An estrogen receptor a suppressor, microRNA-22, is downregulated in estrogen receptor a-positive human breast cancer cell lines and clinical samples Jianhua Xiong 1, *, Dianke Yu 2, *, Na Wei 1 , Hanjiang Fu 3 , Tianjing Cai 1 , Yuanyu Huang 1 , Chen Wu 2 , Xiaofei Zheng 3 , Quan Du 1 , Dongxin Lin 2 and Zicai Liang 1 1 Laboratory of Nucleic Acid Technology, Institute of Molecular Medicine, Peking University, Beijing, China 2 Department of Etiology and Carcinogenesis, State Key Laboratory of Molecular Oncology, Cancer Institute, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, China 3 Beijing Institute of Radiation Medicine, China Introduction MicroRNAs (miRNAs), a class of endogenous short ( 22 nucleotides) noncoding RNAs, have been reported to be capable of suppressing the expression of protein-coding genes at the post-transcriptional level by cleaving target mRNAs and ⁄ or repressing their translation [1]. Aberrant expression of miRNAs is known to be involved in various human diseases, including cancer [2,3]. In two recent studies, more than 50% of human miRNA genes have been mapped to the cancer-related chromosomal regions with high Keywords breast carcinoma; estrogen receptor a; microRNA-22; proliferation Correspondence Z. Liang, Laboratory of Nucleic Acid Technology, Institute of Molecular Medicine, Peking University, Beijing 100871, China Fax: +86 10 62769862 Tel: +86 10 62769862 E-mail: [email protected] Dongxin Lin, Department of Etiology and Carcinogenesis, Cancer Institute, Chinese Academy of Medical Sciences, Beijing 100021, China Fax: +86 10 67722460 Tel: +86 10 87788491 E-mail: [email protected] *These authors contributed equally to this work (Received 30 September 2009, revised 5 January 2010, accepted 25 January 2010) doi:10.1111/j.1742-4658.2010.07594.x Previous studies have suggested that microRNAs (miRNAs) may play important roles in tumorigenesis, but little is known about the functions of most miRNAs in cancer development. In the present study, we set up a cell-based screen using a luciferase reporter plasmid carrying the whole 4.7 kb 3¢-UTR of estrogen receptor a (ERa) mRNA cotransfected with a synthetic miRNA expression library to identify potential ERa-targeting miRNAs. Among all the miRNAs, miR-22 was found to repress robustly the luciferase signal in both HEK-293T and ERa-positive MCF-7 cells. Mutation of the target site was found to abrogate this repression effect of miR-22, whereas antagonism of endogenous miR-22 in MDA-MB-231 cells resulted in elevated reporter signals. We assessed the miR-22 expression patterns in five breast cancer cell lines and 23 clinical biopsies and revealed that there is a significant inverse association between the miR-22 levels and ERa protein expression. To evaluate the potential of miR-22 as a potential therapeutic intervention, we found that reduction of endogenous ERa pro- tein levels and suppression of cancer cell growth could be achieved in MCF-7 cells by miR-22 overexpression in a way that can be recapitulated by the introduction of specific small interfering RNA against ERa. The phenomena can be rescued by the reintroduction of ERa. Taken together, our data indicate that miR-22 was frequently downregulated in ERa-posi- tive human breast cancer cell lines and clinical samples. Direct involvement in the regulation of ERa may be one of the mechanisms through which miR-22 could play a pivotal role in the pathogenesis of breast cancer. Abbreviations DMEM, Dulbecco’s modified Eagle’s medium; ERa, estrogen receptor a; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; miRNA, microRNA; siRNA, small interfering RNA. 1684 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

An estrogen receptor a suppressor, microRNA-22, isdownregulated in estrogen receptor a-positive humanbreast cancer cell lines and clinical samplesJianhua Xiong1,*, Dianke Yu2,*, Na Wei1, Hanjiang Fu3, Tianjing Cai1, Yuanyu Huang1, Chen Wu2,Xiaofei Zheng3, Quan Du1, Dongxin Lin2 and Zicai Liang1
1 Laboratory of Nucleic Acid Technology, Institute of Molecular Medicine, Peking University, Beijing, China
2 Department of Etiology and Carcinogenesis, State Key Laboratory of Molecular Oncology, Cancer Institute, Chinese Academy of Medical
Sciences and Peking Union Medical College, Beijing, China
3 Beijing Institute of Radiation Medicine, China
Introduction
MicroRNAs (miRNAs), a class of endogenous short
(� 22 nucleotides) noncoding RNAs, have been
reported to be capable of suppressing the expression of
protein-coding genes at the post-transcriptional level
by cleaving target mRNAs and ⁄or repressing their
translation [1]. Aberrant expression of miRNAs is
known to be involved in various human diseases,
including cancer [2,3]. In two recent studies, more than
50% of human miRNA genes have been mapped to
the cancer-related chromosomal regions with high
Keywords
breast carcinoma; estrogen receptor a;
microRNA-22; proliferation
Correspondence
Z. Liang, Laboratory of Nucleic Acid
Technology, Institute of Molecular Medicine,
Peking University, Beijing 100871, China
Fax: +86 10 62769862
Tel: +86 10 62769862
E-mail: [email protected]
Dongxin Lin, Department of Etiology and
Carcinogenesis, Cancer Institute, Chinese
Academy of Medical Sciences, Beijing
100021, China
Fax: +86 10 67722460
Tel: +86 10 87788491
E-mail: [email protected]
*These authors contributed equally to this
work
(Received 30 September 2009, revised 5
January 2010, accepted 25 January 2010)
doi:10.1111/j.1742-4658.2010.07594.x
Previous studies have suggested that microRNAs (miRNAs) may play
important roles in tumorigenesis, but little is known about the functions of
most miRNAs in cancer development. In the present study, we set up a
cell-based screen using a luciferase reporter plasmid carrying the whole
� 4.7 kb 3¢-UTR of estrogen receptor a (ERa) mRNA cotransfected with
a synthetic miRNA expression library to identify potential ERa-targetingmiRNAs. Among all the miRNAs, miR-22 was found to repress robustly
the luciferase signal in both HEK-293T and ERa-positive MCF-7 cells.
Mutation of the target site was found to abrogate this repression effect of
miR-22, whereas antagonism of endogenous miR-22 in MDA-MB-231 cells
resulted in elevated reporter signals. We assessed the miR-22 expression
patterns in five breast cancer cell lines and 23 clinical biopsies and revealed
that there is a significant inverse association between the miR-22 levels and
ERa protein expression. To evaluate the potential of miR-22 as a potential
therapeutic intervention, we found that reduction of endogenous ERa pro-
tein levels and suppression of cancer cell growth could be achieved in
MCF-7 cells by miR-22 overexpression in a way that can be recapitulated
by the introduction of specific small interfering RNA against ERa. The
phenomena can be rescued by the reintroduction of ERa. Taken together,
our data indicate that miR-22 was frequently downregulated in ERa-posi-tive human breast cancer cell lines and clinical samples. Direct involvement
in the regulation of ERa may be one of the mechanisms through which
miR-22 could play a pivotal role in the pathogenesis of breast cancer.
Abbreviations
DMEM, Dulbecco’s modified Eagle’s medium; ERa, estrogen receptor a; GAPDH, glyceraldehyde-3-phosphate dehydrogenase;
miRNA, microRNA; siRNA, small interfering RNA.
