This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Vaccine 29 (2011) 7905– 7912
Contents lists available at SciVerse ScienceDirect
Vaccine
jou rn al h om epa ge: www.elsev ier .com/ locate /vacc ine
A single point mutation in framework region 3 of heavy chain affects viralneutralization dynamics of single-chain Fv against bovine herpes virus type 1
Madhuri Kotia, Eva Nagyb, Azad K. Kaushika,∗
a Department of Molecular and Cellular Biology, University of Guelph, Guelph, Ontario N1G 2W1, Canadab Department of Pathobiology, University of Guelph, Guelph, Ontario N1G 2W1, Canada
a r t i c l e i n f o
Article history:Received 17 June 2011Received in revised form 20 July 2011Accepted 15 August 2011Available online 26 August 2011
Keywords:AntibodySingle chain fragment variableBovine herpes virus-1Virus neutralization
a b s t r a c t
We constructed functional recombinant single chain Fv (scFv) against bovine herpes virus type 1 (BoHV-1), aetiological agent of respiratory and genital diseases in cattle for which available vaccines do notprovide adequate protection. The scFv against BoHV-1 with 18 amino acid long flexible linker (scFv3-18L;monomeric form) recognized target antigen and, also, neutralized BoHV-1 in vitro. A comparison withrecombinant scFv with 7 amino acid linker against BoHV-1 (scFv1-7L), capable of forming diabodies,indicated that a relatively higher concentration (two-fold) of monomer scFv3-18L is needed for virusneutralization as compared to scFv1-7L. A single point replacement mutation (Asp98 to Gly98) in theframework-3 (FR3) variable-region of scFv with 18 amino acid linker (scFv4m-18L), however, affectedthe viral neutralization in a dose-dependent manner where 2.7 fold higher mutant scFv concentrationwas required to achieve virus neutralization. Despite differences in dose-dependent viral neutralization ofthe mutant scFv-18L, it detected viral antigen in an immunofluorescent assay. The outlined experimentsdemonstrate that recombinant scFv against BoHV-1, whether expressed as scFv or diabody, provide aneffective antibody based therapeutic and immunodiagnostic protein. Further, single point substitutionmutation in the FR3 can affect viral neutralization dynamics without affecting qualitative viral antigenrecognition.
© 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Molecular diversification of immunoglobulins via somaticrecombination [1] followed by mutations upon encounter with anantigen in the periphery generates specific antibody-mediated pro-tective humoral immunity. Indeed, the use of somatic cell fusiontechniques in generating hybrid cell lines that secrete monoclonalantibodies of desired specificity revolutionized immunodiagnosticsand therapeutics. In humans, the development of human anti-murine antibody (HAMA) response limited use of monoclonalantibodies in clinical applications [2]. To overcome such compli-cations, recombinant minimal antigen binding fragments, apartfrom chimeric antibodies [3–5], CDR grafting [6] and transgenichumanized mice [7–9], have been developed and expressed inmicrobes, such as, phage [10], E. coli [11] and yeast [12]. Whilecomplete antibody has a low blood clearance and tissue penetra-tion, the single chain Fv (scFv) diffuses readily through vasculatureand, also, gets rapidly cleared from blood circulation [13–15]. Suchpharmacokinetic advantages have led to construction of scFvs for
∗ Corresponding author at: Department of Molecular and Cellular Biology,SCIE4248, University of Guelph, Guelph, Ontario N1G 2W1, Canada.
E-mail address: [email protected] (A.K. Kaushik).
application in immunotherapy and clinical diagnosis across speciesincluding humans [16,17] and a number of these are currently inpreclinical trials [17–19]. While the focus of generating recombi-nant therapeutic antibodies is on humans, some effort has beenmade for developing therapeutic antibody in domestic species ofeconomic significance such as cattle. Initially, the constant regiongenes (C�, C� and C�) were expressed as transfectomas to producebovine × mouse chimeric antibodies [5]. Now recombinant scFvs ofbovine origin have been developed against foot-and-mouth disease[20] and prion protein [21–23].
Earlier we constructed functional scFv [24] against bovine her-pes virus type-1 (BoHV-1) expressed in secretory form in Pichiapastoris [25–27] under the influence of methanol regulated pro-moters. The recombinant scFv against BoHV-1 recognized viralantigen in an immunofluorescence assay and also neutralizedBoHV-1 in vitro. These scFv were constructed in VL–VH orienta-tion that provides 5–6 A greater bridging distance as comparedto VH–VL configuration [28,29] with a flexible protease resistantseven amino acid linker. The linker composition and size is criticalfor affinity, multimerization, conformational stability and foldingkinetics of the recombinant scFv [30,31]. The linkers composed ofglycine, serine residues with a size greater than 12 amino acidshave been suggested to provide sufficient flexibility for the assem-bly of VL and VH domains into a monovalent antigen combining site
0264-410X/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2011.08.077

Author's personal copy
7906 M. Koti et al. / Vaccine 29 (2011) 7905– 7912
Fig. 1. Overlapping PCR amplification of VDJ (a) and VJ (b) recombinations from cDNA isolated from HB-9907 hybridoma secreting anti-BoHV-1 IgG1. (a) Lane 1, 1-kb DNAMW ladder; Lane 2, Positive VDJ control (BLV10H8 hybridoma); Lane 3, Negative control; Lane 4, VDJ amplicon. (b) Lane 1, 1-kb MW DNA ladder; Lane 2, Positive VJ control(BLV10H8 hybridoma); Lane 3, Negative control; Lane 4, VJ PCR amplicon. (c) Lane 1, 1-kb MW DNA ladder; Lane 2, Overlap (VJ-18 codon linker-VDJ) amplicon; Lane 3,Negative control.
comparable to Fab fragment [32–34]. By contrast, linkers shorterthan 12 residues have been shown to prevent interaction betweenthe linked VL and VH domains. In such instances, complementaryVL–VH pairs from two scFv molecules are forced to form inter-molecular bivalent dimer, termed ‘diabody’, where VH domain ofone chain is known to pair with VL domain of another chain and viceversa [35]. For these reasons, recombinant scFv against BoHV-1 withan 18 amino acid long linker (monomer scFv) were constructed todetermine if differences existed with regard to viral neutralizationfunction when compared with recombinant dimerized scFv, i.e.,diabody, with higher avidity. Whether a substitution mutation inFR3 of recombinant scFv influenced viral neutralization was alsoinvestigated. The outlined experiments demonstrate that recom-binant scFv against BoHV-1, whether expressed as monomer scFvor diabody, recognized the viral antigens but in vitro viral neu-tralization dynamics differed. A single point mutation resulting inreplacement mutation (Asp98 to Gly98) in the FR3 of heavy chain ofscFv permitted antigen recognition but affected viral neutralizationin dose dependent manner.
