Amplification of Telomerase Reverse Transcriptase Gene in Human Mammary Epithelial Cells with Limiting Telomerase RNA Expression Levels Ying Cao, Lily I. Huschtscha, Amanda S. Nouwens, Hilda A. Pickett, Axel A. Neumann, Andy C-M. Chang, Christian D. Toouli, Tracy M. Bryan, and Roger R. Reddel Children’s Medical Research Institute, Westmead, and University of Sydney, Sydney, New South Wales, Australia Abstract Activation of telomerase is a crucial step during cellular immortalization, and in some tumors this results from amplification of the human telomerase reverse transcriptase (hTERT ) gene. Immortalization of normal human cells has been achieved by transduction with hTERT cDNA under the control of a strong heterologous enhancer/promoter, but this is sometimes an inefficient process, with periods of poor growth or even crisis occurring before immortalization. Here, we showed that normal human mammary epithelial cells expressing exogenous hTERT amplified the transgene exten- sively and expressed high levels of hTERT mRNA and protein. Paradoxically, the cells had low levels of telomerase activity and very short telomeres, indicating that telomerase activity did not correlate with hTERT expression. These cells con- tained only f20 human telomerase RNA (hTR) molecules/cell (compared with f120 hTR molecules per 293 cell). Expression of exogenous hTR caused increased telomerase activity and telomere lengthening. These data indicate that some hTERT- transduced normal cells may express high levels of the transgene but fail to up-regulate endogenous hTR expression sufficiently to enable expression of robust levels of telomerase activity. [Cancer Res 2008;68(9):3115–23] Introduction Human telomeres are nucleoprotein complexes located at the ends of linear chromosomes (1). They shorten at each cell division, which limits the number of times a cell can divide (2). Telomerase is a ribonucleoprotein enzyme complex that adds telomeric repeat sequences to the ends of chromosomes to counteract this shortening (3). It has recently been found that the active human telomerase enzyme is composed of human telomerase reverse transcriptase (hTERT), human telomerase RNA (hTR), and dyskerin (4): hTERT is the catalytic reverse transcriptase component (5), hTR serves as the RNA template for the addition of telomeric repeats (6), and dyskerin is an RNA-binding protein. Mutations in any of these components may result in dyskeratosis congenita, a human disease syndrome associated with short telomeres (reviewed by Kirwan et al. in ref. 7). Telomerase is not detectable in most normal human somatic cells, whereas it is expressed in >85% of human tumors (8). How telomerase levels are regulated in these tumors is still poorly understood. A major focus of investigation has been the control of hTERT transcription (reviewed by Horikawa et al. in ref. 9). Amplification of the hTERT gene has been observed in various human cancer cell lines and tumors, and is also thought to be a mechanism of telomerase activation (10–13). In view of the observations that hTR is ubiquitously expressed (6, 14), whereas hTERT is expressed only in telomerase-positive cells (5), and that expression of exogenous hTERT alone can immortalize normal human cells (15, 16), abundance of hTERT was previously thought to be the sole limiting factor for telomerase activity. Although immortalization of many types of normal human cells has been achieved by transducing them with exogenous hTERT, for some strains of normal human cells, this is an ineffi- cient process, with periods of poor growth or even crisis being experienced before immortalization (17–19). Data are accumulat- ing in support of the notion that hTR levels can also be limiting for telomerase activity. For example, hTR mutations have been found to cause autosomal dominant dyskeratosis congenita (20) and can reduce telomerase activity via haploinsufficiency (21). Furthermore, overexpression of both hTERT and hTR elongates telomeres better than overexpression of hTERT alone in X-linked and autosomal dominant dyskeratosis congenita (22, 23). It has also been shown that concomitant overexpression of hTERT and hTR was necessary to substantially increase telomerase activity in both cancer and primary cells (24). However, none of these studies quantified the number of hTR molecules in the cells. This is important because a previous study has reported that there are 11,000 to 13,500 hTR molecules per cell in the telomerase-negative BJ, IMR90, and SW13 cell strains (25), which would be inconsistent with a limiting role for hTR in light of the 20 to 50 active telomerase molecules per cell in 293 cells (4). In this study, we explore mechanisms of telomerase activity regulation. Human mammary epithelial cells (HMEC) have been transduced with hTERT in our laboratory, and four independent hTERT-immortalized mass cultures (B80-TERT1, 2, 3a, and 3b) were obtained (26). In the present study, we found that the B80- TERT1 cell line had undergone extensive amplification of the hTERT transgene and was expressing high levels of hTERT mRNA and protein but had low telomerase activity. Furthermore, we showed that hTR levels limit telomerase activity and telomere length in this cell line. More importantly, our quantitation of hTR molecules per cell provides an explanation for why hTR is limiting: the low abundance of hTR (f20 molecules of hTR per cell) is Note: Supplementary data for this article are available at Cancer Research Online (http://cancerres.aacrjournals.org/). Current address for A.S. Nouwens: School of Molecular and Microbial Sciences, and Australian Institute of Biotechnology and Nanotechnology, The University of Queensland, St Lucia, Queensland 4072, Australia; Current address for C.D. Toouli, Bio-Link Partners Ltd., Suite 7/G11, Locomotive Workshop, Australian Technology Park, Eveleigh, New South Wales 1430, Australia. Requests for reprints: Roger R. Reddel, Children’s Medical Research Institute, 214 Hawkesbury Road, Westmead, Sydney, New South Wales 2145 Australia. Phone: 61-2- 9687-2800; Fax: 61-2-9687-2120; E-mail: [email protected]. I2008 American Association for Cancer Research. doi:10.1158/0008-5472.CAN-07-6377 www.aacrjournals.org 3115 Cancer Res 2008; 68: (9). May 1, 2008 Research Article Research. on June 5, 2015. © 2008 American Association for Cancer cancerres.aacrjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Amplification of Telomerase Reverse Transcriptase Gene in
Human Mammary Epithelial Cells with Limiting
Telomerase RNA Expression Levels
Ying Cao, Lily I. Huschtscha, Amanda S. Nouwens, Hilda A. Pickett, Axel A. Neumann,Andy C-M. Chang, Christian D. Toouli, Tracy M. Bryan, and Roger R. Reddel
Children’s Medical Research Institute, Westmead, and University of Sydney, Sydney, New South Wales, Australia
Abstract
Activation of telomerase is a crucial step during cellularimmortalization, and in some tumors this results fromamplification of the human telomerase reverse transcriptase(hTERT) gene. Immortalization of normal human cells hasbeen achieved by transduction with hTERT cDNA under thecontrol of a strong heterologous enhancer/promoter, but thisis sometimes an inefficient process, with periods of poorgrowth or even crisis occurring before immortalization. Here,we showed that normal human mammary epithelial cellsexpressing exogenous hTERT amplified the transgene exten-sively and expressed high levels of hTERT mRNA and protein.Paradoxically, the cells had low levels of telomerase activityand very short telomeres, indicating that telomerase activitydid not correlate with hTERT expression. These cells con-tained only f20 human telomerase RNA (hTR) molecules/cell(compared with f120 hTR molecules per 293 cell). Expressionof exogenous hTR caused increased telomerase activity andtelomere lengthening. These data indicate that some hTERT-transduced normal cells may express high levels of thetransgene but fail to up-regulate endogenous hTR expressionsufficiently to enable expression of robust levels of telomeraseactivity. [Cancer Res 2008;68(9):3115–23]
