Adenovirus Mediated Alpha Interferon (IFN-α) Gene Transfer into CD34 + Cells and CML Mononuclear Cells E. FELDMAN, a T. AHMED, a J.D. LUTTON, a T. FARLEY, a K. TANI, b M. FREUND, c S. ASANO, b N.G. ABRAHAM a a Departments of Pharmacology and Medicine, New York Medical College, Valhalla, New York, USA; b The Institute of Medical Science, The University of Tokyo, Tokyo, Japan; c University of Rostock, Division of Hematology and Oncology, Rostock, Germany Key Words. Gene transfer · α interferon · Leukemia · Stem cells ABSTRACT Gene transfer or gene therapy has advantages in the treatment of a variety of disorders due to its selective expression within specific mammalian cells. Interferon-α (IFN-α) has been used in the management of leukemia but its diverse adverse activities with multiple potential side effects, possibly unrelated to therapeutic targets, may neg- atively influence the ability of IFN-α to treat this disorder. Therefore, we examined the ability of adenovirus (Ad)- IFN-α gene construct to transfect normal (CD34 + cells) and chronic myelogenous leukemia (CML) bone marrow mononuclear cells (BMMNC) and the transient overex- pression of IFN-α in these cells. Ad-cytomegalovirus pro- moter driven IFN-α (AdCMV-IFN-α29 at multiple doses was assessed to transfect highly purified CD34 + cells in liq- uid culture, and optimal transduction of CD34 + cells was achieved using 120 plaque forming units. Flow cytometric determinations revealed that there was no significant dif- ference in cell viability for the 4 h or 24 h transfection periods. Immunoassay of IFN-α produced by CD34 + cells shows that IFN-α levels increased several fold in transfected cells. Transient expression of the IFN-α gene did not suppress proliferation of CD34 + progeni- tors as indicated by BFU-E or colony forming units- granulocyte-macrophage (CFU-GM) growth. Reverse transcriptase/polymerase chain reaction analysis of RNA from CD34 + harvested CFU-GM progenitor cells demonstrated transient IFN-α mRNA expression. Similarly, CML BMMNC were transfected with AdCMV-IFN-α under similar conditions as described for CD34 + cells. BMMNC cells exposed to adenovirus for 24 h and 48 h were found to express IFN-α at a sub- stantial level. This in vitro data suggest that Ad-medi- ated gene transfer of IFN-α into hematopoietic stem cells can be achieved and that the IFN-α gene can be translated into its specific mRNA in CD34 progenitor cells. Stem Cells 1997;15:386-395 STEM CELLS 1997;15:386-395 Correspondence: Dr. Nader G. Abraham, Department of Pharmacology, New York Medical College, Valhalla, NY 10595, USA. Accepted for publication June 9, 1997. ©AlphaMed Press 1066-5099/97/$5.00/0 INTRODUCTION Recently, this laboratory has utilized retrovirus vectors as vehicles for gene transfer into a variety of cells including hematopoietic stem cells in vitro and in vivo [1]. Furthermore, we have established novel conditions in which stromal adher- ent cells permit high efficiency of retrovirus-mediated gene transfer of adenosine deaminase into bone marrow (BM) hematopoietic progenitor cells [1]. Successful transplantation of the genes was further documented by their appearance in hematopoietic clones of cells from animals many months after reconstitution. Over the past several years, investigators have begun to apply retroviral gene transfer technology directed at hematopoietic stem cells in preliminary human clinical trials [2]. Recombinant adenoviruses are the second most com- monly used viral vector for gene delivery [3-4], thus mini- mizing the risk of permanently altering the cellular genotype or of insertional mutagenesis. Adenoviruses are preferentially used when the transient expression of a transgene is suffi- cient to achieve the therapeutic goals [5-6]. Investigators found that adenovirus vectors can infect various BM hematopoietic cells including early progenitors, and that transduced cells are functionally normal in vitro. Several pro- cedures have been reported using different exposure times with adenovirus vectors for in vitro gene transduction [7-8].

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Adenovirus Mediated Alpha Interferon (IFN-α) Gene Transfer into CD34+ Cells and CML Mononuclear Cells
E. FELDMAN ,a T. AHMED,a J.D. LUTTON,a T. FARLEY ,a K. TANI ,b M. FREUND,c S. ASANO,b N.G. ABRAHAM a
aDepartments of Pharmacology and Medicine, New York Medical College, Valhalla, New York, USA;bThe Institute of Medical Science, The University of Tokyo, Tokyo, Japan;
cUniversity of Rostock, Division of Hematology and Oncology, Rostock, Germany
Key Words. Gene transfer · α interferon · Leukemia · Stem cells
ABSTRACT
Gene transfer or gene therapy has advantages in thetreatment of a variety of disorders due to its selectiveexpression within specific mammalian cells. Interferon-α(IFN-α) has been used in the management of leukemia butits diverse adverse activities with multiple potential sideeffects, possibly unrelated to therapeutic targets, may neg-atively influence the ability of IFN-α to treat this disorder.Therefore, we examined the ability of adenovirus (Ad)-IFN-α gene construct to transfect normal (CD34+ cells)and chronic myelogenous leukemia (CML) bone marrowmononuclear cells (BMMNC) and the transient overex-pression of IFN-α in these cells. Ad-cytomegalovirus pro-moter driven IFN-α (AdCMV-IFN- α) at multiple doseswas assessed to transfect highly purified CD34+ cells in liq-uid culture, and optimal transduction of CD34+ cells wasachieved using 120 plaque forming units. Flow cytometricdeterminations revealed that there was no significant dif-ference in cell viability for the 4 h or 24 h transfection
periods. Immunoassay of IFN-α produced by CD34+
cells shows that IFN-α levels increased several fold intransfected cells. Transient expression of the IFN-αgene did not suppress proliferation of CD34+ progeni-tors as indicated by BFU-E or colony forming units-granulocyte-macrophage (CFU-GM) growth. Reversetranscriptase/polymerase chain reaction analysis ofRNA from CD34+ harvested CFU-GM progenitor cellsdemonstrated transient IFN-α mRNA expression.Similarly, CML BMMNC were transfected withAdCMV-IFN- α under similar conditions as describedfor CD34+ cells. BMMNC cells exposed to adenovirusfor 24 h and 48 h were found to express IFN-α at a sub-stantial level. This in vitro data suggest that Ad-medi-ated gene transfer of IFN-α into hematopoietic stemcells can be achieved and that the IFN-α gene can betranslated into its specific mRNA in CD34 progenitorcells. Stem Cells1997;15:386-395
STEM CELLS 1997;15:386-395
Correspondence: Dr. Nader G. Abraham, Department of Pharmacology, New York Medical College, Valhalla, NY 10595,USA. Accepted for publication June 9, 1997. ©AlphaMed Press 1066-5099/97/$5.00/0
INTRODUCTION
Recently, this laboratory has utilized retrovirus vectors asvehicles for gene transfer into a variety of cells includinghematopoietic stem cells in vitro and in vivo [1]. Furthermore,we have established novel conditions in which stromal adher-ent cells permit high efficiency of retrovirus-mediated genetransfer of adenosine deaminase into bone marrow (BM)hematopoietic progenitor cells [1]. Successful transplantationof the genes was further documented by their appearance inhematopoietic clones of cells from animals many months afterreconstitution. Over the past several years, investigators havebegun to apply retroviral gene transfer technology directed at
hematopoietic stem cells in preliminary human clinical trials[2]. Recombinant adenoviruses are the second most com-monly used viral vector for gene delivery [3-4], thus mini-mizing the risk of permanently altering the cellular genotypeor of insertional mutagenesis. Adenoviruses are preferentiallyused when the transient expression of a transgene is suffi-cient to achieve the therapeutic goals [5-6]. Investigatorsfound that adenovirus vectors can infect various BMhematopoietic cells including early progenitors, and thattransduced cells are functionally normal in vitro. Several pro-cedures have been reported using different exposure timeswith adenovirus vectors for in vitro gene transduction [7-8].

