An anti-steroidogenic inhibitory primer pheromone in male sea lamprey (Petromyzon marinus) Yu-Wen Chung-Davidson, Huiyong Wang, Mara B. Bryan 1 , Hong Wu 2 , Nicholas S. Johnson 3 , Weiming Li ⇑ Department of Fisheries and Wildlife, Michigan State University, 480 Wilson Road, East Lansing, MI 48824, USA article info Article history: Received 2 November 2012 Revised 11 April 2013 Accepted 14 April 2013 Available online 30 April 2013 Keywords: GnRH Steroid Bile salt Priming Agnathan abstract Reproductive functions can be modulated by both stimulatory and inhibitory primer pheromones released by conspecifics. Many stimulatory primer pheromones have been documented, but relatively few inhibitory primer pheromones have been reported in vertebrates. The sea lamprey male sex phero- mone system presents an advantageous model to explore the stimulatory and inhibitory primer phero- mone functions in vertebrates since several pheromone components have been identified. We hypothesized that a candidate sex pheromone component, 7a, 12a-dihydroxy-5a-cholan-3-one-24-oic acid (3 keto-allocholic acid or 3kACA), exerts priming effects through the hypothalamic-pituitary–gona- dal (HPG) axis. To test this hypothesis, we measured the peptide concentrations and gene expressions of lamprey gonadotropin releasing hormones (lGnRH) and the HPG output in immature male sea lamprey exposed to waterborne 3kACA. Exposure to waterborne 3kACA altered neuronal activation markers such as jun and jun N-terminal kinase (JNK), and lGnRH mRNA levels in the brain. Waterborne 3kACA also increased lGnRH-III, but not lGnRH-I or -II, in the forebrain. In the plasma, 3kACA exposure decreased all three lGnRH peptide concentrations after 1 h exposure. After 2 h exposure, 3kACA increased lGnRH- I and -III, but decreased lGnRH-II peptide concentrations in the plasma. Plasma lGnRH peptide concentra- tions showed differential phasic patterns. Group housing condition appeared to increase the averaged plasma lGnRH levels in male sea lamprey compared to isolated males. Interestingly, 15a-hydroxyproges- terone (15a-P) concentrations decreased after prolonged 3kACA exposure (at least 24 h). To our knowl- edge, this is the only known synthetic vertebrate pheromone component that inhibits steroidogenesis in males. Ó 2013 Elsevier Inc. All rights reserved. 1. Introduction Reproductive functions can be modulated by both stimulatory and inhibitory primer pheromones released by conspecifics (Reyes et al., 2004). In insects, inhibitory primer pheromones are well doc- umented. The queen retinue pheromone inhibits the ovary devel- opment of worker bees (Butler et al., 1959; Slessor et al., 2005; Trhlin and Rajchard, 2011; Wanner et al., 2007). Similar queen inhibitory pheromones are also found in the ants Camponotus floridanus and Lasius niger (Endler et al., 2004; Holman et al., 2010), and in the termite Reticulitermes speratus (Matsuura et al., 2010). Larval surface chemical cues (brood pheromones) inhibit the activation of the worker bee ovaries (Le Conte et al., 1990, 2001; Maisonnasse et al., 2009; Mohammedi et al., 1998). In addi- tion, termite kings can inhibit the reproduction in males (Fujita and Watanabe, 2010). Many stimulatory primer pheromones have been documented in vertebrates, but relatively few reports have been focused on inhibitory primer pheromones. In mice, juvenile females showed accelerated onset of puberty when exposed to urine from adult males or pregnant females, but showed delayed puberty when ex- posed to urine from females maintained in groups (Drickamer, 1977, 1983; Vandenbergh et al., 1972). Mature female mice, grouped in the absence of a male stimulus, exhibit a suppressed estrus cycle (so-called Lee–Boot effect) (Lee and van der Boot, 1955, 1956; Ma et al., 1998; McClintock, 1983). Isolated female house mice tend to display shorter estrus cycle, while females housed under crowded conditions exhibit irregular or prolonged cycles (Lee and van der Boot, 1955, 1956). The estrus inhibition 0016-6480/$ - see front matter Ó 2013 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.ygcen.2013.04.023 ⇑ Corresponding author. Address: Department of Fisheries and Wildlife, Michigan State University, 13 Natural Resources Building, 480 Wilson Road, East Lansing, MI 48824, USA. Fax: +1 517 432 1699. E-mail address: [email protected] ( Weiming Li). 1 Present address: Energy Biosciences Institute, University of California, 130 Calvin Laboratory, MC 5230, Berkeley, CA 94720, USA. 2 Present address: Department of Microbiology & Immunology, School of Medicine, Emory University, Rollins Research Center G214, 201 Dowman Drive, Atlanta, GA 30322, USA. 3 Present address: U.S. Geological Survey, Great Lakes Science Center, Hammond Bay Biological Station, 11188 Ray Road, Millersburg, MI 49759, USA. General and Comparative Endocrinology 189 (2013) 24–31 Contents lists available at SciVerse ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

General and Comparative Endocrinology 189 (2013) 24–31
Contents lists available at SciVerse ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
An anti-steroidogenic inhibitory primer pheromone in male sealamprey (Petromyzon marinus)
Yu-Wen Chung-Davidson, Huiyong Wang, Mara B. Bryan 1, Hong Wu 2, Nicholas S. Johnson 3,Weiming Li ⇑Department of Fisheries and Wildlife, Michigan State University, 480 Wilson Road, East Lansing, MI 48824, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 2 November 2012Revised 11 April 2013Accepted 14 April 2013Available online 30 April 2013
Keywords:GnRHSteroidBile saltPrimingAgnathan
0016-6480/$ - see front matter � 2013 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.ygcen.2013.04.023
⇑ Corresponding author. Address: Department of FisState University, 13 Natural Resources Building, 480 W48824, USA. Fax: +1 517 432 1699.
E-mail address: [email protected] (Weiming Li).
