AFLP analysis of genetic relationships in the genus Fosterella L.B. Smith (Pitcairnioideae, Bromeliaceae) Martina Rex, Kerstin Patzolt, Katharina Schulte, Georg Zizka, Roberto Va ´ squez, Pierre L. Ibisch, and Kurt Weising Abstract: The neotropical genus Fosterella L.B. Smith (Pitcairnioideae, Bromeliaceae) comprises about 30 species, with a centre of diversity in semiarid to humid habitats of the Andean slopes and valleys of Bolivia. Morphologic differentiation of species is difficult because of a paucity of diagnostic characters, and little is known about the infrageneric phylogeny. Here, we present the results of an amplified fragment length polymorphism (AFLP) analysis of 77 Fosterella specimens, covering 18 recognized species and 9 as-yet undescribed morphospecies. Eight primer combinations produced 310 bands, which were scored as presence/absence characters. Neighbour-joining tree reconstruction revealed 12 clusters (A–L) with various levels of support. Well-supported species groups were also recovered by a principal coordinates analysis. With few exceptions, morphologically defined species boundaries were confirmed by the molecular data. Phylogenetic relationships between species groups remained ambiguous, however, because of short internal branch lengths. The AFLP data were complemented by a survey of the leaf anatomy of 19 Fosterella species. Species concepts and assemblages are discussed in the context of molecular, morphologic, anatomic, ecologic, and biogeographic data. The data suggest that accidental long-distance dispersal and founder events have been important for Fosterella speciation. Key words: Fosterella, Bromeliaceae, Pitcairnioideae, AFLP, molecular phylogeny, leaf anatomy, biogeography. Re ´sume ´: Le genre ne ´otropical Fosterella L.B. Smith (Pitcairnioideae, Bromeliaceae) compte environ 30 espe `ces dont le centre de diversite ´ est situe ´ dans les habitats semi-arides a ` humides des flancs et valle ´es ande ´ens en Bolivie. La diffe ´ren- ciation morphologique des espe `ces est difficile en raison du faible nombre de caracte `res diagnostiques et peu de choses sont connues quant a ` la phyloge ´nie infrage ´ne ´rique. Les auteurs pre ´sentent ici les re ´sultats de l’analyse du polymorphisme de longueur des fragments amplifie ´s (AFLP) sur 77 spe ´cimens de Fosterella couvrant 18 espe `ces reconnues et 9 morpho- espe `ces ine ´dites. Huit combinaisons d’amorces ont produit 310 bandes qui ont e ´te ´ note ´es comme e ´tant pre ´sentes ou absen- tes. La reconstruction d’arbres ‘neighbor-joining’ a re ´ve ´le ´ 12 groupes (A–L) supporte ´s a ` des degre ´s divers. Des groupes d’espe `ces bien supporte ´s ont aussi e ´te ´ obtenus par analyse de coordonne ´es principales. A ` quelques exceptions pre `s, les frontie `res entre les espe `ces de ´finies sur des bases morphologiques ont e ´te ´ confirme ´es suite aux analyses mole ´culaires. Les relations phyloge ´ne ´tiques entre les groupes d’espe `ces demeuraient cependant ambigue ¨s en raison de la faible longueur des branches internes. Les donne ´es AFLP ont e ´te ´ comple ´te ´es par un examen de l’anatomie des feuilles chez 19 espe `ces du genre Fosterella. Les concepts d’espe `ce et de groupes d’espe `ces sont discute ´s dans le contexte de donne ´es mole ´culaires, morphologiques, anatomiques, e ´cologiques et bioge ´ographiques. Il est sugge ´re ´ que la dispersion accidentelle sur de longues distances et l’effet fondateur ont e ´te ´ importants au cours de la spe ´ciation chez le genre Fosterella. Mots cle ´s : Fosterella, Bromeliaceae, Pitcairnioideae, AFLP, phyloge ´nie mole ´culaire, anatomie des feuilles, bioge ´ographie. [Traduit par la Re ´daction] Introduction Bromeliaceae (Poales) are one of the largest neotropical families of flowering plants, comprising more than 2 600 species in 56 genera (Smith and Till 1998). Bromeliads are renowned for their high ecological plasticity, allowing them to occupy a great variety of terrestrial and epiphytic habitats (e.g., arid coastal plains, humid montane forests, and high Received 7 July 2006. Accepted 6 October 2006. Published on the NRC Research Press Web site at http://genome.nrc.ca on 14 March 2007. Corresponding Editor: B. Golding. M. Rex and K. Weising. 2 Plant Molecular Systematics, Institute of Biology, Dept. of Sciences, University of Kassel, Heinrich-Plett-Str. 40, D-34132 Kassel, Germany. K. Patzolt, K. Schulte, and G. Zizka. Research Institute Senckenberg and J.W. Goethe-University, Frankfurt am Main, Germany. R. Va ´squez. Sociedad Boliviana de Bota ´nica, Casilla 3822, Santa Cruz, Bolivia. P.L. Ibisch. 1 Fundacio ´n Amigos de la Naturaleza (FAN), Dept. de Ciencias, Santa Cruz de la Sierra, Bolivia. 1 Present address: Faculty of Forestry, University of Applied Sciences, Eberswalde, Germany. 2 Corresponding author (e-mail: [email protected]). 90 Genome 50: 90–105 (2007) doi:10.1139/G06-141 # 2007 NRC Canada

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AFLP analysis of genetic relationships in thegenus Fosterella L.B. Smith (Pitcairnioideae,Bromeliaceae)
Martina Rex, Kerstin Patzolt, Katharina Schulte, Georg Zizka, Roberto Vasquez,Pierre L. Ibisch, and Kurt Weising
Abstract: The neotropical genus Fosterella L.B. Smith (Pitcairnioideae, Bromeliaceae) comprises about 30 species, with acentre of diversity in semiarid to humid habitats of the Andean slopes and valleys of Bolivia. Morphologic differentiationof species is difficult because of a paucity of diagnostic characters, and little is known about the infrageneric phylogeny.Here, we present the results of an amplified fragment length polymorphism (AFLP) analysis of 77 Fosterella specimens,covering 18 recognized species and 9 as-yet undescribed morphospecies. Eight primer combinations produced 310 bands,which were scored as presence/absence characters. Neighbour-joining tree reconstruction revealed 12 clusters (A–L) withvarious levels of support. Well-supported species groups were also recovered by a principal coordinates analysis. With fewexceptions, morphologically defined species boundaries were confirmed by the molecular data. Phylogenetic relationshipsbetween species groups remained ambiguous, however, because of short internal branch lengths. The AFLP data werecomplemented by a survey of the leaf anatomy of 19 Fosterella species. Species concepts and assemblages are discussedin the context of molecular, morphologic, anatomic, ecologic, and biogeographic data. The data suggest that accidentallong-distance dispersal and founder events have been important for Fosterella speciation.
Key words: Fosterella, Bromeliaceae, Pitcairnioideae, AFLP, molecular phylogeny, leaf anatomy, biogeography.
Resume : Le genre neotropical Fosterella L.B. Smith (Pitcairnioideae, Bromeliaceae) compte environ 30 especes dont lecentre de diversite est situe dans les habitats semi-arides a humides des flancs et vallees andeens en Bolivie. La differen-ciation morphologique des especes est difficile en raison du faible nombre de caracteres diagnostiques et peu de chosessont connues quant a la phylogenie infragenerique. Les auteurs presentent ici les resultats de l’analyse du polymorphismede longueur des fragments amplifies (AFLP) sur 77 specimens de Fosterella couvrant 18 especes reconnues et 9 morpho-especes inedites. Huit combinaisons d’amorces ont produit 310 bandes qui ont ete notees comme etant presentes ou absen-tes. La reconstruction d’arbres ‘neighbor-joining’ a revele 12 groupes (A–L) supportes a des degres divers. Des groupesd’especes bien supportes ont aussi ete obtenus par analyse de coordonnees principales. A quelques exceptions pres, lesfrontieres entre les especes definies sur des bases morphologiques ont ete confirmees suite aux analyses moleculaires. Lesrelations phylogenetiques entre les groupes d’especes demeuraient cependant ambigues en raison de la faible longueur desbranches internes. Les donnees AFLP ont ete completees par un examen de l’anatomie des feuilles chez 19 especes dugenre Fosterella. Les concepts d’espece et de groupes d’especes sont discutes dans le contexte de donnees moleculaires,morphologiques, anatomiques, ecologiques et biogeographiques. Il est suggere que la dispersion accidentelle sur de longuesdistances et l’effet fondateur ont ete importants au cours de la speciation chez le genre Fosterella.
Mots cles : Fosterella, Bromeliaceae, Pitcairnioideae, AFLP, phylogenie moleculaire, anatomie des feuilles, biogeographie.
[Traduit par la Redaction]
Introduction
Bromeliaceae (Poales) are one of the largest neotropicalfamilies of flowering plants, comprising more than 2 600
species in 56 genera (Smith and Till 1998). Bromeliads arerenowned for their high ecological plasticity, allowing themto occupy a great variety of terrestrial and epiphytic habitats(e.g., arid coastal plains, humid montane forests, and high
Received 7 July 2006. Accepted 6 October 2006. Published on the NRC Research Press Web site at http://genome.nrc.ca on 14 March2007.
Corresponding Editor: B. Golding.
M. Rex and K. Weising.2 Plant Molecular Systematics, Institute of Biology, Dept. of Sciences, University of Kassel, Heinrich-Plett-Str.40, D-34132 Kassel, Germany.K. Patzolt, K. Schulte, and G. Zizka. Research Institute Senckenberg and J.W. Goethe-University, Frankfurt am Main, Germany.R. Vasquez. Sociedad Boliviana de Botanica, Casilla 3822, Santa Cruz, Bolivia.P.L. Ibisch.1 Fundacion Amigos de la Naturaleza (FAN), Dept. de Ciencias, Santa Cruz de la Sierra, Bolivia.
1Present address: Faculty of Forestry, University of Applied Sciences, Eberswalde, Germany.2Corresponding author (e-mail: [email protected]).
90
Genome 50: 90–105 (2007) doi:10.1139/G06-141 # 2007 NRC Canada

Andean savannahs). Their great ecologic versatility appearsto be linked to several key innovations, including unique fo-liar trichomes capable of water absorption, succulence, foliarimpoundment, and crassulacean acid metabolism photosyn-thesis (reviewed by Benzing 2000). In spite of the speciesrichness of bromeliads and their abundance in neotropicalhabitats, updated revisions are lacking for most of the gen-era, and our understanding of speciation in this group ispoor.
The genus Fosterella comprises small to medium-sized(up to 1 m in the flowering stage) meso- to xerophytic ter-restrial bromeliad species. The usually stemless (acaules-cent) plants carry rosulate leaves. Flowers areinconspicuous and mostly whitish; fruits are capsules thatrelease large numbers of minute, appendaged seeds at ma-turity. Fosterella species are distinguished from other, pre-sumably adjacent, genera by naked petals, basifixed anthers(which are coiled at anthesis), and inner filaments adnate tothe petals. The genus is distributed from the Peruvian Andesand the Brazilian Amazon in the north to Paraguay andnorthern Argentina in the south, with a centre of diversityin arid and semihumid habitats of the northeast Andeanslopes of Bolivia (Ibisch et al. 2002). Many of its speciesare characterized by only small distribution ranges. Adisjunct occurrence in Central America is marked byF. micrantha. Fosterella has traditionally been assignedto the subfamily Pitcairnioideae of Bromeliaceae, mainlybecause of its fruit characters. In the most recent mono-graph of this subfamily, Smith and Downs (1974) recog-nized 13 Fosterella species. Since then, additional taxahave been formally described, raising the species numberto 30 (Rauh 1979, 1987; Luther 1981, 1997; Smith andRead 1992; Ibisch et al. 1997, 1999, 2002; Kessler etal. 1999). Little is known so far about phylogenetic rela-tionships within the genus.
Recent molecular studies, based on chloroplast DNA se-quences, have demonstrated that Pitcairnioideae, in their tra-ditional circumscription, are paraphyletic, and actuallyrepresent 6 independent evolutionary lineages (Terry et al.1997; Horres et al. 2000, 2007; Crayn et al. 2004; Givnishet al. 2004, 2007). Fosterella remained with Pitcairnioideaes. str. and proved to be monophyletic in all molecular stud-ies. According to the best-resolved chloroplast trees avail-able, its closest relatives are Deuterocohnia, Dyckia, andEncholirium (Crayn et al. 2004; Givnish et al. 2004). Sev-eral reasons account for the lack of phylogenetic informationat the infrageneric level. First, morphological differentiationof species is notoriously difficult, and the few diagnosticcharacters are only available in the flowering stage. Second,many of the Fosterella species are rare, endemic, and (or)display narrow ecologic amplitudes. Some species areknown only from type collections (Kromer et al. 1999;Ibisch et al. 2002). Third, Fosterella collections are poorlyrepresented in herbaria and botanical gardens. Fourth, sev-eral undescribed taxa are awaiting taxonomic treatment, andthe discovery of additional species is likely.