1684 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS

frequencies of amplification or deletion, and frequent
genomic alterations of miRNAs were observed in can-
cers [4,5]. Emerging evidence shows that miRNAs
function as oncogenes or tumor suppressors to modu-
late multiple oncogenic cellular processes, including
cell proliferation, apoptosis, invasion and migration
[6–8]. For example, it has been shown that p53-depen-
dent miR-34b and miR-34c cooperate to inhibit the
proliferation of neoplastic epithelial ovarian cells [6],
and miR-15 and miR-16 simulate apoptosis in chronic
lymphocytic leukemia by targeting BCL2 [7]. On the
other hand, miR-10b initiates breast cancer invasion
and metastasis by repressing homeobox D10 [8]. More-
over, miRNA expression profiling has been used as a
signature to distinguish different cancer types and to
provide an accurate classification of poorly differenti-
ated tumors [9]. In view of the roles that miRNAs play
in human diseases, including cancer, miRNAs have
been considered to be potential drug candidates or
therapeutic targets [10].
Breast cancer is one of the most common and preva-
lent cancers in women and a leading cause of cancer-
related death [11]. As in other common cancers, the
formation and progression of breast cancer is a multi-
step process involving genetic and epigenetic altera-
tions that drive unrestrained cell proliferation and
growth [12,13]. Several aberrantly expressed miRNAs
have been identified in breast cancer, such as miR-21,
miR-205 and miR-17-5p, which were shown to be
involved in the formation of breast cancer by targeting
the TPM1, HER3 and AIB1 genes, respectively [14–
16]. However, the functional roles of most miRNAs in
the development of breast cancer remain unknown.
In this study, we identified miR-22 as a potent regu-
lator of ESR1 encoding estrogen receptor a (ERa) anddemonstrated that miR-22 is frequently downregulated
in ERa-positive human breast cancer cell lines and
clinical samples. In addition, further functional
studies showed that ERa plays an important role in
miR-22-mediated growth retardation of tumor cells.
Results
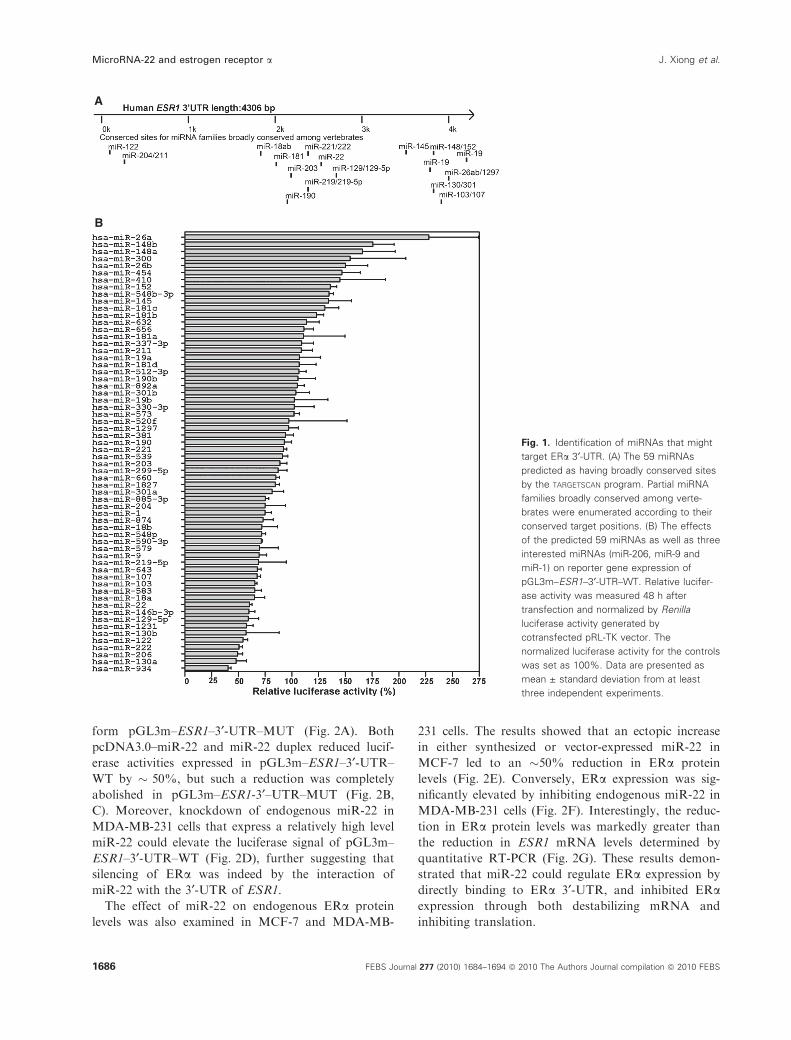
Identification of miRNAs that might target ERa3¢-UTR
To identify human miRNAs that might target ERa3¢-UTR, we used the targetscan program (http://
www.targetscan.org/) to predict miRNAs that have the
interaction with 3¢-UTR of ESR1 mRNA. Along the
� 4.3 kb full length of 3¢-UTR of ESR1 mRNA, 59
miRNAs had conserved target sites and partial
miRNA families broadly conserved among vertebrates
were enumerated according to their conserved target
positions (Fig. 1A). In addition, miR-206, which has
two target sites on 3¢-UTR of ESR1 mRNA, was pre-
viously reported as a negative regulator of ERa [17].
miR-9 and miR-1 were implicated in crucial cancer-
related cell signaling regulation [18,19]. The 62
miRNAs were chosen as our preferred candidates
for ERa regulators. To evaluate comprehensively
miRNAs–ESR1 mRNA interactions, we used the
screening system based on a luciferase reporter plasmid
carrying the full-length 3¢-UTR of ESR1 mRNA. As a
result, nine miRNAs were found to suppress the
expression of the reporter by more than 40%, and 25
miRNAs were found to suppress the expression of the
reporter by more than 20% (Fig. 1B). This might
constitute the major category of miRNAs that play
regulatory roles on ERa through interactions with
3¢-UTR of ESR1 mRNA. As indicated in Fig. 1B,
miR-22 could induce an �40% reduction in the
luciferase signal.
Direct regulation of ERa expression by miR-22
We compared three popular miRNA target prediction
programs and found that miR-22 was highly scored in
all three algorithms [targetscan, miranda (http://
www.microrna.org/microrna/home.do) and pictar
(http://pictar.mdc-berlin.de/)] for targeting ESR1
(Table S1) [20,21]. The targetscan prediction sug-
gested that ESR1 has an extremely conserved miR-22
target site (position 2292–2298 of human ESR1
3¢-UTR) in human and other mammalian species,
including the chimpanzee, rhesus monkey, mouse, rat,
dog and rabbit (Fig. 2A). The predicted DG of 70 bp
5¢- and 3¢-flanking regions of neighboring potential
conserved miR-22 target site was determined by mfold
and the resulting DG values ()10.70 and )8.40kcalÆmol)1, respectively) suggested that miR-22 may
have access to its conserved target ESR1 mRNA site
[22,23].