2. Materials and methods
2.1. Hybridoma
The mouse × cattle hetero-hybridoma (HB-9907; ATCC,Rockville, MD, USA; [36]) secreting anti-BoHV-1 IgG1 antibody(0.12 mg/ml) was grown in RPMI1640 (GIBCO BRL, Gaithersburg,MD, USA) supplemented with 20% (v/v) horse serum, 5 mMsodium pyruvate, 0.5 mM MEM non essential amino acids, 1 mMglutamine and 1% (v/v) 100× antibiotic–antimycotic and 50 �M2-mercaptoethanol (GIBCO BRL, Gaithersburg, MD, USA).
2.2. cDNA synthesis, PCR and cloning
Total cellular RNA was isolated from HB-9907 hybridoma cellsusing Trizol reagent (Invitrogen, Canada) and cDNA synthesized(Amersham Biosciences) as described [37,38]. The rearranged VDJwas amplified from cDNA using primers designed from the heavychain FR1 (PDHL18; 5′ GGTCAGTCCTCTAGATCTTCCGGCGGTG-GTGGCAGCTCCGGTGGTGGCGGTTCCCAGGTGCAGCTGCG 3′) andFR4 (PDHRM; 5′ CTGGCCGGCTTGGCCACTAGTGGAGGAGACGGT-GACCAG 3′) with built-in SfiI restriction sites. The PCR was
performed with 1.5 mM MgCl2, 0.8 �M each primer, 10 mM dNTPsand 2.5 U Taq polymerase. The PCR conditions included a hotstart followed by 30 cycles of denaturation at 95 ◦C for 1 min,annealing at 68 ◦C for 1 min and extension at 72 ◦C for 1 minwith a final extension of 72 ◦C for 7 min. The VJ was ampli-fied using primers designed from the light chain FR1 (PDLLM; 5′
GTGGCCCAGCCGGCCCAGGCTGTGCTGACTCAG 3′) and FR4 (PDLR;5′ GGAAGATCTAGAGGACTGACCTAGGACGGTCAGTGTGGT 3′) withbuilt-in SfiI restriction sites. The PCR conditions were similar tothose for VDJ amplification except for the annealing temperatureof 68 ◦C. The cDNA synthesized from RNA isolated from BLV10H8hybridoma [37] was used as a positive control. The purified VJ andVDJ amplicons (Qiaquick, Qiagen Inc., USA) were combined in anoverlap extension PCR using a linker sequence encoding 18 aminoacids (GQSSRSSSGGGSSGGGGS) using primers from FR1 of VJ andFR4 of VDJ. The PCR conditions included, denaturation at 94 ◦Cfor 30 s, 30 cycles of 94 ◦C for 2 min, annealing at 65 ◦C for 15 s,extension at 72 ◦C for 15 s, followed by a final extension at 72 ◦Cfor 30 min. The purified V�-linker-VH amplicon was cloned intopPICZ� expression vector (Invitrogen, Canada) as described [24]and ligate was used to transform Top 10 E. coli (Invitrogen, Canada)by heat shock at 42 ◦C for 30 s. Recombinant plasmids, pscFv3-18Land pscFv4m-18L, was isolated from transformed E. coli (Qiaprep,Qiagen Inc., USA) and were sequenced using 5′AOX1 and 3′AOX1primers (MOBIX, McMaster University, Hamilton, Ontario, Canada).
Since ‘IMGT Collier de Perles’ program (http://imgt.cines.fr),based on unique IMGT numbering system, provides standardizedtwo-dimensional representation of any immunoglobulin heavy-or light chain domain that defines the limits of FRs and CDRs forany antigen receptor of any species relevant to the analysis ofimmunoglobulin superfamily ([39]), it was used to analyze thestructure of recombinant scFv construct from functional perspec-tive.
2.3. Transformation of P. pastoris with recombinant scFv
A single colony of KM71H P. pastoris strain was used to inoc-ulate 10 ml YPD medium and grown overnight at 30 ◦C. The cellswere diluted to an OD600 of 0.1–0.2 in YPD medium and incu-bated for 4–6 h at 30 ◦C until the OD600 reached 0.6–1.0. The cellswere pelleted by centrifugation at 500 × g for 5 min and resus-pended in 10 ml of Solution I (sorbitol containing ethylene glycol
Author's personal copy
M. Koti et al. / Vaccine 29 (2011) 7905– 7912 7907
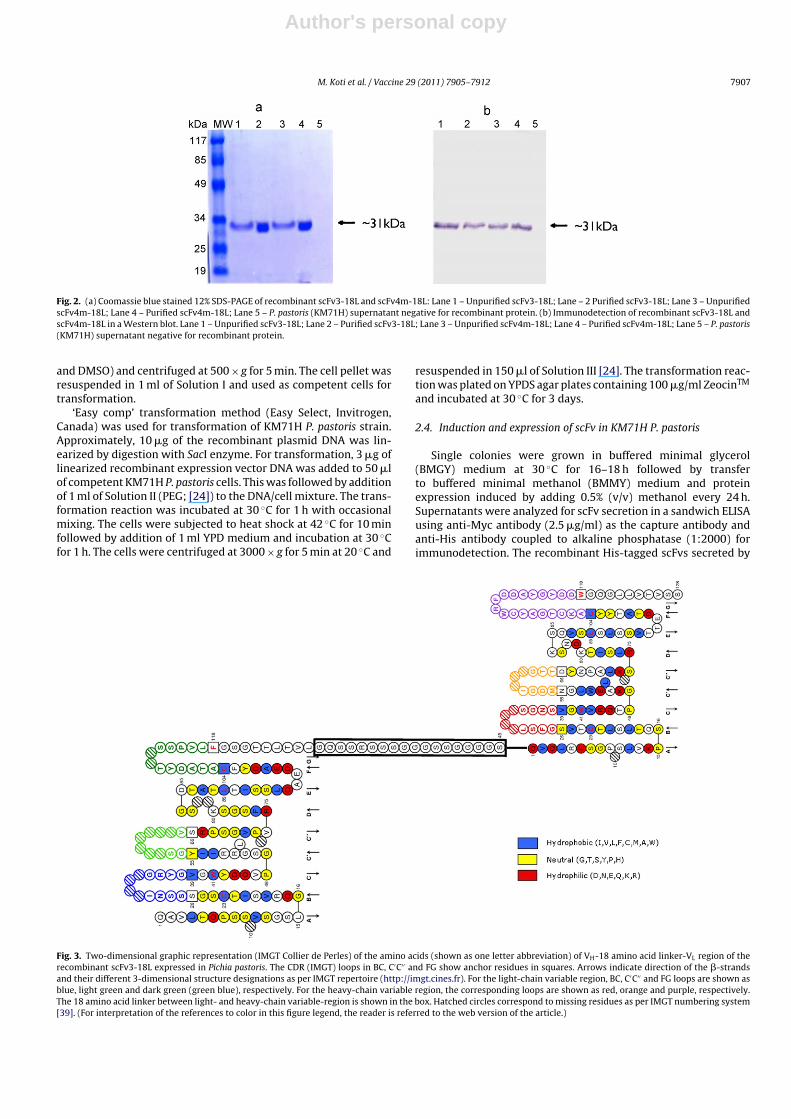
Fig. 2. (a) Coomassie blue stained 12% SDS-PAGE of recombinant scFv3-18L and scFv4m-18L: Lane 1 – Unpurified scFv3-18L; Lane – 2 Purified scFv3-18L; Lane 3 – UnpurifiedscFv4m-18L; Lane 4 – Purified scFv4m-18L; Lane 5 – P. pastoris (KM71H) supernatant negative for recombinant protein. (b) Immunodetection of recombinant scFv3-18L andscFv4m-18L in a Western blot. Lane 1 – Unpurified scFv3-18L; Lane 2 – Purified scFv3-18L; Lane 3 – Unpurified scFv4m-18L; Lane 4 – Purified scFv4m-18L; Lane 5 – P. pastoris(KM71H) supernatant negative for recombinant protein.