In a first attempt to shed light on the infrageneric phylog-eny of Fosterella, a random amplified polymorphic DNA(RAPD) analysis of 26 accessions from 12 species was per-formed (Rex 2001; Ibisch et al. 2002). Neighbour-joining(NJ) analysis of the RAPD dataset identified a set of species
groups, most notably the so-called penduliflora alliance(F. penduliflora, F. latifolia, F. chiquitana), which was sup-ported by a 99% bootstrap value. However, there were alsoinstances where specimens did not group according to theirspecies assignment, or occupied a more isolated position.Most species were represented by single accessions, andtherefore the general validity of conclusions from thisRAPD investigation suffered from limited sampling. The po-tential of various noncoding chloroplast DNA sequences forresolving relationships within Fosterella was also tested.Unfortunately, infrageneric chloroplast (cp)DNA sequencevariation proved to be relatively low, as it is in other generaof the presumably young crown group of Bromeliaceae (e.g.,Horres et al. 2000). Attempts to amplify the nuclear riboso-mal internal transcribed spacer (ITS) region with PCR, usingstandard universal primers, were unsuccessful. GeneratingITS sequence data from Bromeliaceae is known to be diffi-cult (Barfuss et al. 2004; Tuthill and Brown 2004) and, toour knowledge, no published ITS phylogenies of Bromelia-ceae currently exist.
It has been argued that amplified fragment length poly-morphisms (AFLPs) (Vos et al. 1995) are the method ofchoice for analyzing relationships between closely relatedtaxa, where DNA sequencing reveals hardly any variation(Hodkinson et al. 2000; Despres et al. 2003). AFLPs are ap-plicable to all organisms without a priori sequence informa-tion, represent a reasonably random sample of the totalgenome, and often show good reproducibility with the appli-cation of stringent PCR conditions (Hansen et al. 1999). Inthe current investigation, we used AFLP profiling to findout whether Fosterella species, recognized on the basis ofthe few morphologic characters available, also represent ge-netically distinct units; to address genetic relationshipsamong Fosterella species; and to identify the taxonomic af-finities of specimens with yet-unknown species designation.Genetic relationships suggested by the molecular data arediscussed in the context of morphology, leaf anatomy, ecol-ogy, and biogeography.
Material and methods
Plant materialA total of 77 Fosterella accessions, representing 18 of
the 30 recognized Fosterella species, were included in theAFLP analysis (Table 1). Nine accessions that do not corre-spond morphologically to any described species were provi-sionally assigned to morphospecies F. spec. 1 to 9. Thirty-five accessions from 19 Fosterella species were subjectedto a complementary leaf anatomical study (Table 1). Allbut 2 specimens (F. micrantha from Mexico and F. spec. 8from Brazil) were collected in Bolivia, or represent materialof Bolivian species taken from living collections elsewhere.For molecular studies, leaves were used fresh or werestored frozen until DNA extraction. Attempts to quick-drythe moderately succulent leaves by treating them with silicagel were unsuccessful and resulted in degraded DNA prep-arations. Voucher specimens and duplicates have been de-posited in various herbaria and living collections. Detailson collection numbers of voucher specimens, accessionnumbers of living material, and collection sites are givenin Table 1.
Rex et al. 91
# 2007 NRC Canada
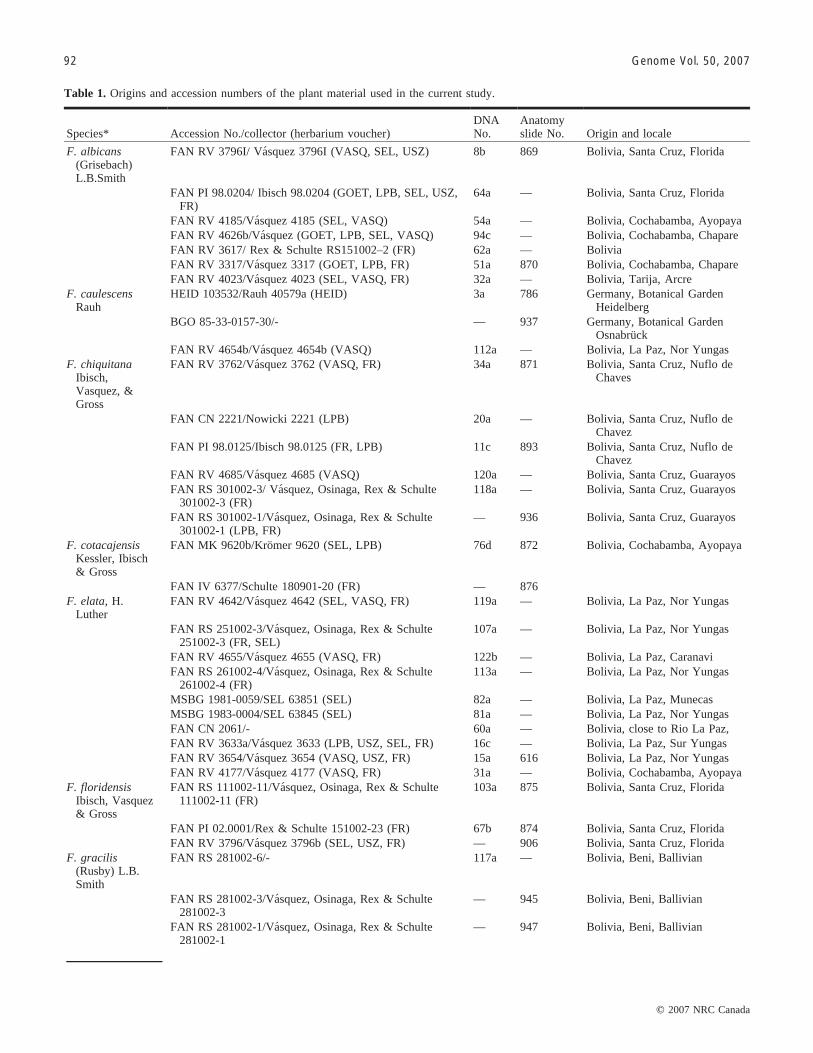
Table 1. Origins and accession numbers of the plant material used in the current study.
Species* Accession No./collector (herbarium voucher)DNANo.
Anatomyslide No. Origin and locale
F. albicans(Grisebach)L.B.Smith
FAN RV 3796I/ Vasquez 3796I (VASQ, SEL, USZ) 8b 869 Bolivia, Santa Cruz, Florida
FAN PI 98.0204/ Ibisch 98.0204 (GOET, LPB, SEL, USZ,FR)
64a — Bolivia, Santa Cruz, Florida
FAN RV 4185/Vasquez 4185 (SEL, VASQ) 54a — Bolivia, Cochabamba, AyopayaFAN RV 4626b/Vasquez (GOET, LPB, SEL, VASQ) 94c — Bolivia, Cochabamba, ChapareFAN RV 3617/ Rex & Schulte RS151002–2 (FR) 62a — BoliviaFAN RV 3317/Vasquez 3317 (GOET, LPB, FR) 51a 870 Bolivia, Cochabamba, ChapareFAN RV 4023/Vasquez 4023 (SEL, VASQ, FR) 32a — Bolivia, Tarija, Arcre
F. caulescensRauh
HEID 103532/Rauh 40579a (HEID) 3a 786 Germany, Botanical GardenHeidelberg
BGO 85-33-0157-30/- — 937 Germany, Botanical GardenOsnabruck
FAN RV 4654b/Vasquez 4654b (VASQ) 112a — Bolivia, La Paz, Nor YungasF. chiquitana
Ibisch,Vasquez, &Gross
FAN RV 3762/Vasquez 3762 (VASQ, FR) 34a 871 Bolivia, Santa Cruz, Nuflo deChaves
FAN CN 2221/Nowicki 2221 (LPB) 20a — Bolivia, Santa Cruz, Nuflo deChavez
FAN PI 98.0125/Ibisch 98.0125 (FR, LPB) 11c 893 Bolivia, Santa Cruz, Nuflo deChavez
FAN RV 4685/Vasquez 4685 (VASQ) 120a — Bolivia, Santa Cruz, GuarayosFAN RS 301002-3/ Vasquez, Osinaga, Rex & Schulte
301002-3 (FR)118a — Bolivia, Santa Cruz, Guarayos
FAN RS 301002-1/Vasquez, Osinaga, Rex & Schulte301002-1 (LPB, FR)
— 936 Bolivia, Santa Cruz, Guarayos
F. cotacajensisKessler, Ibisch& Gross
FAN MK 9620b/Kromer 9620 (SEL, LPB) 76d 872 Bolivia, Cochabamba, Ayopaya
FAN IV 6377/Schulte 180901-20 (FR) — 876F. elata, H.
LutherFAN RV 4642/Vasquez 4642 (SEL, VASQ, FR) 119a — Bolivia, La Paz, Nor Yungas
FAN RS 251002-3/Vasquez, Osinaga, Rex & Schulte251002-3 (FR, SEL)
107a — Bolivia, La Paz, Nor Yungas
FAN RV 4655/Vasquez 4655 (VASQ, FR) 122b — Bolivia, La Paz, CaranaviFAN RS 261002-4/Vasquez, Osinaga, Rex & Schulte
261002-4 (FR)113a — Bolivia, La Paz, Nor Yungas
MSBG 1981-0059/SEL 63851 (SEL) 82a — Bolivia, La Paz, MunecasMSBG 1983-0004/SEL 63845 (SEL) 81a — Bolivia, La Paz, Nor YungasFAN CN 2061/- 60a — Bolivia, close to Rio La Paz,FAN RV 3633a/Vasquez 3633 (LPB, USZ, SEL, FR) 16c — Bolivia, La Paz, Sur YungasFAN RV 3654/Vasquez 3654 (VASQ, USZ, FR) 15a 616 Bolivia, La Paz, Nor YungasFAN RV 4177/Vasquez 4177 (VASQ, FR) 31a — Bolivia, Cochabamba, Ayopaya
F. floridensisIbisch, Vasquez& Gross
FAN RS 111002-11/Vasquez, Osinaga, Rex & Schulte111002-11 (FR)
103a 875 Bolivia, Santa Cruz, Florida
FAN PI 02.0001/Rex & Schulte 151002-23 (FR) 67b 874 Bolivia, Santa Cruz, FloridaFAN RV 3796/Vasquez 3796b (SEL, USZ, FR) — 906 Bolivia, Santa Cruz, Florida
F. gracilis(Rusby) L.B.Smith
FAN RS 281002-6/- 117a — Bolivia, Beni, Ballivian
FAN RS 281002-3/Vasquez, Osinaga, Rex & Schulte281002-3
— 945 Bolivia, Beni, Ballivian
FAN RS 281002-1/Vasquez, Osinaga, Rex & Schulte281002-1
— 947 Bolivia, Beni, Ballivian
92 Genome Vol. 50, 2007
# 2007 NRC Canada
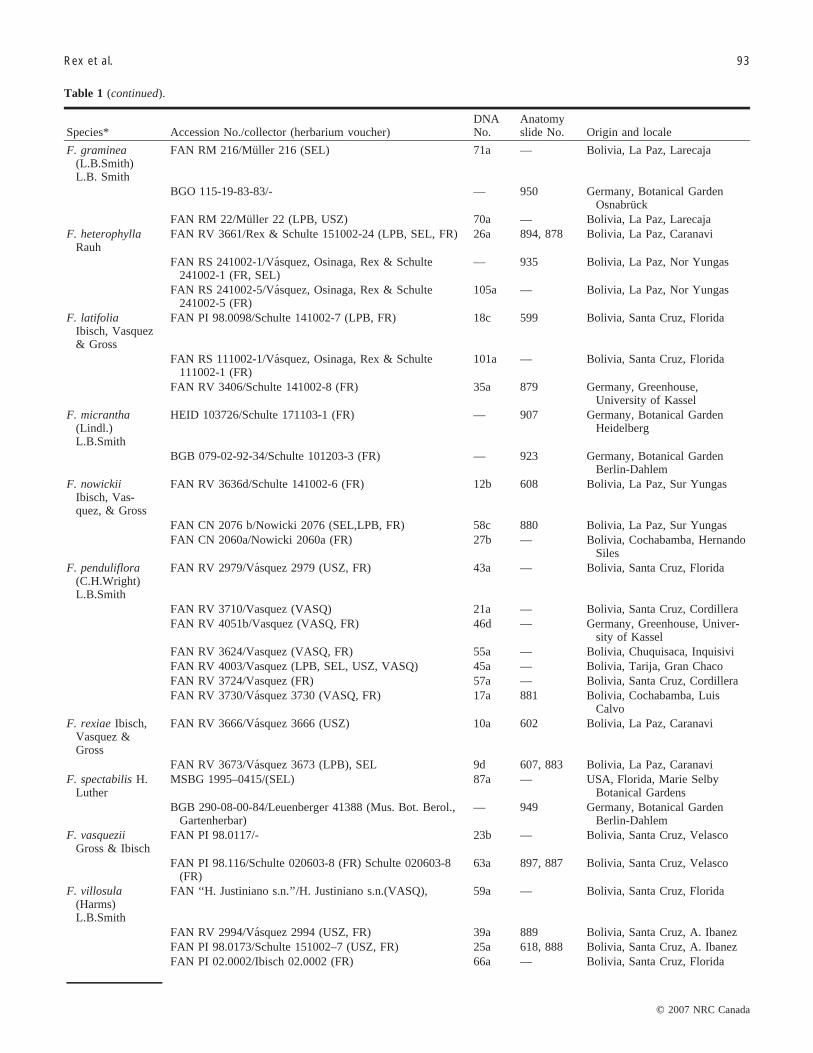
Table 1 (continued).
