R179.
Matilla, M.A., Ramos, J.L., Bakker, P.A.H., Doornbos, R.,Badri, D.V., Vivanco, J.M., and Ramos-González, M.I.(2010) Pseudomonas putida KT2440 causes induced sys-temic resistance and changes in Arabidopsis root exuda-tion. Environ Microbiol Rep 2: 381–388.
Matilla, M.A., Pizarro, P., Roca, A., Fernández, M., Duque, E.,Molina, L., et al. (2011) Complete genome of the plant-growth promoting rhizobacterium Pseudomonas putidaBIRD-1. J Bacteriol 193: 1290.
Meschke, H., and Schrempf, H. (2010) Streptomyces lividansinhibits the proliferation of Arabidopsis thaliana the fungusVerticillium dahliae on seeds and roots. Microb Biotechnol3: 428–443.
Miller, S.H., Browne, P., Prigent-Combaret, C., Combes-Meynet, E., Morrissey, J.P., and O’Gara, F. (2010) Bio-chemical and genomic comparison of inorganic phosphatesolubilization in Pseudomonas species. Environ MicrobiolRep 2: 403–411.
Molina, L., Ramos, C., Duque, E., Ronchel, M.C., García,J.M., Wyke, L., and Ramos, J.L. (2000) Survival of Pseu-domonas putida KT2440 in soil and in the rhizosphere ofplants under greenhouse and environmental conditions.Soil Biol Biochem 32: 315–321.
Molina-Henares, M.A., Ramos, J.L., and Espinosa-Urgel, M.(2006) A two-partner secretion system is involved in seedand root colonization and iron uptake by Pseudomonasputida KT2440. Environ Microbiol 8: 639–747.
Molina-Henares, M.A., García-Salamanca, A., Molina-Henares, A.J., Torre, J., Herrera, M.C., Ramos, J.L., andDuque, E. (2009) Functional analysis of aromatic biosyn-thetic pathways in Pseudomonas putida KT2440. MicrobBiotechnol 2: 91–100.
Muñoz-Rojas, J., Bernal, P., Duque, E., Godoy, P., Segura,A., and Ramos, J.L. (2006) Involvement of cyclopropanefatty acids in the response of Pseudomonas putida KT2440to freeze-drying. Appl Environ Microbiol 72: 472–477.
Murphy, J., and Riley, J.P. (1958) A single-solution method forthe determination of soluble phosphate in sea water. J MarBiolog Assoc UK 37: 9–14.
Naik, P.R., Sahoo, N., Goswani, D., Ayyadurai, N., and Sak-thivel, N. (2008) Genetic and functional diversity amongfluorescent Pseudomonads isolated from the rhizosphereof Banana. Microb Ecol 56: 492–504.
Nakkeeran, S., Dilantha, W.G., and Siddiqui, Z.A. (2005)Plant growth-promoting rhizobacteria formulations and itsscope in commercialization for the management of pestsand diseases. In PGPR: Biocontrol and Biofertizilization.Siddiqui, Z. (ed.). Dordrecht, the Netherlands: Springer, pp.257–296.
Nautilaya, C.S. (1999) An efficient microbiological growthmedium for screening phosphate solubilizing microorgan-isms. FEMS Microbiol Lett 170: 265–270.
Nelson, K.E., Weinel, C., Paulsen, I.T., Dodson, R.J., Hilbert,H., Martins dos Santos, V.A., et al. (2002) Completegenome sequence and comparative analysis of the meta-bolically versatile Pseudomonas putida KT2440. EnvironMicrobiol 4: 799–808.
Ohtake, H., Wu, H., Imazu, K., Anbe, L., Kato, J., and Kuroda,A. (1996) Bacterial phosphonate degradation, phosphiteoxidation and polyphosphate accumulation. ResourConserv Recy 18: 125–134.
Pal, K.K., Tilak, K.V.B., Saxena, A.K., Dey, R., and Singh,C.S. (2001) Suppression of maize root diseases caused byMacrophomina phaseolina, Fusarium moniliforme andFusarium germinearum by plant growth promoting rhizo-bacteria. Microbiol Res 156: 209–223.
Patten, C.L., and Glick, B.R. (1996) Bacterial biosynthesis ofindole-3-acetic acid. Can J Microbiol 42: 207–220.
Patten, C.L., and Glick, B.R. (2002) Role of Pseudomonasputida indole-acetic acid in development of the hostplant root system. Appl Environ Microbiol 68: 3795–3801.
Penrose, D.M., and Glick, B.R. (2001) Levels of ACC andrelated compounds in exudate and extracts of canolaseeds treated with ACC deaminase-containing plantgrowth-promoting bacteria. Can J Microbiol 47: 368–372.