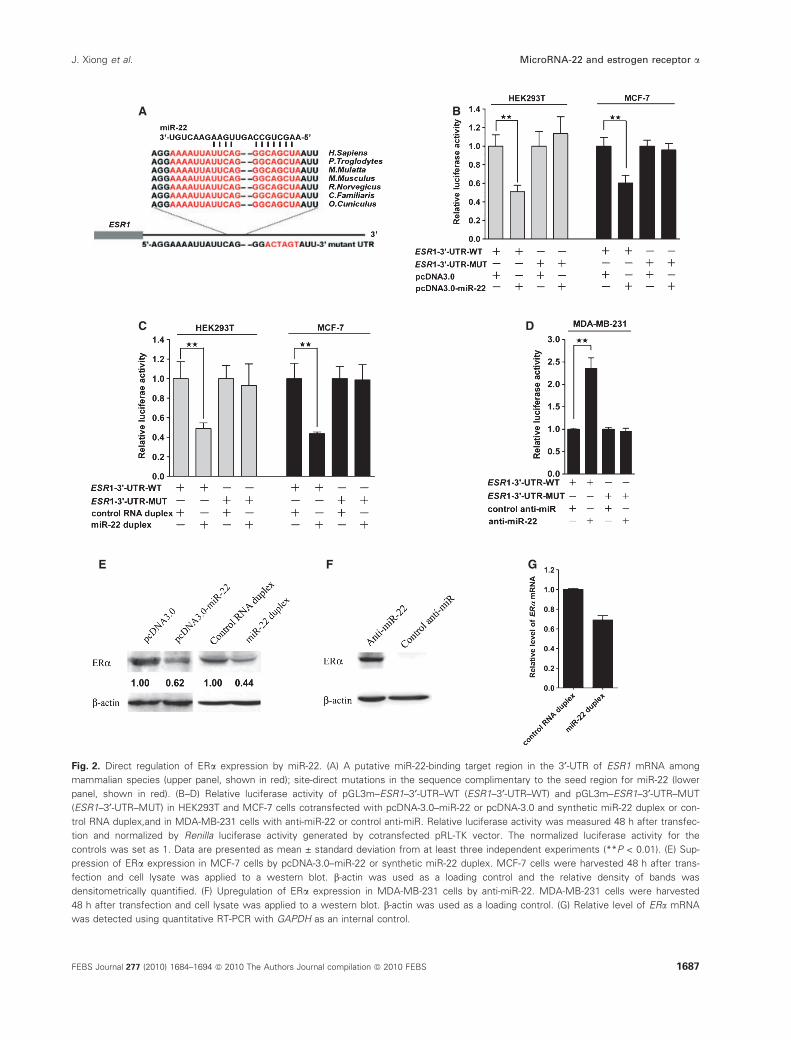
We therefore constructed a reporter plasmid
(pGL3m–ESR1–3¢-UTR–WT) with the 4.3 kb ESR1
3¢-UTR cloned downstream to a firefly luciferase
reporter gene and used both vector-expressed miR-22
and synthetic miR-22 to evaluate the suppression
effects of the miRNAs on the reporter gene expression.
It was found that in HEK293T and ERa-positiveMCF-7 cells, miR-22 had a potent inhibitory effect on
the expression of the reporter gene with the ESR1
3¢-UTR tag (Fig. 2B, C). To examine whether the ERasilencing is mediated by specific and direct interaction
of miR-22 with the ESR1 target site, the complemen-
tary site for the miR-22 seed region was mutated to
J. Xiong et al. MicroRNA-22 and estrogen receptor a
FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS 1685
form pGL3m–ESR1–3¢-UTR–MUT (Fig. 2A). Both
pcDNA3.0–miR-22 and miR-22 duplex reduced lucif-
erase activities expressed in pGL3m–ESR1–3¢-UTR–
WT by � 50%, but such a reduction was completely
abolished in pGL3m–ESR1-3¢–UTR–MUT (Fig. 2B,
C). Moreover, knockdown of endogenous miR-22 in
MDA-MB-231 cells that express a relatively high level
miR-22 could elevate the luciferase signal of pGL3m–
ESR1–3¢-UTR–WT (Fig. 2D), further suggesting that
silencing of ERa was indeed by the interaction of
miR-22 with the 3¢-UTR of ESR1.
The effect of miR-22 on endogenous ERa protein
levels was also examined in MCF-7 and MDA-MB-
231 cells. The results showed that an ectopic increase
in either synthesized or vector-expressed miR-22 in
MCF-7 led to an �50% reduction in ERa protein
levels (Fig. 2E). Conversely, ERa expression was sig-
nificantly elevated by inhibiting endogenous miR-22 in
MDA-MB-231 cells (Fig. 2F). Interestingly, the reduc-
tion in ERa protein levels was markedly greater than
the reduction in ESR1 mRNA levels determined by
quantitative RT-PCR (Fig. 2G). These results demon-
strated that miR-22 could regulate ERa expression by
directly binding to ERa 3¢-UTR, and inhibited ERaexpression through both destabilizing mRNA and
inhibiting translation.
A
B
Fig. 1. Identification of miRNAs that might
target ERa 3¢-UTR. (A) The 59 miRNAs
predicted as having broadly conserved sites
by the TARGETSCAN program. Partial miRNA
families broadly conserved among verte-
brates were enumerated according to their
conserved target positions. (B) The effects
of the predicted 59 miRNAs as well as three
interested miRNAs (miR-206, miR-9 and
miR-1) on reporter gene expression of
pGL3m–ESR1–3¢-UTR–WT. Relative lucifer-
ase activity was measured 48 h after
transfection and normalized by Renilla
luciferase activity generated by
cotransfected pRL-TK vector. The
normalized luciferase activity for the controls
was set as 100%. Data are presented as
mean ± standard deviation from at least
three independent experiments.
MicroRNA-22 and estrogen receptor a J. Xiong et al.
1686 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS
A B
C D
GFE
Fig. 2. Direct regulation of ERa expression by miR-22. (A) A putative miR-22-binding target region in the 3¢-UTR of ESR1 mRNA among
mammalian species (upper panel, shown in red); site-direct mutations in the sequence complimentary to the seed region for miR-22 (lower
panel, shown in red). (B–D) Relative luciferase activity of pGL3m–ESR1–3¢-UTR–WT (ESR1–3¢-UTR–WT) and pGL3m–ESR1–3¢-UTR–MUT
(ESR1–3¢-UTR–MUT) in HEK293T and MCF-7 cells cotransfected with pcDNA-3.0–miR-22 or pcDNA-3.0 and synthetic miR-22 duplex or con-
trol RNA duplex,and in MDA-MB-231 cells with anti-miR-22 or control anti-miR. Relative luciferase activity was measured 48 h after transfec-
tion and normalized by Renilla luciferase activity generated by cotransfected pRL-TK vector. The normalized luciferase activity for the
controls was set as 1. Data are presented as mean ± standard deviation from at least three independent experiments (**P < 0.01). (E) Sup-
pression of ERa expression in MCF-7 cells by pcDNA-3.0–miR-22 or synthetic miR-22 duplex. MCF-7 cells were harvested 48 h after trans-
fection and cell lysate was applied to a western blot. b-actin was used as a loading control and the relative density of bands was
densitometrically quantified. (F) Upregulation of ERa expression in MDA-MB-231 cells by anti-miR-22. MDA-MB-231 cells were harvested
48 h after transfection and cell lysate was applied to a western blot. b-actin was used as a loading control. (G) Relative level of ERa mRNA
was detected using quantitative RT-PCR with GAPDH as an internal control.