Introduction
Human telomeres are nucleoprotein complexes located at theends of linear chromosomes (1). They shorten at each cell division,which limits the number of times a cell can divide (2). Telomeraseis a ribonucleoprotein enzyme complex that adds telomeric repeatsequences to the ends of chromosomes to counteract thisshortening (3). It has recently been found that the active humantelomerase enzyme is composed of human telomerase reversetranscriptase (hTERT), human telomerase RNA (hTR), and dyskerin(4): hTERT is the catalytic reverse transcriptase component (5),hTR serves as the RNA template for the addition of telomericrepeats (6), and dyskerin is an RNA-binding protein. Mutations in
any of these components may result in dyskeratosis congenita, ahuman disease syndrome associated with short telomeres(reviewed by Kirwan et al. in ref. 7).Telomerase is not detectable in most normal human somatic
cells, whereas it is expressed in >85% of human tumors (8). Howtelomerase levels are regulated in these tumors is still poorlyunderstood. A major focus of investigation has been the controlof hTERT transcription (reviewed by Horikawa et al. in ref. 9).Amplification of the hTERT gene has been observed in varioushuman cancer cell lines and tumors, and is also thought to be amechanism of telomerase activation (10–13).In view of the observations that hTR is ubiquitously expressed
(6, 14), whereas hTERT is expressed only in telomerase-positivecells (5), and that expression of exogenous hTERT alone canimmortalize normal human cells (15, 16), abundance of hTERT waspreviously thought to be the sole limiting factor for telomeraseactivity. Although immortalization of many types of normal humancells has been achieved by transducing them with exogenoushTERT, for some strains of normal human cells, this is an ineffi-cient process, with periods of poor growth or even crisis beingexperienced before immortalization (17–19). Data are accumulat-ing in support of the notion that hTR levels can also be limiting fortelomerase activity. For example, hTR mutations have been foundto cause autosomal dominant dyskeratosis congenita (20) and canreduce telomerase activity via haploinsufficiency (21). Furthermore,overexpression of both hTERT and hTR elongates telomeres betterthan overexpression of hTERT alone in X-linked and autosomaldominant dyskeratosis congenita (22, 23). It has also been shownthat concomitant overexpression of hTERT and hTR was necessaryto substantially increase telomerase activity in both cancer andprimary cells (24). However, none of these studies quantified thenumber of hTR molecules in the cells. This is important because aprevious study has reported that there are 11,000 to 13,500 hTRmolecules per cell in the telomerase-negative BJ, IMR90, and SW13cell strains (25), which would be inconsistent with a limiting rolefor hTR in light of the 20 to 50 active telomerase molecules per cellin 293 cells (4).In this study, we explore mechanisms of telomerase activity
regulation. Human mammary epithelial cells (HMEC) have beentransduced with hTERT in our laboratory, and four independenthTERT-immortalized mass cultures (B80-TERT1, 2, 3a, and 3b)were obtained (26). In the present study, we found that the B80-TERT1 cell line had undergone extensive amplification of thehTERT transgene and was expressing high levels of hTERT mRNAand protein but had low telomerase activity. Furthermore, weshowed that hTR levels limit telomerase activity and telomerelength in this cell line. More importantly, our quantitation of hTRmolecules per cell provides an explanation for why hTR is limiting:the low abundance of hTR (f20 molecules of hTR per cell) is
Note: Supplementary data for this article are available at Cancer Research Online(http://cancerres.aacrjournals.org/).
Current address for A.S. Nouwens: School of Molecular and Microbial Sciences, andAustralian Institute of Biotechnology and Nanotechnology, The University ofQueensland, St Lucia, Queensland 4072, Australia; Current address for C.D. Toouli,Bio-Link Partners Ltd., Suite 7/G11, Locomotive Workshop, Australian TechnologyPark, Eveleigh, New South Wales 1430, Australia.
Requests for reprints: Roger R. Reddel, Children’s Medical Research Institute, 214Hawkesbury Road, Westmead, Sydney, New South Wales 2145 Australia. Phone: 61-2-9687-2800; Fax: 61-2-9687-2120; E-mail: [email protected].
I2008 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-07-6377
www.aacrjournals.org 3115 Cancer Res 2008; 68: (9). May 1, 2008
Research Article
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

insufficient for significant telomerase activity and telomeremaintenance even in the presence of a vast excess of hTERT. Toour knowledge, our study is the first to report the low abundanceof hTR in telomerase-negative cells (approximately three hTRmolecules in each telomerase-negative B80 cell), and that hTRlimits telomerase activity and telomere maintenance in the contextof hTERT amplification.
Materials and Methods
Cells and cell culture. HMECs were cultured in MCDB 170 medium
(Invitrogen) as described previously (26). PA317 packaging cells were grownin Dulbecco’s Modified Eagles Medium with 10% fetal bovine serum in a 5%
CO2 incubator at 37jC.Fluorescence in situ hybridization analysis. Chromosome prepara-
tions from colcemid-arrested cells were obtained according to standardcytogenetic methods (27). A 3.4-kb full-length hTERT cDNA excised from
pCIneo-hTERT plasmid (28) or a 650-bp NH2-terminal hTERT fragment
excised from pJJ101-hTERT1-182aa was labeled with bio-16-dUTP using the
Biotin-Nick Translation Mix (Roche) according to the manufacturer’sinstructions. Approximately 30 ng/AL of the full-length hTERT or the 650-bp
NH2-terminal hTERT fragment was hybridized onto separately denatured
chromosome preparations for 16 to 18 h in a humidified chamber at 37jC.The hybridized probe was detected with Fluorescein Avidin DCS (VectorLaboratories) followed by signal amplification with biotinylated anti-avidin
antibody (Vector Laboratories) and another layer of Fluorescein Avidin DCS.
Chromosomes were counterstained with diamidino-phenyl-indole-dihydro-chloride (DAPI; final concentration, 0.6 Ag/mL; Sigma-Aldrich) for
chromosome identification, and slides were evaluated on a Leica DMLB
fluorescence microscope with appropriate filter sets. Fluorescein and DAPI
images were captured separately with a cooled charge-coupled devicecamera (SPOT2; Diagnostic Instruments), merged using SPOT2 software,
and further processed using Adobe Photoshop version 6.0 software.
Southern blot analysis. Genomic DNA was digested with EcoR I and
Sal I, separated through a 0.8% agarose gel with Tris-borate-EDTA (TBE)buffer at pH 7.5, and transferred onto Hybond N+ membrane (Amersham
Biosciences) by capillary action in 0.4 mol/L NaOH for 4 h. The membrane
was blocked in Church buffer [1% bovine serum albumin, 1 mmol/L EDTA,
0.5 mol/L NaPO4 (pH 7.2), and 7% SDS] at 65jC for 1 h, followed byhybridization (in Church buffer at 65jC overnight) with the 3.4-kb full-
length hTERT cDNA excised from pCIneo-hTERT plasmid and radiolabeled
with a-32P-dCTP. The membrane was washed twice in 2�SSC/0.1%SDS at65jC for 20 min, followed by another two washes in 0.2�SSC/0.1%SDS
at 65jC for 20 min. Washed membrane was exposed to Kodak BioMax MS
X-ray film (Sigma-Aldrich).