Species* Accession No./collector (herbarium voucher)DNANo.
Anatomyslide No. Origin and locale
F. graminea(L.B.Smith)L.B. Smith
FAN RM 216/Muller 216 (SEL) 71a — Bolivia, La Paz, Larecaja
BGO 115-19-83-83/- — 950 Germany, Botanical GardenOsnabruck
FAN RM 22/Muller 22 (LPB, USZ) 70a — Bolivia, La Paz, LarecajaF. heterophylla
RauhFAN RV 3661/Rex & Schulte 151002-24 (LPB, SEL, FR) 26a 894, 878 Bolivia, La Paz, Caranavi
FAN RS 241002-1/Vasquez, Osinaga, Rex & Schulte241002-1 (FR, SEL)
— 935 Bolivia, La Paz, Nor Yungas
FAN RS 241002-5/Vasquez, Osinaga, Rex & Schulte241002-5 (FR)
105a — Bolivia, La Paz, Nor Yungas
F. latifoliaIbisch, Vasquez& Gross
FAN PI 98.0098/Schulte 141002-7 (LPB, FR) 18c 599 Bolivia, Santa Cruz, Florida
FAN RS 111002-1/Vasquez, Osinaga, Rex & Schulte111002-1 (FR)
101a — Bolivia, Santa Cruz, Florida
FAN RV 3406/Schulte 141002-8 (FR) 35a 879 Germany, Greenhouse,University of Kassel
F. micrantha(Lindl.)L.B.Smith
HEID 103726/Schulte 171103-1 (FR) — 907 Germany, Botanical GardenHeidelberg
BGB 079-02-92-34/Schulte 101203-3 (FR) — 923 Germany, Botanical GardenBerlin-Dahlem
F. nowickiiIbisch, Vas-quez, & Gross
FAN RV 3636d/Schulte 141002-6 (FR) 12b 608 Bolivia, La Paz, Sur Yungas
FAN CN 2076 b/Nowicki 2076 (SEL,LPB, FR) 58c 880 Bolivia, La Paz, Sur YungasFAN CN 2060a/Nowicki 2060a (FR) 27b — Bolivia, Cochabamba, Hernando
SilesF. penduliflora
(C.H.Wright)L.B.Smith
FAN RV 2979/Vasquez 2979 (USZ, FR) 43a — Bolivia, Santa Cruz, Florida
FAN RV 3710/Vasquez (VASQ) 21a — Bolivia, Santa Cruz, CordilleraFAN RV 4051b/Vasquez (VASQ, FR) 46d — Germany, Greenhouse, Univer-
sity of KasselFAN RV 3624/Vasquez (VASQ, FR) 55a — Bolivia, Chuquisaca, InquisiviFAN RV 4003/Vasquez (LPB, SEL, USZ, VASQ) 45a — Bolivia, Tarija, Gran ChacoFAN RV 3724/Vasquez (FR) 57a — Bolivia, Santa Cruz, CordilleraFAN RV 3730/Vasquez 3730 (VASQ, FR) 17a 881 Bolivia, Cochabamba, Luis
CalvoF. rexiae Ibisch,
Vasquez &Gross
FAN RV 3666/Vasquez 3666 (USZ) 10a 602 Bolivia, La Paz, Caranavi
FAN RV 3673/Vasquez 3673 (LPB), SEL 9d 607, 883 Bolivia, La Paz, CaranaviF. spectabilis H.
LutherMSBG 1995–0415/(SEL) 87a — USA, Florida, Marie Selby
Botanical GardensBGB 290-08-00-84/Leuenberger 41388 (Mus. Bot. Berol.,
Gartenherbar)— 949 Germany, Botanical Garden
Berlin-DahlemF. vasquezii
Gross & IbischFAN PI 98.0117/- 23b — Bolivia, Santa Cruz, Velasco
FAN PI 98.116/Schulte 020603-8 (FR) Schulte 020603-8(FR)
63a 897, 887 Bolivia, Santa Cruz, Velasco
F. villosula(Harms)L.B.Smith
FAN ‘‘H. Justiniano s.n.’’/H. Justiniano s.n.(VASQ), 59a — Bolivia, Santa Cruz, Florida
FAN RV 2994/Vasquez 2994 (USZ, FR) 39a 889 Bolivia, Santa Cruz, A. IbanezFAN PI 98.0173/Schulte 151002–7 (USZ, FR) 25a 618, 888 Bolivia, Santa Cruz, A. IbanezFAN PI 02.0002/Ibisch 02.0002 (FR) 66a — Bolivia, Santa Cruz, Florida
Rex et al. 93
# 2007 NRC Canada

DNA isolationTotal genomic DNA, from 100 to 150 mg of fresh or fro-
zen leaves of individual plants, was isolated, either with aDNeasy Plant Mini Kit (Qiagen, Hilden, Germany) or withvarious modifications of the cetyl trimethylammonium bro-mide (CTAB) procedure (Weising et al. 2005). Final DNAconcentrations were determined electrophoretically, usingknown amounts of �-DNA as standards.
AFLP assaysAFLP analyses were performed in accordance with the
protocols described by Debener and Mattiesch (1999) andBanfer et al. (2004), with some modifications. In short,250 ng of genomic DNA were double-digested in a finalvolume of 30 mL at 37 8C with HindIII and MseI overnight,and ligated to HindIII and MseI adapters in the same reac-
tion. Two consecutive PCR amplifications with primers,containing either 1 (+1) or 3 (+3) selective nucleotides attheir 3’ ends (Table 2), were then carried out in a BiometraT-Gradient Cycler (Biometra, Gottingen, Germany) or aBio-Rad i-Cycler (Bio-Rad Laboratories, Munich, Ger-many).
In the first, preselective PCR, 5 mL of the restriction-liga-tion assay were combined with 2.5 ng/mL of unlabeledMseI+1 primer, 1.5 ng/mL of unlabeled HindIII+1 primer,1.5 mmol/L MgCl2, 0.2 mmol/L of each deoxynucleosidetriphosphate (dNTP), 20 mmol/L Tris–HCl (pH 8.0),50 mmol/L KCl, and 1 U Taq DNA polymerase (Invitrogen,Karlsruhe, Germany). Assays were adjusted to 50 mL withdistilled water and subjected to 20 cycles of amplification,each consisting of 94 8C for 30 s, 60 8C for 30 s, and72 8C for 60 s. Final extension was at 72 8C for 7 min. Se-
Table 1 (concluded).
Species* Accession No./collector (herbarium voucher)DNANo.
Anatomyslide No. Origin and locale
FAN RV 4623/Vasquez 4623 (VASQ, FR) 104a — Bolivia, Cochabamba, ChapareFAN RV 3318/Vasquez 3318 (VASQ) 24c — Bolivia, Cochabamba, ChapareFAN RV 4650a/Vasquez 4650a (VASQ, FR) 126a — Bolivia, La Paz, Nor YungasFAN RV 3322/Vasquez 3322 (VASQ) 40a — Bolivia, Cochabamba, Chapare
F. weberbaueri(Mez)L.B.Smith
FAN RV 3570/Vasquez 3570 (VASQ, SEL, LPB, USZ,FR)
48a — Bolivia, Cochabamba, Chapare
FAN RM 217*2/(SEL) 95a 896 Bolivia, La Paz, LarecajaFAN RV 4656/Schulte 020603–4 (FR) 121a 895 Bolivia, La Paz, CaranaviFAN RS 261002-18/Vasquez, Osinaga, Rex & Schulte
261002-18 (FR)114a — Bolivia, La Paz, Nor Yungas
F. cf. weber-baueri
FAN RV 4650/- 122b — Bolivia, La Paz, Caranavi
F. weddelliana(Brongniart exBaker)L.B.Smith
FAN CN 2034/Nowicki 2034 (FR) 37a — Bolivia
FAN RV 3612/Vasquez 3612 (VASQ, FR) 13a 611 Bolivia, La Paz, InquisiviFAN RV 3620/Vasquez 3620 (SEL, VASQ) 56a — Bolivia, La Paz, InquisiviFAN RV 3627/Vasquez (VASQ, FR) 36a — Bolivia, La Paz, Inquisivi
F. spec 1 FAN IV 6387/Vargas 6387 (FR) 30a — Bolivia, Cochabamba, AyopayaF. spec 2 FAN RS 261002-10/Vasquez, Osinaga, Rex & Schulte
261002-10 (FR)111a — Bolivia, La Paz, Nor Yungas
F. spec. 2 FAN RV 3652/Vasquez 3652 (VASQ, FR) 29a — Bolivia, La Paz, Nor YungasF. spec. 3 FAN TK 1398b/Kromer 1398b (USZ) 28d 948, 898 Bolivia, La Paz, CaranaviF. spec. 4 FAN RV 3729a/Vasquez 3729a (VASQ) 128a — Bolivia, Chuquisaca, Luis CalvoF. spec. 5 MSBG 1980-0854/- 88a — USA, Florida, Marie Selby
Botanical GardensF. spec. 6 FAN RS 271002-1/Vasquez, Osinaga, Rex & Schulte
261002-10 (FR)116a — Bolivia, Beni, Ballivian
F. spec. 7 FAN RV 3674a/Vasquez 3674a (SEL, VASQ, FR) 49b — Bolivia, La Paz, CaranaviF. spec. 8 J. B. Fernandes da Silva s.n. (cult. E. Leme 5078/HB); 129a — Brazil, Para, ItaitubaF. spec. 9 MSBG 1980-1733/ 79a — USA, Florida, Marie Selby
Botanical Gardens
Note: BGB, Botanical Garden Berlin-Dahlem; BGO, Botanical Garden Osnabruck, Germany; FAN, living collections of the Fundacion Amigos de laNaturaleza, Santa Cruz, Bolivia; FR, Forschungsinstitut Senckenburg, Frankfurt/Main, Germany; GOET, Herbarium of the Georg-August-University, Gottin-gen, Germany; HB, Herbarium Bradeanum, Rio de Janeiro, Brazil; HEID, Botanical Garden of the University of Heidelberg, Germany; LPB, Herbario Na-cional de Bolivia, La Paz, Bolivia; MSBG and SEL, Marie Selby Botanical Garden, Sarasota, US; USZ, Universdad Autonoma Gabriel Rene Moreno, SantaCruz, Bolivia; VASQ, Herbarium Vasquezianum.*Fosterella species are arranged alphabetically. Nomenclature follows Smith and Downs (1974), and for later described species Ibisch et al. (1997,
1999, 2002), Kessler et al. (1999), Luther (1981, 1997), Rauh (1979, 1987), and Smith and Read (1992). Abbreviations of author names follow Brummitand Powell (1992). Fosterella spec. 1–9 are as yet unnamed morphospecies.