J. Xiong et al. MicroRNA-22 and estrogen receptor a
FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS 1687
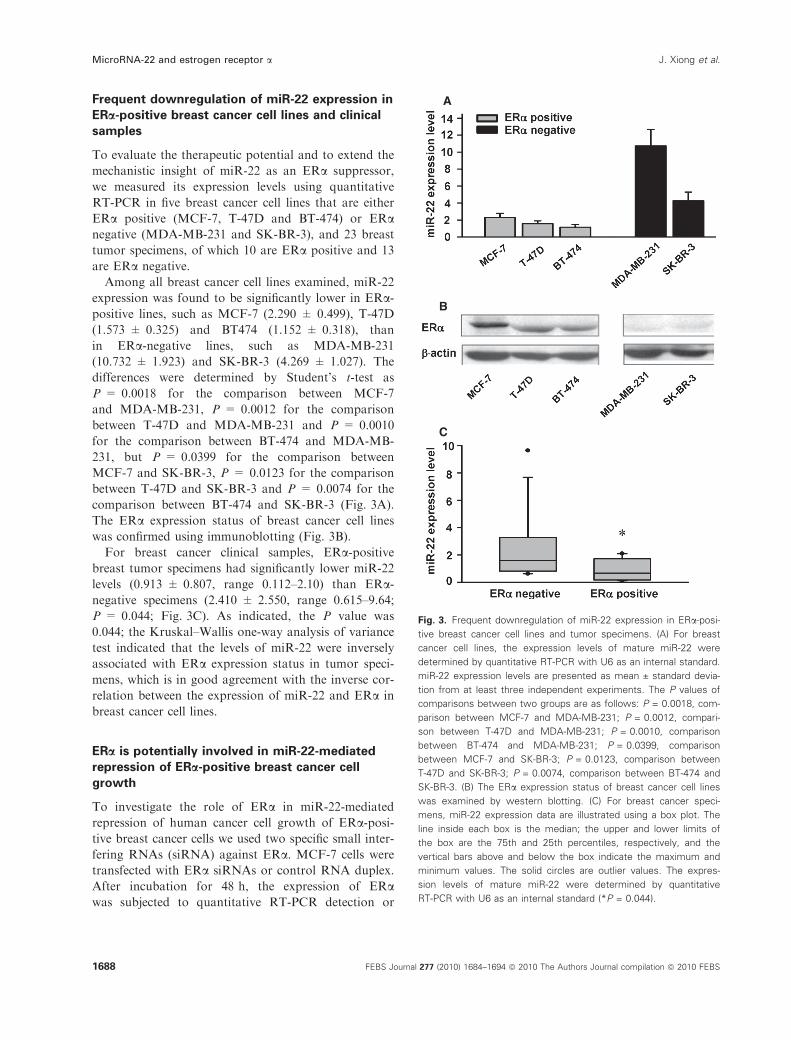
Frequent downregulation of miR-22 expression in
ERa-positive breast cancer cell lines and clinical
samples
To evaluate the therapeutic potential and to extend the
mechanistic insight of miR-22 as an ERa suppressor,
we measured its expression levels using quantitative
RT-PCR in five breast cancer cell lines that are either
ERa positive (MCF-7, T-47D and BT-474) or ERanegative (MDA-MB-231 and SK-BR-3), and 23 breast
tumor specimens, of which 10 are ERa positive and 13
are ERa negative.
Among all breast cancer cell lines examined, miR-22
expression was found to be significantly lower in ERa-positive lines, such as MCF-7 (2.290 ± 0.499), T-47D
(1.573 ± 0.325) and BT474 (1.152 ± 0.318), than
in ERa-negative lines, such as MDA-MB-231
(10.732 ± 1.923) and SK-BR-3 (4.269 ± 1.027). The
differences were determined by Student’s t-test as
P = 0.0018 for the comparison between MCF-7
and MDA-MB-231, P = 0.0012 for the comparison
between T-47D and MDA-MB-231 and P = 0.0010
for the comparison between BT-474 and MDA-MB-
231, but P = 0.0399 for the comparison between
MCF-7 and SK-BR-3, P = 0.0123 for the comparison
between T-47D and SK-BR-3 and P = 0.0074 for the
comparison between BT-474 and SK-BR-3 (Fig. 3A).
The ERa expression status of breast cancer cell lines
was confirmed using immunoblotting (Fig. 3B).
For breast cancer clinical samples, ERa-positivebreast tumor specimens had significantly lower miR-22
levels (0.913 ± 0.807, range 0.112–2.10) than ERa-negative specimens (2.410 ± 2.550, range 0.615–9.64;
P = 0.044; Fig. 3C). As indicated, the P value was
0.044; the Kruskal–Wallis one-way analysis of variance
test indicated that the levels of miR-22 were inversely
associated with ERa expression status in tumor speci-
mens, which is in good agreement with the inverse cor-
relation between the expression of miR-22 and ERa in
breast cancer cell lines.
ERa is potentially involved in miR-22-mediated
repression of ERa-positive breast cancer cell
growth
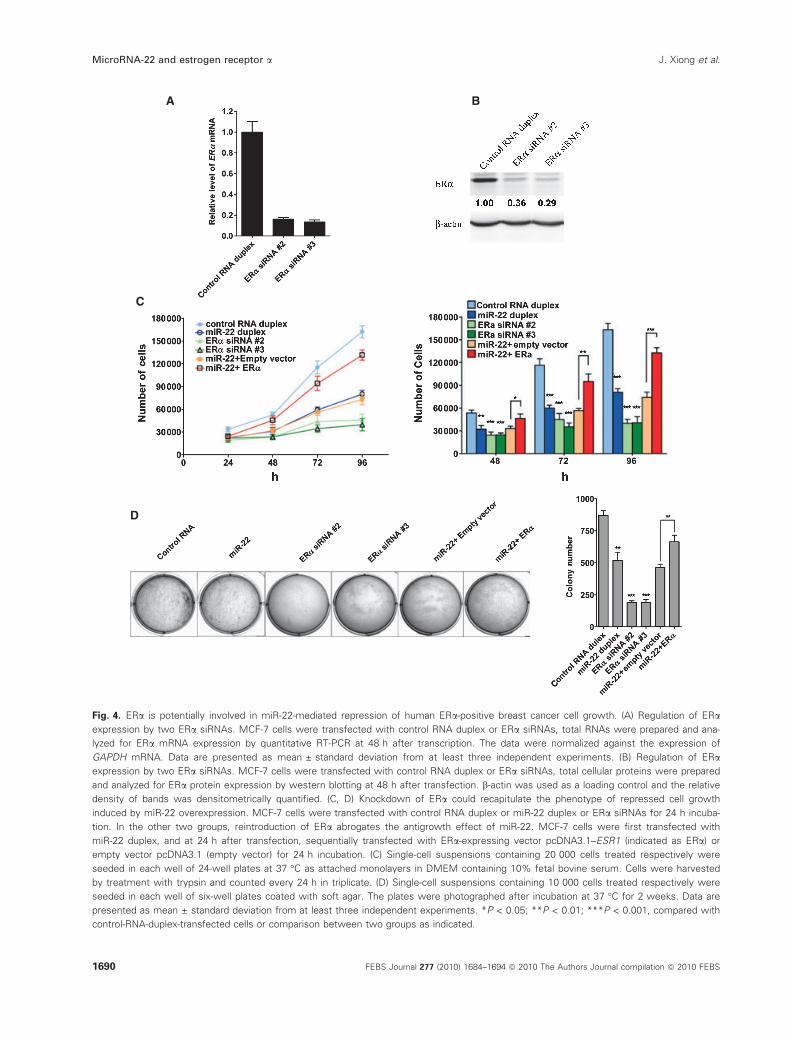
To investigate the role of ERa in miR-22-mediated
repression of human cancer cell growth of ERa-posi-tive breast cancer cells we used two specific small inter-
fering RNAs (siRNA) against ERa. MCF-7 cells were
transfected with ERa siRNAs or control RNA duplex.