1 Present address: Energy Biosciences Institute, UnivLaboratory, MC 5230, Berkeley, CA 94720, USA.
2 Present address: Department of Microbiology & ImmEmory University, Rollins Research Center G214, 20130322, USA.
3 Present address: U.S. Geological Survey, Great LakBay Biological Station, 11188 Ray Road, Millersburg, M
Reproductive functions can be modulated by both stimulatory and inhibitory primer pheromonesreleased by conspecifics. Many stimulatory primer pheromones have been documented, but relativelyfew inhibitory primer pheromones have been reported in vertebrates. The sea lamprey male sex phero-mone system presents an advantageous model to explore the stimulatory and inhibitory primer phero-mone functions in vertebrates since several pheromone components have been identified. Wehypothesized that a candidate sex pheromone component, 7a, 12a-dihydroxy-5a-cholan-3-one-24-oicacid (3 keto-allocholic acid or 3kACA), exerts priming effects through the hypothalamic-pituitary–gona-dal (HPG) axis. To test this hypothesis, we measured the peptide concentrations and gene expressions oflamprey gonadotropin releasing hormones (lGnRH) and the HPG output in immature male sea lampreyexposed to waterborne 3kACA. Exposure to waterborne 3kACA altered neuronal activation markers suchas jun and jun N-terminal kinase (JNK), and lGnRH mRNA levels in the brain. Waterborne 3kACA alsoincreased lGnRH-III, but not lGnRH-I or -II, in the forebrain. In the plasma, 3kACA exposure decreasedall three lGnRH peptide concentrations after 1 h exposure. After 2 h exposure, 3kACA increased lGnRH-I and -III, but decreased lGnRH-II peptide concentrations in the plasma. Plasma lGnRH peptide concentra-tions showed differential phasic patterns. Group housing condition appeared to increase the averagedplasma lGnRH levels in male sea lamprey compared to isolated males. Interestingly, 15a-hydroxyproges-terone (15a-P) concentrations decreased after prolonged 3kACA exposure (at least 24 h). To our knowl-edge, this is the only known synthetic vertebrate pheromone component that inhibits steroidogenesis inmales.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction floridanus and Lasius niger (Endler et al., 2004; Holman et al.,
Reproductive functions can be modulated by both stimulatoryand inhibitory primer pheromones released by conspecifics (Reyeset al., 2004). In insects, inhibitory primer pheromones are well doc-umented. The queen retinue pheromone inhibits the ovary devel-opment of worker bees (Butler et al., 1959; Slessor et al., 2005;Trhlin and Rajchard, 2011; Wanner et al., 2007). Similar queeninhibitory pheromones are also found in the ants Camponotus
ll rights reserved.
heries and Wildlife, Michiganilson Road, East Lansing, MI
ersity of California, 130 Calvin
unology, School of Medicine,Dowman Drive, Atlanta, GA
es Science Center, HammondI 49759, USA.
2010), and in the termite Reticulitermes speratus (Matsuura et al.,2010). Larval surface chemical cues (brood pheromones) inhibitthe activation of the worker bee ovaries (Le Conte et al., 1990,2001; Maisonnasse et al., 2009; Mohammedi et al., 1998). In addi-tion, termite kings can inhibit the reproduction in males (Fujitaand Watanabe, 2010).
Many stimulatory primer pheromones have been documentedin vertebrates, but relatively few reports have been focused oninhibitory primer pheromones. In mice, juvenile females showedaccelerated onset of puberty when exposed to urine from adultmales or pregnant females, but showed delayed puberty when ex-posed to urine from females maintained in groups (Drickamer,1977, 1983; Vandenbergh et al., 1972). Mature female mice,grouped in the absence of a male stimulus, exhibit a suppressedestrus cycle (so-called Lee–Boot effect) (Lee and van der Boot,1955, 1956; Ma et al., 1998; McClintock, 1983). Isolated femalehouse mice tend to display shorter estrus cycle, while femaleshoused under crowded conditions exhibit irregular or prolongedcycles (Lee and van der Boot, 1955, 1956). The estrus inhibition

Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31 25
appears to be mediated by olfaction (Champlin, 1971), and theinhibitory signals were presumed to be excreted in the urine(Wolff and Powell, 1979) and controlled by the adrenal gland inmammals (Ma et al., 1998). However, the molecular identities ofthese inhibitory primer pheromones remain elusive (Ma et al.,1998).
The sea lamprey male sex pheromone system presents anadvantageous model to explore the stimulatory and inhibitory pri-mer pheromone functions in vertebrates since several pheromonecomponents have been identified (Li et al., 2012, 2002; Yun et al.,2003). Sexually mature male sea lamprey release sex pheromonescontaining a main component, 7a, 12a, 24-trihydroxy-5a-cholan-3-one 24-sulfate (3 keto-petromyzonol sulfate or 3kPZS), andseveral minor components including 7a, 12a-dihydroxy-5a-cho-lan-3-one-24-oic acid (3 keto-allocholic acid or 3kACA) asconfirmed by behavioral tests, electro-olfactograms and chemicalanalyses (Johnson et al., 2009; Li et al., 2002; Siefkes et al., 2005;Yun et al., 2003). The role of the major component 3kPZS as a relea-ser and stimulatory primer pheromone is well established (Chung-Davidson et al., 2013; Johnson et al., 2009; Li et al., 2002; Siefkeset al., 2005). Waterborne 3kPZS increased plasma 15a-P, forebrainlGnRH-I and -III peptide concentrations and gene expressions, andaltered plasma lGnRH-I and -III peptide levels (Chung-Davidsonet al., 2013). The function of the minor component 3kACA hasyet to be determined.
In this report, we sought to examine the changes in brain geneexpressions and neuroendocrine output in response to synthetic3kACA. We examined the time-course and dose–response effectsof waterborne 3kACA on the plasma concentrations of 15a-P,lGnRH-I and -III, and brain mRNA concentrations of neuronal acti-vation markers such as jun and JNK, three lGnRH-I splice variants(GAP49, 50, and 58; Suzuki et al., 2000) and lGnRH-III. Immediateearly genes Jun and JNK were chosen due to their role in the signal-ing cascades of GnRH receptors (Burger et al., 2009; Haisenlederet al., 2008; Harris et al., 2002; Yokoi et al., 2000). We found that3kACA altered lGnRH synthesis and release, and decreased 15a-Pconcentrations in the plasma. We conclude that 3kACA is an inhib-itory primer pheromone in male sea lamprey.
2. Materials and methods
2.1. Animals
Migratory adult sea lamprey were collected by agents of the U.S.Fish and Wildlife Service Marquette Biological Station (Marquette,MI, USA) and Department of Fisheries and Oceans Canada Sea Lam-prey Control Centre (Sault Ste. Marie, ON, Canada) in early May.Water temperature was around 15 �C when the animals were cap-tured (Binder et al., 2010). Animal treatment and sample collec-tions were conducted at the U.S. Geological Survey, Great LakesScience Center, Hammond Bay Biological Station (Millersburg, MI,USA). For each set of experiments, all test subjects were capturedfrom the same stream on the same day to reduce variation inmaturity. Standard operating procedures for transporting, main-taining, handling, anesthetizing, and euthanizing sea lamprey wereapproved by the Institutional Committee on Animal Use and Careof Michigan State University.
2.2. Pheromone exposure experiment 1
To examine the time course of hormonal responses of immaturemale sea lamprey exposed to waterborne 3kACA at a wide range ofconcentrations, 216 immature males were acclimated for 2 days(test tank: 200 L, 16 �C, water replenishment 1 L/min). 16 �C was
within the optimal temperature range for lamprey reproduction(Hanson and Manion, 1980). Each tank was randomly assigned tovehicle (0.91 ppm methanol), 10�11 M, 10�10 M or 10�9 M 3kACA.During the treatment period, water was aerated but not replen-ished. 1 ml of the vehicle or concentrated pheromone was dec-anted into the tank to achieve the final concentration of 10�11 M,10�10 M or 10�9 M 3kACA. The pheromone was quite stable inthe aquatic environment for up to 1 week (unpublished data).Immediately before and after pheromone exposure, 1 ml of bloodwas drawn from the caudal vein for plasma 15a-P analyses (Bryanet al., 2004, 2006). One-way ANOVA was used to compare thechanges in plasma 15a-P concentration [D15a-P = (post-treatment15a-P level) – (pre-treatment 15a-P level)] among groups treatedwith the same pheromone concentration but sampled at differenttime point. Fisher’s PLSD post hoc tests were performed if the AN-OVA showed significant effects (p < 0.05).