Feldman, Ahmed, Lutton et al. 387
The current paper investigates the development of genetransfer vectors and assesses the effect of adenovirus-mediatedgene transfer into CD34+ cells. We have previously utilized anadenovirus to mediate gene transfer of heme oxygenase intononhematopoietic stem cells [9]. A similar recombinant aden-oviral vector was used for interferon-α (IFN-α) gene transfer.The viral construct with the cytomegalovirus (CMV) promoterwas also described by Ohwada et al. [10] and used for transferof thrombopoietin into hematopoietic stem cells. A similarmetabolic approach to construct adenovirus CMV-IFN-α genewas used to transfect both normal and chronic myelogenousleukemia (CML) hematopoietic stem cells. Successful aden-ovirus-CMV promoter driven IFN-α (Ad-CMV-IFN-α) genetransfer into CD34+ cells is monitored by Southern blot analy-sis and Northern blot analysis of genomic DNA and RNA,respectively, from the CD34+ cells and from hematopoieticclones, BFU-E and colony forming units-granulocyte-macrophage (CFU-GM) seeded by transfected CD34+ cells.The results allowed us to determine the feasibility of develop-ing an adenovirus-mediated gene transfer into CD34+ cells.Since IFN-α has been used in the management of tumorgrowth and treatment of CML, we decided to use CML cellsas a model tumor system for IFN-α gene transfer.
CML arises from a clonal expansion of a transformedstem cell capable of differentiation into mature hematopoieticcells [11-13]. The Philadelphia (Ph) chromosome is charac-teristic of the disease in which the reciprocal translocationt(9;22) results in the creation of a chimeric bcr/ablgene whichplays a central role in leukemogenesis [14, 15]. Induction ofclonal expansion associated with bcr/abl expression may bedue in part to increased tyrosine kinase activity [16]. In addi-tion, CD34+ marrow cells from patients with CML respond tocolony-stimulating factors, but their adhesion to the stroma isimpaired, resulting in a loss of sensitivity to stromal inhibitorysignals [17-21].
Numerous clinical trails have documented the ability ofIFN to induce hematologic and cytogenetic remissions inpatients with CML [22]. A number of in vitro effects ofIFN-α on CML cells and stroma have been well docu-mented, including inhibition of CML marrow progenitorgrowth [23, 24], restoration of the adherence to stroma [25-27], and regulation of stromal cytokine production [28] andcellular immune surveillance [29-33] important in the controlof growth of the leukemic clone in CML.
The systemic administration of IFN-α following cyto-toxic therapy has been disappointing, due in part to theinability of patients to tolerate the exogenous doses required.It is possible that endogenous production of IFN-α by thehost may be more effective at lower levels as compared tothe levels required exogenously. One possible means ofimproving the efficacy of IFN-α is to deliver the agent as a
gene directly into the hematopoietic progenitor cells that arereinfused (following ablative therapy) by means of a viralvector in which one could achieve efficient and constantexpression of the gene or product over a specific period oftime. In this way, overexpression of IFN-α may provideantileukemic effects that suppress CML growth beyondwhat is possible with systemic administration.
The present report is concerned with the development ofefficient adenovirus-mediated gene transfer of the IFN-αgene into hematopoietic CD34+ cells and also CML bonemarrow mononuclear cells (BMMNCs) in vitro. Such in vitrostudies may then provide insight on how to effectively deliverthe IFN-α gene into cells with possible therapeutic potential.
MATERIALS AND METHODS
The replication-deficient adenovirus vector encodingIFN-α (AdCMV-IFN-α) was constructed in our laboratory,as described previously [9]. The recombinant adenoviral vec-tor was constructed by hemologous recombination betweenthe E3 deleted adenovirus, dL700 1 and the plasmid contain-ing the human IFN-α. The AdCMV-IFN-α plasmid was con-structed as follows: Xho1-Xho1 fragment of the humanIFN-α cDNA deleted from the BMGNeo IFN-α plasmid wasinserted into the hind III site of the pRC/CMV (Invitrogen;San Diego, CA) to obtain CMV-IFN-α. A similar approachwas used in our laboratory with expression of the vector [9].The Nru I-Bam HI fragment containing CMV-IFN-α wasinserted into the Stu 1-Bam H1 sites of the pBacPac 8(Clontech; Palo Alto, CA) to obtain pBacPac 8-CMV-IFN-α.The Bam HI-Bgl III fragment containing CMV-IFN-α frompBacPac8-CMV-IFN-α was inserted into the Bgl II of thepAdv.Bgl II plasmid to obtain the AdCMV-IFN-α. A humanembryonic cell line that expressed E1 (293 cells ATCC #1573- CRL) was cotransfected with 10 mg of ECOR I-digestedAdCMV-IFN-α and 1 mg of Cla I-digested dL7001 DNAusing a mammalian transfection kit (Stratagene; La Jolla,CA). The virus was replicated and encapsidated into aninfectious virus. After five days, plaque locations weremarked on the plate, and the resultant cytopathic effect tothe monolayer was observed microscopically until theplaque reached an adequate size (usually one week). Theplaques were purified by ultracentrifugation throughcesium chloride gradients and checked for the presence ofhuman IFN-α by polymerase chain reaction (PCR) using IFN-α specific primers. The primers for IFN-α are 5′CAG TTCCAG AAG GCT CAA GC3′ and 5′ACC TCC TGC ATCATA CAG GC3′. These primers give a PCR product with asize of 222 bp. The virus was released from infected cells twodays after infection by five freeze-thaw cycles, concentratedby centrifugation using Ultrafree-MC filters (Millipore;Bedford, MA), and dialyzed against phosphate-buffered

388 Gene Transfer in Leukemia
saline (PBS). Titers of viral stocks were determined byplaque assay using 293 cells as previously described [9, 34].Titers of the viral stocks used for transfection were 1 to 5 ×1010 infectious units/ml. Cells were incubated in triplicatedishes with AdCMV-IFN-α at 40, 80, 120, 160 and 200plaque forming units (pfu)/cell. The viral stock was diluted1:1 with glycerol and stored at –20°C. Concentration of thevirus was determined by measuring its optical density (OD)at 260 nm. Viral particle content was then calculated from itsOD at 260 nm under the assumption that 1 OD equals 1 ×1012 viral particles/ml.