and DMSO) and centrifuged at 500 × g for 5 min. The cell pellet wasresuspended in 1 ml of Solution I and used as competent cells fortransformation.
‘Easy comp’ transformation method (Easy Select, Invitrogen,Canada) was used for transformation of KM71H P. pastoris strain.Approximately, 10 �g of the recombinant plasmid DNA was lin-earized by digestion with SacI enzyme. For transformation, 3 �g oflinearized recombinant expression vector DNA was added to 50 �lof competent KM71H P. pastoris cells. This was followed by additionof 1 ml of Solution II (PEG; [24]) to the DNA/cell mixture. The trans-formation reaction was incubated at 30 ◦C for 1 h with occasionalmixing. The cells were subjected to heat shock at 42 ◦C for 10 minfollowed by addition of 1 ml YPD medium and incubation at 30 ◦Cfor 1 h. The cells were centrifuged at 3000 × g for 5 min at 20 ◦C and
resuspended in 150 �l of Solution III [24]. The transformation reac-tion was plated on YPDS agar plates containing 100 �g/ml ZeocinTM
and incubated at 30 ◦C for 3 days.
2.4. Induction and expression of scFv in KM71H P. pastoris
Single colonies were grown in buffered minimal glycerol(BMGY) medium at 30 ◦C for 16–18 h followed by transferto buffered minimal methanol (BMMY) medium and proteinexpression induced by adding 0.5% (v/v) methanol every 24 h.Supernatants were analyzed for scFv secretion in a sandwich ELISAusing anti-Myc antibody (2.5 �g/ml) as the capture antibody andanti-His antibody coupled to alkaline phosphatase (1:2000) forimmunodetection. The recombinant His-tagged scFvs secreted by
Fig. 3. Two-dimensional graphic representation (IMGT Collier de Perles) of the amino acids (shown as one letter abbreviation) of VH-18 amino acid linker-VL region of therecombinant scFv3-18L expressed in Pichia pastoris. The CDR (IMGT) loops in BC, C′C′′ and FG show anchor residues in squares. Arrows indicate direction of the �-strandsand their different 3-dimensional structure designations as per IMGT repertoire (http://imgt.cines.fr). For the light-chain variable region, BC, C′C′′ and FG loops are shown asblue, light green and dark green (green blue), respectively. For the heavy-chain variable region, the corresponding loops are shown as red, orange and purple, respectively.The 18 amino acid linker between light- and heavy-chain variable-region is shown in the box. Hatched circles correspond to missing residues as per IMGT numbering system[39]. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)

Author's personal copy
7908 M. Koti et al. / Vaccine 29 (2011) 7905– 7912
Fig. 4. Two-dimensional graphic representation (IMGT Collier de Perles) of the amino acids (shown as one letter abbreviation) of VH-18 amino acid linker-VL region of therecombinant scFv4m-18L expressed in Pichia pastoris. The CDR (IMGT) loops in BC, C′C′′ and FG show anchor residues in squares. Arrows indicate direction of the �-strandsand their different 3-dimensional structure designations as per IMGT repertoire (http://imgt.cines.fr). For the light-chain variable region, BC, C′C′′ and FG loops are shown asblue, light green and dark green (green blue), respectively. For the heavy-chain variable region, the corresponding loops are shown as red, orange and purple, respectively.The 18 amino acid linker between light- and heavy-chain variable-region is shown in the box. Hatched circles correspond to missing residues as per IMGT numbering system[39]. Note amino acid substitution D89 to G89 in the FR3 region marked an asterisk. (For interpretation of the references to color in this figure legend, the reader is referredto the web version of the article.)
KM71H P. pastoris were purified on a Ni-chelating resin (Probond;Invitrogen, Canada) and protein concentration determined (Bio-Rad Protein Assay Kit; Bio-Rad, California, USA).
2.5. SDS-PAGE and Western immunoblot
Purified recombinant scFv3-18L and scFv4m-18L were gelfractionated in 12% SDS-PAGE [40,41] and electrophoreticallytransferred onto nitrocellulose (Schleicher and Schuell Inc., USA)using transfer buffer (25 mM Tris, 192 mM glycine and 20%methanol). The recombinant scFvs were detected using anti-Hisantibody coupled to alkaline phosphatase (1:2000; Invitrogen,Canada) and revealed by NBT-BCIP chromogen (Roche, Canada).Negative controls included KM71H P. pastoris supernatant that didnot express recombinant protein. In parallel, the scFv3-18L andscFv4m-18L proteins were stained by coomassie blue (0.25%, w/v,Coomassie Blue R-250 in 50:40:10 methanol:distilled water:glacialacetic acid).