Reverse transcriptase-PCR. Total RNA was isolated from cell pellets byusing TRI-zol reagent (Invitrogen). One microgram of total RNA was reverse
transcribed by using Moloney murine leukemia virus reverse transcriptase
(Promega). Taq polymerase (Roche) was used for the amplification. See
Supplementary Data for PCR conditions and primer sequences.Quantitative real-time reverse transcriptase-PCR. The Qiagen RNeasy
Mini kit (Qiagen) was used to extract total RNA from cells that had been
pelleted by centrifugation. Total RNA (1 Ag) was treated with DNase I(Invitrogen), followed by reverse transcription [SuperScript III First-Strand
Synthesis System for reverse transcriptase-PCR (RT-PCR); Invitrogen]
according to the manufacturer’s instructions. Real-time RT-PCR was
performed using SYBR Green PCR Master Mix (Applied Biosystems) in aCorbett RotorGene-6000 thermal cycler. Cycle parameters were as follows:
incubation at 95jC for 10 min, followed by 40 cycles of 95jC for 15 sec and
Figure 1. hTERT amplification in B80-TERT HMEC lines.A , FISH analysis of B80-TERT1 cells using the full-lengthhTERT cDNA as a probe. Top, the fluorescein-avidindetection of hTERT (green ). Middle, DAPI staining of thenucleus (blue ). Bottom , merged picture of hTERT signaland DAPI staining. B , Southern blot analysis of B80,B80-TERT1, 2, 3a, and 3b cells using the full-length hTERTcDNA as a probe. Top , detection of hTERT. Bottom ,shows the agarose gel stained with ethidium bromide toindicate DNA loading. Note that only 1 Ag of B80-TERT1DNA was loaded because of the very high hTERT genecopy number in these cells, whereas 10 Ag of DNA wasloaded for the other cell lines. Untransfected B80 cells arethe control.
Cancer Research
Cancer Res 2008; 68: (9). May 1, 2008 3116 www.aacrjournals.org
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

60jC for 1 min. Housekeeping gene glyceraldehyde-3-phosphate dehydroge-nase (GAPDH) was used as the reference gene. Primer pairs were matched
for PCR efficiencies. See Supplementary Data for primer sequences.
Northern blot analysis. Total RNA from 5 � 106 or 1 � 107 cells was
prepared using an RNeasy Mini kit (Qiagen), TRI-zol reagent (Invitrogen), orphenol/chloroform extraction from whole cell lysate to compare potential
yield differences. Total RNA extracted by any of the above methods was
precipitated, resuspended in 10 AL of water and 10 AL of formamide loading
buffer, followed by heating at 70jC for 5 min, loaded on a 1.5-mm thick4% polyacrylamide, 8M urea gel, and electrophoresed at 25 W for 1 h in
1� TBE buffer. RNA was transferred onto Hybond N+ membrane
(Amersham Biosciences) by electroblotting at 1.5A for 2 h in 0.5� TBE
buffer, with the buffer cooled to 4jC. The membrane was crosslinked with254 nm UV light (Stratalinker; Stratagene) and prehybridized in Church
buffer (see above) at 55jC for 1 h, followed by hybridization in Church
buffer at 55jC overnight with 32P 5¶-end–labeled hTR probe (5¶-CGG TGGAAG GCG GCA GGC CGA GGC-3¶). The membrane was washed three times
in 0.1�SSC/0.1%SDS at 55jC for 10 min, exposed to a phosphor screen, and
scanned with a Storm 860 optical scanner with ImageQuant software
(Molecular Dynamics).Western blot analysis. Cell pellets were lysed in freshly made radioim-
munoprecipitation assay buffer [10 mmol/L Tris-HCl (pH8.0), 140 mmol/L
NaCl, 1% Triton X-100, 0.1% SDS, 1� complete protease inhibitors (Roche),
10 mmol/L phenylmethylsulfonylfluoride, and 0.1% sodium deoxycholate].Lysates equivalent to 50 Ag of protein were separated on an 8%
polyacrylamide/SDS gel and transferred to Hybond enhanced chemilumi-
nescence (ECL) nitrocellulose (Amersham Biosciences) by electroblotting.The membrane was blocked with 1� PBS containing 2% skim milk and
0.05% Tween 20 at room temperature for 1 h. The membrane was then
incubated with the primary antibody (goat polyclonal anti-hTERT; sc-7215;
1:1,000 dilution; Santa Cruz) in 1�PBS containing 1% skim milk and0.05% Tween 20 at 4jC overnight. The membrane was washed in
1�PBS containing 0.05% Tween 20, followed by the incubation with the
secondary antibody (rabbit anti-goat immunoglobulin horseradish perox-
idase; 1:10,000 dilution; DAKO Cytomation) in 1�PBS containing 1% skimmilk and 0.05% Tween 20 at room temperature for 1 h. Amersham ECL plus
Western blotting detection system (Amersham Biosciences) and high
performance chemiluminescence film (Amersham Biosciences) were usedfor detection.
Direct telomerase activity assay. A direct telomerase activity assay
recently developed in our laboratory (29) was used. Cells (1 � 107) were
lysed and the lysate was incubated with purified anti-hTERT antibody (4).Protein G/Agarose beads were added and the immunoprecipitate was
collected into a microspin column. The beads were washed, and antigenic
peptide (4) was mixed with the immunoprecipitate to elute telomerase. The
activity of immunopurified telomerase was analyzed as described inSupplementary Data.
Production of retroviruses and infections. hTR construct pBABE-
puro-U3-hTR (kindly provided by Dr. Kathleen Collins, Department of
Molecular and Cell Biology, University of California, Berkeley, CA) andvector alone (pBABE-puro) were transfected into PA317 packaging cells
separately using siPORT XP-1 transfection reagent (Ambion). After 48 h,
medium was harvested and filtered through a 0.45-Am membrane, andpolybrene (Sigma-Aldrich) was added to a final concentration of 4 Ag/mL.
B80-TERT1 cells were grown to 50% confluence and then incubated with
retroviral supernatant at 37jC for 8 h, followed by incubation with fresh
medium overnight. Forty-eight hours after infection, cells were passagedand grown in selection medium containing 0.6 Ag/mL puromycin (Sigma-
Aldrich). Clones were obtained and maintained in medium containing
0.6 Ag/mL puromycin.
Terminal restriction fragment analysis. Hin f I- and Rsa I-digestedgenomic DNA (1.5 Ag) was fractionated on a pulsed-field electrophoresis
apparatus (Bio-Rad) as previously described (30). The gel was dried,
denatured, and hybridized to a [g-32P] dATP 5¶-end–labeled telomeric oligo-nucleotide probe, (TTAGGG)3. The gel was exposed to a phosphor screen
and scanned with a Storm 860 optical scanner with ImageQuant software.
Median telomere length was obtained with Telometric software (31).
See Supplementary Data for Preparation of hTR standards and in vitrohTERT translation.