BM SamplesHeparinized BM samples were aspirated from the pos-
terior iliac crest of normal volunteers or CML patients afterinformed consent was obtained according to the guidelinesestablished by the Human Investigation Committee of NewYork Medical College. Specimens of normal and CML BMcells were also obtained from the National Disease ResearchInterchange (The National Resources Center). Low-densityBMMNC were isolated by density centrifugation ofheparinized marrow layered over Ficoll-Paque (PharmaciaFine Chemicals; Piscataway, NJ) at 500 g for 25 min.
Selection of Purified Progenitor PopulationsBMMNC were prepared by sequential counterflow cen-
trifugation elutriation [35], sheep erythrocyte rosetting [36]and immunomagnetic bead depletion [37]. Alternatively,CD34+ cells were selected from BMMNC using avidin-biotinimmunoadsorbtion columns (CellPro Inc; Bothell, WA) [38].The selected populations were labeled with anti-CD34-phy-coerythrin and anti-HLA-DR-fluorescein isothiocyanate anti-bodies (Becton-Dickinson; San Jose, CA) and sorted on anEPICS Elite II laser flow cytometry system (Coulter; Hialeah,FL). Cells were selected for low vertical and horizontal lightscatter properties and for expression of CD34 and HLA-DRantigens based on isotype control stains [37, 39]. In somecases, purified CD34+ cells were obtained from the HudsonValley Blood Services (Valhalla, NY).
Transfection Protocol for CD34+ Cells and BMMNC inLiquid Culture
CD34+ cells were resuspended in Iscove’s modifiedDulbecco’s medium (IMDM) containing 0.5% fetal bovineserum in T-25 flasks supplemented with cytokines (interleukin6 [recombinant human interleukin 6, 50 U/ml]; stem cell factor[recombinant human stem cell factor, 100 ng/ml]; interleukin 3;5 ng/ml, G-CSF [50 U/ml]; GM-CSF [50 U/ml] [Amgen;Thousand Oaks, CA]; and megakaryocyte growth factor [10U/ml] [Immunex; Seattle, WA]). Various pfu/cell (20, 40, 80,120, 160 and 240) of AdCMV-IFN-α were addeddirectly to
the cultures of CD34+. The cocultivation of adenovirus withCD34+ was continued for 4, 8, 12, 24 and 48 h. After eachincubation time, samples were removed for measurement ofcell viability by propidium iodide (PI) staining or for clonalefficiency in methylcellulose cultures. IFN-α mRNA levelswere assessed on CD34+ samples by reverse transcription(RT)/PCR using human specific primers to yield a PCRproduct of 222 bp. A similar protocol was followed for trans-fection of progenitor cells obtained from CML donors. CMLBMMNC from fresh leukapheresis samples were separatedby counterflow centrifugation and sheep erythrocyte roset-ting as described above. BMMNC were then plated in T-25flasks containing IMDM and 0.5% fetal calf serum and ade-novirus was added to the flasks at 120-300 pfu/cell for 24 h.After this time period, samples were removed and assessedfor clonal efficiency and expression of IFN-α mRNA inCFU-GM and BFU-E after 12 and 14 days.
Human IFN-αThe presence of IFN-α in CD34+ cell media was deter-
mined using a commercially available quantitative enzyme-linkedimmunosorbent assay (ELISA) (Endogen Inc.; Cambridge, MA).After 24 h incubation, the culture medium was removed andcentrifuged at 1,000 rpm (4ºC) to remove floating cells. Theresultant supernatant was passed through a 0.22 m filter andstored at –70ºC. Human IFN-α levels in the supernatant weredetermined by radioimmunoassays using antihuman IFN-αmonoclonal antibodies following the manufacturer’s instruc-tions. CD34+ cells were placed in fresh IMDM at a concentra-tion of 5 × 106 cell/ml and assayed for IFN-α after 24 and 48 h.
Hematopoietic Colony AssaysMethylcellulose culture technique was used to assess the
clonal efficiency of the transduced cells with the IFN-α gene.One thousand CD34+ cells were plated in methylcellulose(final concentration 1.12% [Fisher Scientific; Fairlawn, NY])with IMDM supplemented with 15% fetal bovine serum and100 U/ml GM-CSF for CFU-GM colony assay or in the pres-ence of 2 U/ml erythropoietin ([EPO] Toyobo; Osaka, Japan)and antibiotics as described previously [1, 29]. Cells wereincubated at 37ºC in a humidified atmosphere containing 5%CO2 in air. Myeloid colonies consisting of 50 cells or morewere counted as CFU-GM and more than two clusters of 20or more hemoglobinized cells were counted as erythroidBFU-E using an inverted microscope. CFU-GM and BFU-Eclones were picked from methylcellulose plates and used formeasurement of IFN-α mRNA by RT/PCR.
Quantitative Measurement of IFN-α mRNA by RT/PCRRNA is reverse-transcribed using the first strand cDNA
synthesis kit from Clontech, Inc. (Palo Alto, CA). Briefly,

Feldman, Ahmed, Lutton et al. 389
RNA in 13.5 ml of DEPC-treated H2O containing 1 ml ofoligo (dT)18 (final concentration 0.2 mM) is denatured at70ºC for two min. The denatured RNA is placed on ice, and6.5 ml of the reverse transcription mixture containing 50mM Tris-HCl, pH 8.3, 75 mM KCl, 3 mM MgCl2, 0.5 mMof each dNTP, 1 U/ml of RNase inhibitor, and 200 UMoloney murine leukemia virus reverse transcriptase(Bethesda Research Laboratories; Gaithersburg, MD) areadded. The reaction tube is then incubated at 42ºC for oneh followed by heating to 95ºC to stop the reaction and thenplaced on ice.
The PCR reaction is performed by adding the PCRmixture to a final volume of 100 ml to the RT reactiontube [40]. The PCR mixture contained 10 mM Tris-HCl,pH 8.3, 50 mM KCl, 0.001% gelatin, 1.5 mM MgCl2, 250mM of each dNTP, 1 mM of sense and antisense primers,2.5 U of Taq DNA polymerase, and 1 mCi of 32P-dCTP[α-32P] dCTP (3,000 Ci/mmol and Ci-37 Gbq are fromAmersham Corporation; Arlington Heights, IL; DNAmolecular weight marker VI is from BoehringerMannheim Biochemicals; Indianapolis, IN; dNTPs [100mM in sterile deionized H2O] are from PromegaCorporation; Madison, WI; Taq DNA polymerase andrestriction enzymes PstI and PvuII are from Stratagene,and Nusieve GTG and SeaKem LE agarose are fromFMC BioProducts; Rockland, ME; standard and primersare from Promega). The reaction mixture is overlaid withtwo drops of mineral oil and subjected to 40 cycles as fol-lows: 95ºC for 1 min, 55ºC for 1 min and 72ºC for 2 min.After the last cycle, a final extension will be performed at72ºC for 10 min.