94 Genome Vol. 50, 2007
# 2007 NRC Canada

lective PCRs were then performed with 2.5 mL of a 1:20diluted preamplified DNA as a template, and various combi-nations of an unlabeled MseI+3 primer (3 ng/mL) (Roth,Karlsruhe, Germany) and a fluorescence-labeled HindIII+3primer (either 3 ng/mL of IRDye700 or 3.6 ng/mL of IR-Dye800-labeled primer) (MWG Biotech, Ebersberg,Germany) (Table 2). Each 30 mL assay also contained2 mmol/L MgCl2, 0.2 mmol/L of each dNTP, 0.3% TritonX-100, 20 mmol/L Tris–HCl (pH 8.0), 50 mmol/L KCl, and0.25 U Taq DNA polymerase (Invitrogen). A touchdownprotocol was followed for 47 cycles, each consisting of94 8C for 30 s, 658 to 56 8C for 30 s, and 72 8C for 60 s.Starting at 65 8C, the annealing temperature was reduced by0.8 8C per cycle during the first 11 cycles, and then left con-stant at 56 8C. Final extension was at 72 8C for 7 min.
Final products of the selective PCR were mixed with anequal volume of loading buffer, containing 98% (v/v) forma-mide, and denatured at 80 8C for 5 min. Samples (0.5 mL) ofeach reaction were electrophoresed on denaturing 41 cm �0.2 mm polyacrylamide gels (6% Sequagel XR (NationalDiagnostics, Atlanta, Ga.) in 1æ TBE buffer (Sambrook andRussell 2001)), using an automated sequencer (LI-COR4200 IR2; LI-COR Inc., Lincoln, Nebr.). Fragment mobilitywas measured with real-time laser scanning. Gel imageswere stored electronically for further analysis.
Data analysisAFLP banding patterns were recorded as the presence (1)
or absence (0) of a band at a particular position, with thehelp of GenImageIR version 4.02 software (ScanalyticsInc., Fairfax, Va.). The raw data were edited manually, aftervisual inspection. Faint or fuzzy bands were generallyignored. Scored fragment sizes ranged from 40 to 575 nu-cleotides. Standard lanes carrying identical samples wererun on each gel, which facilitated the combination of data-sets from 2 or more independent runs.
For tree construction, the binary character matrix wasconverted into a distance matrix based on the Nei and Li(1979) ( = Dice) index of similarity, using NTSYSpc 2.10p(Rohlf 2000) or Treecon 1.3b software (Van de Peer and DeWachter 1994). Phenograms were generated either by un-weighted pair group method with arithmetic mean(UPGMA) cluster analysis (Sneath and Sokal 1973) or bythe neighbour-joining (NJ) algorithm (Saitou and Nei 1987),using the same program packages. An unweighted maximum
parsimony analysis of the AFLP character matrix was con-ducted using PAUP* 4.0b10 (Swofford 2002). Characterchanges were interpreted under ACCTRAN optimizationon, random addition, tree bisection-reconnection swappingoptions, and Maxtrees set to increase without limits.Statistical support for individual furcations in both the NJand maximum parsimony trees was evaluated withbootstrapping (Felsenstein 1985). Bootstrap analyses used1000 replicates. The complemented programs INTERVALDATA, DCENTER, and EIGEN implemented in theNTSYSpc 2.10p software package (Rohlf 2000) were usedto perform a principle coordinates analysis.
Leaf blade anatomyMature leaves were used fresh, or fixed in formalin/acetic
acid/alcohol (FAA) and stored in 70% ethanol. Completetransverse-hand sections of the leaf blade (about halfwaybetween the leaf tip and base) were prepared from unstainedfresh material. Microtome sections were obtained frommaterial either embedded in Paraplast and stained withsafranine-astrablue or embedded in HEMA (Igersheim andCichocki 1996) and stained with toluidine. The sections wereinvestigated and documented with light microscopy (LeicaDialux 22), a digital camera system (Leica DC 300), andcamera lucida drawings. For measuring and analyzing thesections, IM 1000 software (Leica) was applied.
Morphology and biogeographyThe assessment of morphologic, ecologic, and biogeo-
graphic characters was derived from a database comprisingmore than 360 specimens of all known Fosterella species.The database includes data from the literature (Smith andDowns 1974; Rauh 1979, 1987; Luther 1981, 1997; Smithand Read 1992; Ibisch et al. 1997, 1999, 2002; Kessler etal. 1999), information obtained from the study of cultivatedplants (mainly in the Living Collection of the FundacionAmigos de la Naturaleza, Santa Cruz, Bolivia), and personalobservations that were made during numerous field trips toall ecoregions of Bolivia (Pierre L. Ibisch, 1993–2003).The classification of the Bolivian ecoregions follows thatdescribed by Ibisch et al. (2004).
Results
AFLP dataIn pilot experiments, 54 combinations of labeled HindIII-
primer and unlabeled MseI-primer with +3 selective baseseach were screened with a small set of template DNAs.Well-resolved banding patterns were obtained with 13 ofthese combinations, and 8 primer pairs were eventually re-tained to analyse the entire set of 112 samples (Table 2). Atotal of 77 template DNA samples produced distinct AFLPprofiles with all primer sets, and were included in the phylo-genetic analysis (Table 1). Unfortunately, we were not ableto generate consistent AFLP patterns from F. micrantha,which is the only Fosterella species present in CentralAmerica. The reproducibility of banding patterns was exem-plarily tested with 2 primer combinations and 61 taxa. Num-bers of unequivocally scorable band positions varied from30 to 52, depending on the primer pair. Of 310 band posi-tions totally scored, 96% were variable (Table 2). AFLP fin-
Table 2. Primer combinations used for selectiveamplification.
Primer combinationTotalbands
Polymorphicbands (%)
Mse+CAA/Hind+ACC 52 94Mse+CAC/Hind+ACC 34 94Mse+CAC/Hind+AGC 32 97Mse+CAA/Hind+ACA 36 94Mse+CAA/Hind+AAG 42 100Mse+AAG/Hind+AAC 32 94Mse+AGC/Hind+ACG 30 100Mse+AAG/Hind+ACA 52 96Total 310 96
Rex et al. 95
# 2007 NRC Canada
gerprints generated by the primer combination Mse+CAAand Hind+AAG from a subset of samples are exemplarilyshown in Fig. 1.
Phenetic analysesPairwise Nei and Li (1979) similarity values ranged from
0.39 to 0.95 for all samples, and generally exceeded 0.66 atthe species level. The single plant of F. spectabilis and 1 ofthe 2 investigated samples of F. graminea exhibited thelargest distances from the other specimens and were arbitra-rily chosen to root the trees. Defining alternative outgroupsor using the midpoint rooting option had no major influenceon tree topology (not shown). Both the UPGMA and NJ treereceived cophenetic correlation coefficients > 0.83, and ex-hibited almost identical branching patterns. Only the NJ treeis shown here (Fig. 2). It is subdivided into 12 clusters thatreceive various levels of bootstrap support (BS). The moder-ately well-supported cluster A (73% BS) comprises allspecimens of F. elata. Clusters B and C each unite 2 con-specific accessions from closely adjacent localities (i.e.,F. vasquezii (87% BS) and F. floridensis (100% BS), re-spectively). Cluster D (87% BS) harbours 2 accessions ofF. heterophylla and 2 accessions of the unclassified F. spec.2. The monophyly of these 2 species also receives good sup-port. Cluster E is again moderately well-supported (69% BS)
and contains all specimens of F. nowickii andF. weddelliana, with the clonotype specimen of F. cotaca-jensis in a basal position. The monospecific cluster F (58%BS) is formed by the 2 accessions of F. caulescens. F. re-xiae, together with the unclassified F. spec. 3, makes upcluster G (73% BS). All but 1 of the investigated accessionsof F. albicans, plus the unclassified F. spec. 4, constitute theunsupported cluster H. Species groups A–H are united in alarge (but unsupported) supercluster, with F. graminea(RM216), F. gracilis, and F. albicans (RV4023) forming abasal grade.
The weakly defined cluster I contains a second F. grami-nea (RM22) and the unclassified F. spec. 5 and F. spec. 6,whereas the well-supported cluster J (98% BS) comprises allaccessions of F. villosula. Internal bootstrap values withincluster J are also relatively high, suggesting considerable sub-division within F. villosula. Cluster K (72% BS) comprisesall specimens of F. penduliflora, F. latifolia, and F. chiqui-tana, plus the unclassified F. spec. 7. The samples from these4 apparently closely related species are intermingled witheach other. Cluster L (57% BS) harbours all sampled plantsof F. weberbaueri, together with an unclassified species (F.spec. 8), which was collected in Brazil.
With few exceptions, relationships between the clustersoutlined above remain obscure. Clusters D and E are sister
Fig. 1. Amplified fragment length polymorphism (AFLP) profiles generated from a subset of the investigated Fosterella specimens afterselective amplification with a combination of unlabeled Mse+CAA and IRDye700-labeled Hind+AAG primers. PCR products were sepa-rated on a 6% denaturing polyacrylamide gel in an automated Li-COR sequencer.
96 Genome Vol. 50, 2007
# 2007 NRC Canada
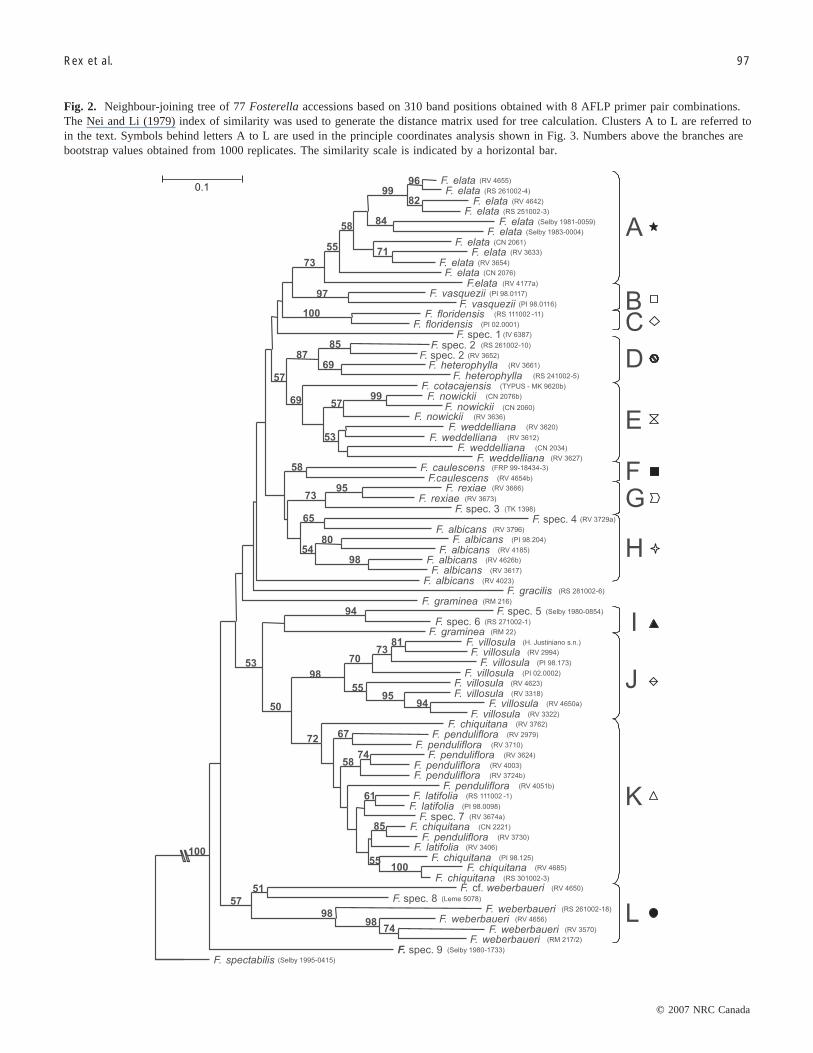
Fig. 2. Neighbour-joining tree of 77 Fosterella accessions based on 310 band positions obtained with 8 AFLP primer pair combinations.The Nei and Li (1979) index of similarity was used to generate the distance matrix used for tree calculation. Clusters A to L are referred toin the text. Symbols behind letters A to L are used in the principle coordinates analysis shown in Fig. 3. Numbers above the branches arebootstrap values obtained from 1000 replicates. The similarity scale is indicated by a horizontal bar.
Rex et al. 97
# 2007 NRC Canada

groups, as are J and K, but each with very low support (BS57% and 50%, respectively). The latter relationship was alsoobserved in our preliminary RAPD study, where these 2clusters were referred to as villosula and pendulifloragroups, respectively (Ibisch et al. 2002). Finally, clusters I,J, and K are united in a larger cluster that receives 53%bootstrap support.