After incubation for 48 h, the expression of ERawas subjected to quantitative RT-PCR detection or
A
B
C
Fig. 3. Frequent downregulation of miR-22 expression in ERa-posi-
tive breast cancer cell lines and tumor specimens. (A) For breast
cancer cell lines, the expression levels of mature miR-22 were
determined by quantitative RT-PCR with U6 as an internal standard.
miR-22 expression levels are presented as mean ± standard devia-
tion from at least three independent experiments. The P values of
comparisons between two groups are as follows: P = 0.0018, com-
parison between MCF-7 and MDA-MB-231; P = 0.0012, compari-
son between T-47D and MDA-MB-231; P = 0.0010, comparison
between BT-474 and MDA-MB-231; P = 0.0399, comparison
between MCF-7 and SK-BR-3; P = 0.0123, comparison between
T-47D and SK-BR-3; P = 0.0074, comparison between BT-474 and
SK-BR-3. (B) The ERa expression status of breast cancer cell lines
was examined by western blotting. (C) For breast cancer speci-
mens, miR-22 expression data are illustrated using a box plot. The
line inside each box is the median; the upper and lower limits of
the box are the 75th and 25th percentiles, respectively, and the
vertical bars above and below the box indicate the maximum and
minimum values. The solid circles are outlier values. The expres-
sion levels of mature miR-22 were determined by quantitative
RT-PCR with U6 as an internal standard (*P = 0.044).
MicroRNA-22 and estrogen receptor a J. Xiong et al.
1688 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS

immunoblot analysis. The result showed that the two
siRNAs could reduce ERa mRNA and protein level
significantly (Fig. 4A,B). Further functional studies
showed that knockdown of ERa by the two siRNAs
could mimic the inhibitory effect of miR-22 on the
proliferation and colony formation of breast cancer
cells, whereas a control siRNA duplex did not show
an effect (Fig. 4C,D).
We then went further to determine whether overex-
pression of ERa could counterbalance the antigrowth
effect of miR-22 on MCF-7 cells. We forced MCF-7
cells to express ERa constitutively using a construct
encoding the entire encoding region of ERa mRNA, but
lacking the ERa 3¢-UTR, thus yielding an mRNA that
is resistant to miR-22-mediated inhibition of translation.
Indeed, we found that the miR-22-induced cell growth
repression phenotype was partially rescued by the intro-
duction of this vector expressing an miRNA-resistant
ERa transcript (Fig. 4C, D). These findings suggest that
ERa plays an important role in miR-22-retarded growth
of ERa-positive breast cancer cells.
Discussion
To date, more than 700 human miRNAs have been
identified using experiment-driven methods and compu-
tation-driven approaches [24,25]. miRNAs have diverse
expression patterns in different cell types and it is well
accepted that miRNAs regulate numerous physiological
and pathological processes [1,26]. The biological func-
tion of most miRNAs is, however, largely unknown.
miRNAs have been relatively better investigated in
tumor cells and it has already been shown that
miRNAs can function as both tumor suppressors and
oncogenes by directly regulating genes involved in
related pathways. Unrestrained cell proliferation and
deregulated cell death underlie neoplastic progression in
almost all cancer types [13,27]. An increasing number of
miRNAs have been implicated in tumorigenesis via the
regulation of cancer cell proliferation and growth. For
instance, let-7 can inhibit proliferation of lung and liver
cancer cells by targeting multiple cell cycle oncogenes
[28] and miR-34b and miR-34c have a cooperative
negative effect on proliferation and colony formation
of ovarian cancer cells [6], whereas overexpression of
the miR-17-92 cluster miRNAs enhance lung cancer
cell proliferation and growth as oncogenes [29].
The highly conserved human miR-22 gene is located
at a fragile cancer-relevant genomic region in chromo-
some 17 (17p13.3), and mapped to an exon of the
C17orf91 gene [4,30]. To date, several genes, including
HOXA6, HOXA4, HSPG2, GPNMB, CLIC4 and SP1,
have been predicted as targets of miR-22 [31–33],
whereas ERa has been suggested as a direct target of
this miRNA in a recent work [34]. miRNA expression
profiling data revealed that miR-22 had a great reduc-
tion in acute myeloid leukemia with mutations in
NPM1 compared with acute myeloid leukemia without
NPM1 mutations, and HOXA6–HOXA4 were pre-
dicted as targets of miR-22 [31]. miR-22 has been
detected with a distinct expression pattern in human
Duchenne muscular dystrophy, where HSPG2,
GPNMB and CLIC4 were predicted as its potential
targets [32]. HSPG2 has been reported to contribute to
tumor growth and angiogenesis in vivo [35], and
GPNMB was identified as a pathological and diagnos-
tic marker in melanocyte tumor progression [36].
Moreover, CLIC4 was found to participate in stress-
induced apoptosis in human osteosarcoma cells [37].
Flow cytometry analysis showed that overexpression
of miR-22 could reduce ERa and SP1 protein levels in
pancreatic cancer cells [33].
Estrogen receptors (mainly ERa and ERb) constitutea group of ligand-activated nuclear receptors that are
activated by estrogen. Human ERa is a transcription
factor that regulates diverse gene expression, and is
implicated in cancers by stimulating cell proliferation
and tumor growth [38,39]. An miRNA library-based
screening with miR-206, miR-18a and miR-221 ⁄ 222 as
putative positive controls [17,40,41] demonstrated that
miR-22 could robustly suppress the luciferase signal of
ERa 3¢-UTR tethered vector. By mutating the comple-
mentary site for the miR-22 seed region, we showed that
repression of ERa by miR-22 was almost completely
abolished. Conversely, the ERa signal intensities were
significantly elevated by knockdown of endogenous
miR-22 in MDA-MB-231 cells that expressed a rela-
tively high level of miR-22. These results demonstrate
that ERa is a direct target of miR-22. miR-22 treatment
was found to dramatically reduce the endogenous trans-
lational yield of ERa, and knockdown of endogenous
miR-22 could elevate ERa protein expression.
Because ERa expression is routinely monitored in
breast cancer samples as a prognostic marker, we went
further to assess the correlation between miR-22
expression and ERa protein levels in breast cancer cell
lines and surgical specimens. It was interesting to find
that downregulation of miR-22 expression occurs fre-
quently, not only in ERa-positive human breast cancer
cell lines, but also in surgical specimens compared with
ERa-negative counterparts. This result made it appeal-
ing to examine whether miR-22 could also be used as
a marker for the identification of breast cancer sub-
types in addition to ERa itself, as miR-22 probably
regulates a different set of genes in comparison with
the regulatory profile of ERa.