Results
Amplification of exogenous hTERT in HMECs. Four indepen-dent hTERT-immortalized mass cultures B80-TERT1, 2, 3a, and 3bwere previously obtained in our laboratory by transfecting B80HMECs with pCIneo-hTERT (26). In the present study, we detectedextensive amplification of hTERT DNA in B80-TERT1 cells byfluorescence in situ hybridization (FISH) analysis of metaphase
Figure 2. hTERT mRNA and protein expression correlates with hTERT genecopy number. hTERT mRNA expression was detected by RT-PCR (A) andquantitative real-time RT-PCR (B ; using GAPDH as the reference gene). Errorbars, ranges for duplicate PCR reactions. hTERT protein expression wasdetected by Western blot using hTERT antibody sc7215 (C ). In vitro translatedhTERT was used as a positive control in the hTERT Western; it migrates alittle more slowly in the gel due to the presence of NH2-terminal tags on theprotein. hTERT was not detectable in untransfected B80 cells by either RT-PCRor Western blot. B80 refers to untransfected B80, and 1, 2, 3a, and 3b refer toB80-TERT1, B80-TERT2, B80-TERT3a, and B80-TERT3b cells, respectively.
hTERT Amplification in Cells with Limiting hTR Levels
www.aacrjournals.org 3117 Cancer Res 2008; 68: (9). May 1, 2008
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
spreads using both a 3.4-kb full-length hTERT cDNA (Fig. 1A) and a650-bp NH2-terminal hTERT probe (data not shown). Amplificationof hTERT was observed in every metaphase (Fig. 1A) andinterphase nucleus (Supplementary Fig. S1), which indicates thatthis immortalized cell line has become clonal, although it wasoriginally established as a mass culture. A similar pattern ofhybridization in B80-TERT1 cells was also observed by FISHanalysis when pCIneo vector alone was used as a probe (data notshown). These observations indicate that the hTERT transgene andits accompanying vector sequence were extensively amplified inB80-TERT1 cells.Furthermore, hTERT amplification was also detected in B80-
TERT3b cells by FISH analysis (Supplementary Fig. S2), althoughto a much lesser extent than in B80-TERT1 cells. No hTERT ampli-fication was detectable by FISH in B80-TERT2 or 3a cells (data notshown).To confirm the hTERT amplification seen by FISH in B80-TERT1
and B80-TERT3b cells, Southern blot analysis was performed usingthe full-length hTERT cDNA as a probe. A very high hTERT copynumber was detected in only 1 Ag of B80-TERT1 genomic DNA,whereas only a small amount of hTERT DNA was detected in 10 Agof genomic DNA from B80-TERT2 and B80-TERT3a. An interme-diate TERT copy number was observed in B80-TERT3b (Fig. 1B).The genomic DNA was digested with EcoR I and Sal I restrictionenzymes, which excise the 3.4-kb hTERT cDNA from plasmidpCIneo-hTERT with which the B80-TERT lines were generated. Thepresence of a dominant 3.4-kb band (Fig. 1B, top) showed that itwas the exogenous hTERT that was amplified. This was confirmedby the observation that a high copy number of the vector backbonewas detected in B80-TERT1 cells when the pCIneo vector alone wasused as a probe in Southern blot analysis (data not shown).
Correlation between hTERT RNA/protein expression andhTERT gene copy number. Expression of hTERT mRNA wasmeasured by conventional and real-time RT-PCR. As shown inFig. 2A , the highest level of hTERT mRNA was detected in B80-TERT1 cells, less hTERT mRNA was detected in B80-TERT3b, andthe least amount of hTERT mRNA was detected in B80-TERT2 andB80-TERT3a cells. No hTERT mRNA could be detected in untrans-fected B80 cells. Similar amounts of GAPDH mRNA were detectedin all five HMEC cultures (Fig. 2A). Quantitation by real-timeRT-PCR is shown in Fig. 2B . No hTERT mRNA was detected inuntransfected B80 cells (data not shown). All hTERT immortalizedlines showed a significantly higher level of hTERTmRNA comparedwith telomerase-positive 293 cells, with the highest level of hTERTmRNA detected in B80-TERT1 (>30-fold greater than in 293 cells).To determine whether hTERT protein is being expressed and
whether the protein level correlates with the hTERT gene copynumber and hTERT mRNA level, Western blot analysis using anantibody against hTERT was performed, with in vitro translatedhTERT as a positive control (Fig. 2C). No hTERT could be detectedin untransfected B80 cells (data not shown) and the highest hTERTprotein level was found in B80-TERT1 cells. Therefore, we concludethat hTERT copy number in this panel of HMEC cell linescorrelates with expression levels of hTERT mRNA and protein.Lowest telomerase activity and lowest hTR in the highest
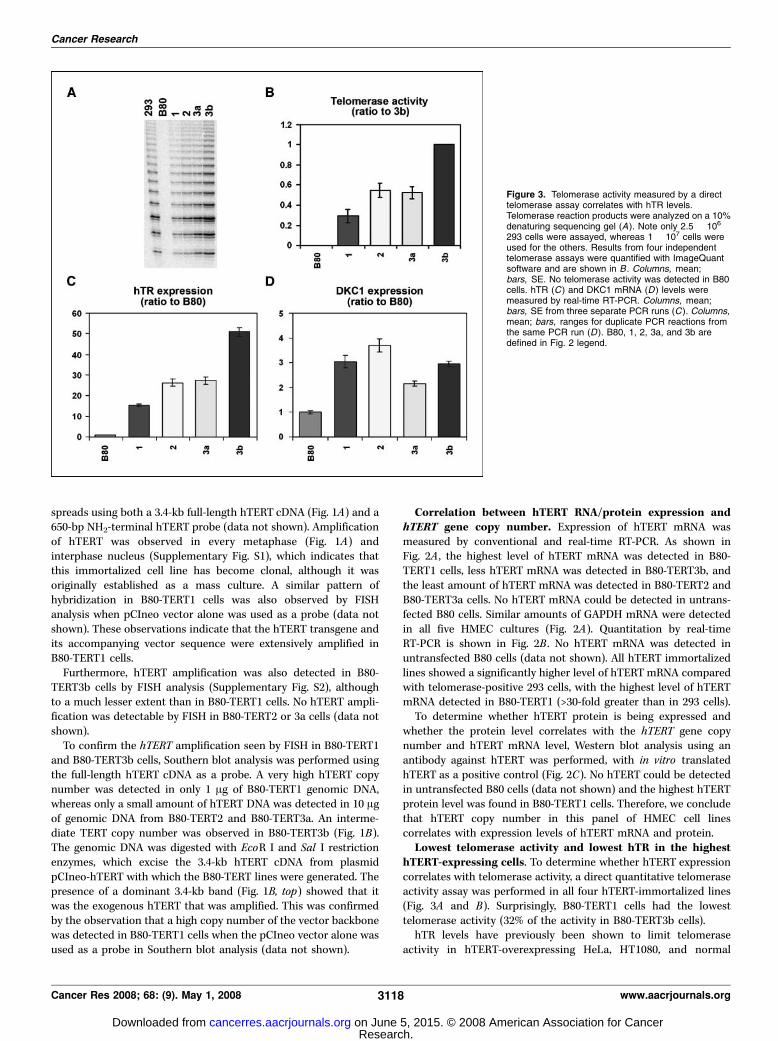
hTERT-expressing cells. To determine whether hTERT expressioncorrelates with telomerase activity, a direct quantitative telomeraseactivity assay was performed in all four hTERT-immortalized lines(Fig. 3A and B). Surprisingly, B80-TERT1 cells had the lowesttelomerase activity (32% of the activity in B80-TERT3b cells).hTR levels have previously been shown to limit telomerase
activity in hTERT-overexpressing HeLa, HT1080, and normal
Figure 3. Telomerase activity measured by a directtelomerase assay correlates with hTR levels.Telomerase reaction products were analyzed on a 10%denaturing sequencing gel (A). Note only 2.5 � 106
293 cells were assayed, whereas 1 � 107 cells wereused for the others. Results from four independenttelomerase assays were quantified with ImageQuantsoftware and are shown in B . Columns, mean;bars, SE. No telomerase activity was detected in B80cells. hTR (C ) and DKC1 mRNA (D ) levels weremeasured by real-time RT-PCR. Columns, mean;bars, SE from three separate PCR runs (C ). Columns,mean; bars, ranges for duplicate PCR reactions fromthe same PCR run (D ). B80, 1, 2, 3a, and 3b aredefined in Fig. 2 legend.