Phylogenetic analysisThe original binary character matrix was also subjected to
a phylogenetic analysis using maximum parsimony as an op-timality criterion, as was also done in other AFLP studies(e.g., Kardolus et al. 1998; Koopman et al. 2001; Despres etal. 2003). Defining F. spectabilis as an outgroup, PAUP*found 4 trees with 2491 steps, a consistency index of 0.12and a retention index of 0.5164 (not shown). The low con-sistency index value indicates a high level of homoplasy, aswould be expected with the application of AFLPs to a largedataset (see Despres et al. 2003 for a discussion). The strictconsensus tree displays a basal polytomy of various clades,the species compositions of which are generally congruentwith the clusters defined above in the NJ tree. The followingclades (clusters) receive bootstrap supports above 50% inthe maximum parsimony tree: A (56%), B (81%), C (99%),D (57%), E (65%), J (63%), K (97%), and L (53%). Otherclades, and relationships between clades, are not resolved.
Principal coordinates analysisRelationships among Fosterella specimens investigated by
AFLP are further illustrated in the results of a principal co-ordinates analysis (Fig. 3). The first 3 coordinates togetheraccount for 52% (i.e., 28.4%, 12.2%, and 11.4%) of the totalvariance. Essentially, the same groups are revealed as in theNJ tree, but resolution varies between the individual groups.Whereas clusters A, B, D, J, K, and L are each relativelywell resolved in the 3-dimensional principle coordinatesplot, other groups show considerable overlap.
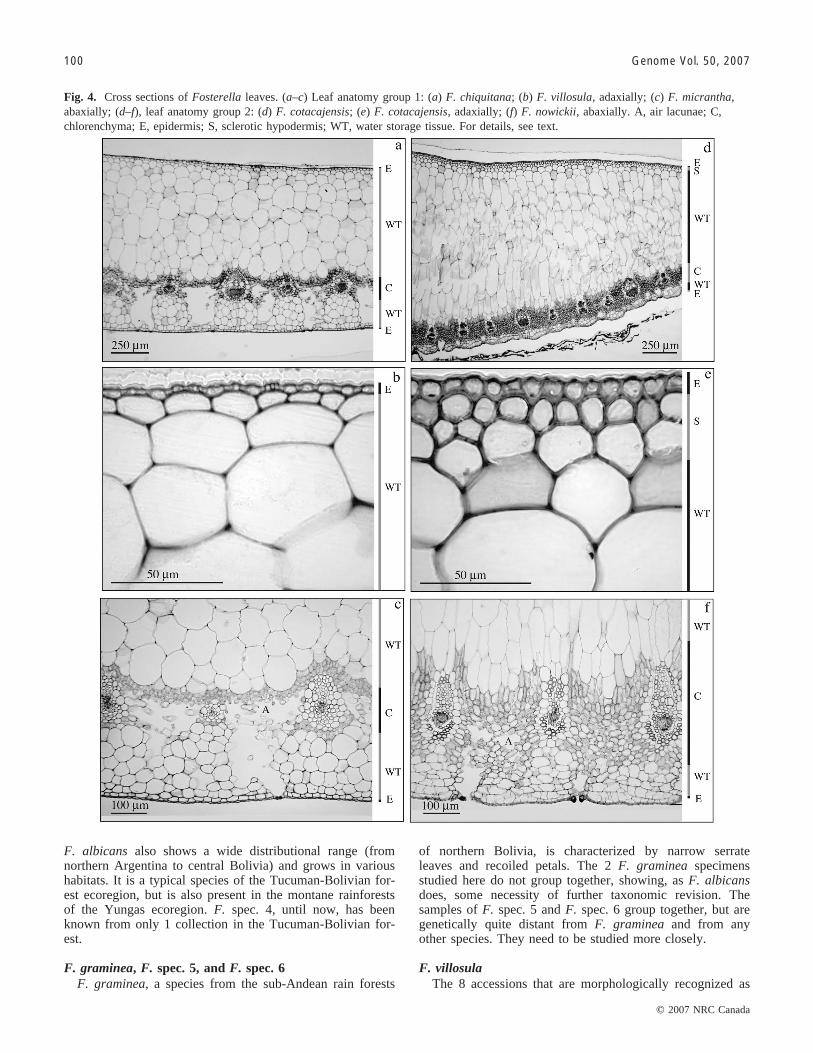
Leaf blade anatomyOne group of 5 species (F. villosula, F. micrantha,
F. penduliflora, F. latifolia, and F. chiquitana), referred toas leaf anatomy group 1, is characterized by a set of com-mon leaf anatomical characters (exemplified in Figs. 4a–4c): cell walls of the adaxial epidermis are uniform andhardly thickened; sclerotic hypodermis is adaxially absent(Fig. 4b); chlorenchyma consists of isodiametric cells only,and is sharply demarcated from the adaxial water storagetissue; vascular bundles (first order) are adaxially not adja-cent to water storage tissue and are embedded in chloren-chyma (Fig. 4c); aerenchyma is loosely packed; abaxialwater storage tissue forms very large cushion-like groups(Fig. 4c); and the abaxial epidermis is planar. With the ex-ception of F. micrantha, where no AFLP data are available,the species of leaf anatomy group 1 fully correspond to clus-ters J and K of the AFLP tree (see Fig. 2).
The remainder of the investigated species are anatomi-cally quite heterogeneous. However, a group of 3 species(F. nowickii, F. cotacajensis, F. weddelliana (leaf anatomygroup 2, exemplified in Figs. 4d–4f)) share a number ofcharacters, as follows: cell walls of the adaxial epidermisare uniform and slightly thickened; sclerotic hypodermis is
adaxially present (Fig. 4e); chlorenchyma consists of isodia-metric and palisade-like cells; chlorenchyma is not sharplydemarcated from the adaxial water storage tissue but isseparated from the latter by an area of poorly differentiatedcells; vascular bundles (first order) are adaxially adjacent towater storage tissue (Fig. 4f); aerenchyma is more denselypacked; abaxial water storage tissue forms small tomedium-sized cushion-like groups (Fig. 4f); and abaxialepidermis has slight depressions. The species of this groupare also closely related to each other in the AFLP tree,where they form cluster E (see Fig. 2).
To sum up, the diacritic leaf anatomical characters are:the shape and thickening of cells of the adaxial epidermis;the absence/presence of adaxial sclerotic hypodermis; theform of cells of the chlorenchyma; the demarcation of chlor-enchyma from the adaxial water storage tissue; the positionof vascular bundles within the mesophyll; and the form andsize of abaxial water storage tissue.
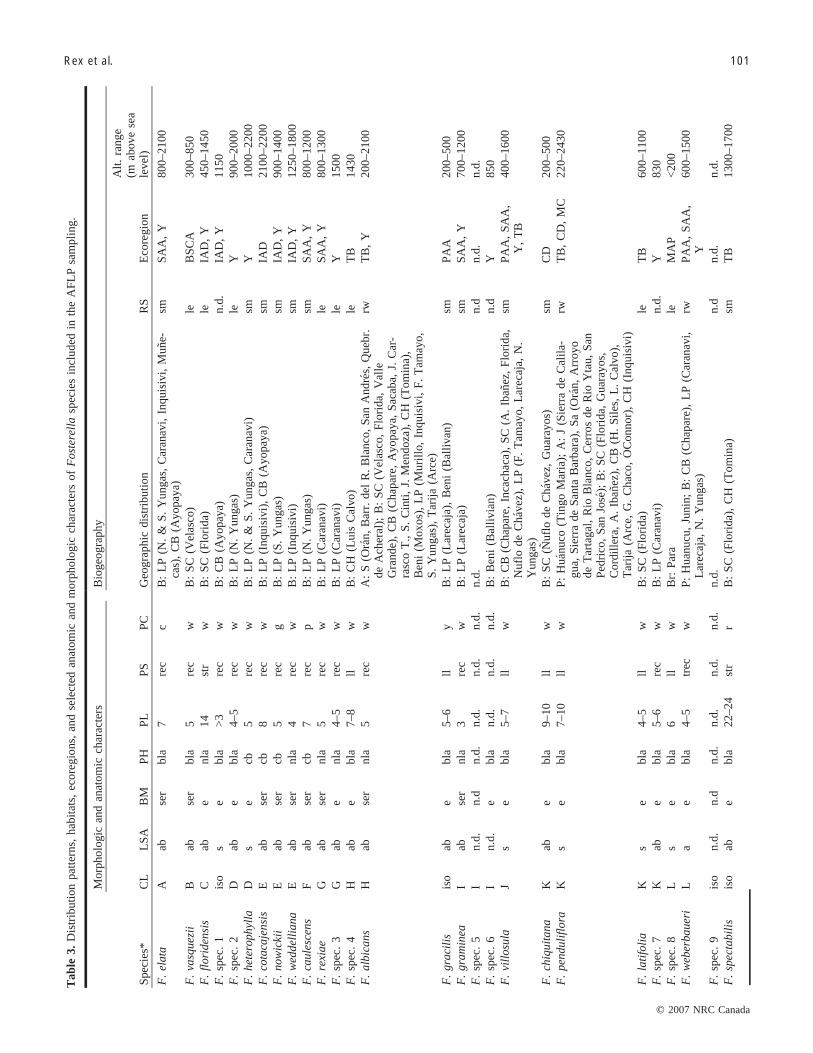
Morphology, ecology, and biogeographyTable 3 summarizes the distributional patterns and some
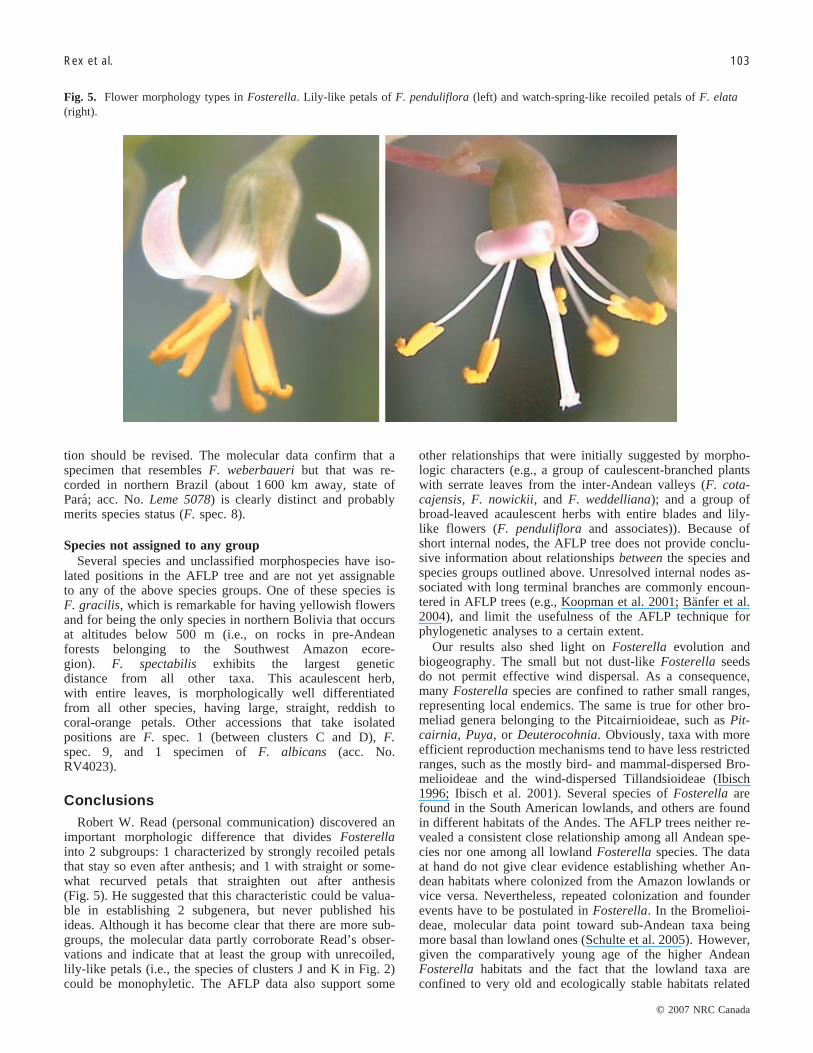
selected characters of Fosterella species that were includedin the AFLP sampling. Several groups and individual spe-cies can be discerned by morphology. One group comprisespurely Andean species, with a caulescent-branched habit,serrate leaves, and petals that are recoiled like watch springs(and that remain so even after anthesis) (Fig. 5, right panel).This group contains 4 species: F. cotacajensis, F. nowickii,F. weddelliana, and F. caulescens. It is morphologically re-lated to some Andean acaulescent species with serrate leavesand recoiled petals, such as F. rexiae and F. albicans. An-other group is formed by several species with more-or-lessbroad-leaved acaulescent flat rosettes, entire leaves, andlily-like petals, such as F. chiquitana, F. latifolia,F. penduliflora, and F. villosula (Fig. 5, left panel). Speciesthat are morphologically clearly distinct are F. floridensis,which are narrow-leaved, acaulescent, and have a more-or-less upright rosette and straight petals, and F. spectabilis,which have rather long and straight reddish-orange petals.All other Fosterella species have whitish or cream-colouredpetals (yellow in F. gracilis). F. spec. 8 is remarkable be-cause of its disjunct occurrence in the northern Amazon,but shows morphologic affinities to F. weberbaueri. TheFosterella species with the most disjunct distribution, F. mi-crantha from Central America, is morphologically similar toF. villosula. With 1 exception (i.e., the inter-Andean groupwith F. cotacajensis and others), morphologic groups are re-flected by neither geographic distribution nor by habitat andecology.