Southern and Northern Blot Analysis of Transfected CellsGenomic DNA was extracted from CD34+ cells, and
CFU-GM and BFU-E clones. The cell preparations (2-5 ×106 cells) were digested with Bam HI, Hind III and XhoI. The digested DNA was electrophoresed on an agarosegel and then transferred to nitrocellulose filters as previ-ously described [34]. Total RNA from 2-5 × 106 CD34+
cells was extracted according to the method ofChomczyskiand Sacchi[41] and used for RT/PCR. IFN-α transcripts were amplified by RT/PCR using 2 ml of100 ml cDNA mixture of total reaction volume of 25 ml.Aliquots of the reaction mixture (10 ml) were subjectedto electrophoresis in a 0.18% agarose gel. Nucleic acidswere transfected to nitrocellulose by blotting and probedwith IFN-α cDNA probes. Filters were exposed to x-rayfor 48 h at –80°C. The Bgl I fragment of IFN-α cDNAwas labeled with [α-32P] dCTP using a mulitprimer DNA-labeling system. The filters were then hybridized with the32P-labeled IFN-α cDNA.
RESULTS
In Vitro Expression of IFN-α cDNA Assessment of IFN-αProtein and mRNA
In preliminary experiments, we examined the expres-sion/overexpression of IFN-α in transfected CD34+ cells.Multiple doses of AdCMV-IFN-α were used to transfecthuman CD34+ cells. A schematic representation of the ade-noviral vector containing the IFN-α gene is represented inFigure 1. We next evaluated the ability of the AdvCMV-IFN-α vector to express the IFN-α cDNA and direct pro-duction of IFN-α protein. After 24 h of induction, the cellswere spun down, the medium harvested and IFN-α proteinmeasured by radioimmunoassay. Results are presented inTable 1. As seen in Table 1, AdCMV-IFN-α caused eleva-tion of IFN-α protein in the conditioned medium, suggest-ing that the IFN-α gene is expressed in CD34+ cellsresulting in the release of IFN-α into the medium (4.1U/ml). On the other hand, AdCMV-HO-1 (heme oxygenasegene) in control cells does not express significant levels ofIFN-α. The value listed for these two cells represents onlyabout 0.3 U/ml.
Adenoviral vector containingthe human interferon alpha gene
pAdBg1 II
CMVAd (Pkg) IFN-α HRF
NruI/StuIEcoRI Bg1II HindIII/XhoI BamHI/Bg1II
Figure 1. A schematic diagram of the adenoviral vector containingthe IFN-α gene.
Table 1. Secretion of IFN-α by AdCMV-IFN-α transduced humanCD34+ cells
Condition Unit/ml
Control CD34+ 0.3 ± 0.1
Transduced CD34+ AdCMV-IFN-α 4.1 ± 0.6
Transduced CD34+ AdCMV-HO-1 0.3 ± 0.1
Concentration of released IFN-α in U/106 cells/24 h. Value below assaysensitivity limit was recorded as <0.3 U/control CD34+ cells/24 h.

390 Gene Transfer in Leukemia
Viability of the cells was determined using PI and flowcytometric analysis. Cells were stained with 20 ml of CD34-fluorescein isothiocyanate and CD45-PerCP at 4°C for 30min. Usually 10,000 cells were analyzed for each condition.PI (5 mg) was added five min before completion of the anti-body incubation. The cells were washed once, resuspendedin PBS, and analyzed immediately by flow cytometry. Theflow cytometric analysis was performed using an EPICSProfile II (Coulter). The viability of the CD34+ and CD34–
cell populations was determined by restricting analysis basedupon antibody reactivity (fluorescence). Histograms depictlight scatter characteristics CD34+ analysis and PI uptake ofthe CD34+ cell population. Results are from the flow cyto-metric determination of CD34+ cell viability at 4, 8, 16 and24 h transfection periods.
As shown in Figure 2, viabilities were determined forpurified CD34+ cell populations after 4 h (Fig. 2A-C), 8 h(Fig. 2D-F), 16 h (Fig. 2G-I), and 24 h (Fig. 2J-L) incubationperiods. The loss of membrane integrity (nonviable) was cal-culated by measurement of PI dye uptake for both CD34+ andCD34– cell populations. The size (forward scatter) and inter-nal complexity (side scatter) for each event were plotted forthe four incubation periods (Fig. 2A, D, G, J). The percentageof CD34+ cells in the samples, which was corrected for debris,was 96.3% at 4 h (Fig. 2B), 94.8% at 8 h (Fig. 2E), 67.2% at16 h (Fig. 2H), and 57.0% at 24 h (Fig. 2K). The percentageof cells that were CD34+ and maintained membrane integritywas 98.8% at 4 h (Fig. 2C), 97.0% at 8 h (Fig. 2F), 83.3% at16 h (Fig. 2I), and 73.5% at 24 h (Fig. 2L). This was done byanalysis of data in Figure 2 (B, E, H and K) in conjunctionwith PI uptake, represented in Figure 2 (C, F, I and L).