Pérez, J., Muñoz-Dorado, J., Braña, A.F., Shimkets, L.J.,Sevillano, L., and Santamaría, R.I. (2011) Myxococcusxanthus induces actinorhodin overproduction and aerialmycelium formation by Streptomyces coelicolor. MicrobBiotechnol 4: 175–183.
Prentki, P., and Krisch, H.M. (1984) In vitro insertional muta-genesis with a selectable DNA fragment. Gene 29: 303–313.
Ramos-González, M.I., Campos, M.J., and Ramos, J.L.(2005) Analysis of Pseudomonas putida KT2440 geneexpression in the maize rhizosphere: in vivo expressiontechnology capture and identification of root-activated pro-moters. J Bacteriol 187: 4033–4041.
Reid, M. (1987) Ethylene in plant growth, development andsenescence. In Plant Hormones and Their Role in PlantGrowth and Development. Davies, P.J. (ed.). Boston, MA,USA: Martinus Nijhoff, pp. 257–279.
Richardson, A.E. (2001) Prospect for using soil microorgan-isms to improve to improve the acquisition of phosphorusby plants. Aust J Plant Physiol 28: 897–906.
Richardson, A.E., and Hadobas, P.A. (1997) Soil isolates ofPseudomonas spp. that utilize inositol phosphates. Can JMicrobiol 43: 509–516.
PGPR properties 13
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Rodríguez, H., and Fraga, R. (1999) Phosphate solubilizingbacteria and their role in plant growth promotion. Biotech-nol Adv 17: 319–339.
Rodríguez-Herva, J.J., García, V., Hurtado, A., Segura, A.,and Ramos, J.L. (2007) The ttgGHI solvent efflux pumpoperon of Pseudomonas putida DOT-T1E is located ona large self-transmissible plasmid. Environ Microbiol 9:1550–1561.
Saleem, M., Arshad, M., Hussain, S., and Bhatti, A.S. (2007)Perspective of plant growth promoting rhizobacteria(PGPR) containing ACC deaminase in stress agriculture. JInd Microbiol Biotechnol 34: 635–648.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) MolecularCloning: A Laboratory Manual, 2nd edn. New York, NY,USA: Cold Spring Harbor Laboratory Press.
Sandermann, H. (2004) Molecular ecotoxicology of plants.Trends Plant Sci 9: 406–414.
Schalk, I.J., Hannauer, M., and Braud, A. (2011) New roles ofbacterial siderophores in metal transport and tolerance.Environ Microbiol 13: 2844–2854.
Segura, A., Rodríguez-Conde, S., Ramos, C., and Ramos,J.L. (2009) Bacterial responses and interactions with plantsduring rhizoremediation. Microb Biotechnol 4: 452–464.
Shekar, C., and Nautiyal, C.S. (1999) An efficient micro-biological growth medium for screening phosphoroussolubilizing microorganisms. FEMS Microbiol Lett 170:265–270.
Simmons, M., Permentier, H.P., de Weger, L.A., Wijffelman,C.A., and Lugtenberg, B. (1997) Amino acid synthesis isnecessary for tomato root colonization by Pseudomonasfluorescens strain WCS365. Mol Plant Microbe Interact 10:102–106.
Skrary, F.A., and Cameron, D.C. (1998) Purification and char-acterization of a Bacillus licheniformis phosphatase spe-cific for D-alphaglycerophosphate. Arch Biochem Biophys349: 27–35.
Somers, E., and Vanderleyden, J. (2004) Rhizosphere bac-terial signaling: a love parade beneath our feet. Crit RevMicrobiol 30: 205–240.
Spaepen, S., Vanderleyden, J., and Remans, R. (2007)Indole-3-acetic acid in microbial and microorganism-plantsignaling. FEMS Microbiol Rev 31: 425–448.
Stajner, D., Gasaic, O., Matkovic, B., and Vargas, S.Z.I.(1995) Metolachlor effect on antioxidants enzyme activitiesand pigments content in seeds and young leaves of wheat(Triticum aestivum L.). Agr Med 125: 267–273.
Stajner, D., Kevreaan, S., Gasaic, O., Mimica-Dudic, N., andZongli, H. (1997) Nitrogen and Azotobacter chroococcumenhance oxidative stress tolerance in sugar beet. Biol Plant39: 441–445.
Steenhoudt, O., and Vanderleyden, J. (2000) Azospirillum, afree-living nitrogen-fixing bacterium closely associated withgrasses: genetic, biochemical and ecological aspects.FEMS Microbiol Rev 24: 487–506.