Discussion
Whereas the monophyly and generic circumscription ofFosterella have never been questioned, species boundarieshave been difficult to define, and little is known about infra-generic relationships. The current AFLP study generallysupports species concepts that have been based on morpho-logic characters, habitat preferences, and geographic distri-bution (Ibisch et al. 1997, 1999, 2002). Thus, with theexception of F. graminea, F. albicans, and the species ofthe ‘‘penduliflora’’ alliance, all investigated accessions
98 Genome Vol. 50, 2007
# 2007 NRC Canada
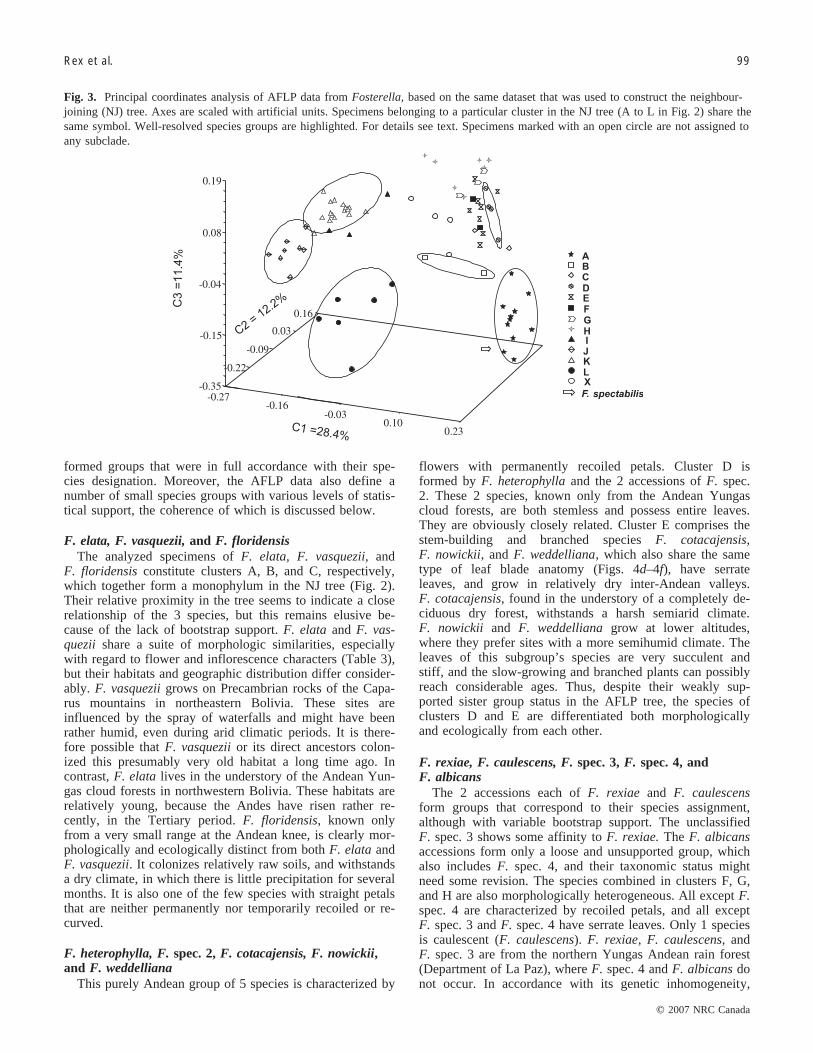
formed groups that were in full accordance with their spe-cies designation. Moreover, the AFLP data also define anumber of small species groups with various levels of statis-tical support, the coherence of which is discussed below.
F. elata, F. vasquezii, and F. floridensisThe analyzed specimens of F. elata, F. vasquezii, and
F. floridensis constitute clusters A, B, and C, respectively,which together form a monophylum in the NJ tree (Fig. 2).Their relative proximity in the tree seems to indicate a closerelationship of the 3 species, but this remains elusive be-cause of the lack of bootstrap support. F. elata and F. vas-quezii share a suite of morphologic similarities, especiallywith regard to flower and inflorescence characters (Table 3),but their habitats and geographic distribution differ consider-ably. F. vasquezii grows on Precambrian rocks of the Capa-rus mountains in northeastern Bolivia. These sites areinfluenced by the spray of waterfalls and might have beenrather humid, even during arid climatic periods. It is there-fore possible that F. vasquezii or its direct ancestors colon-ized this presumably very old habitat a long time ago. Incontrast, F. elata lives in the understory of the Andean Yun-gas cloud forests in northwestern Bolivia. These habitats arerelatively young, because the Andes have risen rather re-cently, in the Tertiary period. F. floridensis, known onlyfrom a very small range at the Andean knee, is clearly mor-phologically and ecologically distinct from both F. elata andF. vasquezii. It colonizes relatively raw soils, and withstandsa dry climate, in which there is little precipitation for severalmonths. It is also one of the few species with straight petalsthat are neither permanently nor temporarily recoiled or re-curved.
F. heterophylla, F. spec. 2, F. cotacajensis, F. nowickii,and F. weddelliana
This purely Andean group of 5 species is characterized by
flowers with permanently recoiled petals. Cluster D isformed by F. heterophylla and the 2 accessions of F. spec.2. These 2 species, known only from the Andean Yungascloud forests, are both stemless and possess entire leaves.They are obviously closely related. Cluster E comprises thestem-building and branched species F. cotacajensis,F. nowickii, and F. weddelliana, which also share the sametype of leaf blade anatomy (Figs. 4d–4f), have serrateleaves, and grow in relatively dry inter-Andean valleys.F. cotacajensis, found in the understory of a completely de-ciduous dry forest, withstands a harsh semiarid climate.F. nowickii and F. weddelliana grow at lower altitudes,where they prefer sites with a more semihumid climate. Theleaves of this subgroup’s species are very succulent andstiff, and the slow-growing and branched plants can possiblyreach considerable ages. Thus, despite their weakly sup-ported sister group status in the AFLP tree, the species ofclusters D and E are differentiated both morphologicallyand ecologically from each other.
F. rexiae, F. caulescens, F. spec. 3, F. spec. 4, andF. albicans
The 2 accessions each of F. rexiae and F. caulescensform groups that correspond to their species assignment,although with variable bootstrap support. The unclassifiedF. spec. 3 shows some affinity to F. rexiae. The F. albicansaccessions form only a loose and unsupported group, whichalso includes F. spec. 4, and their taxonomic status mightneed some revision. The species combined in clusters F, G,and H are also morphologically heterogeneous. All except F.spec. 4 are characterized by recoiled petals, and all exceptF. spec. 3 and F. spec. 4 have serrate leaves. Only 1 speciesis caulescent (F. caulescens). F. rexiae, F. caulescens, andF. spec. 3 are from the northern Yungas Andean rain forest(Department of La Paz), where F. spec. 4 and F. albicans donot occur. In accordance with its genetic inhomogeneity,
Fig. 3. Principal coordinates analysis of AFLP data from Fosterella, based on the same dataset that was used to construct the neighbour-joining (NJ) tree. Axes are scaled with artificial units. Specimens belonging to a particular cluster in the NJ tree (A to L in Fig. 2) share thesame symbol. Well-resolved species groups are highlighted. For details see text. Specimens marked with an open circle are not assigned toany subclade.
Rex et al. 99
# 2007 NRC Canada
F. albicans also shows a wide distributional range (fromnorthern Argentina to central Bolivia) and grows in varioushabitats. It is a typical species of the Tucuman-Bolivian for-est ecoregion, but is also present in the montane rainforestsof the Yungas ecoregion. F. spec. 4, until now, has beenknown from only 1 collection in the Tucuman-Bolivian for-est.
F. graminea, F. spec. 5, and F. spec. 6F. graminea, a species from the sub-Andean rain forests
of northern Bolivia, is characterized by narrow serrateleaves and recoiled petals. The 2 F. graminea specimensstudied here do not group together, showing, as F. albicansdoes, some necessity of further taxonomic revision. Thesamples of F. spec. 5 and F. spec. 6 group together, but aregenetically quite distant from F. graminea and from anyother species. They need to be studied more closely.
F. villosulaThe 8 accessions that are morphologically recognized as
Fig. 4. Cross sections of Fosterella leaves. (a–c) Leaf anatomy group 1: (a) F. chiquitana; (b) F. villosula, adaxially; (c) F. micrantha,abaxially; (d–f), leaf anatomy group 2: (d) F. cotacajensis; (e) F. cotacajensis, adaxially; (f) F. nowickii, abaxially. A, air lacunae; C,chlorenchyma; E, epidermis; S, sclerotic hypodermis; WT, water storage tissue. For details, see text.
100 Genome Vol. 50, 2007
# 2007 NRC Canada
Tab
le3.
Dis
trib
utio
npa
ttern
s,ha
bita
ts,
ecor
egio
ns,
and
sele
cted
anat
omic
and
mor
phol
ogic
char
acte
rsof
Fos
tere
lla
spec
ies
incl
uded
inth
eA
FLP
sam
plin
g.
Mor
phol
ogic
and
anat
omic
char
acte
rsB
ioge
ogra
phy
Spec
ies*
CL
LSA
BM
PHPL
PSPC
Geo
grap
hic
dist
ribu
tion
RS
Eco
regi
on
Alt.
rang
e(m
abov
ese
ale
vel)
F.
elat
aA
abse
rbl
a7
rec
cB
:L
P(N
.&
S.Y
unga
s,C
aran
avi,
Inqu
isiv
i,M
une-
cas)
,C
B(A
yopa
ya)
smSA
A,
Y80
0–21
00
F.
vasq
uezi
iB
abse
rbl
a5
rec
wB
:SC
(Vel
asco
)le
BSC
A30
0–85
0F
.fl
orid
ensi
sC
abe
nla
14st
rw
B:
SC(F
lori
da)
leIA
D,
Y45
0–14
50F
.sp
ec.
1is
os
ebl
a>
3re
cw
B:
CB
(Ayo
paya
)n.
d.IA
D,
Y11
50.
F.
spec
.2
Dab
ebl
a4–
5re
cw
B:
LP
(N.
Yun
gas)
leY
900–
2000
F.
hete
roph
ylla
Ds
ecb
5re
cw
B:
LP
(N.
&S.
Yun
gas,
Car
anav
i)sm
Y10
00–2
200
F.
cota
caje
nsis
Eab
ser
cb8
rec
wB
:L
P(I
nqui
sivi
),C
B(A
yopa
ya)
smIA
D21
00–2
200
F.
now
icki
iE
abse
rcb
5re
cg
B:
LP
(S.
Yun
gas)
smIA
D,
Y90
0–14
00F
.w
edde
llia
naE
abse
rnl
a4
rec
wB
:L
P(I
nqui
sivi
)sm
IAD
,Y
1250
–180
0F
.ca
ules
cens
Fab
ser
cb7
rec
pB
:L
P(N
.Y
unga
s)sm
SAA
,Y
800–
1200
F.
rexi
aeG
abse
rnl
a5
rec
wB
:L
P(C
aran
avi)
leSA
A,
Y80
0–13
00F
.sp
ec.
3G
abe
nla
4–5
rec
wB
:L
P(C
aran
avi)
leY
1500
.F
.sp
ec.
4H
abe
bla
7–8
llw
B:
CH
(Lui
sC
alvo
)le
TB
1430
.F
.al
bica
nsH
abse
rnl
a5
rec
wA
:S
(Ora
n,B
arr.
del
R.B
lanc
o,Sa
nA
ndre
s,Q
uebr
.de
Ach
eral
);B
:SC
(Vel
asco
,Fl
orid
a,V
alle
Gra
nde)
,C
B(C
hapa
re,
Ayo
paya
,Sa
caba
,J.
Car
-ra
sco
T.,
S.C
inti,
J.M
endo
za),
CH
(Tom
ina)
,B
eni
(Mox
os),
LP
(Mur
illo,
Inqu
isiv
i,F.