At 0 h, 2 h, 4 h, 8 h, 24 h and 48 h after pheromone exposure,animals were anesthetized with 0.05% MS222 (Sigma, St. Louis,MO, USA) (McVicar and Rankin, 1983), and euthanized by a knifecut at the position of the first gill opening to sever the head andthe body. Six brain samples were snap frozen and analyzed fortranscripts of lGnRH-I (GAP49, GAP50 and GAP58 (Suzuki et al.,2000) and -III, jun, JNK, and 40S ribosomal RNA. These experimentswere performed in 2006 and 2007.
2.3. Real time quantitative PCR (RTQ-PCR)
RTQ-PCR followed the procedure described by Chung-Davidsonet al. (2008). Synthetic oligos were used as standards and ran onthe sample plate. 40s ribosomal RNA was used as an internal stan-dard and confirmed to have no change in the expression level in allexperiments. RTQ-PCR data (among groups treated with samepheromone concentration but sampled at different time point)were analyzed by one-way ANOVA followed by Fisher’s PLSD posthoc tests if the ANOVA showed significant time effects (p < 0.05).The sequence for primers and TaqMan RGB probe (AppliedBiosystems) for each mRNA was listed below. 40S ribosomalRNA: 50 primer (50ACCTACGCAGGAACAGCTATGAC30), probe(50ATCTCGAGCAGCTGAA30), 30 primer (50CGACGAATTCCACCACATTG30). Jun: 50 primer (50CATGGCCGCAAACTTTGG30), probe (50CAC-GAACCTGACCAGC30), 30 primer (50CCACCTCCCTGCTGATGCT30).JNK: 50 primer (50TCAGGCGTGTGGCCAAGT30), probe (50CCATGACTTGATCGAATGT30), 30 primer (50GAATCAAATTGAGAACGCAAACG30). Lamprey GnRH-III: 50 primer (50TGACACGAACCCTGTCAATGA30), probe (50ATGCCCTCGCTGTGGT30), 30 primer(50ACAAAGGGTCTAAGAGACGTCACA30). To analyze three splicevariants of lGnRH-I, we used the same 50 primer (50TGAATTACGCG-CAGCACTACTC30) and probe (50TGGAATGGAAACCCGG30), but the 30
primers were designed so that they were located at the splice junc-tion of each corresponding transcript (GAP49: 50CTCCTCCAGGTCTCGTTTGC30; GAP50: 50CTCCTGCTCCAGGTCTCGTT30; GAP58: 50CCAGCTCTCGTGTGTGACTGA30).
2.4. Pheromone exposure experiment 2
Since the effects of waterborne 3kACA on lGnRH gene expres-sions started after 2 h exposure, and waterborne 3kPZS also in-duced robust GnRH release after 2 h exposure in male sealamprey (Chung-Davidson et al., 2013), we examined GnRH releasein immature males exposed to vehicle or 10�10 M 3kACA at twotime points (1 h and 2 h). In 2011, 56 immature males were accli-mated as described above. Immediately after pheromone exposure,animals were euthanized and blood was drawn by cardiac punc-ture for plasma lGnRH-I, -II and -III analyses using UPLC-MS/MS

26 Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31
(Chung-Davidson et al., 2013). Brain samples were snap-frozen andanalyzed for lGnRH-I, -II and -III peptide concentrations usingUPLC-MS/MS (Chung-Davidson et al., 2013). Analyses of lGnRH-II(Kavanaugh et al., 2008) were added in this experiment. Studentt-tests and F-tests were used to compare the plasma concentra-tions of lGnRHs between the control and 3kACA-treated groupsampled at different time point.
2.5. UPLC-MS/MS analyses of lGnRH-I, -II and -III
Lamprey GnRH-I (HPLC purity 99.8%) and -II (HPLC purity98.9%) standards were custom synthesized from GenScript USAInc. (Piscataway, NJ, USA). lGnRH-III (HPLC purity > 96%) and mam-malian GnRH (HPLC purity > 96%) were purchased from BAChemAmericas, Inc. (Torrance, CA, USA). Each compound was dissolvedin 50% methanol/H2O (v/v) to make 1 mg/ml stock solution, andstored at �20 �C until use. Subsequent dilutions were made in50% methanol/H2O. Calibration standard spiking solutions wereprepared in a range from 0.01 to 10 ng/ml by spiking appropriatestock solutions to brain tissue or plasma extracts. Internal standard(mammalian GnRH) solution (50 ng/ml) was prepared in 50%methanol/H2O and 20 ll was added to each sample (1 ng persample).
Brain samples were weighed and homogenized with 400 ll of1% formic acid in cold (�20 �C) acetonitrile with 1 ng internal stan-dard. 1 ml of 1% formic acid in cold (�20 �C) acetonitrile was addedto the homogenate, incubated at �20 �C for 15 min, and centri-fuged at 15,800�g for 20 min at 4 �C. The supernatant was trans-ferred to a new tube, freeze-dried using a CentriVap Cold TrapConcentrator (Labconco Co., Kansas City, MO, USA), and reconsti-tuted in 1 ml water solution with 3% acetic acid and 1% TFA.500 ll plasma samples were processed with similar procedurewithout the homogenization step. The reconstituted supernatantwas transferred to a SPE HLB cartridge preconditioned with 3 mlmethanol followed by 3 ml loading solution (water/acetic acid/TFA, 96:3:1, v/v), washed with 3 ml loading solution and followedby 3 ml loading solution/methanol (70:30, v/v). Samples wereeluted with 3 ml 3% acetic acid/methanol (30:70, v/v), freeze-driedand reconstituted in 100 ll 3% acetic acid/methanol (50:50, v/v),vortexed for 30 s and transferred to glass autosampler vials forUPLC-MS/MS analyses (Chung-Davidson et al., 2013).
Fig. 1. Synthetic pheromone component 3kACA decreased plasma 15a-hydroxy-progesterone (15a-P) concentrations in immature male sea lamprey. D 15a-P = (plasma 15a-P concentration after treatment) – (plasma 15a-P concentrationbefore treatment). � indicates significant difference between treatment group andvehicle control (p < 0.05). Data are presented as mean ± s.e.m. Sample size, n = 6 forall treatment groups except the groups exposed to 10�10 M 3kACA for 24 h or 48 h(n = 5).
2.6. Plasma GnRH episodic release
Since the lGnRH plasma concentrations fluctuated in the controlgroup, it was difficult to interpret the results of Pheromone Expo-sure Experiment 2. We investigated the episodic release of plasmalGnRHs in male sea lamprey without any pheromone treatment. In2012, 16 immature males were housed in group or isolated in flow-through tanks (200 L, 15 ± 1 �C). Plasma samples were collected byrandom blood draw over 7 days. Prior to each blood draw, animalswere lightly anesthetized with 0.02% MS222 and gills were coveredwith a wet cloth. 1 ml of blood was drawn from the caudal vein.The wound was sealed with VetBond (3 M, Maplewood, MN,USA) and animals were released back to their housing tank. Plasmasamples were extracted immediately and stored at �80 �C untilUPLC-MS/MS analyses for lGnRH-I, -II and -III (Chung-Davidsonet al., 2013). ANOVA was used to compare the plasma concentra-tions of lGnRHs among different housing conditions and timepoints. ‘‘Time’’ and ‘‘housing condition’’ effects were statisticallysignificant (p < 0.05). We then compared the average and the var-iance of lGnRH plasma concentrations between different housingconditions (group vs. isolated, pooled time points) using studentt-tests and F-tests.