2.6. Virus neutralization assay
The BoHV-1 neutralizing ability of recombinant scFv3-18L andscFv4m-18L was tested using plaque reduction assay [24,42]. First,BoHV-1 virus titer was determined to be 5 × 107 pfu/ml usingplaque assay in Madin Darby bovine kidney cells (MDBK). Briefly,100 pfu BoHV-1 was mixed with 100 �l of various concentrationsof purified scFv3-18L and scFv4m-18L and incubated at 20 ◦C for1 h following which the MDBK cells were allowed to adsorb withthe virus alone or scFv treated virus for 1 h at 20 ◦C. The HB-9907 hybridoma supernatant containing monoclonal anti-BHV-1IgG1 was used as a positive control. The infected monolayers were
washed three times with sterile PBS and overlaid with 0.7% agarosein DMEM containing 3% FBS. The cells were incubated at 37 ◦C for4 days under 5% CO2 atmosphere and fixed with 10% formalin. Thecells were then stained with 0.75% crystal violet and the result-ing plaques counted. The virus neutralization was calculated onthe basis of non-specific inhibition by heterologous protein BSA.A minimum of 50% plaque reduction was considered positive forvirus neutralization activity.
2.7. Indirect immunofluorescence
A mixture of uninfected and infected BoHV-1 MDBK cells(VMRD Inc., USA) were incubated with recombinant scFv3-18Land scFv4m-18L at the final concentration of 10 �g at 37 ◦C for30 min. The cells were washed and stained with anti-Myc anti-body (1 �g; Invitrogen, Canada), followed by detection with ProteinA conjugated to FITC (1 �g; Sigma–Aldrich, Canada). Heterologousbovine IgG1 (Cedarlane, Burlington, Canada), RPMI1640 media andphosphate buffered saline provided negative control. The parentHB9907 IgG1 monoclonal antibody against BoHV-1 was used as apositive control. The cells were washed and examined under a flu-orescence microscope (400×; Leica Microsystems Inc., IL, USA) at495 nm.
3. Results
3.1. Construction and expression of recombinant scFv with 18amino acid linker
The strategy for synthesis of V�-linker-VH involved VDJ (Fig. 1a)and VJ (Fig. 1b) recombinations, isolated from hetero-hybridomasecreting IgG1 against BoHV-1, co-amplified (Fig. 1c) together
Author's personal copy
M. Koti et al. / Vaccine 29 (2011) 7905– 7912 7909
Mean Plaque Count
Mol
ar C
once
ntra
tion
250200150100500BoHV-1 Only
PBSBSA (0.15 µM)BSA (0.45 µM)BSA (1.35 µM)
scFv3-18L (0.35 µM)scFv3-18L (1.05 µM)scFv3-18L (3.15 µM)
scFv4m-18L (0.48 µM)scFv4m-18L (1.44 µM)scFv4m-18L (4.32 µM)
HB9907 IgG1 (0.073 µM)HB9907 IgG1 (0.73 µM)
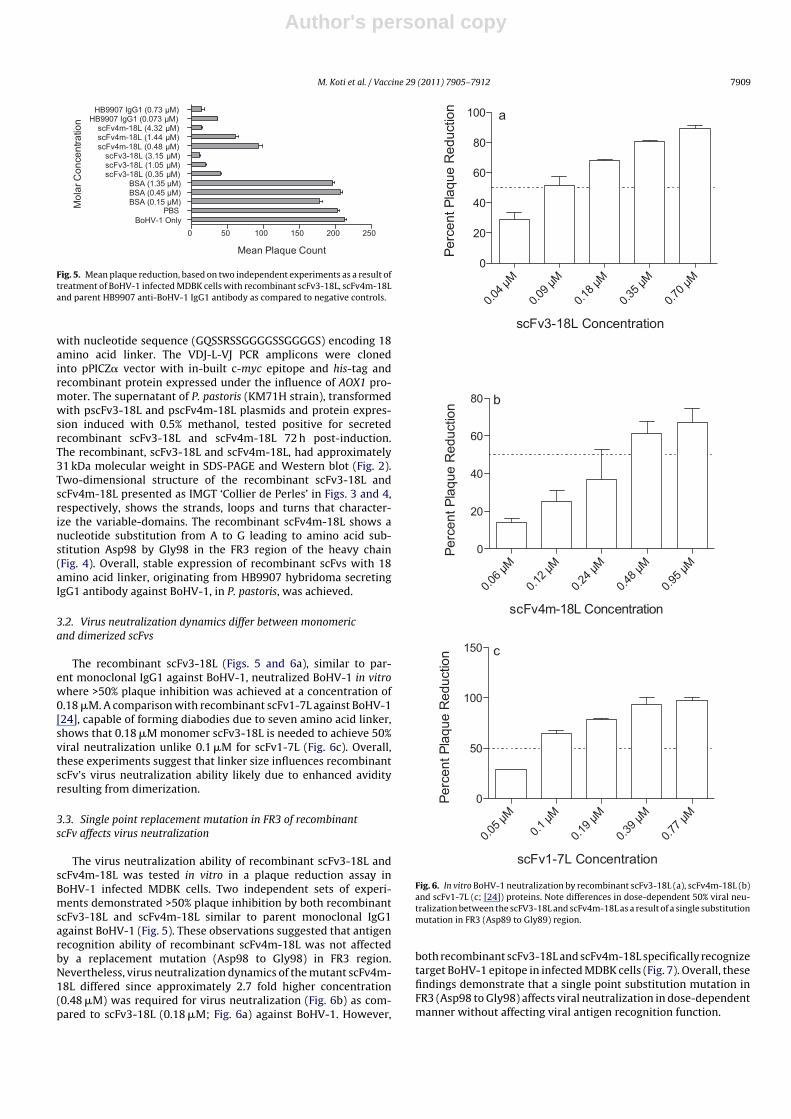
Fig. 5. Mean plaque reduction, based on two independent experiments as a result oftreatment of BoHV-1 infected MDBK cells with recombinant scFv3-18L, scFv4m-18Land parent HB9907 anti-BoHV-1 IgG1 antibody as compared to negative controls.
with nucleotide sequence (GQSSRSSGGGGSSGGGGS) encoding 18amino acid linker. The VDJ-L-VJ PCR amplicons were clonedinto pPICZ� vector with in-built c-myc epitope and his-tag andrecombinant protein expressed under the influence of AOX1 pro-moter. The supernatant of P. pastoris (KM71H strain), transformedwith pscFv3-18L and pscFv4m-18L plasmids and protein expres-sion induced with 0.5% methanol, tested positive for secretedrecombinant scFv3-18L and scFv4m-18L 72 h post-induction.The recombinant, scFv3-18L and scFv4m-18L, had approximately31 kDa molecular weight in SDS-PAGE and Western blot (Fig. 2).Two-dimensional structure of the recombinant scFv3-18L andscFv4m-18L presented as IMGT ‘Collier de Perles’ in Figs. 3 and 4,respectively, shows the strands, loops and turns that character-ize the variable-domains. The recombinant scFv4m-18L shows anucleotide substitution from A to G leading to amino acid sub-stitution Asp98 by Gly98 in the FR3 region of the heavy chain(Fig. 4). Overall, stable expression of recombinant scFvs with 18amino acid linker, originating from HB9907 hybridoma secretingIgG1 antibody against BoHV-1, in P. pastoris, was achieved.