Tam
ayo,
S.Y
unga
s),
Tar
ija(A
rce)
rwT
B,
Y20
0–21
00
F.
grac
ilis
iso
abe
bla
5–6
lly
B:
LP
(Lar
ecaj
a),
Ben
i(B
alliv
an)
smPA
A20
0–50
0F
.gr
amin
eaI
abse
rnl
a3
rec
wB
:L
P(L
arec
aja)
smSA
A,
Y70
0–12
00F
.sp
ec.
5I
n.d.
n.d
n.d.
n.d.
n.d.
n.d.
n.d.
n.d
n.d.
n.d.
F.
spec
.6
In.
d.e
bla
n.d.
n.d.
n.d.
B:
Ben
i(B
alliv
ian)
n.d
Y85
0.F
.vi
llos
ula
Js
ebl
a5–
7ll
wB
:C
B(C
hapa
re,
Inca
chac
a),
SC(A
.Ib
anez
,Fl
orid
a,N
uflo
deC
have
z),
LP
(F.
Tam
ayo,
Lar
ecaj
a,N
.Y
unga
s)
smPA
A,
SAA
,Y
,T
B40
0–16
00
F.
chiq
uita
naK
abe
bla
9–10
llw
B:
SC(N
uflo
deC
have
z,G
uara
yos)
smC
D20
0–50
0F
.pe
ndul
iflo
raK
se
bla
7–10
llw
P:H
uanu
co(T
ingo
Mar
ia);
A:
J(S
ierr
ade
Cal
ila-
gua,
Sier
rade
Sant
aB
arba
ra),
Sa(O
ran,
Arr
oyo
deT
arta
gal,
Rio
Bla
nco,
Cer
ros
deR
ioY
tau,
San
Pedr
ico,
San
Jose
);B
:SC
(Flo
rida
,G
uara
yos,
Cor
dille
ra,
A.
Iban
ez),
CB
(H.
Sile
s,L
.C
alvo
),T
arija
(Arc
e,G
.C
haco
,O
Con
nor)
,C
H(I
nqui
sivi
)
rwT
B,
CD
,M
C22
0–24
30
F.
lati
foli
aK
se
bla
4–5
llw
B:
SC(F
lori
da)
leT
B60
0–11
00F
.sp
ec.
7K
abe
bla
5–6
rec
wB
:L
P(C
aran
avi)
n.d.
Y83
0..F
.sp
ec.
8L
se
bla
6ll
wB
r:Pa
rale
MA
P<
200.
F.
web
erba
ueri
La
ebl
a4–
5tr
ecw
P:H
uanu
cu,
Juni
n;B
:C
B(C
hapa
re),
LP
(Car
anav
i,L
arec
aja,
N.
Yun
gas)
rwPA
A,
SAA
,Y
600–
1500
F.
spec
.9
iso
n.d.
n.d
n.d.
n.d.
n.d.
n.d.
n.d.
n.d
n.d.
n.d.
F.
spec
tabi
lis
iso
abe
bla
22–2
4st
rr
B:
SC(F
lori
da),
CH
(Tom
ina)
smT
B13
00–1
700
Rex et al. 101
# 2007 NRC Canada

F. villosula form a well-supported group of acaulescentherbs with entire more-or-less broad leaves, lily-like nonre-coiled petals (Fig. 5, left panel), and a shared leaf anatomy(Figs. 4d–4f). Morphologic, leaf anatomical, and preliminarycpDNA sequence data suggest that F. micrantha from Cen-tral America also belongs to this group (see Figs. 4a–4c).The arising hypothesis is that F. micrantha is the product ofa relatively recent long-distance dispersal event from centralSouth America to the dry forests of El Salvador, westernGuatemala, and Mexico. F. villosula itself, as currently de-fined, possibly includes 2 distinct species. Some strikingmorphologic and ecologic intraspecific differences had beennoticed before (Ibisch et al. 2002), which have now beenconfirmed by the AFLP data. Thus, the specimens collectedclose to the type locality of F. villosula, in the highly humidrainforests of the Chapare region in central Bolivia, grouptogether, and so do the more southern specimens from semi-humid forests at the Andean knee near Santa Cruz. The mo-lecular and ecologic differences should be sufficient toestablish a new taxon for the southern specimens (e.g.,P. Ibisch & C. Nowicki 98.0173).
F. penduliflora, F. latifolia, F. chiquitana, and F. spec. 7This group of accessions is characterized by lily-like pet-
als and a common leaf blade anatomy that is shared withF. villosula and F. micrantha (Figs. 4d to 4f). Whereas thewhole group (i.e., cluster K in Fig. 2) is moderately sup-ported by AFLP data, its internal topology does not reflecttaxonomic boundaries. F. penduliflora is a widespread spe-cies, occurring from northern Argentina to central and east-ern Bolivia, in several ecoregions, such as the Tucuman-Bolivian forests, the montane Chaco, and the Chiquitanodry forest. Although seemingly a mesophytic herb, it with-stands considerable phases of drought. Under extremedrought, the plants can shed their leaves entirely, and thebulbous young shoots are able to endure several monthswithout precipitation. The species occurs in the understoryof Andean dry forests, and can survive on sun-exposed rockswith very limited water supply. F. chiquitana andF. latifolia were recently described as new taxa from thelowlands of the Chiquitano dry forest and from semihumidAndean forests, respectively (Ibisch et al. 1999). Their statusas separate species is not confirmed by the AFLP data, andmight require revision. If F. chiquitana and F. latifolia (andthe yet-unclassified F. spec. 7) are included inF. penduliflora, the latter taxon becomes even more variablethan previously thought. For the time being, we still con-sider F. latifolia to be a geographically and morphologicallydistinct taxon with a very small range.
F. weberbaueri and F. spec. 8The accessions combined in cluster L are acaulescent
herbs with entire leaves and only temporarily recoiled petalsthat become straight again after anthesis (see Ibisch et al.2002 for differentiation of F. weberbaueri from F. schido-sperma). They live in the pre- and sub-Andean rain forestsand even reach the montane Yungas forests. According tothe AFLP data, a specimen from northern La Paz (Nor Yun-gas province; acc. No. RS 261002-18) is clearly distinctfrom plants collected in the southern provinces of Caranavi,Larecaja, and Chapare. A potential morphologic differentia-T
able
3(c
oncl
uded
). Mor
phol
ogic
and
anat
omic
char
acte
rsB
ioge
ogra
phy
Spec
ies*
CL
LSA
BM
PHPL
PSPC
Geo
grap
hic
dist
ribu
tion
RS
Eco
regi
on
Alt.
rang
e(m
abov
ese
ale
vel)
Not
e:A
lt.,
altit
ude.
Mor
phol
ogic
alan
dan
atom
ical
char
acte
rs:
n.d.
,no
tde
term
ined
;C
L,
clus
ter;
iso,
isol
ated
posi
tion,
not
arra
nged
ina
clus
ter;
LSA
,le
afsc
ales
abax
iall
y(a
b,ab
unda
nt;
a,ab
sent
;s,
scar
ce);
BM
,bl
ade
mar
gin
(e,
entir
e,se
r,se
rrat
eat
the
base
);PH
,pl
ant
habi
t(b
la,
broa
d-le
aved
acau
lesc
ent
flat
rose
ttes;
nla,
narr
ow-l
eave
dac
aule
scen
t±
upri
ght
rose
ttes;
cb,
caul
esce
nt-b
ranc
hed)
;PL
,m
axim
alpe
tal
leng
th(m
m);
PS,
peta
lsh
ape
(rec
,re
coile
d,ev
enaf
ter
anth
esis
;ll,
lily-
like;
trec
,te
mpo
rari
lyre
coile
d,st
raig
htaf
ter
anth
esis
;st
r,st
raig
ht);
PC,
peta
lco
lor
(w,
whi
te;
r,re
ddis
hto
cora
l-or
ange
;c,
crea
man
dro
sesp
ots;
p,pa
legr
een,
then
slig
htly
redd
ish;
g,gr
eeni
sh;
y,ye
llow
).G
eogr
aphi
cdi
stri
butio
n(c
ount
ries
and
dist
rict
s):
B,
Bol
ivia
;P,
Peru
;B
r,B
razi
l;A
,A
rgen
tina;
J,Ju
juy;
LP,
La
Paz;
S,Sa
lta;
SC,
Sant
aC
ruz;
CB
,C
ocha
bam
ba;
CH
,C
huqu
isac
a.R
S,ra
nge
size
(le,
loca
lly
ende
mic
;sm
,sm
all;
rw,
rela
tivel
yw
ides
prea
d).
Eco
regi
ons:
BSC
A,
Sout
hwes
tA
maz
on/B
eni
and
Sant
aC
ruz
Am
azon
Fore
sts;
CD
,C
hiqu
itano
Dry
Fore
st;
IAD
,In
ter-
And
ean
Dry
Fore
sts;
MA
P,M
oist
Am
azon
Fore
sts
ofPa
ra;
PAA
,So
uthw
est
Am
azon
/Pre
-And
ean
Am
azon
Fore
sts;
SAA
,So
uthw
est
Am
azon
/Sub
-And
ean
Am
azon
Fore
sts;
TB
,T
ucum
an-B
oliv
ian
Fore
sts;
Y,
(Per
uvia
n-B
oliv
ian)
Yun
gas.
*Sp
ecie
sar
ear
rang
edac
cord
ing
toth
eir
occu
rren
ce(c
lust
ers
Ato
L)
inth
ene
ighb
our-
join
ing
tree
(see
Fig.
2).
102 Genome Vol. 50, 2007
# 2007 NRC Canada
tion should be revised. The molecular data confirm that aspecimen that resembles F. weberbaueri but that was re-corded in northern Brazil (about 1 600 km away, state ofPara; acc. No. Leme 5078) is clearly distinct and probablymerits species status (F. spec. 8).
Species not assigned to any groupSeveral species and unclassified morphospecies have iso-
lated positions in the AFLP tree and are not yet assignableto any of the above species groups. One of these species isF. gracilis, which is remarkable for having yellowish flowersand for being the only species in northern Bolivia that occursat altitudes below 500 m (i.e., on rocks in pre-Andeanforests belonging to the Southwest Amazon ecore-gion). F. spectabilis exhibits the largest geneticdistance from all other taxa. This acaulescent herb,with entire leaves, is morphologically well differentiatedfrom all other species, having large, straight, reddish tocoral-orange petals. Other accessions that take isolatedpositions are F. spec. 1 (between clusters C and D), F.spec. 9, and 1 specimen of F. albicans (acc. No.RV4023).
ConclusionsRobert W. Read (personal communication) discovered an
important morphologic difference that divides Fosterellainto 2 subgroups: 1 characterized by strongly recoiled petalsthat stay so even after anthesis; and 1 with straight or some-what recurved petals that straighten out after anthesis(Fig. 5). He suggested that this characteristic could be valua-ble in establishing 2 subgenera, but never published hisideas. Although it has become clear that there are more sub-groups, the molecular data partly corroborate Read’s obser-vations and indicate that at least the group with unrecoiled,lily-like petals (i.e., the species of clusters J and K in Fig. 2)could be monophyletic. The AFLP data also support some
other relationships that were initially suggested by morpho-logic characters (e.g., a group of caulescent-branched plantswith serrate leaves from the inter-Andean valleys (F. cota-cajensis, F. nowickii, and F. weddelliana); and a group ofbroad-leaved acaulescent herbs with entire blades and lily-like flowers (F. penduliflora and associates)). Because ofshort internal nodes, the AFLP tree does not provide conclu-sive information about relationships between the species andspecies groups outlined above. Unresolved internal nodes as-sociated with long terminal branches are commonly encoun-tered in AFLP trees (e.g., Koopman et al. 2001; Banfer et al.2004), and limit the usefulness of the AFLP technique forphylogenetic analyses to a certain extent.
Our results also shed light on Fosterella evolution andbiogeography. The small but not dust-like Fosterella seedsdo not permit effective wind dispersal. As a consequence,many Fosterella species are confined to rather small ranges,representing local endemics. The same is true for other bro-meliad genera belonging to the Pitcairnioideae, such as Pit-cairnia, Puya, or Deuterocohnia. Obviously, taxa with moreefficient reproduction mechanisms tend to have less restrictedranges, such as the mostly bird- and mammal-dispersed Bro-melioideae and the wind-dispersed Tillandsioideae (Ibisch1996; Ibisch et al. 2001). Several species of Fosterella arefound in the South American lowlands, and others are foundin different habitats of the Andes. The AFLP trees neither re-vealed a consistent close relationship among all Andean spe-cies nor one among all lowland Fosterella species. The dataat hand do not give clear evidence establishing whether An-dean habitats where colonized from the Amazon lowlands orvice versa. Nevertheless, repeated colonization and founderevents have to be postulated in Fosterella. In the Bromelioi-deae, molecular data point toward sub-Andean taxa beingmore basal than lowland ones (Schulte et al. 2005). However,given the comparatively young age of the higher AndeanFosterella habitats and the fact that the lowland taxa areconfined to very old and ecologically stable habitats related
Fig. 5. Flower morphology types in Fosterella. Lily-like petals of F. penduliflora (left) and watch-spring-like recoiled petals of F. elata(right).