3. Results
3.1. Exposure to waterborne 3kACA decreased plasma 15a-Pconcentrations in immature males
Exposure to waterborne 3kACA for at least 24 h decreased plas-ma 15a-P concentrations in immature males (Fig. 1). Exposure to10�11 M or 10�10 M 3kACA decreased 15a-P concentrations after24 h whereas exposure to 10�9 M 3kACA decreased 15a-P concen-trations after 48 h (Fig. 1). There were no apparent dose-dependentresponses. In fact, treatment of 3kACA at higher concentration(10�9 M) seemed to induce delayed response (48 h) when com-pared to lower concentrations (24 h, Fig. 1).
3.2. Exposure to waterborne 3kACA exerted differential effects on braingene expressions
After exposure to waterborne 3kACA at different concentration(10�11 M, 10�10 M or 10�9 M), lGnRH-III mRNA concentrationsshowed rapid and prolonged decreased in the forebrain (Fig. 2).On the other hand, only one of the lGnRH-I splice variants(GAP49) showed decrease in the forebrain after 8 h exposure(Fig. 2). JNK mRNA levels in the forebrain decreased after exposureto 10�10 M or 10�9 M 3kACA for 48 h (Fig. 2), whereas jun mRNAconcentrations increased after exposure to 10�11 M 3kACA for48 h (Fig. 2). Exposure to 10�10 M 3kACA for 8 h increased all threelGnRH-I splice variants in the hindbrain (Fig. 2). Exposure to10�9 M 3kACA for 48 h decreased GAP48, lGnRH-III, jun and JNKmRNA concentrations in the hindbrain (Fig. 2).
3.3. Exposure to waterborne 3kACA increased lGnRH-III peptideconcentrations in the forebrain but exerted differential effects inplasma
Exposure to 10�10 M 3kACA increased lGnRH-III but had no ef-fect on lGnRH-I or -II peptide concentrations in the forebrain(Fig. 3a–c). Exposure to different concentrations of 3kACA had noeffect in the hindbrain of immature males (Fig. 3d–f). Controlmales showed difference in plasma lGnRH-I and -III peptide con-centrations, but not in plasma lGnRH-II at the time point mea-sured. Waterborne 3kACA decreased plasma lGnRH-II peptideconcentrations, but exerted differential effects on plasma lGnRH-Iand -III (Fig. 3g–i).
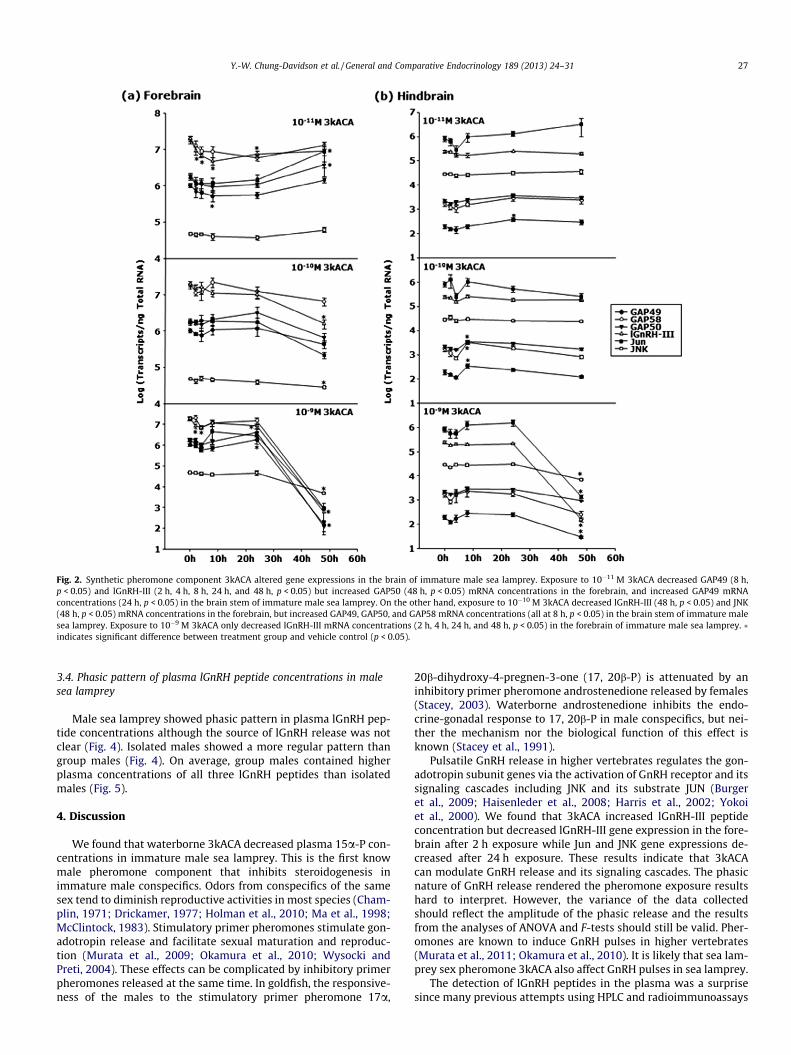
Fig. 2. Synthetic pheromone component 3kACA altered gene expressions in the brain of immature male sea lamprey. Exposure to 10�11 M 3kACA decreased GAP49 (8 h,p < 0.05) and lGnRH-III (2 h, 4 h, 8 h, 24 h, and 48 h, p < 0.05) but increased GAP50 (48 h, p < 0.05) mRNA concentrations in the forebrain, and increased GAP49 mRNAconcentrations (24 h, p < 0.05) in the brain stem of immature male sea lamprey. On the other hand, exposure to 10�10 M 3kACA decreased lGnRH-III (48 h, p < 0.05) and JNK(48 h, p < 0.05) mRNA concentrations in the forebrain, but increased GAP49, GAP50, and GAP58 mRNA concentrations (all at 8 h, p < 0.05) in the brain stem of immature malesea lamprey. Exposure to 10�9 M 3kACA only decreased lGnRH-III mRNA concentrations (2 h, 4 h, 24 h, and 48 h, p < 0.05) in the forebrain of immature male sea lamprey. �indicates significant difference between treatment group and vehicle control (p < 0.05).
Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31 27
3.4. Phasic pattern of plasma lGnRH peptide concentrations in malesea lamprey
Male sea lamprey showed phasic pattern in plasma lGnRH pep-tide concentrations although the source of lGnRH release was notclear (Fig. 4). Isolated males showed a more regular pattern thangroup males (Fig. 4). On average, group males contained higherplasma concentrations of all three lGnRH peptides than isolatedmales (Fig. 5).