Cancer Research
Cancer Res 2008; 68: (9). May 1, 2008 3118 www.aacrjournals.org
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

human lung fibroblasts (24). To determine whether telomeraseactivity is limited by hTR level in hTERT-immortalized B80 cells,relative hTR level was examined using real-time RT-PCR. As shownin Fig. 3C , the hTERT-transduced B80 cells had increased hTRexpression (15- to 51-fold) compared with the untransfected con-trols. The lowest level of hTR was detected in B80-TERT1 (15-foldincrease), and the highest level of hTR was detected in B80-TERT3b(51-fold). Different primer sets gave similar real-time RT-PCRresults (Supplementary Fig. S3).To determine the number of hTR molecules in each cell, we
performed both Northern blot analysis and real-time RT-PCR usingserially diluted in vitro transcribed hTR as standards. Both assaysshowed excellent linearity of the standards over the range used(Fig. 4, bottom). Three Northern blot analyses with 293 cells (usingthree different methods to extract total RNA to compare potentialRNA yield difference; see Materials and Methods section) gave veryreproducible results, and the number of hTR molecules in each293 cell was 123 F 23 (mean F SE; data not shown). Northern blotanalysis showed that the number of hTR molecules per cell was 23,28, 43, and 90 in B80-TERT1, 2, 3a, and 3b, respectively (Fig. 4A).A repeat Northern gave a 2-fold lower estimate of hTR moleculesper cell (data not shown). Three separate real-time RT-PCRanalyses with duplicates in each PCR run gave a slightly higherestimate of hTR copy number as follows: 45, 76, 79, and 148molecules of hTR in B80-TERT1, 2, 3a, and 3b, respectively, andf300 molecules of hTR in each 293 cell (Fig. 4B).The f2-fold difference in hTR molecules per cell quantified by
our Northern and real-time RT-PCR could be due to the fact thatNorthern blot analysis only detects the mature 451 nucleotide hTR,whereas real-time RT-PCR would detect the mature hTR in
addition to any form of precursor, incompletely synthesized andeven partially degraded hTR, provided the target 150-bp ampliconwas intact. There are also some technical reasons that may con-tribute to the differences between the two methods. For example,the signal to noise ratio may confound quantitation by Northernmore than by real-time RT-PCR. In addition, this difference issimilar to the 3-fold difference in TLC1 (yeast telomerase RNA)copy number examined by Northern and real-time RT-PCR analysis(32). We consider that the copy number of functional hTR mole-cules is more likely to correspond to the number obtained byNorthern because this technique detects the mature full-lengthhTR. Regardless of which technique is more accurate, the dataindicate that telomerase activity correlates with hTR, rather thanwith hTERT expression levels in these hTERT-overexpressingHMECs.As dyskerin (encoded by the DKC1 gene) is part of the active
telomerase enzyme in addition to hTERTand hTR (4), we examinedDKC1 mRNA expression to determine whether DKC1 levelcorrelates with levels of hTERT, hTR, or telomerase activity. Asshown in Fig. 3D , hTERT overexpression resulted in increasedDKC1 expression from 2- to 4-fold in the four B80-TERT lines.However, no consistent correlation was found between expressionof DKC1 mRNA and either hTERT or hTR expression, or telomeraseactivity.Increased telomerase activity and telomere lengthening
resulted from hTR overexpression. We next addressed whetheroverexpression of hTR would increase telomerase activity in B80-TERT1 cells. An hTR-containing retroviral vector, pBABE-U3-hTR,was used to induce hTR expression, with pBABE vector alone as acontrol. B80-TERT1 cells were infected with the retrovirus and
Figure 4. hTR molecules per cell determined byboth quantitative Northern blot analysis andreal-time RT-PCR. A, a representative Northernblot analysis using in vitro transcribed hTR(i.v. hTR ) as standards (top ). Total RNA from5 � 106 293 cells and 1 � 107 B80-TERT1, 2, 3a,and 3b cells was loaded on the gel. RC, a recoverycontrol added before RNA precipitation(32P-labeled 100-mer DNA oligonucleotide). About40% of the RNA was recovered after RNAprecipitation (data not shown), which was takeninto account when calculating hTR molecules percell (given beneath the gel) by interpolation fromthe standard curve shown at bottom. B, hTRratio (to 293) from a representative real-timeRT-PCR analysis (top ). The standard curve(bottom ) was from the same real-time RT-PCRanalysis. hTR molecules per cell calculated fromthree independent experiments are shown in theright column (mean F SE). Each RT-PCRreaction used 1 Ag total cellular RNA; the numberof hTR molecules per cell was adjusted for theproportion of total RNA yield represented by this1 Ag. Serially diluted in vivo hTR was included tomake the standard curve (bottom ). To achievesimilar reverse transcription and PCR efficiency,in vivo hTR was reverse transcribed togetherwith 1 Ag of VA13 RNA (hTR-negative; ref. 49),followed by PCR amplification.