J. Xiong et al. MicroRNA-22 and estrogen receptor a
FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS 1689
A
C
D
B
Fig. 4. ERa is potentially involved in miR-22-mediated repression of human ERa-positive breast cancer cell growth. (A) Regulation of ERa
expression by two ERa siRNAs. MCF-7 cells were transfected with control RNA duplex or ERa siRNAs, total RNAs were prepared and ana-
lyzed for ERa mRNA expression by quantitative RT-PCR at 48 h after transcription. The data were normalized against the expression of
GAPDH mRNA. Data are presented as mean ± standard deviation from at least three independent experiments. (B) Regulation of ERa
expression by two ERa siRNAs. MCF-7 cells were transfected with control RNA duplex or ERa siRNAs, total cellular proteins were prepared
and analyzed for ERa protein expression by western blotting at 48 h after transfection. b-actin was used as a loading control and the relative
density of bands was densitometrically quantified. (C, D) Knockdown of ERa could recapitulate the phenotype of repressed cell growth
induced by miR-22 overexpression. MCF-7 cells were transfected with control RNA duplex or miR-22 duplex or ERa siRNAs for 24 h incuba-
tion. In the other two groups, reintroduction of ERa abrogates the antigrowth effect of miR-22. MCF-7 cells were first transfected with
miR-22 duplex, and at 24 h after transfection, sequentially transfected with ERa-expressing vector pcDNA3.1–ESR1 (indicated as ERa) or
empty vector pcDNA3.1 (empty vector) for 24 h incubation. (C) Single-cell suspensions containing 20 000 cells treated respectively were
seeded in each well of 24-well plates at 37 �C as attached monolayers in DMEM containing 10% fetal bovine serum. Cells were harvested
by treatment with trypsin and counted every 24 h in triplicate. (D) Single-cell suspensions containing 10 000 cells treated respectively were
seeded in each well of six-well plates coated with soft agar. The plates were photographed after incubation at 37 �C for 2 weeks. Data are
presented as mean ± standard deviation from at least three independent experiments. *P < 0.05; **P < 0.01; ***P < 0.001, compared with
control-RNA-duplex-transfected cells or comparison between two groups as indicated.
MicroRNA-22 and estrogen receptor a J. Xiong et al.
1690 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS

In summary, we showed that frequent downregula-
tion of miR-22 expression is associated with ERa-posi-tive human breast cancer cells, and miR-22 can
directly regulate ERa expression. We further showed
that ERa is potentially involved in miR-22-mediated
repression of human cancer cell growth of ERa-posi-tive breast cancer cells. It would then be interesting to
explore whether miR-22 could serve as a potential
therapeutic reagent in the treatment of cancer in which
ERa plays an important role.
Materials and methods
Cell lines and cultures
Five breast carcinoma cell lines were obtained from the Cell
Resource Center of Peking Union Medical College (Beijing,
China) and maintained in our laboratory. HEK293T
(American Type Culture Collection, Manassas, VA, USA)
and MCF-7 cells were maintained in 10% fetal bovine
serum-supplemented Dulbecco’s modified Eagle’s medium
(DMEM) (Hyclone, Logan, UT, USA); MDA-MB-231 cells
were maintained in 10% fetal bovine serum-supplemented
L-15 (Gibco, Grand Island, NY, USA); SK-BR-3 cells were
maintained in 10% fetal bovine serum-supplemented
RPMI-1640 (Hyclone); T-47D and BT-474 cells were
maintained in RPMI-1640 (Hyclone) plus 10% fetal bovine
serum and 0.2 UÆmL)1 insulin.
Vector construction
A DNA segment encompassing the mature miR-22
sequence and its 5¢- and 3¢-flanking regions (130 and
80 bp, respectively) was cloned into the BamHI and XhoI
sites in pcDNA3.0 (Invitrogen, Carlsbad, CA, USA) to
create the miR-22 expression vector pcDNA3.0–miR-22.
pGL3m was modified from a firefly luciferase-expressing
vector pGL3-control (Promega, Madison, WI, USA) by
inserting a multiple cloning sequence downstream of the
XbaI site, including EcoRV, ApaI, SacII, NdeI, PstI,
EcoRI and NruI sites. The insertion site is immediately
downstream of the stop codon of the firefly luciferase
reporter gene. A 4.3 kb fragment encoding the full-length
3¢-UTR of human ESR1 mRNA (Genbank accession no.
NM_000125) was cloned between the SacII and EcoRI
sites in pGL3m, forming pGL3m–ESR1–3¢-UTR–WT, in
which, site-specific mutations were performed to disrupt
the binding site of miR-22, forming pGL3m–ESR1–3¢-UTR–MUT. The ERa-expressing vector (pcDNA3.1–
ESR1) was created by cloning the ESR1 coding sequence
into the EcoRI and NheI sites of pcDNA3.1 (Invitrogen).
The primers used in the subcloning experiments are
included in Table S2; all the construct products were
validated by sequencing.
RNA isolation and quantitative RT-PCR detection
Total RNAs from cultured cells were isolated using TRI
Reagent (Sigma, St Louis, MO, USA) and converted into
cDNA using ImPro-II reverse transcriptase (Promega).
Detection of the mature form of miR-22 was performed
using Quantitect SYBR Green PCR Kit (Qiagen, Hilden,
Germany) and quantitative RT-PCR Primer Sets (Ribo-
bio, Guangzhou, China) with the U6 small nuclear RNA
as an internal control. Detection of the ERa mRNA was
performed using Quantitect SYBR Green PCR Kit
(Qiagen), with glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) mRNA as an internal control. The detection
primers for ESR1 and GAPDH are included in Table S2.
RNA oligoribonucleotides and cell transfections
The miRNA mimic library was obtained from Ribobio
(Guangzhou, China); miR-22 duplex and the negative con-
trol RNA duplex (indicated as control RNA duplex) were
obtained from GenePharma (Shanghai, China) (Fig. S1).
The control RNA duplex was used to eliminate the poten-
tial nonsequence-specific effects and its sequences were non-
homologous to any human genome sequences. MCF-7 cells
were transfected with 50 nm RNA duplex using lipofecta-
mine 2000 (Invitrogen) and counted 24 h after transfection
for plating wells to observe proliferation and colony forma-
tion. The anti-miR-22 was a 2¢-O-methyl-modified oligori-
bonucleotide designed as an inhibitor of miR-22, and its
sequence is 5¢-ACAGUCUUCAACUGGCAGCUU-3¢. Thenegative control for anti-miR-22 in the antagonism experi-
ments was control anti-miR, with a sequence of 5¢-GUG
GAUAUUGUUGCCAUCA-3¢. The sequences of two siR-
NAs for ESR1 are as follows: ERa siRNA #2 sense strand
5¢-UCAUCGCAUUCC UUGCAAAdTdT-3¢, antisense
strand 5¢- UUUGCAAGGAAUGCGAUGAdTdT-3¢; ERasiRNA #3 sense strand 5¢- GGAGAAUGUUGAAACA
CAAdTdT-3¢, antisense strand 5¢- UUGUGUUUCAA
CAUUCUCCdTdT-3¢. The transfection efficiency was
monitored by fluorescence-activated cell sorting, using a
carboxyfluorescein (FAM)-labeled siRNA.