Rex et al. 103
# 2007 NRC Canada

to different kinds of pre-Cambric rocks (e.g., F. chiquitana,F. hatschbachii, F. penduliflora, F. rojasii, F. windischii,F. vasquezii, and F. yuvinkae), a lowland origin can be hy-pothesized. Apparently, the genus also reached remote re-gions, such as Central America, by rare events of long-distance dispersal, perhaps through seeds attached to birds.
Further interdisciplinary studies on diversity, biogeogra-phy, and phylogeny of this genus seem promising. The re-sults will likely facilitate further taxonomic treatment byconfirming hypotheses about new taxa and the genetic dif-ferentiation according to geographic provenances.
AcknowledgementsThe authors acknowledge the support of the Deutsche
Forschungsgemeinschaft (DFG grants We 1830/5-1, IB 85/1-1 and ZI 557/6-1). Part of the plant material was kindlysupplied by the Palmengarten Frankfurt am Main, the Bota-nical Gardens of Heidelberg, Berlin and Osnabruck; theMarie Selby Botanical Garden of Sarasota, Fla., and by theliving collection of FAN, where Arturo Osinaga provided in-valuable help. We also thank Elton Leme for support, andChristoph Nowicki, Robert Muller, and Jule Peters for helpwith the data and tables.
ReferencesBanfer, G., Fiala, B., and Weising, K. 2004. AFLP analysis of phy-
logenetic relationships among myrmecophytic species of Macar-anga (Euphorbiaceae) and their allies. Plant Syst. Evol. 249:213–231. doi:10.1007/s00606-004-0219-4.
Barfuss, M.H.J., Samuel, M.R., and Till, W. 2004. Molecular phy-logeny in subfamily Tillandsioideae (Bromeliaceae) based on sixcpDNA markers: an update. J. Bromeliad Soc. 54: 8–16.
Benzing, D.H. 2000. Bromeliaceae: profile of an adaptive radiation.Cambridge University Press, Cambridge.
Brummit, R.K., and Powell, C.E. 1992. Authors of plant names.Royal Botanic Gardens, Kew. Whitstable Litho Ltd., Kent.
Crayn, D.M., Winter, K., and Smith, J.A.C. 2004. Multiple origins ofcrassulacean acid metabolism and the epiphytic habit in the Neo-tropical family Bromeliaceae. Proc. Natl. Acad. Sci. U.S.A. 101:3703–3708. doi:10.1073/pnas.0400366101. PMID:14982989.
Debener, T., and Mattiesch, L. 1999. Construction of a geneticlinkage map for roses using RAPD and AFLP markers. Theor.Appl. Genet. 99: 891–899. doi:10.1007/s001220051310.
Despres, L., Gielly, L., Redoutet, B., and Taberlet, P. 2003. UsingAFLP to resolve phylogenetic relationships in a morphologicallydiversified plant species complex when nuclear and chloroplastsequences fail to reveal variability. Mol. Phylogenet. Evol. 27:185–196. PMID:12695084.
Felsenstein, J. 1985. Confidence limits on phylogenies: an ap-proach using the bootstrap. Evolution, 39: 783–791. doi:10.2307/2408678.
Givnish, T.J., Millam, K.C., Evans, T.M., Hall, J.C., Pires, J.C.,Berry, P.E., and Sytsma, K.J. 2004. Ancient vicariance or recentlong-distance dispersal? Inferences about phylogeny and SouthAmerican-African disjunctions in Rapateaceae and Bromeliaceaebased on ndhF sequence data. Int. J. Plant Sci. 165: S35–S54.doi:10.1086/421067.
Givnish, T.J., Millam, K.C., Evans, T.M., Pires, J.C., Berry, P.E.,and Sytsma, K.J. 2007. Phylogeny, biogeography, and ecologicalevolution in Bromeliaceae: insights from ndhF sequences. InMonocots: comparative biology and evolution. Edited by J.T.
Columbus, E.A. Friar, C.W. Hamilton, J.M. Porter, L.M. Prince,and M.G. Simpson. In press.
Hansen, M., Kraft, T., Christiansson, M., and Nilsson, N.O. 1999.Evaluation of AFLP in Beta. Theor. Appl. Genet. 98: 845–852.doi:10.1007/s001220051143.
Hodkinson, T.R., Renvoize, S.A., Chonghaile, G.N., Stapleton,C.M.A., and Chase, M.W. 2000. A comparison of ITS nuclearrDNA sequence data and AFLP markers for phylogenetic studiesin Phyllostachys (Bambusoideae, Poaceae). J. Plant Res. 113:259–269. doi:10.1007/PL00013936.
Horres, R., Zizka, G., Kahl, G., and Weising, K. 2000. Molecularphylogenetics of Bromeliaceae: evidence from trnL(UAA) in-tron sequences of the chloroplast genome. Plant Biol. 2: 306–315. doi:10.1055/s-2000-3700.
Horres, R., Schulte, K., Weising, K., and Zizka, G. 2007. Systema-tics of Bromelioideae (Bromeliaceae) - molecular, anatomicaland morphological Studies. In Monocots: comparative biologyand evolution. Edited by J.T. Columbus, E.A. Friar, C.W. Ha-milton, J.M. Porter, L.M. Prince, and M.G. Simpson, M.G. Inpress.
Ibisch, P.L. 1996. Neotropische Epiphytendiversitat - das BeispielBolivien. Martina-Galunder-Verlag, Wiehl.
Ibisch, P.L., Gross, E., Rauer, G., and Rudolph, D. 1997. On thediversity and biogeography of the genus Fosterella L.B. Smith(Bromeliaceae) with the description of a new species from East-ern Bolivia. J. Bromel. Soc. 47: 211–217.
Ibisch, P.L., Vasquez, R., and Gross, E. 1999. More novelties ofFosterella L.B. Smith (Bromeliaceae, Pitcairnioideae) from Bo-livia. Rev. Soc. Boliv. Bot. 2: 117–132.
Ibisch, P.L., Nowicki, C., and Vasquez, R. 2001. Towards an un-derstanding of diversity patterns and conservation requirementsof the Bolivian Bromeliaceae. J. Bromel. Soc. 51: 99–113.
Ibisch, P.L., Vasquez, R., Gross, E., Kromer, T., and Rex, M. 2002.Novelties in Bolivian Fosterella (Bromeliaceae). Selbyana, 23:204–219.
Ibisch, P.L., Beck, S.G., Gerkmann, B., and Carretero, A. 2004.Ecoregions and ecosystems. In Biodiversity: the richness of Bo-livia. State of knowledge and conservation. Edited by P.L. Ibischand G. Merida. Ministerio de Desarrollo Sostenible y Planifica-cion/Editorial FAN, Santa Cruz. pp. 47–88.
Igersheim, A., and Cichocki, O. 1996. A simple method for micro-tome sectioning of prehistorical charcoal specimens, embeddedin 2-hydroxyethyl methacrylate (HEMA). Rev. Palaeobot. Paly-nol. 92: 389–393.
Kardolus, J.P., van Eck, H.J., and van den Berg, R.G. 1998. Thepotential of AFLPs in biosystematics: a first application in Sola-num taxonomy (Solanaceae). Plant Syst. Evol. 210: 87–103.doi:10.1007/BF00984729.
Kessler, M., Ibisch, P.L., and Gross, E. 1999. Fosterella cotacajen-sis, una nueva especie de Bromeliaceae de los valles secos andi-nos de Bolivia. Rev. Soc. Boliv. Bot. 2: 111–116.
Koopman, W.J.M., Zevenbergen, M.J., and van den Berg, R.G.2001. Species relationships in Lactuca s.l. (Lactuceae, Astera-ceae) inferred from AFLP fingerprints. Am. J. Bot. 88: 1881–1887.
Kromer, T., Kessler, M., Holst, B.K., Luther, H.E., Gouda, E.J.,Ibisch, P.L., Till, W., and Vasquez, R. 1999. Checklist of Boli-vian Bromeliaceae with notes on species distribution and levelsof endemism. Selbyana, 20: 201–223.
Luther, H.E. 1981. Miscellaneous new taxa of Bromeliaceae (I).Selbyana, 5: 310–311.
Luther, H.E. 1997. A showy new Fosterella from Bolivia. J. Bro-mel. Soc. 47: 118–119.
Nei, M., and Li, W.-H. 1979. Mathematical model for studying ge-
104 Genome Vol. 50, 2007
# 2007 NRC Canada

netic variation in terms of restriction endonucleases. Proc. Natl.Acad. Sci. U.S.A. 76: 5269–5273. doi:10.1073/pnas.76.10.5269.PMID:291943.
Rauh, W. 1979. Bromelienstudien. I. Neue und wenig bekannte Ar-ten aus Peru und anderen Landern (9.Mitteilung). 4. Fosterella.Trop. Subtrop. Pflanzenwelt, 31: 23–29.
Rauh, W. 1987. Bromelienstudien. 4. Fosterella L.B. Smith. Trop.Subtrop. Pflanzenwelt, 60: 24–26.
Rex, M. 2001. Molekulare Analyse von Verwandtschaftsbeziehun-gen in ausgewahlten Gattungen der Pitcairnioideae (Bromelia-ceae). Diploma Thesis, University of Kassel.
Rohlf, F.J. 2000. NTSYSpc version 2.10p. Numerical taxonomyand multivariate analysis system. Exeter, New York.
Saitou, N., and Nei, M. 1987. The neighbour joining method: anew method for reconstructing phylogenetic trees. Mol. Biol.Evol. 4: 406–425. PMID:3447015.
Sambrook, J., and Russell, D.W. 2001. Molecular Cloning. A La-boratory Manual. 3rd ed. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, New York.
Schulte, K., Horres, R., and Zizka, G. 2005. Molecular phylogenyof Bromelioideae and its implications on biogeography and theevolution of CAM in the family. Senckenb. Biol. 85: 113–125.
Smith, L.B., and Downs, R.J. 1974. 13. Fosterella. In Flora Neotro-pica, Monograph no. 14, Pitcairnioideae. Hafner Press, NewYork. pp. 199–209.
Smith, L.B., and Read, R.W. 1992. Flora Neotropica, MonographNo. 14 (1), suppl. no. 3. Fosterella. Bradea, 6: 134–140.
Smith, L.B., and Till, W. 1998 Bromeliaceae. In The Families andGenera of Vascular Plants. Edited by K. Kubitzki. Vol. 4.Springer, Berlin, Heidelberg, New York. pp. 74–99.
Sneath, P.H.A., and Sokal, R.O. 1973. Numerical Taxonomy. Free-man, San Francisco.
Swofford, D.L. 2002. PAUP*: Phylogenetic analysis using parsi-mony, (*and other methods) v. 4.0b10 [computer program]. Si-nauer Assoc., Sunderland.
Terry, R.G., Brown, G.K., and Olmstead, R.G. 1997. Examinationof subfamilial phylogeny in Bromeliaceae using comparative se-quencing of the plastid locus NDHF. Am. J. Bot. 84: 664–670.doi:10.2307/2445903.
Tuthill, D., and Brown, G.K. 2004. Bromeliad endophytes and theserendipity of science. J. Bromeliad Soc. 54: 28–32.
Van de Peer, Y., and De Wachter, R. 1994. TREECON for Win-dows: a software package for the construction and drawing ofevolutionary trees for the Microsoft Windows environment.Comput. Appl. Biosci. 10: 569–570. PMID:7828077.
Vos, P., Hogers, R., Bleeker, M., Reijans, M., Van de Lee, T.,Hornes, M., et al. 1995. AFLP: a new technique for DNA fin-gerprinting. Nucleic Acids Res. 23: 4407–4414. PMID:7501463.
Weising, K., Nybom, H., Wolff, K., and Meyer, W. 2005. DNAFingerprinting in Plants: Principles, Methods, and Applications.2nd ed. CRC Press, Taylor & Francis Group, Boca Raton, Fla.
Rex et al. 105
# 2007 NRC Canada
Related Documents