3.2. Virus neutralization dynamics differ between monomericand dimerized scFvs
The recombinant scFv3-18L (Figs. 5 and 6a), similar to par-ent monoclonal IgG1 against BoHV-1, neutralized BoHV-1 in vitrowhere >50% plaque inhibition was achieved at a concentration of0.18 �M. A comparison with recombinant scFv1-7L against BoHV-1[24], capable of forming diabodies due to seven amino acid linker,shows that 0.18 �M monomer scFv3-18L is needed to achieve 50%viral neutralization unlike 0.1 �M for scFv1-7L (Fig. 6c). Overall,these experiments suggest that linker size influences recombinantscFv’s virus neutralization ability likely due to enhanced avidityresulting from dimerization.
3.3. Single point replacement mutation in FR3 of recombinantscFv affects virus neutralization
The virus neutralization ability of recombinant scFv3-18L andscFv4m-18L was tested in vitro in a plaque reduction assay inBoHV-1 infected MDBK cells. Two independent sets of experi-ments demonstrated >50% plaque inhibition by both recombinantscFv3-18L and scFv4m-18L similar to parent monoclonal IgG1against BoHV-1 (Fig. 5). These observations suggested that antigenrecognition ability of recombinant scFv4m-18L was not affectedby a replacement mutation (Asp98 to Gly98) in FR3 region.Nevertheless, virus neutralization dynamics of the mutant scFv4m-18L differed since approximately 2.7 fold higher concentration(0.48 �M) was required for virus neutralization (Fig. 6b) as com-pared to scFv3-18L (0.18 �M; Fig. 6a) against BoHV-1. However,
a
0.04 µ
M
0.09 µ
M
0.18 µ
M
0.35 µ
M
0.70 µ
M0
20
40
60
80
100
Perc
ent P
laqu
e R
educ
tion
scFv3-18L Concentration
b
0.06 µ
M
0.12 µ
M
0.24 µ
M
0.48 µ
M
0.95 µ
M0
20
40
60
80
Perc
ent P
laqu
e R
educ
tion
scFv4m -18L Con cen tratio n
c
0.05 µ
M0.1
µM
0.19 µ
M
0.39 µ
M
0.77 µ
M0
50
100
150
Perc
ent P
laqu
e R
educ
tion
scFv1-7L Concentrat ion
Fig. 6. In vitro BoHV-1 neutralization by recombinant scFv3-18L (a), scFv4m-18L (b)and scFv1-7L (c; [24]) proteins. Note differences in dose-dependent 50% viral neu-tralization between the scFV3-18L and scFv4m-18L as a result of a single substitutionmutation in FR3 (Asp89 to Gly89) region.
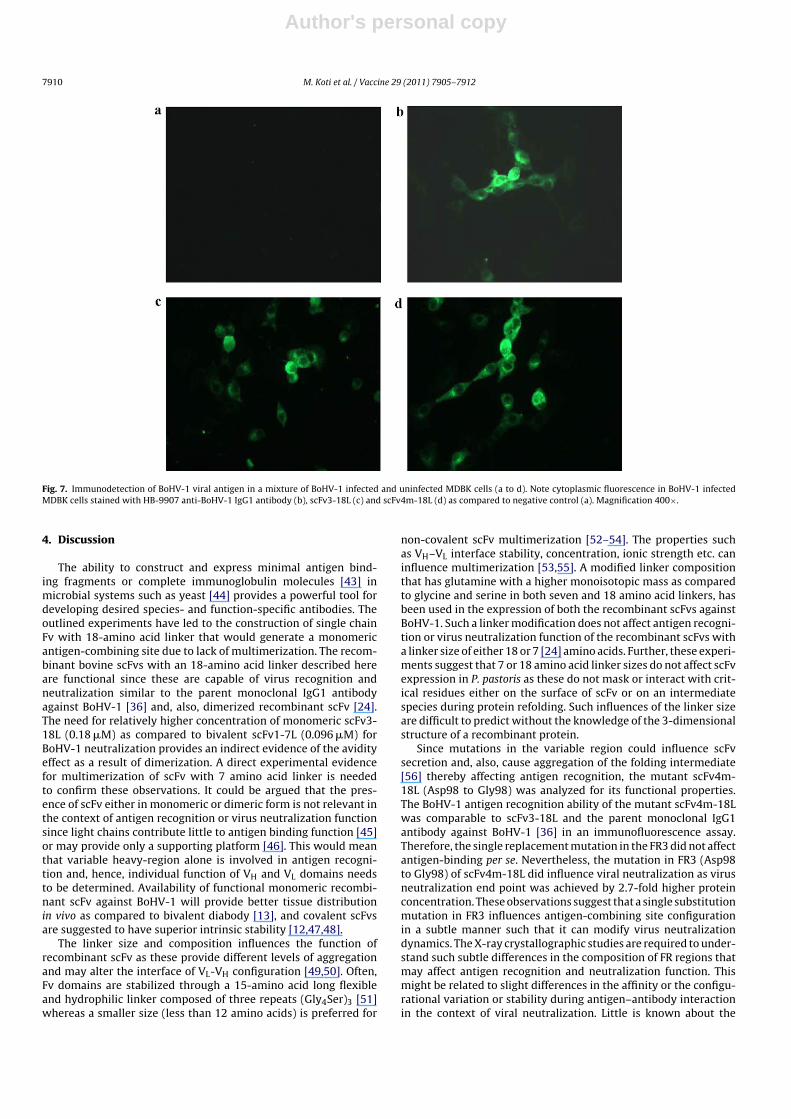
both recombinant scFv3-18L and scFv4m-18L specifically recognizetarget BoHV-1 epitope in infected MDBK cells (Fig. 7). Overall, thesefindings demonstrate that a single point substitution mutation inFR3 (Asp98 to Gly98) affects viral neutralization in dose-dependentmanner without affecting viral antigen recognition function.