Establishment of the Dose Dependent Expression of AdCMV-IFN- α in CD34+ Cells
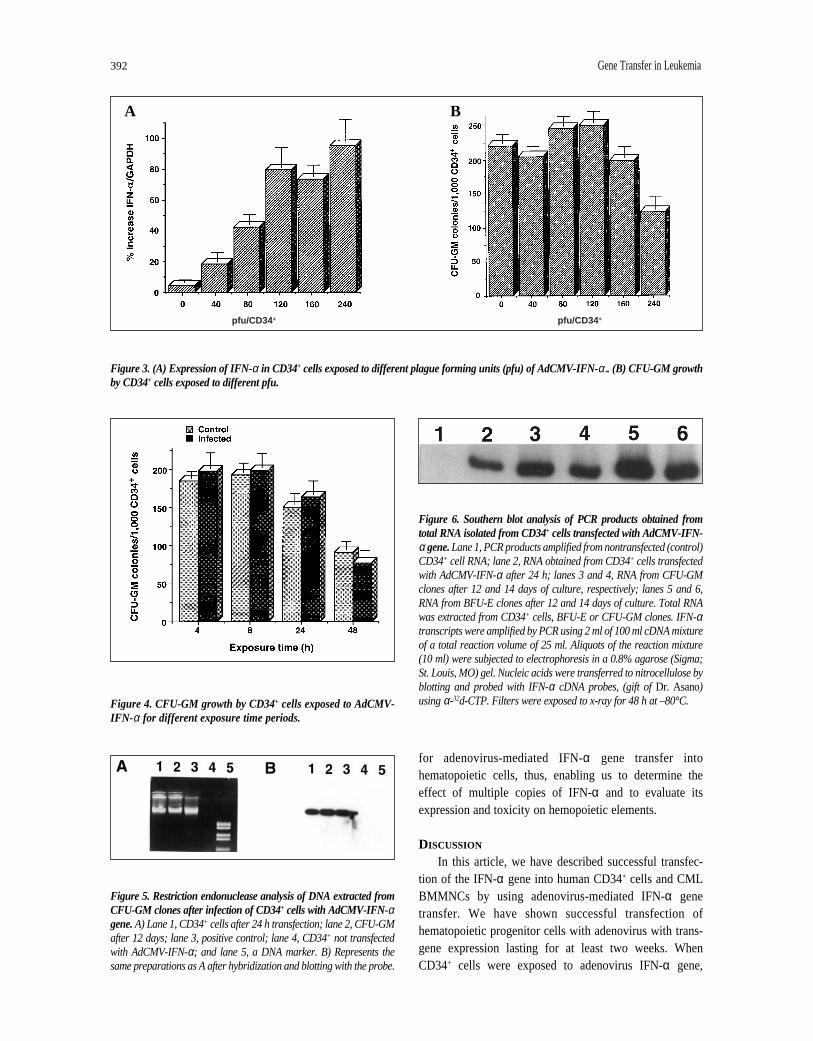
CD34+ cells were plated in IMDM at 2 × 106 cells per 25cm2 flask and incubated with AdCMV-IFN-α at different lev-els of viral particles (40, 80, 120, 160, 240, pfu/cell) for 1-48 h(37°C, 5% CO2) and then washed and used for assessment ofmRNA and clonal efficiency. IFN-α mRNA was measuredby RT/PCR in CD34+ cells. As seen in Figure 3A, optimalexpression of IFN-α in CD34+ cells could be achieved with120 pfu/CD34+/cell, (81%), and there was little or no signifi-cant increase with 160-240 pfu (76%-91%). Cells exposed to30-40 pfu yielded about 15%-27% increase in IFN-α (datanot shown).
Toxicity of AdCMV-IFN- α on CD34+ CellsWe tested the effect of the dose response from 120-240
pfu/cell on CD34+ progenitor cell growth and differentiation.After 24 h incubation with AdCMV-IFN-α, aliquots of CD34+
cells (1,000 cells) were plated in methylcellulose cultures
containing GM-CSF. In all experiments the number of CFU-GM clones of more than 50 cells were counted. Results showthat cells transfected with 240 pfu grow less CFU-GM whenplated in culture as contrasted with cells that were exposed to120 pfu (Fig. 3B). Furthermore, 120 pfu yielded colonygrowth similar or equal to control cells. These results showthat an optimal expression of IFN-α can be achieved withCD34+ cells exposed to 120 pfu without any noticeable toxiceffect on CFU-GM growth derived from the CD34+ cells.
Clonogenic Potential of IFN-α Transduced CD34+ CellsTo evaluate the effect of AdCMV-IFN-α on CFU-GM
growth, we compared the amount of CFU-GM growth byinfected and noninfected cells. Results in Figure 4 show thatthere was no significant difference in CFU-GM growth ofinfected versus control cells for any of the exposure times.Although colony numbers were slightly less for the 24 h cellgroups as compared with four or eight h, maximal gene trans-fer occurred with the cells exposed for 24 h. However, theoverall clonogenic potential of both transduced and nontrans-duced cells in 48 h groups was less than that of cells exposedto adenovirus for 24 h in the liquid culture. Experiments arebeing performed using adenovirus vectors carrying the bacte-rial lacZ gene to accurately monitor transduction efficiency.However, the AdCMV heme oxygenase gene is available inour laboratory, and we used it for comparison to the AdCMV-IFN-α gene. Preliminary results have shown, by in vitrohybridization, that the rate of transfection varied from 5% to41% (data not shown).
Presence of CMV cDNA in Transfected CellsRestriction endonuclease analysis of DNA extracted
from CFU-GM clones after infection of CD34+ cells withAdCMV-IFN-α gene was done. Results are depicted inFigure 5 where lane 1 represents CD34+ cells after 24 h trans-fection; lane 2 represents CFU-GM after 12 days; lane 3 is apositive control; lane 4 shows CD34+ not transfected and lane5 is a DNA marker. It can be seen that the DNA patterns aresimilar in lanes 1, 2 and 3, indicating an adequate presence ofCMV cDNA of adenovirus construct which transfectedCD34+ cells as well as CD34+ progenitors (CFU-GM) derivedfrom the CD34+ cells after 14 days of culture.
Detection of IFN-α mRNA in Transduced CellsSouthern blot analysis of the PCR product obtained from
total RNA isolated from cells (and colonies) transfected withAdCMV-IFN-α gene is presented in Figure 6. Total RNA wasextracted from CD34+ cells, BFU-E and CFU-GM clones.Results in Figure 6 show that transfected CD34+ cells (lane 2)and clones of CFU-GM (lanes 3, 4) and BFU-E (lanes 5, 6)(grown for 12-14 days) all show the appropriate RNA bands.

Feldman, Ahmed, Lutton et al. 391
Lane 1 is a nontransfected control. The experiments could notdetermine how many copies of IFN-α mRNA are transcribed.However, quantitative RT/PCR methods are being developedto answer this question.
CFU-GM Clonogenic Potential ofTransduced CML-BMMNC
Previous results using adenovirus-mediated IFN-α transfer as a reporter genewithout significant suppression of CD34+
growth encouraged us to test the ability ofadenovirus on transfections of progenitorcells obtained from CML patients. For thispurpose, we used similar doses of aden-ovirus (120-240 pfu) to transfect BMMNCin liquid culture for 24 h in the presence ofcytokines. Aliquots of cells were removedand tested for the transfection ability andclonal efficiency of CML progenitor cells.The clonal efficiency of transfected cellswas then assayed by plating BMMNC inmethylcellulose. As seen in Table 2,BMMNC from CML transduced withIFN-α (120 pfu) generated 46.0 ± 5.9CFU-GM as compared to 48.0 ± 5.0 innontransduced cells. In contrast, aden-ovirus CMV-IFN-α transfection causes adecreased number of CFU-GM when thepfu/cell were 240 compared to the unin-fected control cells. A substantial inhibi-tion of CFU-GM was also seen whenBMMNC were transfected with 300pfu/cell. Expression of IFN-α mRNA inCFU-GM generated from CML was nextdetermined. In an effort to assess theexpression of IFN-α mRNA in progenitorcells, we performed RT/PCR on the RNAextracted from various clones similar tothat performed on CD34+ progenitor cells.As seen in Figure 7, results of RT/PCR oftotal RNA of CFU-GM and BFU-Edemonstrate IFN-α mRNA was expressedin CFU-GM clones after 12 and 14 days ofculture (lanes 3 and 4) as compared to thecontrol (lane 1). Similarly, BFU-E clonesexpressed a substantial level of IFN-αmRNA. These results demonstrate thatrecombinant adenovirus is an effectivevehicle for transiently expressing genes inboth hematopoietic BMMNC obtainedfrom CML donors and in CD34+ cellsobtained from normal donors. Further, the
IFN-α gene is effectively carried to clones of hematopoieticelements (CFU-GM) derived from both primitive cells ofnormal and CML donors. Although the expression of thegene is transient, we have been able to identify conditions
Figure 2. Results from the flow cytometric determination of CD34+ cell viability at 4 h(A, B, C), 8 h (D, E, F), 16 h (G, H, I), and 24 h (J, K, L) transfection periods. Cells werestained with 20 ml of CD34-fluorescein isothiocyanate and CD45-PerCP (Becton-Dickinson) at 4°C for 30 min. PI (5 mg) was added five min before completion of the anti-body incubation. The cells were washed once, resuspended in PBS, and analyzedimmediately by flow cytometry. Histograms depict light scatter characteristics (Fig. 2A, D,G and J), CD34+ analysis (Fig. 2B, E, H and K) and PI uptake of the CD34+ cell popula-tion (Fig. 2C, F, I and L).