Western blot
Forty-eight hours after transfection, the cells were lysed
using cell lysis buffer (Cell Signaling Technology, Beverly,
MA, USA). Isolated proteins were separated in 10% SDS
polyacrylamide gels, transferred to polyvinylidene difluoride
membranes (Bio-Rad Laboratories, Hercules, CA, USA),
and detected with antibodies for human ERa (Cell Signal-
ing Technology), b-actin (Santa Cruz Biotechnology, Santa
Cruz, CA, USA) and ECL kit (Santa Cruz Biotechnology).
The intensity of protein bands was quantified using image j
software (National Institutes of Health, Bethesda, MD,
USA).
J. Xiong et al. MicroRNA-22 and estrogen receptor a
FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS 1691
new
高亮
new
高亮

Luciferase reporter assay
For miRNA duplex library analysis, HEK293T and MCF-7
cells per well in 24-well plates were cotransfected with 50 nm
miRNA duplex, 120 ng pGL3m–ESR1–3¢-UTR–WT and
8 ng pRL-TK (Promega) in duplicate. For miR-22 analysis,
cells were cotransfected with plasmids (300 ng pcDNA3–
miR-22 or pcDNA3.0) or duplexes (33 nm miR-22 duplex or
control RNA duplex), 120 ng pGL3m–ESR1–3¢-UTR–WT
or pGL3m–ESR1–3¢-UTR–MUT and 8 ng pRL-TK in trip-
licate. In MDA-MB-231 cells, cells were cotransfected with
200 nm anti-miR-22 or control anti-miR, 120 ng pGL3m–
ESR1–3¢-UTR–WT or pGL3m–ESR1–3¢-UTR–MUT and
8 ng pRL-TK in triplicate. For the above analyses, cell
lysates were analyzed 48 h after transfection using the Dual-
Luciferase Reporter Assay System (Promega) and the experi-
ments were independently repeated at least three times.
Luciferase activity was detected using the Synergy HT micro-
plate fluorescence reader (Bio-Tek Instruments, Winooski,
VT, USA). The pRL-TK vector constitutively expressing
Renilla luciferase was cotransfected as an internal control to
minimize experimental variability caused by the differences
in cell viability or transfection efficiencies.
Tissue specimens and RNA extraction
Fresh breast cancer tissues of 23 individual patients were
procured from surgical resection specimens collected in the
Cancer Hospital, Chinese Academy of Medical Sciences
(Beijing, China) in 2009. The clinical characteristics of
patients with breast carcinoma are shown in Table 1. No
patients received treatment before surgery and they signed
informed consent forms for sample collection. ERa protein
expression status was confirmed at diagnosis using standard
immunohistochemistry procedures. Total RNA was isolated
and then converted to cDNA using miR-22 RT primer
(Ribobio) and ImPro-II reverse transcriptase (Promega).
Soft-agar colony assay
Anchorage-independent growth was carried out in six-well
plates coated with 0.6% soft agar in DMEM plus 10% fetal
bovine serum. Twenty-four hours after transfection, 1 · 104
transfected cells were plated into each well of six-well plates
and maintained in DMEM plus 10% fetal bovine serum for
2 weeks. Colonies were stained with 1.25 mgÆmL)1 nitroblue
tetrazolium for 16 h before imaging.
Statistical analysis
Data are presented as mean ± standard deviation from at
least three independent experiments and differences were
assessed using Student’s t test. The Kruskal–Wallis one-way
analysis of variance test was used to test the significance of
association between ERa status and the levels of miR-22 in
tumor specimens. These statistical analyses were imple-
mented in statistic analysis system software (version 8.0,
SAS Institute). P < 0.05 was used as the criterion for
statistical significance; all statistical tests were two-sided.
Acknowledgements
We thank Dr Yangming Wang for critical reading of
the manuscript. This work was supported by
the National High-tech R&D Program of China
(2007AA02Z165, 2008DFA30770), the National Basic
Research Program of China (2007CB512100), and the
National Foundation of Natural Science (grant
30871385).
References
1 Bartel DP (2004) MicroRNAs: genomics, biogenesis,
mechanism, and function. Cell 116, 281–297.
2 Lu M, Zhang Q, Deng M, Miao J, Guo Y, Gao W &
Cui Q (2008) An analysis of human microRNA and
disease associations. PLoS ONE 3, e3420.
3 Esquela-Kerscher A& Slack FJ (2006) Oncomirs-microR-
NAs with a role in cancer.Nat Rev Cancer 6, 259–269.
4 Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch
E, Yendamuri S, Shimizu M, Rattan S, Bullrich F,
Negrini M et al. (2004) Human microRNA genes are
frequently located at fragile sites and genomic regions
involved in cancers. Proc Natl Acad Sci USA 101,
2999–3004.
5 Zhang L, Huang J, Yang N, Greshock J, Megraw MS,
Giannakakis A, Liang S, Naylor TL, Barchetti A, Ward
MR et al. (2006) microRNAs exhibit high frequency
genomic alterations in human cancer. Proc Natl Acad
Sci USA 103, 9136–9141.
6 Corney DC, Flesken-Nikitin A, Godwin AK, Wang W
& Nikitin AY (2007) MicroRNA-34b and MicroRNA-
34c are targets of p53 and cooperate in control of cell
Table 1. Clinical characteristics of patients with breast carcinoma
Variable
ERa positive ERa negative
n (%) n (%)
Total no. 10 (43.5) 13 (56.5)
Age
< 50 years 3 (30.0) 6 (46.2)
‡ 50 years 7 (70.0) 7 (53.8)
Tumor size
£ 2 cm 5 (50.0) 7 (53.8)
> 2 cm 5 (50.0) 6 (46.2)
Lymph node metatasis
No 4 (40.0) 8 (61.5)
Yes 6 (60.0) 5 (38.5)
Histology
Invasive ductal carcinoma 10 (100) 12 (92.3)
Medullary carcinoma 0 (0) 1 (7.7)
MicroRNA-22 and estrogen receptor a J. Xiong et al.
1692 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS
new
高亮

proliferation and adhesion-independent growth. Cancer
Res 67, 8433–8438.
7 Cimmino A, Calin GA, Fabbri M, Iorio MV, Ferracin
M, Shimizu M, Wojcik SE, Aqeilan RI, Zupo S, Dono M
et al. (2005) miR-15 and miR-16 induce apoptosis by tar-
geting BCL2. Proc Natl Acad Sci USA 102, 13944–13949.
8 Ma L, Teruya-Feldstein J & Weinberg RA (2007)
Tumour invasion and metastasis initiated by micro-
RNA-10b in breast cancer. Nature 449, 682–688.
9 Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J,
Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferran-
do AA et al. (2005) MicroRNA expression profiles clas-
sify human cancers. Nature 435, 834–838.
10 Castanotto D & Rossi JJ (2009) The promises and pit-
falls of RNA-interference-based therapeutics. Nature
457, 426–433.
11 Parkin DM, Bray F, Ferlay J & Pisani P (2005) Global
cancer statistics, 2002. CA Cancer J Clin 55, 74–108.
12 Wooster R & Weber BL (2003) Breast and ovarian can-
cer. N Engl J Med 348, 2339–2347.
13 Hanahan D & Weinberg RA (2000) The hallmarks of
cancer. Cell 100, 57–70.
14 Zhu S, Si ML, Wu H & Mo YY (2007) MicroRNA-21
targets the tumor suppressor gene tropomyosin 1
(TPM1). J Biol Chem 282, 14328–14336.