hTERT Amplification in Cells with Limiting hTR Levels
www.aacrjournals.org 3119 Cancer Res 2008; 68: (9). May 1, 2008
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

selected with puromycin. Individual colonies were picked andcultured for further analysis. Seven hTR-overexpressing clones(hTR15, hTR17, and hTR19-23), three pBABE vector-alone clones(BABE2, BABE6, and BABE8), and parental B80-TERT1 cells wereexamined for hTR, hTERT, and DKC1 expression using real-timeRT-PCR.In the hTR-infected cells, hTR was overexpressed from 35- to
125-fold compared with parental B80-TERT1 cells, whereas in the
vector control clones, hTR levels varied from 0.8- to 1.9-foldcompared with the parental cells (Fig. 5A). The difference in hTRlevel between hTR-infected and vector control clones was highlysignificant (P < 0.0001). hTERT expression varied from 1.2- to4.3-fold in the three vector control clones and from 1- to 6.1-fold inthe seven hTR-overexpressing clones compared with parentalB80-TERT1 cells (Fig. 5B). DKC1 RNA expression varied from 1.5- to2.7-fold in the three vector control clones and from 1.7- to 3-fold inthe seven hTR-overexpressing clones (Fig. 5C). No significantdifference was observed in either hTERT level (P = 0.68) or DKC1level (P = 0.25) between hTR-infected and vector control clones(Fig. 5).To measure telomerase activity levels, we used a direct
telomerase activity assay. Telomerase activity was increased from33- to 63-fold in hTR-overexpressing clones in comparison with1.1- to 1.6-fold in vector-alone controls (Fig. 6A). We then deter-mined whether the increased telomerase activity resulting fromhTR overexpression in B80-TERT1 cells would cause telomerelengthening. As shown in Fig. 6B , B80-TERT1 cells had very shorttelomeres (median telomere length, 5 kb), whereas B80-TERT3bcells had very long telomeres (median telomere length, 14 kb),correlating with the telomerase activities in those cells (Fig. 3A). Asexpected, all hTR-overexpressing clones had extensively elongatedtelomeres (median telomere length varied from 29–36 kb in 7hTR-overexpressing clones; Fig. 6B), compared with the vectorcontrols that exhibited short telomeres (median telomere lengthvaried from 4.5–5.2 kb in three vector control clones), similar tothe parental B80-TERT1 cells. We therefore conclude that hTRoverexpression in B80-TERT1 cells resulted in increased telomeraseactivity and consequent telomere lengthening.
Discussion
hTERT amplification has previously been observed in humancancer cell lines (10), neuroblastomas, and tumors of lung, cervix,and breast (10–13). There seems to be an inconsistent relationshipbetween amplification of the hTERT gene and the levels of hTERTmRNA, protein expression, and telomerase activity (10–13). Ourstudy has identified a reason for lack of correlation between hTERTexpression levels and telomerase activity, namely inadequate up-regulation of hTR expression.There was a very low abundance of hTR in telomerase-negative
B80 cells before hTERT overexpression, approximately three mole-cules per cell as measured by real-time RT-PCR (and undetectableby Northern blot; data not shown). Therefore, even a 15-fold up-regulation of hTR in B80-TERT1 (Fig. 3C) was associated with atelomerase activity level that resulted only in maintenance of veryshort telomeres. That hTR levels were limiting in the B80-TERT1cells was confirmed by showing that expression of exogenoushTR resulted in high levels of telomerase activity and very longtelomeres.Although until recently it has been assumed that telomerase
activity is controlled solely by the availability of hTERT, data areaccumulating in support of the notion that hTR and hTERT areboth limiting. Notably, overexpression of both hTR and hTERT inHeLa cells and fibroblasts resulted in high levels of telomeraseactivity and massive telomere elongation (24). Moreover, theimportant contribution of hTR levels to telomerase activity isindicated by the long-standing observation that hTR levels aresubstantially elevated in a wide variety of human tumors (33–37)and immortal human cell lines (5). hTR amplification has also been
Figure 5. hTR, hTERT mRNA, and DKC1 mRNA expression inhTR-overexpressing B80-TERT1 clones quantitated by real-time RT-PCR.The log of ratios to parental B80-TERT1 cells is presented on the Y-axis.Columns, mean; bars, ranges for duplicate PCR reactions. Unpaired t testwas used to compare BABE empty vector control clones (n = 3) andhTR-transduced clones (n = 7). P values are shown in each panel.
Cancer Research
Cancer Res 2008; 68: (9). May 1, 2008 3120 www.aacrjournals.org
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

observed in various tumors (38–40), and it may be one mechanismof increased hTR expression in the tumors. More recently, a viralform of telomerase RNA (vTR) with 88% homology to chickentelomerase RNA was found to be encoded by oncogenic strains ofMarek’s disease virus (MDV) but not by nononcogenic MDV strains(41). Furthermore, the wild-type vTR was shown to facilitate aviantumorigenesis (42). More importantly, the decreased hTR levelsobserved in human dyskeratosis congenita patients due either tohTR or DKC1 mutations are associated with abnormally shorttelomeres (reviewed by Kirwan et al. in ref. 7), which can berescued by overexpression of both hTERT and hTR (22, 23). Itseems that the hTR mutations associated with dyskeratosis conge-nita reduce telomerase activity via haploinsufficiency (21). Studiesin mice also suggest telomerase RNA can be limiting (43, 44).However, none of the above mentioned studies quantitated the
number of hTR molecules per cell. It has been suggested that thereare 20 to 50 active telomerase molecules per cell in 293 cells (4).B80-TERT1 cells are predicted to have many more molecules of
hTERT per cell than 20 to 50 because hTERT is readily detectableby our Western conditions in this cell line (Fig. 2C) and not in 293cells (data not shown). Thus, our quantification of hTR moleculesper cell explains the very short telomeres maintained by this verylarge number of hTERT molecules: f20 molecules of hTR areinsufficient for significant telomere elongation (Fig. 6B), even in thepresence of a vast excess of hTERT.A previous study has reported that there are 11,000 to 13,500 hTR
molecules per cell in the telomerase-negative BJ, IMR90, and SW13cell strains (25). Assuming that other normal human somatic cellshave hTR levels within an order of magnitude of the levels inHMECs, our estimate of approximately three molecules of hTR perB80 cell is more consistent than the previous study (25) with theobservation that a 2-fold change of hTR abundance (hTR haplo-insufficiency) can lead to dyskeratosis congenita (reviewed byMarrone et al. in ref. 45) because a 2-fold decrease in a vast excessof hTR would not be expected to affect telomere length. Our studyis also in agreement with the observation of a low TLC1 abundance
Figure 6. hTR overexpression in B80-TERT1results in increased telomerase activity andelongated telomeres. Telomerase assay productswere analyzed on a 10% denaturing sequencinggel (A). Note that only 2.5 � 106 293 cells wereassayed, whereas 1 � 107 cells were used for theothers. Telomere length was measured byterminal restriction fragment (TRF) analysis usingpulsed field gel electrophoresis (B ). HeLa andGM847 are examples of telomerase-positive andAlternative Lengthening of Telomeres–positivecells, respectively. The correlation betweentelomerase activity and median telomere restrictionfragment length is shown in C (the correlationvalue R was obtained using MS Excel software);points, mean values for three BABE clones andseven hTR clones; bars, SE.