for adenovirus-mediated IFN-α gene transfer intohematopoietic cells, thus, enabling us to determine theeffect of multiple copies of IFN-α and to evaluate itsexpression and toxicity on hemopoietic elements.
DISCUSSION
In this article, we have described successful transfec-tion of the IFN-α gene into human CD34+ cells and CMLBMMNCs by using adenovirus-mediated IFN-α genetransfer. We have shown successful transfection ofhematopoietic progenitor cells with adenovirus with trans-gene expression lasting for at least two weeks. WhenCD34+ cells were exposed to adenovirus IFN-α gene,
392 Gene Transfer in Leukemia
Figure 3. (A) Expression of IFN-α in CD34+ cells exposed to different plague forming units (pfu) of AdCMV-IFN-α.. (B) CFU-GM growthby CD34+ cells exposed to different pfu.
Figure 4. CFU-GM growth by CD34+ cells exposed to AdCMV-IFN- α for different exposure time periods.
Figure 5. Restriction endonuclease analysis of DNA extracted fromCFU-GM clones after infection of CD34+ cells with AdCMV-IFN-αgene. A) Lane 1, CD34+ cells after 24 h transfection; lane 2, CFU-GMafter 12 days; lane 3, positive control; lane 4, CD34+ not transfectedwith AdCMV-IFN-α; and lane 5, a DNA marker. B) Represents thesame preparations as A after hybridization and blotting with the probe.
Figure 6. Southern blot analysis of PCR products obtained fromtotal RNA isolated from CD34+ cells transfected with AdCMV-IFN-α gene.Lane 1, PCR products amplified from nontransfected (control)CD34+ cell RNA; lane 2, RNA obtained from CD34+ cells transfectedwith AdCMV-IFN-α after 24 h; lanes 3 and 4, RNA from CFU-GMclones after 12 and 14 days of culture, respectively; lanes 5 and 6,RNA from BFU-E clones after 12 and 14 days of culture. Total RNAwas extracted from CD34+ cells, BFU-E or CFU-GM clones. IFN-αtranscripts were amplified by PCR using 2 ml of 100 ml cDNA mixtureof a total reaction volume of 25 ml. Aliquots of the reaction mixture(10 ml) were subjected to electrophoresis in a 0.8% agarose (Sigma;St. Louis, MO) gel. Nucleic acids were transferred to nitrocellulose byblotting and probed with IFN-α cDNA probes, (gift of Dr. Asano)using α-32d-CTP. Filters were exposed to x-ray for 48 h at –80°C.
A B
pfu/CD34 + pfu/CD34 +

Feldman, Ahmed, Lutton et al. 393
IFN-α protein was detected in the incubation mediumwithin 24 h, indicating that IFN-α protein was released intothe medium. Analysis of IFN-α gene in CD34+ cellsdemonstrated that adenovirus was able to deliver functionalDNA since IFN-α mRNA was detected in both CD34+ cellsand in colonies generated from the CD34+ cells. At the pre-sent time, we do not know the efficacy of adenoviral vec-tors for transduction of CML BMMNCs as compared withnormal CD34+ cells. It is possible that virus infection maybe taken up at a higher level in BMMNCs than in CD34+
cells. Experiments to determine the number of copies ofIFN-α mRNA in the more phenotypically primitiveCD34+/CD38– CML BMMNCs are to be done in the nearfuture. However, it is possible that a different receptor ispresent on more mature cells, such as BMMNC, whichcould then facilitate the uptake of adenovirus. In fact, Huanget al. [42] have shown that upregulation of certain receptorssuch as integrins on monocytes, facilitates adenovirus medi-ated gene delivery. Identification and utilization of a specificreceptor on CML cells may prove to be an excellent tool forinduction of cell death by IFN-α. It is not know what theeffectiveness of microenvironmental production (endoge-nous) of IFN-α will have on progenitor cell growth and dif-ferentiation. It remains possible that local production ofIFN-α in the stromal microenvironment may be more effec-tive than much higher levels of IFN-α supplied exogenously.
The antitumor effects of IFN-α have been extensivelystudied by many investigators, and the effectiveness of IFNas a therapeutic modality has been confirmed in the treat-ment of renal cell carcinoma, hairy cell leukemia, malig-nant melanoma and myeloma [4]. In addition to theseneoplasms, recent studies have revealed that IFN-α is alsoeffective in the treatment of Ph+ CML [22]. However, long-term continual parenteral administration of IFN-α isrequired to maintain therapeutic efficacy against these dis-eases. IFN-α therapy has been shown to be of great value tocontrol the levels of bcr/abl mRNA, adhesion moleculeexpression and enhancement of cell differentiation. It
would be beneficial for the patient if an appropriate amountof IFN-α could be expressed locally over a long period oftime. Gene transfer of IFN-α into human CD34+ cells mayprove to be of clinical value for continuous overexpressionof IFN-α in CML patients.
In this report, we used adenovirus as a transient means ofgene transfer. Adenovirus vectors provide an ideal vehiclefor transient expression by which to deliver IFN-α for thepurpose of abrogating Ph+ levels induced in CML cells.Results suggest that the transgene of IFN-α can be achievedusing the AdCMV vector with a varied efficiency rangingfrom 5%-41%. Further, there is a direct relationship betweenincreased adenovirus dose and toxicity, especially in MNCsof CML donors. The objective of these studies was to defineconditions that enhance in vitro adenovirus-mediated genetransfer to hematopoietic cells and to examine the effects ofvarious growth factors on transduction to improve transduc-tion efficiency. We have applied the adenovirus IFN-α-mediated gene transfer into CD34+ cells with good success.These studies in our laboratory have shown that there is nosuppressive effect on CD34+ cell growth and differentiationafter transduction with IFN-α gene.