4. Discussion
We found that waterborne 3kACA decreased plasma 15a-P con-centrations in immature male sea lamprey. This is the first knowmale pheromone component that inhibits steroidogenesis inimmature male conspecifics. Odors from conspecifics of the samesex tend to diminish reproductive activities in most species (Cham-plin, 1971; Drickamer, 1977; Holman et al., 2010; Ma et al., 1998;McClintock, 1983). Stimulatory primer pheromones stimulate gon-adotropin release and facilitate sexual maturation and reproduc-tion (Murata et al., 2009; Okamura et al., 2010; Wysocki andPreti, 2004). These effects can be complicated by inhibitory primerpheromones released at the same time. In goldfish, the responsive-ness of the males to the stimulatory primer pheromone 17a,
20b-dihydroxy-4-pregnen-3-one (17, 20b-P) is attenuated by aninhibitory primer pheromone androstenedione released by females(Stacey, 2003). Waterborne androstenedione inhibits the endo-crine-gonadal response to 17, 20b-P in male conspecifics, but nei-ther the mechanism nor the biological function of this effect isknown (Stacey et al., 1991).
Pulsatile GnRH release in higher vertebrates regulates the gon-adotropin subunit genes via the activation of GnRH receptor and itssignaling cascades including JNK and its substrate JUN (Burgeret al., 2009; Haisenleder et al., 2008; Harris et al., 2002; Yokoiet al., 2000). We found that 3kACA increased lGnRH-III peptideconcentration but decreased lGnRH-III gene expression in the fore-brain after 2 h exposure while Jun and JNK gene expressions de-creased after 24 h exposure. These results indicate that 3kACAcan modulate GnRH release and its signaling cascades. The phasicnature of GnRH release rendered the pheromone exposure resultshard to interpret. However, the variance of the data collectedshould reflect the amplitude of the phasic release and the resultsfrom the analyses of ANOVA and F-tests should still be valid. Pher-omones are known to induce GnRH pulses in higher vertebrates(Murata et al., 2011; Okamura et al., 2010). It is likely that sea lam-prey sex pheromone 3kACA also affect GnRH pulses in sea lamprey.
The detection of lGnRH peptides in the plasma was a surprisesince many previous attempts using HPLC and radioimmunoassays
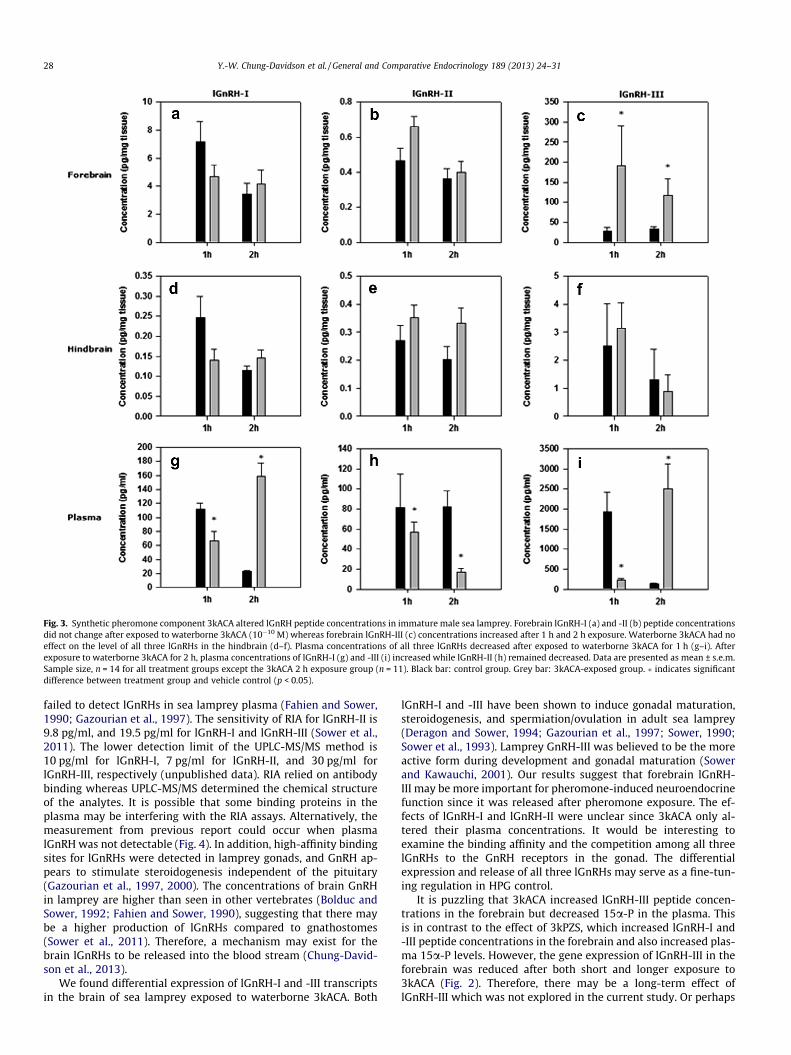
Fig. 3. Synthetic pheromone component 3kACA altered lGnRH peptide concentrations in immature male sea lamprey. Forebrain lGnRH-I (a) and -II (b) peptide concentrationsdid not change after exposed to waterborne 3kACA (10�10 M) whereas forebrain lGnRH-III (c) concentrations increased after 1 h and 2 h exposure. Waterborne 3kACA had noeffect on the level of all three lGnRHs in the hindbrain (d–f). Plasma concentrations of all three lGnRHs decreased after exposed to waterborne 3kACA for 1 h (g–i). Afterexposure to waterborne 3kACA for 2 h, plasma concentrations of lGnRH-I (g) and -III (i) increased while lGnRH-II (h) remained decreased. Data are presented as mean ± s.e.m.Sample size, n = 14 for all treatment groups except the 3kACA 2 h exposure group (n = 11). Black bar: control group. Grey bar: 3kACA-exposed group. � indicates significantdifference between treatment group and vehicle control (p < 0.05).
28 Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31
failed to detect lGnRHs in sea lamprey plasma (Fahien and Sower,1990; Gazourian et al., 1997). The sensitivity of RIA for lGnRH-II is9.8 pg/ml, and 19.5 pg/ml for lGnRH-I and lGnRH-III (Sower et al.,2011). The lower detection limit of the UPLC-MS/MS method is10 pg/ml for lGnRH-I, 7 pg/ml for lGnRH-II, and 30 pg/ml forlGnRH-III, respectively (unpublished data). RIA relied on antibodybinding whereas UPLC-MS/MS determined the chemical structureof the analytes. It is possible that some binding proteins in theplasma may be interfering with the RIA assays. Alternatively, themeasurement from previous report could occur when plasmalGnRH was not detectable (Fig. 4). In addition, high-affinity bindingsites for lGnRHs were detected in lamprey gonads, and GnRH ap-pears to stimulate steroidogenesis independent of the pituitary(Gazourian et al., 1997, 2000). The concentrations of brain GnRHin lamprey are higher than seen in other vertebrates (Bolduc andSower, 1992; Fahien and Sower, 1990), suggesting that there maybe a higher production of lGnRHs compared to gnathostomes(Sower et al., 2011). Therefore, a mechanism may exist for thebrain lGnRHs to be released into the blood stream (Chung-David-son et al., 2013).