Taghavi, S., Garafola, C., Monchy, S., Newman, L., Hoffman,A., Weyens, N., et al. (2009) Genome survey and charac-terization of endophytic bacteria exhibiting a beneficialeffect on growth and development of poplar trees. ApplEnviron Microbiol 75: 748–757.
Uroz, S., Buée, M., Murat, C., Frey-Klett, P., and Martin, F.(2010) Pyrosequencing reveals a contrasted bacterial
diversity between oak rhizosphere and surrounding soil.Environ Microbiol Rep 2: 281–288.
Vial, L., Chapalain, A., Groleau, M.C., and Déziel, E. (2011)The various lifestyles of the Burkholderia cepacia complexspecies: a tribute to adaptation. Environ Microbiol 13:1–12.
Viruel, E., Lucca, M.E., and Sineriz, F. (2011) Plant growthpromotion traits of phosphobacteria isolated from puna,Argentina. Arch Microbiol 193: 489–496.
Vyas, P., and Gulati, A. (2009) Organic acid production invitro and plant growth promotion in maize under controlledenvironment by phosphate-solubilizing fluorescent Pseu-domonas. BMC Microbiol 9: 174.
Weinel, C., Nelson, K.E., and Tümmler, B. (2002) Globalfeatures of the Pseudomonas putida KT2440 genomesequence. Environ Microbiol 4: 809–818.
Wu, X., Monchy, S., Taghavi, S., Zhu, W., Ramos, J.L., andvan der Lelie, D. (2010) Comparative genomics and func-tional analysis of niche specific adaptation in Pseu-domonas putida. FEMS Microbiol Rev 35: 299–323.
Yang, S.F., and Hoffman, N.E. (1984) Ethylene biosynthesisand its regulation in higher plants. Annu Rev Plant Physiol35: 155–189.
Yousef-Coronado, F., Travieso, M.L., and Espinosa-Urgel, M.(2008) Different, overlapping mechanisms for colonizationof abiotic and plant surfaces by Pseudomonas putida.FEMS Microbiol Lett 288: 118–124.
Zhang, P., and Yuen, G. (2000) The role of chitinase produc-tion by Stenotrophomonas maltophilia strain C3 in biologi-cal control of bipolaris sorokiniasa. Phytopathology 41:117–153.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article:
Fig. S1. Growth of Pseudomonas putida BIRD-1 with differ-ent sources of inorganic phosphorous. We used A mediumwith the indicated C source and insoluble tricalcium phos-phate (500 mg l-1) as a source of phosphate. Viable cells atthe beginning of the assay were 2 to 3 ¥ 107 cfu ml-1. After24 h incubation at 30°C the cfu ml-1 was determined afterspreading serial dilutions on LB medium. The control is M9medium with 50 mM soluble phosphate.Fig. S2. Pikovskaya test with BIRD-1 and mutants inthe Entner–Doudoroff pathway. Plates were prepared asdescribed in Experimental procedures. We inoculated cellsusing a toothpick in a single point on the agar surface. Plateswere incubated for 48 h at 30°C and then the solubilizationhalo was visualized. (A) Wild-type; (B) zwf mutant and (C)eda mutant.Fig. S3. Growth of P. putida with phytate. Assays were per-formed with A medium containing 1 g l-1 phytate. At the indi-cated times cfu ml-1 were determined. As a control, cellsgrowing on M9 minimal medium were used. �, growth in M9medium; �, growth with phytate.Fig. S4. Production of IAA by wild-type BIRD-1 and differentisogenic mutants. IAA concentration was determined asdescribed in Experimental procedures. The solid back bar
14 A. Roca et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

represents the wild-type strains and the other bars theindicated mutants.Fig. S5. Corn root development with mutants in the IAA bio-synthetic pathways. The picture shows controls without bac-teria (A), with wild-type BIRD-1 (B) and KT2440 (C) strains,and three isogenic mutants of BIRD-1 with knockouts inPPU_BIRD-1_0418, 1202 and 3125 genes (D, E and Frespectively).
Fig. S6. Corn root development in a control seed withoutbacteria (A), and inoculated seeds with BIRD-1 (B) and amutant deficient in the acc deaminase gene (C). Conditionswere as described for Fig. 2 except that a knockout(PPU_BIRD-1_3642) acc mutant was used.Table S1. Genes involved in adhesion to biotic and abioticsurfaces in Pseudomonas putida strain BIRD-1 deduced fromthe identification of reciprocal genes in P. putida KT2440.
PGPR properties 15
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
Related Documents