hTERT Amplification in Cells with Limiting hTR Levels
www.aacrjournals.org 3121 Cancer Res 2008; 68: (9). May 1, 2008
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

in yeast (32). The reason for the discrepancy between our data andthose of Yi et al. (25) is likely to be technical rather than an intrinsicdifference between B80 cells on the one hand and telomerase-negative BJ, IMR90, and SW13 cells on the other because hTR levelswere quantitated in 293 cells in both studies and reported to be120 to 300 molecules per cell (Northern and real-time RT-PCR; thisstudy) and f69,000 molecules per cell (competitor RNA RT-PCRmethod; ref. 25). The number of RNA molecules per cell reportedhere may be an underestimate due to the yield of RNA during cellextraction being <100%, but this is unlikely to account for a 200- to600-fold difference; furthermore, three different methods of RNAextraction gave similar results (see Materials and Methods).The limiting hTR levels in the B80-TERT1 cells may have been
the selection pressure that resulted in a growth advantage for aclone of cells within the original mass culture that had acquiredamplification of hTERT. Before amplification of hTERT, it ispossible that the cellular or local concentrations of both moleculeswere below the Kd for the interaction, in which case, a higherhTERT concentration would favor the interaction between themolecules and lead to increased telomerase activity, regardless ofstoichiometry. Another possibility is that before amplification,hTERT may have been poorly expressed, as it is in B80-TERT3a(Fig. 2C), such that the number of hTERTmolecules was lower thanhTR, and this limiting hTERT expression may have selected foramplification of hTERT. Alternatively, cells with increased hTERTmay have a growth advantage due to putative telomere length-independent functions of hTERT (46, 47).Up-regulation of hTR has previously been observed in cells
expressing exogenous hTERT (22, 23, 48). The increased hTR wassuggested to be due to an increased half-life as a result of theassociation of hTR with hTERT (48). Our results indicate that thiscannot be the whole explanation because the cells with the highesthTERT protein levels had the lowest hTR levels.We found a very consistent correlation between telomere length
and telomerase activity (Fig. 6C) as measured by the directtelomerase activity assay. This assay includes a step in whichtelomerase is partially purified by immunoprecipitation from cellextracts with an hTERT antibody, which greatly increases the
sensitivity of the assay (e.g., telomerase activity was readilydetectable in 2.5 � 106 293 cells without overexpression of hTRor hTERT). In the series of cell lines generated from B80 cells, therank order according to telomerase activity was the same as thatfor telomere length, and in the hTR-transduced B80-TERT1 cells,up-regulation of telomerase activity was accompanied by acorresponding increase in telomere length. In agreement withdata from a previous study (24), this indicates that in human cells,telomerase activity level is a major determinant of telomere length.Our observation that hTERT amplification correlates with hTERT
mRNA/protein expression but not telomerase activity in hTERT-immortalized HMECs indicates that hTERT amplification andhTERT expression are not good surrogate markers for telomeraseactivity levels. Insufficient hTR up-regulation could be one of theexplanations for the previously observed nonassociation betweenhTERT amplification and telomerase activity in some tumors (10–13). Our study is the first to examine the expression of othertelomerase components, hTR and dyskerin, in the context ofhTERT-immortalized HMECs, and provides further direct evidencethat hTR is limiting for telomerase activity and telomere lengthmaintenance. Furthermore, our quantitation of hTR levels(approximately three hTR molecules per cell in telomerase-negativeB80 cells) provides an explanation for this observation, which wasat odds with the previously reported vast excess of hTR intelomerase-negative cells.
Acknowledgments
Received 11/25/2007; revised 2/7/2008; accepted 2/21/2008.Grant support: National Health and Medical Research Council (NHMRC) Peter
Doherty Postdoctoral Fellowships (228413, Y. Cao; 228412, A.S. Nouwens), NHMRCHealthy Ageing Research Grant (219306, L.I. Huschtscha and R.R. Reddel), and aWellcome Trust Senior Research Fellowship (GRO66727MA, T.M. Bryan).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Kathleen Collins for the construct pBABE-puro-U3-hTR; KarenMacKenzie and Laura Veas for the hTR2 and B2M primers; Julie Jurczyluk for theconstruct pJJ101-hTERT1-182aa, in vitro transcribed hTR, and technical assistance;Scott Cohen for the construct pUC-hTR and the direct telomerase activity assay; andJeremy Henson, Akira Nguyen, Ze Huai Zhong, Jane Noble, and Elizabeth Collins fortechnical assistance and advice.
References
1. Moyzis RK, Buckingham JM, Cram LS, et al. A highlyconserved repetitive DNA sequence, (TTAGGG)n, pres-ent at the telomeres of human chromosomes. Proc NatlAcad Sci U S A 1988;85:6622–6.
2. Harley CB, Futcher AB, Greider CW. Telomeresshorten during ageing of human fibroblasts. Nature1990;345:458–60.
3. Greider CW, Blackburn EH. Identification of a specifictelomere terminal transferase activity in Tetrahymenaextracts. Cell 1985;43:405–13.
4. Cohen SB, Graham ME, Lovrecz GO, Bache N,Robinson PJ, Reddel RR. Protein composition ofcatalytically active human telomerase from immortalcells. Science 2007;315:1850–3.
5. Nakamura TM, Morin GB, Chapman KB, et al.Telomerase catalytic subunit homologs from fissionyeast and human. Science 1997;277:955–9.
6. Feng J, Funk WD, Wang SS, et al. The RNA componentof human telomerase. Science 1995;269:1236–41.
7. Kirwan M, Dokal I. Dyskeratosis congenita: a geneticdisorder of many faces. Clin Genet 2008;73:103–12.
8. Kim NW, Piatyszek MA, Prowse KR, et al. Specificassociation of human telomerase activity with immortalcells and cancer. Science 1994;266:2011–5.
9. Horikawa I, Barrett JC. Transcriptional regulation ofthe telomerase hTERT gene as a target for cellular andviral oncogenic mechanisms. Carcinogenesis 2003;24:1167–76.
10. Zhang A, Zheng C, Lindvall C, et al. Frequentamplification of the telomerase reverse transcriptasegene in human tumors. Cancer Res 2000;60:6230–5.
11. Fan X, Wang Y, Kratz J, et al. hTERT geneamplification and increased mRNA expression in centralnervous system embryonal tumors. Am J Pathol 2003;162:1763–9.
12. Palmqvist R, Zhang A, Xu D, et al. hTERT gene copynumber is not associated with hTERT RNA expressionor telomerase activity in colorectal cancer. Int J Cancer2005;116:395–400.
13. Zhu CQ, Cutz JC, Liu N, et al. Amplification oftelomerase (hTERT) gene is a poor prognostic marker innon-small-cell lung cancer. Br J Cancer 2006;94:1452–9.
14. Avilion AA, Piatyszek MA, Gupta J, Shay JW, BacchettiS, Greider CW. Human telomerase RNA and telomeraseactivity in immortal cell lines and tumor tissues. CancerRes 1996;56:645–50.
15. Bodnar AG, Ouellette M, Frolkis M, et al. Extension oflife-span by introduction of telomerase into normalhuman cells. Science 1998;279:349–52.
16. Vaziri H, Benchimol S. Reconstitution of telomerase
activity in normal human cells leads to elongation oftelomeres and extended replicative life span. Curr Biol1998;8:279–82.
17. MacKenzie KL, Franco S, May C, Sadelain M, MooreMA. Mass cultured human fibroblasts overexpressinghTERT encounter a growth crisis following an extendedperiod of proliferation. Exp Cell Res 2000;259:336–50.
18. Noble JR, Zhong ZH, Neumann AA, Melki JR, Clark SJ,Reddel RR. Alterations in the p16(INK4a) and p53 tumorsuppressor genes of hTERT-immortalized human fibro-blasts. Oncogene 2004;23:3116–21.
19. Wen VW, Wu K, Baksh S, et al. Telomere-drivenkaryotypic complexity concurs with p16INK4a inactiva-tion in TP53-competent immortal endothelial cells.Cancer Res 2006;66:10691–700.