We found differential expression of lGnRH-I and -III transcriptsin the brain of sea lamprey exposed to waterborne 3kACA. Both
lGnRH-I and -III have been shown to induce gonadal maturation,steroidogenesis, and spermiation/ovulation in adult sea lamprey(Deragon and Sower, 1994; Gazourian et al., 1997; Sower, 1990;Sower et al., 1993). Lamprey GnRH-III was believed to be the moreactive form during development and gonadal maturation (Sowerand Kawauchi, 2001). Our results suggest that forebrain lGnRH-III may be more important for pheromone-induced neuroendocrinefunction since it was released after pheromone exposure. The ef-fects of lGnRH-I and lGnRH-II were unclear since 3kACA only al-tered their plasma concentrations. It would be interesting toexamine the binding affinity and the competition among all threelGnRHs to the GnRH receptors in the gonad. The differentialexpression and release of all three lGnRHs may serve as a fine-tun-ing regulation in HPG control.
It is puzzling that 3kACA increased lGnRH-III peptide concen-trations in the forebrain but decreased 15a-P in the plasma. Thisis in contrast to the effect of 3kPZS, which increased lGnRH-I and-III peptide concentrations in the forebrain and also increased plas-ma 15a-P levels. However, the gene expression of lGnRH-III in theforebrain was reduced after both short and longer exposure to3kACA (Fig. 2). Therefore, there may be a long-term effect oflGnRH-III which was not explored in the current study. Or perhaps
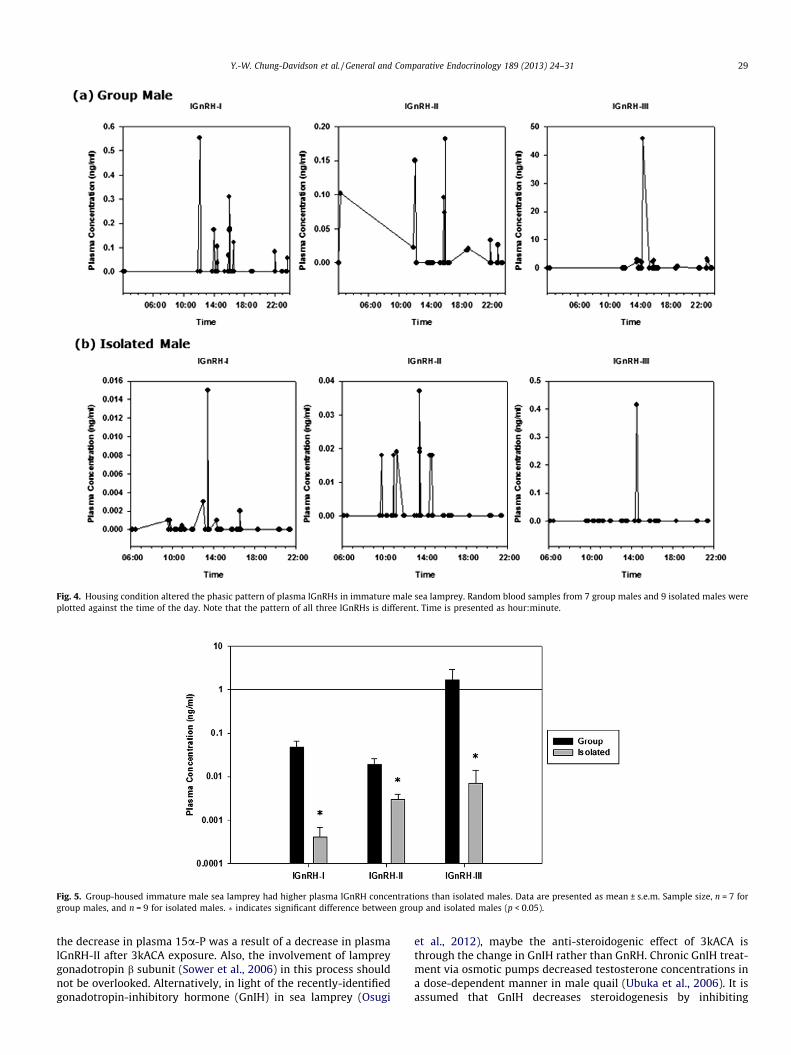
Fig. 4. Housing condition altered the phasic pattern of plasma lGnRHs in immature male sea lamprey. Random blood samples from 7 group males and 9 isolated males wereplotted against the time of the day. Note that the pattern of all three lGnRHs is different. Time is presented as hour:minute.
Fig. 5. Group-housed immature male sea lamprey had higher plasma lGnRH concentrations than isolated males. Data are presented as mean ± s.e.m. Sample size, n = 7 forgroup males, and n = 9 for isolated males. � indicates significant difference between group and isolated males (p < 0.05).
Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31 29
the decrease in plasma 15a-P was a result of a decrease in plasmalGnRH-II after 3kACA exposure. Also, the involvement of lampreygonadotropin b subunit (Sower et al., 2006) in this process shouldnot be overlooked. Alternatively, in light of the recently-identifiedgonadotropin-inhibitory hormone (GnIH) in sea lamprey (Osugi
et al., 2012), maybe the anti-steroidogenic effect of 3kACA isthrough the change in GnIH rather than GnRH. Chronic GnIH treat-ment via osmotic pumps decreased testosterone concentrations ina dose-dependent manner in male quail (Ubuka et al., 2006). It isassumed that GnIH decreases steroidogenesis by inhibiting

30 Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31
gonadotropin synthesis and release in birds (Ubuka et al., 2006).However, our results indicated that steroidogenesis is not solelycontrolled by gonadotropin. There must be an additional mecha-nism that controls steroidogenesis independent of gonadotropinsynthesis and release. An alternative explanation may be that GnIHinhibits steroidogenesis directly in sea lamprey. In fact, GnIHreceptor mRNA was found in chicken ovaries and testes (Maddin-eni et al., 2008), which demonstrated that GnIH may exert aninhibitory effect directly in gonads. Furthermore, the human GnIHortholog RFRP-3 suppressed gonadotropin-induced progesteroneproduction in human granulosa cell culture (Oishi et al., 2012).The anti-steroidogenic effect of 3kACA maybe via GnIH effect inthe gonad, and further investigation is warranted.
Acknowledgments
We thank all summer assistants, especially David Partyka, Jo-seph Bednark, Christine N. Bedore, and Aaron Smuda, for their helpin sample collection. We thank all members of the U.S. Fish andWildlife Service Marquette Biological Station (Marquette, MI,USA) and the Department of Fisheries and Oceans Canada Sea Lam-prey Control Centre. This article is contribution number 1738 ofthe U.S. Geological Survey, Great Lakes Science Center. This studyis supported by U.S. National Science Foundation grantIOB0517491, U.S. National Institute of General Medical Sciencesgrant 5R24GM83982 and the Great Lakes Fishery Commission.
References
Binder, T.R., McLaughlin, R.L., McDonald, D.G., 2010. Relative importance of watertemperature, water level, and lunar cycle to migratory activity in spawning-phase sea lamprey in Lake Ontario. Trans. Am. Fish. Soc. 139, 700–712.
Bolduc, T.G., Sower, S.A., 1992. Changes in brain gonadotropin-releasing hormone,plasma estradiol 17-beta, and progesterone during the final reproductive cycleof the female sea lamprey, Petromyzon marinus. J. Exp. Zool. 264, 55–63.