Author's personal copy
7910 M. Koti et al. / Vaccine 29 (2011) 7905– 7912
Fig. 7. Immunodetection of BoHV-1 viral antigen in a mixture of BoHV-1 infected and uninfected MDBK cells (a to d). Note cytoplasmic fluorescence in BoHV-1 infectedMDBK cells stained with HB-9907 anti-BoHV-1 IgG1 antibody (b), scFv3-18L (c) and scFv4m-18L (d) as compared to negative control (a). Magnification 400×.
4. Discussion
The ability to construct and express minimal antigen bind-ing fragments or complete immunoglobulin molecules [43] inmicrobial systems such as yeast [44] provides a powerful tool fordeveloping desired species- and function-specific antibodies. Theoutlined experiments have led to the construction of single chainFv with 18-amino acid linker that would generate a monomericantigen-combining site due to lack of multimerization. The recom-binant bovine scFvs with an 18-amino acid linker described hereare functional since these are capable of virus recognition andneutralization similar to the parent monoclonal IgG1 antibodyagainst BoHV-1 [36] and, also, dimerized recombinant scFv [24].The need for relatively higher concentration of monomeric scFv3-18L (0.18 �M) as compared to bivalent scFv1-7L (0.096 �M) forBoHV-1 neutralization provides an indirect evidence of the avidityeffect as a result of dimerization. A direct experimental evidencefor multimerization of scFv with 7 amino acid linker is neededto confirm these observations. It could be argued that the pres-ence of scFv either in monomeric or dimeric form is not relevant inthe context of antigen recognition or virus neutralization functionsince light chains contribute little to antigen binding function [45]or may provide only a supporting platform [46]. This would meanthat variable heavy-region alone is involved in antigen recogni-tion and, hence, individual function of VH and VL domains needsto be determined. Availability of functional monomeric recombi-nant scFv against BoHV-1 will provide better tissue distributionin vivo as compared to bivalent diabody [13], and covalent scFvsare suggested to have superior intrinsic stability [12,47,48].
The linker size and composition influences the function ofrecombinant scFv as these provide different levels of aggregationand may alter the interface of VL-VH configuration [49,50]. Often,Fv domains are stabilized through a 15-amino acid long flexibleand hydrophilic linker composed of three repeats (Gly4Ser)3 [51]whereas a smaller size (less than 12 amino acids) is preferred for
non-covalent scFv multimerization [52–54]. The properties suchas VH–VL interface stability, concentration, ionic strength etc. caninfluence multimerization [53,55]. A modified linker compositionthat has glutamine with a higher monoisotopic mass as comparedto glycine and serine in both seven and 18 amino acid linkers, hasbeen used in the expression of both the recombinant scFvs againstBoHV-1. Such a linker modification does not affect antigen recogni-tion or virus neutralization function of the recombinant scFvs witha linker size of either 18 or 7 [24] amino acids. Further, these experi-ments suggest that 7 or 18 amino acid linker sizes do not affect scFvexpression in P. pastoris as these do not mask or interact with crit-ical residues either on the surface of scFv or on an intermediatespecies during protein refolding. Such influences of the linker sizeare difficult to predict without the knowledge of the 3-dimensionalstructure of a recombinant protein.
Since mutations in the variable region could influence scFvsecretion and, also, cause aggregation of the folding intermediate[56] thereby affecting antigen recognition, the mutant scFv4m-18L (Asp98 to Gly98) was analyzed for its functional properties.The BoHV-1 antigen recognition ability of the mutant scFv4m-18Lwas comparable to scFv3-18L and the parent monoclonal IgG1antibody against BoHV-1 [36] in an immunofluorescence assay.Therefore, the single replacement mutation in the FR3 did not affectantigen-binding per se. Nevertheless, the mutation in FR3 (Asp98to Gly98) of scFv4m-18L did influence viral neutralization as virusneutralization end point was achieved by 2.7-fold higher proteinconcentration. These observations suggest that a single substitutionmutation in FR3 influences antigen-combining site configurationin a subtle manner such that it can modify virus neutralizationdynamics. The X-ray crystallographic studies are required to under-stand such subtle differences in the composition of FR regions thatmay affect antigen recognition and neutralization function. Thismight be related to slight differences in the affinity or the configu-rational variation or stability during antigen–antibody interactionin the context of viral neutralization. Little is known about the

Author's personal copy
M. Koti et al. / Vaccine 29 (2011) 7905– 7912 7911
fine functional outcome of somatic mutations in antibody variableregion, though somatic mutations are known to influence affin-ity of antigen recognition [57]. A recent study demonstrated thata naturally occurring 3 amino acid insertion in the heavy-chainvariable region of influenza virus-specific human antibody 2D1reconfigured the antibody-combining site resulting in increasedanti-viral potency [58]. The structure of the wild-type antibodyrevealed that the insertion at the base of CDRH2 led to the loopenlargement causing distortion of CDRH1 architecture and displac-ing it away from the antigen-combining site. Further, removal ofthe insertion resulted in markedly reduced antigen-binding andneutralization activity because of steric conflict of CDRH1 withthe target hemagglutinin epitope. These observations are consis-tent with the differences noted with regard to viral neutralizationactivity between scFv3-18L and scFv4m-18L as a result of con-figurational variation in antigen-combining site originating froma single substitution mutation (Asp98 to Gly98). Whether secre-tion of mutant scFv4m-18L was enhanced or decreased is not yetdetermined since mutations in FR residues are suggested to triggeraggregation of a folding intermediate and toxicity to the bacteria[56]. Overall, the experimental evidence suggests that recombi-nant scFv with a single substitution mutation in FR3 may havesubtle influence on the architecture of the antigen-combining siteaffecting the affinity or functional activity against a target epitope.Availability of BoHV-1 neutralizing scFvs provide a useful anti-viralagent for possible passive immunization that may be administeredvia mucosal route, e.g., intra-nasal sprays, as available vaccinesagainst this pathogen are ineffective.
Acknowledgement
The outlined research in this manuscript was supported byNSERC Canada Discovery grant to Azad K. Kaushik.
References
[1] Tonegawa S. Somatic generation of antibody diversity. Nature 1983;3023(April(5909)):575–81.