20. Vulliamy T, Marrone A, Goldman F, et al. The RNAcomponent of telomerase is mutated in autosomal do-minant dyskeratosis congenita. Nature 2001;413:432–5.
21. Marrone A, Stevens D, Vulliamy T, Dokal I, Mason PJ.Heterozygous telomerase RNA mutations found indyskeratosis congenita and aplastic anemia reducetelomerase activity via haploinsufficiency. Blood 2004;104:3936–42.
22. Wong JM, Collins K. Telomerase RNA level limitstelomere maintenance in X-linked dyskeratosis conge-nita. Genes Dev 2006;20:2848–58.
Cancer Research
Cancer Res 2008; 68: (9). May 1, 2008 3122 www.aacrjournals.org
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

hTERT Amplification in Cells with Limiting hTR Levels
www.aacrjournals.org 3123 Cancer Res 2008; 68: (9). May 1, 2008
23. Westin ER, Chavez E, Lee KM, et al. Telomererestoration and extension of proliferative lifespan indyskeratosis congenita fibroblasts. Aging Cell 2007;6:383–94.
24. Cristofari G, Lingner J. Telomere length homeostasisrequires that telomerase levels are limiting. EMBO J2006;25:565–74.
25. Yi X, Shay JW, Wright WE. Quantitation of telomerasecomponents and hTERT mRNA splicing patterns inimmortal human cells. Nucleic Acids Res 2001;29:4818–25.
26. Toouli CD, Huschtscha LI, Neumann AA, et al.Comparison of human mammary epithelial cells im-mortalized by simian virus 40 T-Antigen or by thetelomerase catalytic subunit. Oncogene 2002;21:128–39.
27. Perrem K, Colgin LM, Neumann AA, Yeager TR,Reddel RR. Coexistence of alternative lengthening oftelomeres and telomerase in hTERT-transfected GM847cells. Mol Cell Biol 2001;21:3862–75.
28. Colgin LM, Wilkinson C, Englezou A, Kilian A,Robinson MO, Reddel RR. The hTERTa splice variantis a dominant negative inhibitor of telomerase activity.Neoplasia 2000;2:426–32.
29. Cohen SB, Reddel RR. A sensitive direct humantelomerase activity assay. Nat Methods. In press 2008.
30. Bryan TM, Englezou A, Gupta J, Bacchetti S, ReddelRR. Telomere elongation in immortal human cellswithout detectable telomerase activity. EMBO J 1995;14:4240–8.
31. Grant JD, Broccoli D, Muquit M, Manion FJ, Tisdall J,Ochs MF. Telometric: a tool providing simplified, repro-ducible measurements of telomeric DNA from constantfield agarose gels. Biotechniques 2001;31:1314–8.
32. Mozdy AD, Cech TR. Low abundance of telomerase
in yeast: implications for telomerase haploinsufficiency.RNA 2006;12:1721–37.
33. Soder AI, Going JJ, Kaye SB, Keith WN. Tumourspecific regulation of telomerase RNA gene expressionvisualized by in situ hybridization. Oncogene 1998;16:979–83.
34. Heine B, Hummel M, Demel G, Stein H. Demonstra-tion of constant upregulation of the telomerase RNAcomponent in human gastric carcinomas using in situhybridization. J Pathol 1998;185:139–44.
35. Paradis V, Dargere D, Laurendeau I, et al. Expressionof the RNA component of human telomerase (hTR) inprostate cancer, prostatic intraepithelial neoplasia, andnormal prostate tissue. J Pathol 1999;189:213–8.
36. Yashima K, Litzky LA, Kaiser L, et al. Telomeraseexpression in respiratory epithelium during the multi-stage pathogenesis of lung carcinomas. Cancer Res 1997;57:2373–7.
37. Morales CP, Lee EL, Shay JW. In situ hybridization forthe detection of telomerase RNA in the progressionfrom Barrett’s esophagus to esophageal adenocarcino-ma. Cancer 1998;83:652–9.
38. Soder AI, Hoare SF, Muir S, Going JJ, Parkinson EK,Keith WN. Amplification, increased dosage and in situexpression of the telomerase RNA gene in humancancer. Oncogene 1997;14:1013–21.
39. Heselmeyer-Haddad K, Janz V, Castle PE, et al.Detection of genomic amplification of the humantelomerase gene (TERC) in cytologic specimens as agenetic test for the diagnosis of cervical dysplasia. Am JPathol 2003;163:1405–16.
40. Andersson S, Wallin KL, Hellstrom AC, et al. Frequentgain of the human telomerase gene TERC at 3q26 incervical adenocarcinomas. Br J Cancer 2006;95:331–8.
41. Fragnet L, Blasco MA, Klapper W, Rasschaert D. TheRNA subunit of telomerase is encoded by Marek’sdisease virus. J Virol 2003;77:5985–96.
42. Trapp S, Parcells MS, Kamil JP, et al. A virus-encodedtelomerase RNA promotes malignant T cell lymphoma-genesis. J Exp Med 2006;203:1307–17.
43. Hathcock KS, Hemann MT, Opperman KK, StrongMA, Greider CW, Hodes RJ. Haploinsufficiency of mTRresults in defects in telomere elongation. Proc Natl AcadSci USA 2002;99:3591–6.
44. Chiang YJ, Hemann MT, Hathcock KS, et al.Expression of telomerase RNA template, but not telo-merase reverse transcriptase, is limiting for telomerelength maintenance in vivo . Mol Cell Biol 2004;24:7024–31.
45. Marrone A, Walne A, Dokal I. Dyskeratosis congenita:telomerase, telomeres and anticipation. Curr OpinGenet Dev 2005;15:249–57.
46. Stewart SA, Hahn WC, O’Connor BF, et al. Telomer-ase contributes to tumorigenesis by a telomere length-independent mechanism. Proc Natl Acad Sci U S A 2002;99:12606–11.
47. Cao Y, Li H, Deb S, Liu JP. TERT regulates cellsurvival independent of telomerase enzymatic activity.Oncogene 2002;21:3130–8.
48. Yi X, Tesmer VM, Savre-Train I, Shay JW, Wright WE.Both transcriptional and posttranscriptional mecha-nisms regulate human telomerase template RNA levels.Mol Cell Biol 1999;19:3989–97.
49. Bryan TM, Marusic L, Bacchetti S, Namba M,Reddel RR. The telomere lengthening mechanism intelomerase-negative immortal human cells does notinvolve the telomerase RNA subunit. Hum Mol Genet1997;6:921–6.
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

2008;68:3115-3123. Cancer Res Ying Cao, Lily I. Huschtscha, Amanda S. Nouwens, et al. RNA Expression LevelsHuman Mammary Epithelial Cells with Limiting Telomerase Amplification of Telomerase Reverse Transcriptase Gene in
Updated version
http://cancerres.aacrjournals.org/content/68/9/3115
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2008/04/30/68.9.3115.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/68/9/3115.full.html#ref-list-1
This article cites 48 articles, 23 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/68/9/3115.full.html#related-urls
This article has been cited by 4 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on June 5, 2015. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
Related Documents