Data from different laboratories suggest that normalhematopoietic stem cells (HSCs), present in CML marrow,can be separated from leukemic HSCs by fluorescence-acti-vated cell sorting [39, 43]. These findings suggest thatCML progenitors may be targets for vectors which expressantisense bcr/abl [44]. Also, retroviral vectors which conferdrug resistance to autologous bcr/abl-negative HSCs maybe of potential therapeutic benefit for CML. Retrovirus-mediated antisense of bcr/abl is currently being considered
Table 2. Clonogenic potential (CFU-GM) of CML BMMNCtransfected with AdCMV-IFN-α*
AdCMV-IFN- α pfu/cell No. of CFU-GM % Change
0 48.0 ± 5.0 –
120 46.0 ± 5.9 4
240 31.7 ± 5.9 34
300 26.3 ± 4.9 46
*Samples of BMMNC were plated (5 × 103/well) in methylcellulosemedium to determine CFU-GM progenitor content. Each data point isderived from triplicate samples in the presence of 100 U/ml GM-CSF.
Figure 7. Southern blot analysis of PCR products obtained fromtotal RNA isolated from CML hematopoietic stem cells(BMMNC) transfected with AdCMV-IFN-α gene. Lane 1, PCRproducts amplified from nontransfected (control) cell RNA; lane2, RNA obtained from cells transfected with AdCMV-IFN-α after24 h; lane 3, RNA from CFU-GM clones after 12 days; lane 4,CFU-GM clones after 14 days of culture; lane 5, no RNA wasadded; and lane 6, positive control. Total RNA was extracted fromstem cells or CFU-GM clones by one single extraction. cDNA wassynthesized from total RNA. IFN-α transcripts were amplified byPCR using 2 ml of 100 ml cDNA mixture of a total reaction vol-ume of 25 ml. Aliquots of the reaction mixture (10 ml) were sub-jected to electrophoresis in a 0.8% agarose (Sigma) gel. Nucleicacids were transferred to nitrocellulose by blotting and probedwith IFN-α cDNA probes, using α-32P-CTP. Filters were exposedto x-ray for 48 h at –80°C.

394 Gene Transfer in Leukemia
for treatment of CML [45]. Retroviral vectors are currentlythe only gene transfer system with the appropriate charac-teristics of chromosomal integration and the concurrent useof stable helper-free producer cell lines that can be usedclinically in protocols targeted at HSCs. In rodent models,investigators have demonstrated efficient and reproduciblegene transfer to a high percentage of long-term repopulat-ing stem cells and achieved long-term expression of intro-duced genes in appropriate lineages [1, 46]. Efficient genetransfer to primitive human progenitor cells, such as CFU-granulocyte-erythroid-macrophage-megakaryocyte or long-term culture initiating cells, has been reported, with genetransfer efficiencies greatly increased by exposing targetcells to hematopoietic growth factors during transductionwith viral vectors [46].
Results in this study show that we can efficientlytransfer the IFN-α gene into normal (CD34+) cells andCML primitive hematopoietic mononuclear stem cells,and that the gene transfer has little or no effect on nor-mal CD34+ hematopoietic growth but does reduce
growth by CML BMMNC. This in vitro data using tran-sient gene transfer IFN-α suggest that selective expres-sion of IFN-α may be of beneficial value for CMLtherapy.
In summary, this report demonstrates and confirms otherinvestigations that adenovirus vector can be used to mediategene transfer into CD34+ cells. We applied this approach toan important clinical setting in which IFN is of general use.We have shown that adenovirus-mediated IFN-α can beeffective in gene transfer. However, the level of IFN-α to beused at therapeutic levels to enhance adhesion molecules orsuppress bcr/abl gene expression is an important debate forupcoming investigations.
ACKNOWLEDGMENTS
This work was supported, in part, by NationalInstitutes of Health Grants RO1-HL-5438. We thank Dr.R. Shadduck(University of Pittsburgh School of Medicine,Pittsburgh, PA) for his helpful suggestions and review ofthe manuscript.
REFERENCES
1 Chertkov JL, Jiang S, Lutton JD et al. The hematopoietic stromalmicroenvironment promotes retrovirus-mediated gene transferinto hematopoietic stem cells. STEM CELLS 1993;11:218-227.
2 Dunbar CE, Emmons RV. Gene transfer to hematopoieticprogenitor and stem cells: progress and problems. STEM CELLS1994;12:563-570.
3 Mulligan RC. The basic science of gene therapy. Science1994;260:926-932.
4 Hanania EG, Kavanagh J, Hortogagyi G et al. Recentadvance in the application of gene therapy to human disease.Am J Med 1995;99:537-552.
5 Christal RG, McElvaney NG, Rosenfeld MA et al.Administration of an adenovirus containing the human CFTRcDNA to the respiratory tract of individuals with cystic fibrosis.Nat Genet 1994;8:42-51.
6 Watanabe T, Kiszynski C, Ino K et al. Gene transfer into humanbone marrow hematopoietic cells mediated by adenovirusvectors. Blood 1996;87:5032-5036.
7 Haddada H, Lopez M, Martinache C et al. Efficient aden-ovirus-mediated gene transfer into human blood monocyte-derived macrophages. Biochem Biophy Res Commun1993;195:1174-1183.
8 Mitani K, Graham FL, Caskey CT. Transduction of humanbone marrow by adenoviral vector. Hum Gene Ther1994;5:941-948.
9 Abraham NG, da Silva J-L, Lavrovsky Y et al. Adenovirus-mediated heme oxygenase-1 gene transfer into rabbit ocular tis-sues. Invest Ophthalmol Vis Sci 1995;36:2202-2210.
10 Ohwada A, Rafii S, Moore MAS et al. In vivo adenovirus vec-tor-mediated transfer of the human thrombopoietin cDNA main-tains platelet levels during radiation- and chemotherapy-inducedbone marrow suppression. Blood 1996;88:778-784.
11 Fialkow PJ, Jacobson RJ, Papayannopoulou T. Chronic mye-locytic leukemia: clonal origin in a stem cell common to thegranulocyte, erythrocyte, platelet and monocyte/macrophage.Am J Med 1977;63:125-130.
12 Martin PJ, Najfield V, Hansen JA et al. Involvement of the B-lymphoid system in chronic myelogenous leukemia. Nature1980;287:49-50.
13 Nitta M, Kato Y, Strife A et al. Incidence of involvement of theB and T lymphocyte lineages in chronic myelogenous leukemia.Blood 1985;66:1053-1061.
14 McGlave PB, Bartsch G, Anasetti C et al. Unrelated donorbone marrow transplantation therapy for chronic myelogenousleukemia: initial experience of the National Marrow DonorProgram (NMDP). Blood 1993;81:543-550.
15 Snyder DS, McGlave PB. Treatment of chronic myelogenousleukemia with bone marrow transplantation. Hematol OncolClin North Am 1990;4:53-57.
16 McWhirter JR, Wang JYI. Activation of tyrosine kinase andmicrofilament-binding functions by c-able by bcr sequencesin bcr/abl fusion protein. Mol Cell Biol 1991;11:1553-1565.