[2] Klee GG. Human anti-mouse antibodies. Arch Pathol Lab Med 2000;124(June(6)):921–3.
[3] Morrison SL, Johnson MJ, Herzenberg LA, Oi VT. Chimeric human antibodymolecules: mouse antigen-binding domains with human constant regiondomains. Proc Natl Acad Sci USA 1984;81(November (21)):6851–5.
[4] Boulianne GL, Hozumi N, Shulman MJ. Production of functional chimaericmouse/human antibody. Nature 1984;312(December (5995)):643–6.
[5] Knight KL, Suter M, Becker RS. Genetic engineering of bovine Ig. Constructionand characterization of hapten-binding bovine/murine chimeric IgE, IgA, IgG1,IgG2, and IgG3 molecules. J Immunol 1988;140(May (10)):3654–9.
[6] Jones PT, Dear PH, Foote J, Neuberger MS, Winter G. Replacing thecomplementarity-determining regions in a human antibody with those from amouse. Nature 1986;321(May–June (6069)):522–5.
[7] Co MS, Queen C. Humanized antibodies for therapy. Nature 1991;351(June(6326)):501–2.
[8] Winter G, Harris WJ. Humanized antibodies. Immunol Today 1993;14(June(6)):243–6.
[9] Rader C, Cheresh DA, Barbas III CF. A phage display approach for rapid antibodyhumanization: designed combinatorial V gene libraries. Proc Natl Acad Sci USA1998;95(July (15)):8910–5.
[10] Barbas III CF, Kang AS, Lerner RA, Benkovic SJ. Assembly of combinatorial anti-body libraries on phage surfaces: the gene III site. Proc Natl Acad Sci USA1991;88(September (18)):7978–82.
[11] Glockshuber R, Malia M, Pfitzinger I, Pluckthun A. A comparison of strategiesto stabilize immunoglobulin Fv-fragments. Biochemistry 1990;29(February(6)):1362–7.
[12] FitzGerald K, Holliger P, Winter G. Improved tumour targeting by disulphidestabilized diabodies expressed in Pichia pastoris. Protein Eng 1997;10(October(10)):1221–5.
[13] Colcher D, Bird R, Roselli M, Hardman KD, Johnson S, Pope S, et al. In vivo tumortargeting of a recombinant single-chain antigen-binding protein. J Natl CancerInst 1990;82(July (14)):1191–7.
[14] Yokota T, Milenic DE, Whitlow M, Schlom J. Rapid tumor penetration of asingle-chain Fv and comparison with other immunoglobulin forms. Cancer Res1992;52(June (12)):3402–8.
[15] Adams GP, Schier R, McCall AM, Crawford RS, Wolf EJ, Weiner LM, et al. Pro-longed in vivo tumour retention of a human diabody targeting the extracellulardomain of human HER2/neu. Br J Cancer 1998;77(May (9)):1405–12.
[16] Beck A, Wurch T, Bailly C, Corvaia N. Strategies and challenges for the nextgeneration of therapeutic antibodies. Nat Rev Immunol 2010;10(May (5)):345–52.
[17] Dubel S. Handbook of therapeutic antibodies: technologies, emerging develop-ments and approved therapeutics. Wiley–Blackwell; 2009.
[18] Holliger P, Hudson PJ. Engineered antibody fragments and the rise of singledomains. Nat Biotechnol 2005;23(September (9)):1126–36.
[19] Chan AC, Carter PJ. Therapeutic antibodies for autoimmunity and inflammation.Nat Rev Immunol 2010;10(May (5)):301–16.
[20] ShengFeng C, Ping L, Tao S, Xin W, GuoFeng W. Construction, expression, purifi-cation, refold and activity assay of a specific scFv fragment against foot andmouth disease virus. Vet Res Commun 2003;27(April (3)):243–56.
[21] Cardinale A, Filesi I, Vetrugno V, Pocchiari M, Sy MS, Biocca S. Trapping prionprotein in the endoplasmic reticulum impairs PrPC maturation and preventsPrPSc accumulation. J Biol Chem 2005;280(January (1)):685–94.
[22] Donofrio G, Heppner FL, Polymenidou M, Musahl C, Aguzzi A. Paracrine inhi-bition of prion propagation by anti-PrP single-chain Fv miniantibodies. J Virol2005;79(July (13)):8330–8.
[23] Padiolleau-Lefevre S, Alexandrenne C, Dkhissi F, Clement G, Essono S, BlacheC, et al. Expression and detection strategies for an scFv fragment retaining thesame high affinity than Fab and whole antibody: implications for therapeuticuse in prion diseases. Mol Immunol 2007;44(March (8)):1888–96.
[24] Koti M, Farrugia W, Nagy E, Ramsland PA, Kaushik AK. Construction of single-chain Fv with two possible CDR3H conformations but similar inter-molecularforces that neutralize bovine herpesvirus 1. Mol Immunol 2010;47(February(5)):953–60.
[25] Tschopp JF, Brust PF, Cregg JM, Stillman CA, Gingeras TR. Expression of the lacZgene from two methanol-regulated promoters in Pichia pastoris. Nucleic AcidsRes 1987;15(May (9)):3859–76.
[26] Cregg JM, Madden KR, Barringer KJ, Thill GP, Stillman CA. Functional character-ization of the two alcohol oxidase genes from the yeast Pichia pastoris. Mol CellBiol 1989;9(March (3)):1316–23.
[27] Cereghino GP, Cereghino JL, Ilgen C, Cregg JM. Production of recombinant pro-teins in fermenter cultures of the yeast Pichia pastoris. Curr Opin Biotechnol2002;13(August (4)):329–32.
[28] Huston JS, Tai MS, McCartney J, Keck P, Oppermann H. Antigen recognition andtargeted delivery by the single-chain Fv. Cell Biophys 1993;22(January–June(1–3)):189–224.
[29] Borrebaeck CA. Antibody engineering. 2nd ed. USA: Oxford University Press;1994.
[30] Freund C, Ross A, Guth B, Pluckthun A, Holak TA. Characterization of the linkerpeptide of the single-chain Fv fragment of an antibody by NMR spectroscopy.FEBS Lett 1993;320(April (2)):97–100.
[31] Freund C, Ross A, Pluckthun A, Holak TA. Structural and dynamic propertiesof the Fv fragment and the single-chain Fv fragment of an antibody in solutioninvestigated by heteronuclear three-dimensional NMR spectroscopy. Biochem-istry 1994;33(March (11)):3296–303.