Bryan, M.B., Scott, A.P., Cerny, I., Young, B.A., Li, W., 2004. 15 Alpha-hydroxyprogesterone in male sea lampreys, Petromyzon marinus L. Steroids69, 473–481.
Bryan, M.B., Young, B.A., Close, D.A., Semeyn, J., Robinson, T.C., Bayer, J., Li, W., 2006.Comparison of synthesis of 15 alpha-hydroxylated steroids in males of fourNorth American lamprey species. Gen. Comp. Endocrinol. 146, 149–156.
Burger, L.L., Haisenleder, D.J., Aylor, K.W., Marshall, J.C., 2009. Regulation of Lhb andEgr1 gene expression by GnRH pulses in rat pituitaries is both c-Jun N-terminalkinase (JNK)- and extracellular signal-regulated kinase (ERK)-dependent. Biol.Reprod. 81, 1206–1215.
Butler, C.G., Callow, R.K., Johnston, N.C., 1959. Extraction and purification of ‘queensubstance’ from queen bees. Nature 184, 1871.
Champlin, A.K., 1971. Suppression of oestrus in grouped mice, the effects of variousdensities and the possible nature of the stimulus. J. Reprod. Fertil. 27, 233–241.
Chung-Davidson, Y.-W., Bryan, M.B., Teeter, J., Bedore, C.N., Li, W., 2008.Neuroendocrine and behavioral responses to weak electric fields in adult sealampreys (Petromyzon marinus). Horm. Behav. 54, 34–40.
Chung-Davidson, Y.-W., Wang, H., Siefkes, M.J., Bryan, M.B., Wu, H., Johnson, N.S., Li,W., 2013. Pheromonal bile acid 3-ketopetromyzonol sulfate primes theneuroendocrine system in sea lamprey. BMC Neurosci. 14, 11.
Deragon, K.L., Sower, S.A., 1994. Effects of lamprey gonadotropin-releasinghormone-III on steroidogenesis and spermiation in male sea lampreys. Gen.Comp. Endocrinol. 95, 363–367.
Drickamer, L.C., 1977. Delay of sexual maturation in female house mice by exposureto grouped females or urine from grouped females. J. Reprod. Fertil. 51, 77–81.
Drickamer, L.C., 1983. Male acceleration of puberty in female mice (Mus musculus).J. Comp. Physiol. 97, 191–200.
Endler, A., Liebig, J., Schmitt, T., Parker, J.E., Jones, G.R., Schreier, P., Hölldobler, B.,2004. Surface hydrocarbons of queen eggs regulate worker reproduction in asocial insect. Proc. Natl. Acad. Sci. USA 101, 2945–2950.
Fahien, C.M., Sower, S.A., 1990. Relationship between brain gonadotropin-releasinghormone and final reproductive period of the adult male sea lamprey,Petromyzon marinus. Gen. Comp. Endocrinol. 80, 427–437.
Fujita, A., Watanabe, H., 2010. Inconspicuous matured males of worker form areproduced in orphaned colonies of Reticulitermes speratus (Isoptera,Rhinotermitidae) and participate in reproduction. J. Insect Physiol. 56, 1510–1515.
Gazourian, L., Deragon, K.L., Chase, C.F., Pati, D., Habibi, H.R., Sower, S.A., 1997.Characteristics of GnRH binding in the gonads and effects of lamprey GnRH-Iand -III on reproduction in the adult sea lamprey. Gen. Comp. Endocrinol. 108,327–339.
Gazourian, L., Evans, E.L., Hanson, L., Chase, C.F., Sower, S.A., 2000. The effects oflamprey GnRH-I, -III and analogs on steroidogenesis in the sea lamprey(Petromyzon marinus). Aquaculture 188, 147–165.
Haisenleder, D.J., Burger, L.L., Walsh, H.E., Stevens, J., Aylor, K.W., Shupnik, M.A.,Marshall, J.C., 2008. Pulsatile gonadotropin-releasing hormone stimulation ofgonadotropin subunit transcription in rat pituitaries: evidence for theinvolvement of Jun N-terminal kinase but not p38. Endocrinology 149, 139–145.
Hanson, L.H., Manion, P.J., 1980. Sterility method of pest-control and its potentialrole in an integrated sea lamprey (Petromyzon marinus) control program. Can. J.Fish. Aquat. Sci. 37, 2108–2117.
Harris, D., Bonfil, D., Chuderland, D., Kraus, S., Seger, R., Naor, Z., 2002. Activation ofMAPK cascades by GnRH: ERK and Jun N-terminal kinase are involved in basaland GnRH-stimulated activity of the glycoprotein hormone LHb-subunitpromoter. Endocrinology 143, 1018–1025.
Holman, L., Jørgensen, C.G., Nielsen, J., d’ettorre, P., 2010. Identification of an antqueen pheromone regulating worker sterility. Proc. R. Soc. London B 277, 3793–3800.
Johnson, N.S., Yun, S.-S., Thompson, H.T., Brant, C.O., Li, W., 2009. A syntheticpheromone induces upstream movement in female sea lamprey and summonsthem into traps. Proc. Natl. Acad. Sci. USA 106, 1021–1026.
Kavanaugh, S.I., Nozaki, M., Sower, S.A., 2008. Origins of gonadotropin-releasinghormone (GnRH) in vertebrates: identification of a novel GnRH in a basalvertebrate, the sea lamprey. Endocrinology 149, 3860–3869.
Le Conte, Y., Arnold, G., Trouiller, J., Masson, C., 1990. Identification of a broodpheromone in honeybees. Naturwissenschaften 81, 462–465.
Le Conte, Y., Mohammedi, A., Robinson, G.E., 2001. Primer effects of a broodpheromone on honeybee behavioural development. Proc. R. Soc. London B 268,163–168.
Lee, S., van der Boot, L.M., 1955. Spontaneous pseudopregnancy in mice. ActaPhysiol. Pharmacol. Neerl. 4, 442–443.
Lee, S., van der Boot, L.M., 1956. Spontaneous pseudopregnancy in mice II. ActaPhysiol. Pharmacol. Neerl. 5, 213–214.
Li, W., Scott, A.P., Siefkes, M.J., Yan, H.G., Liu, Q., Yun, S.S., Gage, D.A., 2002. Bile acidsecreted by mate sea lamprey that acts as a sex pheromone. Science 296, 138–141.
Li, K., Siefkes, M.J., Brant, C.O., Li, W., 2012. Isolation and identification ofpetromyzesterosterol, a polyhydroxysteroid from sexually mature male sealamprey (Petromyzon marinus L.). Steroids 77, 806–810.
Ma, W., Miao, Z., Novotny, M.V., 1998. Role of the adrenal gland and adrenal-mediated chemosignals in suppression of estrus in the house mouse, the Lee–Boot effect revisited. Biol. Reprod. 59, 1317–1320.
Maddineni, S.R., Oco9n-Grove, O.M., Krzysik-Walker, S.M., Hendricks III, G.L.,Ramachandran, R., 2008. Gonadotropin-inhibitory hormone (GnIH) receptorgene is expressed in the chicken ovary, potential role of GnIH in follicularmaturation. Reproduction 135, 267–274.