15 Iorio MV, Casalini P, Piovan C, Di Leva G, Merlo A,
Triulzi T, Menard S, Croce CM & Tagliabue E (2009)
microRNA-205 regulates HER3 in human breast can-
cer. Cancer Res 69, 2195–2200.
16 Hossain A, Kuo MT & Saunders GF (2006) Mir-17-5p
regulates breast cancer cell proliferation by inhibiting
translation of AIB1 mRNA. Mol Cell Biol 26,
8191–8201.
17 Adams BD, Furneaux H & White BA (2007) The
micro-ribonucleic acid (miRNA) miR-206 targets the
human estrogen receptor-a (ERa) and represses ERamessenger RNA and protein expression in breast cancer
cell lines. Mol Endocrinol 21, 1132–1147.
18 Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo
R, Sabbioni S, Magri E, Pedriali M, Fabbri M,
Campiglio M et al. (2005) MicroRNA gene expression
deregulation in human breast cancer. Cancer Res 65,
7065–7070.
19 Datta J, Kutay H, Nasser MW, Nuovo GJ, Wang B,
Majumder S, Liu CG, Volinia S, Croce CM, Schmitt-
gen TD et al. (2008) Methylation mediated silencing of
microRNA-1 gene and its role in hepatocellular carcino-
genesis. Cancer Res 68, 5049–5058.
20 Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP &
Burge CB (2003) Prediction of mammalian microRNA
targets. Cell 115, 787–798.
21 Rajewsky N (2006) MicroRNA target predictions in
animals. Nat Genet 38(Suppl), S8–S13.
22 Zhao Y, Samal E & Srivastava D (2005) Serum
response factor regulates a muscle-specific microRNA
that targets Hand2 during cardiogenesis. Nature 436,
214–220.
23 Kuhn DE, Martin MM, Feldman DS, Terry AV Jr,
Nuovo GJ & Elton TS (2008) Experimental validation
of miRNA targets. Methods 44, 47–54.
24 Berezikov E, Cuppen E & Plasterk RH (2006)
Approaches to microRNA discovery. Nat Genet 38
(Suppl), S2–S7.
25 Griffiths-Jones S, Saini HK, van Dongen S & Enright
AJ (2008) miRBase: tools for microRNA genomics.
Nucleic Acids Res 36, D154–D158.
26 He L & Hannon GJ (2004) MicroRNAs: small RNAs
with a big role in gene regulation. Nat Rev Genet 5,
522–531.
27 Evan GI & Vousden KH (2001) Proliferation, cell cycle
and apoptosis in cancer. Nature 411, 342–348.
28 Johnson CD, Esquela-Kerscher A, Stefani G, Byrom
M, Kelnar K, Ovcharenko D, Wilson M, Wang X,
Shelton J, Shingara J et al. (2007) The let-7 microRNA
represses cell proliferation pathways in human cells.
Cancer Res 67, 7713–7722.
29 Hayashita Y, Osada H, Tatematsu Y, Yamada H,
Yanagisawa K, Tomida S, Yatabe Y, Kawahara K,
Sekido Y & Takahashi T (2006) A polycistronic micr-
oRNA cluster, miR-17-92, is overexpressed in human
lung cancers and enhances cell proliferation. Cancer Res
65, 9628–9632.
30 Lagos-Quintana M, Rauhut R, Lendeckel W & Tuschl
T (2001) Identification of novel genes coding for small
expressed RNAs. Science 294, 853–858.
31 Garzon R, Garofalo M, Martelli MP, Briesewitz R,
Wang L, Fernandez-Cymering C, Volinia S, Liu CG,
Schnittger S, Haferlach T et al. (2008) Distinctive micr-
oRNA signature of acute myeloid leukemia bearing
cytoplasmic mutated nucleophosmin. Proc Natl Acad
Sci USA 105, 3945–3950.
32 Eisenberg I, Eran A, Nishino I, Moggio M, Lamperti
C, Amato AA, Lidov HG, Kang PB, North KN, Mitra-
ni-Rosenbaum S et al. (2007) Distinctive patterns of
microRNA expression in primary muscular disorders.
Proc Natl Acad Sci USA 104, 17016–17021.
33 Sun M, Estrov Z, Ji Y, Coombes KR, Harris DH &
Kurzrock R (2008) Curcumin (diferuloylmethane) alters
the expression of microRNAs in human pancreatic can-
cer cells. Mol Cancer Ther 7, 464–473.
34 Pandey DP & Picard D (2009) MiR-22 inhibits estrogen
signaling by directly targeting the estrogen receptor
alpha mRNA. Mol Cell Biol 29, 3783–3790.
35 Sharma B, Handler M, Eichstetter I, Whitelock JM,
Nugent MA & Iozzo RV (1998) Antisense targeting of
perlecan blocks tumor growth and angiogenesis in vivo.
J Clin Invest 102, 1599–1608.
36 Weterman MA, Ajubi N, van Dinter IM, Degen WG,
van Muijen GN, Ruitter DJ & Bloemers HP (1995)
nmb, a novel gene, is expressed in low-metastatic
J. Xiong et al. MicroRNA-22 and estrogen receptor a
FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS 1693

human melanoma cell lines and xenografts. Int J Cancer
60, 73–81.
37 Fernandez-Salas E, Suh KS, Speransky VV, Bowers
WL, Levy JM, Adams T, Pathak KR, Edwards LE,
Hayes DD, Cheng C et al. (2002) mtCLIC ⁄CLIC4, anorganellular chloride channel protein, is increased by
DNA damage and participates in the apoptotic
response to p53. Mol Cell Biol 22, 3610–3620.
38 Deroo BJ & Korach KS (2006) Estrogen receptors and
human disease. J Clin Invest 116, 561–570.
39 Clarke RB, Anderson E & Howell A (2004) Steroid
receptors in human breast cancer. Trends Endocrinol
Metab 15, 316–323.
40 Liu WH, Yeh SH, Lu CC, Yu SL, Chen HY, Lin CY,
Chen DS & Chen PJ (2009) MicroRNA-18a prevents
estrogen receptor-alpha expression, promoting prolifera-
tion of hepatocellular carcinoma cells. Gastroenterology
136, 683–693.
41 Zhao JJ, Lin J, Yang H, Kong W, He L, Ma X,
Coppola D & Cheng JQ (2008) MicroRNA-221 ⁄ 222negatively regulates estrogen receptor alpha and is
associated with tamoxifen resistance in breast cancer.
J Biol Chem 283, 31079–31086.
Supporting information
The following supplementary material is available:
Fig. S1. The sequences of synthetic miR-22 duplex and
negative control RNA duplex.
Table S1. Partial predicted miRNAs that target ESR1
3¢-UTR by targetscan, miranda or pictar.
Table S2. Primers for cloning miR-22, ESR1 3¢-UTR
and ESR1 coding sequence and for the detection of
ESR1 and GAPDH mRNA by quantitative RT-PCR.
This supplementary material can be found in the
online version of this article.
Please note: As a service to our authors and readers,
this journal provides supporting information supplied
by the authors. Such materials are peer-reviewed and
may be re-organized for online delivery, but are not
copy-edited or typeset. Technical support issues arising
from supporting information (other than missing files)
should be addressed to the authors.
MicroRNA-22 and estrogen receptor a J. Xiong et al.
1694 FEBS Journal 277 (2010) 1684–1694 ª 2010 The Authors Journal compilation ª 2010 FEBS
Related Documents