17 Gordon MY, Dowding CR, Riley GP et al. Altered adhesiveinteractions with marrow stroma of haematopoietic cells inchronic myeloid leukemia. Nature 1987;328:342-345.
18 Verfaillie CM, McCarthy JB, McGlave PB. Mechanismsunderlying abnormal trafficking of malignant progenitorsin chronic myelogenous leukemia. J Clin Invest1992;90:1232-1241.
19 Wetzler M, Kurzrock R, Lowe DG et al. Alteration in bonemarrow adherent layer growth factor expression: a novel mech-anism of chronic myelogenous leukemia progression. Blood1991;78:2400-2406.

Feldman, Ahmed, Lutton et al. 395
20 Bhatia R, McGlave PB, Dewald GW et al. Abnormal functionof the bone marrow microenvironment in chronic myelogenousleukemia: role of malignant stromal macrophages. Blood1995;85:3636-3645.
21 Bhatia R, McCarthy JB, Varfaillie CM. Interferon-α restoresnormal b1 integrin-mediated inhibition of hematopoieticprogenitor proliferation by the marrow microenvironment inchronic myelogenous leukemia. Blood 1996;87:3883-3891.
22 Italian Cooperative Study Group on Chronic MyeloidLeukemia. Interferon alfa-2a as compared with conventionalchemotherapy for the treatment of chronic myeloid leukemia.N Engl J Med 1994;330:820-825.
23 Broxmeyer HE, Lu E, Platzer E et al. Comparative analysesof the influences of human gamma, alpha and beta inteferonson human multipotential (CFU-GEMM), erythroid (BFU-E)and granulocyte macrophage (CFU-GM) progenitor cells. JImmunol 1983;131:1330-1335.
24 Geissler D, Gastl G, Aulitzky W et al. Recombinant interferon-alpha-2C in chronic myelogenous leukemia: relationship ofsensitivity of committed haematopoietic precursor cells in vitro(BFU-E, CFU-GM, CFU-Meg) and clinical response. LeukRes 1990;14:629-636.
25 Upadhyaya G, Guba SC, Sih SA et al. Interferon-alpharestores the deficient expression of the cytoadhesion mole-cule lymphocyte function antigen-3 by chronic myelogenousleukemia progenitor cells. J Clin Invest 1991;88:2131-2136.
26 Dowding C, Guo AP, Osterholz J et al. Interferon-alpha over-rides the deficient adhesion of chronic myelogenous leukemiaprimitive progenitor cells to bone marrow stromal cells.Blood 1991;78:499-505.
27 Bhatia R, McGlave P, Verfaillie CM. Treatment of marrowstroma with interferon-α restores normal b1 integrin-depen-dent adhesion of chronic myelogenous leukemia hematopoi-etic progenitors. Role of MIP-1α. J Clin Invest 1995;96:931.
28 Aman MJ, Keller U, Derigs G et al. Regulation of cytokineexpression by interferon alpha in human bone marrow stromacells: inhibition of hematopoietic growth factors and induc-tion of IL-1 receptor antagonist. Blood 1994;84:4142-4150.
29 Bhatia R, McGlave PB, Verfaillie CM. Interferon-α treat-ment of marrow stroma results in enhanced adhesion ofchronic myelogenous leukemia progenitors via mechanisminvolving MIP-1α and TGF-β. Exp Hematol 1994;22:797a.
30 Zoumbos N, Gascon P, Young NS. The induction of lympho-cytes in normal and suppressed hematopoiesis. Blut 1984;48:1-9.
31 Emerson SG, Antin JH. Bone marrow progenitor cells inducea regulatory autologous proliferative T lymphocyte response.J Immunol 1989;142:766-772.
32 Eaves CJ, Cashman JD, Eaves AC et al. Mechanisms that regu-late the cell cycle status of very primitive hematopoietic cells inlong-term human marrow cultures. II. Analysis of positive andnegative regulators produced by stromal cells within the adher-ent layer. Blood 1991;78:110-117.
33 Hanabuchi S, Koyanagi M, Kawasaki A et al. Fas and its lig-and is a general mechanism of T-cell-mediated cytotoxicity.Proc Natl Acad Sci USA 1994;91:4930-4934.
34 Ausubel FM, Brent R, Kingston RE et al. Current Protocols inMolecular Biology. New York: John Wiley & Sons, 1987:121-132.
35 Brandt J, Srour EF, van Besien K et al. Cytokine dependent longterm culture of highly enriched precursors of hematopoieticprogenitor cells from human bone marrow. J Clin Invest1990;86:932-941.
36 Verfaillie C, McGlave PB. Leukemia inhibitory factor/humaninterleukin for DA cells: a growth factor that stimulates the invitro development of multipotential human hemopoieticprogenitors. Blood 1991;77:263-270.
37 Verfaillie C, Blakholmer K, McGlave P. Purified primitivehuman hematopoietic progenitors with long term in vitrorepopulating capacity adhere selectively to irradiated bonemarrow stroma. J Exp Med 1990;179:509-520.
38 Berenson RJ, Andrews RG, Bensinger WI et al. AntigenCD34+ marrow cells engraft lethally irradiated baboons. J ClinInvest 1988;81:951-956.
39 Verfaillie CM, Miller WJ, Boylan K et al. Selection of benignprimitive hematopoietic progenitors in chronic myelogenousleukemia on the basis of DR antigen expression. Blood1992;79:1003-1008.
40 Goodman AI, Choudhury M, da Silva J-L et al. Quantitativemeasurement of heme oxygenase-1 in the human adenocarci-noma. J Cell Biochem 1996;63:342-348.
41 Chomczyski R, Sacchi N. Single-step method of RNA isola-tion by acid guanidium isothiocyanate-phenol-chloroformextraction. Anal Biochem 1987;162:156-159.
42 Huang S, Endo RI, Nemerow GR. Upregulation of integrinsalpha v beta 3 and alpha v beta 5 on human monocytes andT lymphocytes facilitates adenovirus-mediated gene delivery.J Virol 1995;69:2257-2263.
43 Talpaz M, Kantarjiian H, Kurzrock R et al. Interferon-alpha pro-duced sustained cytogenetic responses in chronic myelogenousleukemia. Ann Intern Med 1991;114:532-538.
44 Leemhuis T, Liebowitz D, Cox G et al. Identification ofBCR/ABL-negative primitive hematopoietic progenitorcells within chronic myeloid leukemia marrow. Blood1993;81:801-807.
45 Martiat P, Lewalle P, Taj AS et al. Retrovirally transducedantisense sequences stably suppress P210BCR-ABL expressionand inhibit the proliferation of BCR/ABL-containing cell lines.Blood 1993;81:502-509.
46 Conneally E, Bardy P, Eaves CJ et al. Rapid and efficient selec-tion of human hematopoietic cells expressing murine heat stateantigen as an indicator of retroviral-mediated gene transfer.Blood 1996;87:456-464.
Related Documents