Maisonnasse, A., Lenoir, J.C., Costagliola, G., Baslay, D., Choteau, F., Crauser, D.,Becard, J.M., Plettner, E., Le Conte, Y., 2009. A scientific note on E-b-ocimene, anew volatile primer pheromone that inhibits worker ovary development inhoneybees. Apidologie 40, 562–564.
Matsuura, K., Himuro, C., Yokoi, T., Yamamoto, Y., Vargo, E.L., Keller, L., 2010.Identification of a pheromone regulating caste differentiation in termites. Proc.Natl. Acad. Sci. USA 107, 12963–12968.
McClintock, M.K., 1983. Pheromone regulation of the ovarian cycle, enhancement,suppression and synchrony. In: Vandenbergh, J.G. (Ed.), Pheromones andReproduction in Mammals. Academic Press, New York, pp. 113–149.
McVicar, A.J., Rankin, J.C., 1983. Renal function in unanaesthetized river lampreys(Lampetra fluviatilis L.): effects of anaesthesia, temperature and environmentalsalinity. J. Exp. Biol. 105, 351–362.
Mohammedi, A., Paris, A., Crauser, D., Le Conte, Y., 1998. Effect of aliphatic esters onovary development of queenless bees (Apis mellifera L.). Naturwissenschaften85, 455–458.
Murata, K., Wakabayashi, Y., Kitago, M., Ohara, H., Watanabe, H., Tamogami, S.,Warita, Y., Yamagishi, K., Ichikawa, M., Takeuchi, Y., Okamura, H., Mori, Y., 2009.Modulation of gonadotrophin-releasing hormone pulse generator activity bythe pheromone in small ruminants. J. Neuroendocrinol. 21, 346–350.
Murata, K., Wakabayashi, Y., Sakamoto, K., Tanaka, T., Takeuchi, Y., Mori, Y.,Okamura, H., 2011. Effect of brief exposure of male pheromone on multiple-unitactivity at close proximity to kisspeptin neurons in the goat arcuate nucleus. J.Reprod. Dev. 57, 197–202.
Oishi, H., Klausen, C., Bentley, G.E., Osugi, T., Tsutsui, K., Gilks, C.B., Yano, T., Leung,P.C.K., 2012. The human gonadotropin-inhibitory hormone ortholog RFamide-related peptide-3 suppresses gonadotropin-induced progesterone production inhuman granulosa cells. Endocrinology 153, 3435–3445.
Okamura, H., Murata, K., Sakamoto, K., Wakabayashi, Y., Ohkura, S., Takeuchi, Y.,Mori, Y., 2010. Male effect pheromone tickles the gonadotrophin-releasinghormone pulse generator. J. Neuroendocrinol. 22, 825–832.
Osugi, T., Daukss, D., Gazda, K., Ubuka, T., Kosugi, T., Nozaki, M., Sower, S.A., Tsutsui,K., 2012. Evolutionary origin of the structure and function of gonadotropin-inhibitory hormone, insights from lampreys. Endocrinology 153, 2362–2374.
Reyes, R., Mendoza, J., Ballesteros, J., Moffatt, C., 2004. Male chemosignals inhibitthe neural responses of male mice to female chemosignals. Brain Res. Bull. 63,301–308.
Siefkes, M.J., Winterstein, S.R., Li, W., 2005. Evidence that 3-keto petromyzonolsulphate specifically attracts ovulating female sea lamprey, Petromyzonmarinus. Anim. Behav. 70, 1037–1045.

Y.-W. Chung-Davidson et al. / General and Comparative Endocrinology 189 (2013) 24–31 31
Slessor, K.N., Winston, M.L., Le Conte, Y., 2005. Pheromone communication in thehoneybee (Apis mellifera L.). J. Chem. Ecol. 31, 2731–2745.
Sower, S.A., 1990. Neuroendocrine control of reproduction in lampreys. Fish Physiol.Biochem. 8, 365–374.
Sower, S.A., Kawauchi, H., 2001. Update, brain and pituitary hormones of lampreys.Comp. Biochem. Physiol. 129, 291–302.
Sower, S.A., Chiang, Y.C., Lovas, S., Conlon, J.M., 1993. Primary structure andbiological activity of a third gonadotropin-releasing hormone from lampreybrain. Endocrinology 132, 1125–1131.
Sower, S.A., Moriyama, S., Kasaraha, M., Takahashi, A., Nozaki, M., Uchida, K.,Dahlstrom, J.M., Kawauchi, H., 2006. Identification of sea lamprey GTHb-likecDNA and its evolutionary implications. Gen. Comp. Endocrinol. 148, 22–32.
Sower, S.A., Balz, E., Aquilina-Beck, A., Kavanaugh, S.I., 2011. Seasonal changes ofbrain GnRH-I, -II, and -III during the final reproductive period in adult male andfemale sea lamprey. Gen. Comp. Endocrinol. 170, 276–282.
Stacey, N.E., 2003. Hormones, pheromones and reproductive behavior. Fish Physiol.Biochem. 28, 229–235.
Stacey, N.E., Sorensen, P.W., Dulka, J.G., Cardwell, J.R., Irvine, A.S., 1991. Fish sexpheromones, current status and potential applications. Bull. Inst. Zool. Acad.Sinica Monogr. 16, 189–227.
Suzuki, K., Gamble, R.L., Sower, S.A., 2000. Multiple transcripts encoding lampreygonadotropin-releasing hormone-I precursors. J. Mol. Endocrinol. 24, 365–376.
Trhlin, M., Rajchard, J., 2011. Chemical communication in the honeybee (Apismellifera L.), a review. Vet. Med. 56, 265–273.
Ubuka, T., Ukena, K., Sharp, P.J., Bentley, G.E., Tsutsui, K., 2006. Gonadotropin-inhibitory hormone inhibits gonadal development and maintenance bydecreasing gonadotropin synthesis and release in male quail. Endocrinology147, 1187–1194.
Vandenbergh, J.G., Drickamer, L.C., Colby, D.R., 1972. Social and dietary factors inthe sexual maturation of female mice. J. Reprod. Fertil. 28, 397–405.
Wanner, K.W., Nichols, A.S., Walden, K.K.O., Brockmann, A., Luetje, C.W., Robertson,H.M., 2007. A honeybee odorant receptor for the queen substance 9-oxo-2-decenoic acid. Proc. Natl. Acad. Sci. USA 104, 14383–14388.
Wolff, P.R., Powell, A.J., 1979. Urination patterns and estrous cycling in mice. Behav.Neural Biol. 27, 379–383.
Wysocki, C.J., Preti, G., 2004. Facts, fallacies, fears, and frustrations with humanpheromones. Anat. Rec. Part A 281A, 1201–1211.
Yokoi, T., Ohmichi, M., Tasaka, K., Kimura, A., Kanda, Y., Hayakawa, J., Tahara, M.,Hisamoto, K., Kurachi, H., Murata, Y., 2000. Activation of the luteinizinghormone b promoter by gonadotropin-releasing hormone requires c-Jun NH2-terminal protein kinase. J. Biol. Chem. 275, 21639–21647.
Yun, S.S., Scott, A.P., Li, W., 2003. Pheromones of the male sea lamprey, Petromyzonmarinus L., structural studies on a new compound, 3-keto allocholic acid, and 3-keto petromyzonol sulfate. Steroids 68, 297–304.
Related Documents