[32] Bird RE, Hardman KD, Jacobson JW, Johnson S, Kaufman BM, Lee SM,et al. Single-chain antigen-binding proteins. Science 1988;242(October(4877)):423–6.
[33] Whitlow M, Bell BA, Feng SL, Filpula D, Hardman KD, Hubert SL, et al. Animproved linker for single-chain Fv with reduced aggregation and enhancedproteolytic stability. Protein Eng 1993;6(November (8)):989–95.
[34] Kortt AA, Malby RL, Caldwell JB, Gruen LC, Ivancic N, Lawrence MC, et al.Recombinant anti-sialidase single-chain variable fragment antibody. Charac-terization, formation of dimer and higher-molecular-mass multimers and thesolution of the crystal structure of the single-chain variable fragment/sialidasecomplex. Eur J Biochem 1994;221(April (1)):151–7.
[35] Perisic O, Webb PA, Holliger P, Winter G, Williams RL. Crystal structureof a diabody, a bivalent antibody fragment. Structure 1994;2(December(12)):1217–26.
[36] Levings RL, Stoll IR, inventor. Bovine monoclonal antibodies to bovine her-pesvirus 1 from sequential fusion heterohybridomas. U.S.A.; 1991.
[37] Saini SS, Allore B, Jacobs RM, Kaushik A. Exceptionally long CDR3H region withmultiple cysteine residues in functional bovine IgM antibodies. Eur J Immunol1999;29(August (8)):2420–6.
[38] Saini SS, Hein WR, Kaushik A. A single predominantly expressed polymorphicimmunoglobulin VH gene family, related to mammalian group, I, clan, II, isidentified in cattle. Mol Immunol 1997;34(June (8–9)):641–51.
[39] Lefranc MP, Pommie C, Ruiz M, Giudicelli V, Foulquier E, Truong L, et al.IMGT unique numbering for immunoglobulin and T cell receptor variabledomains and Ig superfamily V-like domains. Dev Comp Immunol 2003;27(Jan-uary (1)):55–77.
[40] Laemmli UK. Cleavage of structural proteins during the assembly of the headof bacteriophage T4. Nature 1970;227(August (5259)):680–5.
[41] Silva SV, Little PB, Kaushik A. An immunodominant epitope on 40 kilodaltonouter membrane protein is conserved among different strains of Haemophilus(Histophilus) somnus. Zentralbl Bakteriol 1995;282(October (4)):449–56.
[42] Martin SW, Nagy E, Armstrong D, Rosendal S. The associations of viral andmycoplasmal antibody titers with respiratory disease and weight gain in feed-lot calves. Can Vet J 1999;40(August (8)):560–7, 70.
[43] Carter PJ. Potent antibody therapeutics by design. Nat Rev Immunol2006;6(May (5)):343–57.

Author's personal copy
7912 M. Koti et al. / Vaccine 29 (2011) 7905– 7912
[44] Boder ET, Wittrup KD. Yeast surface display for screening combinatorialpolypeptide libraries. Nat Biotechnol 1997;15(June (6)):553–7.
[45] Sinclair MC, Gilchrist J, Aitken R. Molecular characterization of bovine V lambdaregions. J Immunol 1995;155(September (6)):3068–78.
[46] Saini SS, Farrugia W, Ramsland PA, Kaushik AK. Bovine IgM antibodies withexceptionally long complementarity-determining region 3 of the heavy chainshare unique structural properties conferring restricted VH + Vlambda pairings.Int Immunol 2003;15(July (7)):845–53.
[47] Olafsen T, Cheung CW, Yazaki PJ, Li L, Sundaresan G, Gambhir SS, et al. Covalentdisulfide-linked anti-CEA diabody allows site-specific conjugation and radio-labeling for tumor targeting applications. Protein Eng Des Sel 2004;17(January(1)):21–7.
[48] Cochlovius B, Kipriyanov SM, Stassar MJ, Schuhmacher J, Benner A,Moldenhauer G, et al. Cure of Burkitt’s lymphoma in severe combined immun-odeficiency mice by T cells, tetravalent CD3 × CD19 tandem diabody, and CD28costimulation. Cancer Res 2000;60(August (16)):4336–41.
[49] Turner DJ, Ritter MA, George AJ. Importance of the linker in expression of single-chain Fv antibody fragments: optimisation of peptide sequence using phagedisplay technology. J Immunol Methods 1997;205(June (1)):43–54.
[50] Dall’Acqua W, Carter P. Antibody engineering. Curr Opin Struct Biol1998;8(August (4)):443–50.
[51] Huston JS, Levinson D, Mudgett-Hunter M, Tai MS, Novotny J, Margolies MN,et al. Protein engineering of antibody binding sites: recovery of specific activityin an anti-digoxin single-chain Fv analogue produced in Escherichia coli. ProcNatl Acad Sci USA 1988;85(August (16)):5879–83.
[52] Kortt AA, Lah M, Oddie GW, Gruen CL, Burns JE, Pearce LA, et al. Single-chainFv fragments of anti-neuraminidase antibody NC10 containing five- and ten-residue linkers form dimers and with zero-residue linker a trimer. Protein Eng1997;10(April (4)):423–33.
[53] Arndt KM, Muller KM, Pluckthun A. Factors influencing the dimer tomonomer transition of an antibody single-chain Fv fragment. Biochemistry1998;37(September (37)):12918–26.
[54] Atwell JL, Breheney KA, Lawrence LJ, McCoy AJ, Kortt AA, Hudson PJ. scFv mul-timers of the anti-neuraminidase antibody NC10: length of the linker betweenVH and VL domains dictates precisely the transition between diabodies andtriabodies. Protein Eng 1999;12(July (7)):597–604.
[55] Desplancq D, King DJ, Lawson AD, Mountain A. Multimerization behaviour ofsingle chain Fv variants for the tumour-binding antibody B72.3. Protein Eng1994;7(August (8)):1027–33.
[56] Knappik A, Pluckthun A. Engineered turns of a recombinant antibody improveits in vivo folding. Protein Eng 1995;8(January (1)):81–9.
[57] Ohlin M, Borrebaeck CA. Insertions and deletions in hypervariable loopsof antibody heavy chains contribute to molecular diversity. Mol Immunol1998;35(March (4)):233–8.
[58] Krause JC, Ekiert DC, Tumpey TM, Smith PB, Wilson IA, Crowe Jr JE. An insertionmutation that distorts antibody binding site architecture enhances function ofa human antibody. MBio 2011;2(1).
Related Documents











